
Detection of Human Cytomegalovirus in Different Histopathological Types of Glioma in Iraqi Patients

ences 267 (2008) 76–85www.elsevier.com/locate/jns
Journal of the Neurological Sci
Therapeutic effects of combined treatment with ribavirin and tiazofurinon experimental autoimmune encephalomyelitis development:
Clinical and histopathological evaluation
Danijela Stojkov a,⁎, Irena Lavrnja a, Sanja Pekovic a, Sanja Dacic b, Ivana Bjelobaba a,Marija Mostarica-Stojkovic c, Stanislava Stosic-Grujicic d, Sasa Jovanovic a,
Nadezda Nedeljkovic b, Ljubisav Rakic a, Mirjana Stojiljkovic a
a Department of Neurobiology, Institute for Biological Research “Sinisa Stankovic”, Bulevar Despota Stefana 142, Belgrade, 11000, Serbiab Institute of Physiology and Biochemistry, School of Biology, Studentski trg 3, Belgrade, 11000, Serbiac Institute of Microbiology and Immunology, School of Medicine, Dr Subotica 8, Belgrade, 11000, Serbia
d Department of Immunology, Institute for Biological Research “Sinisa Stankovic”, Bulevar Despota Stefana 142, Belgrade, 11000, Serbia
Received 8 June 2007; received in revised form 28 September 2007; accepted 4 October 2007Available online 8 November 2007
Abstract
Experimental autoimmune encephalomyelitis (EAE) is an animal model of multiple sclerosis (MS) and the helpful tool in preclinicaltesting of various substances considered for treatment of this human CNS disease. Ribavirin (R) and tiazofurin (T) are purine nucleosideanalogues, with the broad spectrum of anti-viral, anti-tumoral and anti-inflammatory properties. We proposed that combined treatment withRT, administrated during the effector phase of EAE, would attenuate disease severity, both clinically and pathologically. Ribavirin was givendaily at a dosage of 30 mg/kg and tiazofurin was given at a dosage of 10 mg/kg every other day for 15 days. We detected amelioration ofclinical signs and faster recovery in the RT group compared to the control group. Immunohistochemical analyses revealed that RT treatmentdecrease the number of T cells, macrophages and microglia. In the controls, we detected reactive type of microglia, while in the RT group wenoticed ramified/resting form. Demyelination areas and axonal damage were not recorded in the RT group, in contrast to the control groupwhere multiple areas of demyelination zones and axonal loss were found. RT combination treatment suppresses ongoing EAE, preventsdemyelination and axonal loss, and therefore may well be the potential therapy for the treatment of MS.© 2007 Elsevier B.V. All rights reserved.
Keywords: Experimental autoimmune encephalomyelitis; Multiple sclerosis; Immunosupression; Ribavirin; Tiazofurin; Combination therapy
1. Introduction
Multiple sclerosis (MS) is a chronic demyelinatinginflammatory disease of the human central nervous system(CNS) and the most frequent disabling neurological disorderof young adults. The main hallmarks of this disease areinflammation in the CNS, demyelination and variable degreeof axonal loss. The generally accepted concept of pathogen-esis in both MS and EAE suggests that encephalitogenic
⁎ Corresponding author. Tel.: +381 11 2078340; fax: +381 11 2761433.E-mail address: [email protected] (D. Stojkov).
0022-510X/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jns.2007.10.010
proteins or cross-reactive antigens in the periphery primeautoreactive CD4+ T cells which differentiate into effectorcells and migrate to the CNS. Upon reactivation by targetantigen in the CNS, T cells initiate effector functions throughthe variety of cytokines and chemokines that activate bloodderived macrophages and the resident cells such asmicroglia. These cells produce second wave of proinflam-matory mediators, which lead to the oligodendrocyte, myelinand axon damage [1].
Nevertheless, existing knowledge about EAE andMS doesnot offer satisfactory therapies, either immunomodulatory orneuroprotective. Therefore, during MS despite remissions and

Table 1Influence of therapeutic treatment with ribavirin and tiazofurin on extent ofactively induced EAEaParameters of disease C b RTc
No. of animals 15 19Mean days to onset 9.2±0.4 9.8±0.4Duration of paralysis 3.1±0.4 0.6±0.2⁎⁎
Mean maximum severity score 3.6±0.2 2.5±0.3⁎
Mortality rate 4/15 (27%) 2/19 (11%)
a Parameters of disease were evaluated as described in the Materials andmethods. Data are presented as mean±SEM.b C group was immunized and given 0.2 ml saline as a daily i.p. injection
from the onset of first clinical signs.c RT group was immunized and given combination of ribavirin(R)+
tiazofurin(T) at a dosages 30 mg/kg/day+10 mg/kg/every other day,respectively, from EAE onset to 15 following days. ⁎Pb0.05.; ⁎⁎Pb0.005.
