
Precipitating Polyelectrolyte-Surfactant Systems by Admixing ...
Upload
independentCategory
view
0download
0
R1
Title: The physiologic and inflammatory response to instillation of an oxidized surfactant in a rat model of surfactant deficiency.
Authors: Timothy C. Bailey1, Keith A. Da Silva1, James F. Lewis1,3, Karina Rodriguez-Capote2, Fred Possmayer2 and Ruud A.W. Veldhuizen1,3
Affiliations: Departments of 1Physiology and Pharmacology, 2Biochemistry and3Medicine, Lawson Health Research Institute, University of Western Ontario, London, Ontario, Canada, N6A 4V2
Running Title: Physiologic effect of in vitro oxidized surfactant
Corresponding Author:Timothy C. BaileyLawson Health Research Institute, H417268 Grosvenor Street, London, ONT, CanadaN6A 4V2e-mail: [email protected]: 519-646-6100 ext. 64110fax: 519-646-6110
Articles in PresS. J Appl Physiol (December 29, 2003). 10.1152/japplphysiol.01143.2003
Copyright (c) 2003 by the American Physiological Society.

2
Abstract:Pulmonary surfactant is a mixture of phospholipids (~90%) and surfactant
associated proteins (SP’s) (~10%) that stabilize the lung by reducing the surface tension.
One proposed mechanism by which surfactant is altered during acute lung injury is via
direct oxidative damage to surfactant. In vitro studies have revealed that the surface
activity of oxidized surfactant was impaired and that this effect could be overcome by
adding SP-A. Based on this information, we hypothesized that animals receiving oxidized
surfactant preparations would exhibit an inferior physiologic and inflammatory response
and the addition of SP-A to the oxidized preparations would ameliorate this response. To
test this hypothesis mechanically ventilated, surfactant deficient rats were administered
either bovine lipid extract surfactant (BLES) or in vitro oxidized BLES of three doses;
10mg/kg, 50mg/kg or 10mg/kg + SP-A. When instilled with 10mg/kg normal surfactant
the rats had a significantly superior PaO2 responses compared to the rats receiving
oxidized surfactant. Interestingly, increasing the five times dose mitigated this
physiologic effect and the addition of SP-A to the surfactant preparation had little impact
on improving oxygenation. There were no differences in alveolar surfactant pools and the
indices of pulmonary inflammation between the 10mg/kg dose groups, neither was there
any differences observed between either of the groups supplemented with SP-A.
However, there was significantly more surfactant and more inflammatory cytokines in the
50mg/kg oxidized BLES group compared to the 50mg/kg BLES group. We conclude that
instillation of an in vitro oxidized surfactant causes an inferior physiologic response in a
surfactant deficient rat.
Key words: alveolar metabolism, biophysical function and acute lung injury

3
Introduction:
Pulmonary surfactant is a mixture of phospholipids (~90%) and surfactant
associated proteins (SP’s) (~10%) that stabilize the whole lung by reducing the surface
tension at the air- liquid interface of each alveolus (34). This important function of
pulmonary surfactant is impaired in the lungs of patients with acute lung injury (ALI),
thereby contributing to lung dysfunction (24, 25, 39). The specific alterations to the
surfactant system in patients with ALI include changes in its characteristic phospholipids
and protein composition, a reduction of the relative amount of the active subtype of
surfactant, the large aggregates (LA), and an impaired surface tension reducing activity
(21, 26).
One proposed mechanism by which surfactant metabolism and function are
altered during ALI is via direct oxidative damage to surfactant. Two lines of evidence
support this concept. First, increased production of reactive oxygen species (ROS) has
been detected in the lungs of patients with ALI (22, 23, 36, 41). These ROS are produced
during the process of inflammation when phagocytic cells are recruited to the lung where
they subsequently release a wide variety of highly reactive oxidative species. This
oxidative damage is likely exacerbated via the common supportive intervention for
patients with ALI, mechanical ventilation, which is often utilized in combination with
high concentrations of oxygen to stabilize gas exchange in these patients. In this setting
mechanical ventilation has been shown to cause further increases in pulmonary
inflammation (37) and the increased oxygen provides more substrate for the production
of oxidative species.

4
The second line of evidence for a potential role of oxidized surfactant in the
development of surfactant and lung dysfunction stems from in vitro experiments. Several
studies have exposed various isolated surfactant preparations to different oxidizing
conditions and have demonstrated the production of phospholipid peroxidation products
and impaired surface tension reducing properties of the oxidized preparations (1, 14, 29,
32). For example, we have recently exposed a commercial exogenous surfactant that
contains a normal phospholipid contingent and only the hydrophobic surfactant
associated proteins (SP-B and SP-C), bovine lipid extract surfactant (BLES), to
hypochlorous acid and to the Fenton reaction. Analysis of the resulting oxidized
surfactant in a captive bubble surfactometer revealed that the surface activity was
impaired and that this effect could be overcome by adding surfactant protein A (SP-A) to
the oxidized preparation (35). Unfortunately, the in vitro analyses do not take into
account all the complex metabolic aspects of surfactant synthesis, secretion and
catabolism that could potentially influence its activity within the lung.
We hypothesized that animals receiving oxidized surfactant preparations would
exhibit an inferior physiologic and inflammatory response and the addition of SP-A to the
oxidized preparations would ameliorate this response. These studies addressed the
following 3 specific questions: what is the physiologic response to oxidized surfactant in
a surfactant deficient lung? How is the oxidized surfactant metabolized within the
airspace? And how does oxidized surfactant affect the inflammatory mediators associated
with lung dysfunction?

