
Curing of foot-and-mouth disease virus from persistently infected cells by ribavirin involves...
11
Curing of foot-and-mouth disease virus from persistently infected cells by ribavirin involves enhanced mutagenesis Antero Airaksinen, Nonia Pariente, Luis Mene ´ndez-Arias, and Esteban Domingo* Centro de Biologı ´a Molecular “Severo Ochoa” (CSIC-UAM), Universidad Auto ´noma de Madrid, Cantoblanco, 28049 Madrid, Spain Received 16 December 2002; returned to author for revision 28 January 2003; accepted 6 February 2003 Abstract BHK-21 cells persistently infected with foot-and-mouth disease virus (FMDV) can be cured of virus by treatment with the antiviral nucleoside analogue ribavirin. To study whether the process involved an increase in the number of mutations in the mutant spectrum of the viral population, viral genomes were cloned from persistently infected cells treated or untreated with ribavirin. An increase of up to 10-fold in mutation frequencies associated with ribavirin treatment was observed in the viral genomes from the treated cultures as compared with parallel, untreated cultures. To address the possible mechanisms of enhanced mutagenesis, we investigated the mutagenic effects of ribavirin together with guanosine, and mycophenolic acid in the presence or absence of guanosine. Changes in the intracellular nucleotide concentrations were determined for all treatments. The results suggest that the increased mutation frequencies were not dependent on nucleotide pool imbalances or due to selection of preexisting genomes but they were produced by a mutagenic action of ribavirin. © 2003 Elsevier Science (USA). All rights reserved. Keywords: Picornavirus; Foot-and-mouth disease virus; Persistent infection; Mutagenesis; Error catastrophe; Lethal mutagenesis; Ribavirin; Mycophenolic acid Introduction Picornaviruses are the causative agents of many impor- tant human and animal diseases, associated with either acute or persistent infections (Racaniello, 2001; Semler and Wim- mer, 2002). Foot-and-mouth disease virus (FMDV) is a picornavirus of the genus Aphthovirus that causes the eco- nomically most important disease of farm animals world- wide (Sobrino et al., 2001). FMDV usually causes an acute infection in animals, but it may also establish long-term persistent infections in ruminants (van Bekkum et al., 1959; Salt, 1993) and in cell culture (De la Torre et al., 1985). During persistent infections of cattle established with bio- logical clones of FMDV, continuous genetic and antigenic evolution of the virus, with rates of fixation of mutations of up to 7 10 2 substitutions per nucleotide and year, was recorded (Gebauer et al., 1988). A system of BHK-21 cells persistently infected with FMDV clone C-S8c1 was estab- lished (De la Torre et al., 1985). The characterization of this system suggested a steady-state persistent infection, with the majority of cells displaying viral antigens, and the virus and cells undergoing a coevolutionary process in which the cells became gradually resistant to FMDV and the virus gradually more virulent for the parental BHK-21 cells (De la Torre et al., 1988, 1989; Dı ´ez et al., 1990; Martin- Hernandez et al., 1994). The nucleoside analogue ribavi- rin (1--D-ribofuranosyl-1,2,3-triazole-3-carboxamide; Rib) was effective in eliminating FMDV from persistently in- fected cells (De la Torre et al., 1987) without significant decrease of cell viability, but the mechanism of its anti- FMDV activity is not known. Rib has shown antiviral ac- tivity against a number of RNA and DNA viruses (Sidwell et al., 1972; reviewed in Snell, 2001). Rib is phosphorylated by cellular enzymes into ribavirin mono-, di-, and triphos- phate (RMP, RDP and RTP, respectively) (Streeter et al., 1973; Zimmerman and Deeprose, 1978) and exerts its an- tiviral effect through various mechanisms: competitive in- hibition of inosine monophosphate dehydrogenase (IM- PDH; EC 1.1.1.205), which reduces the intracellular * Corresponding author. Fax: 34-91-397-4799. E-mail address: [email protected] (E. Domingo). R Available online at www.sciencedirect.com Virology 311 (2003) 339 –349 www.elsevier.com/locate/yviro 0042-6822/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved. doi:10.1016/S0042-6822(03)00144-2
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Curing of foot-and-mouth disease virus from persistently infected cells by ribavirin involves...

Curing of foot-and-mouth disease virus from persistently infected cellsby ribavirin involves enhanced mutagenesis
Antero Airaksinen, Nonia Pariente, Luis Mene´ndez-Arias, and Esteban Domingo*Centro de Biologıa Molecular “Severo Ochoa” (CSIC-UAM), Universidad Autonoma de Madrid, Cantoblanco, 28049 Madrid, Spain
Received 16 December 2002; returned to author for revision 28 January 2003; accepted 6 February 2003
Abstract
BHK-21 cells persistently infected with foot-and-mouth disease virus (FMDV) can be cured of virus by treatment with the antiviralnucleoside analogue ribavirin. To study whether the process involved an increase in the number of mutations in the mutant spectrum of theviral population, viral genomes were cloned from persistently infected cells treated or untreated with ribavirin. An increase of up to 10-foldin mutation frequencies associated with ribavirin treatment was observed in the viral genomes from the treated cultures as compared withparallel, untreated cultures. To address the possible mechanisms of enhanced mutagenesis, we investigated the mutagenic effects of ribavirintogether with guanosine, and mycophenolic acid in the presence or absence of guanosine. Changes in the intracellular nucleotideconcentrations were determined for all treatments. The results suggest that the increased mutation frequencies were not dependent onnucleotide pool imbalances or due to selection of preexisting genomes but they were produced by a mutagenic action of ribavirin.© 2003 Elsevier Science (USA). All rights reserved.
Keywords: Picornavirus; Foot-and-mouth disease virus; Persistent infection; Mutagenesis; Error catastrophe; Lethal mutagenesis; Ribavirin; Mycophenolicacid
Introduction
Picornaviruses are the causative agents of many impor-tant human and animal diseases, associated with either acuteor persistent infections (Racaniello, 2001; Semler and Wim-mer, 2002). Foot-and-mouth disease virus (FMDV) is apicornavirus of the genusAphthovirus that causes the eco-nomically most important disease of farm animals world-wide (Sobrino et al., 2001). FMDV usually causes an acuteinfection in animals, but it may also establish long-termpersistent infections in ruminants (van Bekkum et al., 1959;Salt, 1993) and in cell culture (De la Torre et al., 1985).During persistent infections of cattle established with bio-logical clones of FMDV, continuous genetic and antigenicevolution of the virus, with rates of fixation of mutations ofup to 7� 10�2 substitutions per nucleotide and year, wasrecorded (Gebauer et al., 1988). A system of BHK-21 cellspersistently infected with FMDV clone C-S8c1 was estab-
lished (De la Torre et al., 1985). The characterization of thissystem suggested a steady-state persistent infection, withthe majority of cells displaying viral antigens, and the virusand cells undergoing a coevolutionary process in which thecells became gradually resistant to FMDV and the virusgradually more virulent for the parental BHK-21 cells (Dela Torre et al., 1988, 1989; Dı´ez et al., 1990; Martin-Hernandez et al., 1994). The nucleoside analogue ribavi-rin (1-�-D-ribofuranosyl-1,2,3-triazole-3-carboxamide; Rib)was effective in eliminating FMDV from persistently in-fected cells (De la Torre et al., 1987) without significantdecrease of cell viability, but the mechanism of its anti-FMDV activity is not known. Rib has shown antiviral ac-tivity against a number of RNA and DNA viruses (Sidwellet al., 1972; reviewed in Snell, 2001). Rib is phosphorylatedby cellular enzymes into ribavirin mono-, di-, and triphos-phate (RMP, RDP and RTP, respectively) (Streeter et al.,1973; Zimmerman and Deeprose, 1978) and exerts its an-tiviral effect through various mechanisms: competitive in-hibition of inosine monophosphate dehydrogenase (IM-PDH; EC 1.1.1.205), which reduces the intracellular
* Corresponding author. Fax:�34-91-397-4799.E-mail address: [email protected] (E. Domingo).
R
Available online at www.sciencedirect.com
Virology 311 (2003) 339–349 www.elsevier.com/locate/yviro
0042-6822/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved.doi:10.1016/S0042-6822(03)00144-2

concentrations of guanine nucleotides (Franklin and Cook,1969; Streeter et al., 1973), inhibition of some viral RNA-dependent RNA polymerases (Eriksson et al., 1977; Wray etal., 1985a,b; Toltzis et al., 1988; Fernandez-Larsson et al.,1989), inhibition of mRNA cap formation (Goswami et al.,1979; Scheidel and Stollar, 1991), and enhancement of Th1antiviral immune responses (Hultgren et al., 1998; Ning etal., 1998). Recently, it has been shown that Rib exerts anantiviral action against poliovirus and hepatitis C virus(HCV) associated with enhanced mutagenesis, as a result ofincorporation of RMP by poliovirus and HCV polymerases(Crotty et al., 2000, 2001; Maag et al., 2001). The muta-genic activity resulted from equal pairing of Rib with cyti-dine and uridine during viral RNA synthesis.