77D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85
medical treatment, there is irreversible axonal damage leadingto cumulative disability and invalidity [2,3]. Current treat-ments include drugs such as interferon-β, glatiramer acetate,mitoxantrone, and natalizumab. Although these drugs alter thenatural course of disease, their long-term impact on clinicallyrelevant disability progression is uncertain. Therefore, manytreatment protocols, which include new agents or combinationtherapies, are currently under the trial. In the past few years inthe special focus are the inhibitors of the enzymes catalyzingthe processes involved in the metabolism of purine andpyrimidine nucleosides and nucleotides [4]. Inosine-mono-phosphate-dehydrogenase (IMPDH) is a rate-limiting enzymein de novo purine synthesis pathway [5]. It was shown thatIMPDH inhibitors induce anti-viral, anti-tumor and immuno-suppresant effect [6–8] and some of them are approved asclinical drugs. Synthetic nucleoside analogues, ribavirin (1-β-D-ribofuranosyl-1,2,4-triazole-3-carboxamide) and tiazofurin(2-β-D-ribofuranosylthiasole-4-carboxamide) inhibit IMPDHvia their active metabolites, ribavirin monophosphate (RMP)and thiazole-4-carboxamide adenine nucleotide (TAD), whichbind on different ligand sites of this enzyme [9]. Inhibition ofIMPDH reduces cellular guanilate concentration and causesinterruption of cell metabolism [10]. The fact that the sameenzyme is inhibited by different pathways offers possibility touse ribavirin and tiazofurin in combination, which providesseveral advantages of combined therapy in comparison tosingle drug therapy, such as potentiation of inhibitory effect onguanilate synthesis, reduction of cell resistance to drugs andthemost important— handlingwith smaller dosages to reducetoxicity. Currently there is no adequate method to revealMS inearly stages, so most of the patients are diagnosed with MSwhen the symptoms start to express and disease is already ineffector phase. For that reason the present study is focused onthe effect of combined ribavirin+tiazofurin (RT) therapyadministrated from the appearance of the first EAE clinicalsign, based on the findings from our previous results [11,12].Since our preliminary study has shown variability in EAEonset, we administrated RT combination at each animalindividually. We carried out clinical and histopathologicalevaluations of EAE to reveal the effect of these drugs cocktail.
2. Materials and methods
2.1. Animals and EAE induction
Male Dark Agouti (DA) rats (12–16 weeks old), wereobtained from the Institute for Biological Research (Belgrade,Serbia). Two to five animals were kept per cage, matched byage and weight (180–260 g). They were housed underconventional conditions with laboratory chow and water adlibitum, and were watered by hand during the period ofparalysis. The rats were immunized by the encephalitogenicemulsion prepared by mixing equal amounts of rat spinal cordhomogenate (SCH, 50% w/v in saline) and complete Freund'sadjuvant containing 0.5 mg/ml Mycobacterium tuberculosis(CFA; ICN Pharmaceuticals Costa Mesa, CA). Each animal
was given 0.1 ml of the SCH–CFA emulsion by a single i.d.injection in the right hind footpad. Immunization wasperformed under anesthesia with ether. Animals were dailymonitored for EAE clinical signs, which were scored from 0 to5 using the following scale: (0) no clinical signs; (1) flaccidtail; (2) hind limb paresis; (3) hind limb paralysis; (4)moribund state, (5) death. Several parameters of the diseasewere examined to evaluate severity of EAE: mean day to onset(the mean day on which affected rats within a group firstdeveloped clinical signs of the disease); mean clinical score(average clinical scores for all rats within a group on a givenday); mean maximal severity score (the mean of the maximalclinical score that each rat in a group developed over the courseof the experiment); duration of paralysis (the mean number ofdays for which the rats had a score of 3 or more). All animalswere observed for 22–27 days after the immunization.Experimental protocols were according to the recommenda-tions given in “Guide for the Care and Use of LaboratoryAnimals” (National Academy Press, Washington, D.C. 1996)and approved by the Local Animal Care Committee and theOffice of Laboratory Animal Welfare (OLAW), NIH, USA.
2.2. Treatment procedure
Ribavirin and tiazofurin were provided by ICN Pharma-ceuticals (Costa Mesa, CA, USA). In order to analyze theirtherapeutic effect on EAE development the immunized ratswere randomly divided at the beginning of experiment in 2groups: control–saline treated group (C) and ribavirin+ tiazofurin treated group (RT). RT group was treated withribavirin+ tiazofurin combination i.p., ribavirin was givendaily at a dosage of 30 mg/kg and tiazofurin was given at adosage of 10 mg/kg every other day. In C-group rats received0.2 ml/kg/day of saline i.p. In all groups treatment wasindividual for each animal and started when the first signs ofEAE appeared (from day 7 to day 12 after immunizations).The treatment lasted for 15 days. After cessation oftreatment, the animals were sacrificed under deep anesthesia,from day 22 to day 27 depending on the disease onset foreach animal.