5
Materials and Methods:
Experimental Groups:
To examine the physiologic impact of an oxidized surfactant system in vivo, rats
were made surfactant deficient by repetitive saline lavage and instilled with either BLES
or in vitro oxidized BLES (OX). 10mg/kg phospholipid was chosen as one dose because
it has been demonstrated to return alveolar levels of phospholipids back to pre- lavage
levels following two hours of mechanical ventilation (6). Two other doses were also
used; 50 mg/kg phospholipid and a 10mg/kg phospholipid dose that was supplemented
with 2.5% bovine SP-A.
Surfactant preparations:
BLES was obtained from BLES Biochemicals (London, Ontario, Canada) and the
oxidation process with hypochlorous acid according to previously established conditions
(35). To avoid variable levels of oxidation, this oxidization process was performed on the
entire amount of surfactant required for our studies. The non-oxidized control surfactant
utilized was from the same production batch as the oxidized material. The SP-A was
extracted from natural bovine alveolar surfactant and purified by chromatography with a
mannose column as described by Cockshutt et al (9).
Animal Experimentation:
Male Sprague-Dawley rats, weighing between 350-500g were used for these
experiments (Charles River, Constant, PQ, Canada). Animals were acclimatized for 3
days during which time they were group-housed and allowed free access to water and
standard chow. The mass was recorded and the animals were anaesthetized (75 mg/kg
Ketamine / 5mg/kg Xylazine in sterile 0.15M saline) with an intra-peritoneal injection.

6
Once anaesthetized the animal received analgesic (Buprenorphine: 0.05-0.1mg/kg) intra-
muscularly. The right jugular vein and the right carotid artery were exposed and
cannulated with P50 tubing. The venous line was used for further administration of drugs.
The carotid line was used to take measures of heart rate, blood pressures, to obtain
samples for arterial blood gas measurements and for the instillation of fluids (sterile
0.15M saline and 1000 i.u. heparin/l) via an infusion pump at a rate of 1mL/100g/hr. The
trachea was exposed and an endotracheal tube (14 gauge) was secured in place with 2.0
surgical silk. Immediately upon securing the endotracheal tube the animal was
administered a neuro-muscular paralytic intravenously (pancurium bromide: 2 mg/kg) to
inhibit spontaneous respiratory movements. The endotracheal tube was connected to a
rodent, volume cycle mechanical ventilator (Harvard Instruments, Saint Laurent, PQ,
Canada) set at a tidal volume of 7ml/kg, 5cmH20 positive end expiratory pressure
(PEEP), respiratory rate of 58 breaths / minute and 100% oxygen. In parallel with the
ventilator was an airway pressure monitor (Sechrist Industries Inc., Anaheim CA, USA).
Sodium pentobarbital (20-40mg/kg) was administered i.v. as further anaesthetic and to
stabilize systemic blood pressure below 100mmHg. The initial inclusion criteria were an
arterial partial pressure of oxygen (PaO2) greater than 400mmHg, as measured in arterial
blood samples (ABL500 Radiometer, Copenhagen, Denmark) and peak inspiratory
pressure (PIP) less than 15cmH20.
Upon meeting the initial criteria the animal was subjected to a series of whole
lung lavages, as previously described (5, 20). Briefly, PIP was recorded and the animal
was immediately disconnected from the ventilator and a 10c.c. syringe filled with 37oC
saline (0.15M NaCl) was connected to the endotracheal tube and flushed into and gently

7
pulled back out of the lungs of the animal. The animal was re-connected to the ventilator
and the volume of the recovered material was recorded. This procedure was repeated four
times with 5 minutes between each lavage and another blood gas measure was taken.
Following this blood gas the lavage procedure was repeated, with subsequent blood gases
measured after every two lavages until the animal met the following inclusion criteria; a
PaO2 less than 100mmHg that was stable for 10 minutes and an increase of greater than
3cmH20 from the baseline PIP. Based upon previously published data by our laboratory
these physiologic parameters, following repetitive saline lavage, are indicative of a
surfactant deficient state (5). Immediately after reaching these secondary inclusion
criteria the animal was randomized into one of the following: instillation of a 1ml air
bolus (no-Rx), 10mg/kg BLES (10 BLES), 10mg/kg oxidized BLES (10 OX), 50mg/kg
BLES (50 BLES), 50mg/kg oxidized BLES (50 OX), 10mg/kg BLES + SP-A (2.5% of
phospholipids by weight) (BLES + SP-A) or 10mg/kg oxidized BLES + SP-A (OX +
SP-A). All administration volumes were 1ml, regardless of group. At the time of
instillation the animal was disconnected from the mechanical ventilator, supported in an
upright position and the surfactant preparation was administered intra-tracheally,
followed immediately by a 1.5mL air bolus to ensure distal distribution. The animal was
then placed back in the supine position and the endotracheal tube was reconnected to the
ventilator.
The animal was ventilated for 2 hours and blood gases were taken 5, 15, 30, 60,
90 and 120 minutes post-surfactant administration, during which time blood pressure was
maintained below 100mmHg with sodium pentobarbital i.v. and pancurium bromide
(2mg/kg) was administered every hour following the initial dose. Upon completion of the

