
THE UNIVTRSITY OF I4ANITOBA SUBMITTED TO THË FACULTY ...
214
THE UNIVTRSITY OF I4ANITOBA NEURAL CONTROL OF TRACHEAI. SMOOTH MUSCLE FROM CONTROL AND IMMUNOLOGICALLY SENSITIZËD DOGS BY J. M. BERGEN A THTSIS SUBMITTED TO THË FACULTY OF GRADUATË STUDIES IN PARTIAL FULFILMENT OF THE REQUTREMENTS I.OR THE DEGREE DOCTOR OF PHILOSOPHY ÜEPARTMINT OF PHYSIOLOGY WINNIPEG, MANITOBA FEBRUARY, 1983
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of THE UNIVTRSITY OF I4ANITOBA SUBMITTED TO THË FACULTY ...

THE UNIVTRSITY OF I4ANITOBA
NEURAL CONTROL OF TRACHEAI. SMOOTH MUSCLE
FROM CONTROL AND IMMUNOLOGICALLY SENSITIZËD DOGS
BY
J. M. BERGEN
A THTSIS
SUBMITTED TO THË FACULTY OF GRADUATË STUDIES
IN PARTIAL FULFILMENT OF THE REQUTREMENTS
I.OR THE DEGREE DOCTOR OF PHILOSOPHY
ÜEPARTMINT OF PHYSIOLOGY
WINNIPEG, MANITOBA
FEBRUARY, 1983

NEURAL CONTROL OF TRACHEAL SMOOTH MUSCLE
FROM CONTROL AND IMMUNOLOGICALLY SENSITIZED DOGS
BY
J.M. BERGEN
A thesis subnlitted to the Faculty of Graduate Studies ofthe university of Manitoba in partial fulfillment of the requirenrerrtsof the degree of
DOCTOR OF PHILOSOPHY
o t983
Pernrission has been grarìted to the L¡BRARy oF THE un-lvER-slrY oF MANITOBA to lend or seil copies of this trresis. tothe NATIONAL LIBRARy oF CANADA to microfilnr thisthesis a'd to lend or sell copies of the fihn, and UNIVERSITYMICROFILMS to publish an abstract of this thesis.
The author reserves other publicatio¡l rights, and neither the
thesis nor exte¡rsive extracts fronl it may be printed or other-wise reproduced without the author's writteu permission.

ACKNOl,'JL EDGEI4ENTS
I wish to express thanks to my supervisor Dr. E.A. Kroeger for
the opportunity to work in his laboratory, for his time and patience
and mostly for his constructive critic'ism and encouragement in the
development of this thesis. I feel I have learned a great dea'l from
him both with'in and outside the bounds of science and for this I
am grateful .
I wish to thank those co-workers in the laboratory who have also
become friends over the years; Blanka Kucera, Richard þlitchell,
David Tesarowski and Lou Antonissen.
My thanks aÈe also extended to my examin'ing committee for their
perseverance in reading and criticizìng this manuscript; Drs. D. Bose,
N.L. Stephens, [''¡.S. Dhalla and J. Marshall.
Over the years many friends and fam'ily have contributed greatly
to support and enrich life outs'ide of my work and to these persons
whom I will not name lest I forget one at this moment I express a sincere
thanks.
Lastly and most important I wish to thank my wife Rita for her support
and patience during these years.

ABSÏRACT

-iABSTRACT
The precise role of airway smooth muscle in pulmonary physiology is
not known; however its regulation of airway calibre and therefore re-
sistance to airflow Ís appreciated. The regu'lation of its contractile
state Ís dependent largely on neural and pharmacological mechanisms which
ard poorly understood. Our ignorance of the interactÍons of regulation
mechanisms is particu'lar1y acute as it relates to disease states such as
asthma in which the airway smooth muscle may hyperreact to certain
stÍmul i .
The present studies were designed to assess the functional contri-
butíons of endogenous neural el ements, thei r prej unctional control
mechanÍsms and the relevant post-junctional receptors in canine tracheal
smooth muscle (TSM). The specific objectives were to: 1) Examine adreno-
ceptor mediated responses in isolated TSM, and the relationshÍp of these
responses to active tone, 2l compare adrenoceptor mediated responses of
TSM taken from dogs immunologica'lly sensitized to ovallbumin (04) with a
control population, 3) examine the effect of histamine and 5-hydroxytryp-
tamine (5-HT) on isometric tensÍon development by muscles from control
and 0A-sensitized dogs, and 4) determine the effect of a variety of
stimulatory conditions on overflow of acety'lcholine and norepinephrine
from non electricalìy stimulated endogenous cholinergic and adrenergic
nerves respectively. The effect of propranolol on acetylcholine overflow
uJas al so studied. Isometric tension measurements demonstrated
physíoìogica'l importance of neuromodulation'

- t1
The effect of tone on responses of canine tracheal smooth muscle
(TSM) to norepinephrine (NE) was studied to elucidate the role of
sympathetic innervation and adrenoceptors in the control of the aírways.
¡ectrical field stimulation produced contraction of TSM in vitro which
1,,,as augmented by eserine, depressed by phentolamine and almost eliminated
by TTX or hyoscyamíne. Resting TSM did not contract in response to NE
(1É - lÉM)) normal'ly or in the presence of propranolo'l (lÉM).
The addition of NE (1É - tO-6N) at the p'lateaux of contractions
produced by K+ Q2.8 mM), histamÍne (lÉM) or acetylcholÍne (RCh' 5
x tO-Sm) produced a further phentolami ne-sensi tÍve contraction which
was potentiated by beta-adrenoceptor blockade with propranolol (lÉM).
The addi tion of tyrarni ne (1É or lÉM) at the pì ateau of contrac-
tion produced by K+ (22.8 mM) produced a further contraction which was
potentiated by propranolol (10-5M) and reduced by phentolamine
(10-5M). Following cold storage (4"C for 3 days) tne response to
tyramine was inhibited while responses to NE remaÍned. While NE in the
presence of active tone caused a further increase in tension at low con-
centration (1ù€ to t0{N), a propranolol-sensitive relaxant response
was el ici ted at higher concentrations (tO-5 and 1ÉM) . Maximum
contractile responses to NE in the absence or presence of beta-blockade
v,rere dependent on the tone of the muscl e. These f i ndi ngs suggest a
functional adrenergic innervation of canine TSM and the presence of
al pha- and betaadrenoceptors which rnedi ate contractil e and rel axant
responses respectively. The functiona'l dominance of either of the latter
is related to the active tone in the muscle and the concentration of Nt.

- iii -
The bi phasic responses, i nvol vi ng al pha-adrenoceptor medi ated
contraction and beta-adrenoceptor medÍated relaxation in 0A sensitized
and control TSM were both qua'l itatively and quantÍtatively simil ar.
Unlike control TSM, sensítized tissue at basal tone contracted when
exposed to tyramine tO-4N or the combination of propranolol (lÉM)
and norepi nephrine (lÉM) . Beta-bl ockade wi th propranol ol (1ÉM)
enhanced the tyramine-stirnulated contraction whíle phentolamine (1ÉM)
abolíshed it. These studies attribute the increased alpha-adrenoceptor-
mediated responsiveness of sensitized TSM to an increased resting basal
t,one although this was not tneasured directly.
The observation that the histamine (1Û-sM)-stimulated contraction
of TSM is about 507, atropine-sensitive suggests a possible actÍon of his-
tamine in promoting acetylcholine release. The addition of eserÍne
(1ù-6M, an anticholinesterase) prior to histamine enhanced the contrac-
tion to 703% of the initial histamine-induced contraction. The further
additíon of hyoscyamine reduced the contraction to 4L% of the initial
histarnine contraction" A similar pattern although not as pronounced was
noted in the atropine-sensitive (107,) component of the 5-HT tO{m con-
traction and enhancement Q45% of initial ) by eserine tÉN. Eserine
(tO{N) was al so observed to enhance ç+ (23mM) and el ectrical ly stím-
ulated isometric tension development an effect blocked by hyoscyamine.
The addition of eserine to unstimulated muscles was without effect. The
addition of phentolamine (1ÉM) reduced isometric tension of both
histamine (1ÉM) and 5-HT (tO-6m)-induced contractions suggesting a
rol e for these substances i n the neuromodul ation of norepi nephrine

- iv -
release from adrenergic nerves in addition to their dÍrect contractile
effect on TSM
A simil ar protoco'l for hi stami ne and 5-HT i n 0A-sensi tized TSM
demonstrated a qual i tative and quanti tative simil arity to control
ti ssue s .
Acetylcholine (1ÉM)-contracted TSM was partia'lly inhibited (57%
of initial ) by phentolamine (1ÉM) and further addition of atropine
(10-7) abolished the contraction. Eserine tO{U enhanced isometric
tension developed fo'l1owíng acetylcholine or K+ (23 mM) stimulation.
These studies suggest a role of 5-HT, hístamine and acetylcholine in
prejunctional neuromodulation on both cholinergic and adrenergic nerves
in TSM. Further st,udies utilize TSM preincubated in either 3H-nore-
pinephrine or 14C-choline to demonstrate overflow ín the unstimulated
preparation" 3H-norepi nephrine overfl ow was increased by el ectrical
fiel d stimulation (EFS, 15V, 60 Hz, AC) , phentoìamine (1ÉM) , 23mM
6+ nonepi nephri ne (lÉM) and acetyl chol i ne tO-6m i n cocai ne
(1ÉM)-pretreated muscles. When cocaine was absent onìy el ectrical and
K+ (Z3mM) stimul ation produced measureable increases in 3H-Uf over-
fl ow Tyrami ne (1ÉM) b,as wi thout ef fect on 3U-rue overfl ow i n the
presence or àbsence of cocaíne. 14C-choline overflow was increased by
el ectrical fi el d stimul ati on and K+ (29 mM) . l,{hen eseri ne (1ÉM) was
present throughout the experiment hi stami ne (1ÉM) , 5-HT (1ÉM) ,
and norepi nephri ne (1r7M) al so enhanced l40-chol i ne overfl ow. The
further addition of tetrodotoxin (TTX, tO-6m) abolished the effect of
histamine and 5-HT. Norepinephrine (lÉM) and propranolo'l (lÉM)

v
had a signÍficant inhibitory action on l40-choline overflow appl ied
in the presence of eserine tO{U and tetrodotoxin (tO{U).
These st,udi es suggest the presence of prej unctional a'l pha and
beta-adrenoceptors on both adrenergic and cholinergic nerves, prejunc-
tional muscarinic receptors on adrenergíc nerves and 5-HT and histarnine
receptors at the level of the ganglia on cholinergic nerves'

-vi
TABLE ()F CONTENTS
LIST OF FIGURTS
LIST OF TABLTS
REVITI^J OF LITERATUREFunction of airwaY smooth muscleTracheal smooth muscleStructure, ultrastructure and electrophysiology
of tracheal smooth muscleNeural control of air$Jay smooth muscle
ParasYmPatheti c sYstemEffects of ACh on the postjunctional membrane
Sympathetic sYstemPotass i um
Preiunctional Neuromodul ationAdrenergic transmitter release
Airway Smooth Muscl e HyperreactivityI Abnormal Mediator Stimuli
A) Extrinsic Factors
B)
A'l I ergyIrritant receptor reflex
trinsic FactorsChol Í nergic 0veractivi tYAlteratÍons ín mediator productionand/or rel ease
3) SymPathetic sYstem4) Nonadrenergic sYstem
II Abnormal Responses of Smooth Muscle1) Changes in muscle mass2l Changes at recePtor I evel3 ) Changes at i ntracel I ul ar I evel
STATEI4TNT OF THE PROBLEM
METHODS' Sensitized canine TSM
Di ssecti onIsometric tension measurementsStatistics usedRadio labelled transmitter efflux experiments
t3Hl - Norepinephrine effluxtl4Cl - Acetycholine efflux
High Performance 1Íquid chromatography ofnorepinephrine efflux
Page
X
xii
B
1111151516?325252528
30
313334
4243434447475051
1
35
1)2)In1)z)
353639
40
51

- vii -
TABLT 0F CONTENTS (Contínued)
RESULTS
A) Electrical field stimulatÍon of canÍneTSM, i n_ld_!ro
Effect of tone on adrenoceptor mediatedresponses in canine TSM
Adrenoceptor mediated responses incanÍne TSM sensítized to ovalbumin (04)
Page
56
96
TT2
L?8
130
134
136
145
148
L72
B)
c)
D) Effect,s of atropÍne and phentolamine oncontractiìe responses produced byhistamÍne and serotonin in control andoval bumi n-sensitized cani ne TSM
E) Effects of eserine, propranolo'l andphentolamine on the contracti'le responsesto histamine, 5-hyroxy-tryptamine,acetylcholine and potassium in canine TSM
in vitroF) Flcï,õFwhich may modulate [3H] -
norepinephrine overflow in canine TSM
G) Neuromodulating influence of severalfators on the cholinergic nervous systemin canine TSM Ín vitro
DISCUSSIONA) ElectrÍcal fÍeld stimulation and isometric
tensi onB) Effect of tone on adrenoceptor mediated
responsesC) Adrenoceptor medÍated responses in
sensÍtized canine TSM
D) Actions of histamine and serotonin on
cani ne TSM
E) Autoregulation of transmÍtter overflow
RTFERENCËS
AP PEND IX
61
76
B4
95

- viii -
LIST OF FIGURES
FI GURE
1. Effect of eserine and hyoscyamine on isometric tensiondeveloped by TSM following electrical field stimulation(EFS).
2. Effect of phentolamine and tetrodotoxin on isometrictension devel oped by TSM fol'lowi ng EFS.
3. Effect of propranolol and phentolamine on isometrictension developed by TSM following EFS.
4. Effect of norepinephrÍne and propranolol on isometrictension devel oped by TSM fol'l owi ng EFS.
5. Effect of norepinephrine on isometric tension developedwith 22.8 mM potassium solution Ín the presence andabsence of phentolamine.
6. Effect of atropine on the response of TSM tonorepinephrine in the presence of tone established withK+(22.8 mM).
7. Effect of propranolol pretreatment on the response ofK-contracted TSM to norepinephrine.
8. Effect of atropine and tyrarnitle on the K-contracture ofTSM.
9. Representative mechanogram showing the effect ofpropranolol and atropine on the response of TSM tonoräpÍnephrine in thä presence of 48.2 mM K+.
10. Responses of atropinized TSM to norepinephrine atvarÍous IK]o. Effect of propranolol on theAl pha-adrenoceptor mediated contractíon.
11. Effect of norepinephrine (NE) on isometric tensiondeveloped with acetylchol ine.
L2. Effect of NE on TSM pretreated wi th propranol ol and
contracted wi th acetyl chol Í ne.
13. Effect of NE on TSM contracted isometrical'ly withhi stami ne.
L4. Effect of NE on TSM contracted isometrical'ly withhistamine and pretreated with propranolol.
Page
57
58
59
60
62
63
65
66
68
69
72
73
74
75

-ix-LIST 0F FIGURES (Continued)
F I GURE
15. Effect of NE on isometric tension deveìoped by TSM from
control and sensitized TSNI, following induction ofactive tone.
16. Effect of atropine on the response of sensitized and
control TSM to norePinePhrine'
17. Effect of NE on control and sensitized TSM contractedi sometrical ly wi th 23 mM K+ and pretreated wi thphentol ami ne.
18. Effect of atropine and tyramine on control and
sensitized TSM contracted isolnetrically with 23 mM
ç+.
19. Effect of tyranine on control and sensitized TSM
pretreate¿ wiür atropine and contracted isometricalìyi,,ritn ZA mM K+.
20. Effect of tyramine on TSM frorn control and 0A
sensi ti zed dogs.
2L. Effect of cumulative addition of norepinephrinetyrarnine and phentolamÌne on 0A-sensitized and controlTSM.
22, Effect of sequentia'l exposure to phentolamine and
atropine on isometric tension developed by
hi stämine-treated cani ne TSM'
2g" Effect of atropÍne and phentolamine on ísometrictension develoþed by histamine-treated TSM'
24. Effect of atropine and phentoìamine on isometrictensÍon develoþed by 5-hydroxytryptamine treated TSM.
25. Effect of phentolamine and atropine on isometrictension deieloped by 5-hydroxytryptamine treated TSM.
26. Effect of phentolamine and atropine on isometrictension developed by control and 0A sensit,ized TSM
contracted isometrica'l 1y wi th histamine'
27. Effect of atropine and phentolamine on isornetrictension develoþed by control and 0A sensitized TSM
contracted wí th 5-hydroxytryptami ne'
Page
77
7f3
79
BO
B1
B3
85
B6
8B
89
90
91
92

-x-LIST 0F FIGURES (Continued)
F I GURI Page
105
106
107
108
109
110
111
113
114
116
LL7
lBB
119
L21.
28. Effect of time on active i sometric tension and
¡3¡l-norepinephrine overflow from TSM.
29. Effect of histarnÍne and electrical field stimulation on
isometríc tension and ¡3¡11-norepinephrine overflowfrom TSM.
30. Effect of 23 mM K+ on isometric tension and
t3Hl-norepi nephri ne overfl ow frorn TSM.
31. Effect of phentolamine on isometric tension and
[3H]-norepinephrine overfl ow from TSM.
32, Effect of norepinephrÍne on isometric tension and
[3H]-norepi nephrine overfl ow from TStvl.
33. Effect of acetylcholine on isometric tension and
[3H]-norepinephrine overfl ow from TSM.
34. Effect of 5-hydroxytryptamine on isometric tension and
¡3¡11-norepi nephrine overfl ow frorn TSM.
35. Summary of effect of various experimental condÍtions on
the efllux of radioactivity from JH-Norepinephrineequibrated TSM.
36. Effect of tírne on active tension and 14C - overfl ow
from TSM.
37. Effect of histamine and electrical field stimulation on
isometric tension and 14C overflow from TSM'
38. Ëffect of 5-hydroxytryplamíne and potassium solution on
isometric tension and r4C - overflow from TSM.
39. Effect of norepinephripç (1ù-7N and lÉM) on
isometric tension and r4C - overflow from TSM'
40. Effect of histamine on 14C - overflow from TSM inthe presence of eserine or eserine plus tetrodotoxin.
41. Effect of S-hydroytryptarnine on 14C - overflow fromTSM in the presence of eserÍne or eseríne pìustetrodotoxi n.
42. Effect of atropine and acetyìcholine on 14C -overflow from TSM in the presence of eserine pìustetrodotoxi n.
122

-xiLIST 0F FIGURES (Contínued)
F I GURE
43. Effect of norepinephrine (10-7u) on 14c -overflow Ín the presence of eserÍne and effect oftyrarnine on L4C - overflow frorn TSM in thepresence of eserine ¡rlus tetrodotoxin.
44. Ef fect of norepi nephri ne (10-4tvt) on 14C
overflow from TSM in the presence of eserine or thepresence of eserine pìus tetrodotoxin"
45. Effect of phentolamine and propranolol on 14C -overflow frotn TSM Ìn the presence of eserine pìustetrodotoxi n.
46. Model of innervation of canine tracheal smooth muscle.
Page
L?3
L24
1.25
L?7

- xii -
LIST OF TABLES
TABLE
Page
46
54
70
93
94
97
99
100
101
102
L72
A Drugs used in exPeriments
B tffect of electrical field stimulation on norepinephrineand epinephrine overflow from TSM
1. Effect of cumulative dose addition of norepinephrine(10-8 to 10-4M) on TSM isometricalìy contractedfollowing exposure to various potassium concentrationshistamine and acetylchol ine.
2. Effect of cumulative dose addition of norepinephrine(10-B to 10-4M) on TSM contractetl wìth exposureto 48 mM K+ and the presence of atropine.
3. Effect of atropine on 5-hydroxytryptamine contractedova'lbumin sensitized TSM pretreated with phentolamine.
4. Effect of eserine and propranolol on TSM isometrical]ycontracted following eiposure to 5-hydroxytryptamine.
5. Effect of eserine and hyoscyamine on TSM isometrica'l]ycontracted fol'lowing exposure to histamine.
6. Effect of histamine on isometric tension in TSM
depolarized with 127 mM K+ and pretreated withatropine, proprano'lol and phentolamine.
7. Effect of eserine and hyoscyamine on isometric tensiondeveloped by TSM fol'lowing exposure to acetylcholine.
B. Effect of phenotolamine and atropine on isometric tensiondeveloped by TSM following exposure to acetylcholine'
9. Effect of eserine and phentolamine on isometric tensiondeveloped by TSM following exposure to 23 mM K+'
10. Ëffect of eserine and proprano'lo'l on isometríc tensiondeveloped by TSM followinô exposure to 23 mM K+'
APPENDIX
82
98

REVIEI'J 0F LITTRATURE

FUNCTION 0F AIRI^IAY SMOOTH MUSCLE
The physiological significance of smooth muscle in the tracheo-
bronchial tree is not well understood. Several theories have been put
forward to explain the possible roles for this muscle in normal "heaìthy"
states. Firstly, airway smooth muscìe, by shortening and therefore pro-
ducing various degrees of bronchoconstriction may divert ventilation to
areas of the lung which are well perfused so as to tnaintain homogeneity
of ventilation/perfusion (lllilliams, 1981). This does occur ín patients
with pulmonary ernbolism; quantitatively, however, this is not adequate to
be of physiologic benefit as measured by lung scanning (l,li1liams,1981).
The bronchoconstriction is a'lso short lived and is unlikeìy to provide
adequate adjustment to a chronic clinical condition.
A second theory put forward by Widdicombe and Nadel (1963) suggested
that airtvay smooth muscle tone may balance the amount of anatomical dead
space and airway resistance. Macklem et. al. (1969) support this theory
for larger airways (>Zmm insíde diameter) in dogs. It should be noted
however, that humans prefer to breathe through the nasal passage (high
resistance) as opposed to the lower resistance oral passage (Macklem and
Ënge1, lg75). An additional consideration is that the volume of anatomi-
cal dead space is small compared to tidal volume and reduction in dead
space occurs at the expense of great increases in airway resistance; it
is therefore difficult to appreciate the physiologic benefit of such an
al terati on .
A third theory relates to the effect of smooth muscle on airway
rigidity. 0l sen et. âl ., (1967 ) have demonstrated that increasing
bronchomotor tone provides cartilage-contaÌning airways with íncreased

-2-rigidíty and therefore less compressibí'lity. This increased stabí'lity
may prevent airway closure during forced expiratory flow with high intra-
thoracic pressures. This benefit is litnited however, in that smaller,
non-cartilage containing aÍrways rnay narrow the airway sufficient'ly to
reduce maximum expiratory flow rates (Macklem and Engel, 1975). lllilliams
(1981) has suggested a dual action for muscle in the aÍrways: one to
stabilize larger airways during forced expiration and another to regulate
maximum expiratory flow. When bronchodilating agents are administered
airway resistance continues to drop past the point, which permits maximum
expiratory flow and the further relaxation of the airway smooth muscle
allows some mechanical instability of the larger airways which then
become the flow-limiting segments.
Another hypothesis regarding the physioìogica'l significance of air-
way smooth muscle suggests that constriction of aírways may function to
pu11 open connecting alveoli and therefore maintain isotropic airways and
parenchyma with Ìdentical specific compliances (Macklem and Enge1, 1975).
The benefit of this effect ís likely to occur at 1ow lung volumes where
collapse of alveoli is more like'ly.
That, bronchoconstriction and increased aírway resistance occur in
disease states such as chronic bronchitis, hay fever and asthma is well
known (Hogg, l-981 ; Boushey, 1.981 ; 0rehek , êt. al . , 1977 ) .
Recently, McFaclden (1981) has demonstrated that excessive heat loss
from airways may induce bronchoconstriction in asthmatics and normals.
It has been suggested that the bronchoconstriction ¡neasured may be a
mechanism of thermoregulation especially in cold climates (l,lilliams,
1e8l ).

-3-Finaì 1y and perhaps mosE importantìy airway smooth muscle may
function to protect the alveoli from irritant inhaled partÍculate or
gaseous matter. Air$ray constriction may in itself mechanical'ly stimulate
irritant receptors in the mucosa and produce cough. Narrower airways
(depending on degree of resistance) will result in a higher linear velo-
city of aírflow and t,herefore a more efficient cough (Macklem and Mead,
1968). Bronchoconstriction further results in the formation of intra-
lumÍnal folds and with increased ve'locity the turbulence produced will
likely Ímpact particu'late matter in the larger airways where turbulence
ís greatest (l,lilliams,1981). Smaldone and Bergofsky (1976) have demon-
strated that bronchoconstriction in small airways would move the flow
'limiting segment towards the trachea. This should increase the velocity
of fìow Ìn the periphery and aid in the clearance of particu'late matter
from small aÍrways.
The mechanisms by which overactivity in airway smooth muscle occurs
are seen from the anatomic Ímmunol ogic, physiol ogic , phannacol ogic,
genetic, biochemical and pathologic perspectives. However, since it is
unlikely that the function of airway smooth muscle is to produce disease
states, a more complete understanding of normal physíological mechanÍsms
of thÍs muscle are required in order to evaluate the homeostatic signi-
fÍcance of changes observed in dísease states.
TRACHIAL SMOOTH MUSCLE
Previous studies have demonstrated di fferences in airway smooth
muscle bet'ween and within species in terms of neural control (Richardson,
L979; Richardson and Ferguson, 1980) receptor status (Fleisch, 1980) and

-4-site of response aìong the tracheobronchial tree (Fleisch and Calkins,
1976). Qther factors such as age dependence (Pandya" L977 ) multi unit
and single unit, behavior (Kroeger and Stephens, 1975) membrane potential
(Stephens and Kroeger,1980) resting tone (Bergen and Kroeger, L9B0; 0hno
et. â.|., 1981) and mechanical responsiveness (stephens, êt. â1., 1975;
Stephens and Kroeger, 1970) all contribute to the interpretation of air-
way smooth muscle function. Therefore in making comparisons between
species and between smooth muscles (e.g. vascular and bronchial) one must
keep in mind the various factors mentioned above. The focus of this
thesi s wi I I be prirnari'ly concerned wi t,h the neural and pharmaco'logical
regu'lation of canine tracheal smooth muscle (Stephens et. al., 1979).
Since it is al so possib'le to sensitize dogs ímmunological'ly to
allergen such that they respond to specific antigen challenge wíth bron-
choconstriction as measured by airway resistance in vivo (Kepron et.
â1., Hirshman and Downes,1981; Kessler et. â1., L973; Krell et. â1.,
1976) and smooth muscle contraction in _vitlg (Antonissen, et. ô1., L979;
Bergen and Kroeger, 1979) this provides a model analogous to allergic
asthma Ín humans (yu et. ôl ., L972). The observat,ion that the intact
human (Permutt, 1973; Nadel, L973) and dog (Stephens and Kroeger, 1980)
show similar responses of both large and srnall resistance airways to
various agonists and nerve stimulation suggests that cattine tracheal
smooth muscle is a useful model in studying normal and disease stat'es
such as asthma. This model provides a unique experimental tool in gaining
an understanding of alterations in smooth muscle receptor status and
innervation of the ìarge aírways with a'llergic disease and an apprecia-
tion of the actions of the wide variety of agents administered in the
treatment of asthma.

-5-RELATION OF STRUCTURE TO ELECTROPHYSIOLOGY OF TRACHEAL SMOOTH MUSCLE
The structure and ultrastructure of canine tracheal smooth muscle
has been revíewed by Stephens, êt. al . 1979), Stephens and Kroeger
( 1980 ), Daniel et. al . Ã979 ) and Richardson and Ferguson ( 1980 ). A
systematic description of this subiect is beyond the scope of the present
treatment and only those aspects r¡/nich rel ate to phenomena of interest
will be considered. Canine TSM shows multiunit propertíes in vitro in
that no spontaneous electrica'l or mechanical activity ís observed and a
myogenic response to stretch is absent (stephens and Kroeger, 1975). Gap
junctions and e'lectrotonic current spread (as measured by the space con-
stant) suggest good cel l -to-cel 'l coup'l i ng as tni ght be expected i n si ngl e
uni t smoo'bh muscl e. Si ngl e uni t srnooth muscl e general ly demonstrates
spontaneous electrica'l activity and myogenic responses (Boz1er, 1948;
Prosser, êt. âl ., 1960). In vivo studies (Akasaka, êt. â.l., 1975) have
shown the presence of spontaneous action potentials in canine and human
bronchial smooth muscìe, the frequency of which was increased in human
asthmatics during bronchospasm. Kroeger and Stephens (1975) and Bose and
Bose (L977\ were able to convert airv,Jay smooth muscle multiunit behaviour
in vitro to sing'le unit responses (spontaneous phasic activity) by the
use of a potassium channel blocker (tetraethylammonium) or glucose deple-
tion respectively. KirkpatrÍck (1975) demonstrated spontaneous phasic
mechanica'l activity wi th bovine tracheal smooth muscle in vitrq- in
response to histamine. These results suggest that while tracheal smooth
muscle appears to resemble multiunÍt type histologicaì'ly by its sparse
i nnervatíon (Daniel et. ôl ., 1979) and in vitro responses (absence of
myogenic activity; Stephens and Kroeger,1975) appropriate condítions may

-6-alter the response to that of single unit behaviour. Whether there ís a
change to spontaneous mechanical and electrical activity or perhaps an
increase of this activity in disease states such as asthrna Ín intact
humans is not known. Antonissen et. al. (1979) have demonstrated that in
aì'lergen sensitized dogs, basal resting tone ís increased, spontaneous
mechanical activity and myogenic response are present in vitro, but not
in littermate controls.
The resting membrane potentia'l of canÍne TSM is -50 + 2 mV (SE)
(Stephens, et a'|., I979) and demonstrates no phasic electrical fluctua-
tions. The contractile state of canine TSM is ìarge'ly dependent on the
degreeofdepoìarization.Suzuki,êt'âl',(L976)demonstratedthata4
mV depol arization from rest produced measurable contraction ( i.e.
mechanical threshold) in canine trachealis while Cameron and Kirkpatrick
(L977 ) using bovine trachealis showed that graded depoìarization produced
grade¿ mehanical responses. Stephens et. al. (1979) used increasing
concentrations of ç+ in the bathing solution to obtajn both graded
mechanical responses and graded depolarization" These graded responses
to 6+ and TEA are abolished in Ca++- free solutions demonstrating
that the mechanical responses to phasic and tonic depolarization are
dependant on extracel I ul ar Ca++. ( Stephens, êt. ôl . , 1975; Stephens,
1976 ) .
The unmasking of a potential for spontaneity with TEA is of specia'l
ínterest regarding the single unít characteristics of the airways in
disease (Akasaka et al., L975). in addition to bìocking potassium con-
ductance (Kroeger and Stephens, L975), TEA íncreases the number of gap
junctions (Kannan and Daniel, 1978), and increases the space constant

-7-(Stephens, êt. â1., 1979). The action potentials recorded in canine TSM
following treatment with TEA (Suzuki, êt. ô1., L976; Kirkpatrick, 1975;
Kroeger and Stephens, l975) were blocked when D-600 (a calcium channel
blocker) was added. These findings suggest an important roìe for a Ca-
current in the development of action potentials'
Graded stimulatÍon usíng a 60 Hz AC electrical source app'lied with
platinum plate electrodes, produced maximal isometric tetanus at 13V in
canine trachealis iq vitro (stephens and Kroeger, 1980). These contrac-
tions, which are atropine-sensitive, are not sustaíned with continuous
electrical field stimulation as observed by a 75% reduction from peak
tension after approxímately 20 seconds of applying the stÍ¡nulus. This is
thought to be the result of acetylchoìine depletion in cholinergíc
nerves. In contrast exogenous carbamylcholine or acetylcholine produces a
sustained isometric contraction wttich gradually decreases to approximate-
ly BS% of the initial response after four hours. Also of interest ís the
finding that the time to peak tension development is the same' regardless
of vo'ltage applied which implies a voìtage independent rate of contrac-
tion. Thís finding suggests a functional similarity among the contrac-
tile units of canine TSM and that as more voltage is applied, cells with
lower excítabilities, or cells farther from the electrode are recruited
in the response. Therefore unlike the all-or-none response observed for
cardiac muscle in which the depolarization spreads throughout the
f unctÍona'l syncyti um the tracheal Ì s appears to be al¡l e to mai ntaÍ n
various degrees of tonic activity dependant on the stimulus input. The
results with carbamylcholine also suggest that airway smooth muscle is
capable of rnaintaining a prolonged tonic contraction as observed in

-8-
asthma where there is prolonged bronchoconstriction'
Si nce thi s thesi s focusses on the neura'l and pharmaco'logic control
of tracheal smooth muscle the reader is referred to other reviews for
further ínformation on the biochemistry, biophysics and ultrastructure
(Richardson and Ferguson, 1980; Danie'l , et. ôl ., 1979; Stephens and
Kroeger, 1980) of srnooth muscle in airways.
NEURAL CONTROL OF AIRWAY SMOOTH MUSCLE
Parasympathetic SYstem
The parasympathetic system is the maior neural tnechanism for bron-
chocontriction in humans (Richardson and Beland, L976) dogs (Brown, êt.
ô1., 1982; Russelì, 1978) cats (0lsen et al., 1965) guinea pigs (Coburn
and Tomita , 1.973) baboons (Middendorf and Russel 1 , 1980) and cows
(Kirkpatrick, 1975). Stimulation of the vagus nerve results in wide-
spread airway narrowing (l^loolcock, et. â1., 1969) thus increasing resis-
tance to airflow. The latter can be enhanced by cholinesterase inhibitors
or blocked at the postsynaptíc muscarÍnic receptor by atropine (Colebatch
and Halmagyi, 1963), Stimulation of vaga'l fibers is also blocked by
tetrodotoxin (TTX) and hexamethonium. The blockade by hexamethonium
(which l¡locks nicotínic receptors) suggests the involvement of ganglia ín
the parasympathetic pathway. Hi stoì ogical ly these gangl i a have been
located in airway muscle or adjacent to it (Richardson and Ferguson,
1e80 ) .
Cholinergic nerve terminals or varicosities can be identified with
electron microscopy by demonstrating small agranuìar vesicles (SAV) of

-9-
50-70 nm diameter (Burnstock, 1970), oF by histochemical techniques
demonstrating the presence of acetylcholinesterase (Mann, 1971). The
gang'l i a receive pregangì ionic chol i nergic and adrenergíc fibers
(Richardson and Ferguson, l9B0) and some may have purinergic fibers (non-
adrenergic fibers) depending on the species. In the human the cholinergic
and nonadrenergic fibers (containing large agranu'lar vesicles, B0-120 nm
in diameter (Burnstock, 1970; Burnstock, 197?)) predominate, úile in the
dog both cholinergíc and adrenergic nerves (as demonstrated by histo-
chemical staining for norepÍnephrine (Fil'1en2,1970), oF small dense
cored granular vesícles of 40-70 nrn diameter (Burnstock, L97?)) are
present. Present evirlence suggests a paucity of adrenergic fibers in the
human (Richardson,1979) and no purinergic fibres in the dog tracheal
smooth musc'le. Postgangl ionic vagal fibers leave the gang'li a, which are
situated just outsíde the smooth muscle and cartilage of the airways to
travel through the smooth muscle giving off branches with multiple neuro-
transmitter-containing varicosities (Richardson and Ferguson, 1980)'
Direct innervation of airway smooth muscle or associated blood vesse'ls by
adrenergic fibers has not been cìearly shown and it has been suggested
that adrenergic fibers may not control airway srnooth muscle' Several
studies, however, using electrical stinulation of nerves have suggested a
functional role of adrenergic nerves in airway smooth muscle tone (Suzuki
et. ô.|., !g76, Cabezas et. â.|., 1971) Cholinergic, adrenergic and non-
adrenergÍc fibers may be i nfl uenced by pregangl ionic vagal fiber
stimul i .
InteractÍons involving reflexes (Coìeridge and Coìeridge, t977;
Russell and Lai-Fook, 1979; Widdecombe, 1975) and modulating infìuences
of ci rcul ati ng adrenal i ne, serotoni n, 1 ocal ly rel eased prostag'l andi ns ,

-10-
histamine and slow reacting substance of anaphylaxis (SRS-A) at the level
of the ganglia are not presentìy understood. The importance of controls
at the level of the gang'lia in the regulation of the effector tissue is
relevant for the understanding of neural mechanisms regulating airway
smooth muscle.
As was ment,ioned above, vagal stimul ation rel eases acetychol i ne
(ACh) from small agranular or electron-transparent vesicles. The ACh is
rapidly hydrolysed by acetylcholinesterase forming choline and acetate;
choline is taken up active'ly by the nerve varicosity to synthesize ACh
and thereby recycles and conserves choline (Burnstock, 1979).
Acetylcholine is released from cholÍnergic nerves following electri-
cal stimulation (Colebatch and Halmagyi, 1963). The contractile response
acompanying ACh release is blocked by atropine but not hexamethonium thus
demonstrati ng that the contracti on i s medi ated by post gang'l i onic
chol i nergíc nerve stimulation. In vitro studies in the dog have also
demonstrated simil ar responses (Russel I , 1978; Stephens and Kroeger'
l9g0). In vitro studies of guinea pig trachea (Carly1e,1963) show that
at rest, cholinergic nerves release ACh at a constant rate. It is likely
that in viyq the airways "at rest" are also under the influence of some
contÍnuous ACh release w|rich may serve to provide a resting tone in air-
way smooth muscle. Addition of atropine or ipratropium bromide results
in bronchodilation ín human airways frorn healthy individuals (Vincent et.
â1.,1970) and asthmatics (Gold,1975; Gandevia, l975) as well as dogs
(Severinghaus and Stupfel, 1955). This basal or rest,ing tone was also
abolished by vagotomy or cooling the vagus, thus demonstrating that a low
I evel of cholinergÍc activity maíntains airway smooth muscle tone
(Severinghaus and StuPfel, 1955).