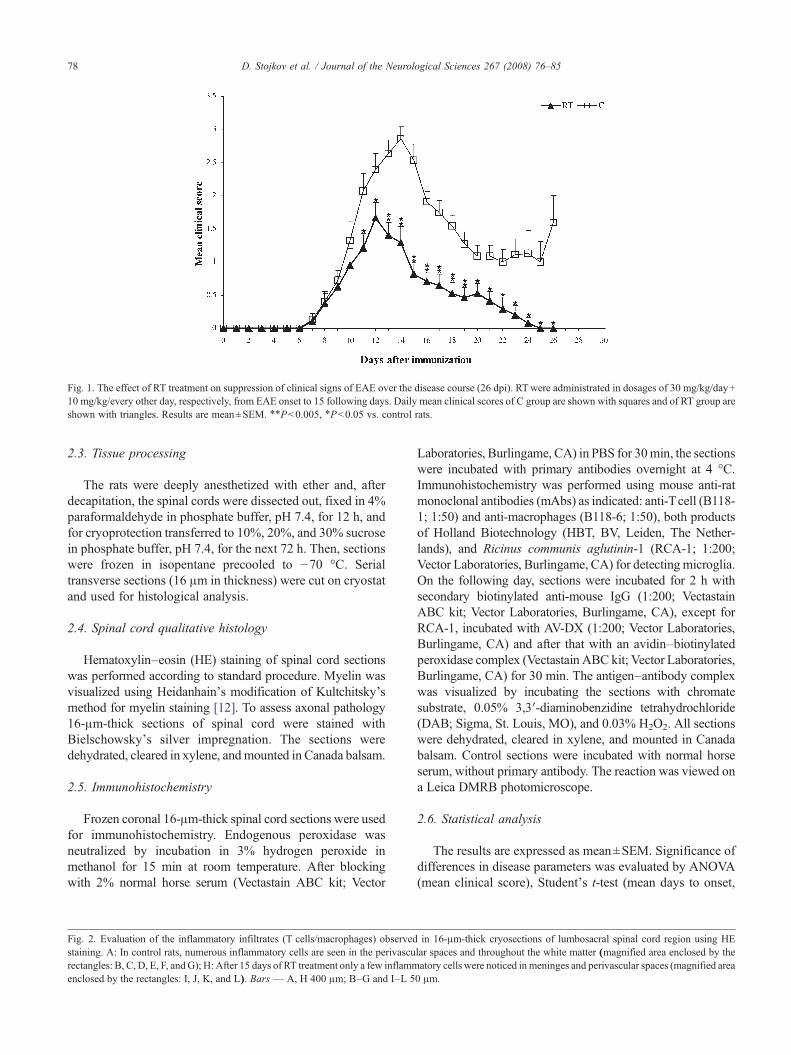
Fig. 1. The effect of RT treatment on suppression of clinical signs of EAE over the disease course (26 dpi). RTwere administrated in dosages of 30 mg/kg/day+10 mg/kg/every other day, respectively, from EAE onset to 15 following days. Daily mean clinical scores of C group are shown with squares and of RT group areshown with triangles. Results are mean±SEM. ⁎⁎Pb0.005, ⁎Pb0.05 vs. control rats.
78 D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85
2.3. Tissue processing
The rats were deeply anesthetized with ether and, afterdecapitation, the spinal cords were dissected out, fixed in 4%paraformaldehyde in phosphate buffer, pH 7.4, for 12 h, andfor cryoprotection transferred to 10%, 20%, and 30% sucrosein phosphate buffer, pH 7.4, for the next 72 h. Then, sectionswere frozen in isopentane precooled to −70 °C. Serialtransverse sections (16 μm in thickness) were cut on cryostatand used for histological analysis.
2.4. Spinal cord qualitative histology
Hematoxylin–eosin (HE) staining of spinal cord sectionswas performed according to standard procedure. Myelin wasvisualized using Heidanhain's modification of Kultchitsky'smethod for myelin staining [12]. To assess axonal pathology16-μm-thick sections of spinal cord were stained withBielschowsky's silver impregnation. The sections weredehydrated, cleared in xylene, andmounted in Canada balsam.
2.5. Immunohistochemistry
Frozen coronal 16-μm-thick spinal cord sections were usedfor immunohistochemistry. Endogenous peroxidase wasneutralized by incubation in 3% hydrogen peroxide inmethanol for 15 min at room temperature. After blockingwith 2% normal horse serum (Vectastain ABC kit; Vector
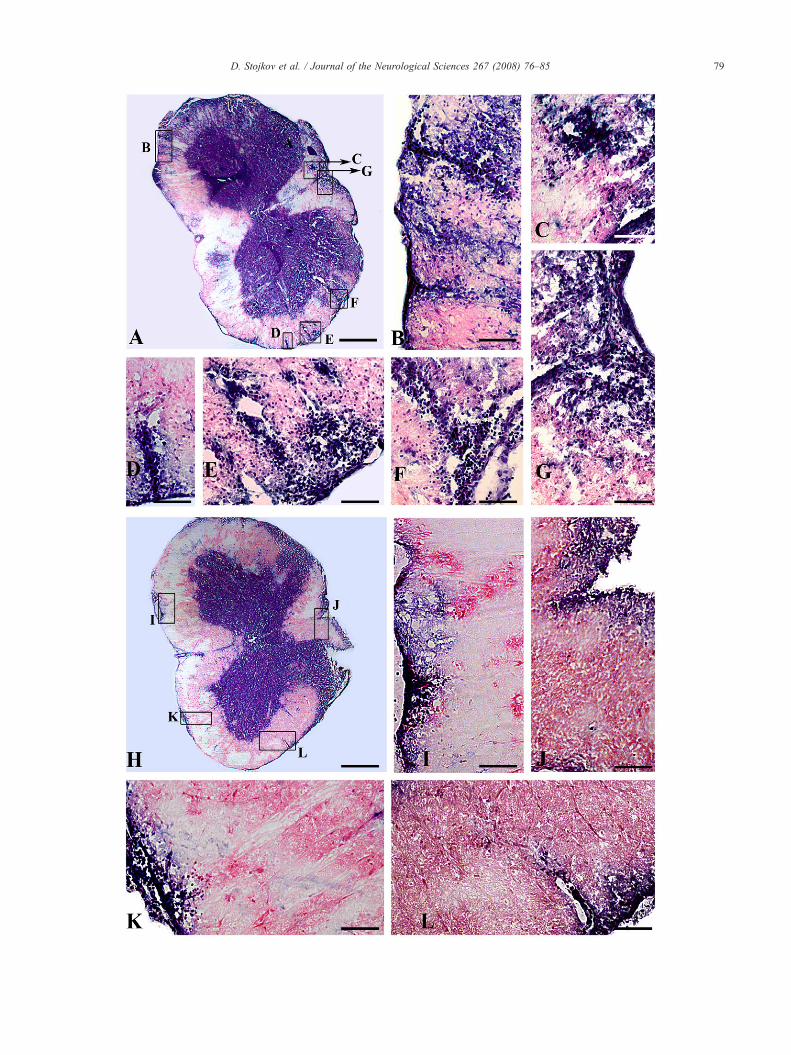
Fig. 2. Evaluation of the inflammatory infiltrates (T cells/macrophages) observedstaining. A: In control rats, numerous inflammatory cells are seen in the perivascurectangles: B, C, D, E, F, andG); H: After 15 days of RT treatment only a few inflammenclosed by the rectangles: I, J, K, and L). Bars — A, H 400 μm; B–G and I–L 5
Laboratories, Burlingame, CA) in PBS for 30min, the sectionswere incubated with primary antibodies overnight at 4 °C.Immunohistochemistry was performed using mouse anti-ratmonoclonal antibodies (mAbs) as indicated: anti-Tcell (B118-1; 1:50) and anti-macrophages (B118-6; 1:50), both productsof Holland Biotechnology (HBT, BV, Leiden, The Nether-lands), and Ricinus communis aglutinin-1 (RCA-1; 1:200;Vector Laboratories, Burlingame, CA) for detectingmicroglia.On the following day, sections were incubated for 2 h withsecondary biotinylated anti-mouse IgG (1:200; VectastainABC kit; Vector Laboratories, Burlingame, CA), except forRCA-1, incubated with AV-DX (1:200; Vector Laboratories,Burlingame, CA) and after that with an avidin–biotinylatedperoxidase complex (Vectastain ABC kit; Vector Laboratories,Burlingame, CA) for 30 min. The antigen–antibody complexwas visualized by incubating the sections with chromatesubstrate, 0.05% 3,3′-diaminobenzidine tetrahydrochloride(DAB; Sigma, St. Louis, MO), and 0.03% H2O2. All sectionswere dehydrated, cleared in xylene, and mounted in Canadabalsam. Control sections were incubated with normal horseserum, without primary antibody. The reaction was viewed ona Leica DMRB photomicroscope.
2.6. Statistical analysis
The results are expressed as mean±SEM. Significance ofdifferences in disease parameters was evaluated by ANOVA(mean clinical score), Student's t-test (mean days to onset,
in 16-μm-thick cryosections of lumbosacral spinal cord region using HElar spaces and throughout the white matter (magnified area enclosed by theatory cells were noticed in meninges and perivascular spaces (magnified area0 μm.