8
ventilation protocol the animal was sacrificed with an overdose of sodium pentobarbital
and exsanguination by transection of the dorsal aorta.
A mid-line sternectomy was performed to remove the chest wall and the lungs
were lavaged with 5 x 10ml of 0.15M NaCl. Each lavage was instilled and withdrawn
three times and all the lavages were combined and the total volumes were recorded.
Lavage Analysis:
One ml was aliquoted, from the first 10ml lavage, for cytokine measurements as
described below. The remaining lavage material was centrifuged at 150 x g for 10
minutes to remove any cellular debris. The supernatant from this centrifugation was
termed the Total Surfactant (TS) from which 5ml was saved for phospholipid analysis.
The remaining TS was subjected to 15 minutes of 40,000 x g centrifugation, from which
the supernatant was termed the Small Aggregates (SA) and the pellet was re-suspended in
2ml of saline to produce a Large Aggregate (LA) suspension. A Duck-Chong
phosphorous assay was performed to determine the amount of phospholipid within each
of the TS, LA and SA fractions (11). A Lowry protein assay was performed on the TS to
determine the amount of total protein in the lavage (27).
Cytokine Measurements:
The 1ml aliquot of lavage material was centrifuged for 10 minutes at 200 x g. The
supernatant was separated into three equal aliquots, which were immediately snap frozen
in liquid nitrogen and stored at –80oC until analysis. The pro-inflammatory cytokines
Tumor Necrosis Factor – alpha (TNF-α) and Interleukin – 6 (Il-6) were both measured by
using opti-EIA ELISA kits and following the manufacturers instructions (Pharmigen San
Diego, CA, USA).

9
Surface Tension Reducing Activity:
Based on phosphorous analysis, LA samples were re-suspended at 2.5 mg PL/ml
in 0.15 M NaCl and 1.5mM CaCl2. The samples were incubated for 1hour at 37ºC and
surface tension reduction properties were assessed on a pulsating bubble surfactometer
(12).
Statistical Analysis: All data are presented as mean " SEM. Between 2 experimental
groups statistical analysis was performed using a Student’s T-Test and between 3
experimental groups a one-way ANOVA with a Tukey’s post hoc test was performed
with SPSS software package for Windows, version 9.0.0 (SPSS Chicago IL, USA). All
comparisons were considered statistically significant at probability values lower than
0.05.
Results:
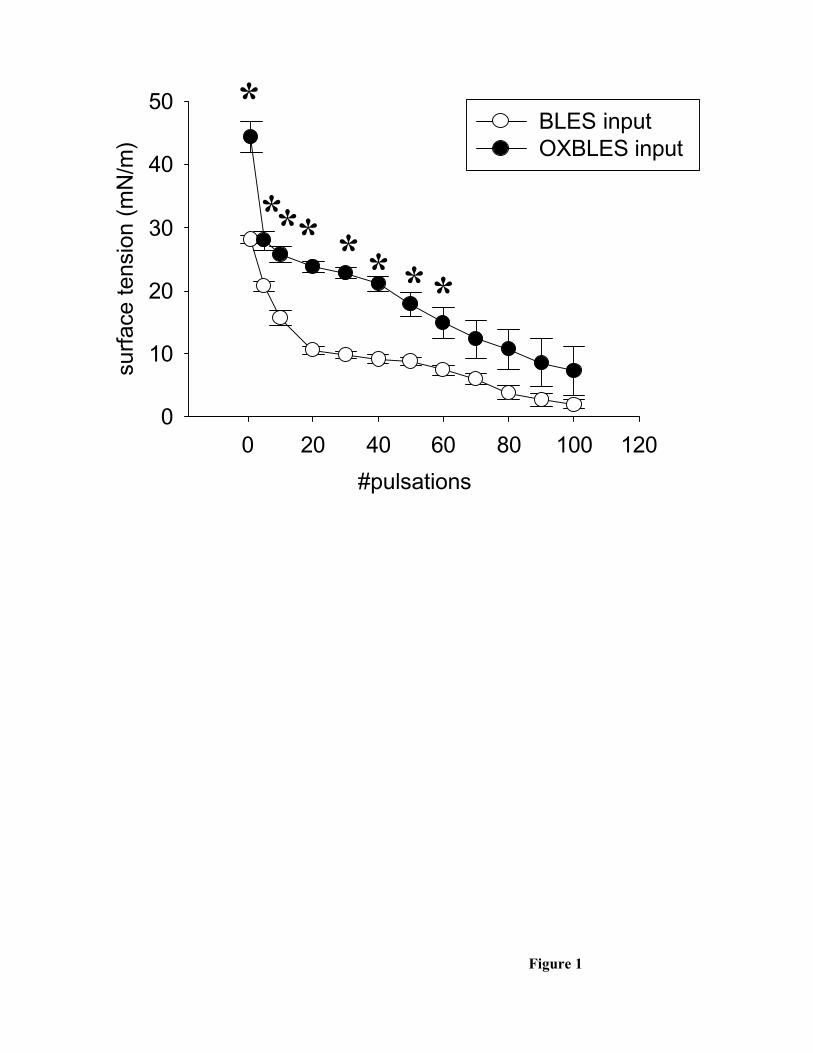
There were significant differences in surface tension reducing ability between the
inputs of normal BLES and the oxidized BLES as analyzed on the pulsating bubble
surfactometer (figure 1). Following the brief adsorption time the BLES samples had a
significantly lower surface tension, which remained significantly lower through out the
subsequent 60 pulsations. Although the trend persisted beyond 60 pulsations, no further
significant differences were observed between the two input samples.
The mean mass of all the animals utilized in these studies was 436 ± 6 grams and
was not different among the experimental groups. The total number of pre-administration
lavages and the volume of total lavaged material recovered were not different between all
groups (data not shown). All animals that met the inclusion criteria had similar