The present study was undertaken to investigate whetherRib-mediated curing of virus from cells persistently infectedwith FMDV involved enhanced mutagenesis of the virus.Since the effects of IMPDH inhibition can be reversed byaddition of 100 �M guanosine, compensating for the re-duced de novo synthesis of guanine nucleotides (Zimmer-man and Deeprose, 1978; Kerr, 1987; Landford et al.,2001), we analyzed the effects of (1) Rib alone, (2) Ribtogether with guanosine, to measure the effects under con-ditions in which cellular GTP concentrations were not de-pleted, (3) mycophenolic acid (MPA), a noncompetitiveinhibitor of IMPDH with apparently no other mechanismsof action, to measure the effects of IMPDH inhibition(Lowe et al., 1977; Lee et al., 1985; Sintchak et al., 1996),and (4) MPA together with guanosine to study whetherMPA had effects other than IMPDH inhibition. Nucleotidesequences of the regions coding for VP1 (capsid protein)and 3D (polymerase) of viral molecular clones were deter-mined, and changes in intracellular nucleotide pools wereanalyzed. The results support a mutagenic action of Ribassociated with curing of FMDV from persistently infectedcells.
Results
Effects of antiviral treatments on viral production ofpersistent FMDV
Rib and MPA are inhibitors of the cellular enzymeIMPDH, and the effects of this inhibition can be reversed byaddition of guanosine (Zimmerman and Deeprose, 1978;Kerr, 1987; Lanford et al., 2001). In order to compare theantiviral efficiencies of these two IMPDH inhibitors, per-sistently infected cells of passage 40 were treated for 3 dayswith 100 or 500 �M Rib, 15 �M MPA, or standard culturemedium, each of these in the presence or absence of up to100 �M guanosine (Fig. 1). None of the treatments had aclear antiviral effect during the first 24 h. High concentra-tions of guanosine in combination with 100 �M Rib or 15�M MPA resulted in an increase of virus production during
the first 24 h. After 2 days, the antiviral effect was evidentin all treatments containing Rib or MPA. This antiviraleffect was significantly suppressed by guanosine in a dose-dependent manner (Fig. 1). After 3 days, viral titers becamebarely detectable in treatments involving 100 or 500 �MRib together with 0 to 20 �M guanosine, while 100 �Mguanosine was able to suppress the antiviral effect consid-erably. Treatments with MPA in combination withguanosine had similar results, but viral titers were somewhathigher in all cases. Hence, the antiviral effect of Rib wasstronger than that of MPA against FMDV in persistentlyinfected cells.
Effects of antiviral treatments on intracellular nucleotidepools
The drug concentrations used were adjusted in prelimi-nary measurements so that the effects of the treatments with
Fig. 1. FMDV production by persistently infected cells subjected to varioustreatments. Samples were taken from the media at 1, 2, and 3 days afteradding the drugs to the media on confluent cell monolayers. Culturemedium was changed daily after samples were taken. A parallel controlexperiment without drugs cannot be described here, since in the absence ofdrugs, the high virus production in a confluent cell monolayer produced acytopathic effect (de la Torre et al., 1985). Results are the average of twoexperiments, and differences between corresponding samples did not ex-ceed 1 logarithm. Procedures for cell culture and titration of FMDV aredescribed under Materials and methods.
340 A. Airaksinen et al. / Virology 311 (2003) 339–349
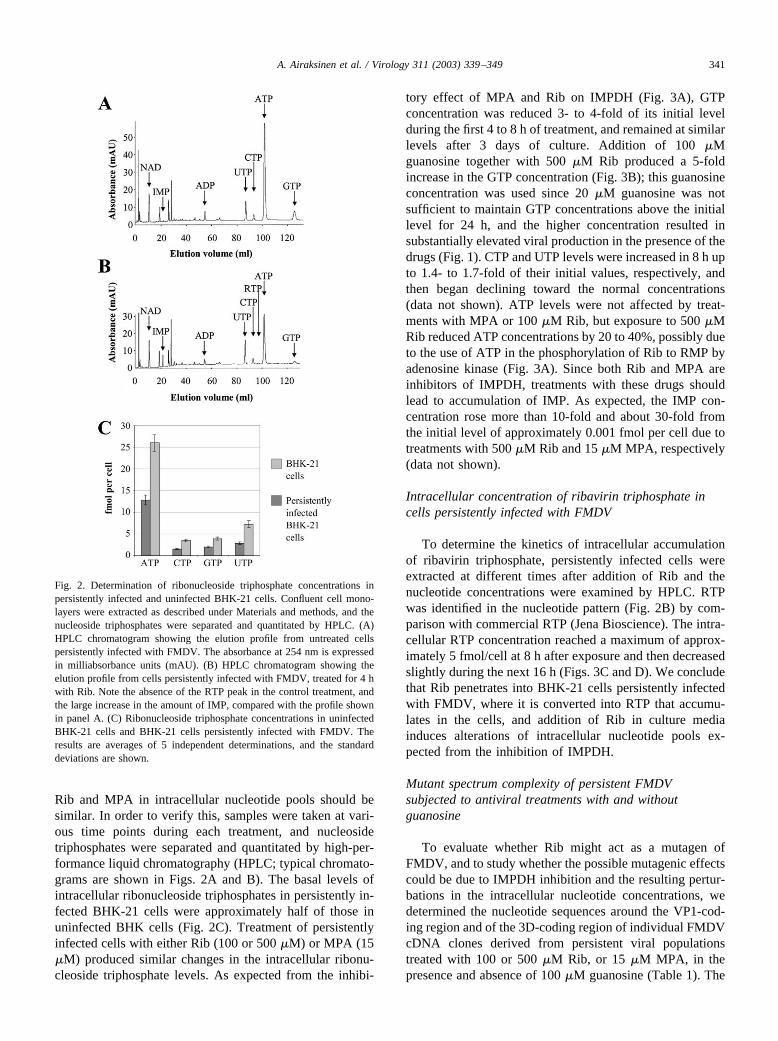
Rib and MPA in intracellular nucleotide pools should besimilar. In order to verify this, samples were taken at vari-ous time points during each treatment, and nucleosidetriphosphates were separated and quantitated by high-per-formance liquid chromatography (HPLC; typical chromato-grams are shown in Figs. 2A and B). The basal levels ofintracellular ribonucleoside triphosphates in persistently in-fected BHK-21 cells were approximately half of those inuninfected BHK cells (Fig. 2C). Treatment of persistentlyinfected cells with either Rib (100 or 500 �M) or MPA (15�M) produced similar changes in the intracellular ribonu-cleoside triphosphate levels. As expected from the inhibi-
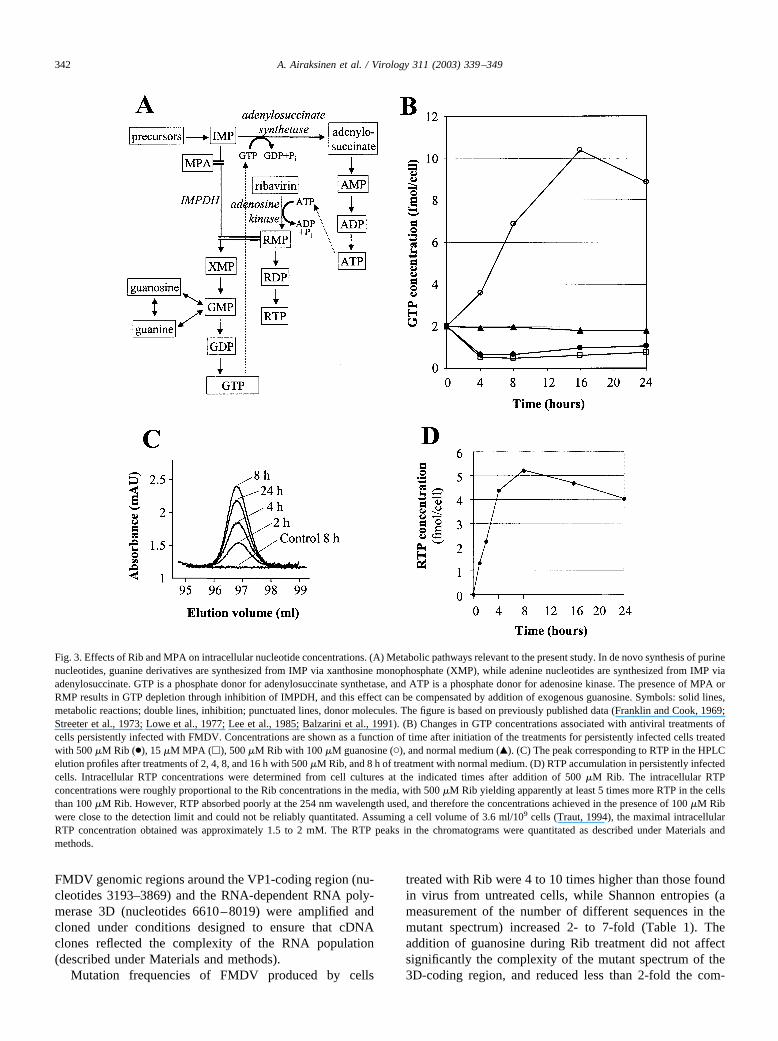
tory effect of MPA and Rib on IMPDH (Fig. 3A), GTPconcentration was reduced 3- to 4-fold of its initial levelduring the first 4 to 8 h of treatment, and remained at similarlevels after 3 days of culture. Addition of 100 �Mguanosine together with 500 �M Rib produced a 5-foldincrease in the GTP concentration (Fig. 3B); this guanosineconcentration was used since 20 �M guanosine was notsufficient to maintain GTP concentrations above the initiallevel for 24 h, and the higher concentration resulted insubstantially elevated viral production in the presence of thedrugs (Fig. 1). CTP and UTP levels were increased in 8 h upto 1.4- to 1.7-fold of their initial values, respectively, andthen began declining toward the normal concentrations(data not shown). ATP levels were not affected by treat-ments with MPA or 100 �M Rib, but exposure to 500 �MRib reduced ATP concentrations by 20 to 40%, possibly dueto the use of ATP in the phosphorylation of Rib to RMP byadenosine kinase (Fig. 3A). Since both Rib and MPA areinhibitors of IMPDH, treatments with these drugs shouldlead to accumulation of IMP. As expected, the IMP con-centration rose more than 10-fold and about 30-fold fromthe initial level of approximately 0.001 fmol per cell due totreatments with 500 �M Rib and 15 �M MPA, respectively(data not shown).