79D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85

Fig. 3. Myelin visualizations and axonal staining (Bielschowsky silver impregnation) of 16-μm-thick cryosections of lumbosacral spinal cord region. Degradedmyelin was present all over white matter in control rats (A: magnified area enclosed by the rectangles: C, D, F and G), whereas after RT treatment myelinremained intact (B, magnified area enclosed by the rectangles: E, H and I). Myelin was visualized using Kultchitsky's method for myelin staining: gray matter(asterisk), unstained (gray); normal myelin, dark blue; degraded myelin, darker than gray matter but lighter than normal myelin. In control rats axonal loss wasobserved all over spinal cord parenchyma (J, black arrows), while in RT treated rats, axonal loss was not detected (K). Bars — A, B, J and K 400 μm; C–I100 μm.
80 D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85

81D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85
mean maximal severity score, duration of paralysis) andFisher's exact probability test (mortality rate). Significantdifferences were accepted at ⁎Pb0.05 and ⁎⁎Pb0.005.
3. Results
3.1. Therapeutic RT treatment during effector phase of EAEsuppresses disease
To examine the therapeutic efficacy of RT treatment on theestablished autoimmune encephalitogenic process, we ap-plied a model of actively induced EAE in highly susceptibleDA rats. As shown previously [13] all animals immunizedwith SCH–CFA developed clinical signs of disease. Themean day of onset in the groups of rats foreseen to serve as acontrol and in those rats, which should be treated, did notdiffer significantly (9.2±0.4 vs. 9.8±0.4, respectively,Table 1). Although, on individual level, we observedvariability in disease onset ranging from day 7 up to day 12post immunization (dpi), causing a difference in the cessationof treatment. In order to insure that RT treatment is reallyinitiated at the beginning of disease, it was appliedindividually, starting in each animal with the appearance ofthe first sign of EAE (flaccid tail) and lasted for 15 days. RTtreatment caused amarkedlymilder EAE as judged by severalparameters of disease severity (Table 1, Fig. 1). Thus, themean maximal severity score was significantly lower in RTtreated group compared to controls (2.5±0.3 vs. 3.6±0.2 incontrol rats, Pb0.05) (Table 1). Duration of paralysis wasalso significantly shorter in treated rats in comparison tocontrols (0.6±0.2 vs. 3.1±0.4 in control rats, Pb0.005)(Table 1). In addition, mortality rate (Table 1) was lower in RTgroup (2/19, 11%) compared to C group (4/15, 27%), but thedifference did not reach the level of statistical significanceaccording to Fisher's exact probability test. The majority of RTtreated animals developedmilder form of EAE in comparison tocontrol rats and completely recovered at the end of observationperiod (Fig. 1). Some of control rats spontaneously started torecover after the peak of EAE, but this incomplete recovery wasfollowed by disease deterioration reaching clinical score 2 at theend of experiment (Fig. 1). In both groups, most of the animalshad acute monophasic course of disease, but in several rats,relapses were noticed. In RT group only 9 of 19 animals passedthrough paralysis, while all animals (15) from C group had thissevere disease symptom. At the end of treatment in controlgroup only 2 animals had a clinical score 0, while all the rest (9)had a clinical score 1, 2 or 3. On the contrary, in RT group allanimals (16), except one, completely recovered from thedisease.