10
physiologic indices reflective of a surfactant deficient state; PaO2 of 73.0 ± 1.3 mmHg,
PaCO2 of 53.8 ± 0.8 mmHg and PIP values of 22.1 ± 0.3 cmH20.
10mg/kg Dose
Figure 2A represents the arterial oxygenation (PaO2) during the 120 minutes
mechanical ventilation time course following intra-tracheal instillation of either 10mg/kg
BLES or 10mg/kg oxidized BLES, 2B represents the surfactant aggregate phospholipid
levels and the surface tension reducing activity of the LA sub-fraction is shown in figure
2C. The 10 BLES group had PaO2 levels that were significantly higher than the animals
instilled with 10 OX (figure 2A). This difference was consistent through out the entire
120 minutes of mechanical ventilation. There was no difference in the PaCO2 levels
between the 10 BLES and the 10 OX at 5 minutes (51.1 ± 3.5 vs 56.0 ± 4.8 mmHg) or
120 minutes (42.4 ± 3.4 vs 44.5 ± 4.1mmHg) post instillation. Similarly, no significant
differences were observed in the measurements of PIP between the 10 BLES and 10 OX
groups at 5 minutes post- instillation (17.0 ± 0.5 vs 21.0 ± 1.5 cmH20) and 120 minutes
(16.8 ± 0.8 vs 18.1 ± 1.3 cmH20).
There were no significant differences in the phospholipid levels measured in the
total surfactant (TS), large aggregates (LA) or small aggregates (SA) between the 10
BLES and the 10 OX groups (figure 2B).
When the LA were assessed for surface tension reducing ability the 10 BLES
group to achieved lower surface tensions with fewer pulsations compared to the LA
recovered from the 10 OX group, however this did not reach statistical significance
(figure 2C).

11
Table 1 shows the total protein values, TNF-α and IL-6 concentrations measured
in the lavage of the no-Rx, 10 BLES and 10 OX groups, of which there were no
differences among all three groups.
50 mg/kg Dose
Figure 3A represents the PaO2 response, figure 3B the surfactant aggregate
phospholipid levels and the surface tension reducing activity of the LA sub-fraction,
shown in figure 3C, following instillation with 50mg/kg BLES or 50mg/kg oxidized
BLES and 120 minutes of mechanical ventilation. PaO2 levels were the same between
animals instilled with 50 BLES and 50 OX throughout the entire 120 minutes of
mechanical ventilation (figure 3A).
There was significantly more phospholipid within the TS and LA fractions in the
50 OX group compared to the 50 BLES group and no differences in the levels of small
aggregates (SA) (figure 3B).
The LA recovered from the 50 BLES group achieved lower surface tensions than
the LA recovered from the 50 OX group. This difference was statistically significant at
pulsations 50-80 (figure 3C).
There was no significant difference in the total protein and IL-6 concentrations in
the lavaged material between the 50 BLES group and 50 OX group (table 2). There was
a significantly higher concentration of TNF-α in the lavage material of the 50 OX group
compared to the 50 BLES group.
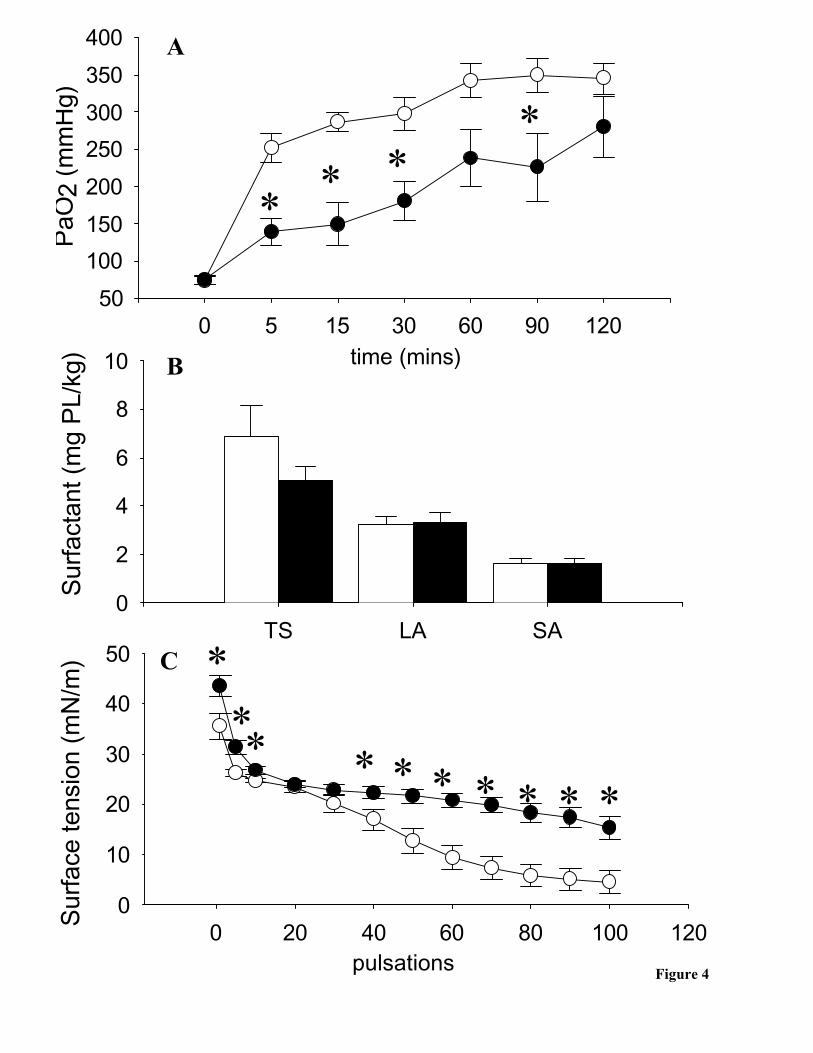
10mg/kg Dose + SP-A
Similar to the 10mg/kg groups, 5 minutes following instillation with BLES + SP-A
PaO2 levels were significantly higher than the animals administered OX + SP-A (figure 4A).

