
Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador
8
Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador) Donatella Mares Elisa Andreotti Maria Elena Maldonado Paola Pedrini Chiara Colalongo Carlo Romagnoli Received: 13 March 2008 / Accepted: 20 April 2008 / Published online: 2 July 2008 Ó Springer Science+Business Media, LLC 2008 Abstract From an undisturbed natural forest soil in Ecuador, three fungal strains of the genus Aspergillus were isolated. Based on molecular and morphological features they are described as three new species, named A. quiten- sis, A. amazonicus, and A. ecuadorensis. Introduction As part of a collaborative project with the University of Quito (Ecuador) to find new microorganisms from undisturbed natural places as possible sources of pharmacologically active compounds, we isolated a few fungal strains from soil samples from the Amazonian forest (Ecuador). The Ama- zonian forest was chosen because it is a place with little anthropogenic influence, where, to date, there is a consid- erable biodiversity of animals, plants, and microorganisms. The first step in this work was to study whether the microorganisms were able to perform Baeyer-Villinger (BV) oxidations. Three fungal isolates were able to induce the oxidation of cyclic ketones to corresponding lactones [2]. These compounds are interesting, since they are starting products for the synthesis of several pharmaceuti- cally active compounds, such as prostaglandins [7]. In this work we studied in detail these three fungal isolates and performed morphological observations and molecular investigations to determine their systematic position. Materials and Methods Microorganisms The three fungi most interesting for their enzymatic prop- erties were isolated in the year 2004 from a soil collected in the Amazonian forest, near the town of Makas (Ecuador). The fungi were arbitrarily assigned the following alpha- numerical codes: E 19 C, E 19 D, and E 19 F. The isolated microorganisms were maintained in vials containing Sab- ouraud dextrose broth (SD; Difco, Detroit, MI) and 20% glycerol in liquid nitrogen at -190°C. Morphological observations were carried out on fungi grown for 20 days on the same solidified culture medium at 26° ± 2°C. Morphological Analyses Macroscopic colony characters (color of the aerial part and of the reverse mycelia, texture, topography, and edges of colonies) and microscopic morphological characteristics were recorded and illustrated using a Leika stereomicro- scope and a Cambridge Stereoscan 360 scanning electron D. Mares P. Pedrini Dipartimento di Biologia ed Evoluzione, Universita ` di Ferrara, C. so Ercole I d’Este, 32, I-44100 Ferrara, Italy E. Andreotti C. Romagnoli (&) Dipartimento del Museo di Paleobiologia e dell’Orto Botanico, Universita ` di Modena e Reggio E, v. le Caduti in Guerra 127, I-41100 Modena, Italy e-mail: [email protected] M. E. Maldonado Centro de Investigacio ´n y Valoracio ´n de la Biodiversidad, Universidad Polite ´cnica Salesiana (CIVABI-UPS), Avenida 12 de Octubre N 24-22 y Wilson, Casilla Postal 17 12 536, Quito, Ecuador C. Colalongo Dipartimento di Scienze e Tecnologie Agroambientali, University of Bologna, Via Fanin 44, 40127 Bologna, Italy 123 Curr Microbiol (2008) 57:222–229 DOI 10.1007/s00284-008-9178-9
-
Upload
juanlapeyre -
Category
Documents
-
view
1 -
download
0
Transcript of Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador

Three New Species of Aspergillus from Amazonian Forest Soil(Ecuador)
Donatella Mares Æ Elisa Andreotti Æ Maria Elena Maldonado ÆPaola Pedrini Æ Chiara Colalongo Æ Carlo Romagnoli
Received: 13 March 2008 / Accepted: 20 April 2008 / Published online: 2 July 2008
� Springer Science+Business Media, LLC 2008
Abstract From an undisturbed natural forest soil in
Ecuador, three fungal strains of the genus Aspergillus were
isolated. Based on molecular and morphological features
they are described as three new species, named A. quiten-
sis, A. amazonicus, and A. ecuadorensis.
Introduction
As part of a collaborative project with the University of Quito
(Ecuador) to find new microorganisms from undisturbed
natural places as possible sources of pharmacologically
active compounds, we isolated a few fungal strains from soil
samples from the Amazonian forest (Ecuador). The Ama-
zonian forest was chosen because it is a place with little
anthropogenic influence, where, to date, there is a consid-
erable biodiversity of animals, plants, and microorganisms.