3.2. Therapeutic RT treatment reduces histological lesionsof EAE
Histological analysis were performed on 16-μm-thick sec-tions of spinal cord segments obtained from rats of C group andRT group after 15 days of treatment. At all spinal cord levels,
perivascular and parenchymal mononuclear cells were detected,but we presented only lumbosacral sections, since inflammationand demyelination were most extensive in that region.
3.3. EAE infiltrates
HE staining of lumbosacral spinal cord sections of controlrats demonstrated inflammation characterized by numerousinflammatory cells localized in the meninges, perivascularspaces and throughout the white matter (Fig. 2: A, magnifiedarea enclosed by the rectangles: B, C, D, E, F, and G). Incontrast, after RT treatment, the inflammation was considerablyreduced and restricted to meninges and perivascular spaces,while in parenchyma of white matter it was not seen (Fig. 2: H,magnified area enclosed by the rectangles: I, J, K, and L).
3.4. Demyelination
In control rats, demyelination was widespread throughoutthe white matter columns (Fig. 3: A, magnified area enclosedby the rectangles: C, D, F and G), judged by Kultchitsky'smethod for myelin staining. Demyelination was found inareas of massive mononuclear infiltration (Fig. 2: A). In RT-treated rats, demyelination plaques were absent (Fig. 3: B,magnified area enclosed by the rectangles: E, H, and I).
3.5. Axonal loss
In addition, large regions of axonal loss were observed inC group (Fig. 3: J, arrows), and were located in/or closely todemyelination areas (Fig. 3: A). On the contrary, in RT groupwe did not detect any signs of axonal loss (Fig. 3: K) asestimated by Bielschowsky silver impregnation.
3.6. Response of CNS-infiltrating T cells, macrophages andmicroglia to RT treatment
Immunohistochemical analysis of frozen coronal lumbo-sacral 16-μm-thick spinal cord sections revealed thatmacrophages were scattered all over white matter parenchy-ma in rats of C group (Fig. 4: A, arrows). In RT treatedanimals only a few macrophages in subpial spinal cord areawere detected (Fig. 4: B, magnified area enclosed by therectangle: E).
Similar picture was obtained for T cells. In control groupthey were spread throughout the white matter (Fig. 4: C,magnified area enclosed by the rectangle: D, arrows), whilein RT treated rats immunoreactivity for T cells was notnoticed (Fig. 4: F).
Staining with lectin RCA-1 revealed prominent presenceof microglia cells in ventral columns of white matter in Cgroup (Fig. 4: G) which had macrophage like large roundbody with short and thick extensions (Fig. 4: I, arrows)pointing to reactive type of these cells. In RT treated ratsventral columns were mostly occupied by sparse RCA-1positive cells (Fig. 4: H). Additionally, morphology of these

82 D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85

83D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85
microglial cells was similar to resting form ofmicroglia. Theyhad a smaller body with long and thin extensions (Fig. 4: J,arrows).
4. Discussion
In this study, we analyzed the effects of combined treatmentwith ribavirin (R) and tiazofurin (T) on the established EAE inDA rats. Obtained results showed that combined RT treatmentsuccessfully attenuated severe clinical deficit and suppressedhistopathological events in EAE.
Primary molecular mechanism that underlies the effects ofR and T is the inhibition of IMPDH. Both drugs interact withIMPDH on different ligand sites with higher affinity than thenatural substrates and products [9]. Consequential reductionsof GTP pools are a major outcome of IMPDH inhibition [10].Importantly, this exhaustion of GTP and dGTP pools had amore potent effect on lymphocytes than on the other cell types,since salvage pathways for the production of guanine nucleo-sides do not operate in T- and B cells [14]. Indeed, it is shownthat R-induced inhibition of lymphocyte proliferation iscaused by depletion of guanylate pools [15,16]. In ourprevious study we have reported the beneficial effects of Rin suppressing of EAE development [12]. On the other hand, Thas anti-proliferative effect in various malignancies especiallyin leukemia through inhibition of the DNA/RNA synthesis[17–19]. The efficacy of Tappears to be related to the ability tokill tumor cells, provoke maturation of surviving cells, as wellas to induce apoptosis, and differentiation, inhibition ofglycosilation and down regulation of signal transduction[5,10,20]. Moreover, it has been recently revealed that T,applied continuously after EAE induction, or following theonset of EAE signs, down-regulates disease severity [11].