12
Superior oxygenation was consistent throughout the ventilation time course, although it was
not statistically different at the 60 and 120 minutes time points.
There were no significant differences between the phospholipid levels within the
total surfactant (TS), large aggregates (LA) and small aggregates (SA) between either of
the BLES + SP-A or OX + SP-A groups (figure 4B).
The LA recovered from the BLES + SP-A group had significantly lower initial
surface tensions and significantly lower surface tensions throughout the pulsation time
course compared to the LA recovered from the OX + SP-A group (figure 4C).
There were no differences in the concentrations of TNF-α and IL-6 in the lavage
material recovered from both the BLES + SP-A and OX + SP-A groups (table 3).
Discussion:
Due to its locale within the airspace, the presence of unsaturated phospholipids
and associated proteins, pulmonary surfactant has been identified as a potential target of
reactive oxygen species (ROS) (1, 8, 35). Furthermore, based on in vitro studies, the
oxidation of surfactant has been proposed as a mechanism by which surfactant
metabolism and biophysical function are altered and can contribute to the
pathophysiology of ALI (1, 14, 29, 30, 35). In this study we examined the physiologic
effects of an in vitro oxidized exogenous surfactant preparation supplemented with and
without SP-A. We observed that when surfactant deficient rats are instilled with 10mg/kg
normal surfactant the PaO2 response is significantly superior compared to the rats
receiving oxidized surfactant. Interestingly, increasing the dose five fold mitigated this
physiologic effect while the addition of SP-A to the surfactant preparation had little

13
impact on improving oxygenation. This study provides in vivo evidence that oxidization
of surfactant may contribute to lung dysfunction in ALI.
Numerous investigators have reported increased oxidative stress as a factor that
contributes to the pathophysiology of ALI (8, 13, 17, 22). Accordingly, in vivo models
have been utilized to mimic the oxidative stress that is accrued during the progression of
ALI, such as exposure to prolonged hyperoxia and ozone (2, 3, 10, 31). A limitation of
these types of in vivo studies is that it is difficult to ascertain the specific contribution of
an altered surfactant system to the lung injury versus the other contributing factors that
are affected by hyperoxia exposure such as oxidative damage to the pulmonary
epithelium or endothelium and the resulting inflammatory response. One approach that
has been utilized to circumvent this issue is to examine the biochemical and biophysical
consequences of an in vitro oxidized surfactant (1, 16, 32). Our experiment has extended
those previous studies by examining the physiological response to oxidized surfactant by
instilling normal and oxidized surfactant preparations into the surfactant deficient lungs
of saline lavaged rats (5, 7, 18). A limitation of this approach is that the lung lavage
protocol may induce some edema formation in addition to the surfactant deficiency.
Although the physiological responses to the surfactant may have been affected by this
additional damage, the observation that the total protein concentrations in the lung
lavages were not different among the groups would indicate that edema formation was
similar in all groups.
Another important aspect of our experiment was the specific oxidizing condition
utilized. We used the previously established conditions of hypochlorous acid exposure
(35), which represented a high pathophysiological exposure level as suggested by Merritt

14
and colleagues (1, 32). It should be considered however that oxidization was limited to
one oxidizing agent, hypochlorous acid, and one surfactant, BLES. Previous studies have
demonstrated that the effect of reactive oxygen and nitrogen species exposure on
surfactant activity may depend on the surfactant composition as well as the specific
oxidizing agent utilized (1, 8, 16, 35). These differences are due to the fact that different
oxidizing agents, such as the Fenton reaction, peroxynitrite or hypochlorous acid, may
have different targets within a specific surfactant preparation. For example, inactivation
of surfactant by peroxynitrite was more severe in a surfactant preparation containing only
SP-B and C as compared to a natural surfactant preparation containing SP-A, B and C
(8). It has also been shown that the impairment of surfactant by peroxynitite was in part
attributed to damage to the small hydrophobic surfactant proteins (16). We have
previously demonstrated that exposure of BLES to hypochlorous acid led to the
formation of oxidized phosphatidylcholine species (35). Thus, although the current study
provides a first indication of the in vivo consequences of an oxidized surfactant, further
studies investigating the surfactant proteins as a potential target for the oxidizing agent as
well as the in vivo responses of other surfactant preparations and/or other oxidizing
conditions are warranted.
The main physiological result from the current study was that the administration
of 10mg/kg BLES resulted in significantly higher PaO2 levels than the instillation of
10mg/kg oxidized BLES. This observation supports the concept that an oxidized
surfactant in the air space is less capable of maintaining lung function than non-oxidized
surfactant. The analysis of the lavage material indicated that at least for this dose,
oxidized and normal BLES were metabolized similarly since surfactant pool sizes were

15
similar. Despite the difference in bloodgas values, the LA’s biophysical function
following the two hours of mechanical ventilation was not significantly different between
the two groups. It is likely that this apparent difference between the activity in vitro and
in vivo results was related to differences in the experimental conditions. In general, the
activity of surfactant in vivo can be affected by distribution of the surfactant within the
lung and the presence of other material in the alveolar space, such as serum proteins. In
contrast, surface activity of LA in vitro is performed with isolated LA at a specific
concentration. Thus it is possible, for example, that oxidized surfactant is more
susceptible to protein inhibition than non-oxidized surfactant. This would affect the in
vivo function, but may not be easily detected when analyzing the activity of isolated LA.
The correlation between physiologic responses to instillation of a specific surfactant
preparation and in vitro activity requires further study.
In addition to the 10 mg/kg dose, we tested a five fold higher dose since studies in
vitro have demonstrated a concentration dependence of surfactant activity when tested by
itself as well as in the presence of inhibitory proteins (15, 19). In support of these in vitro
observations, our results showed that the 50mg/kg dose had similar oxygenation values
between oxidized BLES and BLES and these values were comparable to the 10 BLES.
Interestingly, more surfactant was recovered from rats instilled with 50mg/kg oxidized
BLES compared to BLES. This difference was largely due to an increased recovery of
LA. Although further metabolic studies are required, this result would suggest that
oxidized surfactant is removed more slowly from the airspace. The fact that no difference
in pool sizes were observed in the 10mg/kg groups may be due to the possibility that
these groups had already reached a metabolic equilibrium after 2 hours of ventilation.