- 11 -
EFFECTS OF ACH ON THE POSTJUNCTIONAL MEMBRANE
ACh released from nerve varicosities in tracheal smooth muscle binds
to postiunctional muscarinic receptors with a resultant depolarization of
the muscle mernbrane. Coburn (1979) has demonstrated that while ACh pro-
duces depoìarization it is not required for the production of contrac-
tions. In fact hyperpo'larization of the cell produced by anodal current
pu1 ses did not inhibit ACh-induced contractions and usìng the Ca++
channel blockers verapamil, D-600 or lanthanum had minimal effect 0n
these contractions. This study detnonstrates a pharmacomechanical coup-
'l ing independent, of membrane potential or external Ca++' In con-
trast high-K and 5-hydroxytryptamine (5-HT) stimulated contractions
dernonstrated el ectromechanical coupl i ng ancl a dependence on external
Ca++ (Coburn, 1979).
The action of acetylcholine also appears to stimulate increases in
cellular levels of cycìic guanosine monophosphate (c-GMP) (Schultz et'
â1., tg72\. This response' however, did not' correlate well temporaley
with the increase in intracellular Ca++ which preceeded t'he rÍse in
c-GMP (Lundholrn et. â.l., L976; Andersson et. ô1., 1975)" These data
illustrate the variety of mechanisms producÍng contraction in airway
srnooth muscle. For complete reviews on these mechanisms in smooth muscle
the reader is referred to articles by Bolton Í979) Coburn and Yamaguchi
l,tg77 ) and Simonsson and Svedmyr (]-977)'
SYMPATHETIC SYSTËM
Neural control of tracheal smooth muscl e
throughthepostgangìionicparasympatheticneura]
I ine (ACh) , which stimul ates posti unctional
(TSM) occurs 1 argelY
rel ease of acetYì cho-
muscarinic recePtors

-L2-(l^liddicombe, 1963; Russe]l , 1978; Stephens , !979) to produce contraction'
This appears to be true of most species studied as mentíoned above' The
presence of an adrenergic system innervating airlvay smooth muscle is less
clear. Histoìogical evidence indicates the presence of adrenergic nerves
in airways of cats (Silvia and Ross,1,g74), guinea pigs (Richardson and
Ferguson, LgTg) and dogs (Suzuki, 1976). Evidence for a direct innerva-
tion of TSM by adrenergic nerves in humans is lacking, however adrenergic
fibers are observed in the ganglia (Richardson and Beland, 1976)' The
role of these fibers is not known but it is possible that norepinephrine
released from adrenergic nerves in the gang'lia could act on cholinergic
fibers. Adrenergic fibers in gangìia of the canine lung have been demon-
strated (Jacobowi tz et. âl ., 1973 ). Si nce many adrenergic fjbers
accolnpany blood vessels in the lung' (F'i11enz' 1970' Kadowitz et' al
1976), it is difficult to demonstrate hístologically a direct control
airway smooth muscle-
A functional role of sYmPathetic neurons in canine airwaY smooth
muscle has been suggested by Cabezas et. âl', ft97L) who have shown that
in vivo electrical stimulation of these nerves produces a beta-adreno-
ceptor mediated bronchodilation which is blocked by propranolol ' Russell
(1980) has shown similar results for canine TSM in y'!lro" Further studies
have shown that sectioníng sympathetic fibers to the airways in dogs
resulted in a mild bronchoconstriction and suggested that the bronchodi-
lator activity serves to counterbalance the bronchoconstricting effect of
parasympathetic activity (Green and llliddicombe, 1966)' These resul'Es
suggest a basal rate of sympathetic and parasympathelic activity in aÍr-
v,,ays, and direct measuren¡ents bear thís out (t'liddicombe, 1966)" It is
.,
of

-13-
concl uded that a1 though adrenergic innervation to airways i s sparse
(Mann,1971) a functional role does exist" Controversy as to that role
is abundant. Electrical stimulation of thoracic sympathetic nerves has
been shown to protluce bronchodilation (Castro de la Mata, et. ôl', L962;
Cabezas et. â1.,1971) and this response is dependent on bronchomotor
tone which is abolished by vagotomy (Cabezas et' âl', 1971)' Interest-
ingìy, these studies found that when bronchoconstrictor tone was high the
bronchodil ator effect of sympathetic nerve stirnul ation v',as greatest
(cabezas et. â1.,1971). Similar results were found for cat airways, in
which the admÍnistration of eserine (which increased cholinergic tone)
also enhanced the effects of sympathetic nerve stimulation (Da1y et' âl',
1951). These results illustrate a relationship between active tone and
the bronchodilating action of syrnpathetic nerve stimulation"
A bronchoconstrictor action of the sympathetic system in airways has
al so been suggested (Adol phson, êt. âl ., 1971 ) ' Beta2-adrenoceptors
which mediate relaxation have been identified in the airl'rays (Fleish,
19g0) and thus forms the basis of many bronchodilating therapies (Jack,
Lg73; Wilson and McPhilìips,1978). The existence of alpha-adrenoceptors
metliating contraction of TSM is however, controversial. Evidence for the
exi stence of al pha-adrenoceptors i n the tracheobronchi al tree v''as
obtained for guinea pigs (Everítt and cairncross, 1969) dogs (Beínfield
and sei fter, 1980; Castro de al Mata et. âl . , t96?; Kneussl and
Richardson, 1978; Suzuki et. â1., 1976), rabbits, cats, rats and guinea
pigs (Fleish et. â1., 1970) and humans (Kneussl and Richardson 1978;
Simonsson et. a1., L97?; Mathe, êt. al., 1971)' Foster (1966)' Stone' êt'
al. (1973) and Cabezas et. al. (1971) were unable to find evidence for

-L4-
alpha-adrenoceptor mediated responses in guinea pigs, humans and dogs
respectively. species differences in the amount of contraction produced
by a'lpha-adrenoceptor stimulation, have been noted but could be demon-
strated only af'Eer beta-adrenoceptor blockade (Fleish et' âl', 1970;
Mat,he et. ô.l., 1g71). blhile beta-blockade uras used to demonstrate an
alpha-adrenoceptor mediated contraction in canine trachealis (Suzuki, et'
ô1., 1976) with electrical field stinrulation in vitro, others (Beinfield
and Seifter, lg80) were able to demonstrate phentoìamine-sensitive con-
tractions following low doses of norepinephrine (i.v.) in canine TSM in
the absence of ProPrano'lo1 .
Variabil i ty of the adrenergícal ly medi ated responses noted above
might be attributed to age-related effects" Pandya Í977), on the basis
of observations that the TSM of younger dogs responded more strongly to
norepinephrine than that of older dogs, concluded that the number and/or
activity of a'lpha-adrenoceptors decreased with increasing age' Another
possibility is that the pre-existent tone of the muscles qualitatively
influences the response to alpha-adrenergic stimulation (Ohno et' â1"
1981). Thus Richardson and Ferguson (1979) demonstrated greater a'lpha-
adrenoceptor mediated (phentolamine sensitive) contractions in tracheal
and bronchial smooth muscle strips taken from humans with pneumonia or
chronic obstructive aírway disease. We (Antonissen et' âl', L979) have
also noted that in conditions in wtrich t,he nonnally passive basal tension
is increased, as with immunological sensÍtization, responses to adrener-
gic agents are al tered and have sugges'üed (Bergen and Kroeger, L979) the
possibility of increased alpha-adrenoceptor mediated responses with sen-
sítization" Whether this reflected a prirnary effect of sensitization or

-15-
a consequence of increased basal tone was not clear and pre'liminary
studies with a variety of stimulatory agonists supported the notion of a
controlling influence of pre-existent tone in these responses'
POTASSIUM.INDUCED RESPONSES
potassium-rich saline (high-K) contracts tracheal smooth muscle of
rabbits, guinea pigs, rats (Akcasu, L959), dogs (Kneussl and Richardson'
1978; Coburn " IgTg) and humans (Kneussl and Richardson). High-K treat-
ment resul ts in a depol arization dependent Ca++i nfl ux (Somyl o and
Somlyo , Lg70) producing subsequent smooth muscle contraction' Choliner-
gic receptor antagonists in intestinal smooth muscle and a'lpha-adrenocep-
tor antagonists in vascular smooth muscle reduce the contractile response
to high-K indicating that neural depolarization and transmitter reìease
constitute a component of the response (Lundholm et. al., 1976)'
PREJUNCTIONAL NEUROMODULATION
The frequency of action potentials arriving at the axon terminal or
varicosit,ies was thought to be the only determinant of neurotransmitter
release untÍl recently when it was shown that for the adrenergic system
hormones, neuroLransmitters and local microenvironmental conditions coul d
influence this quantal release (Westfal,l, 19771. The adrenergic system
has come to be the most widely studied with respect to neuromodulation"
and four general mechanisms have been described (Westfall, L9771'
1) Locally, norepinephrine released may bind to presynaptic or pre-
junctional receptors and either increase (positive feedback) or
decrease (negative feedback) further norepinephrine released by
action potentials" This provides a means of autoregu'lation'

-16-2l Transynaptic regulation, where postsynaptic stimulatÍon by
neurotransmitter binding to postjunctional receptors results in
an increased production of a substance or substances (e.g' pro-
stag'landin or adenosine containÍng compounds) which provide
feedback and neuromodulate further transmitter release.
3) Contralateral neuronal controls whereby neurotransmitters from
different nerves (cholinergic, dopaminergic or serotonergic)
adjacent to adrenergic fibers may neuromodulate norepinephrine
release and vice versa.
4) Neuromodulation of norepinephrine release l¡y substances in the
blood such as angiotensÍon, epinephrine or 5-hydroxy-tryptamine.
These mechanisms of neuromodulation for the adrenergic system'
especial'ly the vascular component, illustrate an important regulatory
function. These controls, in other neural networks (e.g: cholinergÍc)
are presen¡y not well known. For purposes of this revieb, several of the
factors important ín prejunctional neuromodulation will be considered,
a'lthough these factors have not been studÍed in airway smooth muscle.
ADRENERGIC TRANSI4ITTTR RELIASE
Norepinephrine is synthesÍzed from tyrosine in adrenergic nerves and
stored in granules until an increase of intracellular calcium promotes
release of the granules by exocytosis (Katz and Miledi,1970; Kirpekar'
prat and Wakade, 1975). Thus action potentials, increased extracellular
potassium concentra't,ion or calcium ionophore A23L87 increase intracelìu-
I ar Ca++ and al I rel ease norepi nephrine (Mel I ow, L979; Tauc, 1982 ).
Released norepinephrine may be recyc'led by an active neuronal uptake

-L7-(Uptake l). This rapid uptake is inhibited by cocaine, imipramine and
amitríptyline antidepressant drugs (Iverson, L974; Langer, 1977; Nigro
and Enero, l98l). A small amount of the NE in the nerve is metabolized
via rnitochondrial monamine oxidase system (MAO) to 3, 4, dehydroyphenyl-
glycol (UOpee) and other metabolites. Unstimulated adrenergic nerves
passively release DOPEG continuously and only sma'|1 quantities of NE are
released. Recentìy, it has been shown that NE is released at a basal
rate which may cqme not from the vesicles of the neuron but the cyto-
plasm. This release produces an estimated 1O-BM concentration of NE
in the immedÍate extracellular area of the nerve varicosity and corres-
ponds to the quantal reìease estimated to occur wtren the action potentia'l
frequency is 2 heriüz (Tauc, 1982). St,imulation of the nerve greatly
increases doparnine beta hydroxylase and NE release while D0PEG release
remains low (Vanhoutte, 1978) 'therefore s'bimuli which increase 3¡1-
norepinephrine overflow do not increase NE metabolite overflow substanti-
al1y. Released NE may bind to effector or target cells at specific re-
ceptors ( al pha or beta) or be taken up extraneural ly by a mernbrane t'rans-
port system (uptake 21. uptake 2 has a much lower affinity for NE and
therefore the majority of Nt released is removed frorn the iunctional
cleft by neural uptake (uptake 1) (Langer, L9771. This difference in
transport systems is vital to the loadÍng of adrenergic nerves with
radiolabelled norepinephríne (Iversen, 1,974; b{estfall, L977). The load-
ing of adrenergic nerves with 3H-norepinephrine has been used in many
tissues for the purpose of examining the prejunctiona'l inf'luence of many
substances (Duckles and Rapoport, L979; Nigro and Enero, 1981; Dixon et'
âl ., L979; l¡leiner , L979; Lorenz, Vanhoutte and Shepherd , t979; l¡lerner et'

-18-
ôl ., i,979; Russel I and Bartl ett, 1981). Foster Í975) demonstrated that
¡3¡11-noradrenal ine neural uptake in gui nea pig trachea h'as almost
eìiminated when nerves were destroyed wíth 6-hydroxydopamine. Subsequent
trans¡nural el ectrical stimul ation resul ted in a marked reduction of
relaxant responses ano 3[H]-NE efflux.
Use of 3H-norepinephrine has been applied to the study of nerves
in airways. After loading adrenergic nerves with 3H-norepinephrine
Vermiere and Vanhoutte '1rgTg) were able to demonstrate a release of 3H-
NE rvith electrical stimulation of canine tracheal smooth rnuscle strips jI
vitro. They also demonstrated that exogenous NE reduced contractile
respgnses following parasympathetic stirnulation more so than contraction
to eogenous acetylcholine in canine airways, an effect whích rl,as abol-
ished wÍth propranolol. It was suggested that NE released from adrenergic
nerves may inhibit ACh release via prejunctional mechanisms (Venniere and
Vanhoutte, LgTg). In contrast, a stimulatory role has been described for
serotonin on the cholinergic system in canine airways (Sheller et. ô1.,
1982). Recently, Russel I and Bartl ett (1981 ) measure¿ 3H NE and
norepinephrine metabolites in the superfusate of canine TSM strips
fo'llowing electrical or tyramine stimulation. llJhile both stimuli in-
creased overflow of intact norepinephrine and NE metabolites the percen-
tage increase uJas greater for norepi nephrine. Usi ng 3U-ruf as a qua'l i-
tative indicator of norepinephrine release these authors also demon-
strated an atropine-sensitive inhibitory effect of exogenous acetyl-
choline on eìectrícal1y stimulated 3H-U¡ overflow and that TTX blocked
all overflow produced by electrical stimulation but had no effect on the
action of tyrarnine. Other preparations, most notably those of the
vasculature or vessel-rich organs such as the sp'leen have provided much

-19-
informatÍon about the nature of prejunctional receptors. Preiunctional
al pha-receptors ( termed al pha2) have been wi dely studi ed ' Aì pha2
receptors as opposed to posti unctional al phal receptors have higher
affinit,ies for the agonist clonidine (Doxey et. âl ., L9771 and the
antagoni st yohimbi ne (ìilood et. ôl . , 1'979). Postsynaptic a'l phal recep-
tors bind the agonist phenyìephrine (St,arke, êt'ôl',1975) and antagon-
ist prazosin (cavero, et. ô1., L977; Cambridge, êt. â.|., L977\ with
greater affinity. utilizing these tools to separate pre- from post-
junctional receptors has demonstrated that stÍmul ation of al pha2
receptors decreases Ca++ movement jnto the nerve and thereby reduces
NE rel eased by nerve stimul at,i on . By bl ocki ng presynaptic a'l pha2
receptors with yohímbine, phentoìamine, oF phenoxybenzamine' (Langer'
Lg77) electrically stimulated NE-overflow is increased. 0n the basis of
these studies it has been suggested that when nerves are firing at a'low
frequency the amount of NE released is small and binds to the higher
affi nity prei unctional beta2 receptor to prornote NE rel ease ( i 'e '
positive feedback). This has been demonstrated by Dixon, et' al' (1979)
using isoproterenol (10-7-10-6M) to enhance 3H-ruE overfl ow,
an effect which was propranolol-sensitive in isolated cat spleen' As
acti on potenti a'l frequency i ncreases more NE i s rel eased (Langer, t977't
and upon reachi ng a threshol d concentration stimul ates prej untional
aìpha2 receptors which reduce the amount of transmitter release (nega-
tivefeedback).Thephysiologicimportanceofalpha.mediatedprejunction-
al neuromodulation was demonstrated by Starke (t972) who showed that'
phenoxybenzamine enhanced electrically stimulated NE overfloh' and this
was accompanied by increased heart rate in the ral¡bit. I^lhile the role of

-20-the presynaptÍc beta2 receptor is probably less important than the pre-
synaptic alpha2 receptor Dixon et. al. (1979) have suggested that cir-
culating catecholamÍnes whose levels increase under stress may activate
presynaptic beta2 adrenoceptors thus enhancing NE release from adrener-
gic nerves.
Acetylcholine (ACh) appears to act in a neuromodulating capacity on
adrenergic NE release. Low concentrations of ACh inhÍbit release of NE
ín response to electrical stimulation (Fozard and Muscho'll, L972) and
this effect can be elÍminated by e'levating extracelIular calcium (Dubey,
et. â1., 1975). ¡¡hile atropine antagonises the inhibitory action of ACh
on NE release (Russell and Bartlett,1981) increasing the ACh concentra-
tion ín the presence of atropine results in augmented Nt overflow upon
electrical stimulation (Lindmar, êt. â1., 196S). The physio'logic impor-
tance of muscarinÍc inhibition and nicotínic augmentation of NË release
is not known. 0f interest is the finding that in the gastrointestinal
tract (Manber and Gershon,1979) and airway smooth muscle (Richardson and
Ferguson, 1980; Jacobowi tz, et. âl . , 1973 ) adrenergic and chol i nergic
axons are often seen side by side and even within t'he same Schwann cell
sheath. The role of this close association seen on electron microscopy
is not known, þut presynaptic mechanisms may be irnportant in such areas.
¡ther presynaptic receptors have been postulated and they are listed
here with a brief account of their action on NE release from adrenergic
nerves when stimul ated e1 ectrica'l 1y.
1) DopamÍne receptors which inhibit Nt release (Enero and Langer,
1975; Long, et. al ., L975; Hope, et' al ', 1978)'
Z) gpiate receptors which, when activated by morphine (Hughes, êt.

-2r-âl ., 1975) or pentapeptides met anq 1eu-enkephal in (Langer,
L977\ decrease NE release.
3) Inhibitory prostaglandin receptors stirnulated by prostaglandins
E1 and E2 (Stiarne, 1973; Hedqvíst, 1976)'
4) Adenosine receptors which decrease Nt release (Hedqvist and
Fredholm, L976; Verhaeghe, vanhoutte and shepherd, 19771.
S) Angiotensin II receptors which increase or facilitate NE release
(Zimmerman, 1978).
6) Histamine (H2) receptors wtrich inhibit NE release (McGrath and
Shepherd, !976; Foldes and Hall, L979; Powell, 1979)'
7) 5-hydroxytryptamine (5-HT) receptors'
presynaptic sites for 5-HT are suggested by indirect evidence from
rabbi t heart ( Fozard and Mi val uko , 1976 ) and i sol ated cat sp'l een
( pl uchi no , tg72). llle stfal I n977 ) sugge sted that 5 -HT rnay i ncrease NE
release from nerves during 1ow frequency firing whereas Fenuik, et. â1.,
(tg7g) have demonstrated a reduction of NË release produced by potassium
depoìarization or electrical stímulation" Whether 5-HT does release NE
frorn nerves at rest has not been detennÍned by direct measurement of
3H-rur overfl ow.
This review of presynaptic receptors illustrates the presynaptic
regulation for the adrenergic system in several tissue types and species.
physiologic and pharmacologíc importance are ascribed to these presynap-
tic receptors. The work descrÍbed has mainly centered on adrenergÍc
neuromodulation in t,issues where adrenergic innervation is dqninant. In
the gastrointestinal tract and tracheobronchíal tree the parasympathetic
(cholinergic) system is dominant. Only a few studies examining presynap-

-22-
tic regulat,ion of the cholinergic system have been undertaken, those in
the gut (Powell and Tapper, L979; Vizi and Kno1l, L97L; Drew, 1',9771 and
canirre airways (Vermiere and Vanhoutte, L979; Sheller, et' al ', 1982)'
General ly, depo'larízation of nervous tissue is accompanied by
i ncreases of i ntraneuronal cycl ic AMP (Kaki uchi , et. âl . , 1969) and
cycìic GMP (Ferrende11i, et. ô.|., 1973). These nucleotides have been
associated wi th cal cium movements in a variety of smooth rnuscìes
(Marshall and Kroeger, Lg73; Bolton, L979; Diamond, 1"978) and in nervous
tissue with a release of neurotransmitter that ís dependent on Ca++
movements (hleiner,1,g7g). Since many of the factors known to have a role
i n prej unctional neurornodul ation al so act postsynaptical ly to affect
cyclic-AMp and cyclic-GMP levels in effector tissues, it is conceivable
that they may protluce similiar effects in nerves. This has been studied
for several substances (Westfall, L977), however little attention has
been given to the effects of presynaptíc receptor activatÍon in nerves at
rest which have a basal rate of firing and hence transmitter efflux.
Presynaptic modulation at low frequencies may be quite different
f rorn that at higher stimul us frequency (sax and westfaì'l , 198i ). A
biphasic regulation is found for norepinephrine (Langer, L977; Langer'
et. al ., 197b) and perhaps s-HT (!'lestfa'l \, L977 ) as mentioned previous'ly.
The rel ative importance of these possibl e regul atory ntechani sms
physiologically, pharmacologicaì'ly, therapeutically in diseases such as
asthma are presently unknown. The widespread use of substances such as
propranol ol and clonídine for hypertension have shown the important
app'l ication gai ned by understandi ng presynaptic neurønoclul ation "

-23-Several excellent reviews on presynaptic regu'lation serve to provide
an overall evaluatíon of these mechanisms (Langer, L977; t^lestfall, t977;
Weiner , L979; Starke , 1'977).
AIRWAY SMOOTH MUSCLE HYPERREACTIVITY
That patients wi th asthma show increased bronchoconstrictor
responses to a variety of stimuli has been knov¡n for some time (Curry,
1946; hleiss et. ô.|., Lg32l. Further studies have demonstrated increased
bronchoconstrictor responses of asthmatics to serotonin (Panzani, 1'962\,
acetylcholine, methacholine and carbachol (Curry, L947; Parker, et' âl',
1965; grehek, êt. ôl ., Lg77), prostag'landin F2 a'lpha (Mathe, êt' âl ',1973 ), bradyki nÍn (Varonier and Panzani, 1968), fo'l'lowi ng exerci se
(McNeil1, et. al., 1966), cold air inhalation (simonsson, et' al ', 1967),
dust (Dubois and Dautrebande, L95B) and chemical irritants such as sulfur
dioxide (Nadel, êt. a1.,1965) as welì as drugs (Rebon and Parikh, l9B0).
certain patients deveì op speci fic al'l ergies and when exposed to the
relevant allergen develop narrowing of the airways (Ki1'lian' et' âl',
Lg76). Although bronchíal hyperreactivity has been used to describe the
smooth muscle constriction producing airblay narrowÍng in asthmatics other
factors such as mucosal edema, excessive mucous production, pooìing and
alterations in mucous viscosity all serve to increase resistance to
airflow in the tracheobronchial tree. The focus of this review will be
on mechanisms ínvolved in asthma leading to a bronchoconstrictor response
mecli ated by ai rway srnooth musc 1 e .
Cross sectional area of the airways at different levels reveals that
the resistance to airflow decreases progressively frøn the nasal passages

-?4-
and large airways, to the smaller t¡ronchi (Hogg, 1981). Bronchoconstric-
tor stimuli rnay constrict airways at various leve'ls including the trachea
where large Íncreases in resistance to airflow can occur. In humans and
Ín dogs the major airway narrowing appears to occur at the level of
smal I er ( 1-5mm) bronchi . The si Ee of i ncreased resi stance i s al so qui te
variable as observed in humans (Ruffin, et. âl ., 1978) and dogs
( Wool cock, et. âl . , 1969) . The mechani sms produci ng thi s resi stance to
aírflow have been studied from a variety of perspectives and have led to
several theories concerni ng bronchí al hyperreact,ivi ty. Bronchi al
hyperreactivity is not excìusive to asthma since it occurs Ín patients
with chronic bronchitis (Simonsson et. ôl ., 1970), viral infections of
the respiratory tract (Parker et. ô1.,1965) and hay fever (Townley, et.
â1.,196S). It is tikeìy that the mechanisms involved in the different
disorders which have some increased resÍstance to airflow are different
or overlap each other. Therefore it is simpìistic to view asthma as a
disease involving onìy one mechanism; it should probab'ly be viewed as a
syndrorne of defined symptoms (e.g. íncreased resistance to aÍrflow,
increased reserve volume of the lung) whích may be produced by single or
multip'le mechanisms. Studies on asthmatic patients treated with several
therapies do suggest rnultiple causes since various indivídua'ls respond to
cholinergic antagonÍsts (Snow, et. â1., L979; Lightbody, êt' ôl', 1978)'
beta-adrenoceptor agonísts (}{ilson and McPhilìips, 1978; Laarson and
Svedemyr , 1,977\, adrenoceptor antagonists (Patel and Kerr, 1975)'
steroids such as dexamethasone (Leitch, L9B2), the mast cell stabilizer,
disodium cromoglyca'fe (Bernsteín, 1981 ), antihistamines (Kar1 in, 1972;
popa, Lg77\ ancl prostaglandins (Mathe and l{edqvist, 1975).

-25-Hyperreactivíty of airway smooth rnuscle, as occurs in asthma may be
conveniently divÍded ínto two groups regarding mechanisms: abnormal
stímulatory mediators and abnormal smooth muscle responses to normal
stímuli. blithin thís framework further subdivisions can be made. While
a complete review of all the mechanisms is not the purpose of this
thesis, it is hoped that an outline may give the reader a perspective on
the mul ti p'l e approaches to thí s subi ect.
I ABNORMAL MEDIATOR STIMULI
The first major group refers to alterations in the mediator inputs
to produce either relaxation or contraction of srnooth muscle. It may be
further subdivided into extrinsic factors and intrinsic factors"
A) Extrinsic Factors
1) A'llergy - This approach to the study of asthma involves immune
rnechanísms leading to airway narrowÍng. Allergic reactions may be of the
immediate varíety (Type I) or involve a delayed react,ion (Type IV)'
Immediate hypersensitivity reactions may be further subdivided into
atopic and non-atopic types, the former invo'lving quantitatively and
qualitativeìy abnormal reactivity to concentrations of endogenous sub-
stances or exogenously administered pharmacoìogic mediators which other-
wise produce no reaction. Non-atopic disease Ínvolves a nomal antibody
response to unnatural exposure of antigen" Atopic disease involves over-
productíon of antibody to a wide variety of antigens such as are found in
house dust or tree pollens whereas non-atopic disease occurs only on
exposure to the specific antigen to wtrich the individual is sensitized.

-?6-Both mechanisms involve mast cells located in the epithelium of airways
(Salato, Ig76) which degranulate following crossbridging of immunoglobu-
I in E (IgE) attacherJ via theír F. portions to mast ce'll membranes (Gold
et. â1., L977, Ishizaka and Ishizaka,1,975). Degranulation leads to
release of histamine, eosinophil-chemotactic factor (ECF), prostagland-
ins, SRS-A (slow reacting substance of anaphylaxis now thought to be a
mixture of leukotrienes), various kÍnins and other substances (W'ilson and
Galant, lg74l. llistamine (Yanta, êt. â1., 1981; Loring et. â.|., 1978)
SRS-A (KrelI and Chakrin, 1978) and several prosbag'landins, excepting the
E serÍes (Snapper, êt. â.|., L979; Malo, et. â1., 1982) can all produce
tracheal smooth muscle contraction and airway narrowing. The response to
aìlergen inhalation can be partia'l'ly btocked by H1 antagonists (Kar1Ín,
Lg72l, atropine (Yu, et. ô1., L972; Gold et. â1., L972) and disodium
cromoglycate (Berstein, 1981). Inhibition of mast cell release of media-
tors can be affected by hi stamíne (HZ receptors) , beta-adrenoceptor
activation, pros'bag'landin E1 and g'lucocorticoids actÍng on mast cel I
menrbranes to inhibit degranulation (Líchtenstein and Gil lespie, L973;
Norn and Skov, 1979). The process of degranulation is dependent on
Ca++ movement into mast cells or basophils (KazimÍerczak,1978) and a
decrease in cellular cyclic-AMP (Norn et. â.|., L977). Substances such
as al pha-adrenoceptor agonists, acetyìcholine and prosLagìandin F2
alpha are assocíated with decreasing ce'llular cyclic-AMP levels and
thereby enhance mast cell degranulation (Krell and Chakrin, L976; Tung
and Lichtenstein, 1981). The histamine released may therefore act 0n
severa'l si tes: Hl receptors on ai rway smooth muscl e produci ng con-
traction (Chand et. â.|., L979; Himori and Taisa, 1978) H2 receptors on

-27mast cel l s to increase cycle AMP i ntracel'lul arly and inhillÍt further
h i starni ne rel ease (Li chtensteÍ n and Gi I I espi e, 1,973; Norn , et. âl . ,
L9771; receptors on adjacent supporting cells and smooth muscle cells tostimulate prostaglandin and leukotriene synthesis (Lewis and Austin,
1981; MÍtchell, 1981; 0range, Lg77l, or on irritant receptors located in
airway epithelium (Vidruk, êt. ô1., 1976) to produce bronchoconstrictÍon
by a vaga'l 'ly medi ated refl ex (Drazen and Austen " 1,91s; Gol d et. âl . ,
1977 ; Wasserman, 1975).
sRS-4, present'ly thought to be leukotrienes c4, D4 and Ë.4
(Leitch,19B2) produces prolonged bronchoconstriction. The role of
prostaglandins and other arachidonic acíd metabolites is currently of
great interest as to theír role in many of the bronchoconstrictor
responses, however the subject is too broad for revÍew here. The reader
i s referred to the fol 1 owi ng references ( Nakani shi , et. ôl . , LglB;
Anclerson, êt. â1., 1980; wasserman, Lgl6; Gardiner and co1ìier, 19g0;
Adcock and Garland, 1980; Dah'len, et. a1 ., 1980, BrÍnk et. a] ., 1990).
Late astltmatic al lergic reactions may appear gradua'l'ly over several
hours following exposure to a specific al'lergen" The reaction is thought
to be of the Eype III, immune complex type, and likely Ínvolves broncho-
constrictor prost,ag'landins, histamine and bradykinin (pepys ancl
Hutchcroft, 1975).
Models of a1'lergic asthma have been developed (Hfrshman et. â1.,
1980; wanner et. a1., L979) and serve to provide a means by which il vivo
and in yitro studies can investigate muscle responses to antigen chal-
1enge" The animal s used are sensitized to a specific an'L,igen such as
oval bumin (Antonissen et. âl ., L979; Kepron, êt" ôl ., Lgl7l, ragweed

-28
pollen (Patterson, 1969) or ascaris antigen (Hirshman, êt. â1., 1980;
Krel I antl Chakri n, 1976 ) . Antoni ssen et. ôl . , Í979 ) have demonstrated
that canine tracheal smooth muscle strÌps isolated from ovalbumÍn-sensi-
tized dogs contract when exposed to the specific antÍgen ovalabumin (04).
Mepyramine a specific H1 antagonÍst filocks the response to 0A challenge
(Antonissen, et. â1., 1980). In vivo studies on the dogs sensitized to
0A (Kepron et. ô1., t977) demonstrated a marked increase in airflow
resistance following aerosol challenge with antigen. Antonissen et al.
(tglg) also demonstra'Eed tracheal smooth muscle of dogs sensitized to 0A
had increased rates and amounts of shortening as compared to non-sensi-
ti zed I i tternrate control ti ssue i n vi tro. These resul 'bs il I ustrate t'he
conrplexity of allergic reactions in allergic disease which may involve
not only differences in mediator release frotn mast cells but also changes
in the smooth muscle itself.
2) Irritant Receptor Reflex - The irrit,ant or fast adapting receptor
has been described by l^liddicombe (1954) as being involved in mediating
cough responses to upper airway stimuli (Nadel, L980; Morto'la, et. ôl.,
1975) and bronchoconstriction with lower airway stirnulation. The irritant
receptor Ís thought to be located within the airway epithelium (Mortola,
et. al.,1975) and is stimulated by histamine (sampson and Vidruk,1975;
Vidruk, êt. ô1., L976; Gold et. â1., L9721 ozone (Goldsmith and Nade'|,
1969) dust (Kessler, et. â1., L973\ ammonia (l,lliddÍcombe, L975) sulfur
dioxide (Nadel et. a'l ., 1965) mechanical stirnul i (Boushey, êt. a1 ., L972)
and other substances ( Nadel , 1980 ) . The refì ex bronchoconstrictÍon
produced is mediated by the vagus nerve as demonstrated by its sensi-
tivity to atropÍne (Gold et. â1., L972) hexamethonium (Simonsson et.

-?9
ô1., 1967) or vagotomy (Gold et. â1., 1972; De Kock et. â1., i966).
Since histamine appears to stimulate this reflex strongly in humans
(Simonsson, et. â.|., tg67l and also to some extent in dogs (Yanta et.
â1.,1981; Kessler, et. ô1.,1973) the relationship to allergy and mast
cell degranulatíon is apparent. Other factors such as viral infections
which damage airway epithelium (Dixon, €t. â1., t979) rnay increase expo-
sure or sensitivity of these irrit,ant receptors to the stimuli mentioned.
In fact nrany indíviduals do show an atropine-sensÍtive increase in air-
flow resistance following a viral respiratory tract infection (Empey, êt.
â1., L976; DÍxon, êt. ô1., 19791. The reflex seems to be involved pre-
dominantly wÍth extrinsic factors such as allergens or air contamÍnants.
0ther considerations such as alterations in mucosal permeabi'lity (Hogg
et. ô.|., LgTgl or darnage to airway epithelium (Lee, et. â1., L977) may
explain vúhy only certain persons develop the bronchoconstrictor response
following chronic exposure to ozone or a viral infection and suggest
intrinsic changes, perhaps in the neural activity of the airways. Hista-
mine may interact with vagal efferents to alter activity either at the
level of the ganglia or central nervous system (Loring et. ô1., L97B;
Douglas, et, al., 1973). Qthers (Hahn, êt. â1., I97B; Sheller et. ô.|.,
1gB2) have demonstrated a facílitation by 5-HT 0f bronchoconstriction
produced by vagal stimulat,ion but not exogenous ACh. These results
suggest a possible influence of histamine and 5-HT on vagal efferent
activity, acetylcholine release or postjunctional changes in the smooth
muscle itsel f.
The reflex involving the vagus nerve has also been implicated in
exerci se i nduced asthma (Simonsson, et. al " , L967 ). Col d dry ai r

-30(Strauss, êt. ô1., L977) and rapid respiratory rates (Haynes, Êt. â'|.,
L976) may precipitate l¡ronchoconstriction" Since exercÍse in a humid
warnl environment, such as swímming is associated with rnilder bronchocon-
strÍction in exercise-induced asthrnatics, Ít has been suggested that
cold, drying and mechanical stirnulation of irritant receptors may be the
major factors involved (Strauss, êt. ô.|., 1977:' 1978) although atropine
is not alvrays effective in blocking the response (Deal, êt. â1.,1976).
It has been shown (Gross, Êt. â1., L974l, that sotne exercise-induced
asthmatics respond to treatment with phentolamine, thus showing that
other neural or receptor mechanisms are also like'ly Ínvolved.
B) Intrínsic Factors
I ) Chol inergic 0veractivity - The chol inergic system which is
active in dogs (Severinghaus and Stupfel,1975) and humans (Vincent et.
a1.,1970) to provide a basal tone to airway smooth muscle, may be over-
active due to reflexes, psychogenic stimulÍ or abnormalities in regula-
tion at the ganglia (Boushey, et. a1.,1980). Since the resting baselíne
cal iber of the ai rways can be al tered by chol i nergic stimul ation
(Colebatch and Ha1rnagyi,1963) it seems probable that a wide range of
resting values exist. Boushey et. al. (1980) have suggested that since
resÍstance to airflow is inversely proportional to the fourth power of
the radiuso equal absol ute changes in radi i wil I produce greater
increases in airflow resistance Ín the smaller aÍrways.
Experiments on asthrnatics and normal individual s have indicated that
1 arge di fferences i n bronchi al reactivi ty occur fol I owi ng carbachol
cha'llenge even when baseline values of airway resistance are sÍlnilar
(Rubínfeld and PaÍn, 1,9771. De Kock et. al. (1966) demonstrated that

- 31
changes in baselíne aÍrflow resistance by stimulatÍon of sectioned cervi-
cal vagus nerves in dogs did not al ter the bronchoconstrÍctor response to
histamine aerosol. These studíes indicate that resting airway caliber
does not influence the further contraction of airway smooth rnuscle in
response to histarnÍne or carbachol. lllhether thÍs is true of other
bronchoconstrÍcting agonists is not known"
AsthmatÍcs (Cropp, 1975) and patÍents with chronic bronchitis
(Klock, êt. â1., L9751 may have increased airway smooth muscle tone pro-
duced by increased vagal activity. This conclusion Ís suggested by the
findÍng that atropine reduces airway resistance to airflow and that air-
ways constrict markedly in the presence of a beta antagonist, (which is
blocked by atropine, McNeil I and Ingrarn, 1966). The cause of thÍs
cholinergic overactivity in asthmatics is not known but, as mentioned
above, several tactors ( refl exes, CNS, gangl ionic regul ation) must be
consi dered.
2) Alterations in Mediator Production and/or Release - Since
asthmatics have marked airway hyperreactivity (Orehek, êt. â1., L977;
Curry, 1947) to aerosolized histamÍne or carbachol, much study has sought
to explain these phenomena without considering the possibility of altera-
tion Ín the quantitÍes of mediator released from nerves, mast cel'ls,
platelets, lymphocytes, basophils and eosinophils. Chronic asthrnatics do
have an increased number of eosinophils infiltrating airway nuscle and
rnuscosa especially around blood vessels (Kay et. â1., L97Ll. Their role
i n infl amatory responses i s partÍa'l 1y understood whil e information
concerning meclÍators from eosinophÍ1s which alter muscle contractility
are not known.