Structural similarities between R and T, and their knownability to inhibit IMPDH, motivated the evaluation of theircombinations in order to determine whether they would have apotential benefit over the use of a single compound. Studies inthe rat showed that reduced IMPDH activity in bone marrowcells caused by T treatment was kept in a low range, whentherapy was continued byR [21]. Therefore, we have expectedthat these two nucleoside analogues would amplify EAEsuppression compared to the effects gained with each druggiven alone. Indeed, in our recent paper [22] we haveconfirmed that preventive treatment with lower doses of R andT given in combination had a more potent effect in attenuatingclinical and histological signs of EAE than doses of individualdrugs used in Stosic-Grujicic et al. [11] and Milicevic et al.[12] studies. We have noticed a variable disease onset in DA
Fig. 4. Identification of macrophages, T cells and RCA-1 lectin positive cells in sspread throughout the white matter (A, arrows), whereas in RT treated rats onlyenclosed by the rectangle E). Strong immunoreactions for T cells was observed inrectangle: D, arrows), while in RT treated rats immunoreactivity for T cells was cpositively stained microglia, being the most prominent in ventral columns of whitmorphology similar to macrophages (G, magnified area enclosed by the rectangle:microglial cells were noticed (H, magnified area enclosed by the rectangles: J, arr
rats, and therefore in this experiment we administrated RT toeach animal after appearance of the first EAE sign to beassured that effects of RTcombination are certainly therapeuticand did not affect the priming phase of the disease.
Besides considerable attenuation of clinical signs of disease,this paper revealed that 15 days ofRT treatment caused a generalreduction of all infiltrating cell populations. Several lines ofevidences suggest that once inflammatory cells enter CNS, theystart to produce cytokines and chemokines, triggering variouspathological processes including invasion of CNS withsecondary recruited leucocytes [23]. Secondary CNS invasionsare associated with functional neurological deficit of EAE [24].We observed that RT reduced the degree of inflammation in theCNS. The presence of macrophages was noticed only inmeninges and perivascular spaces, but T cells were not detectedat all, while in control group inflammatory cells were massivelyscattered throughout the entire white matter. These findings arein agreement with the fact that most of the control animals at theend of observation still showed disease symptoms, while all RTtreated rats fully recovered. It was shown that DA rats developan acute, monophasic disease without relapses. Nevertheless,we havemonitored RT treated animals for twomore weeks aftercessation of treatment and EAE symptoms did not reappear.
Themain event in generation of aCNS autoimmune diseaseis activation and proliferation of encephalitogenic cells inperipheral lymph nodes [25]. Through IMPDH inhibition, Rand T could exert anti-proliferative effect on lymphocytes,including subset of T cells that are potential encephalitogens.However, the expansion of autoreactive cells is not the onlyfactor influencing disease development. As demonstrated inthe present study, RT suppressed clinical signs of EAE whentheir application started in the effector phase of disease afterthe appearance of the first clinical symptom and therefore, afteractivation and proliferation of T cells. Therefore, the anti-proliferative effect of RT may not be the main mechanism ofgained beneficial outcome of RT treatment observed in thisstudy. These data lead to assumption that another mechanism(s) should be taken into account.
Based on the numerous studies published over the last fewdecades, it is becoming increasingly evident that adhesionmolecules are also involved in the initiation and effector phasesof EAE. It has been shown that T inhibited adhesion of tumorcells to extracellularmatrix [17]. Another study revealed that themononuclear cells from T-treated immunized animals exhibitedreduced ability for intercellular adhesion or adhesion to plastic[11]. In addition, biochemical studies discovered that T inhibitsglycosilation of a wide range of glycoproteins [26]. Since R andTare IMPDH inhibitors, through depletion of intracellular GTP
pinal cord of control and RT treated rats. In control rats macrophages werein subpial zone a paucity of macrophages were noticed (B, magnified areawhite matter parenchyma of control rats (C, magnified area enclosed by theompletely absent (F). In both control (G) and RT group (H) RCA-1 lectine matter. In control rats, there were a large number of microglial cells withI, arrows). Simultaneously, in RT treated rats a reduced number of ramifiedows). Bars — A, D, E, I, J 25 μm; B, F 100 μm; C, G, H 50 μm.

84 D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85
they could limit the transfer of fucose and mannose tomembrane glycoproteins and inhibit the expression of adhesionmolecules on leukocytes, thus provoking immunosuppressiveand anti-inflammatory activity [14]. Histological analysis in thisstudy confirmed that the late administration of RT combinationlimited the mononuclear cell infiltration. It can be assumed thatR and T could reduce the encephalitogenic potential of alreadyactivated lymphocytes by blocking the interactions betweencomplementary adhesion molecules. Consequently, they willpossibly prevent the recruitment of activated immune cells to thetarget tissue and obstruct second T-cells infiltration, which haseffects that are more deleterious.