16
Our third experiment involved the addition of SP-A to BLES and oxidized BLES
based on the observation that in vitro SP-A could mitigate the biophysical impairment of
oxidized surfactant (8, 16, 35). In view of these in vitro results it was surprising that
surfactant deficient rats administered SP-A containing oxidized surfactant did not have an
improved physiological response. This result was similar to the results observed with the
preparations without supplemental SP-A (compare figure 2 and 4). These results suggest
that SP-A may not improve the function of oxidized surfactant in vivo, however, some
caution is warranted with regards to this conclusion since our experiments utilized bovine
SP-A in rat lungs. Although a variety of studies have demonstrated cross-species activity
of SP-A (4, 28, 33, 38) it is possible that in our experimental set up bovine SP-A behaves
differently than rat SP-A. It should also be noted that the in vitro experiments utilized 5%
(by weight) SP-A in a calcium containing buffer (35) whereas we utilized 2.5% SP-A in
saline due to the quantities required.
For each of the three experiments we also analyzed TNF-α and IL-6 as indices of
pulmonary inflammation and lung injury. Although statistical significance was only
reached for the TNF-α concentrations in the 50mg/kg groups, in general the animals
receiving oxidized surfactant had higher concentration of the two inflammatory
cytokines. It is known that pulmonary surfactant has immuno-modulatory properties (40)
and it is possible that, in addition to the biophysical impairment, oxidization also affects
this secondary function of surfactant.
In summary, our study was based on evidence of oxidative stress in
patients/animals with ALI and on the reported in vitro impairment of oxidized surfactant.
Our study contributes to these observations by demonstrating that oxidized surfactant has

17
an impaired function in vivo. We also provided preliminary indications that oxidized
surfactant had an altered metabolism and affected the concentrations of inflammatory
cytokines in the lung. Overall these observations provide further support for a role of
oxidative damage to surfactant as a contributing mechanism to lung dysfunction in acute
lung injury. Future studies are required to address the occurrence and relative importance
of oxidative damage to surfactant in injured lungs.

18
Acknowledgements: The authors would like to thank Dr. Li-Juan Yao and Anne
Brickenden for technical assistance. We also wish to acknowledge the Ontario Thoracic
Society and Ontario Graduate Scholarship for salary support of Timothy C. Bailey and
the Canadian Institutes of Health Research for grant support.

19
Figure Legends:
Figure 1. The surface tension reducing properties of BLES inputs (open circles,
n=5) and OXBLES inputs (closed circles, n=4) as measured by a pulsating bubble
surfactometer. *=p<0.05 vs BLES input.
Figure 2. 10mg/kg group. A) Arterial oxygenation (PaO2, mmHg) obtained from the
no-Rx group (closed triangles, n=5), 10 BLES group (open circles, n=5) and the 10 OX
group (closed circles, n=6). B) Surfactant poolsizes (mg phospholipid/kg) of total
surfactant (TS), large aggregates (LA) and small aggregates (SA) obtained from 10
BLES group (open bars, n=5) and 10 OX group (closed bars, n=6). C) Surface tension
reducing properties of large aggregates obtained from 10 BLES (open circles, n=3) and
10 OX (closed circles, n=5). *=p<0.05 vs 10 BLES.
Figure 3. 50mg/kg group. A) Arterial oxygenation (PaO2, mmHg) obtained from the
50 BLES group (open circles, n=6) and the 50 OX group (closed circles, n=5). B)
Surfactant poolsizes (mg phospholipid/kg) of total surfactant (TS), large aggregates (LA)
and small aggregates (SA) obtained from 50 BLES group (open bars, n=6) and 50 OX
group (closed bars, n=5). C) Surface tension reducing properties of large aggregates
obtained from 50 BLES (open circles, n=4) and 50 OX (closed circles, n=4). *=p<0.05
vs 50 BLES.

20
Figure 4. 10mg/kg + SP-A group. A) Arterial oxygenation (PaO2, mmHg) obtained
from the BLES + SP-A group (open circles, n=6) and the OX + SP-A group (closed
circles, n=5). B) Surfactant poolsizes (mg phospholipid/kg) of total surfactant (TS), large
aggregates (LA) and small aggregates (SA) obtained from BLES + SP-A group (open
bars, n=3) and OX + SP-A group (closed bars, n=5). C) Surface tension reducing
properties of large aggregates obtained from BLES + SP-A (open circles, n=3) and 10
OX (closed circles, n=5). *=p<0.05 vs BLES + SP-A.

21
Table 1. The total amount of protein, Tumor Necrosis Factor Alpha (TNF-α) and Interleukin 6 (IL-6) in the lavaged material from animals instilled with 10mg/kg dose.
Group Total Protein TNF-α Il-6 (mg/kg) (pg/ml) (pg/ml)
no-Rx 32 ±15 (4) 608 ± 252 (5) 3162 ± 678 (5)10 BLES 37 ± 10 (3) 676 ± 223 (4) 4453 ± 1147 (5)10 OX 45 ± 8 (4) 1218 ± 615 (4) 9562 ± 2916 (5)
Table 2. The total amount of protein, Tumor Necrosis Factor Alpha (TNF-α) and Interleukin 6 (IL-6) in the lavaged material from animals instilled with 50mg/kg dose (*=p<0.05 vs 50 BLES).
Group Total Protein TNF-α Il-6 (mg/kg) (pg/ml) (pg/ml)
50 BLES 32 ± 11 (4) 344 ± 112 (4) 3135 ± 728 (5)
50 OX 65 ± 15 (5) 746 ± 113 (5) * 5024 ± 556 (5)
Table 3. The total amount of protein, Tumor Necrosis Factor Alpha (TNF-α) and Interleukin 6 (IL-6) in the lavaged material from animals instilled with 10mg/kg dose + SP-A.
Group Total Protein TNF-α Il-6 (mg/kg) (pg/ml) (pg/ml)
10 BLES + SP-A 42 ± 15 (5) 336 ± 180 (4) 6753 ± 3649 (4)10 OX + SP-A 55 ± 7 (6) 631 ± 74 (5) 4640 ± 1819 (5)