-32
In some asthmatics, changes in the number of tnast cells may occur
thus allowing for more mediator release following degranuìation and
therefore greater bronchoconstriction. Following antigen challenge cir-
culating levels of hístamine do increase in asthmatics (Gold, et. â.l.,
Lg77 | but these I evel s do not correl ate wel I wi th the severi ty of
bronchoconstriction "
Stgdies measuring catecholamines, cortisol and histamine (Barnes et.
â1.,1980) in asthmatics at various times have suggested that when cir-
culat,ing adrenaline is ìow, aS occurs at night and early morning, the
beta-adrenoceptor activity mediating a negative feedback on mast cell
histamíne release and smooth muscle relaxation is reduced and the greater
amount of circuìating histamine could produce an asthmatic attack or at
I east increase its severitY.
The role of 5-HT in producing bronchoconstriction in yiyo is not
clear although it is a potent pulmonary vasoconstrictive agent (sterling,
et. â1., Lg72l. In rats serotonin appears to play a major role in
anaphyl actic bronchoconstrictÍ on si nce the I atter i s 'largeì y (70/"\
blocked by methysergide (Church, êt. âl ., L972). Exogenous serotonin
administered to dogs (She1ler, et. â1.,1982) or humans (Haios, t96?;
Rodbard and Kira, Lg72l does produce contraction of tracheal smooth
muscle. The action of serotonin appears to be mediated via a D receptor
l ocated on the muscl e membrane (whÍch i s bl ocked by 'lysergíc acid
diethy'lamide, l-SD; Born, 1970) and a M receptor on nerve cells wtrich is
blocked by atropine or morphine and is therefore thought to be located on
cholinergic neurons (simonsson and Svedrnyr, 1977). Evidence for an inter-
action between serotonin and the cholinergic system has been adduced for

-33
the dog (Hahn, êt. ô.|., 1978; Shel'ler, et. â1., 1982) but the importance
or role of this interaction in disease states is unknown. It vivg studies
of humans suggest that normal subiects do not produce bronchoconstrictÍon
follov¡ing inhalation of aerosolized serotonin (Panzani, 1962) whereas the
majority of ¡hose with asthma do respond as measured by a 20% decrease in
FËV1 (Hajos , t962).
3) Sympathetic System - Some controversy as to the presence and
f unctional rol e of adrenergic fil¡ers i n ai rway smoot,h muscl e was
mentioned above. The role of the sympathetics in human airway muscle is
not clear (Richardson and Beland, 1976) whÍ1e in dogs (Cabezas, et' âl',
LgTLl they are thought to be predøninantly invo'lved in effecting broncho-
dilation or counteracting the vagal constrictor influence. When sympa-
thetic nerves to canine airurayS were sectioned (Woolcock, et. ôl ', 1969)
a small increase in aírflow resistance resulted. In humans, pretreatment
with the beta-adrenoceptor blocker propranolo'l potentíates bronchocon'
strictor responses to acetyìcholÍne (0rehek, êt. ô1., 1975) cigarette
smoke (Zuski n, et,. âl ., Ig74l and histamine (P'loy-Song-Sang, 1978) '
These studies suggest, a functionally signifícant inhibitory role for
sympathetÍcs in humans. It is conceivable that alterations in sympathetic
nervous activity are involved in disease states with airway hyperreacti-
vity. The level at wtrich the possible defect in the sympathetic system
occurs may be at the receptor, nerve or ganglion. Defects or changes at
the receptor level are postjunctional and are di scussed as abnonnal
smooth muscle response mechanisms"
Sympathetic nerve changes, as in rate of activity or in the varying
density of innervation may occur. Damage to sympathetic nerves selective-

-34'ly is unlikely, however genetic factors may contribute since it is known
that asthma has some familial tendancy (8ias,1973). Hirschsprung's
disease, for examp'le, in which a section of the colon is devoid of non-
adrenergic nerves thus leading to defects in smooth muscle control is
genetic ín origin, (Frigo, êt. â1., 1973; Hukuhara, et. â.|., 1961).
Whether an analagous situation involving sympathetics occurs in airways
of asthmatics Ís unknown. The density of adrenergic innervatÍon to air-
ways is Iow and therefore changes in density are dífficult to determine
( Mann, 1971 ) .
The presence of adrenergíc nerve endings in ganglia of airways of
several species (Mann, !g7L; Jacobowitz, et. â1., 1973) has led to the
di scovery that noradrenal i ne rel ease i s abl e to infl uence chol i nergic
nerves and reduce ACh release (Vermiere and Vanhoutte, 1979). A loss of
such regul ation coul d conceivably lead to unopposed parasympathetic
bronchoconstrictor effects and may pìay a role in t,he etiology of asthma
but evidence is'lacking.
4) Nonadrenergic System - As mentioned previously, a functional role
of this system in promoting relaxation in humans (Richardson,1977) and
guinea pigs (coburn and Tomita, L973; Co'leman and Levy, L974) is likely,
whereas in canine airways it is absent. Since the role of the nonadrener-
gíc system is greater than that of the adrenergic system in human airway
bronchodilation (Richardson, Ig77) it is more likely to be faulty in
disease states. It has been suggested that, analagous to Hirschsprungs
disease of the colon, a similar disorder occurring in the lungs rnay lead
to a loss of the inhibitory action of the nonadrenergic system and
therefore allow for the increased airway smooth muscle responses of

- 35
bronchial hyperreactivity (Richardson and Bouchard, 1975). Evidence for
changes in the nonadrenergic system in asthma and normals are lacking due
in part to the paucity of information concerning the neurotransmitter
Ínvolved and lack of specific antagonists.
I I ABNORMAL RISPONSES OF SMOOTH MUSCLE
Abnomal responses of smooth muscle include those which involve the
postj unct,ional rnembrane. l^lhÍl e a nerve i n a di sease state rnay rel ease
more or less neurotransmitter than a normal nerve in response to the same
stimuìus, the postjunctional or muscle response itself may be different
even in response to the same quantities of agonist or antagonist.
Abnormal responses of smooth muscle may be subdivided into those vhÍch
occur due to changes at the level of the cell membrane (e.g. receptors
or ion channels) and those which occur intracellu'larly (e.g. enzyme
activities or molecular configurations). The focus of this section will
be prirnarily on those changes in contractile actÍvity which are mediated
by receptors on airway smooth muscle cell mernbranes.
l) Changes in Muscìe Mass - Patients with asthma (Takizawa and
Thurlbeck, 1971) and some with chronic bronchitis (Hossain, 1970) develop
hypertrophy and hyperp'lasia of airway smooth muscle. This increased
muscle mass capable of deve'lopíng more tension, could probab'ly contribute
to excessive airway narrowing. By analogy, wall thickness of arterioles
may a'lso contribute significantly to the increased blood flow resistance
observed in hypertensive rats (Folkow, 1971). l,.lhil e airþray smooth rnuscle
mass may contrÍbute to the chronic asthmatic it has been suggested that
hypertrophy and hyperplasia of smooth muscle are unlikely to occur in a

-36
short period of t,ime as observed in airway hyperreactivity following
vÍral infections (Empey et. ô.|., L976) or air pol'lutants such as ozone
(l{oltzman, êt. âl ., L979). }Jhil e muscle mass is an important feature
contributing to the severity of ühe dÍsease, it is likely not the cause
of the hyperreactivity o't srnooth muscle. Animal rnodel s of asthma suggest
that greater contractile responses (normalized in g/ctn?) are observed
in response to the agonist hístamíne iq yllfq (Antonissen et. â1., 1980)
and therefore implicate factors other than muscle mass in the increased
reactivity of airway smooth muscle in asthma.
Z\ Changes at receptor level - Szentivanyi (1968) proposed that
patients with asthma had reduced numbers of beta-adrenoceptors" Direct
binding studies of beta-receptors in airway smooth muscle were not
carried out, but much support for this concept came fron studies showing
reduced beta-adrenoceptor stimul ated rel axation in some asthmatics
(Cookson and Reed, L967), increased atropine-sensitíve bronchoconstrictor
responses to a nunrber of stimuli (0rehek et. ô1.,1975) changes in beta-
receptors on leukocytes (Parker and Smith,1973) and mast cells (Austen
and grange, 1975) and increased alpha-adrenoceptor mediated constrictÍon
( Adol phson et. al . , 1971 ) . Beta-adrenoceptor bl ockade produced i ncreased
bronchoconstríct,ion in asthmatics (McNeill and Ingram, 1966; Grieco and
Pierson, lgTL) but not in normals (Tattersfield et. â1., 1973; Town'ley
et. al., 1976). Beinfeld and Seifter (1980) have shown ittcreased alpha-
adrenoceptor-mediated bronchoconstriction fol'lowing propranolol adminis-
trat,ion in dogs, while Patel and Kerr (1975) demonstrated beneficial
treatment o't thymoxamine in asthmatics" Whether this indicates reduced
numbers of bela-receptors is not known without direct binding data"

-37
0thers have measured levels of cyclic-AMP in TSM and suggest that for
dogs (Rinard, êt. â1.,1979), asthma may involve lower cyclic-AMP levels
and perhaps fewer associated beta2 receptors. Reduced cyc'lic-AMP at
the level of smooth muscle or mast cell (0range, L97l; Conolly, 1970)
would potentiate bronchoconstriction, but Gold (1975) questions the
cyclic-AMP evidence on the basis of poor ternporaì correlation of broncho-
constricti on wi th ri ses of hi stamí ne i n serum fo1 ì owi ng antigen
challenge. Further studies have shown that long term treatment with beta
agoni sts may themsel ves I ead to reduced beta-adrenoceptor responses
(Busse et. â.¡., L979; Chuang and Costa, L979) while others have shown
that the long tenn use of terbutaline did not alter beta-adrenoceptor
respons'iveness (Tashki n et. ôl ., 1982 ). Recently Szentivanyi (1979),
usi ng [3H]- dihydroal prenol oì and [3H]- dihydroergocryptine to bi nd
to beta- and alpha adrenoceptors respectively, has shown that lung tissue
of patients wÍth reversible airway obstruction (asthma) had increased
alpha-receptor and retluced beta-receptor complements coimpared to persons
without airway obstruction" Receptor binding studies in the guinea pig
model of chronic asthma support the concept of a shift to an increased
number of al phaadrenoceptors and less beta-andrenoceptors in obstructive
airway disease (Barnes, êt. â1., 1980). Szentivanyi $979) also showed
that the adrenoceptor shift occurred in 'lymphocytes of patients with
asthma but not i n nomal s and that hydrocorti sone coul d restore the
normal bal ance i! yi-tt"g (Szentvanyi , et. al ., 1979) '
Kneussl and Richardson (1979) demonstrated increased a'lpha-adreno-
ceptor responses ín tissues obtained frorn patients with airway diseases.
Several studíes on humans (Kneussl and Richardson" 1979) and dogs (Bergen

- 38
and Kroeger,1980;0hno et. â1.,1981) dernonstrated a relationshÍp of
muscle tone to the expression of alpha-adrenoceptor nrediated contraction.
Therefore it appears that numbers of adrenoceptors (both a'lpha and beta)
and the tone of the preparation influence the amount of isometric tension
devel oped.
Asthmatics demonstrate a hyperreactivity in terms of bronchocon-
strict,ion to acetylchol ine or methacholine chal lenge. Whether this is
due to increased numbers of ACh receptors, decreased acetylcholinesterase
or changes in intracellular events leading to actomyosin interaction is
not known. Sensitized canÍne TSM does not show differences ín electrícal
stimulus-response and carbachol dose-response relationships compared to
I i ttermate control s i n lillq, suggesti ng that i f hyperreactivi ty to
acetyl chol i ne were present i't occurs vi a prej uncti onal mechani sms
(Antonissen et. â.|., Lg7gl. Ozone whÍch acts as an irritant may produce
airway hyperreactivity (Gotden et. a'l., 1978) and increased responses to
acetylcholine. Although ozone may have several effects, it is of interest
that ozone decreases acetyl chol i nesterase concentrations i n red bl ood
ce] I s ( Gol dstei n, et. al . , 1968 ) .
Histamine may act via a direct pathway on smooth muscle cells
(Antonissen et. a].,1980; Nathan et. a].,1979) or indirectly on a vagal
reflex (Yanta et. ô.|.,1981). Direct actions are mediated by broncho-
constrictor tl1 (mepyramine-(pyrilamine maleate) sensitive) receptors 0r
H2 (metiamÍde sensitive) bronchoclílator receptors (Antonissen et. â.|.,
1980; Nathan, êt. â1., L979). Variability o't the response to histamine
is dependant on species differences (Eyre, 1977; Fleisch, 1980), distrí-
bution of histamine receptors in the tracheobronchial tree (Chand et.

- 39
ô.¡.,1980; Eyre,1969; Eyre,1973), responses of humans (Habib et. â1.,
1,979) or dogs ( Snapper et. â1 . , 1979 ) to aerosol i zed hi stami ne. H2
receptors are also important ín providing a negative feedback to mast
cells modulatíng histamine release. It is conceivable that changes in
histamine receptor status (number, affinÍty) rnay be altered in asthmatics
since airway hyperreactivity to histamine aerosol challenge is observed
(Mathe et. ô1., L973; Townley et. â1., 1965). The author is unaware of
any studies measuring histamÍne binding or its alteration in airway
dÍsease. gur st,udies on ovalbumin-sensitized dogs demonstnate increased
isometric tension responses to histamÍne cornpared with litterntate control
tracheal smooth rnuscle in vitro (Antonissen, et. a'l ., 1980). t^le interpret
these fi ndi ngs i n terms of a homogeneous ccmp'l ement of H1 receptors í n
control and sensitized muscles with respect to their affinity for hista-
mine but that the greater response of sensitized t,issue results from a
higher ef'ficacy of the agonist or an increased number of H1 receptors
with the same affinity and intrinisic activity as controls, such that the
increased response is due to an additive cellular actíon.
3) Changes at the Intracellular Leveì - Hyperreactivity may occur
to many agents (vide sqp!"q). Alterations which may occur at the intra-
cellular level of the muscle are little known. Increases in velocity of
shortening observed for TSM from allergic dogs suggest changes occurring
at the level of actornyosÍn interactíon (Antonissen' et. â1., L979). A
compìete revÍew of possibìe intrinisic changes is beyond the scope of the
present treatment, so onìy a 'list of possibilities is provided:
I ) abnormal membrane permeabil i ty to Ca++ and other ions or
changes ín membrane leakiness (lrJeiss and Viswanath, 1979).

-402) altered tnembrane potential in dÍsease.
3) changes in Ca++ regulatory proteins such as protein kinases
and cal modul i n.
4) changes in enzyme activÍties regu'lating Ca++ ¡novements
phosphorylation reactions, activation of actin or the myosin
head, the troponín system, cross bridge formation and cycling.
STATTMENT OF THE PROBLEM
¡¡hile a great deal is known concerning neural and pharmacologicaì
control in airway smooth muscle particu'larìy the canine tracheal smooth
muscle (TSM), several areas are poorly understood. One of these concerns
is the role of adrenoceptors in mediating contractile responses, and
the relationship of the receptors to tonus of TSM and possible changes
whích may occur in airway hyperreactivity observed in diseases such as
asthma. Factors which may modulate neurotransmitter releas-e from neurons
innervating the airways is a'lso poorly understood. Present knowledge has
been gleaned primarily from btood vessels and the central nervous system.
A1 though previous studies on airways have uncovered i nteresting
interactions of various isolated agonists and neural stimulation of TSM
in vivo understanding of these interactions is poor'
The purpose of the present studies was to examine adrenoceptor-
mediated responses in isolated strips of canine TSM, and the relationship
of these responses to active tone. Electrical field stimulat,ion which
excites neural elements was considered to be a useful adiunct to the
pharmacological approach. A comparison of adrenoceptor-mediated responses
between a mongreì control population and dogs specifica'l1y sensitized

-4L
to ovalbumÍn will be made.
Several substances wtrich may act as neuromodulators of cholinergic
and or adrenergic nerves i n cani ne TSM wi I I be studi ed wi th
nonelectrically stimu'lated nerves. Changes in neurotransmitter overflow
will be measured by means of appropriate radiolabelled transmitters. It
is hoped that the physioìogÍcal importance of the factors studied can be
demonstrated by mechanical measurements of isometric tension"
The results of this study may provide Ínformation pertinent to the
understanding of neural control in airways smooth muscle"

42
METHODS

-43SENSITIZED CANINE TSM
Dogs were sensítized to ovalbumin (04) by a technique developed by
pinckard et. al. (1972) to increase 0A-specific IgE antibodies, thus re-
sulting in anaphylactic sensitivity t<t 04. Dogs irnmunized at birth and
receiving booster injections of 10 ug of dinitrophenol coniugated to
oval bumin (DNP2-0A) mixed wi th 30 mg of A (0H)3 deve'l oped high
titers of IgE antibodies specific for 0A and DNP (Kepron et. â.|., L9771-
Animals with serum IgE titers greater than 255, as determined by passive
cutaneous anaphylaxis (PCA) in non-sensitized dogs, were used for the
present studi es. Dogs wí th ti ters i n excess of 64 deve'l oped 'large
increases in specific airflow resistance when challenged by 0A given via
a nebulizer (Kepron, êt. a1., L977\-
In vitro experiments with tracheal smooth muscle strips obtained
frorn gA-sensitized dogs (6 mo. to 1 year of age) were treated in similar
manner as control tissue from the mongrel dog popu'lation. At the end of
each experiment TSM from 0A sensítized dogs was exposed to 0A (0'3
mg/ml). The 0A challenge resulted in contraction in TSM from sensitized
dogs only. The response to 0A is specific as was shown (Antonissen, et.
al., L979) by lack of cross reactivity to bovine serum albumín.
DISSECTION
Tracheal smooth muscle (TSM) was obtained from the cervical trachea
of adult rnongreì dogs (8 to 20 kg.). The animals were anesthetized with
30 mg/kg sodium pentobarbíta1 (Nembutal, Abbott) iniected intravenously,
fo1'lowi ng wlrich the trachea was exci sed and pl aced in col d oxygenated
Krebs-Henseleit solution. The anesthetized dog was then killed by intra-

-44cardial injection of saturated KCI solution (10 ml"). After removal of
adhering connective tissue, the tracheal rings (in groups of 3 rings)
ì¡rere separated and bisected through the anterior cartilage. Eversion of
the bisected rings exposed the mucosa over the smooth muscle on the
luminal aspect. Removal of the tuníca fibrosa left a clean strip of
smooth muscle wíth parallel fibers joining the dorsal ends of the bi-
sected cartilage. Incisions, para'|1e1 to the muscle fibers produced 5 to
6 strips of TSM which were of approximately equal size (averaging 0"8 -
1.4 cm. in length,0.1 - 0.2 cm in width, less than 0.1 cm in thickness
and 10 - 25 mg" in weight). These strips were dissected and mounted in
organ baths for isometric tension recording and radiolabelled neurotrans-
mi t'ber ef fl ux studi es.
ISOMETRIC TENSION MTASUREMENTS
The canine TSM strÍps obtained are ideally suited to studies of
isometric tension development due to the ìarge smooth muscle cornponent
(>75"/") and the paral 1e1 nature of the fibers (Stephens et" âl . , 1969) .
The lower ends of the muscle strips were fastened to a rigÍd aerating
tube by means of 000 braided surgical sil k. The upper end was attached
by a 'length of the surgical sil k to a Statham UC-3 f orce trattsducer
through a lever assembìy for isometric tension recording on a Gould-Brush
2400 recorder. The muscle strips were immersed in a modified Krebs-
Hensel ei t sol ution of the fol 1 owi ng composi tion (mM) : NaCl , 115; Na
HC03 , 25; NaH2P04, 1.3B; KCI , 2.5L; MgS04 "71120, 2"46i CaC'|2,
1.91; and dextrose,5"56. Aeration of the organ bath (15 ml) was pro-
vided by a 95% 02 - 5% C}Z mixture which maintained pH a'l 7.40 while

-45the bath temperature !'ras hel d constant at 37"C. Stri ps of TSM were
maintained at a resting tension of 1g (near optimal length; Stephens, et.
â1.,1969), and allowed to equiìibrate for 90 minutes prior to stirnula-
tion. A high KCI solution ft27 mM) was prepared by substituting KCI
isotonically for NaCl and added to the bath quantitatively so as to
provide the desired ç+ concentration while maintaining a final bath
volume of 15 ml.
Electrical field stimulatíon (EFS) was applied by rectangular plati-
num pl ate el ectrodes pl aced paral'leì to the stnooth muscl e stri ps in the
bath and connected to a variable-voltage 60-Hz AC source. All electrical
stimuli were app'lied at an experirnentally detennined optimum voltage (15
V set at source, see also Brown et. a1.,1980) and of duration defined in
the experiments. Electrical stimuli were given at interva'ls greater than
5 minutes to allow for recovery of the muscle and nerve.
The isometric tension results are expressed in g/cn? of muscle
cross-sectÍonal area above resting tension. Due to varia'nility of the
responses with cumulative additions of drugs it was convenient to express
the initial response as 100% and subsequent responses as a percentage 0f
the inítial value. Thís maneuver decreased variability substant,ially and
statistical testing was performed using these percentages"
Drugs used in the experÍments are listerl in table A. All drug con-
centrations expressed in the results are final bath concentrations and
stock solutions were made at a concentration vlhích was 103 greater than
desired bath concentrations so as to minimize the volume of fluid in-
volved in drug additions. Afber addit,ion of a drug to the bath suffici-
ent tinre was allowed for responses to stabilize prior to the addit,ion of
any subsequent, drug. When a washout period occurred a time interval of

TABLE A
P ryg
acetyl chol i ne
46
Drugs Used in Experiments
Abbrevi ati on
ACh
Acti on
muscarinic andnicotinic receptoragoni st
muscarÍnic receptorantagonÍ st
acetyl chol i neprecursor
ÍnhÍbits neuraluptake of NE
anti acetyl chol i ne-sterase
H1 and H2 receptoragoni st
M and D receptoragoni st
muscarinic receptorantagoni st
Al pha and Betaadrenoceptor agonist
tritiated nore-pi nephri ne
A1 pha adrenoceptorantagoni st
Beta adrenoceptorantagoni st
sel ective neuralsodÍum channel blocker
di spì aces NË fromadrenergic nerves
atropi ne ATR
[ 14c ]-chol i ne
cocat ne
eserl ne
h i stami ne HIST
5-hydroxytryptarni ne 5-HT
hyo scyami ne HYO
norepi nephri ne NE
[3n]- norepinephrine 3u-ffi
phentol ami ne PHENT
propranol ol PROP
tetrodotoxi n TTX
Source
S'igma
Si grna
NewEng'landNuc I ear
Heal t,hProtecti onBranch
Si gma
Si gma
Si gma
Si gma
Si grna
NewEngl andNucl ear
Ciba
Si gma
Sigma
tyrami ne TYR Si gma

-47
20 mínutes lvas usual ly al I owed wi th at least 3 rinses of fresh
KrebsHenseleit solution" When muscles were pretreated wÍth an antagonist
such as atropine, phento'lamine or propranolol a period of 20 minutes was
al I owed before addi tion of further drugs, S0 as to permi t
antagoni streceptor equi I ibration.
Statistical means and standard errors are included for all data.
The results for the actions of norepinephine on TSM after development of
tone ( resul ts section B) are graphical'ly presented as schematic
mechanograms. Although standard errors for these studies are not shown,
they can be obtained from the tables in the appendix" The other results
of isometric tension development are graphed as a line graph ioining the
mean values with standard error bars indicated.
STATISTICS USED
Three statistical tests used are as follows: 1) the paired T-test
for experiments excluding sections B, F and G in the resulius,2) the non-
parametric sign test for section 8,3) the Students'T-test for sections
F and G. Significant effects were determÍned at p < 0.05. An appendix
is provided for individual muscle responses for results sections A, B, C
and D.
RADIOLABELLED TRANSMITTER EFFLUX EXPERIMENTS
Tracheal smooth muscle strips of size and weight range sitnilar to
those used i n studi es of i sometric tensÍon v,,ere used . Stri ps were
mounterl in an organ bath with radÍolabelled transnrit,ter (see below) for a
period of L hour, during which a supramaxirnal electrical field stimulus

- 48
(15 V, 60 cycle, 15 sec duration) was appl ied by platinum plate
electrodes at 10 minute intervals as for isometric tension studies in
order to increase transmitter turnover and therefore prornote neural
uptake of the label. After the incubation perÍod a collection period
began which Ínvolved collecting the entire organ bath volume (10 ml) for
each 3 minute sample interval. The organ baths were filled with fresh
Krebs-Henseleit solution immedíately after each samp'le was collected.
Appropriate drugs were added for each specific protocol outlined below.
All specific stimul Í to be tested were given during the collect,ion
interval from 24 to ?-7 minutes from the start of the sampling period.
The first 3 samp'les, representing prirnarily extracellular label, were
discarded such that the first of the ten consecutive samples ana'lyzed
began at the L2 minute mark from the start of the coìlection. It was
found that a reasonably stable basal rate of transmitter efflux (as
measured by radioactivíty) occurred by the síxth sample (15-18 min") from
the start of the col I ect,ion period. After the sampl i ng perÍod al I
samples were allowed to dry over low heat (100"F) in a fume hood for 3
days after which 7 ml of scintillation fluid (Beckman, HP scintillation
cocktail) was added to each sample vial and radíoactivity was measured on
a Beckman LS-350 scintillation counter. CountÍng efficiency vvas 55% for
3¡ and g0"A for 14C. After the last sample the muscles were
removed, weighed and solubÍlízed Ín scintillatíon vials to wltich 250 ul
NCS (Amersham/Searle) were added. The NCS, a surfaceactive organic base
dissolved the tissues over a period of 6 days wt¡ich freed radiolabelled
transmitter in nerve endings. Scintil lation fl uid vvas added to the
muscles following this solutlilization and radio-activity ín counts

-49per minute (cpm) determined as for previous sampìes. A wide range of
radioactivity remaining ín muscle was found, from approximately 2,000 cpnt
to 100,000 cpm. The reasons for this are not known but may be an
indication for the variability of nerve density in canine TSM. In order
to express the release of radioactivity for each sample Ín a normalized
f ashion the rate coeff ici ent (Shanes and Bi anchi , 1959) v',as used. The
rate coefficient was obtained as follows:
number of c rel eased er mi nute ers le x 100num er cpm rema n ng n e musc ea e
start of the sample used in the numerator
This rate coefficient resul ts are expressed in units of % 'Eissue
radioactivity released per minute. It is a good method of norma'lizing
the wide variability obta;ined for cpm released and those cpm remaining in
the muscl e.
Rate coefficients were used to plot efflux in order to graphically
i I I ustrate the effect of the treatments 0n radioì abel I ed transmi tter
overfl ow. An íncrease in rate coefficient therefore indicates an
increase in radioactivity efflux. Efflux curves were obtained by calcul-
ating means for the fÍrst 5 samples (min" L2-24 in the sanp'le period) for
al I the rnuscl es in the same experÍment such tha'ü al 1 49 muscl es for
exampl e, in the t3Hl- norepi nephrine effl ux experiment were used to
calculate these means. The functíon which best described these 5 means
was found to be a pohrer curve. For all curves calculated the correlation
coef'ticient, was r > .96. Frorn the power curve, the number of sampl es
which were used to obtain the means and a theoretica'l t ít was possible
to calculate the 95% confidence lirnÍts about the efflux curve. Fron the

-50power curve function expected values for sarnp'les from 27 to 39 minutes
were calculated and the confidence limíts extended around these poínts.
This allowed a comparison of treatment effects for the interval (24-27
mínutes) to be compared with the expected value by use of a t-test. 0n'ly
the peri ods i n whi ch the treatments !',ere appl i ed are compared wi th
expected val ues.
Control curves, to whích no stimulus was given, were compared with
the expected calculated control curves in order to support the validity
of calculated values. Figure 28 and 36 illustrate these findings for
t3Hl- norepinephrine and t14Cl- acetylcholíne efflux respectively.
Four different radÍolabelled transmítter efflux studies were carried
out. Specifics of each are described below, whi'le the generaì methods
described previous'ly apply to al I of these experiments"
t3Hl NonTPINEPHRINE EFFLUX
The incubation period was carried out in the presence of t3Ul-
norepÍnephrine, specific activity 13"4 Cilmmol at a concentration of
1É M. At I radi ol abel I ed drugs t3Hl- norepí nephri ne or [14C]
choline were obtained from New England Nuclear, Boston, Mass" Aft'er
i ncubation wi th [3H]-norepi nephri ne and el ectrical stimul ation as
prevÍously described, cocaine was added to all muscles to a final bath
concentration of IÛ-.7 M and maintained throughout the sampling períod
(Nigro and Enero, 1981). Isometric tension was recorded thorughout the
experiment.

- 51
[14 c]-ncETYLcHoLINE EFFLUX
TSM strips were incubated with t14 cl choline (1É M and
specific actÍvily 2-5 mCi/mol) for L hour and electrical stimulation as
described previously. Choline rather than acetylcholine v',as used since
only chol i ne Í s taken up by nerve endi ngs which then synthesize
acetylcholine. By electrical stimulation the turnover of acetylcholine
( ACh ) i s í ncreased and radÍ ol abel I ed chol i ne becomes part of the
vesicul ar pool of acetyl chol i ne. [14C]-acetyl chol i ne overfl ow
experiments were undertaken in L) control muscles (done in the absence of
eserine or tetrodotoxin) 2) in eserine (1É M)-treated muscles
(eserine added at the end of the incubation period and throughout the
experiment and 3) in muscles treated with eserine (1É M) and tetro-
dotoxin (TTX, 1É M) throughout the sampling period. Efflux curves
and tests of significance urere determined as for [3H]-norepinephrine
efflux studies" Simultaneous isometric tension measurements were made
only for [14 Ci-acetylcho]ine efflux in the absence of eserine and
TTX.
NOREP I NEPHRINE OVERFLOW
The demonstration of norepinephrine release from adrenergic nerves
is dependent on showing overflow of intact norepinephrine from a stimu-
lated (i.e. electrically) preparation. The present studies with those of
others fulfil these criteria for electrica'lly stimulated canine TSM iI
vÍtro. Foster and 0'Donnelt (1975) demonstrated that adrenergic nerves
i n gui nea pig trachea take up t3Hl noradrenal i ne and that 3H-ttlE i s
dep'leted by electrical stimulation" When 6-hydroxydopamine was used to

-52destroy adrenergic nerves both [3H]-noradrenal i ne uptake and effì ux
foìlowíng electrical st,imulation were reduced. I^lhile not all the radio-
labelled -NE enters nerves the great majori'Ey ís processed via uptake 1
and subsequent stimulation results in an efflux of the labelled trans-
mÍt¡er (Russell and Bartlett, 1981). Studies using canine TSM have also
shown (Russell and Bartlett,1981) that the majority of the efflux is
intact norepinephrine with a lesser quantity of norepinephrine meta-
bolites. Venniere and Vanhoutte Í979) have applied thÍs informatíon to
the study of adrenergic nerves Ín dog trachealis, and shown an increase
of t3Hl norepinephrine released following an electrical stimulus.
In the present studies the release of intact norepinephrine from
adrenergic nerves ín canine TSM was demonstrated using electrical field
stimulation (EFS, L5 V, 60 H7, AC, 3 minute duration) and high perform-
ance liquid chromatography. The total bath volume (?-0 ml) for each
sarnpl e period was subjected to catechol ami ne extraction fo'lì owi ng the
procedure of Hallman et. al. 1978. The catecholamine extracts v{ere
purified on activated alunina, followed by electrochemical detectÍon
using a Spectra-Physics SP8700 solvent de'livery system linked to a LC-44
amperønetric detector (BioanalytÍca1 Systems, Inc.) on an oxidat,ion mode.
The column used was a RPISSPHERI-5 reverse phase (Brownìee Laboratory)
and results recorded on a Hewlett-Packard Integrator 33904.
Table B demonstrates a large Íncrease of norepinephrine overflow
during EFS during sample period 2, while no increase of epinephrine over-
flow is noted" After the EFS, norepinephrine release continues to rÍse
in sample Ínterval 3 accompanied by a srnall increase of epinephrine. The
reason for the continued release of lrlE followíng the stimulus interval is

-53
not known but Ìs likeìy related to a diffusion de1ay. Similar results
are noted for the 3H-norepinephrine overflow studied (rig. 29). The
present study confirms that the use of 3H-norepinephrine tracer can be
used to determine qualitativeìy norepinephrine release frcrn adrenergic
nerves of canine TSM (Russell and Bartlett, 1981; VermÍere and Vahoutte,
1e7e ) .
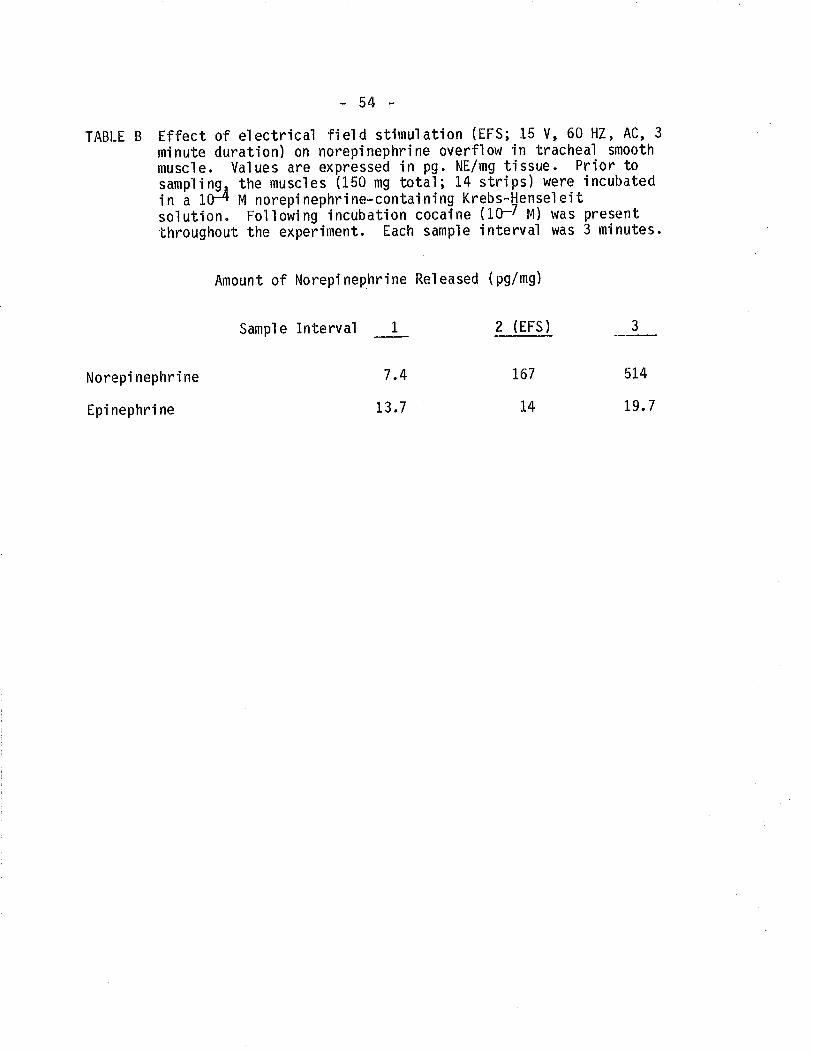
TABI-E B
Norepi nephri ne
Epi nephri ne
-54Effect of electrical fietd stímulation (rfS; 15 V, 60 H7, AC, 3minute duration) on norepinephrine overflow in tracheal smoothmusc'le. Values are expressed in pg. NE/mg tissue. Prior tosamplÍng, the muscles (150 mg total; 14 strips) were incubatedi n a 1tts+ lvl norepi nephri ne-contai ni ng Krebs-$ense1 ei tsol ution. Fol I oi,vi ng' i ncubatíon cocaÍ ne ( 10-l M) was presentthroughout the experitnent. Each sample interval was 3 tninutes.
Amount of Norepinephrine Released (pg/mg)
Sampì e Interval 1 2 (EFS)
7.4 r67
13.7 t4
3
514
L9.7