Intracellular concentration of ribavirin triphosphate incells persistently infected with FMDV
To determine the kinetics of intracellular accumulationof ribavirin triphosphate, persistently infected cells wereextracted at different times after addition of Rib and thenucleotide concentrations were examined by HPLC. RTPwas identified in the nucleotide pattern (Fig. 2B) by com-parison with commercial RTP (Jena Bioscience). The intra-cellular RTP concentration reached a maximum of approx-imately 5 fmol/cell at 8 h after exposure and then decreasedslightly during the next 16 h (Figs. 3C and D). We concludethat Rib penetrates into BHK-21 cells persistently infectedwith FMDV, where it is converted into RTP that accumu-lates in the cells, and addition of Rib in culture mediainduces alterations of intracellular nucleotide pools ex-pected from the inhibition of IMPDH.
Mutant spectrum complexity of persistent FMDVsubjected to antiviral treatments with and withoutguanosine
To evaluate whether Rib might act as a mutagen ofFMDV, and to study whether the possible mutagenic effectscould be due to IMPDH inhibition and the resulting pertur-bations in the intracellular nucleotide concentrations, wedetermined the nucleotide sequences around the VP1-cod-ing region and of the 3D-coding region of individual FMDVcDNA clones derived from persistent viral populationstreated with 100 or 500 �M Rib, or 15 �M MPA, in thepresence and absence of 100 �M guanosine (Table 1). The
Fig. 2. Determination of ribonucleoside triphosphate concentrations inpersistently infected and uninfected BHK-21 cells. Confluent cell mono-layers were extracted as described under Materials and methods, and thenucleoside triphosphates were separated and quantitated by HPLC. (A)HPLC chromatogram showing the elution profile from untreated cellspersistently infected with FMDV. The absorbance at 254 nm is expressedin milliabsorbance units (mAU). (B) HPLC chromatogram showing theelution profile from cells persistently infected with FMDV, treated for 4 hwith Rib. Note the absence of the RTP peak in the control treatment, andthe large increase in the amount of IMP, compared with the profile shownin panel A. (C) Ribonucleoside triphosphate concentrations in uninfectedBHK-21 cells and BHK-21 cells persistently infected with FMDV. Theresults are averages of 5 independent determinations, and the standarddeviations are shown.
341A. Airaksinen et al. / Virology 311 (2003) 339–349
FMDV genomic regions around the VP1-coding region (nu-cleotides 3193–3869) and the RNA-dependent RNA poly-merase 3D (nucleotides 6610–8019) were amplified andcloned under conditions designed to ensure that cDNAclones reflected the complexity of the RNA population(described under Materials and methods).
Mutation frequencies of FMDV produced by cells
treated with Rib were 4 to 10 times higher than those foundin virus from untreated cells, while Shannon entropies (ameasurement of the number of different sequences in themutant spectrum) increased 2- to 7-fold (Table 1). Theaddition of guanosine during Rib treatment did not affectsignificantly the complexity of the mutant spectrum of the3D-coding region, and reduced less than 2-fold the com-
Fig. 3. Effects of Rib and MPA on intracellular nucleotide concentrations. (A) Metabolic pathways relevant to the present study. In de novo synthesis of purinenucleotides, guanine derivatives are synthesized from IMP via xanthosine monophosphate (XMP), while adenine nucleotides are synthesized from IMP viaadenylosuccinate. GTP is a phosphate donor for adenylosuccinate synthetase, and ATP is a phosphate donor for adenosine kinase. The presence of MPA orRMP results in GTP depletion through inhibition of IMPDH, and this effect can be compensated by addition of exogenous guanosine. Symbols: solid lines,metabolic reactions; double lines, inhibition; punctuated lines, donor molecules. The figure is based on previously published data (Franklin and Cook, 1969;Streeter et al., 1973; Lowe et al., 1977; Lee et al., 1985; Balzarini et al., 1991). (B) Changes in GTP concentrations associated with antiviral treatments ofcells persistently infected with FMDV. Concentrations are shown as a function of time after initiation of the treatments for persistently infected cells treatedwith 500 �M Rib (●), 15 �M MPA (�), 500 �M Rib with 100 �M guanosine (�), and normal medium (Œ). (C) The peak corresponding to RTP in the HPLCelution profiles after treatments of 2, 4, 8, and 16 h with 500 �M Rib, and 8 h of treatment with normal medium. (D) RTP accumulation in persistently infectedcells. Intracellular RTP concentrations were determined from cell cultures at the indicated times after addition of 500 �M Rib. The intracellular RTPconcentrations were roughly proportional to the Rib concentrations in the media, with 500 �M Rib yielding apparently at least 5 times more RTP in the cellsthan 100 �M Rib. However, RTP absorbed poorly at the 254 nm wavelength used, and therefore the concentrations achieved in the presence of 100 �M Ribwere close to the detection limit and could not be reliably quantitated. Assuming a cell volume of 3.6 ml/109 cells (Traut, 1994), the maximal intracellularRTP concentration obtained was approximately 1.5 to 2 mM. The RTP peaks in the chromatograms were quantitated as described under Materials andmethods.
342 A. Airaksinen et al. / Virology 311 (2003) 339–349
plexity of the VP1-coding region. Therefore, Rib was foundto be mutagenic to FMDV, and this effect was largelyindependent of the depletion of intracellular GTP concen-tration.
The MPA-induced mutation frequencies were 2.5- to6-fold higher than in the untreated viral population, remain-ing clearly below the ones observed with Rib treatments(Table 1). Shannon entropies increased 3- to 4-fold. Addi-tion of guanosine in the MPA treatment abolished the mu-tagenic effect, suggesting that the mutations were due toIMPDH inhibition (Table 1).