22
Reference List
1. Andersson S, Kheiter A and Merritt TA. Oxidative inactivation of surfactants.
Lung 177: 179-189, 1999.
2. Bailey TC, Cavanagh C, Mehta S, Lewis JF and Veldhuizen RA. Sepsis and
hyperoxia effects on the pulmonary surfactant system in wild- type and iNOS
knockout mice. Eur Respir J 20: 177-182, 2002.
3. Bailey TC, Martin EL, Zhao L and Veldhuizen RA. High oxygen
concentrations predispose mouse lungsto the deleterious effects of high stretch
ventilation. J Appl Physiol 94: 975-982, 2003.
4. Borron P, Veldhuizen RAW, Lewis JF, Possmayer F, Caveney A, Inchley K,
McFadden RG and Fraher LJ. Surfactant associated protein-A inhibits human
lymphocyte proliferation and IL-2 production. Am J Respir Cell Mol Biol 15: 115-
121, 1996.
5. Brackenbury AM, Malloy JL, McCaig LA, Yao LJ, Veldhuizen RA and
Lewis JF. Evaluation of alveolar surfactant aggregates in vitro and in vivo. Eur
Respir J 19: 41-46, 2002.

23
6. Brackenbury AM, Puligandla PS, McCaig LA, Nikore V, Yao LJ,
Veldhuizen RA and Lewis JF. Evaluation of exogenous surfactant in HCL-
induced lung injury. Am J Respir Crit Care Med 163: 1135-1142, 2001.
7. Campbell H, Bosma K, Brackenbury A, McCaig L, Yao LJ, Veldhuizen R
and Lewis J. Polyethylene Glycol (PEG) Attenuates Exogenous Surfactant in
Lung- injured Adult Rabbits. Am J Respir Crit Care Med 165: 475-480, 2002.
8. Cifuentes J, Ruiz-Oronoz J, Myles C, Nieves B, Carlo WA and Matalon S.
Interaction of surfactant mixtures with reactive oxygen and nitrogen species. J
Appl Physiol 78: 1800-1805, 1995.
9. Cockshutt AM, Weitz J and Possmayer F. Pulmonary surfactant-associated
protein A enhances the surface activity of lipid extract surfactant and reverses
inhibition by blood proteins in vitro. Biochemistry 29: 8424-8429, 1990.
10. Currie WD, van Schaik S, Vargas I and Enhorning G. Breathing and
pulmonary surfactant function in mice 24 h after ozone exposure. Eur Respir J 12:
288-293, 1998.
11. Duck-Chong CG. A rapid sensitive method for determining phospholipid
phosphorus involving digestion with magnesium nitrate. Lipids 14: 492-497,
1979.

24
12. Enhorning G. Pulsating bubble technique for evaluating pulmonary surfactant. J
Appl Physiol 43: 198-203, 1977.
13. Galley HF, Howdle PD, Walker BE and Webster NR. The effects of
intravenous antioxidants in patients with septic shock. Free Radic Biol Med 23:
768-774, 1997.
14. Gilliard N, Heldt GP, Loredo J, Gasser H, Redl H, Merritt TA and Spragg
RG. Exposure of the hydrophobic components of porcine lung surfactant to
oxidant stress alters surface tension properties. J Clin Invest 93: 2608-2615, 1994.
15. Haddad IY, Holm BA, Hlavaty L and Matalon S. Dependence of surfactant
function on extracellular pH: mechanisms and modifications. J Appl Physiol 76:
657-662, 1994.
16. Haddad IY, Ischiropoulos H, Holm BA, Beckman JS, Baker JR and Matalon
S. Mechanisms of peroxynitrite- induced injury to pulmonary surfactants. Am J
Physiol 265: L555-L564, 1993.
17. Haddad IY, Pataki G, Hu P, Galliani C, Beckman JS and Matalon S.
Quantitation of nitrotyrosine levels in lung sections of patients and animals with
acute lung injury. J Clin Invest 94: 2407-2413, 1994.

25
18. Hafner D, Germann PG and Hauschke D. Effects of rSP-C surfactant on
oxygenation and histology in a rat- lung- lavage model of acute lung injury. Am J
Respir Crit Care Med 158: 270-278, 1998.
19. Holm BA, Enhorning G and Notter RH. A biophysical mechanism by which
plasma proteins inhibit lung surfactant activity. Chem Phys Lipids 49: 49-55,
1988.
20. Ito Y, Goffin J, Veldhuizen R, Joseph M, Bjarneson D, McCaig L, Yao L-J,
Marcou J and Lewis J. Timing of exogenous surfactant administration in a
rabbit model of acute lung injury. J Appl Physiol 80: 1357-1364, 1996.
21. Jobe AH. Which surfactant for treatment of respiratory-distress syndrome. Lancet
355: 1380-1381, 2000.
22. Lang JD, McArdle PJ, O'Reilly PJ and Matalon S. Oxidant-antioxidant
balance in acute lung injury. Chest 122: 314S-320S, 2002.
23. Laurent T, Markert M, Feihl F, Schaller MD and Perret C. Oxidant-
antioxidant balance in granulocytes during ARDS. Effect of N-acetylcysteine.
Chest 109: 163-166, 1996.
24. Lewis JF and Jobe AH. Surfactant and the adult respiratory distress syndrome.
Am Rev Respir Dis 147: 218-233, 1993.