- 55
RESULTS

-56A) El ec'Erical Fiel d Stilnul ation of canine TSM, in vitry
EletrÍcal field stimulation (15v, 60 Hz, AC, is sec. duration)
applied to tracheal smooth muscle strips showed stable reproducible con-
tractile responses to repeated stimulation" Addition of physostÍgrnine
(eserÍne) 10-7M augrnented the response to EFS to L36% of the initialvalue and the further addition of hyoscyamine (10-5M), still in the
presence of eserÍne reduced the contraction ta 4"/" of the initial (Figure
1 ) . Responses to el ectricat ti el d stimul ation (EFS ) al so showed a 24%
declÍne in isometric tensÍon development in the presence of phentolamine
(10-5M). Addition of tetrodotoxin reduced the response to z% of the
initial e]ectrically stirnulated contraction (Figure 2). Figure 3 indÍ-cates a spontaneous increase of 6+4% over the first 5 isometric tension
responses to EFS which t,'/as not significant and due mainly to increased
responsíveness of two of the nine trachealis strips in the study (Table 3
appendíx). AddÍtion of propranolol (10-5M) non significantly reduced
the response to EFS to 97'14% whereas the further addÍtion of phento'la-
mí ne (10-5M) , stil I i n the presence of propranol ol , signi ficantly( p<0.05) reduced the contraction to 64+g% of the initial response
ft00%, Figure 3).
Addi tion of norepi nephrÍ ne (ruf 10-6M) was wí thout effect on
responses to tFS, but the further addition of NË (10-4M) reduced the
response of four trachealis strips and was without effect 0n the ot,her
one, resultÍng in a mean value of g5+72 (Figure 4). This reduction was
sÍgnificant (p<.05) with the use of a non pararnetrÍc sígn test (used to
eval uate a trend i n response) . Addi tion of propranol o1 (10-5M) and
NE (10-4M) further reduced the response to B5+6% of the initialva'l ue.
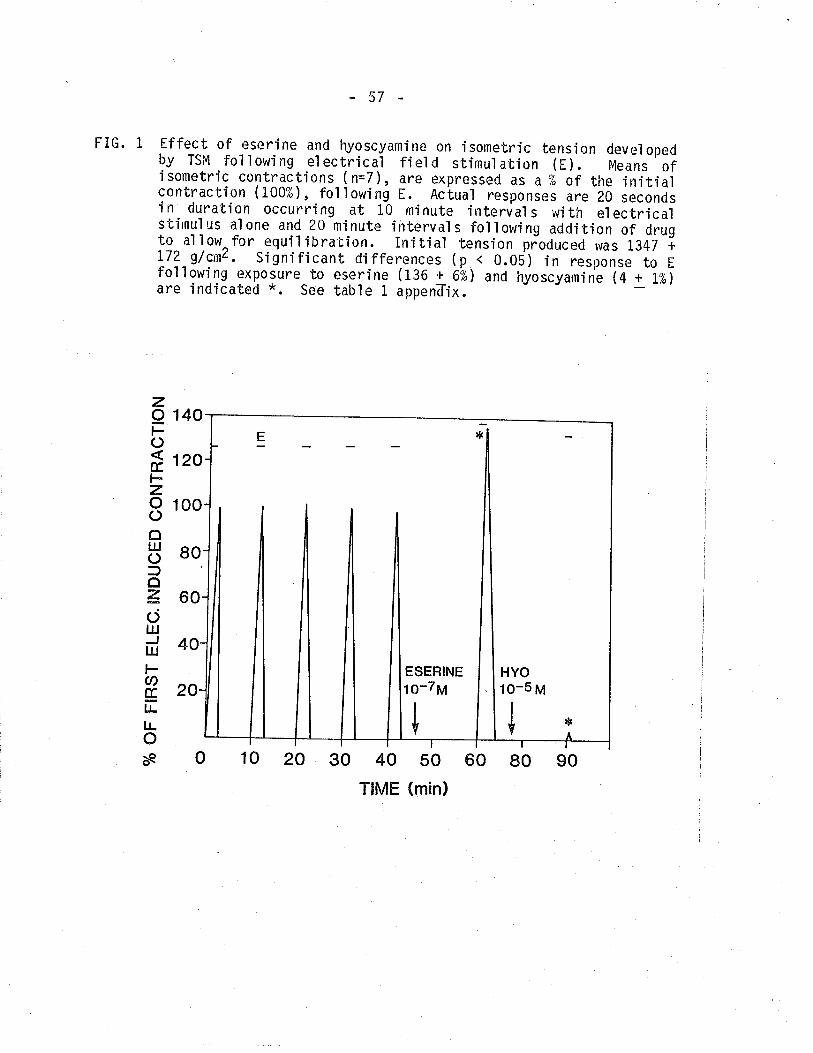
FIG.1 Effect of eserine and hyoscyamÍne on isometric tension develonedby TSM fol 1 owi ng el ectrical fi el d stímul ation ( E ) . N.unr-'órisometric contractions (n=71, are expressed as a % of tire iniliÀlcontraction (100%), fol'lowing E. Actual responies are Z0 seconclsin rluration occurring at 10 mÍnute intervals with electriJàlstirnulus alone and 20 minute inter.vqls following atidi.bion of druglg^ul'low,,for gquil ibraLÍon. Initiar tensíon pñoduced was
-tli,l-+Ll2-glcn¿. signíficant differences (p,< 0.05) in response to Efo]]owi.ng exposure to eseríne (136 + 6bl") and hyoscyamine (-+ + y7¡are indicated *. See table 1 appenTÍx.
57
0 10 20 30 40 50 60 8o 90
TIME (min)
1
zooEf-zooouofo4oIU
IU
F-U)trLL
LLorOo\
120
140
00
BO
60
4
2
E
*
*
ESERINE10-7M
HYO10-5 M
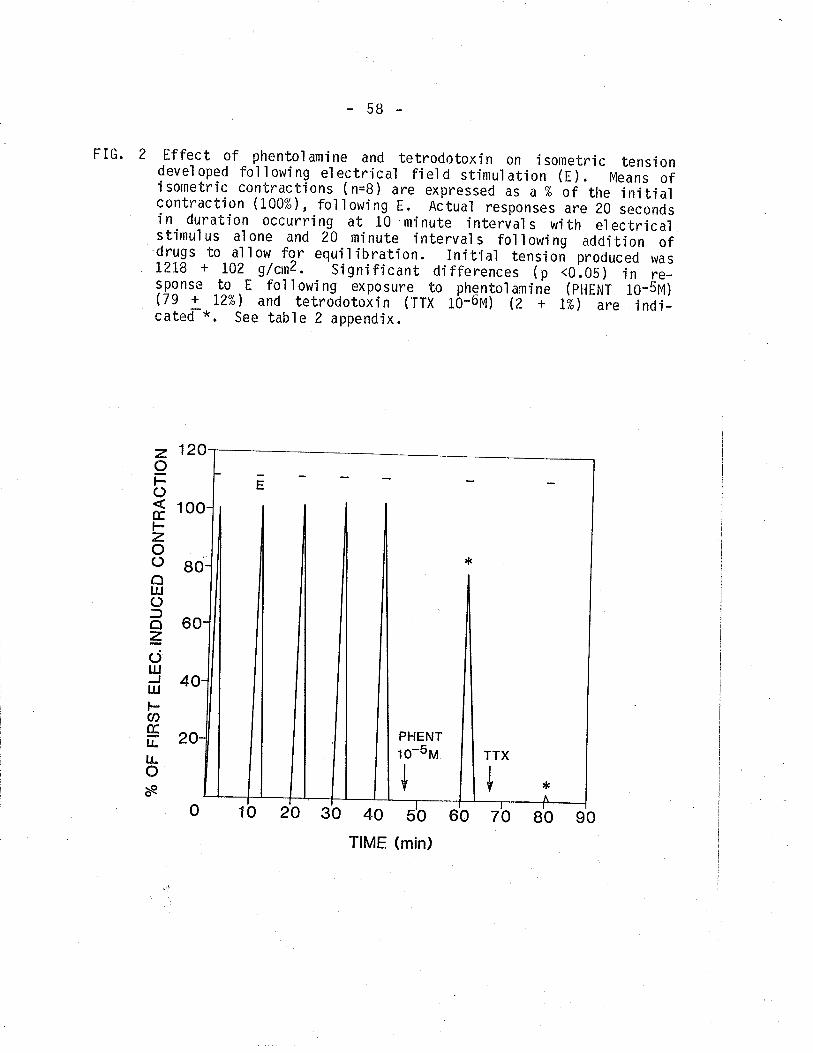
FIG. 2 Effect of phentolamine and tetrodotoxin on Ísometric tensiondeveloped fo'l1owÍng electrical field stímulation (Ei. -Neans orisometrÍc contractions (n=B) are expressed as a% oi ilre initiarcontraction (100%), following E. Aitual responses are 20 seconds
i n duration occurri ng at l0 mÍ nute i nterval s wi th el eiiricalstirnulus alone and z0 mÍnute Íntervals following aoãiiì.ón of!rugs to allow for equilibration. Initial tensioñ pro<luceJ wasrzLB + L02 s/-crf,. .'significant differenc.t fp-iolosj-"in re-sponse to E fol 1 owi ng exposure to phqnto'l amine (pHENT 10-5M)(79 + L2%) and tetrorlotoiin (rrx 1ö-6Mi (2 +- rà/")
-ãre -inoi-cated-*. See table Z appendix.
58
30 40 60 70 80 90
TIME (min)
120
1
I
é-otrOEzooo[rJO:o7oI,U
IJJ
þct)frltIJ.osOo\
60
o 10
2
Ë
*
*tTTX
{
PHENTto-5lr,t
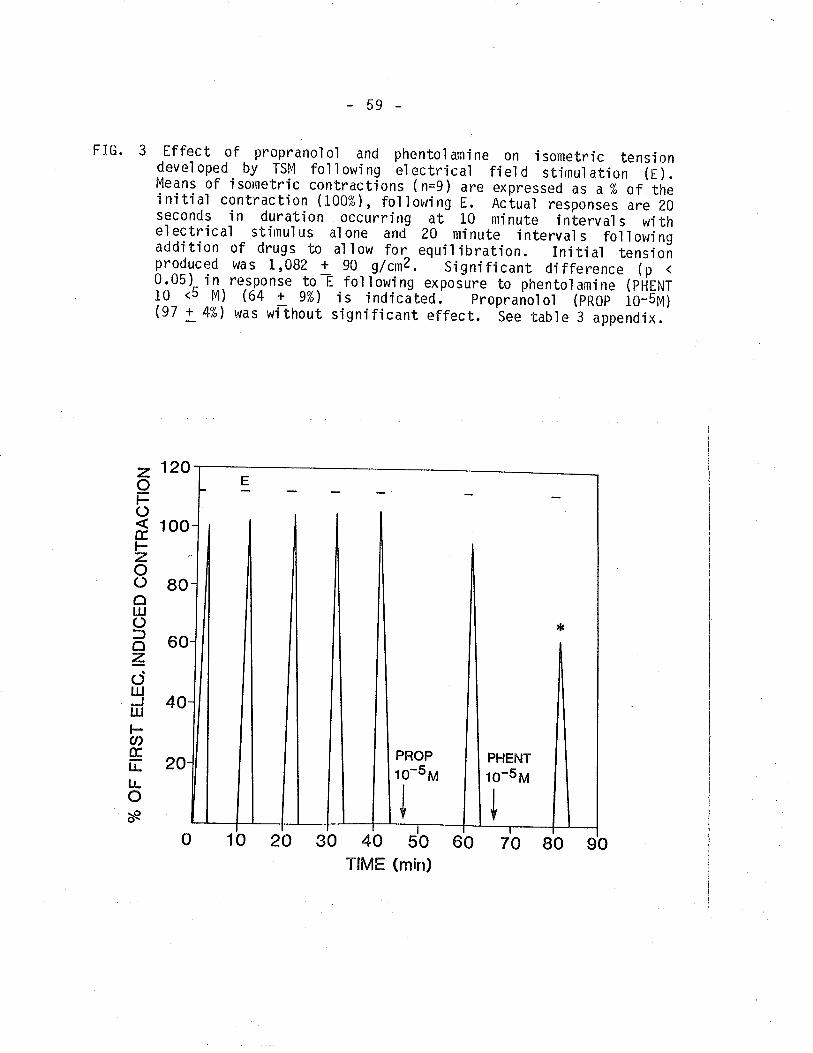
FIG. 3 .tffect of propranol or and phentol amÍne on isometric tensiondevel opqd . by ISl4 fo'l 1 owi ng el ecrrical fí el d stiinul ation- ir ) .Means of isornetric contractions (n=9) are expressed as a % of thei nitial contraction (r00%), foilowing E. Actual rÀtponi.i áre zoseconds in duratÍon occurring at 10 minute interval s wi thel ectrical stirnul us al one and zo minute interval s fol ì ovri ngaddÍtion of drugs to allow fo1 equilíbration. Initial tensionproduced was 1,08? r_ ?0 -p/cnz. SignÍticant difference (p <
9:05L in. re.sponse toT folr-owing exposure to phento'ramÍne (ptie¡¡r10 <5 M) (ø+ 1 e%l i s i ndicãted. nroprunoior-- (pRöp ib::5r'ìi(91 + 4%) was without significant effect. see table 3 appendix.
59
30 40 50 60 70 BO ITIME (m¡n)
120
1
6
oo
zoC)
E,þzoooLUofozoLU
tut-ØtrTL
tLorOo\
80
40
2
o 10 20
E
*
PROP
1o-5MPHENT
ro-5n¡
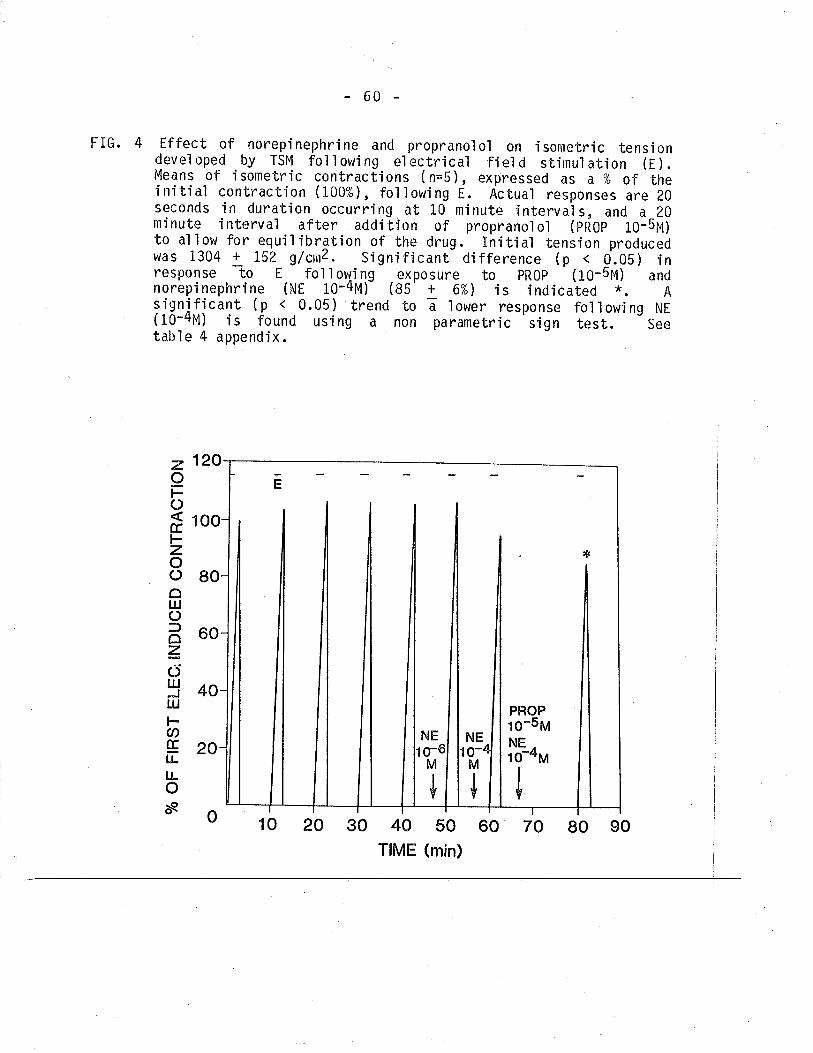
FIG. 4 Effect of norepÍnephrine and propranolol on isornetric tensiondevelope_d by TSM followíng electricar tierd stirnulation (E).Means of isometric contractions (n=S), expressed as a % of'üheinitial contraction (100%), follovring t. Àctual responses are 20seconds in duration occurring at 10 minute intervals. and a z0minute interval after addition of proprano'lo'l (piop 10-5M)to al_lo_w for equil ibration of the drug. Inítial tensíon prorJucedlvas 1304 + L5?- 9/cnt?. Significant dÍfference (p . g.OS) inresponse -to E foì 1 ovli ng exposure to pR0p ' (i0-5M) andnorepi nephrÍ ne (itle 10-+M) (85 + 6%) i s i ndicated *. Asignificant (p < 0.05)'trend to ã lower response fo1ìow'ing Nt(10-414) i s found usi ng a non pararnetric ' sign test. seetable 4 appendix.
60
10 2A 30 40 50 60 70 80 s0T|ME (min)
120
0
80
20
0
1
zoËoEt-zoooLUo:fozö[rJ
TU
t-U)EtLLLo:oo\
60
40
E
M
ü
M
ü
*
NENE66
PROP1o-5MNEro-4¡¡

-61 -value. The trend of increasÍng tension following repeated EFS was mainly
because of exaggerated responses produced by two of the five muscle
strÍps used, and was not significant (Figure 4).
B) Effect of Tone on Adrenoceptor Mecliated Responses in Canine TSM
Unstirnulated tracheal smooth rnuscle strips showed no contractÌle or
rel axant response to norepi nephrine (f ojrq to f o-4rql or tyramine
(10-5U and 1ÉM) even fo] I owí ng beta-adrenoceptor bl ockade wi th pro-
prano'lol (10-5M) . However when i sometrÍc tension u,as deve'l oped to
high-K (22.8 mM) exposure, the subsequent addition of norepi nephrine (tttE
1É - lÉM) aì ways produced a further contractil e response wi th a
mean maximum rise to L7% greater than the response to K alone (flg.5). A
further increase in NE concentra'Lion to tO-5m resul tecl in partial
relaxation whÍch reached a maximum at 6L% of the response to K. Phento-
lamine (tO-5U) further enhanced this relaxation to ?9% (at 10 min) of
the response to K. l^lhen phentolamine uras present throughout the exposure
to NE (flg.5), the results show a re'laxant response to NE with a thres-
hol d at tOjN and maxímum at 1f5M. The response to tf4N Nt
consisted of a small non significant increase in mean tension vuÍtich could
be abolÍshed by the addítion of more phentolamine Q x LO{N).
In order to examine the possibi'lity of a cholinergíc contribution to
't,he increased tensíon deve'loped in response to low doses of NE (in the
presence of 22.8 mM K), atropine (lÉM) uras introduced at the p'lateau
of the K-induced contraction (príor to the admínistration of graded
increases of NË). Figure 6 illustrates the cholinergic conponent of '[he
K contraction in that the tension declined by 20% after treatment with
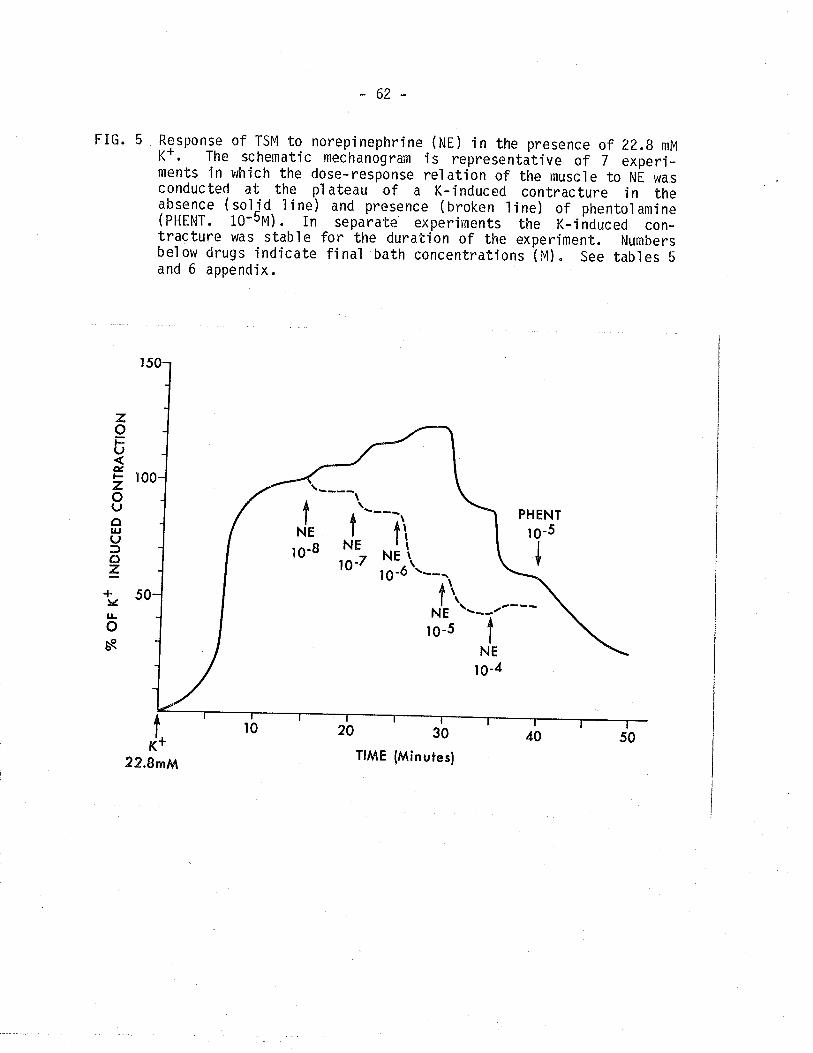
FIG.5 Respons.e of rsM to norepinephrine (NE) in the presence of zz.g mMK'. The schema.tic. mechanograrn is_ representative of 7 experi-ments in vrhích the dose-response relation of the muscle to Ì{Ë wasconducted at the p1 ateau of a K-induced contracture in theabsence (so'ljd lin_e) and presence (broken line) of phento'lum.,tne(PHENT. 10-5M). In sepaiate experiments th; K-Índuced con-tracture was stable for the duration of the experiment. Numbersbelow drugs indicate final bath concentrations ïiq). See tables 5and 6 appendix.
I IPHENT
to-5NE
to-B NE
rc-7 +
62
NE
lo-ó
r5
zotr(Je,Fzo(,ôt¡¡(,Þôz
+}¿rLoñ
I6+
22.8mM
r0
to-5 INE
1o-4
4020 30
TIME (Minutes)50
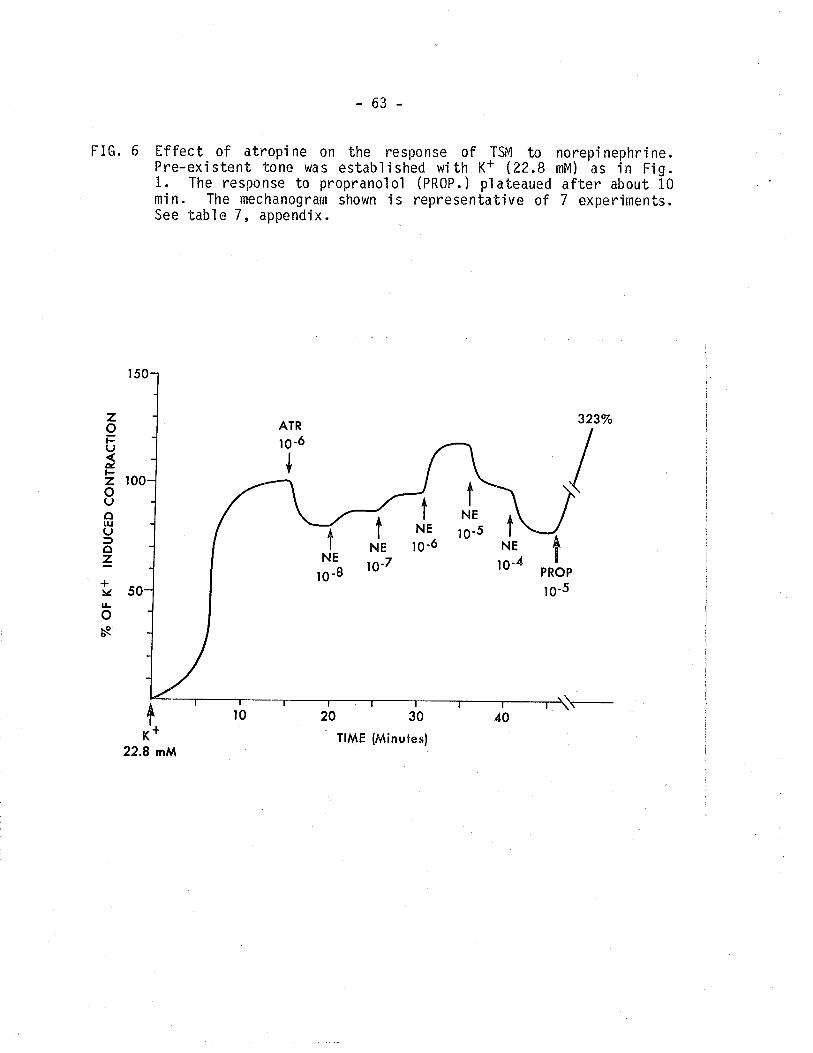
FiG.6
63
Effect of atropi ne on the response of TSM toPre-exi stent tone was establ i slred with ç+ (,?2.81. The response to propranolol (PR0P.) pìateauedmin. The mechanograrn shown is representatíve ofSee table 7, appendÍx.
norepi nephr i ne.mM) as in Fig.after about 107 expe rirnen ts .
INE
9-óI
NEg-7
00
zot-Ud,
zoL,ôll¡L,fôz+V
oÈS
l5
22.8 mM
ATR
lo-ó
{
323Vo
I
NE
lo-B
NE
lo-5 INE
lo-4PROP
to-5
+J-K'
r0 20 30
TIME (Minutes)
40

-64-atropine. The subsequent contractile response to NE (1É to tO{N)
however was not a'ltered following atropine treatment, reaching a mean
maximum of 18% greater than the K-induced contraction. Relaxation pro-
duced by NE (lÉM) r¡,as reversed by beta-adrenoceptor bl ockade wi th
propranoloì (10-5M), resu'lting Ín a mean isometrÍc tension at 323% of
the K-induced contraction" Due to large dífferences in tension develop-
ment (normalized in g/ctn?) between muscles in both group (responses to
NE in the presence and absence of atropÍne) qua'lÍt,ative rneasures of the
responses were conpared by use of the non-parametrÍc sign test. There-
fore, although the schematic mechanograms are based on the means of the
various muscle responses, it is the qualitative aspect (i.e. relaxation
or contraction) which is the major focus of this study.
When the same muscl es (f ig. 6 ) vlere exposed to atropi ne (10-5M)
and propranol o1 (1fsM) 20 minutes prior to K-stirnul ation (Fig. 7 | the
response to NE (1É to 1ÉM) was not biphasic but increased pro-
gressively to a maximum of 257"1 greater than the K-induced contraction.
The contractile response to NE v^Jas reversed to relaxation at all doses of
Nt (1É - lÉM) by al pha-adrenoceptor bl ockade wi th phentol ami ne
(1ÉM). The threshold dose of NE (1ÉM) was not altered by aìpha
or beta-blockers.
Tyramine (10-5N and 1ÉM) , whÍch acts to di spl ace NE from
adrenergic nerve terminals was shown to produce an increase in isometric
tension in addition to that produced by K (22.8 mM). This contractile
response was not blocked by atropine but was enhanced by propranolol and
bl ocked by phentol ami ne (tn5N, Fig. 8 ). To test the possibìe di rect
effects of tyramine on post-iunctional receptors the effectiveness of
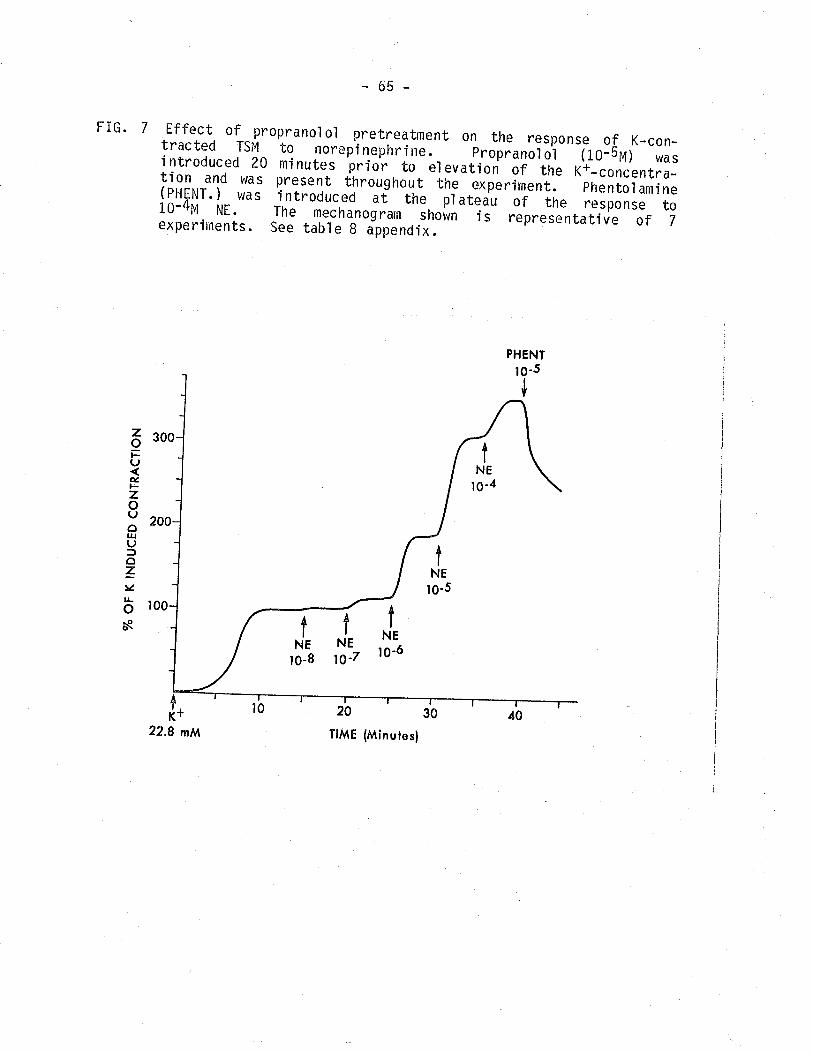
FIG. 7 ,Ef
fe.ct. of -
propranol o] pretreatrnent on the response of K-con-tracted TSM l:-.l:i.p'inephrine. -proó.unbiãi-""iro:ór)'-""u,
introduced 20 minutes 'prio'r to elevatioi ðÏ the K+-concentra-tion and l/"as present ihrougho.ut th_e experirnent. phentol ami ne(PHENT. ) was 'introduceà uT" Û,. pratãaï' äî rhe response to10-4M Nt. Jh. . n'ã-.h*ogrum
..stroirn- -ìi-
räþresentarive of 7experiinents. See table g ippenoix.
-65-
NE
ro-ó
PHENT-50
t
INEg-4
I
200
zot-(J
d,FzoL.,
ôl¡¡(,fôz\¿¡¡-oÈS
INE
to-5
I INE
t0-8NE
to-7
r0 20 30
TIME (Minutes!
4022.8 mM
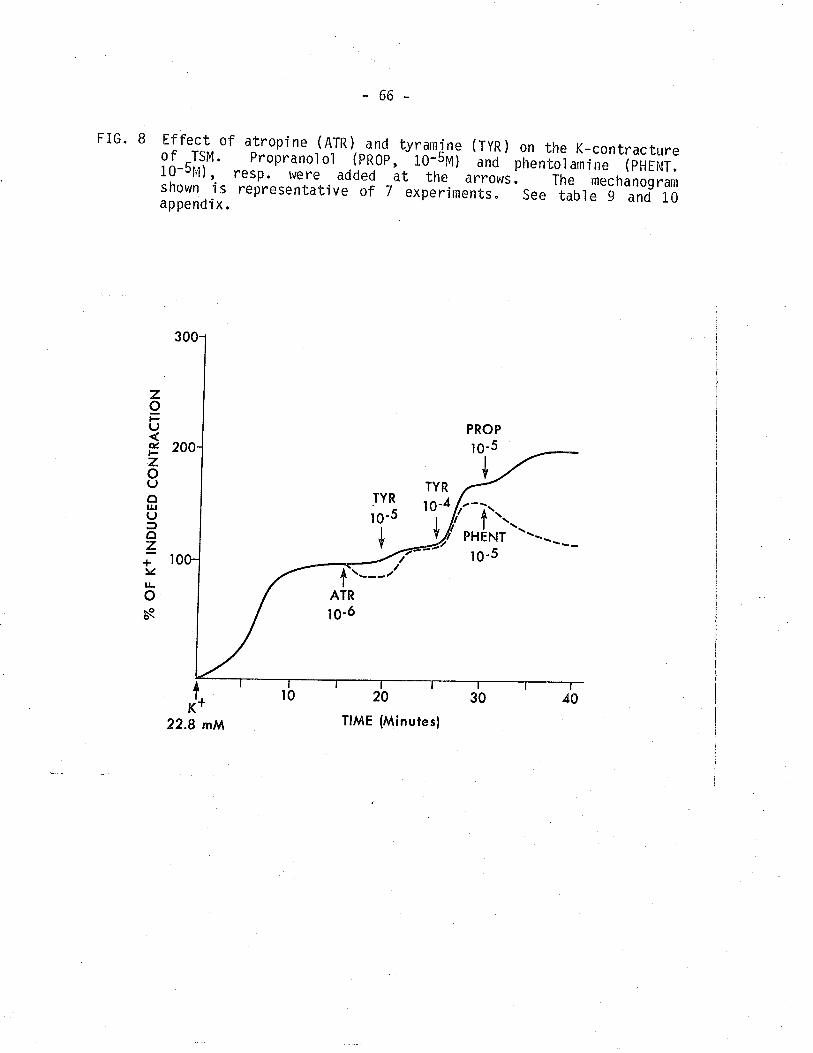
-66-
FIG.8 EIf._*. of atropine lRTR),_uq tvrarnÍne (TyR) on the K-contractureof -TSM. prooranoror .(pR0p, to-5Ni unJ'plåntoramine (pHENT.
19-oNl,. resp. were added ât the a.ronsl"-' The mechanogramshown.is representative of 7 experirnentJ,--see table 9 and 10appendix.
zot-Iu,FzoL)ôlUL)Jôz+
Itoñ
3
200
+TYR
lo-4IYRro-5
++
f
PROP
ro-5
PHEI{T
lo-5
30
tATR
ro-ó
tK+
22.8 mM
r0 20
TIME (Minutes)
40