Types of mutations found
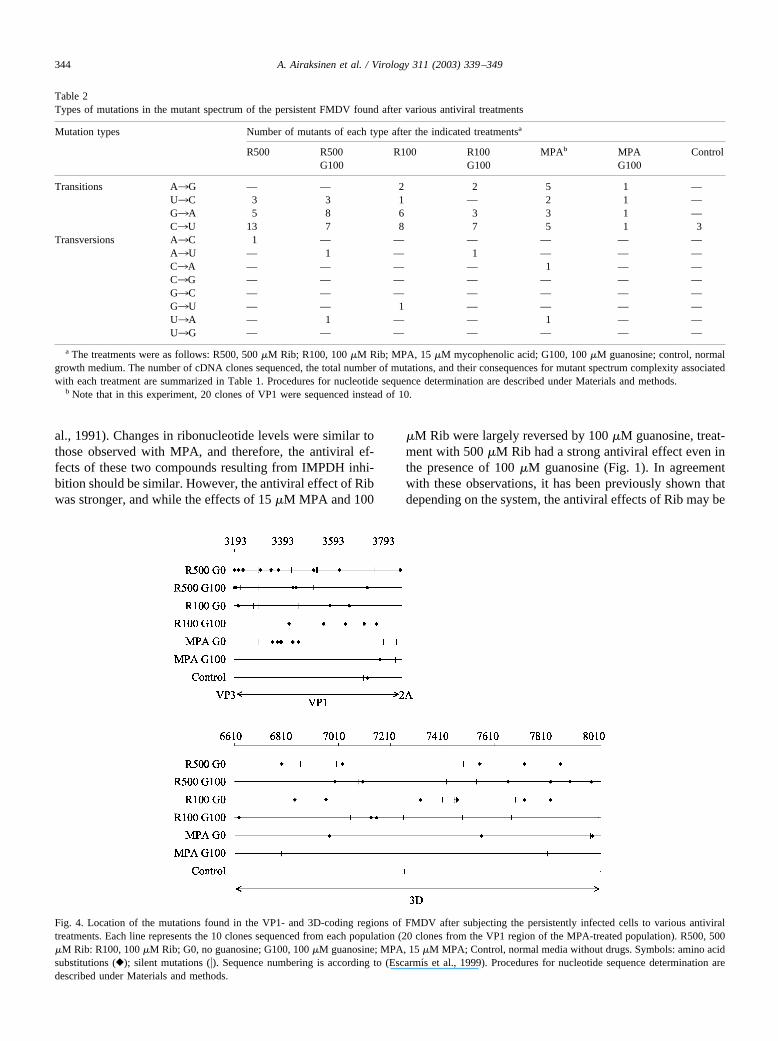
In treatments with 100 or 500 �M Rib, with or withoutguanosine, a total of 57 mutations were found of the typesC3U and G3A, and 11 mutations of the types U3C andA3G (Table 2). Mutations C3U and G3A should beexpected if mutations resulted from Rib incorporation op-posite to C, while mutations U3C and A3G should beexpected if mutations resulted from Rib incorporation op-posite to U. In contrast to the results with Rib-treated pop-ulations, treatment with MPA did not show the same mu-tational bias: there were 10 mutations of the types C3Uand G3A, and 9 mutations of the types U3C and A3G.Synonymous and nonsynonymous mutations were relativelyuniformly distributed along the genomic regions analyzed(Fig. 4). A total of 97 mutations were found in this study, 46in the 3D-coding region (22 of which led to amino acidsubstitutions) and 51 in the VP1 region (25 amino acid
substitutions). Since 3D consists of 470 amino acids andVP1 of 209 amino acids, the results show a lower accept-ability of mutations in the polymerase-coding region. Theresults support a significant mutagenic effect of Rib asso-ciated with curing of FMDV from persistently infectedcells.
Discussion
Persistently infected cells can be cured of FMDV by thenucleoside analogue Rib. Previous studies from several lab-oratories have indicated that Rib exerts its antiviral effectsthrough different biochemical mechanisms, depending onthe virus and the host cell (Malinoski and Stollar, 1981;Jordan et al., 1999), and in many cases the antiviral effectcannot be ascribed to a single mechanism. Rib and MPA areboth inhibitors of the cellular enzyme IMPDH, producing asignificant reduction in the intracellular GTP concentration(Franklin and Cook, 1969; Streeter et al., 1973). Contrary toRib, there are no other known targets for MPA apart fromIMPDH (Franklin and Cook, 1969; Lowe et al., 1977; Leeet al., 1985). Treatment with MPA had a clear antiviraleffect that was suppressed when GTP pools were restoredby adding guanosine exogenously. Thus, MPA was able toreduce viral replication efficiency by altering the host cellribonucleotide pools, as a result of IMPDH inhibition.
Treatment of cells with Rib reduced the intracellularGTP concentration 3- to 4-fold, in agreement with previousreports (Lowe et al., 1977; Wray et al., 1985b; Balzarini et
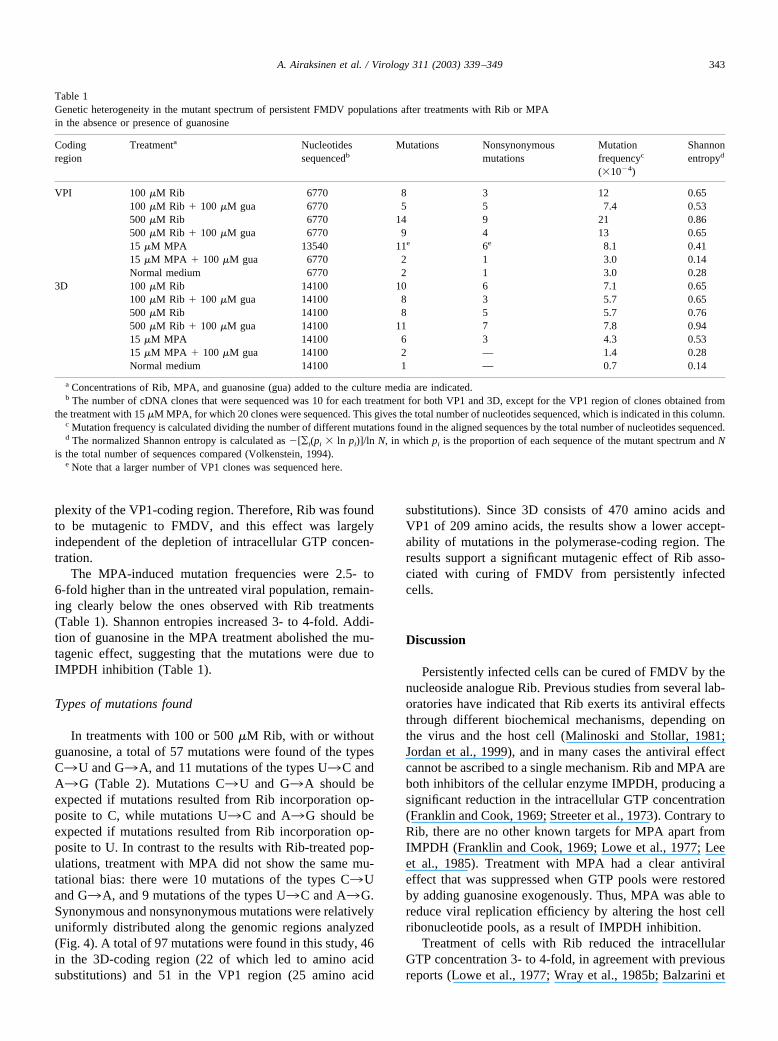
Table 1Genetic heterogeneity in the mutant spectrum of persistent FMDV populations after treatments with Rib or MPAin the absence or presence of guanosine
Codingregion
Treatmenta Nucleotidessequencedb
Mutations Nonsynonymousmutations
Mutationfrequencyc
(�10�4)
Shannonentropyd
VPI 100 �M Rib 6770 8 3 12 0.65100 �M Rib � 100 �M gua 6770 5 5 7.4 0.53500 �M Rib 6770 14 9 21 0.86500 �M Rib � 100 �M gua 6770 9 4 13 0.6515 �M MPA 13540 11e 6e 8.1 0.4115 �M MPA � 100 �M gua 6770 2 1 3.0 0.14Normal medium 6770 2 1 3.0 0.28
3D 100 �M Rib 14100 10 6 7.1 0.65100 �M Rib � 100 �M gua 14100 8 3 5.7 0.65500 �M Rib 14100 8 5 5.7 0.76500 �M Rib � 100 �M gua 14100 11 7 7.8 0.9415 �M MPA 14100 6 3 4.3 0.5315 �M MPA � 100 �M gua 14100 2 — 1.4 0.28Normal medium 14100 1 — 0.7 0.14
a Concentrations of Rib, MPA, and guanosine (gua) added to the culture media are indicated.b The number of cDNA clones that were sequenced was 10 for each treatment for both VP1 and 3D, except for the VP1 region of clones obtained from
the treatment with 15 �M MPA, for which 20 clones were sequenced. This gives the total number of nucleotides sequenced, which is indicated in this column.c Mutation frequency is calculated dividing the number of different mutations found in the aligned sequences by the total number of nucleotides sequenced.d The normalized Shannon entropy is calculated as �[¥i(pi � ln pi)]/ln N, in which pi is the proportion of each sequence of the mutant spectrum and N
is the total number of sequences compared (Volkenstein, 1994).e Note that a larger number of VP1 clones was sequenced here.
343A. Airaksinen et al. / Virology 311 (2003) 339–349
al., 1991). Changes in ribonucleotide levels were similar tothose observed with MPA, and therefore, the antiviral ef-fects of these two compounds resulting from IMPDH inhi-bition should be similar. However, the antiviral effect of Ribwas stronger, and while the effects of 15 �M MPA and 100
�M Rib were largely reversed by 100 �M guanosine, treat-ment with 500 �M Rib had a strong antiviral effect even inthe presence of 100 �M guanosine (Fig. 1). In agreementwith these observations, it has been previously shown thatdepending on the system, the antiviral effects of Rib may be
Table 2Types of mutations in the mutant spectrum of the persistent FMDV found after various antiviral treatments
Mutation types Number of mutants of each type after the indicated treatmentsa
R500 R500 R100 R100 MPAb MPA ControlG100 G100 G100
Transitions A3G — — 2 2 5 1 —U3C 3 3 1 — 2 1 —G3A 5 8 6 3 3 1 —C3U 13 7 8 7 5 1 3
Transversions A3C 1 — — — — — —A3U — 1 — 1 — — —C3A — — — — 1 — —C3G — — — — — — —G3C — — — — — — —G3U — — 1 — — — —U3A — 1 — — 1 — —U3G — — — — — — —
a The treatments were as follows: R500, 500 �M Rib; R100, 100 �M Rib; MPA, 15 �M mycophenolic acid; G100, 100 �M guanosine; control, normalgrowth medium. The number of cDNA clones sequenced, the total number of mutations, and their consequences for mutant spectrum complexity associatedwith each treatment are summarized in Table 1. Procedures for nucleotide sequence determination are described under Materials and methods.
b Note that in this experiment, 20 clones of VP1 were sequenced instead of 10.