The first step in this work was to study whether the
microorganisms were able to perform Baeyer-Villinger
(BV) oxidations. Three fungal isolates were able to induce
the oxidation of cyclic ketones to corresponding lactones
[2]. These compounds are interesting, since they are
starting products for the synthesis of several pharmaceuti-
cally active compounds, such as prostaglandins [7]. In this
work we studied in detail these three fungal isolates and
performed morphological observations and molecular
investigations to determine their systematic position.
Materials and Methods
Microorganisms
The three fungi most interesting for their enzymatic prop-
erties were isolated in the year 2004 from a soil collected in
the Amazonian forest, near the town of Makas (Ecuador).
The fungi were arbitrarily assigned the following alpha-
numerical codes: E 19 C, E 19 D, and E 19 F. The isolated
microorganisms were maintained in vials containing Sab-
ouraud dextrose broth (SD; Difco, Detroit, MI) and 20%
glycerol in liquid nitrogen at -190�C. Morphological
observations were carried out on fungi grown for 20 days
on the same solidified culture medium at 26� ± 2�C.
Morphological Analyses
Macroscopic colony characters (color of the aerial part and
of the reverse mycelia, texture, topography, and edges of
colonies) and microscopic morphological characteristics
were recorded and illustrated using a Leika stereomicro-
scope and a Cambridge Stereoscan 360 scanning electron
D. Mares � P. Pedrini
Dipartimento di Biologia ed Evoluzione, Universita di Ferrara,
C. so Ercole I d’Este, 32, I-44100 Ferrara, Italy
E. Andreotti � C. Romagnoli (&)
Dipartimento del Museo di Paleobiologia e dell’Orto Botanico,
Universita di Modena e Reggio E, v. le Caduti in Guerra 127,
I-41100 Modena, Italy
e-mail: [email protected]
M. E. Maldonado
Centro de Investigacion y Valoracion de la Biodiversidad,
Universidad Politecnica Salesiana (CIVABI-UPS), Avenida 12
de Octubre N 24-22 y Wilson, Casilla Postal 17 12 536,
Quito, Ecuador
C. Colalongo
Dipartimento di Scienze e Tecnologie Agroambientali,
University of Bologna, Via Fanin 44, 40127 Bologna, Italy
123
Curr Microbiol (2008) 57:222–229
DOI 10.1007/s00284-008-9178-9

microscope (Cambridge, UK), owned by the Electron
Microscopic Centre of the University of Ferrara, Italy.
Munsell color charts were used for description of the
colors [6]. Living cultures are deposited at the culture
collection of the Department of Biology and Evolution
(Ferrara University, Italy). Ultrastructural observations
were made by harvesting small samples from a 20-day
cultures of fungi grown on SDA. The samples were pro-
cessed using methods described previously [5].
Isolation and Analysis of Nucleic Acids
Nucleic acids were extracted as described by Griffin et al.
[3] with slight modifications. Briefly, about 50 mg of
fungal mycelia from each isolate was placed in a sterile
1.5-ml microcentrifuge tube. To each tube 400 ll of AP1
buffer (DNeasy Plant Mini Kit; Qiagen) was added and
freeze/thaw was used to lyse fungal cells. Freeze/thaw was
repeated five times using liquid nitrogen and a boiling-
water bath. After the last freezing step samples were boiled
for 30 min with DNA from plant tissue using the DNeasy
Plant Mini Kit. Fungal DNA was eluted in 50 ll AE buffer
and stored at 4�C until use. PCR amplification of the ITS1
and ITS2 regions was performed with universal fungal
primers ITS1 (50-TCCGTAGGTGAACCTGCGG-30) and
ITS4 (50-TCCTCCGCTTATTGATATG-30), as described
by White et al. [12]. The full ITS region was amplified by
PCR in a final volume of 50 ll of the following reaction
mixture: *100 ng template DNA; 0.3 lM (each) forward
(ITS1) and reverse (ITS4) primers; 100 lM (each) dATP,
dCTP, dGTP, and dTTP; 1 9 recombinant Taq DNA
polymerase reaction buffer; 2 mM MgCl2 and 2 U of
recombinant Taq DNA polymerase (Invitrogen Life
Technologies, Carlsbad, CA, USA). Samples were over-
layed with a drop of mineral oil and amplified in a Cetus
Perkin Elmer PCR machine. An initial denaturation step
(94�C for 4 min) was followed by 40 cycles, each con-
sisting of DNA denaturation at 94�C for 30 s, primer
annealing at 50�C for 30 s, and elongation at 72�C for
1 min. The last step was a final extension at 72�C for
10 min. PCR products of *600 bp were visualized by
1.2% agarose gel electrophoresis and ethidium bromide
staining. The amplified ITS regions were then purified from
primers and salts using Nucleospin Extract II spin columns
(Machery-Naghel). Direct sequencing of the fragments was
performed by MWG Biotech using Big Dye terminator
reaction chemistry. Sequences were determined from both
strands using ITS1 and ITS4 as primers.