Histological analysis obtained in this study has shown thatmicroglia in RT-treated animals' resembled resting form ofthese cells. In contrast, in control animals numerousmacrophage-looking microglia cells were observed, pointingto reactive microglia form.Microglial cells are fine sensors ofvarious pathological events in the brain and are rapidlyactivated and recruited to the injured sites. They are involvedin key events of MS and EAE development [27]. Activatedmicroglia acquires functional and morphological features ofmacrophages. During inflammatory or autoimmune CNSdiseases, microglia could exert harmful or protective effectsbecause of the ability to produce pro- and anti-inflammatorycytokines [27,28]. Evidence obtained from in vitro and invivo studies also suggests that microglia may act as competentpresenters of antigen, inducing and regulating the intensity ofT-cell mediated inflammation and tissue injury [27]. Ingeneral, morphological and cellular identity of microglia alsoreflects state of CNS environment [29–31]. Thus, predom-inance of resting type of microglia that we have detected inRT-treated animals pointed out to stable CNS environment incontrast to control animals where predominance of activatedmicroglia reflects unbalanced CNS surrounding. The influ-ence of RT on microglia is probably indirect resulting fromsuppression of T cell and macrophage function. However, itcould not be excluded that R and T have direct effect onmicroglia, since it was shown that both R [32,33] and T[34,35] pass BBB.
Having in mind that axonal loss generates permanentneurological deficit in MS it seems particularly interestingthat in RT treated animals axonal damage was not detectedwhile in control animals multiple areas of axonal injury werefound. RT treatment also prevented the appearance ofdemyelinated lesions. Namely, in contrast to large areas ofdemyelination, which correlated with mononuclear cellinfiltration observed in control animals, in RT-treated rats acomplete absence of degraded myelin was seen. There is acommon opinion that axonal injury is consequence ofdemyelination [36]. Of additional interest is a recent findingthat demyelination is not a prerequisite for axonal loss, but itmay be important for the damage spreading through normalappearing white matter [37]. However, further research onanimals and, more importantly, well controlled, randomizedclinical studies will be necessary to elucidate the trueneuroprotective potential of combined RT therapy.
It should be emphasized that in our study there was noevidence for leukopenia, associated with therapeutic appli-cation of RT (data not shown). Therefore, the potential use ofcombinations to overcome effects of single-compoundtherapy allows consumption of lowered doses, which mayreduce toxicity and decrease cell resistance to drugs.
In conclusion, although the precise mechanisms ofsuppressive effects of combined therapy with R and T inongoing EAE are yet unknown, the results gained in this studysuggest that combination of these two nucleoside analoguesmight act at several steps during disease, and probably theseactions function in concert. Combination of these two nu-cleoside analoguesmight be a useful and relatively inexpensiveapproach for the treatment of MS.
Acknowledgements
This work was supported by the Ministry of Science andEnvironmental Protection, Serbia, Grants 143005, 143029,and 145066.
References
[1] Compston A, Coles A. Multiple sclerosis. Lancet 2002;359(9313):1221–31.
[2] Lovas G, Szilagyi N, Majtenyi K, Palkovits M, Komoly S. Axonalchanges in chronic demyelinated cervical spinal cord plaques. Brain2000;123(Pt 2):308–17.
[3] Grigoriadis N, Ben-Hur T, Karussis D, Milonas I. Axonal damage inmultiple sclerosis: a complex issue in a complex disease. Clin NeurolNeurosurg 2004;106(3):211–7.
[4] Franchetti P, Cappellacci L, Grifantini M. IMP dehydrogenase as atarget of antitumor and antiviral chemotherapy. Farmaco 1996;51(7):457–69.
[5] Weber G, Shen F, Prajda N, Yang H, Li W, Yeh A, et al. Regulation ofthe signal transduction program by drugs. Adv Enzyme Regul1997;37:35–55.
[6] Maisch B, Herzum M, Hufnagel G, Bethge C, Schonian U. Immuno-suppressive treatment for myocarditis and dilated cardiomyopathy. EurHeart J 1995;16(Suppl O):153–61.
[7] Pankiewiez KW. Inhibitors of inosine monophosphate dehidrogenaseas potential chemo therapeutic agents. Exo Opin Ther Patents1999;9:55–65.
[8] Tam RC, Lau JY, Hong Z. Mechanisms of action of ribavirin inantiviral therapies. Antivir Chem Chemother 2001;12(5):261–72.
[9] Yamada Y, Natsumeda Y, Weber G. Action of the active metabolites oftiazofurin and ribavirin on purified IMP dehydrogenase. Biochemistry1988;27(6):2193–6.
[10] Weber G, Shen F, Orban TI, Kokeny S, Olah E. Targeting signaltransduction. Adv Enzyme Regul 2003;43:47–56.
[11] Stosic-Grujicic S, Savic-Radojevic A, Maksimovic-Ivanic D, MarkovicM, Bumbasirevic V, Ramic Z, et al. Down-regulation of experimentalallergic encephalomyelitis in DA rats by tiazofurin. J Neuroimmunol2002;130(1–2):66–77.
[12] Milicevic I, Pekovic S, Subasic S, Mostarica-Stojkovic M, Stosic-Grujicic S, Medic-Mijacevic Lj, et al. Ribavirin reduces clinical signsand pathological changes of experimental autoimmune encephalomy-elitis in dark agouti rats. J Neurosci Res 2003;72(2):268–78.
[13] Vukmanovic S, Mostarica Stojkovic M, Lukic ML. Experimentalautoimmune encephalomyelitis in “low” and “high” interleukin 2producer rats. I. Cellular basis of induction. Cell Immunol 1989;121(2):237–46.

85D. Stojkov et al. / Journal of the Neurological Sciences 267 (2008) 76–85
[14] Allison A, Eugui EM. Mycophenolate mofetil and its mechanisms ofaction. Immunopharmacol 2000;47(2–3):85–118.
[15] Peavy DL, Koff WC, Hyman DS, Knight V. Inhibition of lymphocyteproliferative responses by ribavirin. Infect Immun 1980;29(2):583–9.