Fig. 4. Location of the mutations found in the VP1- and 3D-coding regions of FMDV after subjecting the persistently infected cells to various antiviraltreatments. Each line represents the 10 clones sequenced from each population (20 clones from the VP1 region of the MPA-treated population). R500, 500�M Rib: R100, 100 �M Rib; G0, no guanosine; G100, 100 �M guanosine; MPA, 15 �M MPA; Control, normal media without drugs. Symbols: amino acidsubstitutions (}); silent mutations (�). Sequence numbering is according to (Escarmıs et al., 1999). Procedures for nucleotide sequence determination aredescribed under Materials and methods.
344 A. Airaksinen et al. / Virology 311 (2003) 339–349

either partially or mostly reversed by guanosine (Streeter etal., 1973; Wray et al., 1985b; Scheidel and Strollar, 1991;Jordan et al., 1999), and that the dose-dependent antiviralactivity can increase even when the additional drug has nofurther effect on GTP pools (Wray et al., 1985b). In somesystems the antiviral activity of Rib seems to be eitherindependent of the IMPDH inhibition (Smee and Matthews,1986) or limited to the same level as with other IMPDHinhibitors (Wray et al., 1986; Lanford et al., 2001). There-fore, in some viral systems but not others, Rib possesses anantiviral activity that is independent of the effects on nu-cleotide pools.
Recent studies have raised the possibility that Rib couldexert its action through increased mutagenesis (Crotty et al.,2000, 2001; Maag et al., 2001; Contreras et al., 2002). Wefound an increase in mutation frequencies due to treatmentswith either Rib or MPA, although the latter treatment re-sulted in a less pronounced increase in mutant spectrumcomplexity (Table 1). The mutagenic effect of MPA wasabolished by addition of guanosine, while that of Rib wasonly slightly reduced. The mutations found in MPA-treatedpopulations were presumably due to an increased number oftransition mutations that were favored by changes in therelative intracellular ribonucleotide concentrations, al-though there was no preference for C3U and G3A mu-tations that might be expected due to GTP depletion (Table2). Thus, Rib had an additional source of antiviral activitynot ascribable to IMPDH inhibition that was consistent withmutagenesis through incorporation of Rib into the FMDVgenomic RNA. As for other RNA viruses, FMDV replica-tion is error prone and the mutation rates are close to anerror threshold that sets a limit for maintenance of geneticinformation (Swetina and Schuster, 1982; Eigen andBiebricher, 1988; Holland et al., 1990; Drake and Holland,1999; Eigen, 2000; Sierra et al., 2000; Pariente et al., 2001).Curing of persistently infected cells from FMDV by Ribthus appears to provide yet another example of virus ex-tinction through lethal mutagenesis (Eigen, 2002).
Nucleotide sequencing of the VP1 and 3D genes of viralcDNA clones obtained from the mutant spectrum of persis-tent FMDV after various drug treatments indicates a muta-genic action of Rib. The observed Rib-induced mutationfrequencies are close to those reported for poliovirus andHCV (Crotty et al., 2001; Contreras et al., 2002). Mutationfrequencies in untreated viral populations were similar tothose found earlier in FMDV from persistent or cytolyticinfections (De la Torre et al., 1988; Sierra et al., 2000; Ariaset al., 2001; Pariente et al., 2001), while they were 2- to10-fold higher in populations treated with Rib or MPA. Themutagenic effect of Rib was only mildly suppressed byguanosine addition, indicating that the mutations were notdue to ribonucleotide pool imbalances resulting fromIMPDH inhibition. The reduced amount of GTP might,however, favor incorporation of Rib into the viral RNA(RTP:GTP ratios were up to 10; compare Figs. 3B and D),or increase the number of transition mutations through in-
corporation of adenosine instead of guanosine. The latterpossibility is less likely, since Rib decreased the ATP poolsmore than MPA, and therefore the GTP:ATP ratios wereless affected by Rib treatment. It could be argued that Ribcured FMDV by some other mechanism and that the in-crease in mutant spectrum complexity was the result ofdominance of minority components of the mutant spectrum.This is highly unlikely because minority genomes analyzedfrom FMDV mutant spectra rarely show a mutational biasof the type seen in the Rib-treated FMDV. Furthermore, inno previous case a mutant spectrum of FMDV showed anincrease in complexity unless the population had been sub-jected to mutagenic treatments (Sierra et al., 2000; Parienteet al., 2001). The amino acid substitutions were also ratheruniformly distributed within both VP1 and 3D (Fig. 4). InVP1, 43% of the mutations occurred at residues that werenot accessible to the solvent (Lea et al., 1994; Mateu et al.,1994), implying a mutagenic action since during FMDVevolution, mutations mostly occur in the exposed residuesof the capsid (Semler and Wimmer, 2002, and chapterstherein).
Rib treatment resulted in an increase in C3U and G3Amutations (Table 2), as reported earlier for poliovirus(Crotty et al., 2000, 2001). The frequencies of the other twotypes of transition mutations (U3C and A3G) were muchlower, and similar in all treatments. This is in contrast towhat would be expected if Rib paired equally well withcytidine and uridine, as has been demonstrated for the po-liovirus and HCV polymerases in vitro (Crotty et al., 2000,2001; Maag et al., 2001). Mutations C3U and G3Ashould both occur if Rib is incorporated opposite to cyti-dine, either during negative strand synthesis (C3U) orduring positive strand synthesis (G3A). It would be pos-sible to explain the preference for these two mutations byfavored Rib incorporation when the competing GTP is lim-ited, or as a result of spontaneous transitions due to lowGTP concentration. However, these explanations are notconsistent with the data obtained with treatments involvingRib together with guanosine, since C3U and G3A mu-tations were more abundant in these viral populations aswell. It seems that during FMDV replication Rib does havea preference to be incorporated opposite to cytidine insteadof uridine, or that uridine is incorporated more frequentlythan cytidine when Rib is present in the template.
The mutation frequencies found in all Rib treatments,with or without guanosine, imply an average of 7 mutationsper genome. Due to negative selection of at least the poly-merase-coding region, it is possible that the actual muta-genic effect was stronger than that reflected by the clonessequenced, since the small difference in mutation frequen-cies obtained in treatments with 500 or 100 �M Rib with100 �M guanosine (Table 1) is probably not sufficient toexplain the large difference in the antiviral effects of thesetwo treatments (Fig. 1). Alternatively, other effects of Ribsuch as inhibition of viral RNA synthesis could participatein curing of FMDV (Snell, 2001). Inhibition of cap forma-
345A. Airaksinen et al. / Virology 311 (2003) 339–349

tion should not have an antiviral effect on FMDV, sincepicornaviruses use a cap-independent initiation of transla-tion (Jang et al., 1988; Pelletier and Sonenberg, 1988), andthey do not depend on capped cellular mRNA molecules forreplication (reviews in Semler and Wimmer, 2002).
Rib-mediated mutagenesis has been reported for polio-virus, hepatitis C virus, Hantaan virus and possibly GB virus B(Crotty et al., 2000; Lanford et al., 2001; Maag et al., 2001;Contreras et al., 2002; Severson et al., 2003), while no mu-tagenesis was found for lymphocytic choriomeningitis virus(LCMV) despite a potent antiviral effect of Rib (Ruız-Jarabo etal., 2003). The HCV RNA polymerase was found to incorpo-rate Rib and to induce mutagenic base pairing, albeit lessefficiently than the poliovirus RNA polymerase (Maag et al.,2001), and Rib was mutagenic for HCV in a plasmid-basedreplication system (Contreras et al., 2002). In HCV patients,however, no evidence of mutagenesis has been seen (Gerotto etal., 1999; Querenghi et al., 2001; Sookoian et al., 2001), pos-sibly due to the low Rib concentrations reached at the sites ofviral replication. Consistent with earlier findings (Rankin etal., 1989), we found Rib to accumulate inside the cells,yielding maximal concentrations of approximately 1.5 to 2mM RTP in the presence of 500 �M Rib in the medium(legend to Fig. 3D). The maximal in vivo Rib concentrationat the site of infection has been reported to be 600 to 800�M in respiratory fluids after small-particle aerosol admin-istration (Gilbert and Knight, 1986), while intravenous ad-ministration was estimated to result in peak concentrationsof approximately 20–150 �M in serum, which rapidly de-clined to levels as low as 1 �M (Connor et al., 1993; Jordanet al., 2000). Oral administration resulted in concentrationsbelow 20 �M in serum and 10 �M in cerebrospinal fluid(Crumpacker et al., 1986; Ogle et al., 1989; Connor et al.,1993; Anderson and Rahal, 2002). All these values in vivoare clearly lower than the concentrations that have beenfound to be mutagenic in cell culture, and they may not besufficient for a significant mutagenic activity of RTP.