Data Analysis
Alignment of the three sequences obtained was performed
by ClustalW software (http://www.ebi.ac.uk/clustalw/).
Search comparisons were also made between the three
fungal sequences and other ITS regions available in the
GenBank database using MEGABLAST for sequence
identification.
Results
Our research focused on the three samples of isolated fungi
showing the most interesting enzymatic properties, as
detailed in previously published work [2]. The three iso-
lated fungi were macroscopically very similar. However,
they appeared to be different enzymatically. To verify the
effective diversity among the three isolates, macroscopic
and microscopic analyses were carried out.
On SDA medium all colonies grew slowly, reaching
diameters of from 1.5 to 3.0 cm in 2 weeks at 26� ± 2�C.
SEM observations were made on the 20th day from the
inoculum, when the colonies reached a diameter of 4 cm.
All three fungi showed mycelia flat in topography, granular
in texture, and brown in color, with small floccose and
white areas (Fig. 1). The brown pigmentation of the
mycelia varied lightly among the three strains: in E19C the
surface was blackish-red, in E19D the same color was more
dusky-brown, whereas in E19F the colony showed a
moderate reddish-brown color. Reverse of the cultures
showed a reddish-brown color. Vegetative mycelia were
largely submerged, and surface growth consisted of bran-
ched hyphae with many conidiophores born from an aerial
mycelium (Fig. 2).
All conidiophores had a swollen tip (vesicle) located at
the apex of the stalk, on which conidiogenous cells
Fig. 1 Brown pigmentation of the three Aspergillus mycelia, with
small floccose and white areas
D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador) 223
123

(phialides) and conidia were mounted; this type of conid-
iophore is characteristic of Aspergillales. In all three
isolated strains the conidial heads were globular, radiate,
and typically brown in color. They formed at the end of a
long, straight, colorless, and lightly wrinkled stalk (Fig. 3).
In fact the conidiophore hypha enlarged at its apex to form
a globose structure, known as a vesicle; in the three iso-
lated fungi the vesicles were fertile over the whole surface
(Fig. 4) and were characterized by uni- or biseriate phia-
lides bearing yellow-brown or brown conidia. The
differences among the three fungi in relation to these
characters are reported in Table 1.
In E19C the number of conidiophores, seen at various
development stages, was quite high (Fig. 5). They were
hyaline and 250 ± 5 lm long, and in the upper part they
widened to a cinnamon-brown vesicles, close-woven,
measuring about 50 lm in diameter, with uniseriate phia-
lides over the whole surface (Fig. 6). From phialides, small
dusky-brown conidia were born; they were round, mea-
sured 2.0 ± 0.4 lm in diameter, and were arranged in long
chains (Figs. 7 and 8).
In E19D fungus, the conidiophores were less abundant
in comparison to E19C, but they were longer (about
320 ± 10 lm) and came off with dusky-brown, almost-
black vesicles with tightly adherent conidia, that were tied
to truncated phialides (Fig. 9). The head averaged
48 ± 2 lm and showed only one or two layers of adherent
conidia. These were slightly lemon-shaped and measured
1.9 ± 0.2 9 3.0 ± 0.4 lm, with a kind of spur on both
sides (Fig. 10).