-67 -this agent was examined in cold stored rnuscles. After 3 days of cold
storage (4"C) TSM showed dirninished response to tyramine (1É or
1ÉM), whil e responses to exogenous NE were not markedly affected
(data not shown).
In order to examine the relation between the level of pre-existent
tone and the magnitude of al pha-adrenoceptor medÍated responses, atro-
pi nized (1ÉM) muscl es were exposed to various IK+]o prior to the
addition of NE. At 48.2 mM K+ Ehe contractile response to NE (1É
106M) was greatly reduced in spite of the fact that the cholinergÍc
component could stÍll be seen (fig.9). The relaxation response to NE
(1ÉM) was markedly reduced (rig.9). This relaxation was propranolol-
sensit,ive as was observed for lower IK+]0 (rig.1 and 21. The con-
traction produced by NE (1É to 1ÉM) at the p'lateau of the
K (48.ZmM) contraction t,'ras enhanced after pretreatment wi th propranol o'l
(1h5N, Fig. 7it, but was much less than that observed at ìower
IK+]o (22.8mM, Fig. 7). Similar experiment,s were conducted over the
range of [K+]o from t7.4 to 64.2 mM and responses to Nt (1É to
tn4N, Fig. 10) are expressed as the absol ute 'tension above or bel ow
the K-induced contracture at the respective [K]0. It can be seen that
the opt,irnal [K]o for demonstrating the a'lpha-adrenoceptor mediated con-
traction, both in the presence and absence of propranolol was 2?"8 nM"
Increases Ín K systematically potentiated the relaxant component and
ínhibíted the a1 pha-adrenoceptor-mediated contractions; thus, whil e
1É M NE produced contraction at the lower IKJe, it produced relaxa-
tion at the higher [KJe.
In order to test whether the biphasic response to NE was specific to
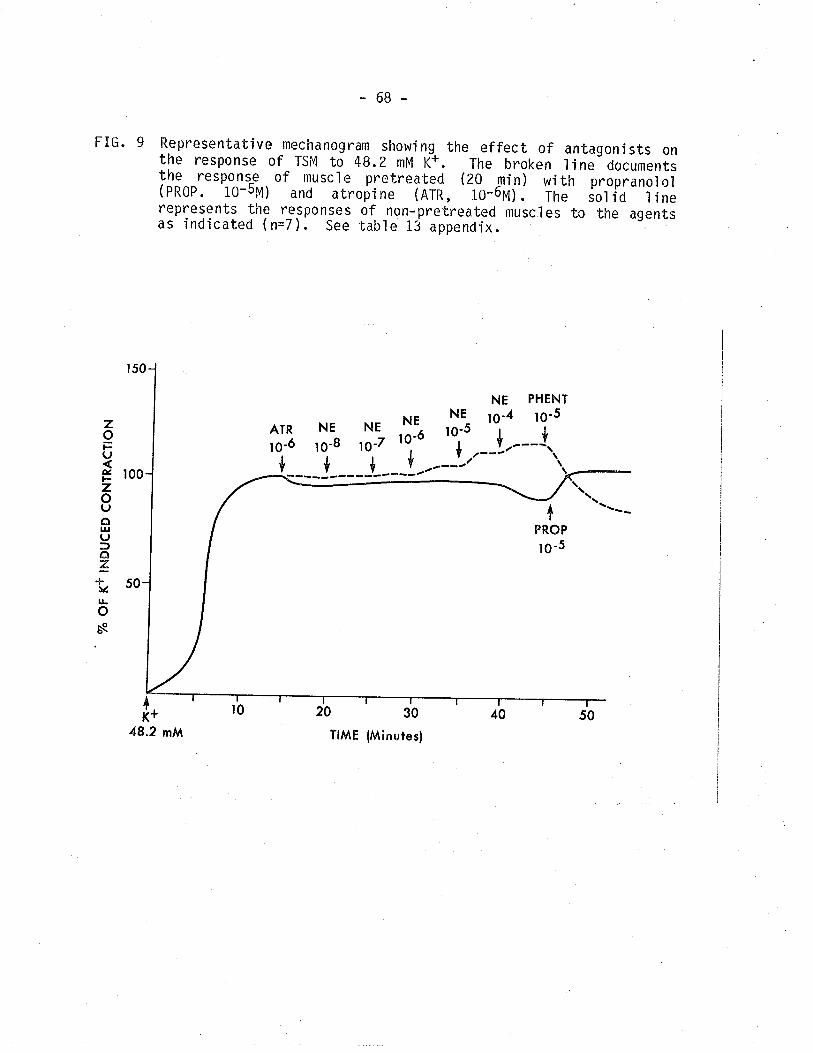
FIG.9
r50
-68-
NE NE
ro-8 rc-7+ü
Representative mechanogram showing the effect of antagonísts onl!,. response of TSM to 48.2 nrM ç+. The broken I i ne documentsth. respons_e of m.uscle pretreated (20 min) wÍttr p.opränoìol(PROP. 10-5M) and atropine (ATR, t0-6Mi. The sol id I inerepresents the r_esponses of non-pretreated muscles to the agentsas indicated (n=7 ) . See tabl e 13 appendÍ,r.
t00
zoË(,d,l-zoUôl¡¡UÐôz
+v¡!oñ
ATR
lo-ó
NE
to-ó
+
NE
lo-5
+
NE
to-4
+
PHENT
lo-5
+PROP
l0-5
50
46+
48.2 mM
20 30
TIME (Minutos)
r0 10 50
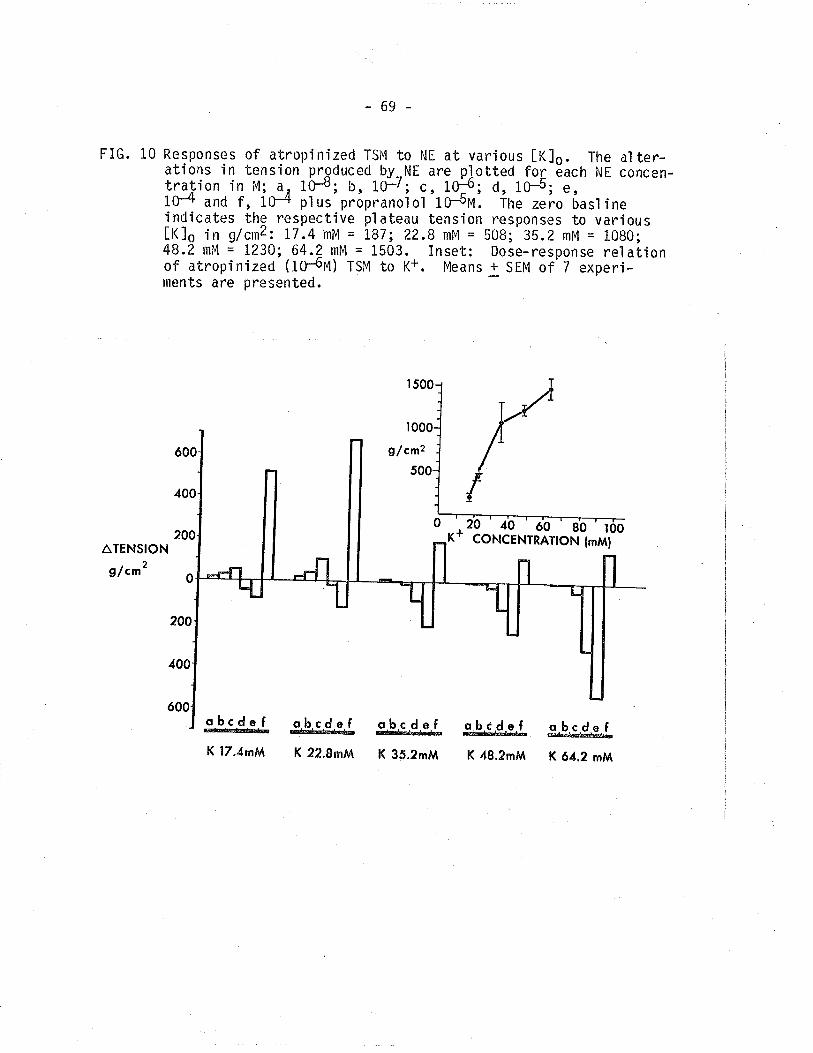
69
FIG. 10 Responses of atropinized TStvl to I'lE at various [K]0. The al ter-ations ín tension prpduced by-Nt are plotted fof ðach I,lE concen-tration in ivl; a, 1(F; b, th/; c, lûj; d, 1h5; e,1É and f, Ífa ptus'própranolol'tnsN. The zero baslineindicates the respective p'lateau tension responses to various[K]o in g/anz: 17.4 mM = 1E7; 22.8 nM = 508t'35.2 mM = 1080;4B.2 mM = 1230; 64.2 ml'1 = 1503. Inset: Dose-response rel ationof atropinized (10áM) TSM to K+. Means + SEM of'7 experi-rnents are presented.
ó00 g/ cm2
0 80 I200 K+ CONCENTRATION lmMfATENSION.2glcm
obcdef@
0
200
400
K 17.AmM
obcdef.&iÀerb!E
K 22.BmM
o.b,c.d.e,f
K 35.2mM
obcdefd*[E*#
K 4B.2mM
qbcdef-*'+e*ù
K ó4.2 mM

TABLE 1
AGONISTINITIALTENSION(gl cnz)
NE
to-6ptNt
to-SN
70
CunrulatÍve dose response relatÍon to NE (IÉ to ldM)following developrnent of active tone (initial te¡sion, g/cn?).Values for Nt and after propropranolol (PROP 10-5M) addÍ-tion are expressed as the percent of the initial contractíon.All muscles whích were contraçted with potassium (K+) werepretreated with atropine (tO-6m). Means + SE for 7 muscles foreach agoni st.
% OF INITIAL TENSIONNE NE
tr€u thTMNE
io-4tvrPROP
thsM
K+ 17.4mM 187+55
K+ 22.8mM 480+140
K+ 35.zmM 1080+263
K+ 4B.2mM 1230+82
K+ 64.2mM 1503+144
Histamine 106M L6B+62
ACh 5x10 8M 449+80
L07+4
103t8
10010
10010
100r0
11616
102+1
1 18+6
109.9
100.1
100.0
100+0
1 16+2 3
L06+2
L24+L6
L?I+?tL
100l1
9B+1
97+L
28+10
93+9
B0+1 5
97+2L
96+4
e1l3
79+3
2L!12
65+18
66+1 4
74+î!L
8S++
8215
65+6
35+?!L
59+1 9
494+L25
24I+97
t27+IL
LL0!2
LL0!?
925+L7 6
t66+29

-7L
K-i nduced tone both hi stami ne (lÉM) and Ach (s x tù-sN) were used.
Table I indícates that the biphasic response is observed with all three
agents and the maximum a'lpha-adrenoceptor mediated contraction is depen-
dent on the amoun'ü of resting tone. Fig. 10 indicates that maximum
alpha-mediate<l responses occur when tone Ís deveìoped to a K+ concen-
tra'bion of 22"8 mM. lrlhen the sarne responses are expressed as a percent-
age of the plateau tension deve'loped to the agonist the maximum percent-
age increase occurs where active tone developed is least, (compare per-
centages in the presence of histamine and propranolol with those in high
-R, 64.2mM and propranolol, Table l).Figures 11 and 13 illustrate the biphasÍc response to norepÍnephrine
(rue 1É to 1ÉM) in TSM contracted wíth acety]choline (ncrr 5 x
1ÉM) and hi stami ne (1ÉM) respectively. The change froln contrac-
tile to relaxant responses however, occurs at a lower concentration of Nt
( lÉM) i n the presence of these two agonÍ sts, than that for an equi va-
I ent K-induced contraction (to{N ruE, fig. 5). Addition of proprano]ol
(10-5M) reversed the rel axation in both cases (Figs. 11 and 13).
l^lhether the biphasíc response deve'loped with norepÍnephrine was ín
some way unique to ç+ stimul ated contractions rvas further studied.
Si nce a speci fic ant,agoní st for potassi um i s not avail ab1 e, Ít was
possible to test the effect of abol ishÍng ínitial tone produced by
acetylcholine (S x tf4N) on the adrenoceptor nrediated contraction.
A<tdi tion of hyoscyarni ne (1ÉM) el imi nated the contraction produced by
acety'lcholine (s x tnSN) and NE (lÉ to 1É) in TSM pretreated
with propranolol (to-5N, Fi9 " Lz) Addition of hyoscyamine (1ÉM) to
histamine-pretreated (1ÉM) rnuscles in the presence of NE ancl prop
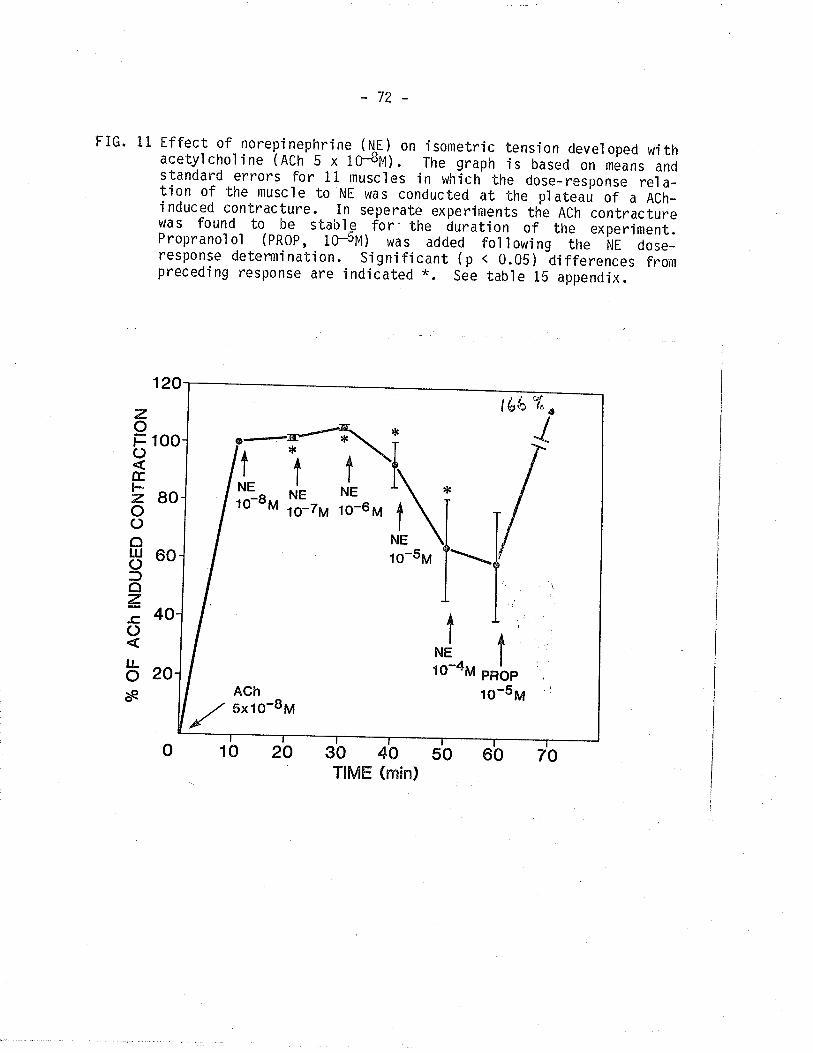
FIG. 11 Effect of_norepinephrine ([E) on isometrÍc tension developed withacetyl chol ine (Ach 5 x tÉu) . The graph is based on ,näans andstandard errors for 11 muscles in whÍch' the dose-response rela-tion of the muscle to Nt was conducted at the -plateaú
oi u Ãcn-induced contracture. In seperate experÍments ilìe ACh contracturetvas found to be stable for the äuration oi the.^p..i*Àñt.Propranolol (pR0p, 10-5M) was added fo1'lowing the NE dose_response determination. Significant (p < 0.0S) differences frompreceding response are indicated *. see table ls appendix.
120
0 10 20
72
30 40 50TltuîË (min)
60 70
100
BO
60
20
zooEt-zo()oUJo:)o4oLLorOo\
4
**
NE *(
1 g-8
I
/
166 "í. u
J
NE
10-5M
ACh5xt0-8M
IhtE
io-4M
A
I
PROP
ro-5Af
M lO-7n¡ 1O-6M
*
11NE NE
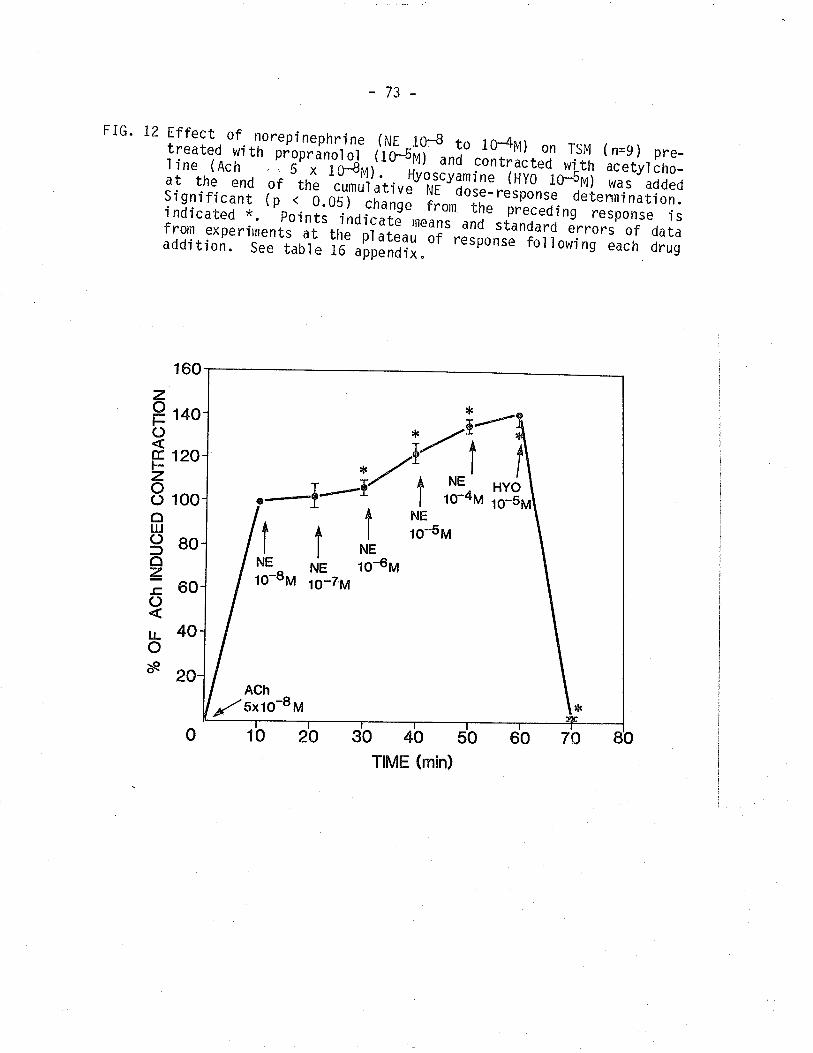
- 1')tJ -
FIG. 12 Effect of..norepinephrine ,tNE ,lo--8 to ,9;]ryl
, ol T.SM (n=9) pre_rffårii.iírh erSpîäi'H .rro-sNi unå'.åntracteo *iti, u..tvrcho-a r rhe une or -thi å,"Ji!ii,lrî',.råil::"Jlif;j_ïúi.m;.llS;:signÍficant (p . o.oll'cnangã i.or-"iñ*
.preceding response isindicated *. 'points ìn¿t.utã-,n.unî"aii stanuar¿ errors of dataIååi.iJå:'îfJï,r1f 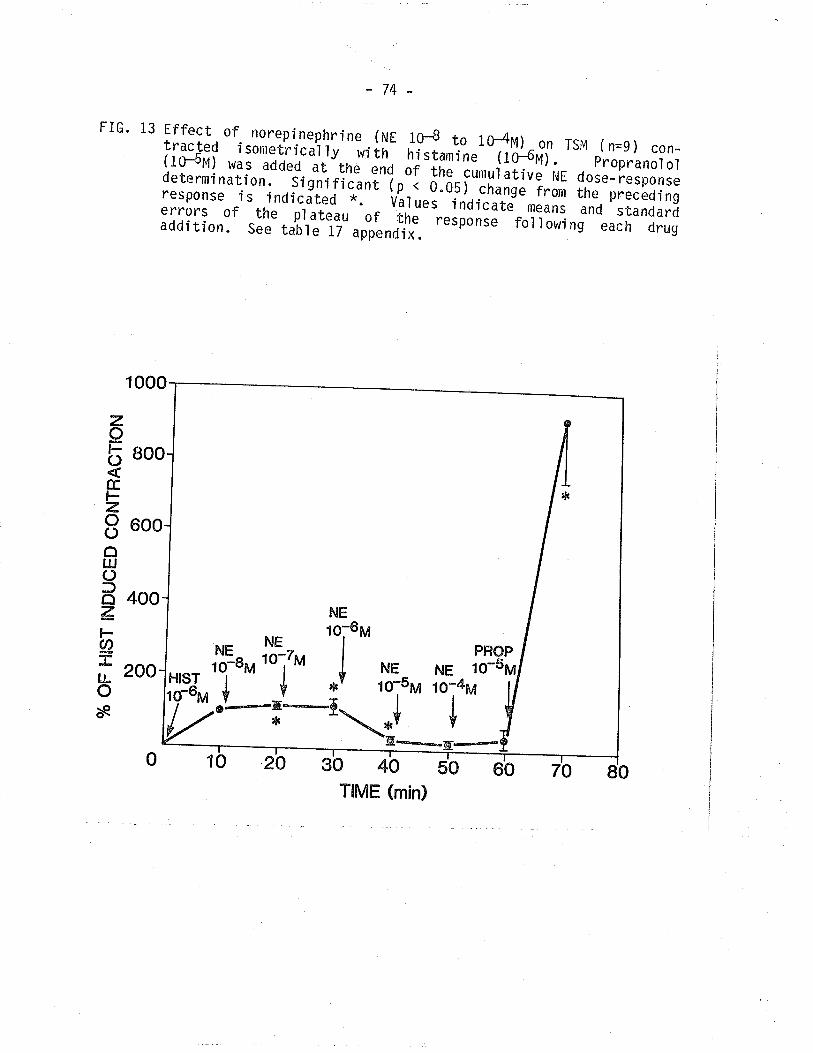
FIG. 13 tffect o1 -lorgUi.nephrine (NE 1É to 1tracled
i:oru,tri.ui ry wî rh nirtu*ì.î.^î[L,oi. trr ',.î;îln!îi;
(tn5N) wg: added aL"rhe 9nq ,i-ü;'ii*urarive NE dose_responsedererminar.ron. signiriåàni j.o_i tj.'osi".r,uÀg.'irä,n- irî."'p....,rinsresponse i s i ndÍcared *. _ vãl ,u, -in¿ù;i;-d;;,; äî srandard
;ääiii,,:t tl[' ,,l|Xtîï .Jot ;t'l-.- '.iöäÅ'u roiìäwì nf "-ea*r
drus
-74-
30 40TüME (min)
7o$ eoo
1 000
0 2A
EF-zoootUOÐozt--aILo:oo\
600
400
200
1 50
*
PROPro-5n¿
+@
,t
NE NE
NË
NE ro¡onr
'o+'* -JNEro-8H¡
to-5n¡ to-4m
.+ {
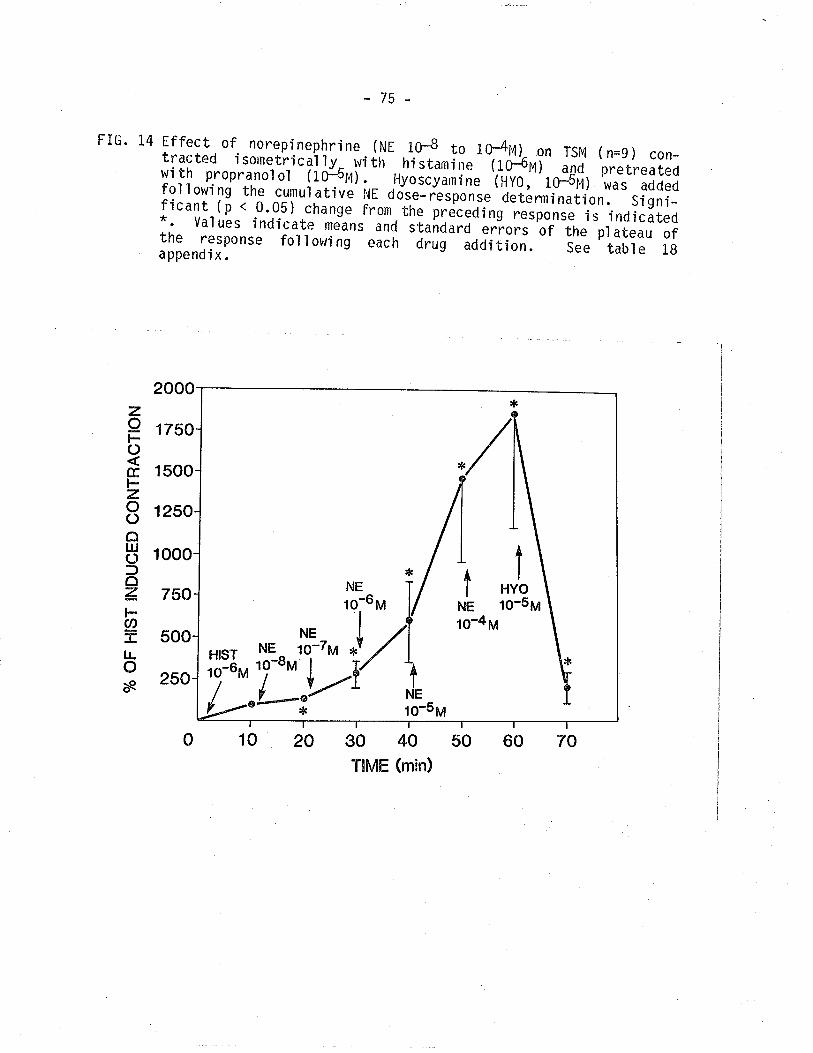
FIG. 14 Effect of. norepinephrine- (NE iÉ to tO-4N)- on TSM (n=9) con_tracted i sornetricg] ly. wt tn ni stamiñe (lÉM) and pretreatedwi rh propranol oì (r Ési'r) . nv.rðv;;i;. 'tHvo, '' rdsr,il Tu, addedfo1 'lowi ng the cumul ative I'lE ¿<ise-íeìü;t9 detänni nuiïonl si gnÍ-f icant (p t .0'0.5) change from, ilre pr-Jceä-ing ,.rponse is indicated*' values indicate mðans and standard errors of the plateau ofthe response fo1 'l owi ng each d;;g ã¿¿i tion. See tabl e 18appendix.
0 10 20
-75-
30 40TIME (min)
50 60 70
0200zIF.ofrt-zoC)atUC)foz.t-U):rTLoa
1750
1 500
1250
100
750
50
250
1o-5M
,1.
*
*I
*
NE
10:6MHYO10-5MNE
tO-4H,t
lå1,* JNE
1o-8MHIST
10-6M