In summary, we have shown that the antiviral effect ofRib on FMDV in persistent infection is caused by at leasttwo mechanisms: (1) direct mutagenesis and (2) imbalancesin ribonucleotide pools resulting from IMPDH inhibition.Our results support Rib incorporation into viral RNA, butsuggest that Rib does not pair equally well with cytidine anduridine. Further studies are needed to clarify the base pair-ing and incorporation properties of RTP by viral poly-merases in vitro and to investigate whether intracellularconcentrations of Rib could be high enough in vivo to allowsignificant incorporation of the drug in the viral RNA.
Materials and methods
Cells, viruses, and treatments
The baby hamster kidney cell line (BHK-21), persis-tently infected with FMDV C-S8c1, and procedures for its
maintenance have been characterized previously (De laTorre et al., 1985, 1987, 1988, 1989). Persistently infectedcells at passage 40 were used. Cells were shedding virus tothe medium with maximal titers of about 105 PFU/ml (De laTorre et al., 1988; Dıez et al., 1990). FMDV C-S8c1 is aplaque-purified derivative of the natural isolate C1 SantaPau-Spain/70 (Sobrino et al., 1983). Drug treatments ofpersistently infected cells were initiated as the cell mono-layers reached confluence. The cells were washed withnormal medium [Dulbecco’s modified Eagle’s medium(Gibco) supplemented with nonessential amino acids(Gibco), 50 mg/ml gentamicin (Sigma), and 5% fetal calfserum (Gibco)], and new media were added, containing Rib(0, 100, or 500 �M) or MPA (0 or 15 �M; Sigma) in thepresence or absence of guanosine (0, 5, 20, or 100 �M;Sigma). The media were changed daily, and samples fromthe media were taken immediately before each change. Inthe experiments with Rib or MPA, the drugs did not causesignificant cytotoxicity, as initially confluent monolayersremained confluent during the 3 days of treatment. After24 h, the number of viable cells had increased by 40 % inthe untreated cultures, 15% in Rib-treated cultures, and 7%in MPA-treated cultures.
Sample preparation for high-performance liquidchromatography
To prepare samples from cell cultures for quantificationof ribonucleotide levels by HPLC, we used the systemdescribed by Pogolotti and Santi (1982), with minor modi-fications. Briefly, the cell monolayers (� 60 mm) werewashed with phosphate-buffered saline, treated with 300 �lof 0.6 M trichloroacetic acid, and incubated on ice for 10min. The supernatants were collected and extracted with1,1,2-trichlorotrifluoroethane (Sigma) containing 0.5 M tri-n-octylamine (Sigma). The mixtures were vortexed for 10 sand centrifuged 30 s at 12,000 g. The upper phase wascollected and immediately frozen at �80°C, and upon thaw-ing, the samples were analyzed immediately. The entiretreatment was performed at 0 to 4°C, using ice-cold solu-tions. The ATP:ADP ratio was consistently above 20, indi-cating minimal loss of the terminal phosphate during samplepreparation.
HPLC analysis of nucleotide pools
Nucleotides were separated using a 4.6 � 250-mm Par-tisil 10 SAX column (Whatman) with a 4.6 � 30-mmPartisil 10 SAX precolumn (Phenomenex). Around 25 pmolof nucleotides in a total volume of 100 �l were analyzed.After washing the column with 18 ml of buffer A (7 mMNH4H2PO4, pH 3.8), nucleotides were eluted with a lineargradient prepared with 45 ml of buffer A and 45 ml ofbuffer B (250 mM NH4H2PO4, 500 mM KCl, pH 4.5),followed by a final wash with 30 ml of buffer B (Pogolottiand Santi, 1982). The flow rate was 0.8 ml/min and the
346 A. Airaksinen et al. / Virology 311 (2003) 339–349

optical density of the eluant was continuously monitored at254 nm. Nucleotide amounts were estimated from a stan-dard curve obtained using samples containing knownamounts of each nucleotide eluted under the same condi-tions. To correct for the fluctuations in the numbers of cellsanalyzed, peak sizes were normalized against the well-isolated and reproducible NAD peak of each sample, theconcentration of which was not affected by any of thetreatments. Chromatograms were analyzed using the Uni-corn 3.00 software (Amersham Pharmacia Biotech). Nucle-otide peaks were identified by comparison with the ob-served retention times of commercial solutions of RTP(Jena Bioscience), IMP, NAD (Sigma), ATP, ADP, CTP,GTP, and UTP (Amersham Pharmacia Biotech). In addi-tion, the identity of the NAD peak was confirmed by massspectroscopy as described below.
Mass spectrometry analysis
The identity of the presumed NAD peak was confirmedby analyzing by matrix-assisted laser desorption/ionizationtime-of-flight (MALDI-TOF) mass spectrometry using anexternally calibrated Reflex III instrument (Brucker-Fran-zen Analytic, Bremen, Germany) operating in the positive-ion reflection mode. HPLC-collected fractions were lyoph-ilized and resuspended in 50 �l of distilled water. Part ofthis sample (5 �l) was then mixed with 10 �l of a 10 mg/mlsolution of 2,4,6-trihydroxyacetophenone matrix in ethanol;1 �l of this mixture was dried down and subjected toanalysis.
PCR, molecular cloning, and sequencing
Viral RNA was purified from 300 �l of persistentlyinfected cell culture supernatant using the RNeasy Mini Kit(Qiagen). Reverse transcription and PCR were performedwith AMV reverse transcriptase (Promega) and high fidel-ity Pwo DNA polymerase (Roche). Primers for PCR am-plification of the 3D polymerase-coding region were5�-CAGAGCTCGACCCTGAACCGCA-3� (positions 6580–6602 of the FMDV genome) and 5�-GCCCCGAATTCAT-TGTGC-3� (positions 8060–8043), designed to create tar-get sites for restriction endonucleases SacI and EcoRI,respectively. Primers for amplification of the VP1-codingregion were 5�-CTTTGAGCTCCGGCTACCTGTG-3� (po-sitions 3171–3192) and 5�-GAGAAGAAGAAGGGC-CCAGGGTTGGAC-3� (positions 3896–3870), with targetsites for restriction endonucleases SacI and BamHI, respec-tively. Numbering of residues in the FMDV genome isaccording to Escarmıs et al. (1999). The PCR products weredigested using the indicated enzymes and ligated into thepolycloning site of plasmid pGEM-4Z (Amersham). Com-petent Escherichia coli DH5� bacteria were transformed bythe plasmid and plated, and individual colonies were pickedfor PCR amplification. PCR of the DNA from bacterialcolonies, together with negative controls, were performed
using AmpliTaq DNA polymerase (Perkin Elmer) and theprimers described above. The PCR products were se-quenced entirely in both orientations with an ABI sequencerusing appropriately spaced primers, as previously described(Sierra et al., 2000; Pariente et al., 2001).
Samples for the analysis of quasispecies complexity
Mutant spectrum heterogeneity was studied with viralpopulations that had been treated with drugs for 48 h, andsome of these populations were approaching extinction atthe time of RNA extraction. It was therefore essential tomake sure that cloning and sequencing were performedreliably from individual clones that reflected the authenticRNA population rather than repeating the analysis withPCR products amplified from the same original viral RNAmolecules. When analyzing n random amplification prod-ucts from an RT-PCR performed with a sample containingD RNA molecules (D can be regarded as the last dilutionfound positive in PCR), the expected number of indepen-dent clones (E) is given by E � D(1 � ((D � 1)/D)n [theformula is modified from Airaksinen and Hovi, 1998]. Inthe present work, 6 RT-PCR reactions were performed fromeach RNA sample. In addition to 3 independent reactionswith undiluted RNA, reactions were also performed usingsamples diluted 1:10, 1:20, and 1:40. In cases where the1:40 dilution was positive in RT-PCR, one of the RT-PCRobtained with undiluted RNA was used as the material forcloning. Under these conditions, sequencing 10 clonesshould give at least 8.9 independent sequences (D � 40, n� 10, E � 8.9). When the 1:40 dilution was not positive but1:20 or 1:10 were, all 3 RT-PCR from undiluted RNA wereused in cloning, and the total of 10 sequences (4 � 3 � 3)were derived from the 3 parallel cloning procedures (E �9.4 or E � 8.9 for dilutions 1:20 and 1:10, respectively).The results demonstrate that these precautions were suffi-cient, since only 3 different mutations, out of the 97 scored,were found twice in a single population. For determinationof the mutation frequencies, generally 10 clones were se-quenced of each of the seven persistent FMDV populationssubjected to various antiviral treatments. The exact numbersof clones analyzed in each experiment are given underResults and Table 1.