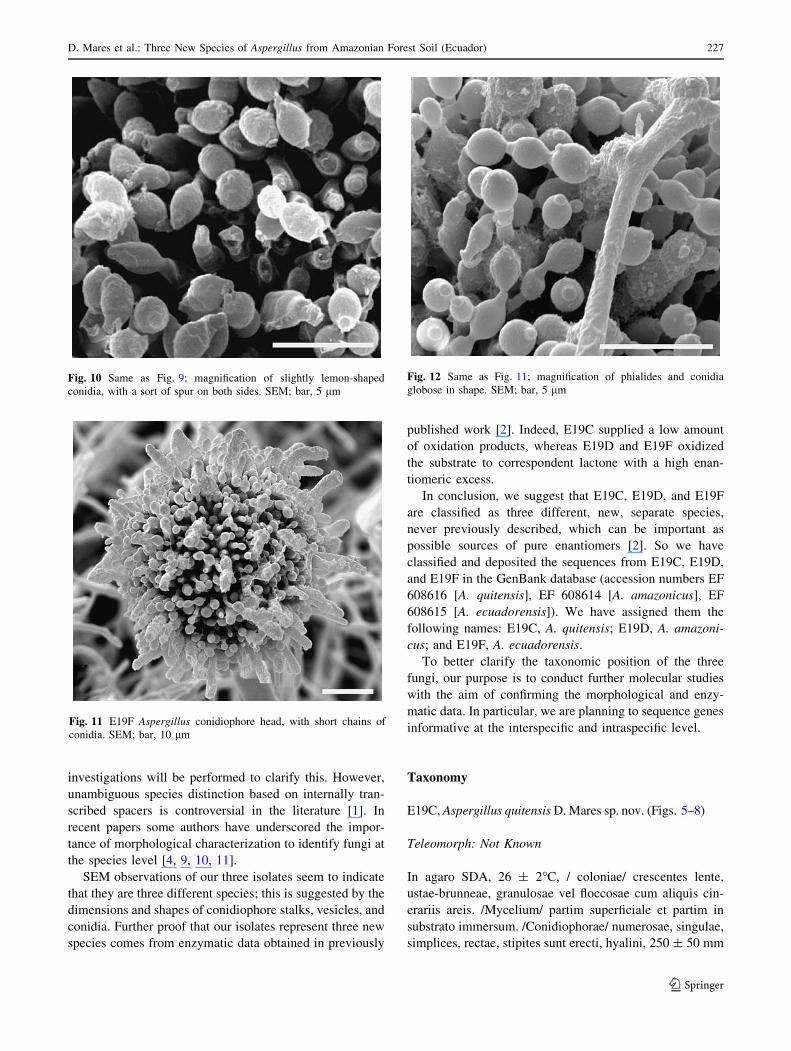
Even E19F had conidiophores with a brown head, from
which many conidia departed, forming short chains
adherent to the head (Fig. 11). Conidiophore stalk length
and vesicle diameter were even smaller: about 200 ± 4 and
36 ± 6 lm, respectively. Conidia were globose in shape
and had an average diameter of 1.65 ± 0.5 lm (Fig. 12).
For molecular identification of our three isolates (E19C,
E19D, E19F) the ITS 1-5.8S-ITS 2 regions were sequenced
using primers ITS 1 and ITS 4. This yielded an identical
sequence of 538 bp for all three strains. The best BLAST
hits of the three identical sequences, obtained after a search
in the GenBank database, were ITS sequences from a few
species of Emericella (E. falconensis, AB249004; E. sim-
ilis, AB248996; E. violacea, AB248985; E. fruticulosa,Fig. 2 Overview of vegetative mycelia showing many conidio-
phores. SEM; bar, 50 lm
Fig. 3 Detail of a conidiophore with globular and radiate conidial
heads, formed at the end of a long, straight, and wrinkled stalk. SEM;
bar, 20 lm
224 D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador)
123

AB248969) and Aspergillus versicolor (AY373882) with
88% sequence identity.
Nucleotide Sequence Accession Numbers
The sequences of the ITS regions of the three fungi
described in this study were deposited in the GenBank
database and assigned accession numbers EF 608616 (A.
quitensis; E19C), EF 608614 (A. amazonicus; E19D), and
EF 608615 (A. ecuadorensis; E19F).
Discussion
The three fungi selected from the Amazonian forest soil
and tested for their BV activity showed different enzymatic
activity [2], although their macroscopical morphology was
very similar. Our morphological observations indicate that
they belong to the Aspergillus group for the presence of
characteristic conidiophores. Since relatively few aspergilli
produce cleistothecia and ascospores [8], we chose to study
other characteristics of the colonies as a basis for their
identification: we used scanning electron microscopy to
characterize morphologically spore-bearing structures and
employed molecular methods by sequencing the ITS1-
5.8S-ITS2 region of rDNA [9].