-76-ranolol reduced the tension drarnatÍca11y fro¡n 1864 + 726% to 205 + 83% of
the initial histamine-Índuced (100%) contraction (rtg. 14). Table 1B
(appendix) indÍcates a wide variabí1ity in the responses to hÍstanrine
(lÉM) and addi tions of NË (1É to tÉM) pt us hyoscyami ne. when
expressed in g/cnz the resul ts ( tanle 1g, appendix) índicate that
hyoscyamine relaxed those muscles to the initial tension developed in the
presence of histarnÍne (tO-6N).
C) Adrenoceptor Mediated Responses in Canine TSM Sensitized to0val bumi n ( 0A )
studi es of the bi phasic response to norepí nephri ne (lÉ to
lÉM) and its dependance on pre-exi sting tone were carrÍed out Ín
canÍne TSM from an a'l 'lergic model of ast,hma" Responses of TSM muscle
from dogs sensitized to ovalbumin (04) were compared to those frorn con-
trol tnongrel dogs. Since maximum alpha-adrenoceptor-mediated contractions
were observed with a potassium concentration of 23mM prior to NE (lÉto 1ÉM) exposure, the same protocol t¡¡as used for sensitized tissue.
The results reveal no significant differences in the biphasic response
(tti9. 15), maximun a1 pha-adrenoceptor-mectiated contraction fo'l'lowing
proprano'lo'l (tf5N) exposure (fig" 16), maximum beta-adrenoceptor-
mediated re'laxation (Fig. L7) or responses to tyramine (Figs.18 and 19).
Responses of a mongrel control popu'lation are simil ar quantítative'ly and
qualitatively to those of OA-sensitized canine TSM with respect to adren-
oceptor-mecli ated responses, when tone v'las f i rst el eva'üed wi th ¡1+
(23mlt1) . Varying the concentration of K+ to 48.2 mM resul ted in a shi'Ft
fro¡n a1 pha-adrenoceptor-medíated contraction to beta-adrenoceptor
medíated relaxation; thís shÍft was not significantly different from that
in controls (Table 2).
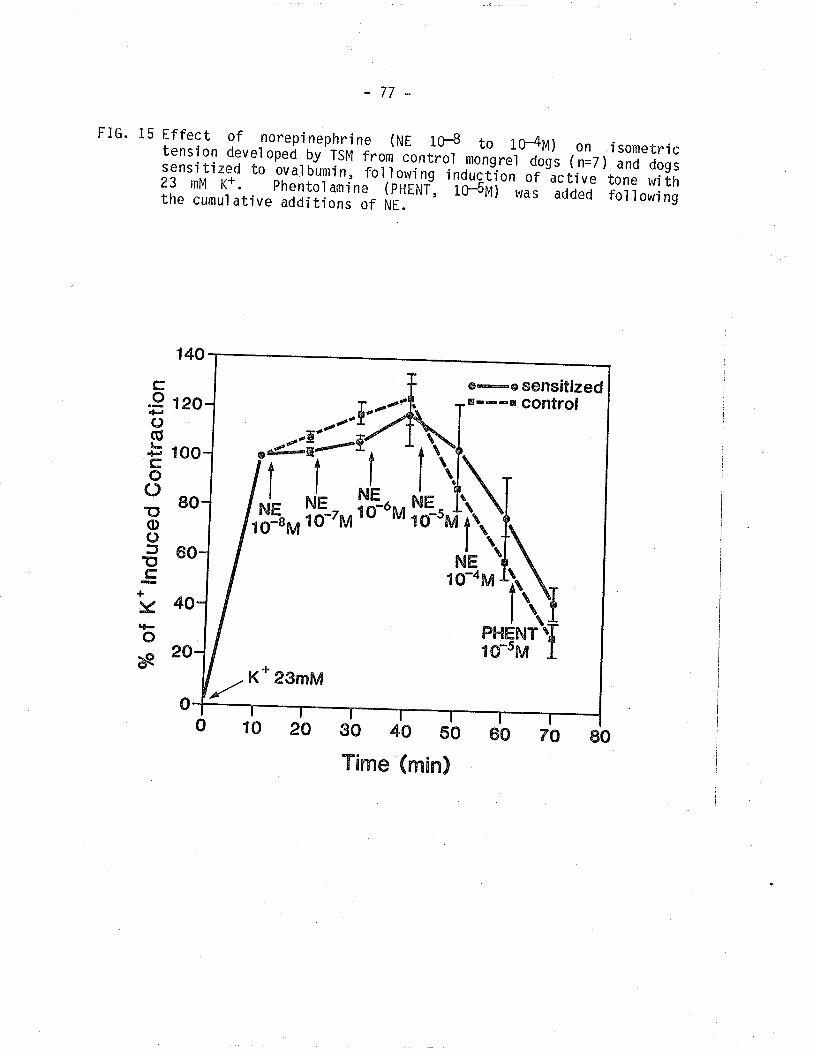
-77 -
FIG. 15 Effect 9f norepinephrine (ue lÉ to lÉM) on isometrictension developed by TSM from .ontrol rnong.rel Oogs (n=Z) anO OogssensÍ tized to ovar bumÍn, for,ì_ogi né Irjr.tion of active tone wÍ thîi. ii_å:.r,,.01åälîlåïl'3. rpHËn¡i",
'iËi¡,- *ii ;;;ï roiio"iñö
140
c.aC)6Lcoc)ÐooËç
+vl'l-o:oeÞ
20
o0
1
1
80
60
40
20
o0 10 20 30 40 50 60 70 80
Time (min)
K" zSmM
T 0-sM
/
E
***iI
N
10-8MNE
6æ@ sensitlzeds-EEñr COntfOl
NEt o-óMNE
1 0-7M
NE1 0-4M
FF{ENT1o-sM
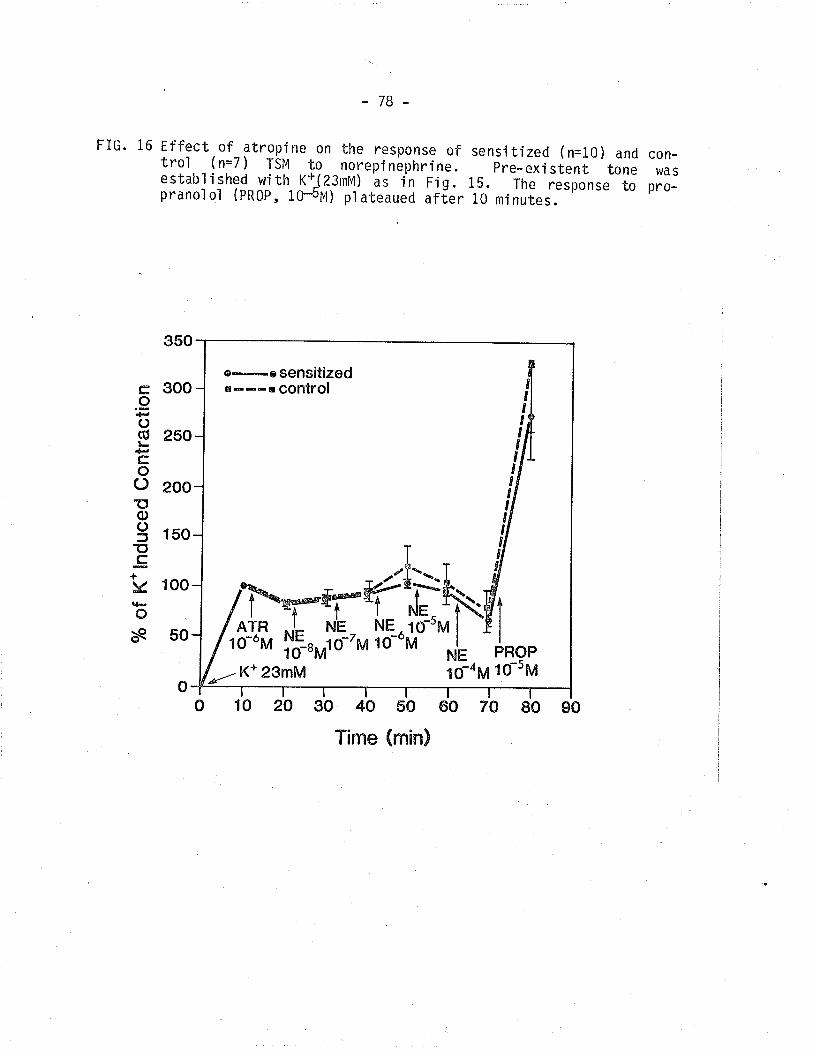
FIG' 16 Effect of-atropine on the response of sensilized (n=10) and con-trol ( n=7 ) TSM to norepi näphrÍne. pre-exi stent tone wasesLabrished with K+(23mfrr) .as ín rlg. 15. Th; .*ponr. to pro-pranol o] (pR0p, r0-5M) pl ateaued afier I'o rinutes. -'- - .--
7B
o102030405060708090Time (m!n)
c.oo6cooEooEc+
o:oo\
300
200
350
250
150
100
50
o á/t-
I,IIt
,
I7
ATR1 0-óM d
K+ 23mM '10-4H¡
@_--e sensitizedEeEEB COntrOl
PROP1o-sM
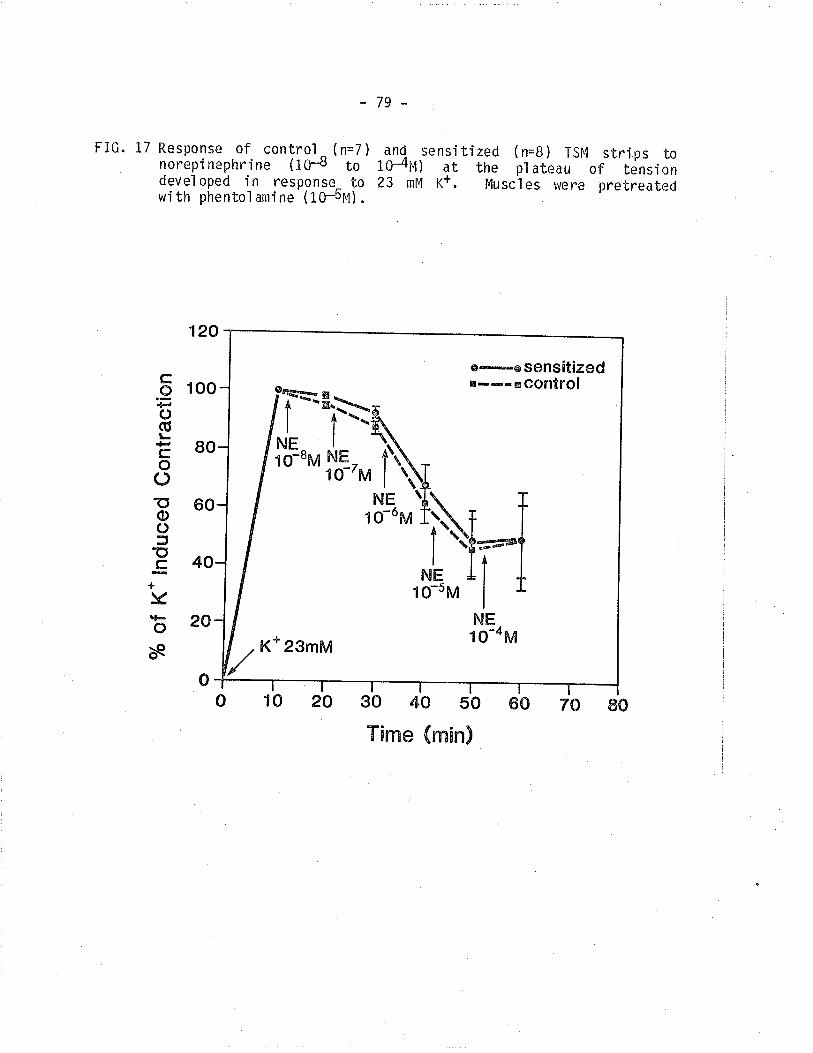
-79-
FIG. 17 Response of control _ (n=7)norepinephrine (1É todevel oped i n response_ towi th phentol arnÍ ne (tû-bN) .
anq sensitized (n=B) TSM1ÉM) at the pt ateau?3 mM K+. Muscies were
stri ps toof tensi on
pretreated
120
4A
20
c.9C)6L
ËoOõooEc
+vo
rOo\
100
BO
60
0o10203040 sCI 60 70 80
Tlrne (¡nin)
NE
K+ 2gmM
tO-BM NE-1 o-rM
6:sgÇnSitizedEEeq a COntnOl
NE'to-óM
NE1 o-sM
NIE10-4M
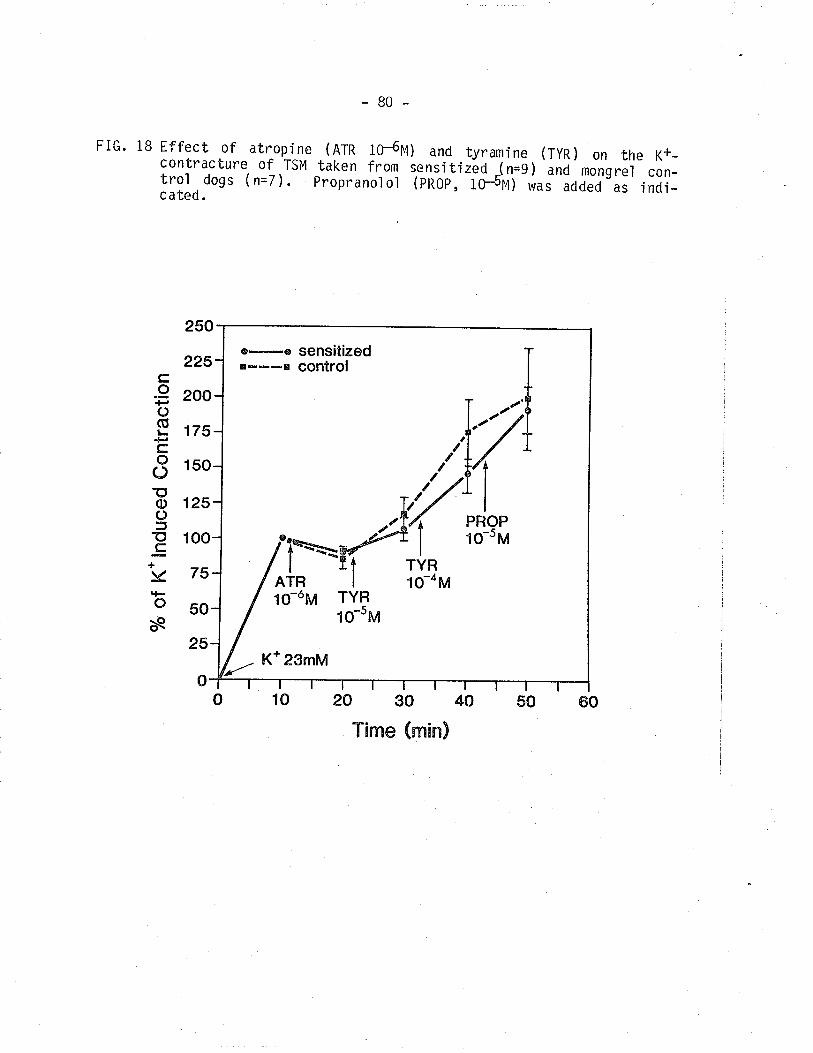
FIG. 18 Effect of atropine (nfn lÉM) and tyramine (TyR) on the K+_contracture of TSM taken from sensÍ tizé¿ ,-(n=9 ) 'anä'mJngret
con_tro'l dogs (n=7 ). propranot ot (pR0p, iO--SM) -*ur "ã¿älî''
u, indi_c ated .
-80-
0102030405060Time (min)
coo(ú
coC)þc)C)
õs+Vo
oìR
002
250
225
175
150
125
100
75
25
50
oK* zgmM
o@@ sensitizedEEEEa control
TYR1o-4MATR
10-óM TYR1o-sM
PROP10-sM
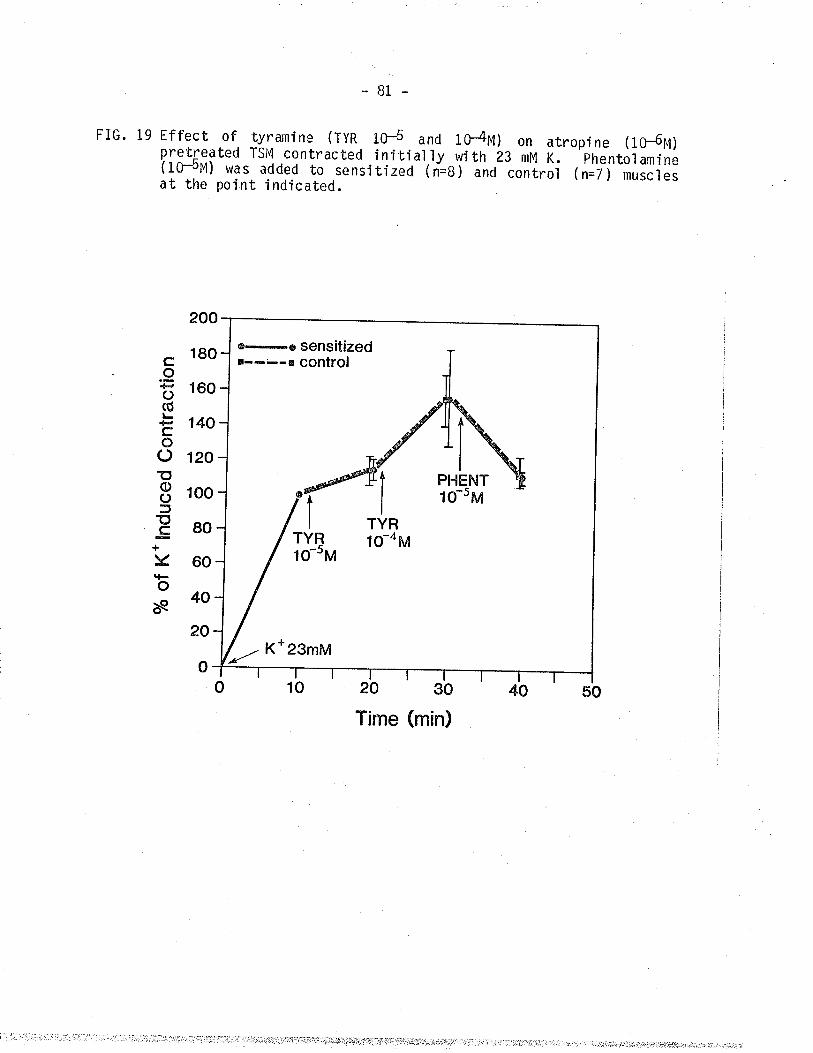
FïG. 19 Ef fect of LyramÍne (TyR 10-5 and tÉM) on atropi ne (lÉM)prefrea Led Tslvl contracted i ni t.i al ìy wi th 23 mM K. 'phentol
ami ne( 10-bM) rva s added to se nsi tÍ zed (ÉA j ãn¿ controi t n=/l musc I esat the point indicated.
-81
20 30
Tlme (min)
40 50
200
180
160
140
coo6coottoo5E-Ë
+
o:oo\
120
100
80
60
40
20
oo 10
TYRto-5n¡
K *
2gmM
e-6 sensitized8--;rE COntfOl
TYR10-4M
PHEhIT1 o-sñd
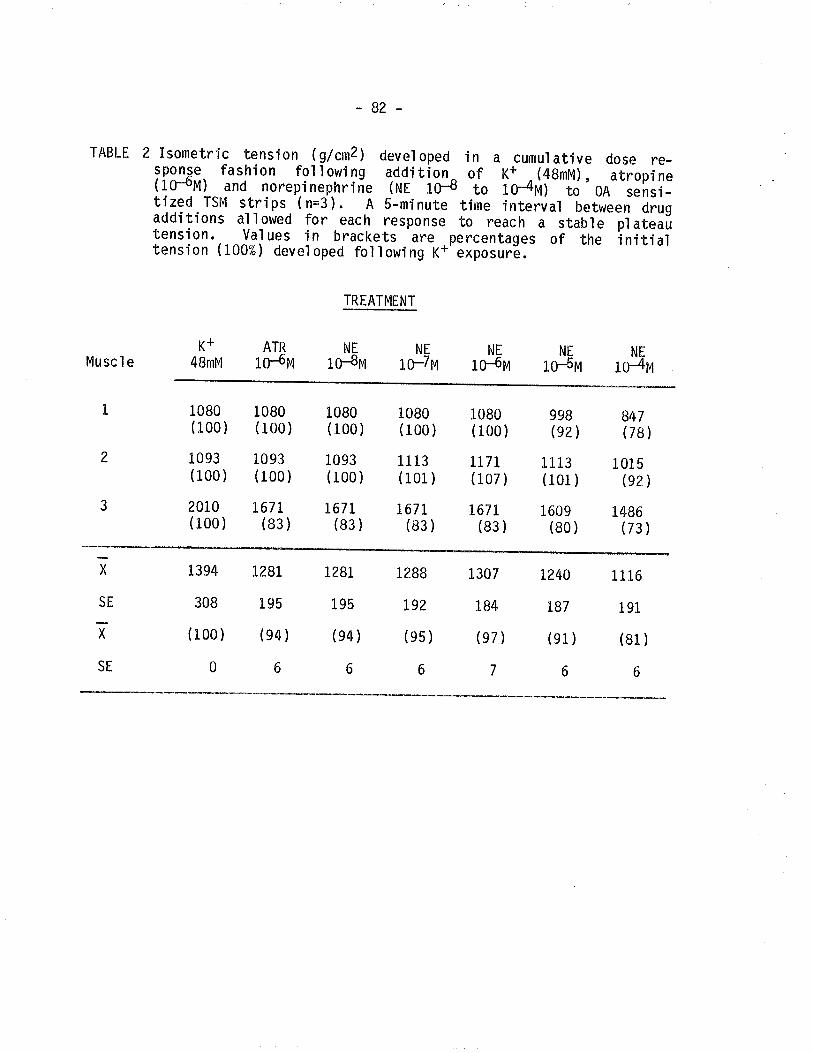
TABLE
Muscle
2 IsometrÍc tensíon (g/cn?) developed in a cumulative dose re-
¡issfiL :i'ol''ii,".;il];il1ifl- iï1"l3%'å iþ,Íi'g' oo..!ållf:tÍ zed rslvl stri ps ( n=3 ) . A 5-mi nute time i nterval between drugadditions allowed for each response to reach a stable ptui.autensÍon. val ues in brackets are . percentages of the initialtension (I00%) devel oped fol I owi ng f+' exposuié.
82
TREATMINT
¡+4Bmttl
ATR
rcÉplNE
to-{NNT NE
tnSN thTMNE NT
10 5M to-4u
1
2
3
1080(100 )
i093(100)
1080(100 )
1093(100 )
L67L(83 )
1080(100)
1093(100 )
L67L(83 )
1080(100)
1113(101)
L67L(83 )
1080(100)
L67L(83 )
998(e2)
1609(80 )
847(78)
1015(e2)
1486(73 )
LLTT(107)
1113(101 )
2010(100)
X
X
SE
SE
1394
308
(100 )
0
LzBL
195
(e4 )
6
L28L
195
(e4 )
6
1288
192
(e5 )
6
1307
184
(e7 )
7
1240
187
(e1)
6
1116
191
(81)
6
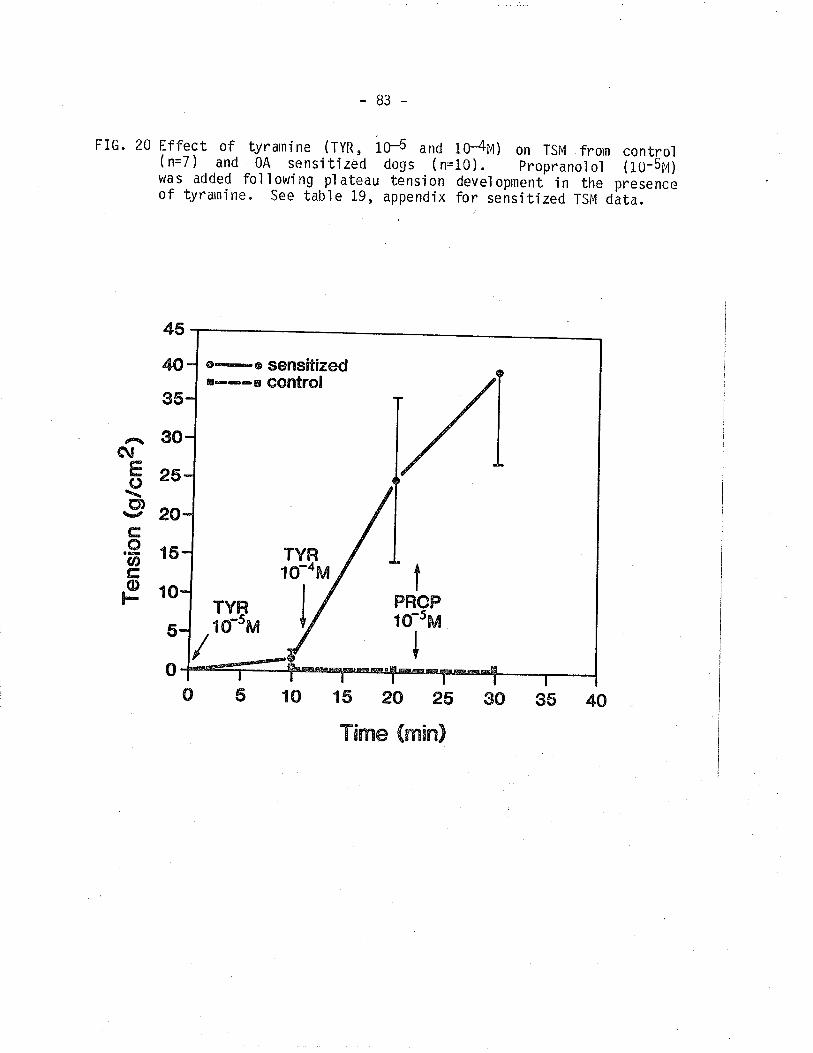
FIG. 20 Ef fect of tyratni ne (TYR, 1t-5 and 1ÉM) on TSIvt froin control( n=7 ) and 0A sensi ti zed dogs ( n=10 ) . propranol ot iiö--bf,j jwas added following qlateau tension development in the presenceof tyramÍne" see table L9, appendix for sensitized rsM däta.
B3
15 20 26 ß0 35 40
TEsT?e (mEn)
45
40
35
26
30
2A
15
10
5
o
n̂tËocÐ\/g.gCnËoþ
50 "t0
FROPtr0-sM
TYR10-1M
TYRt0-5M
/
@@@ SensitizedsoæEE eOntfOl

-84-In unstimul ated control TSM, tyratni ne (1û-5 and tf4N) produced
no response even with the further additÍon of propranolol (tO-5¡1, Fig.
20). 0f the 10 sensitized TSM tissues 5 produced a stnall contractile
response to tyrami ne tO-4N QS L LIg/ cn?) . Thi s wa s enhanced by the
additíon of propranolol (tA-6N, Fig. 20 and table 19 appendix).
Prevíous resul ts dernonstrated that control TSM responds to tyramine
(f f5 and 1ÉM) when active tone has been Ìncreased by 22.8 mM 6+
(fig.8). As noted above, unstimulated control TSM did not respond to
norepí nephri ne (1É to 1ÉM) even i n the presence of beta-adreno-
ceptor blockade rvith propranoloì (tO{N, Fig. 2L and table 20 appen-
dix). Addítion of norepinephrine (1É to tf4U in a cumulative
paradigrn) to 0A-sensítized TSM produced a contractile response with a
mean of 183 + 99 9/cnl (fig. 2L). Further addition of tyramine (1É
and lÉtvl) increased the tension produced to a mean value of 402 + L34
glcn?, which r¡ras abolished by phento'lamine (tt{}4 Fig. 2L and table
2L appendíx).
D) Effects of Atropine and PhentolamÍne on Contractile Responses Pro-duced by Histamine and Serotonin in Control and Ovalbumin-sensitizedCanine TSM
The possible interaction of the antagonists atropÍne and phentolamine
with histamine and serotonín (5-HT)-induced contractions of canine TSM
were studied. A histamine (1ÉM)-induced contraction (100%) was par-
tia11y antagonized by phentolamine (1rsM) to 73 + 6% of the inÍtial
contraction and plateaued 10 minutes fol'lowing phentolamine administra-
tíon at 78 + LI% of the histamine induced active tone (fig. 2?). Pre-
treatment wi th phentol ami ne (1ÉM) al so reduced the response to hi sta-
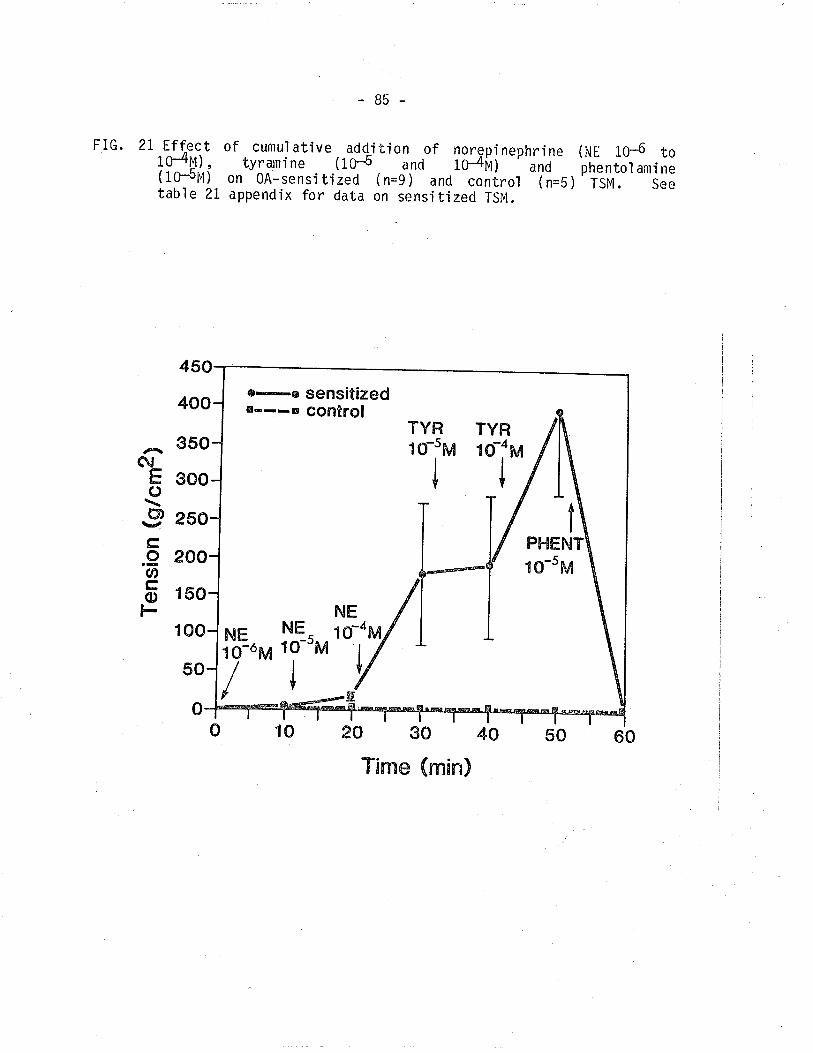
B5
F IG. 2L Effect10-4u),( 1û-5M)table 2
of cumul ative addition of norepi nephrinetYratnine (10-5 and lÉM) andon 0A-sensitized (n=9) and control (n=5)
1 appendix for data on sensi ti zed TSivl.
(Nt 1É tophentol ami neTSM. See
Ã&,
Ëoo\4c.oØcc)F-
300
250
450
400
350
200
150
100
50
0102030 40 50 60
Tlr¡"¡e (mln)
/l
cr@ SenSitized6--EE COntfOl
TYR1 o-sM
TYRt0-4m
I
NE1 o-4M
ffi-o* ffi'*
PHËN
1o-sM
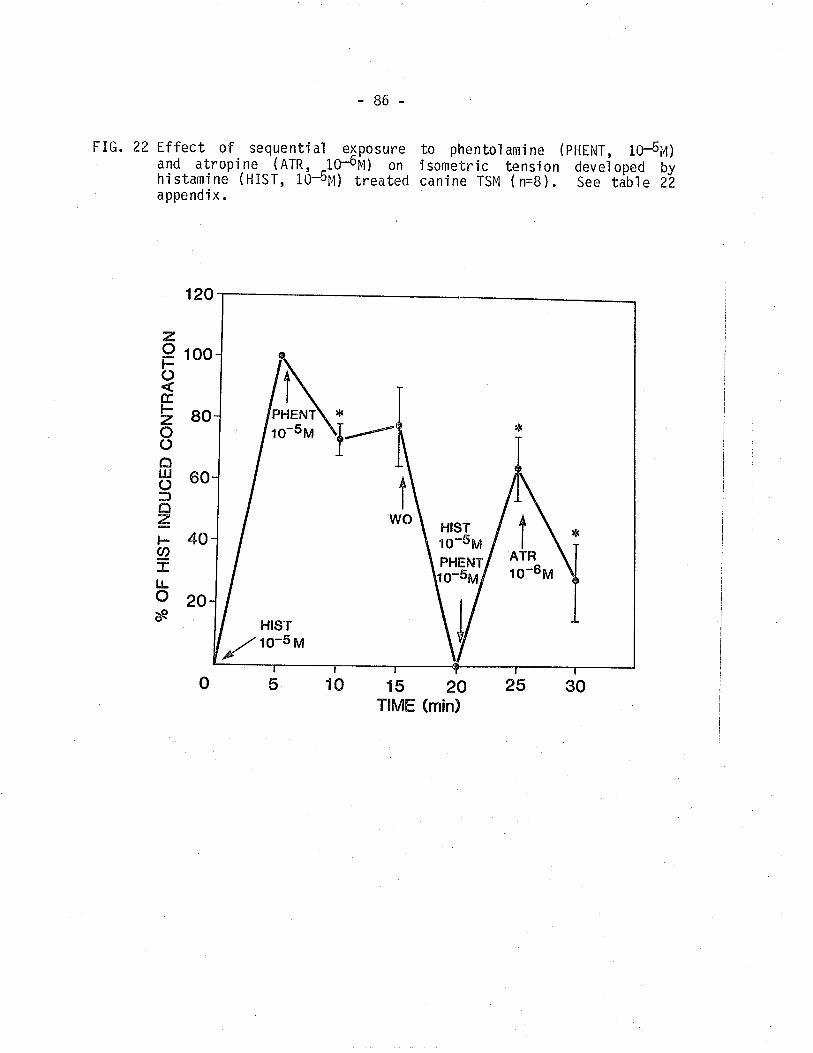
B6
FIG. 22 Effect of sequential e¡posureand atropi ne (ATR, _10-Ô M) onhistamíne (HIST, 10-bM) treatedappendix.
to phentol ami nei sometric tensioncanine TSM ( n=B).
(PHENT, 1r5M)deveì opedSee table
by22
100?otroE,
zooâlUC)Ðâa;U)rLLorOo\
120
BO
60
40
20
0 5 10 15 20TIME (min)
25 30
1o-5M*
*
wo*HIST
lo*5MATR10-6M
HIST10-5 M

-87-mÍ ne ( tO-sN) such that only 65 + LO"/" of the fi rst hÍ stami ne- i nduced
contraction (t00%) rernained. This latter effect was not significant'ly
di f ferent from that resul ti ng when phentoì ami ne (tnsy) uras added at
the plateau of the histamine (lÉM) induced contraction. Following
the pl ateau of tension responses to histamÍne (1fsM) in the presence
of phento'l ami ne (t nsN) , atropi ne (lÉM) further reduced the con-
tracture by 37% so that only 28 + L2% of the first histarnine contraction
(L00"/") remained (fig" 22 and table 22 appendix). l¡,lhen histamine
(tO-5u)-stimulated TSM !,rere treated with atropine (1ÉM) in the
absence of phentol ami ne a drarnati c 46/" decl Í ne i n active tone resul ted
(fig. 23). Pretreating with atropine (IÉM) resul ted in a simil ar
Ínhibition of histamÍne-induced tension development. In the previous
study (fig. 22) phento'lamÍne significantly ( p < 0.05) inhibited hista-
mine-induced tensÍon development, whereas in the present tissues which
had been atropÍnized prÍor to phentolamine treatment, this inhibition was
not observed (Fig. 23).
l'lith serotonin (5-HT)-stimulatetl TSM the actions of the antagonists
atropÍne and phento'lamine were reversed as compared to their effects in
hÍstamine-treated tÍssues. Atropine (1ÉM) produced a smal I but signi-
ficant (p < 0.05) inhibition of a 5-HT (lÉM) induced contraction
whether admínÍstered at the plateau of contractíon or prior to 5-HT
(to{N, Fi 9 . 24 and tabl e 24 appendix) . Thi s Ínhibi tion lvas not signÍ-
ficant when the muscles had been blocked wíth phentolamine (1ÉM)
prior to the addition of atropine (fig.25). These results indicate a
potent Ínhibi tory action of phentol ami ne (1ÉM) on contractions stimu-
I ated by 5-HT (IÑM) i n the presence of atropi ne (f i g. 241 or wi th
5-HT alone (Fig. 25, see also tables 24 and 25 appendix).
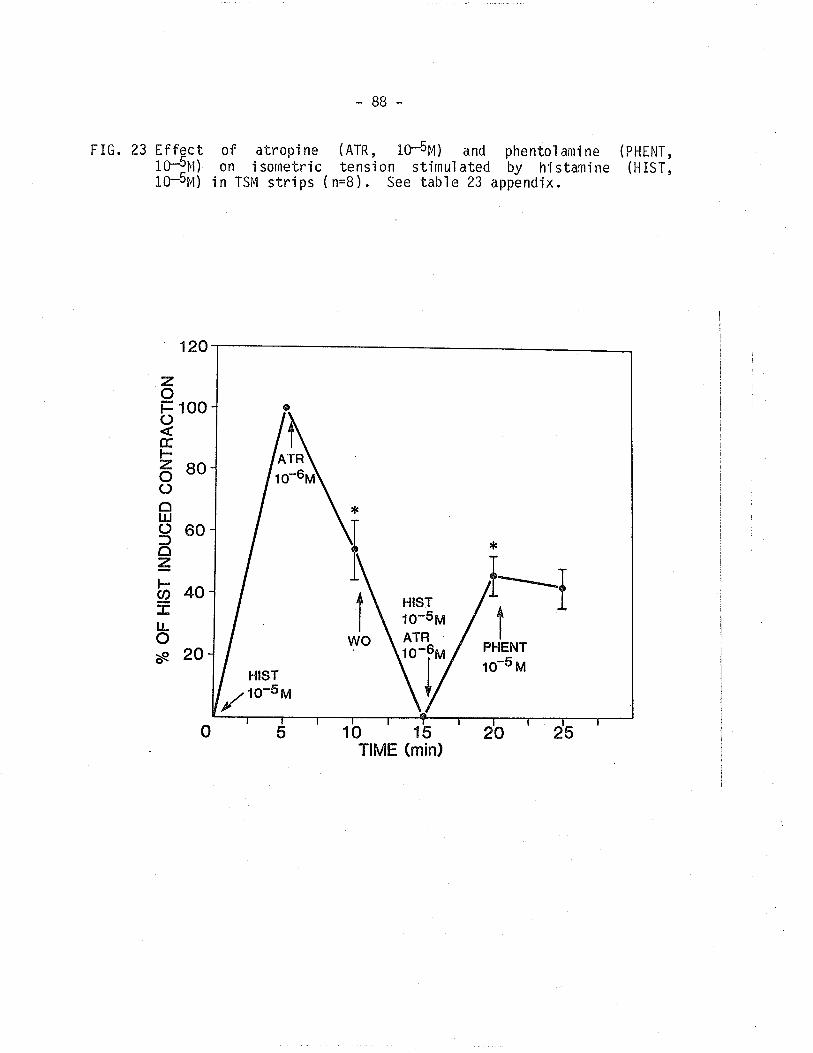
8B
FIG.23 Effectto{r'rlthsM)
of atropi ne (ATR, 10-5M) and phentol ami neon i sometric tensi on stimul ated by hi stami ne
i n TSM stri ps ( n=B ) . See tabl e 23 appendÍx.
(PHENT,(HIST,
zoofEt-zoooTUC):)ÕzF-U)
LLorC'c,\
1 oo
120
BO
60
40
20
0 1 1 52TIME (min)
wo
/
10-6ATR
HIST10-5M
*
t
10 -6M
HIST1o-5MATR
PHENT
1o-5 M
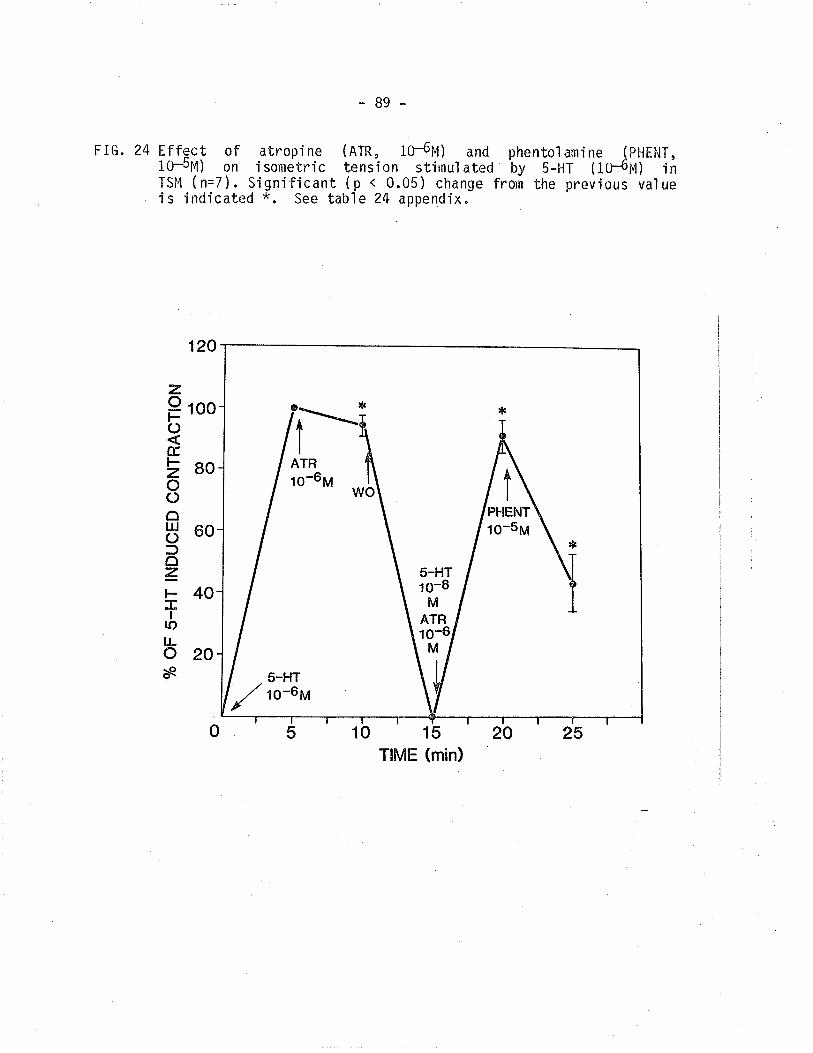
FIG.24 tffqct of atropine (ATR, lÑM) and phentolamine (thSM) on i sometric tensi on stirnul ated 'by 5-HT (1ÉTSl4 (n=7). SignÍticant (p < 0.05) change frorn the previousis indÍcated *. See table 24 appendix.
B9
TIME min)
PHTNT,M) in
val ue
001
zIf--Ofrt--zo()âLU()ozFt
Iro]Loèq
120
80
60
40
20
o 5 10 520251
(
wo
ATR10-6M
5-HT1o-6M
* *
'fi
M
PHENT
10-5M
5-HT10-6
M
ATR10-6