26
25. Lewis JF and Veldhuizen R. The role of exogenous surfactant in the treatment
of acute lung injury. Annu Rev Physiol 65: 613-642, 2003.
26. Lewis JF and Veldhuizen RAW. Exogenous surfactant adminstration for ARDS.
In: Recent advances in critical care 4, edited by Evans and Hinds. London:
Churchill Livingstone, 1996, p. 45-69.
27. Lowry OH, Rosebrough NJ, Farr AL and Randall RJ. Protein measurement
with the Folin reagent. J Biol Chem 193: 265-275, 1951.
28. Manz Keinke H, Egenhofer C, Plattner H and Schlepper Schafer J. Specific
interaction of lung surfactant protein A (SP-A) with rat alveolar macrophages.
Exp Cell Res 192: 597-603, 1991.
29. Mark L and Ingenito EP. Surfactant function and composition after free radical
exposure generated by transition metals. Am J Physiol 276: L491-L500, 1999.
30. Marzan Y, Mora R, Butler A, Butler M and Ingenito EP. Effects of
simultaneous exposure of surfactant to serum proteins and free radicals. Exp Lung
Res 28: 99-121, 2002.
31. Matalon S, Holm BA, Loewen GM, Baker RR and Notter RH. Sublethal
hyperoxic injury to the alveolar epithelium and the pulmonary surfactant system.
Exp Lung Res 14 Suppl: 1021-1033, 1988.

27
32. Merritt TA, Amirkhanian JD, Helbock H, Halliwell B and Cross CE.
Reduction of the surface-tension- lowering ability of surfactant after exposure to
hypochlorous acid. Biochem J 295 ( Pt 1): 19-22, 1993.
33. Oosting RS, Van Iwaarden JF, Van Bree L, Verhoef J, Van Golde LM and
Haagsman HP. Exposure of surfactant protein A to ozone in vitro and in vivo
impairs its interactions with alveolar cells. Am J Physiol 262: L63-L68, 1992.
34. Possmayer F, Yu SH, Weber JM and Harding PG. Pulmonary surfactant. Can
J Biochem Cell Biol 62: 1121-1133, 1984.
35. Rodriguez-Capote K, McCormack FX and Possmayer F. Pulmonary surfactant
protein-A (SP-A) restores the surface properties of surfactant after oxidation by a
mechanism that requires the Cys6 interchain disulfide bond and the phospholipid
binding domain. J Biol Chem 278 (23): 20461-20474, 2003.
36. Sittipunt C, Steinberg KP, Ruzinski JT, Myles C, Zhu S, Goodman RB,
Hudson LD, Matalon S and Martin TR. Nitric oxide and nitrotyrosine in the
lungs of patients with acute respiratory distress syndrome. Am J Respir Crit Care
Med 163: 503-510, 2001.
37. Tremblay LN and Slutsky AS. Ventilator-induced injury: from barotrauma to
biotrauma. Proc Assoc Am Physicians 110: 482-488, 1998.

28
38. Van Iwaarden JF, Teding van Berkhout F, Whitsett JA, Oosting RS and Van
Golde LM . A novel procedure for the rapid isolation of surfactant protein A with
retention of its alveolar-macrophage-stimulating properties. Biochem J 309: 551-
555, 1995.
39. Veldhuizen RA, McCaig LA, Akino T and Lewis JF. Pulmonary surfactant
subfractions in patients with the acute respiratory distress syndrome. Am J Respir
Crit Care Med 152: 1867-1871, 1995.
40. Wright JR. Immunomodulatory functions of surfactant. Physiol Rev 77: 931-962,
1997.
41. Zhu S, Ware LB, Geiser T, Matthay MA and Matalon S. Increased levels of
nitrate and surfactant protein a nitration in the pulmonary edema fluid of patients
with acute lung injury. Am J Respir Crit Care Med 163: 166-172, 2001.
#pulsations
0 20 40 60 80 100 120
surf
ace
tens
ion
(mN
/m)
0
10
20
30
40
50BLES input OXBLES input
*
*** * * * *
Figure 1

0 5 15 30 60 90 120
PaO
2 (m
mH
g)
0
100
200
300
400
500
TS LA SA
Sur
fact
ant (
mg
PL/
kg)
0
2
4
6
8
0 20 40 60 80 100 120
Sur
face
tens
ion
(mN
/m)
0
5
10
15
20
25
30
A
B
C
Figure 2
* * * * *
pulsations
time (mins)

0 5 15 30 60 90 1200
100
200
300
400
500
PaO
2 (m
mH
g)
TS LA SA0
5
10
15
20
25
0 20 40 60 80 100 1200
5
10
15
20
25
30
35
Sur
fact
ant (
mg
PL/
kg)
Sur
face
tens
ion
(mN
/m)
* *
* * **
A
B
C
Figure 3
pulsations
time (mins)
0 5 15 30 60 90 120
PaO
2 (m
mH
g)
50
100
150
200
250
300
350
400 A
***
*
TS LA SA0
2
4
6
8
10
Sur
fact
ant (
mg
PL/
kg)
0 20 40 60 80 100 1200
10
20
30
40
50
Sur
face
tens
ion
(mN
/m)
B
C
Figure 4
*** * * * * * * *
pulsations
time (mins)
Copyright © 2022 FDOKUMEN




