Using the identification keys of Rapel and Fennell [8],
the distinctive morphological characters indicate that all
three isolated samples are members of the Aspergillus
wentii group. A. wentii Wehmer is a cosmopolitan fungus
that lives all over the world in the soil and on a great
variety of organic substrates. Nevertheless, in A. wentii the
Fig. 4 Detail of a vesicle fertile over its whole surface. SEM; bar,
5 lm
Table 1 Characteristics of
conidiophore branches, vesicles,
phialides, and conidia of the
three isolated fungi
Note: Measurements are the
average of 50 SEM
determinations ± standard error
E19C E19D E19F
Conidiophores
Stalks
Length (lm) 250 ± 5 320 ± 10 200 ± 4
Color Colorless Colorless Colorless
Vesicles
Diameter (lm) 50 ± 3 48 ± 2 36 ± 6
Color Cinnamon-brown Dusky-brown/black Brown
Conidia
Arrangement Long chains 1 or 2 layers Short chains
Shape Round Oval Globular
Color Dusky-brown Dusky-brown/black Brown
Measurements (lm) 2 ± 0.4 1.9 ± 0.2 9 3.0 ± 0.4 1.65 ± 0.5
Phialides Uniseriate Biseriate Uniseriate
Fig. 5 Several conidiophores in the E19C strain. SEM; bar, 100 lm
D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador) 225
123

conidiophore stalks (that may reach several millimeters in
length) and the heads (diameter of C500 lm [8] are much
bigger than those observed in our samples. Thus, we think
that our isolated fungi are not A. wentii strain, but they
could be another species belonging to the same group.
These morphological data were confirmed by BLAST
analyses. We found that the ITS regions of our three fungi
were at least 12% different from other fungal ITS
sequences present in the GeneBank database. Samson et al.
[11] suggest ITS sequence comparison as the molecular
marker of choice for species recognition. Indeed, ITS
sequence comparison has been widely used and many
sequences are now available in public databases. Balajee
et al. [1] indicate that the ITS region is the best tool for
intersection-level classification of Aspergillus. Because of
the great sequence diversity we observed with other spe-
cies, we can confidently conclude that our three fungi have
not been described previously by molecular data. The
closest sequence similarity we found was with a limited
number of Emericella sp. and Aspergillus sp., as shown
under Results.
As ITS regions 1 and 2 yielded an identical sequence for
all three strains, we were not able to distinguish the three
fungal strains by the ITS method, and further molecular
Fig. 6 Uniseriate phialides adherent to the surface of E19C conid-
iophores. SEM; bar, 10 lm
Fig. 7 Conidiophore head of E19C with many round conidia. SEM;
bar, 20 lm
Fig. 8 Detail of an E19C conidial head showing conidia arranged in
long chains. SEM; bar, 10 lm
Fig. 9 E19D conidiophore showing tightly adherent conidia, tied to
truncated phialides. SEM; bar, 10 lm
226 D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador)
123
investigations will be performed to clarify this. However,
unambiguous species distinction based on internally tran-
scribed spacers is controversial in the literature [1]. In
recent papers some authors have underscored the impor-
tance of morphological characterization to identify fungi at
the species level [4, 9, 10, 11].
SEM observations of our three isolates seem to indicate
that they are three different species; this is suggested by the
dimensions and shapes of conidiophore stalks, vesicles, and
conidia. Further proof that our isolates represent three new
species comes from enzymatic data obtained in previously
published work [2]. Indeed, E19C supplied a low amount
of oxidation products, whereas E19D and E19F oxidized
the substrate to correspondent lactone with a high enan-
tiomeric excess.
In conclusion, we suggest that E19C, E19D, and E19F
are classified as three different, new, separate species,
never previously described, which can be important as
possible sources of pure enantiomers [2]. So we have
classified and deposited the sequences from E19C, E19D,
and E19F in the GenBank database (accession numbers EF
608616 [A. quitensis], EF 608614 [A. amazonicus], EF
608615 [A. ecuadorensis]). We have assigned them the
following names: E19C, A. quitensis; E19D, A. amazoni-
cus; and E19F, A. ecuadorensis.