FIG. 25 Effect of phentol amÍ ne (PHENT, 10-5M) and atropi ne (nrn1ñM) on i sbrnetríc tensi on oevét opeo úi s:Hì riää'ml- rreare<jTSM (n=81., Significant _(p < 0.0s) ihange-froin previous value isindicated *. See table 25 appendÍx.
5 10
90
15 20T¡ME (min)
2s 30
00
BO
60
40
2
zo-FroÉ,
zooâllJC)3o4r
IK)IJ-oa
120
o
M
*
JPHENTto-5n¡
wo
+
ATR1o-6M
5-HT10-6 M
1
5-HTto-6n
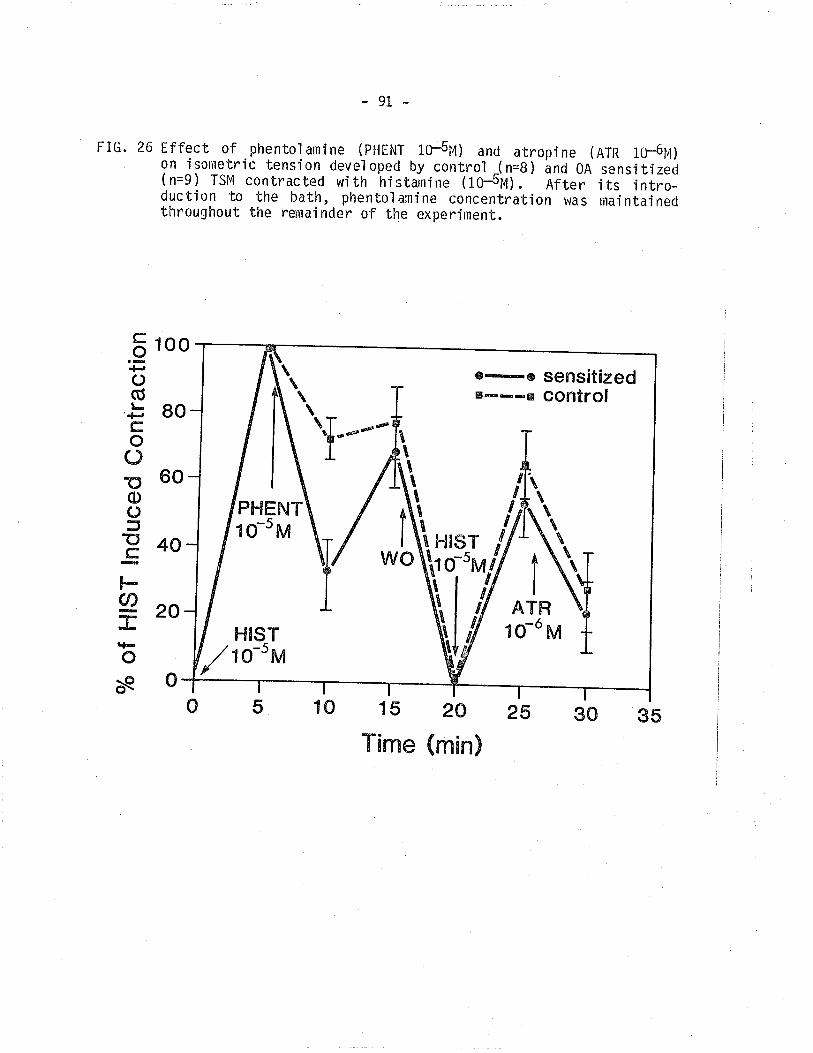
FIG.26 Effect of phentolamÍne (P!{ENT lrsM) and atropine (nrn to-6pl)on isontetric tension developed by control fn=B) änd 0A sensitized( n=9 ) TSM contrac ted wi th 'hi
staini rre ( 1ÉM) . ' Ãiiu. i:ut i ntro-ductíon to the bath, phentol amine concentration was riraíntainedthroughout the remainder of Lhe experÍ¡nent.
91
o 5 10 15 20 25 30 35Time (rnin)
100.oo(úLËoÇëc)O5õsf-.-Ø-Lrþo
qôo\
80
60
40
2A
o
PHENTto-sn
\\\
r \IT
t \\r I \q tðH T
WO 1 0-q
ATR60-1 M
rIt
@k@ sensitizedsaee@ COntfOl
H!ST10-sM
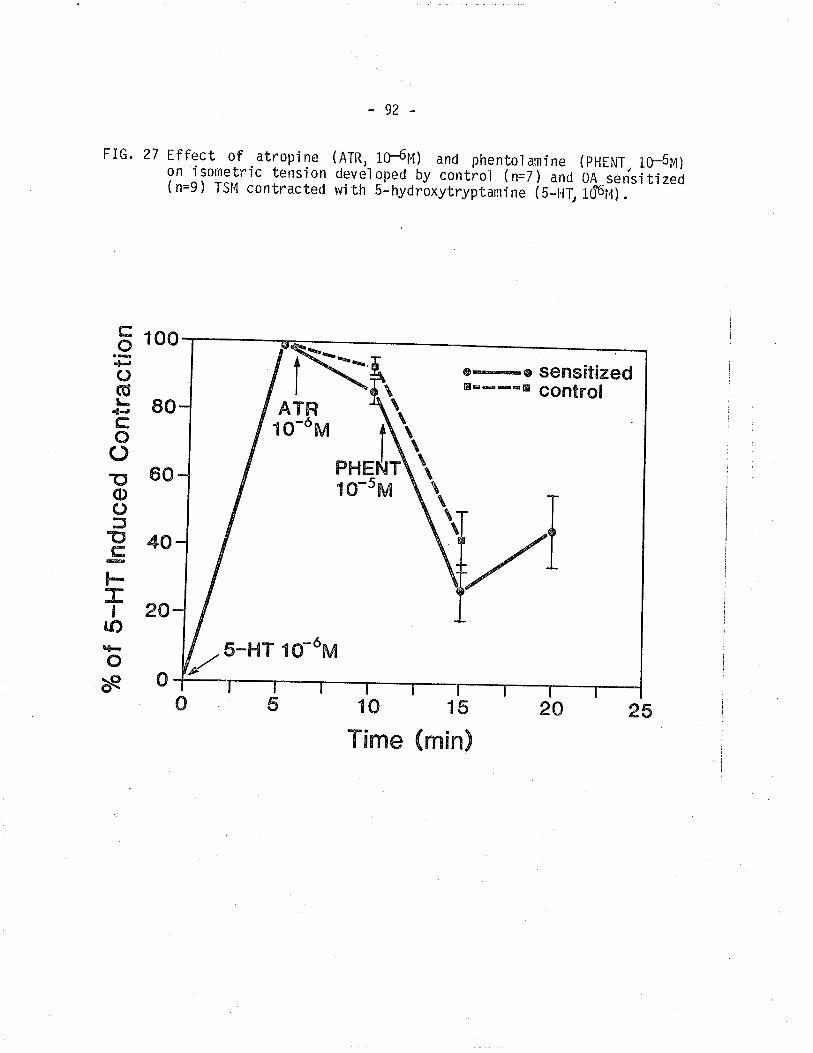
92
FIG. 27 tffect of atropine (ATR,1ryMJ and . phentoramine (PHENT, 10-5M)on isoriretric tension developed by control (n=7) àñ,1 ön lunsitized( n=e) TSM conrracred wirh 5-hy¿röxvrivpiurin.' ís:rii iäor,ï .
c..o+-,C)6tL+tËo
C)EOC)JEL-
ËEF
-LI
roeþo
róo\
100
EO
60
40
20
o0 ËiJ 10 15 2A 25
Time (min)
5-HT î O'
\
6M
TM
6e€Ð sensitized$eeeeg! control
ATR10-óM
PFIFI 0-s
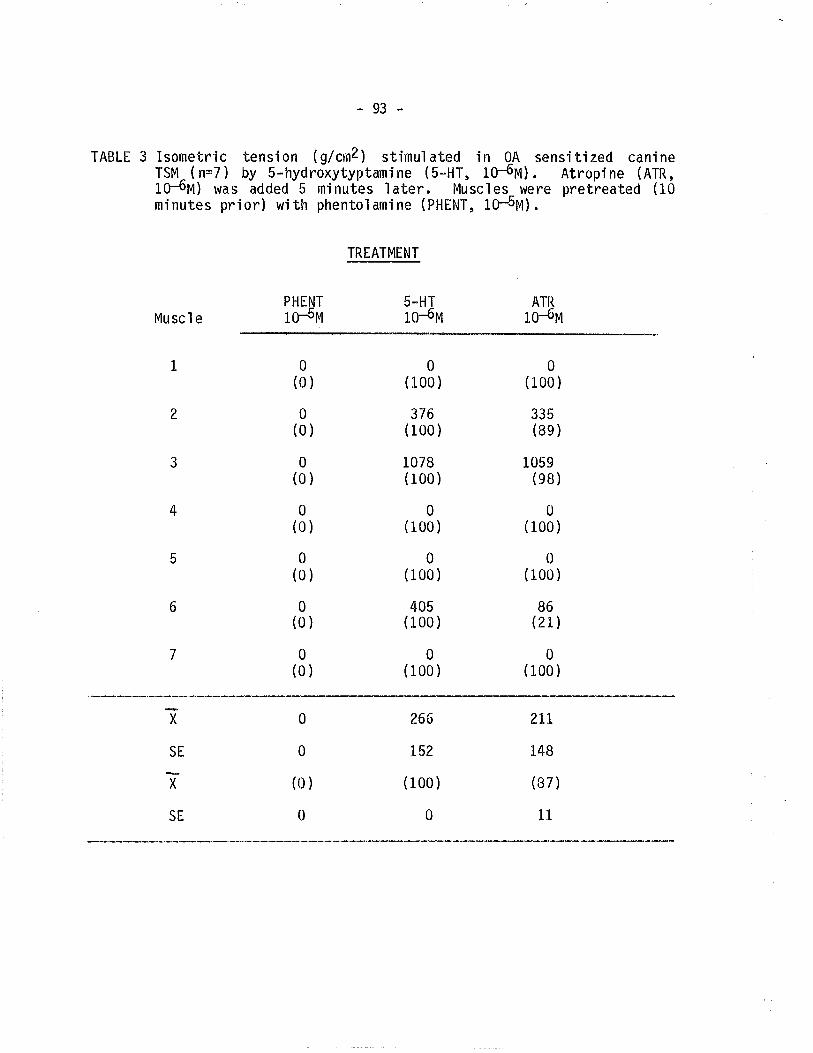
TABLE 3 Isometric tensi on (g/ ctn? ) stirnul ated Í n 0-A sensi tized cani neTSM- ( n=7 ) by 5-hydroxytyptamÍ ne (5-HT, 1ÉM) . AtropÍ ne (ATR,to-6N) was ãdde¿-5 mÍnutes later. Múscles-were preti^eated (1Ó
minutes prÍor) with phentolamÍne (PHENT, 10--5M).
93
TREATMENT
MusclePHENT1rrsM
5.HTlo-6N
ATRtoj¡'l
1 0(0)
0(0)
0(0)
0(0)
0(0)
0(0)
0(0)
0(100 )
376(100)
1078(100)
0(100)
0(100)
405(100)
0(100)
0(100)
335(8e )
1059(eB )
2
3
4
5
0(100)
6
7
0(100)
86(2rl
0(100)
ï
TST
SE
0
0
(0)
0
266
L52
(100)
0
2LL
148
(87 )
11
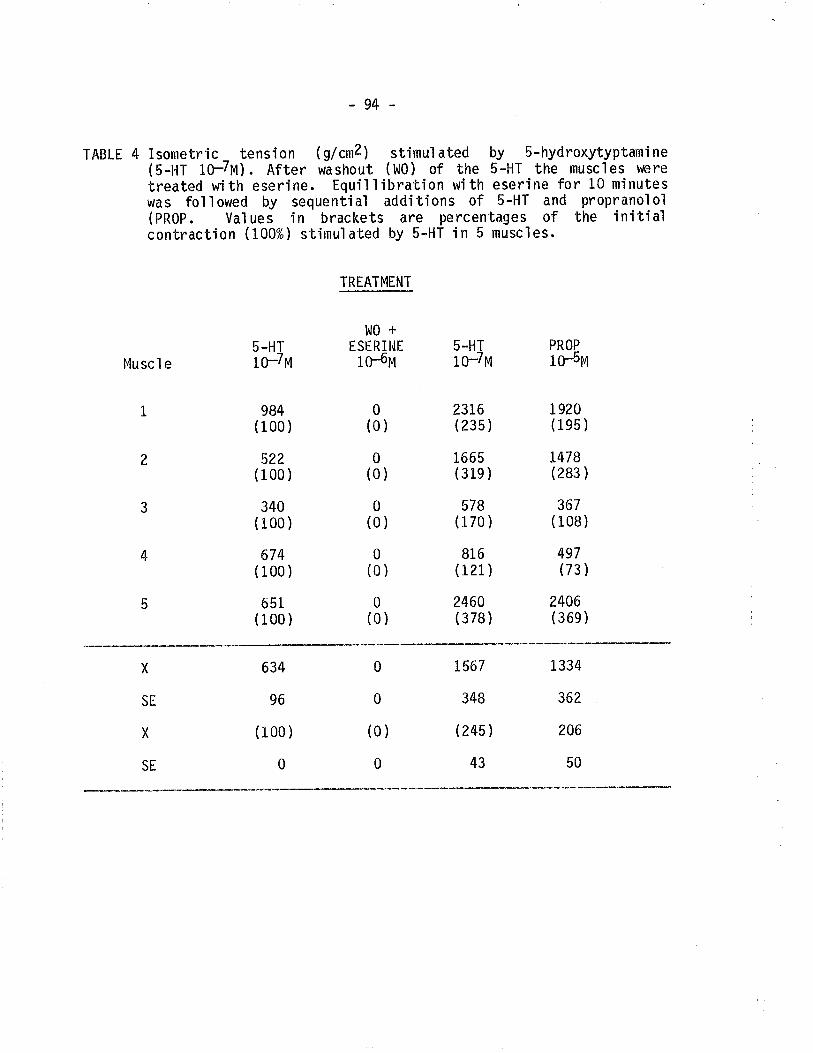
94
TABI-E 4 Isometric - tension (g/ cn?) stimul ated by 5-hydroxytyptami ne(5-HT th/M). After washout (hJO) of the 5-HT the muscles !',eretreated with eserine. Equillibration with eserine for 10 minutes¡¡as fol I owed by sequenti a'l addi tions of 5-HT and proprano'l o1
(pnOp. Val ues in brackets are percentages of the initialcontraction (100%) stirnul ated by s-HT i n 5 muscl es.
TREATMENT
l4uscl e5.HTrn7M
l.J0+TSERINTto-6N
0(0)
0(0)
0(0)
0(0)
0(0)
5.HT1o-7M
PROP
to-SN
1
2
3
4
5
984(100)
522(100)
340(100)
674(100)
1665(31e )
2316(235)
I 920(1e5)
L47B(283 )
367( 108)
651(100)
578(170)
816(121)
2460(378)
+97(73 )
?406(36e)
X
SE
X
SE
634
96
(100)
0
0
0
(0)
0
L567
348
(245)
43
1334
362
206
50

-95-Data obtained in tissues froln 0A-sensitized dogs showed both quali-
tative and quantitative similarity to controls. Figures 26 and 27 denton-
strate that, in histamine (ff-sM)- or 5-HT (lÉM)-stimulated muscles
the actions of phentol ami ne (10-5M) and atropí ne (1ÉM) were simil ar
for sensitized and control groups with the exception that a greater
transi ent í nhibi tory action of phentol ami ne (1f5M) on a hÍ stami ne
(1Û-5M)-induced contractÍon Ín sensitized TSM was observed. Ten
minutes fo'llowíng phentolamÍne exposure the stable plateau values for
sensitized and control tissues were significantly dífferent (Fig. 26).
In sensitized TSM pretreated with phentolamine (lÉM) only 3 out
of 7 rnuscles responded to the subsequent addition of 5-HT (tOifq, Table
3 ) . A1 though the mean contractil e response Q66 + L52 gl cn?-) was not
significantly different from the control response of 245 1 66 9/cn?
(Tatlle 25 appendix), all the control muscles (n=8) contracted in response
to 5-HT (1fsM) exposure after phentolamine pretreatment.
t) Effects of Eserine, Propranolol and Phentolamine on the ContractÍleResponses to Hi stami ne, S-hydroxytryptami ne, Acetyl chol i ne andPotassium Ín Canine TSM itLti!to_
Further studÍes focussed on the possibí1ity that 5-HT and hÍstamine
interact wÍth cholinergÍc or adrenergíca11y mediated responses. Eseríne
(physostigmine, lÉM) signíficant'ly potentiated contractÍons produced
by 5-HT (lor7M) to a mean of 245 + 34% of the initial 5-HT (lr7M)-
induced contraction. Propranolol (1ÉM) reduced the tension to 206 +
50% (Table 4). In eserÍne-pretreated muscles the response to hístamine
(lÉM) rose to 703 + L74% of the val ue produced by hi stami ne al one
(100%). Additíon of hyoscyamÍne (lCÉM) reduced the response to hista-
mine in the presence of eserine to 4L + 32% of the initial histamine-in-
duced contraction (Table 5).

-96-A1 though atropi ne and hyoscyami ne parti al 'ly i nhibi t hi stami ne-
i nduced (1ÉM) contractions, hi stami ne at higher concentration
(1ÉM) i s a potent stimul ator of depol arízed TSM, which has been
bl ocked vli th atropi ne, propranol o1 and phentol ami ne. Hi stamÍ ne ( lÉM)
under these pretreatment conditions produced a contractÍon of 280 + t6%
of that in the presence of K+, L27 mM (Tab'le 6).
In order to test for the specificÍty of eserine action on choliner-
gÍca1 1y medi ated responses, muscari nic bl ockade wi th hysocyami ne was
used. The addition of eserine at the plateau of an acetylcholine-Índuced
contraction produced an increase of 213 + t8%, over the initial response
( 100%); the subsequent addÍtion of hyoscyamine (tO--5N) abol i shed the
response (Tabl e 7 ). Phentol ami ne ( 10-5M) reduced acety'l chol i ne
(lÉM)-induced contraction ÃOO%) to SZ + 3% and the further addition
of atropine (10-7M) abolished the contraction completely (Table B).
Potassium (K+, 23 mM) induced contractions were al so increased
dramatical 1y by eseri ne (lÉM) and occurred to 'ühe same degree lvhether
eserine was app'lied at the plateau of the K+ induced contraction (Table
9) or prior to ç+ addition (Table 10). Although phentolamine (1ù-5M)
slightly reduced (Table 9) and propranolo1 (1f5M) slightly augmented
(Table 10) tne tension stirnul ated by 23 mM ç+ i n the presence of
eserine, neither effect was significant (p < 0.05).
F) Factors tlhich May Modulate t3Hl NorepinephrÍne Overflow in CanineTSM in vitro
The previous studies dernonstrate that phentol amine significantly
reduced isometrÍc tension developed by TSM ín the presence of electrÍcal
field stimulation, (rig. 2\, tyrarnÍne (rlg. I and 21), histamine (rig.
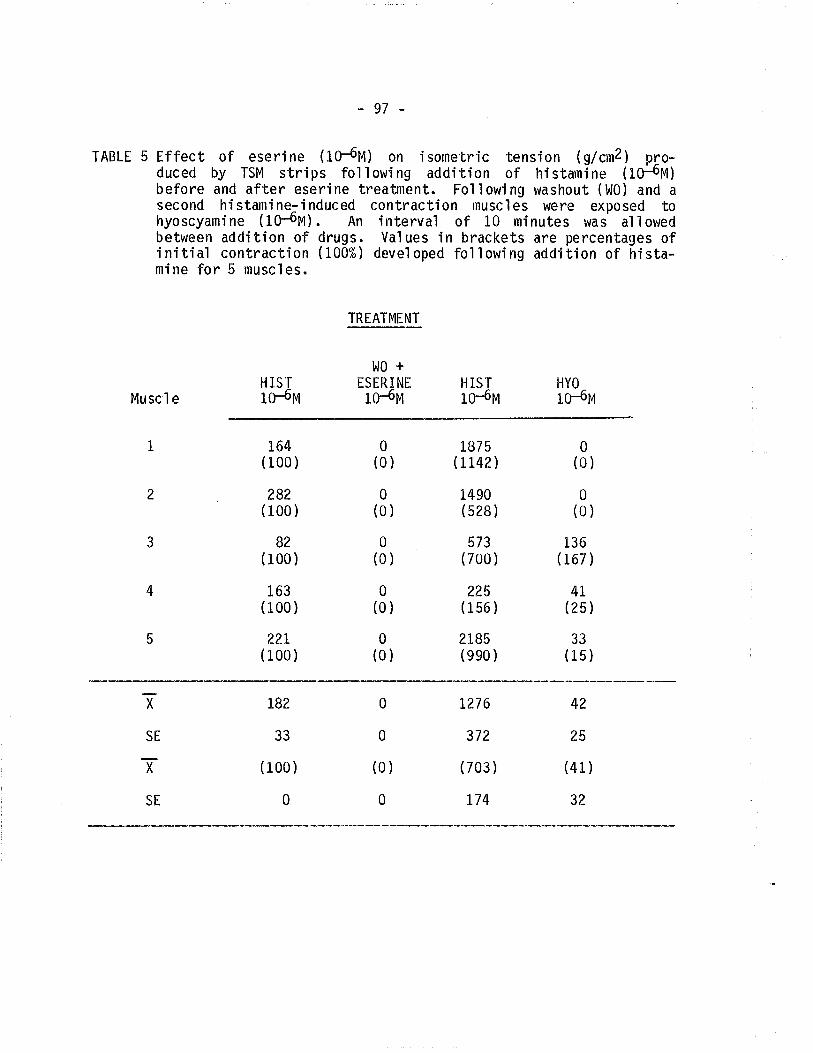
-97-
TABLE 5 Effect of eserine (lÉM) on isometric tensíon (g/cn?) pfo-duced by TSM stri ps fol 1 owi ng addi tion of hi stami ne (1fM)before and after eserÍne treatment. Following washout (l^l0) and asecond histamine;induced contractÍon rnuscles vlere exposed tohyoscyami ne (1ÉM) . An i nterval of 10 mi nutes was al I owedbetween addition of drugs. Values in brackets are percentages ofinitial contractÍon (100%) developed fo1lowíng addition of hista-mine for 5 muscles"
TREATMENT
Muscl eH IS'TtrsN
l¡10 +
ESERI NT
to{NH ISTto-6N
HYO
to{N
1 164(100)
282(100)
82(100)
1875(1142)
1490(528)
573(700)
225(156)
2L85(ee0)
136,167
\
00
00
00
00
0
(
00
00
)
2
3
4
5
163(100) 25\
33(15 )
4L
?-2L(100) (0)
TSE
TSE
I82
33
(100)
0
0
0
(0)
0
t276
372
(703)
L74
42
25
(41)
32
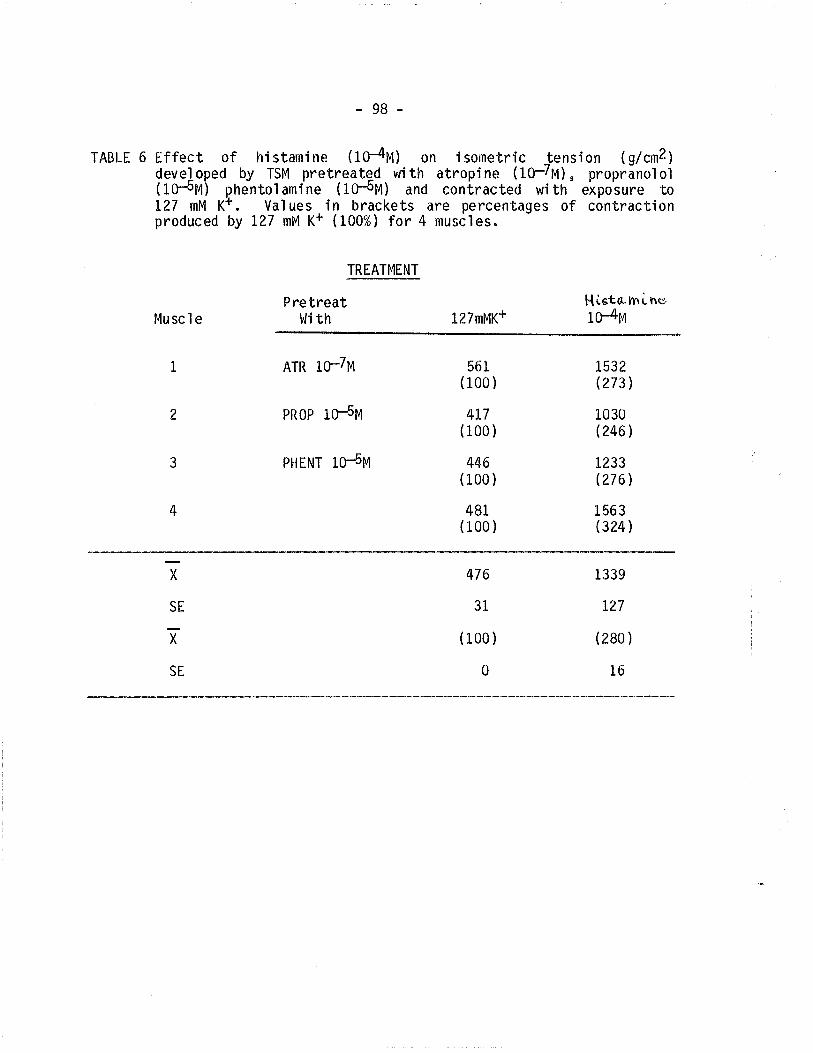
TABLE 6 Ëf fect of hi stamÍ ne (1ÉM) on i sometríc tensi on (g/cn?)developed by TSM pretreate-d with atropine (tO-7m), proprãno1o1(lFbM) phentol amÍ ne (lfbM) and contracted wi th exposure tol?7 mM K+. Values in brackets are percentages of contractionproduced by L27 mM K+ (1007,) for 4 muscles.
-98-
TREATMENT
Muscl ePretreat
hti rh 127mMK+
Hùs'ta.rnr neto-4N
1
2
3
4
ATR 1O-7M
PROP lÉM
PHENT TO{N
561(100)
1532(273)
4r7(100)
1030(2461
t233(276)
1563(324)
446(100)
481(100)
X
X
SE
SE
476
31
(100)
0
1339
127
(280)
16
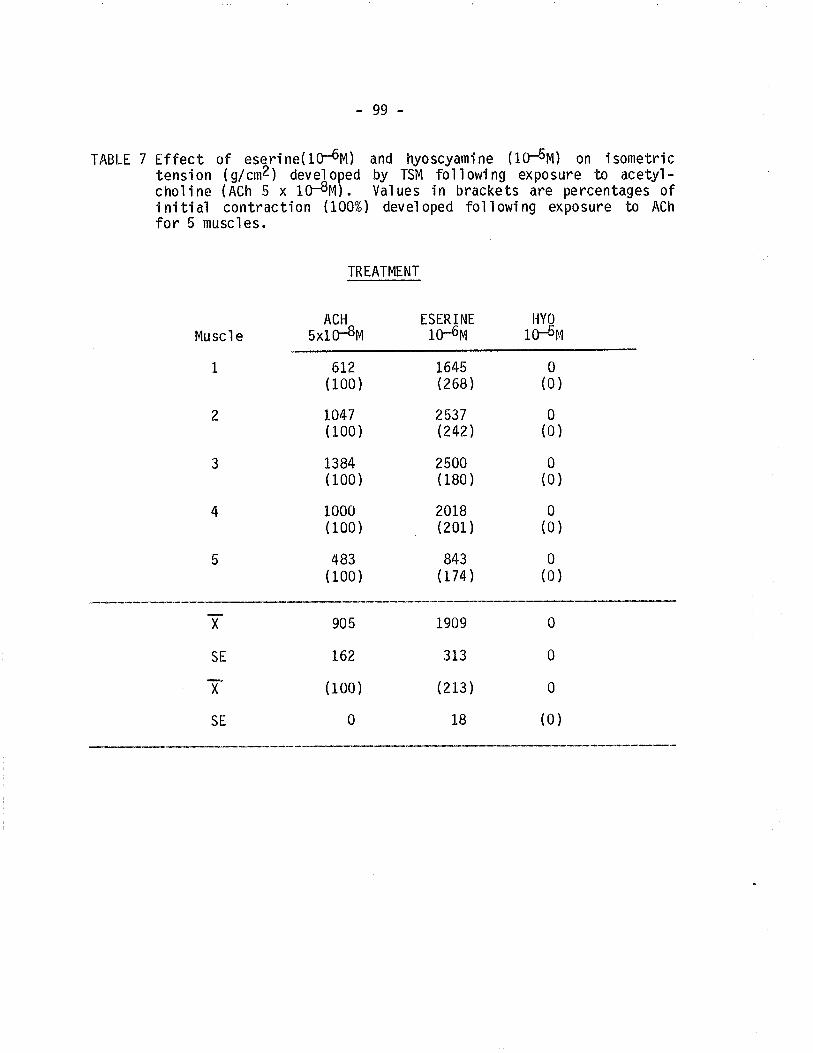
TABLE 7 Ef fect of esqri ne(tO{tut) and hyoscyami ne (1fsM) on i sometrictension (g/cnz) developed by TSM folìowing exposure to acetyl-chol i ne (ACh 5 x 10+M) . Val ues in brackets are percentages ofinitial contractÍon (1007,) developed following exposure to ACh
for 5 muscles.
99
TREATMENT
Muscle
I
ACH
sxtÉNESTRTNE HYOto-srq thsu
6L2(100)
1645(268)
2537(242)
2500(180)
2018(201 )
843(1741
00
00
00
00
00
2
3
4
5
L047(100)
1384(100)
1000(100)
483(100)
0
T
TSE
SE
905
L62
(100)
0
1909
313
(213]'
18
0
0
(0)
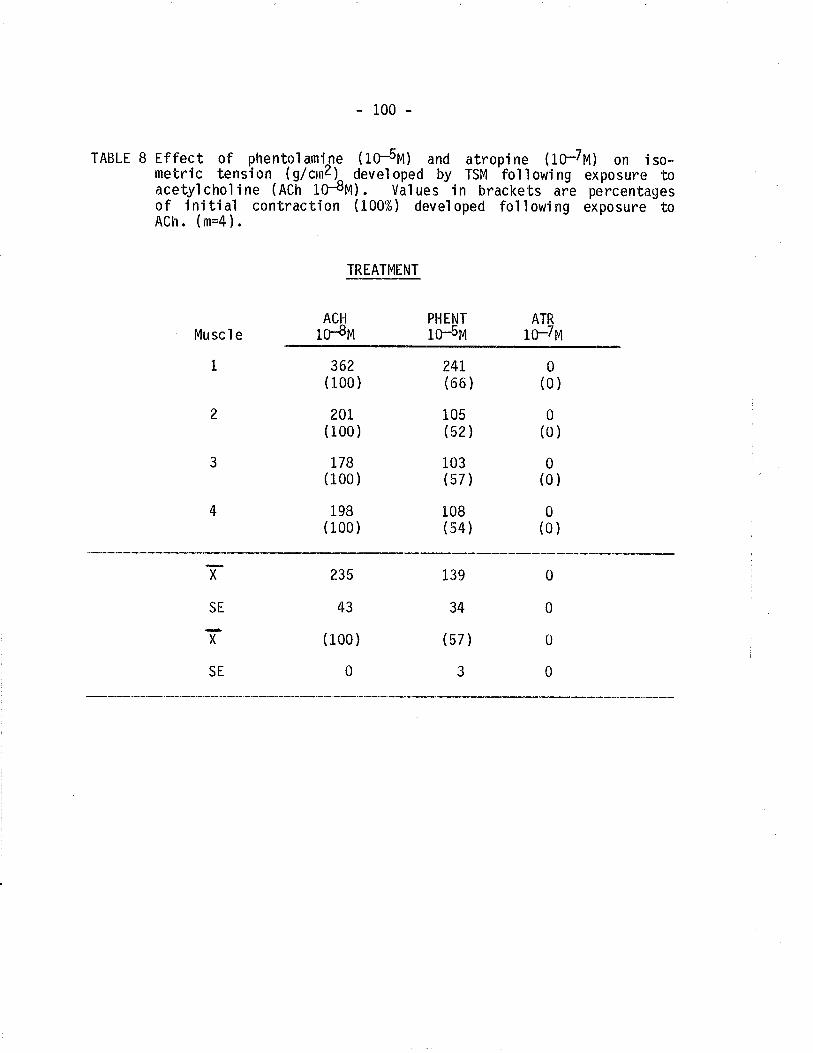
TABLE 8 Effect of nhentolaminemetric tension (glqn?)^acetyl chol i ne (nCfr lNMof initial contractÍonACh. (m=4).
ACH
ro-8N
( lÑM) and atropi ne (10-7M) on i so-developed by TSM following exp0sure to). Values in brackets are percentages(L00%) devel oped fo1 1 owi ng exposure to
-100-
TREATMENT
PHENT ATR1o-5M thTMMuscle
1 0
2
3
4
362(100)
201(100)
L7B(100)
198(100 )
i03(57)
24r(66 )
105(52)
(0)
0(0)
00
00
108(54 ) )
X
X
SE
SE
235
43
(100)
0
139
34
(57 \
3
0
0
0
0
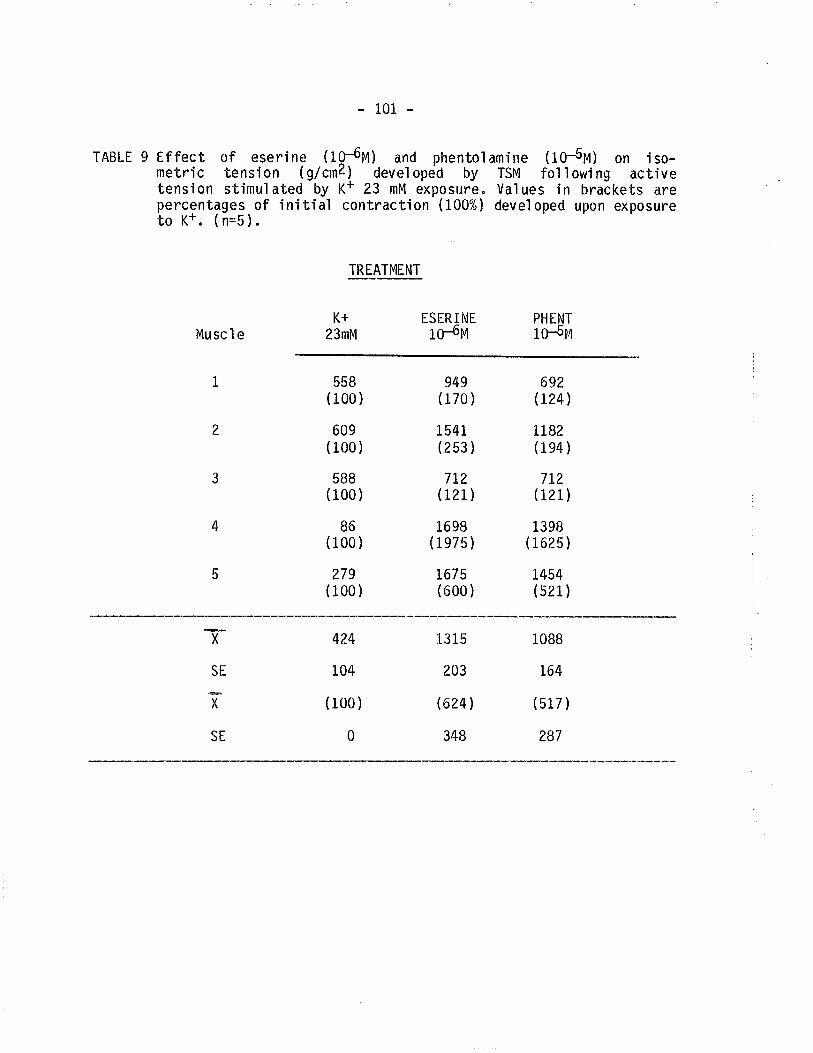
TABLT 9 tffect of eserine (1FM) and phentolamine (1fsM) on iso-metrÍc tension (glcnz) devel oped by TSM fol I owi ng activetension stimulated by K+ 23 mM exposure. Values in braðkets arepercentages of initial contraction (100%) developed upon exposureto K+. (n=5).
-10i-
TREATMENT
MuscleK+
23mM
ESTR I NE
to{NlPHËNT
to-5M
1
2
3
4
5
558(100)
609(100)
5BB(100)
B6(100)
279(100)
1541(253 )
1698( 1e75 )
167 5(600)
TLBz(1e4)
L454(521 )
1398(r625],
949(170)
692(r24)
7L2(L?L)
712Í2Ll
TSE
X
SE
4?_4
104
(100)
0
1315
203
(6241
348
1088
164
(517)
287
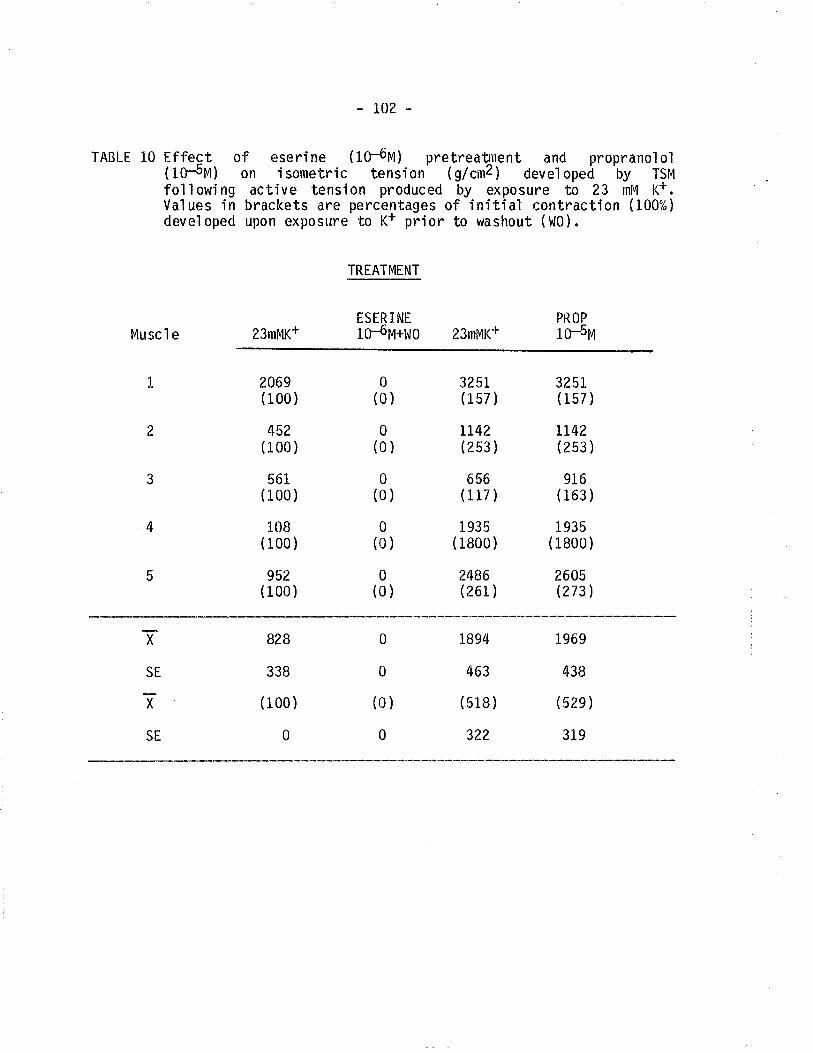
TABLE 10 Ef feqt of eserÍ ne (lÉM) pretreatment and propranol o1( 10-5M) on i sometric tensi on' (g/ cn?) devel opeä 'by
TSM
following active tension produced by exposure to 23 mM K+.Values in braclcets are percentages of initial contraction (100%)developed upon exposure to K+ prior to washout (l^l0).
Muscl e 23mtvlK+
-t02-
TREATMENT
TSER I NE
tr-6N+wo 23mMK+PROP
trsM
)
)
1
2
3
4
5
2069(100)
00
00
00
00
00
(
325L(157)
325L(157)
tL42(253 )
656(117 )
916(163)
452(100)
561(100 )
It42(253)
1 935( 1800 )
?486Q6LI
1 935( 1800 )
2605Q73)
108(100)
952(100)
X
SE
SE
828
338
(100)
0
0
0
(0)
0
1894
463
(518)
322
1969
438
(52e)
319
X