Acknowledgments
We are indebted to J.C. de la Torre for supplying the Ribnucleoside, and to M. Davila for expert technical assistance.We thank M. Alonso for helpful assistance with mass spec-troscopy analyses. This research (A.A.) was supported by aMarie Curie Fellowship of the European Community pro-gramme “Quality of Life and Management of Living Re-sources” under Contract QLK2-CT-1999-51462, and bygrant, BMC 2001-1823-CO2-01 from MCyT and 08.2/0015/2001 from CAM. N.P. was supported by a predoctoral
347A. Airaksinen et al. / Virology 311 (2003) 339–349

fellowship from MCyT (Spain). An institutional grant ofFundacion Ramon Areces is also acknowledged.
References
Airaksinen, A., Hovi, T., 1998. Modified base compositions at degeneratepositions of a mutagenic oligonucleotide enhance randomness in site-saturation mutagenesis. Nucleic Acids Res. 26, 576–581.
Anderson, J.F., Rahal, J.J., 2002. Efficacy of interferon alpha-2b andribavirin against West Nile virus in vitro. Emerg. Infect. Dis. 8, 107–108.
Arias, A., Lazaro, E., Escarmıs, C., Domingo, E., 2001. Molecular inter-mediates of fitness gain of an RNA virus: characterization of a mutantspectrum by biological and molecular cloning. J. Gen. Virol. 82, 1049–1060.
Balzarini, J., Lee, C.K., Herdewijn, P., De Clercq, E., 1991. Mechanism ofthe potentiating effect of ribavirin on the activity of 2�,3�-dideoxyi-nosine against human immunodeficiency virus. J. Biol. Chem. 266,21509–21514.
Connor, E., Morrison, S., Lane, J., Oleske, J., Sonke, R.L., Connor, J.,1993. Safety, tolerance, and pharmacokinetics of systemic ribavirin inchildren with human immunodeficiency virus infection. Antimicrob.Agents Chemother. 37, 532–539.
Contreras, A.M., Hiasa, Y., He, W., Terella, A., Schmidt, E.V., Chung,R.T., 2002. Viral RNA mutations are region specific and increased byribavirin in a full-length hepatitis C virus replication system. J. Virol.76, 8505–8517.
Crotty, S., Cameron, C.E., Andino, R., 2001. RNA virus error catastrophe:direct molecular test by using ribavirin. Proc. Natl. Acad. Sci. USA 98,6895–6900.
Crotty, S., Maag, D., Arnold, J.J., Zhong, W., Lau, J.Y., Hong, Z., Andino,R., Cameron, C.E., 2000. The broad-spectrum antiviral ribonucleosideribavirin is an RNA virus mutagen. Nature Med. 6, 1375–1379.
Crumpacker, C., Bubley, G., Lucey, D., Hussey, S., Connor, J., 1986.Ribavirin enters cerebrospinal fluid. Lancet 2, 45–46.
De la Torre, J.C., Alarcon, B., Martınez-Salas, E., Carrasco, L., Domingo,E., 1987. Ribavirin cures cells of a persistent infection with foot-and-mouth disease virus in vitro. J. Virol. 61, 233–235.
De la Torre, J.C., Davila, M., Sobrino, F., Ortin, J., Domingo, E., 1985.Establishment of cell lines persistently infected with foot-and-mouthdisease virus. Virology 145, 24–35.
De la Torre, J.C., Martınez-Salas, E., Dıez, J., Domingo, E., 1989. Exten-sive cell heterogeneity during persistent infection with foot-and-mouthdisease virus. J. Virol. 63, 59–63.
De la Torre, J.C., Martınez-Salas, E., Dıez, J., Villaverde, A., Gebauer, F.,Rocha, E., Davila, M., Domingo, E., 1988. Coevolution of cells andviruses in a persistent infection of foot-and-mouth disease virus in cellculture. J. Virol. 62, 2050–2058.
Dıez, J., Davila, M., Escarmıs, C., Mateu, M.G., Domınguez, J., Perez, J.J.,Giralt, E., Melero, J.A., Domingo, E., 1990. Unique amino acid sub-stitutions in the capsid proteins of foot-and-mouth disease virus from apersistent infection in cell culture. J. Virol. 64, 5519–5528.
Drake, J.W., Holland, J.J., 1999. Mutation rates among RNA viruses. Proc.Natl. Acad. Sci. USA 96, 13910–13913.
Eigen, M., 2000. Natural selection: a phase transition? Biophys. Chem. 85,101–123.
Eigen, M., 2002. Error catastrophe and antiviral strategy. Proc. Natl. Acad.Sci. USA 99, 13374–13376.
Eigen, M., Biebricher, C.K., 1988. Sequence space and quasispecies dis-tribution, in: Domingo, E., Ahlquist, P., Holland, J.J. (Eds.), RNAGenetics, Vol 3. CRC Press, Boca Raton, FL, pp. 211–245.
Eriksson, B., Helgstrand, E., Johansson, N.G., Larsson, A., Misiorny, A.,Noren, J.O., Philipson, L., Stenberg, K., Stening, G., Stridh, S., Oberg,B., 1977. Inhibition of influenza virus ribonucleic acid polymerase byribavirin triphosphate. Antimicrob. Agents Chemother. 11, 946–951.
Escarmıs, C., Davila, M., Domingo, E., 1999. Multiple molecular pathwaysfor fitness recovery of an RNA virus debilitated by operation of Mul-ler’s ratchet. J. Mol. Biol. 285, 495–505.
Fernandez-Larsson, R., O’Connell, K., Koumans, E., Patterson, J.L., 1989.Molecular analysis of the inhibitory effect of phosphorylated ribavirinon the vesicular stomatitis virus in vitro polymerase reaction. Antimi-crob. Agents Chemother. 33, 1668–1673.
Franklin, T.J., Cook, J.M., 1969. The inhibition of nucleic acid synthesis bymycophenolic acid. Biochem. J. 113, 515–524.
Gebauer, F., De la Torre, J.C., Gomes, I., Mateu, M.G., Barahona, H.,Tiraboschi, B., Bergmann, I., De Mello, P.A., Domingo, E., 1988.Rapid selection of genetic and antigenic variants of foot-and-mouthdisease virus during persistence in cattle. J. Virol. 62, 2041–2049.
Gerotto, M., Sullivan, D.G., Polyak, S.J., Chemello, L., Cavalletto, L.,Pontisso, P., Alberti, A., Gretch, D.R., 1999. Effect of retreatment withinterferon alone or interferon plus ribavirin on hepatitis C virus qua-sispecies diversification in nonresponder patients with chronic hepatitisC. J. Virol. 73, 7241–7247.
Gilbert, B.E., Knight, V., 1986. Biochemistry and clinical applications ofribavirin. Antimicrob. Agents Chemother. 30, 201–205.
Goswami, B.B., Borek, E., Sharma, O.K., Fujitaki, J., Smith, R.A., 1979.The broad spectrum antiviral agent ribavirin inhibits capping ofmRNA. Biochem. Biophys. Res. Commun. 89, 830–836.
Holland, J.J., Domingo, E., De La Torre, J.C., Steinhauer, D.A., 1990.Mutation frequencies at defined single codon sites in vesicular stoma-titis virus and poliovirus can be increased only slightly by chemicalmutagenesis. J. Virol. 64, 3960–3962.
Hultgren, C., Milich, D.R., Weiland, O., Sallberg, M., 1998. The antiviralcompound ribavirin modulates the T helper (Th) 1/Th2 subset balancein hepatitis B and C virus-specific immune responses. J. Gen. Virol. 79,2381–2391.
Jang, S.K., Krausslich, H.G., Nicklin, M.J., Duke, G.M., Palmenberg,A.C., Wimmer, E., 1988. A segment of the 5� nontranslated region ofencephalomyocarditis virus RNA directs internal entry of ribosomesduring in vitro translation. J. Virol. 62, 2636–2643.
Jordan, I., Briese, T., Averett, D.R., Lipkin, W.I., 1999. Inhibition of Bornadisease virus replication by ribavirin. J. Virol. 73, 7903–7906.
Jordan, I., Briese, T., Fischer, N., Lau, J.Y., Lipkin, W.I., 2000. Ribavirininhibits West Nile virus replication and cytopathic effect in neural cells.J. Infect. Dis. 182, 1214–1217.
Kerr, S.J., 1987. Ribavirin induced differentiation of murine erythroleuke-mia cells. Mol. Cell. Biochem. 77, 187–194.
Lanford, R.E., Chavez, D., Guerra, B., Lau, J.Y., Hong, Z., Brasky, K.M.,Beames, B., 2001. Ribavirin induces error-prone replication of GBvirus b in primary tamarin hepatocytes. J. Virol. 75, 8074–8081.
Lea, S., Hernandez, J., Blakemore, W., Brocchi, E., Curry, S., Domingo,E., Fry, E., Abu-Ghazaleh, R., King, A., Newman, J., et al., 1994. Thestructure and antigenicity of a type C foot-and-mouth disease virus.Structure 2, 123–139.