To better clarify the taxonomic position of the three
fungi, our purpose is to conduct further molecular studies
with the aim of confirming the morphological and enzy-
matic data. In particular, we are planning to sequence genes
informative at the interspecific and intraspecific level.
Taxonomy
E19C, Aspergillus quitensis D. Mares sp. nov. (Figs. 5–8)
Teleomorph: Not Known
In agaro SDA, 26 ± 2�C, / coloniae/ crescentes lente,
ustae-brunneae, granulosae vel floccosae cum aliquis cin-
erariis areis. /Mycelium/ partim superficiale et partim in
substrato immersum. /Conidiophorae/ numerosae, singulae,
simplices, rectae, stipites sunt erecti, hyalini, 250 ± 50 mm
Fig. 10 Same as Fig. 9; magnification of slightly lemon-shaped
conidia, with a sort of spur on both sides. SEM; bar, 5 lm
Fig. 11 E19F Aspergillus conidiophore head, with short chains of
conidia. SEM; bar, 10 lm
Fig. 12 Same as Fig. 11; magnification of phialides and conidia
globose in shape. SEM; bar, 5 lm
D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador) 227
123

longi et vesicae sunt globosae, 50 ± 2 mm latae, fertiles in
tota esteriore parte, cinnamum-brunneae. /Phialides/ uni-
verticillati. /Conidia/ globosa 2.0 ± 0,4 mm, atro-brunnea,
sicut longae catenae vinculate.
Anamorph: In SDA, at 26� ± 2�C, slow-growing colo-
nies, flat, blackish-red, granular in texture except in
somewhat floccose gray areas; exudate lacking; reverse
dusky-brown. Mycelium superficial and immersed in the
substratum, composed of septate, branched, and smooth
hyphae. Conidiophores abundant, single, unbranched,
straight, with erect and hyaline stalks, 250 ± 5 lm long.
Vesicles globose, cinnamon-brown, 50 ± 2 lm in diame-
ter. Phialides uniseriate over the whole surface. Conidia
small, round, 2.0 ± 0.4 lm in diameter, dusky-brown, and
arranged in long chains.
Specimen examined: Ecuador, Makas, from soil, 25
April 2004.
Etymology: The specific epithet quitensis refers to
Quito, the capital of Ecuador.
E19D, Aspergillus amazonicus D. Mares sp. nov.
(Figs. 9 and 10)
Teleomorph: Not Known
In agaro SDA, 26 ± 2�C, / coloniae/ crescentes lente,
floccosae atro-brunneae. /Mycelium/ partim superficiale et
partim in substrato immersum. /Conidiophorae/ singulae,
simplices, rectae, stipites sunt erecti, hyalini, 320 ± 10 mm
longi et vesicae sunt globosae, 48 ± 2 mm latae, fertiles in
tota esteriore parte, atro-brunneae vel atrae. /Phialides/
biverticillati et trunci. /Conidia/ ovoidea vel ellipsoidea 1.9
± 0, 2 x 3, 0 ± 0, 4 mm, atro-brunnea vel atra, portata in 1
vel 2 strata adhaerentia exteriori parti vesicae.
Anamorph: In SDA, at 26� ± 2�C, slow-growing colo-
nies, flat, floccose, dusky-brown; exudate clear; reverse
dusky-brown. Mycelium superficial and immersed in the
substratum, composed of septate, branched, and smooth
hyphae. Conidiophores single, unbranched, straight, with
erect and colorless stalks, 320 ± 10 lm long. Vesicles
globose and fertile over the entire surface, almost black,
48 ± 2 lm in diameter. Phialides biseriate and truncated.
Conidia oval or ellipsoidal, 1.9 ± 0.2 9 3.0 ± 0.4 lm,
almost black, tightly adherent to vesicle surface, forming
one or two closely packed layers.
Specimen examined: Ecuador, Makas, from soil, 25
April 2004.
Etymology: The specific epithet amazonicus refers to the
place of origin.
E19F, Aspergillus ecuadorensis D. Mares sp. nov.