[16] Heagy W, Crumpacker C, Lopez PA, Finberg RW. Inhibition of immunefunctions by antiviral drugs. J Clin Invest 1991;87(6):1916–24.
[17] Tovari J, Bocsi J, Ladanyi A, Lapis K, Timar J. The antitumor effect ofTiazofurin (TR) consists of anti-proliferative and anti-invasiveelements. Anticancer Res 1996;16(6A):3307–12.
[18] Timar J, Tovari J, Pogany G, Ladanyi A, Paku S, Rso E, et al. Theantimetabolite Tiazofurin (TR) inhibits glycoconjugate biosynthesisand invasiveness of tumour cells. Eur J Cancer 1996;32A(1):152–9.
[19] Weber G, Prajda N, Abonyi M, Look KY, Tricot G. Tiazofurin:molecular and clinical action. Anticancer Res 1996;16(6A):3313–22.
[20] Franchetti P, Grifantini M. Nucleoside and non-nucleoside IMPdehydrogenase inhibitors as antitumor and antiviral agents. CurrMed Chem 1999;6(7):599–614.
[21] Prajda N, Hata Y, Abonyi M, Singhal RL, Weber G. Sequential impactof tiazofurin and ribavirin on the enzymic program of the bone marrow.Cancer Res 1993;53(24):5982–6.
[22] Lavrnja I, Stojkov D, Pekovic S, Subasic S, Mostarica-Stojkovic M,Stosic-Grujicic S, et al. Combination of nucleoside analoguestiazofurin and ribavirin downregulates experimental autoimmuneencephalomyelitis. Ann N YAcad Sci 2005;1048:392–5.
[23] Steinman L. A few autoreactive cells in an autoimmune infiltratecontrol a vast population of nonspecific cells: a tale of smart bombs andthe infantry. Proc Natl Acad Sci U S A 1996;93(6):2253–6.
[24] Brocke S, Gijbels K, Allegretta M, Ferber I, Piercy C, Blankenstein T,et al. Treatment of experimental encephalomyelitis with a peptideanalogue of myelin basic protein. Nature 1996;379(6563):343–6.
[25] Pender MP. Experimental autoimmune encephalomyelitis. In: PenderMP, McCombe PA, editors. Autoimmune neurological disease.Cambridge: Cambridge University Press; 1995. p. 26–88.
[26] Sokoloski JA, Sartorelli AC. Effects of the inhibitors of IMPdehydrogenase, tiazofurin and mycophenolic acid, on glycoproteinmetabolism. Mol Pharmacol 1985;28(6):567–73.
[27] Raivich G, Banati R. Brain microglia and blood-derived macrophages:molecular profiles and functional roles in multiple sclerosis and animal
models of autoimmune demyelinating disease. Brain Res Brain ResRev 2004;46(3):261–81.
[28] Aloisi F, Ria F, Adorini L. Regulation of T-cell responses by CNSantigen-presenting cells: different roles for microglia and astrocytes.Immunol Today 2000;21(3):141–7.
[29] Bohatschek M, Kloss CU, Kalla R, Raivich G. In vitro model ofmicroglial deramification: ramified microglia transform into amoeboidphagocytes following addition of brain cell membranes to microglia-astrocyte cocultures. J Neurosci Res 2001;64(5):508–22.
[30] Flugel A, Bradl M, Kreutzberg GW, Graeber MB. Transformation ofdonor-derived bone marrow precursors into host microglia duringautoimmune CNS inflammation and during the retrograde response toaxotomy. J Neurosci Res 2001;66(1):74–82.
[31] Flugel A, Matsumuro K, Neumann H, Klinkert WE, Birnbacher R,Lassmann H, et al. Anti-inflammatory activity of nerve growth factorin experimental autoimmune encephalomyelitis: inhibition of mono-cyte transendothelial migration. Eur J Immunol 2001;31(1):11–22.
[32] Gilbert BE, Wyde PR. Pharmacokinetics of ribavirin aerosol in mice.Antimicrob Agents Chemother 1988;32(1):117–21.
[33] Hosoya M, Shigeta S, Mori S, Tomoda A, Shiraishi S, Miike T, et al.High-dose intravenous ribavirin therapy for subacute sclerosingpanencephalitis. Antimicrob Agents Chemother 2001;45(3):943–5.
[34] Ristic S, Tasic J, Piperski V, Dacevic M, Petrovic L, Jokanovic M.HPLC determination of tiazofurin in the rat brain. J Pharm BiomedAnal 2003;33(4):839–43.
[35] Janac B, Pesic V, Veskov R, Ristic S, Tasic J, Piperski V, et al. Theeffects of tiazofurin on basal and amphetamine-induced motor activityin rats. Pharmacol Biochem Behav 2004;77(3):575–82.
[36] Aktas O, Waiczies S, Zipp F. Neurodegeneration in autoimmunedemyelination: recent mechanistic insights reveal novel therapeutictargets. J Neuroimmunol 2007;184(1–2):17–26.
[37] Aboul-Enein F, Weiser P, Hoftberger R, Lassmann H, Bradl M.Transient axonal injury in the absence of demyelination: a correlate ofclinical disease in acute experimental autoimmune encephalomyelitis.Acta Neuropathol 2006;111(6):539–47.
Copyright © 2022 FDOKUMEN




