Lee, H.J., Pawlak, K., Nguyen, B.T., Robins, R.K., Sadee, W., 1985.Biochemical differences among four inosinate dehydrogenase inhibi-tors, mycophenolic acid, ribavirin, tiazofurin, and selenazofurin, stud-ied in mouse lymphoma cell culture. Cancer Res. 45, 5512–5520.
Lowe, J.K., Brox, L., Henderson, J.F., 1977. Consequences of inhibition ofguanine nucleotide synthesis by mycophenolic acid and virazole. Can-cer Res. 37, 736–743.
Maag, D., Castro, C., Hong, Z., Cameron, C.E., 2001. Hepatitis C virusRNA-dependent RNA polymerase (NS5B) as a mediator of the anti-viral activity of ribavirin. J. Biol. Chem. 276, 46094–46098.
Malinoski, F., Stollar, V., 1981. Inhibitors of IMP dehydrogenase preventsindbis virus replication and reduce GTP levels in Aedes albopictuscells. Virology 110, 281–289.
Martın-Hernandez, A.M., Carrillo, E.C., Sevilla, N., Domingo, E., 1994.Rapid cell variation can determine the establishment of a persistentviral infection. Proc. Natl. Acad. Acad. Sci. USA 91, 3705–3709.
Mateu, M.G., Hernandez, J., Martınez, M.A., Feigelstock, D., Lea, S.,Perez, J.J., Giralt, E., Stuart, D., Palma, E.L., Domingo, E., 1994.
348 A. Airaksinen et al. / Virology 311 (2003) 339–349

Antigenic heterogeneity of a foot-and-mouth disease virus serotype inthe field is mediated by very limited sequence variation at severalantigenic sites. J. Virol. 68, 1407–1417.
Ning, Q., Brown, D., Parodo, J., Cattral, M., Gorczynski, R., Cole, E.,Fung, L., Ding, J.W., Liu, M.F., Rotstein, O., Phillips, M.J., Levy, G.,1998. Ribavirin inhibits viral-induced macrophage production of TNF,IL-1, the procoagulant fg12 prothrombinase and preserves Th1 cyto-kine production but inhibits Th2 cytokine response. J. Immunol. 16,3487–3493.
Ogle, J.W., Toltzis, P., Parker, W.D., Alvarez, N., Mcintosh, K., Levin,M.J., Lauer, B.A., 1989. Oral ribavirin therapy for subacute sclerosingpanencephalitis. J. Infect. Dis. 159, 748–750.
Pariente, N., Sierra, S., Lowenstein, P.R., Domingo, E., 2001. Efficientvirus extinction by combinations of a mutagen and antiviral inhibitors.J. Virol. 75, 9723–9730.
Pelletier, J., Sonenberg, N., 1988. Internal initiation of translation ofeukaryotic mRNA directed by a sequence derived from poliovirusRNA. Nature 334, 320–325.
Pogolotti Jr., A.L., Santi, D.V., 1982. High-pressure liquid chromatog-raphy–ultraviolet analysis of intracellular nucleotides. Anal. Biochem.126, 335–345.
Querenghi, F., Yu, Q., Billaud, G., Maertens, G., Trepo, C., Zoulim, F., 2001.Evolution of hepatitis C virus genome in chronically infected patientsreceiving ribavirin monotherapy. J. Viral. Hepatol. 8, 120–131.
Racaniello, V. R., 2001. Picornaviridae: the viruses and their replication,fourth ed., in: (H.P.M. Knipe, D.M., Griffin D.E., Lamb, R.A., Martin,M.A., Roizman, B,, Straus, S.E., Eds.). Fields Virology. LippincottWilliams & Wilkins, Baltimore.
Rankin Jr., J.T., Eppes, S.B., Antczak, J.B., Joklik, W.K., 1989. Studies onthe mechanism of the antiviral activity of ribavirin against reovirus.Virology 168, 147–158.
Ruız-Jarabo, C. M., Ly, C., Domingo, E., De La Torre, J. C., 2003. Lethalmutagenesis of the prototypic arenavirus lymphocytic choriomeningitisvirus (LCMV). Virology, in press.
Salt, J.S., 1993. The carrier state in foot and mouth disease—an immuno-logical review. Br. Vet. J. 149, 207–223.
Scheidel, L.M., Stollar, V., 1991. Mutations that confer resistance tomycophenolic acid and ribavirin on Sindbis virus map to the nonstruc-tural protein nsP1. Virology 181, 490–499.
Semler, B.L., Wimmer, E., 2002. Molecular Biology of Picornaviruses.Ann. Soc. Microbiol, Washington, DC, Eds..
Severson, W.E., Schmaljohn, C.S., Javadian, A., Jonsson, C.B., 2003.Ribavirin causes error catastrophe during Hantaan virus replication.J. Virol. 77, 481–488.
Sidwell, R.W., Huffman, J.H., Khare, G.P., Allen, L.B., Witkowski, J.T.,Robins, R.K., 1972. Broad-spectrum antiviral activity of Virazole:1-beta-D-ribofuranosyl- 1,2,4-triazole-3-carboxamide. Science 177,705–706.
Sierra, S., Davila, M., Lowenstein, P.R., Domingo, E., 2000. Response offoot-and-mouth disease virus to increased mutagenesis: influence ofviral load and fitness in loss of infectivity. J. Virol. 74, 8316–8323.
Sintchak, M.D., Fleming, M.A., Futer, O., Raybuck, S.A., Chambers, S.P.,Caron, P.R., Murcko, M.A., Wilson, K.P., 1996. Structure and mech-anism of inosine monophosphate dehydrogenase in complex with theimmunosuppressant mycophenolic acid. Cell 85, 921–930.
Smee, D.F., Matthews, T.R., 1986. Metabolism of ribavirin in respiratorysyncytial virus-infected and uninfected cells. Antimicrob. Agents Che-mother. 30, 117–121.
Snell, N.J., 2001. Ribavirin—current status of a broad spectrum antiviralagent. Expert Opin. Pharmacother. 2, 1317–1324.
Sobrino, F., Davila, M., Ortin, J., Domingo, E., 1983. Multiple geneticvariants arise in the course of replication of foot-and-mouth diseasevirus in cell culture. Virology 128, 310–318.
Sobrino, F., Saiz, M., Jimenez-Clavero, M.A., Nunez, J.I., Rosas, M.F.,Baranowski, E., Ley, V., 2001. Foot-and-mouth disease virus: a longknown virus, but a current threat. Vet. Res. 32, 1–30.
Sookoian, S., Castano, G., Frider, B., Cello, J., Campos, R., Flichman, D.,2001. Combined therapy with interferon and ribavirin in chronic hep-atitis C does not affect serum quasispecies diversity. Dig. Dis. Sci. 46,1067–1071.
Streeter, D.G., Witkowski, J.T., Khare, G.P., Sidwell, R.W., Bauer, R.J.,Robins, R.K., Simon, L.N., 1973. Mechanism of action of 1-�-D-ribofuranosyl-1,2,4-triazole-3-carboxamide (Virazole), a new broad-spectrum antiviral agent. Proc. Natl. Acad. Sci. USA 70, 1174–1178.
Swetina, J., Schuster, P., 1982. Self-replication with errors. A model forpolynucleotide replication. Biophys. Chem. 16, 329–345.
Toltzis, P., O’Connell, K., Patterson, J.L., 1988. Effect of phosphorylatedribavirin on vesicular stomatitis virus transcription. Antimicrob. AgentsChemother. 32, 492–497.
Traut, T.W., 1994. Physiological concentrations of purines and pyrimi-dines. Mol. Cell. Biochem. 140, 1–22.
Van Bekkum, J.G., Frenke, H.S., Frederiks, H.H.J., Frenkel, S., 1959.Observations on the carrier state of cattle exposed to foot-and-mouthdisease virus. Tijdschr. Diergeneeskd. 84, 1159–1164.
Volkenstein, M.V., 1994. Physical Approaches to Biological Evolution.Springer-Verlag, Berlin.
Wray, S.K., Gilbert, B.E., Knight, V., 1985a. Effect of ribavirin triphos-phate on primer generation and elongation during influenza virus tran-scription in vitro. Antiviral Res. 5, 39–48.
Wray, S.K., Gilbert, B.E., Noall, M.W., Knight, V., 1985b. Mode of actionof ribavirin: effect of nucleotide pool alterations on influenza virusribonucleoprotein synthesis. Antiviral Res. 5, 29–37.
Wray, S.K., Smith, R.H., Gilbert, B.E., Knight, V., 1986. Effects ofselenazofurin and ribavirin and their 5�-triphosphates on replicativefunctions of influenza A and B viruses. Antimicrob. Agents Chemother.29, 67–72.
Zimmerman, T.P., Deeprose, R.D., 1978. Metabolism of 5-amino-1-beta-D-ribofuranosylimidazole-4-carboxamide and related five-memberedheterocycles to 5�-triphosphates in human blood and L5178Y cells.Biochem. Pharmacol. 27, 709–716.
349A. Airaksinen et al. / Virology 311 (2003) 339–349




