(Figs. 11 and 12)
Teleomorph: Not Known
In agaro SDA, 26 ± 2�C, / coloniae/ crescentes lente, floc-
cosae, brunneae. Albi umores exudant. /Mycelium/ partim
superficiale et partim in substrato immersum. /Conidiopho-
rae/ singulae, simplices, rectae, stipites sunt erecti, hyalini,
200 ± 4 mm longi et vesicae sunt globosae, 36 ± 6 mm
latae, fertiles in tota exteriore parte, atro-brunneae vel atrae.
/Phialides/ univerticillati, et ampulliformes. /Conidia/ glob-
osa 1.65 ± 0.5 mm, atro-brunnea vel atra, sicut catenae
vinculata et adhaerentia exteriori parti vesicae.
Anamorph: In SDA, at 26o ± 2�C, slow-growing colo-
nies, flat, floccose, moderate reddish-brown; exudate clear;
reverse dusky-brown. Mycelium superficial and immersed in
the substratum, composed of septate, branched, and smooth
hyphae. Conidiophores single, unbranched, straight, with
erect and colorless stalks, 200 ± 4 lm long. Vesicles glo-
bose and fertile over the entire surface, brown, 36 ± 6 lm in
diameter. Phialides uniseriate. Conidia globular, 1.65 ±
0.5 lm, brown, arranged in short chains and adherent to the
vesicle surface.
Specimen examined: Ecuador, Makas, from soil, 25
April 2004.
Etymology: The specific epithet ecuadorensis refers to
the nation of origin.
Acknowledgments The authors wish to thank Prof. Marisa Ferra-
resi, Dr. Mirco Iotti, and the staff of the Electron Microscopy Center
at Ferrara University for their skillful assistance. This work was
supported by research grants from MIUR-URST (Ministero dell’Ist-
ruzione, dell’Universita e della Ricerca-Area Universita e Ricerca
Scientifica e Tecnologica) of Italy.
References
1. Balajee SA, Houbraken J, Verweij PE, Hong S-B, Yaghuchi T,
Varga J, Samson RA (2007). Aspergillus species identification in
the clinical setting. Stud Mycol 59:39–46
2. Fantin F, Giovannini PP, Guerrini A, Maietti S, Medici A, Pedrini
P (2006) Enantioselective Baeyer-Villiger oxidation of bicy-
cle[3.2.0]hept-2-en-6-one with fungi: optimization of biotransfor-
mation and use of TiO2 as support of cell growth. Biotechnol Lett
28:805–810
3. Griffin DW, Kellogg CA, Peak KK, Shinn EA (2002) A rapid and
efficient assay for extracting DNA from fungi. Lett Appl
Microbiol 34:210–214
4. Hawksworth D, Kirk P, Sutton B, Pegler D (1996) Ainsworth’s and
Bisby’s dictionary of the fungi, 8th edn. CABI, Wallingford, UK
5. Mares D, Romagnoli C, Benvegnu R et al (2002). Mannan-
mediated antifungal activity on Epidermophyton floccosum by 3-
methyl-5-aminoisoxazole-4-thiocyanate, a new azole derivative.
Fung Gen Biol 36:47–57
228 D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador)
123

6. Munsell (1990) Munsell soil color charts. MacBeth Division,
Kollmorgen Instruments Corp. Baltimore, MD
7. Newton RF (1982) In: Roberts SM, Scheimann F (eds) New
synthetic routes to prostaglandins and thromboxanes. Academic
Press, New York, p 61
8. Raper KB, Fennell DI (1965). The genus Aspergillus. Williams
and Wilkins, Baltimore, MD
9. Samson RA, Hong SB, Frisvad JC (2006) Old and new concepts
of species differentiation in Aspergillus. Med Mycol 44:S133–
S148
10. Samson RA, Varga J, Witiak SM, Geiser DM (2007) The species
concept in Aspergillus: recommendations of an international
panel. Stud Mycol 59:71–73
11. Taylor JW, Jacobson DJ, Kroken S Kasuga et al (2000) Phylo-
genetic species recognition and species concepts in fungi. Fung
Gen Biol 31:21–32
12. White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and
direct sequencing of fungal ribosomal RNA genes for phyloge-
netics. In: Innis M, Gelfand D, Sninsky J, White T (eds) PCR
protocols: a guide to methods and application. Academic Press,
San Diego, CA, pp 315–322
D. Mares et al.: Three New Species of Aspergillus from Amazonian Forest Soil (Ecuador) 229
123




