-103-221, serotonin (Fig. 25\ acety'lcholine (Table 8) and potassium in the
presence of eserÍne, (although not significant, 4 of the 5 muscles
responded with a decrease in tension, Table 9). The possibi'lity that the
above agonÍsts àct on adrenergic neural elements to increase norepine-
phrÍne release was studied by incubating muscles wÍth [3H]-norepine-
phrine and stimulating the preparation with electrÍcal field stirnulation
(EFS) to promote NE turnover in nerves (see methods). Follovring the
perÍod of t3Hl- norepÍ nephri ne i ncubation, cocaí ne ( 10-7M) was
added to the bath to prevent NE neural uptake (Uptake 1) and was sub-
sequent'ly added to the bath fol l owi ng each samp'le col I ection throughout
the experiment. Following a períod of washout (24 ninutes) an agonist was
Íntroduced for a 3-minute collection Ínterval after whÍch the muscle was
exposed to Krebs-Henseleit solution alone for the duration of the experi-
ment. Isometric tension was recorded sirnul taneously throughout the
experÍment. All of the studies involving labelled transnitter release
were similar in their sampling of radioactivi'üy overflow and all comparÍ-
sons were ¡nade for the samp'l e duri ng which treatment u,as given ( i .e.
interval between 24 and 27 min,). Significant changes from the expected
control value were determíned for the values at time 27 ninutes. Efflux
curves (of rate coefficient) for unst,imulated muscles demonstrate that,
the calculated expected values are an accurate representation of what
actual ly occurs for control rnusc'les (Fig" 28). The upper panel (Fig" 28)
also indicates a stable resting tone for the entire period.
El ectrical fi el d stÍmul ation (EFS ) whích i s known to stimul ate
nerves, as seen by the sensitivÌty of EFS-induced tension responses to

-104-tetrodotoxin (fig. 2), was observed to increase active tensÍon and
tritium overflow sÌgnificantly (p < 0.05). Histamine (1O_sM) which
Íncreased active tension did not change tritium overflow compared wÍth
the expected control values (flg. 29). The change in rate coefficient
was therefore not an artefact of tension-dependent al teratÍons of the
exchange kinetics in the extracellular space.
A high-potassium (23 mM) solution increased tension development to
669 + 85 g/ç¡12, whích was accompanied by a significant increase in rate
coefficient to 2.34 + 0.45 (Fig. 30). Both ç+ and EFS depolarize
nerves, although by dÍfferent methods, and were expected to increase
tritium overflow. Phentolamine (1fsM) however produced a signÍficant(p < 0.05) increase in rate coeffÍcient ín the absence of any mechanÍcal
effects (ftg. 31). Norepinephrine (1ÑM) al so produced a significant
increase ín tritium overflow without active tension development (fig.
321 . Ace'by1 chol i ne (1ÉM) produced a contractÍon which was parti al lyphentol amÍ ne- sensi Eive (Tabl e B ) , and Ach to-6N was observed to
increase t3Hl norepinephrine overflow signíficantly (flg. 33). previous
experirnents demonstrated a rnarked sensitivi ty of 5-HT (1ÉM)-incluced
contractíons (flg. 25\ to phentolamine, whí1e the present results (Fig.
34) do not demonstrate a significant increase in tritium overfl ovÀ, accorn-
panying the development of isometrÍc tension.
Figure 35 summarizes the results for overflow of tritium and com-
pares each with the control value expected. A signiFicant (p < 0.05)
overflow of trÍtíum was protluced in TSM following exposure to EFS,
phentolamine, K+ 23 ffiM, acetylcholine an<t norepinephrine" Both hista-
mine and 5-HT were without sígnificant effect. A'lthough significant
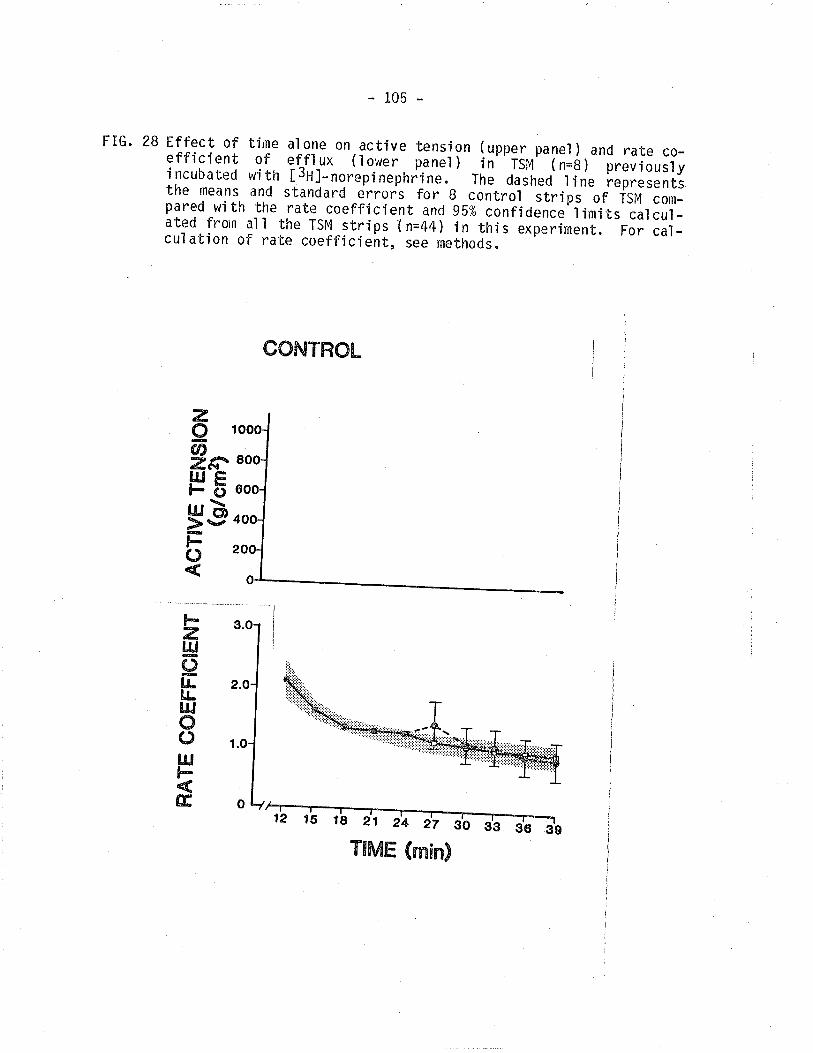
-105-
FIG' 28 Effect of time alone on active tension (upper panel) and rate co-efficient of effl ux (l ower pane] ) ìn' ' TSM--i ;=é ) ireviouslyincubated rvÍth [3H]-norepinephrine. '
ftre dashed lÍne iepresentsthe means and standard errors for B control itripr lr rsiv corn-pared with the rate coefficient and g5% confidence limits calcul-ated frorn all the TSM strips (n=44) in thir.*pà.i**;[:- For cal_culation of ra'Le coeffjcÍent, see methods.
cth¡rROL
æ.ÕØry^&(l{UJtrFe)!ub>\,F--O
I
20
2
þI&UJ
õtJ-TLutÕe)e¿Jþqtr
I
I
I
i
1
o12151 1 33 36
T8turË (min)
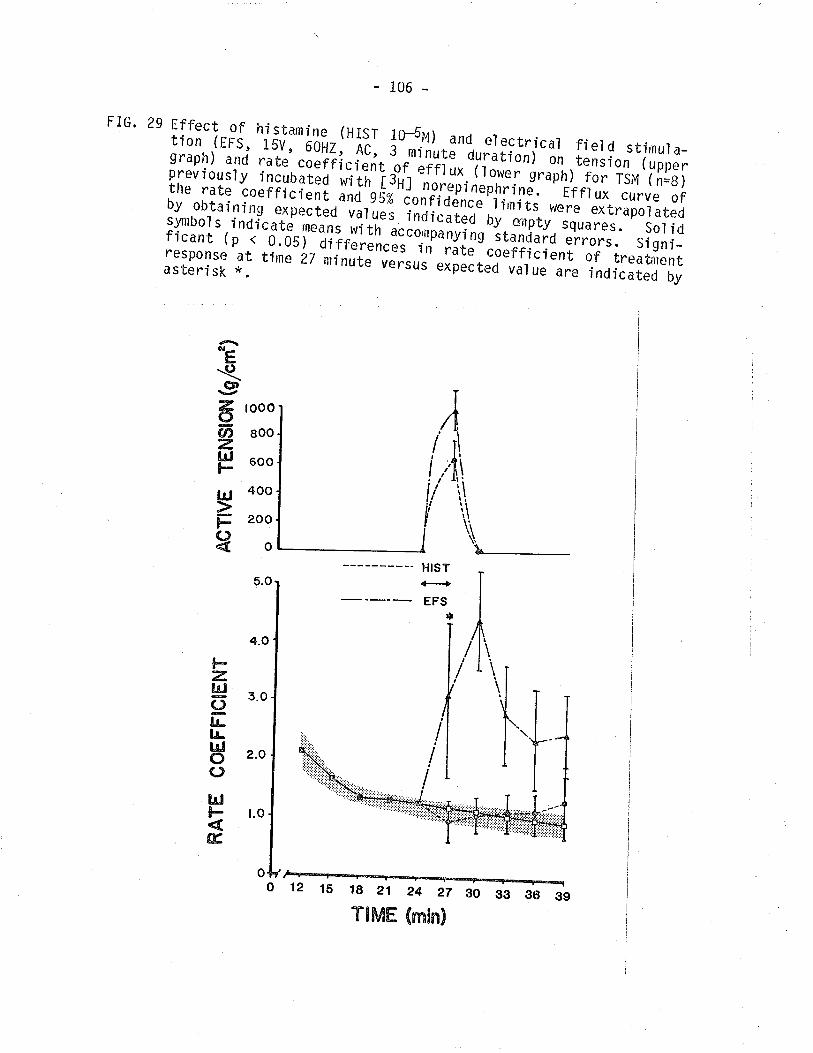
-106-
FIG' 29 Effect oj his.tarnÍne (HIST r0-5Nl and eJectrjcar fierd stirnuJa-tion (EF-si 1qv' aouz)"Åc,, ã--*inrtå"îuxarion) ;n ü;;ion (upper;låiil, lii" iliîr!!:iik li: t,l,l füg, _.*r vrap1¡
-,ä, is v r i,,s rthe rate coefritiuni änd ot* confÍdence ''ltl]is.. Effl ux ct¡rve ofby obtainjng o^puãiä,r-iärru, indicated ,.lirnits
were eitrapotateåsvmbol s indicaru
'n.unì ,uì;ï"^.:'ii::::::^ov, ernpty squares. sol id
I j *l;j o . o . o s i
- äï :r r; ::i.,!!:1i'l#.t
n,¡ :î?l U:,il'¡: ;,. : ä: j ;asterir* fl tine 27 rninute versus u^ôããtuu vatueïr. -in¿i.ared
by
"E-q-
Glt
æ,ö6?&e,F.
[tt
tr(J
Þ7lr"!
{ll
C)ËTL
8 z.o(J
tdF r.o
ffi
looo
800
600
400
200
o
5.O
4.O
I
---------- Htsr<a_+
EFS*
O12iS1g21242To
TIMF (n¡ån)
30 33 38 39
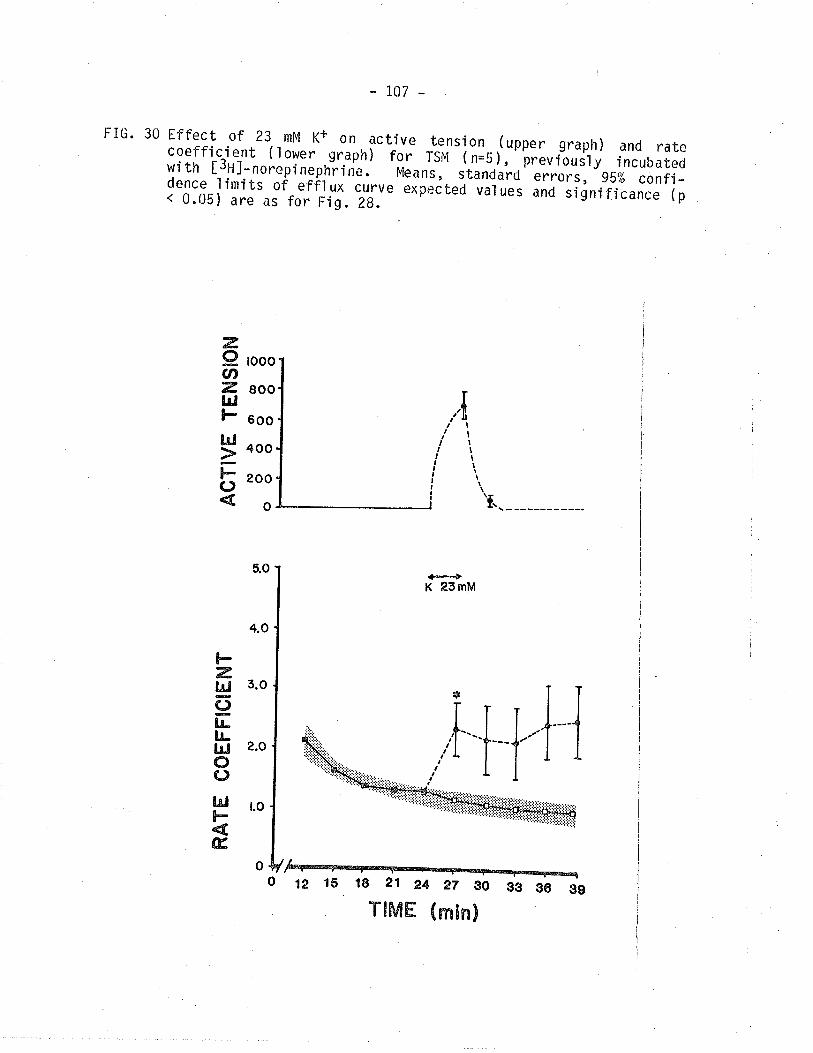
107 -
FIG.30 Effect of 23 mM ç+ on active tension_.(upper graph) and ratecoeffícig.'nt (lower.graph) for TSM (n=¡1, previously incubatedwith [3H]-.norepineprriiìie. Means, sïandard errors, gs% confi-dence rimÍts of effrux curve expected varues and significance (p< 0.05) are as for FÍg. ZB.
?Oqf,&[¿Jtsu,J
F(J
800
r000
,{,,Ltl600
400
200
o
IIItt
,I,,
tËr
5.0
4.O
F-3uJ 3.o
õüt¡-u 2.oÕL}
$¿J r.oÞ-
lr
K 23mM
,,{- I
12 15 18 2't 24 AT gO gg
g
oo
ïlMË (mln)38 39

i08 -
FIG. 31 Effect of phentoìamine (PHENT iù-sM) on active tension (uppergraph) and rate coeffícjçn! (lower pane]) for TSM in=Sl,' þ;;_yi9!:lv incubared wirh [3H]-norepineþhrjne. significance ti -.0.05) and other details are as foi rié " ZB.
A"F
åIgtñqÉ[&¡F-þJÞþc^t4
4.O
Þ-æ,KJ
õ 3'o
Lu-Lrlõ 2.o
C)
rooo
200
o
800
600
400
5.O
<F-+PHENT
¡ TI
,,
rlr
,t,,,,
,
ül.lþ t.o
&
o121518212427o
TIME (rnin)30 33 38 39
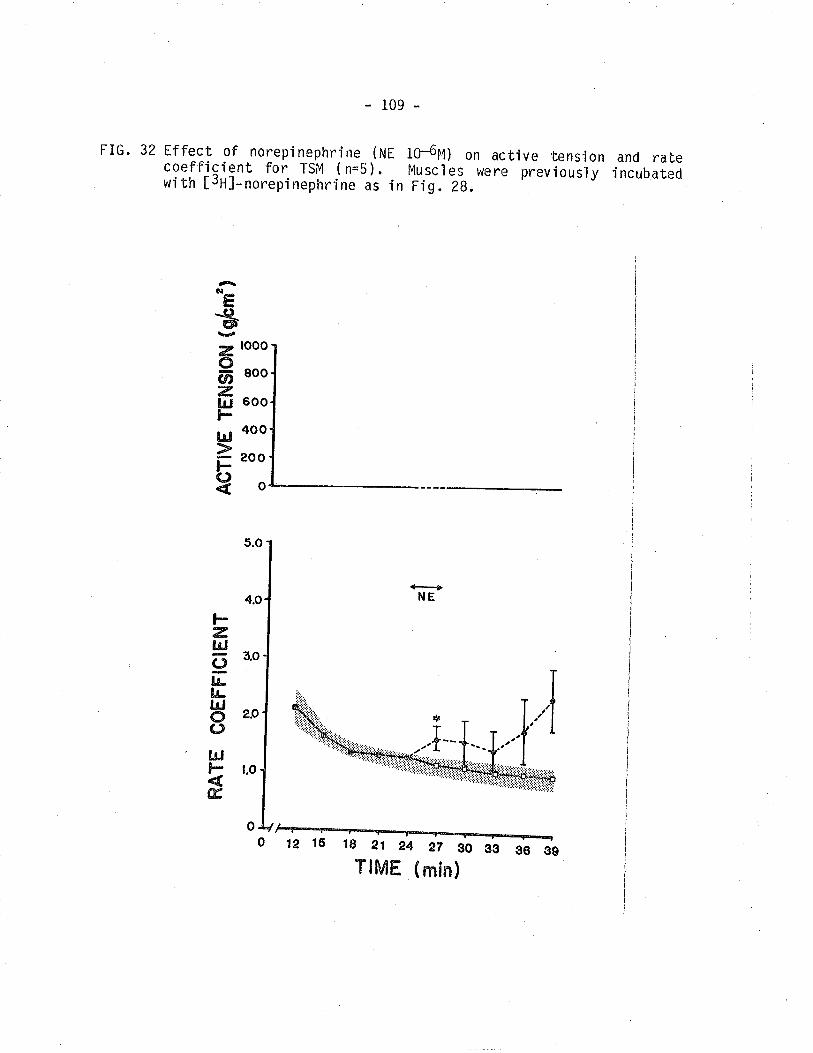
109 -
FIG.32 Effect of norepÍnephrine (¡¡e iñM) on active tensjoncoefticient for TSM (n=5). Muscles rvere prevÍousìywith [3H]-norepinephrine as in fig. Z-8.
20
Aô¡
lô
-.sæ'
?C)ffiz,bJþ&JÞts(.)
5.O
4.O
Þ&l¿J
õ 3.o'
Ë&L
8 zp'(J
looo
600
400
800
o-
o
o
<_'NE
*
t&lF- r.o4ffi
01215182124 27 30 33 36 39
TIME ( min)
and ratei ncubated
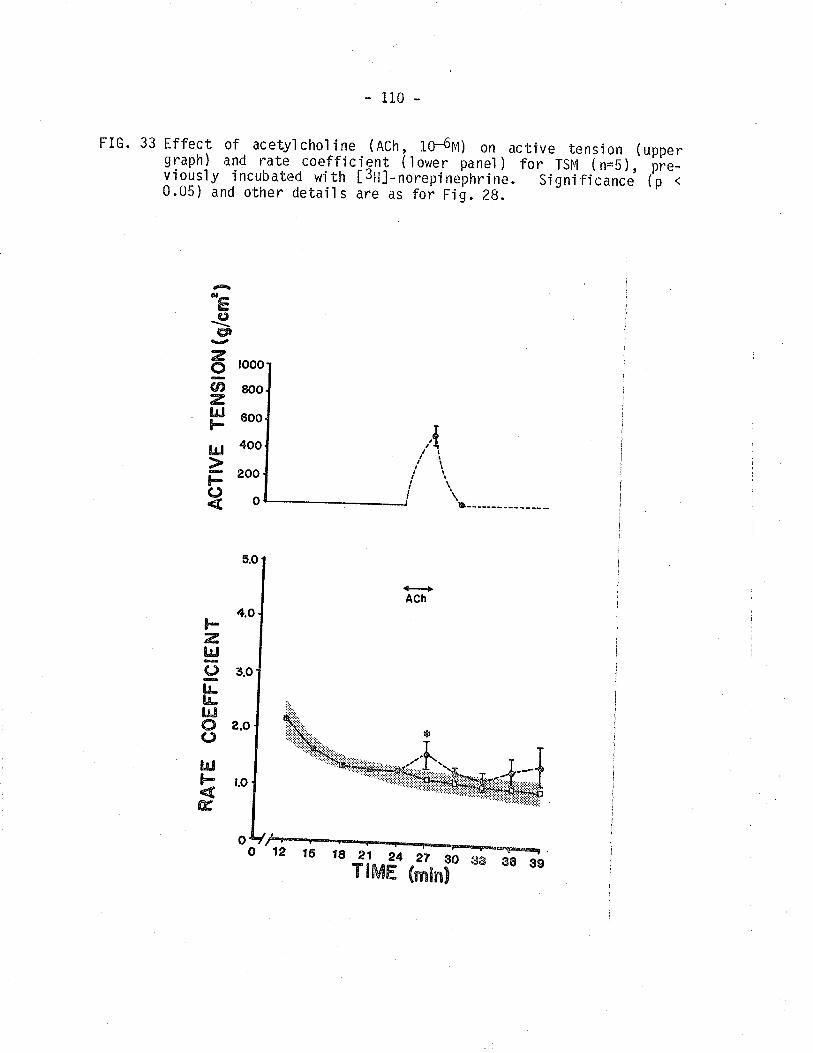
110 -
FIG. 33 Effect of acety'lchol ine (Ach, lÉM) on active tensiongraph) and rate coefficient (lower pane'l) for TSM in=s),yi9!:1v incubated wirh t3Hl-norepineþhrine. signifÍiance0.05) and other details are as foi Fiö. 28
( upperpre-(p <
A'Ë€Þ-,ÊÕ€f)3&dF&¡.l
F-(J4
Þ-æ,ge&Lh-tejÕ("¡
looo
800
600
4o0
200
o
5.O
4.O
3.O
2.o
&JF t.oqffi
<t
-}ACh
o12161821 24 27 s0
s
o
T¡Mg (müm);BS 38 39
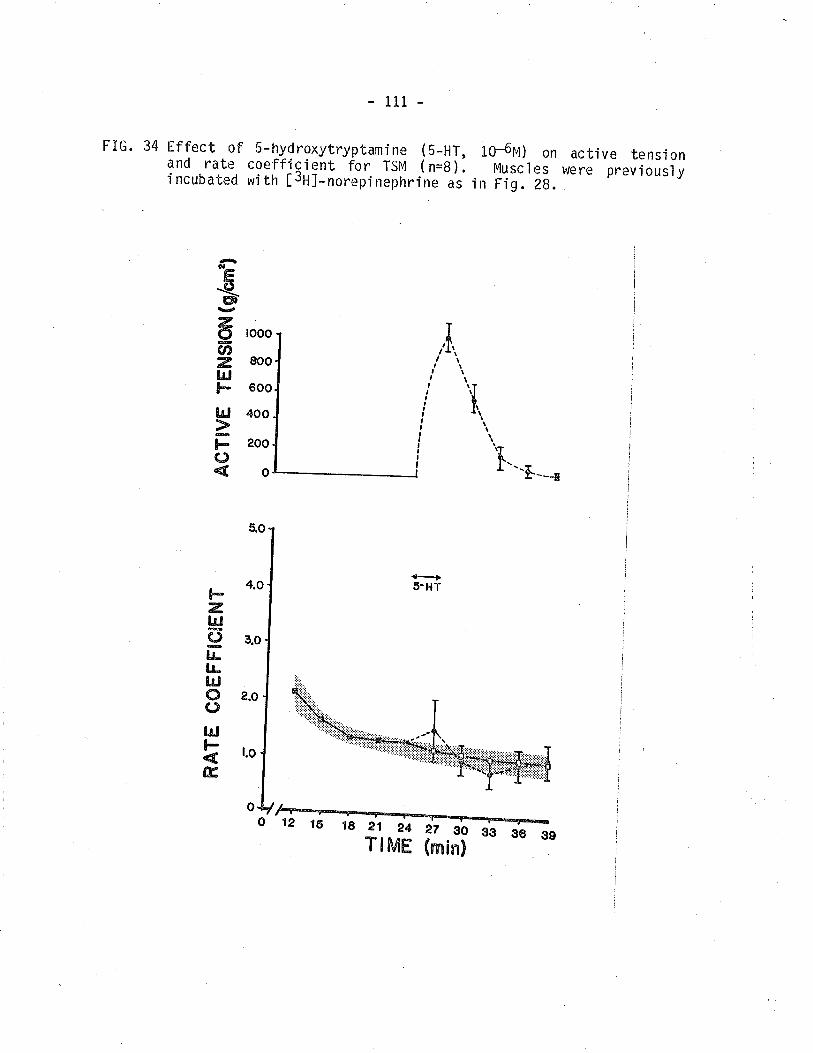
- lil -
FIG.34 E
aÍ
f fect of 5-hydroxytryptarnÍ ne (5-HT, lÉM) onnd rate coetfíçient for TSM (n=B). Musclesncubated wíth [3H]-norepinepnriìre as in fig. ãé.
*-€
GÞ-
.{-gtn-Ð&KJtskl
F.()
þ3H(JËu-UJe(}&Jt-€ffi
to00
800
600
400
200
o
5.O
4.O
IA,,\
l¡/ ',7ihl' 't,
3.O
2.O
'Ë-J- \-ï
Þ*--s
+-t5-HT
21 24 27 33 3g30T lFdE (min¡)
l.O
oo121618
g0
acti ve tensi onrvere previousìy

-Ltz-change in rate coefficient was not observed it was noted that 6 of I
muscles demonstrated an increase in rate coefficient fo'llowing the addi-
tion of 5-HT (1ÉM) . Sínce the method used i s not intended for quan-
titative estimation of transmiter release but rather to indicate
qual i tative changes, the present data suggest that 5 -HT (1ÉM) may
also promote tritium overflow.
In a separate study, the ef fect of tyrami ne (1ÉM) r,las observed
to be without significant (p > 0.05) effect on tritium efflux or active
tension development in canine TSM (n=7). The experiments vlere carried
out in the presence of TTX (1ÉM) and cocaine (1r7M); 'Ehe latter
may account for thÍs negat,Íve result.
G ) Neuromodul ati ng Infl uence of Several Factors on the Chol i nergicNervous System in Canine TSM in vitro
ExperÍments urere carrÍed out to determi ne the ef fects of s-HT
(1ÉM) , K+ (n mM) , EFs (15v , 60 HZ, Ac, 3 mi nute duration) , hi sta-
mi ne (1O-sM) , and norepi nephrine Of1 and tn4N) on the effl ux of
14C- choline. Following an Íncubation period to load the nerves
with radioactivity, a washout curve was obtained similar to the above
procedure for [3H]-norepÍnephrine studies. The first set of experÍments
described was carried out without eserine or TTX present.
Figure 36 il lustrates that a stable basal rest,ing tension in
untreated muscles was maintained for the duration of the experiment and
that the actual experimental values for the rate coefficient were similar
to the values predÍcted by extrapolation. Electrical field stimulation
increased tension and 14C-efflux significantly (p < 0.0S) whíle
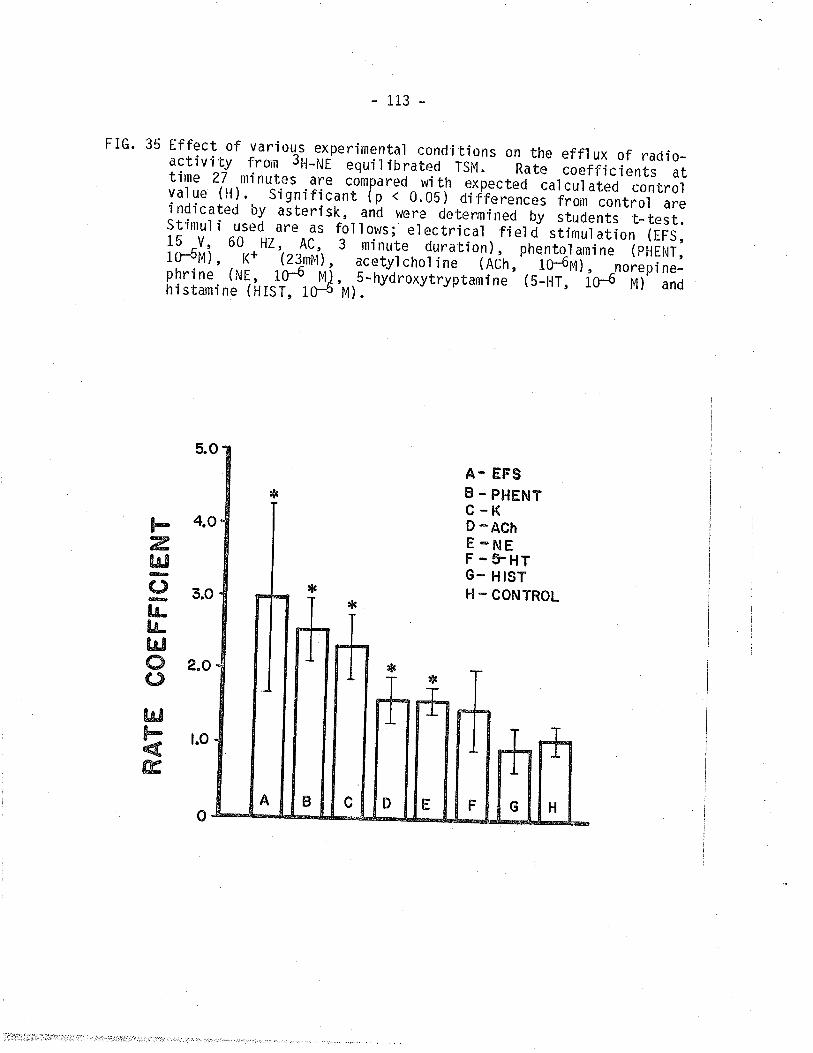
-113-
FIG.35 Effect of varÍous experimental condi'Eions on the efflux of rarlio-activity from 3H-¡¡r equf 1 ibrated TS¡q. - naîä coef ficients attirne 27 mi nures
. are cornpared *ji¡ ãip".tåä iar cur ated controrval,ue
f Hl. significanr (p < 0.0s) ¿îir*äi,i.i rro,n conrror areindicated by asterisk, an¿ were oetermtneä-ol students t-test.Stimuli used are_as foilows; electricar fierí stimuration (EFS,15 -V, 60 HZ, AC,. 3 minute du.ration), in.nto-larnine (plltNT,1ù-5M) , K* (zlrnrvr]., qcetyr ãnol in. iÁir,, ""îñi,41 ;"':r.;;;i;ì¿_phrÍne. (Nq, tF.^MJ,...5-hydroxyrryptamine (5-HT,
.í0_6 -Nj" än¿hÍstamine (HIST, l0-b M). -
þ%€_fuJ@
(-)üu&JÕ(J
k¡þ€ffi
*
*5.O
5.O
4.O
2.O
l.o
A. EFSB - PHENTc-KD -AChE-NEF-5-HTG- H ISTH - CONTROL*
* *
o
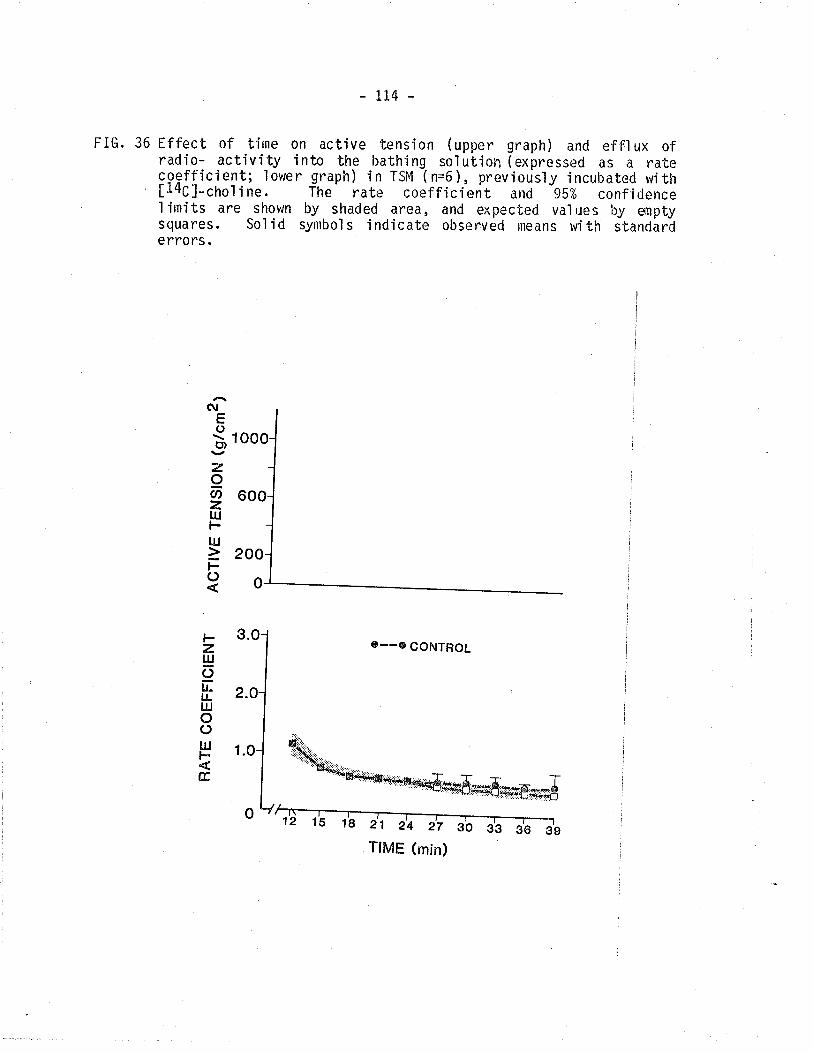
FIG.36 Effect of time on active tension (upper graph) and efflux ofradio- activity i nto th.e bathi ng sol ution (expressed as a ratecgefficient; lower graph) in TSt4 (n=6), previously incubated withti4cl-choliñe. ffre' rate coefficÍent and 95% confidenceIimits are shovrn by shaded area, and expected varues by emptysquares. Sol id symbol s indicate observed rneans rvi th standarderrors.
-114-
(Ð--@ coNTROL
1 1 242
ôtEoo)
zo6zTU
UJ
tro
3
2
ztrJ
õtrltIJJooIU
ftc
1 00o
60
200
1
0 I 1 30 33
TIME (min)

-115-histarnine stimulated isometric tension development without changing the
rate coefficient of l40-efflux, thus demonstrating, as for tritiated
norepinephrine, that contraction alone does not stimulate or increase
acetylcholine release (flg" 37). A similar result was found wTren high-
potassium and 5-HT were tested. Figure 38 illustrates the significant
i ncrease of i sometric tension and 14C-effl ux accompanyi ng a K+-
i nduced contracture and the contractil e effect of 5-HT wi thout a con-
comitant change ín rate coefficient.
Norepinephrine, for wl¡Ích indirect evidence suggests an Ínhibitory
effect on acetylcholine release (Vermiere and Vanhoutte, L979) was tested
at two concentra'L,Íons in the present studies" Figure 39 de¡nonstrates
that both concentrations of norepi nephrine (tO-7 and 1ÉM) t{ere
without sîgnificant effect (p > 0.05) on resting tension or
l4cefft ux.
The second and third types of experiments 0n factors wttich may
modify acetylcholine efflux !,Jere carríed out, in the presence of eserÍne
(lÉM), âr acetylcholinesterase inhibitor, or with eserine (lÉM)
and tetrodotoxi n (1ÉM) throughout the experiment. The anti-acetyl -
cholinesterase effect of eserine was expected to enhance the stímulated
overflow of acetylcholine by varÍous substances. TTX was used to ínhibit
ganglion-inÍtiated action potentials ín nerves" l,lhen eserine v,ras present,
histamine significant'ly (p < 0.05) increased 14c-efflux and this
effect was eliminated by TTX (Fig. a0). In the previous experirnent (Fig"
37) no increase of 14C overflow was observed"
The effect o't 5-HT on 14C-effl ux uras simil ar to that of hi sta-
mine but the amount of the íncrease (0 of the B muscles increased
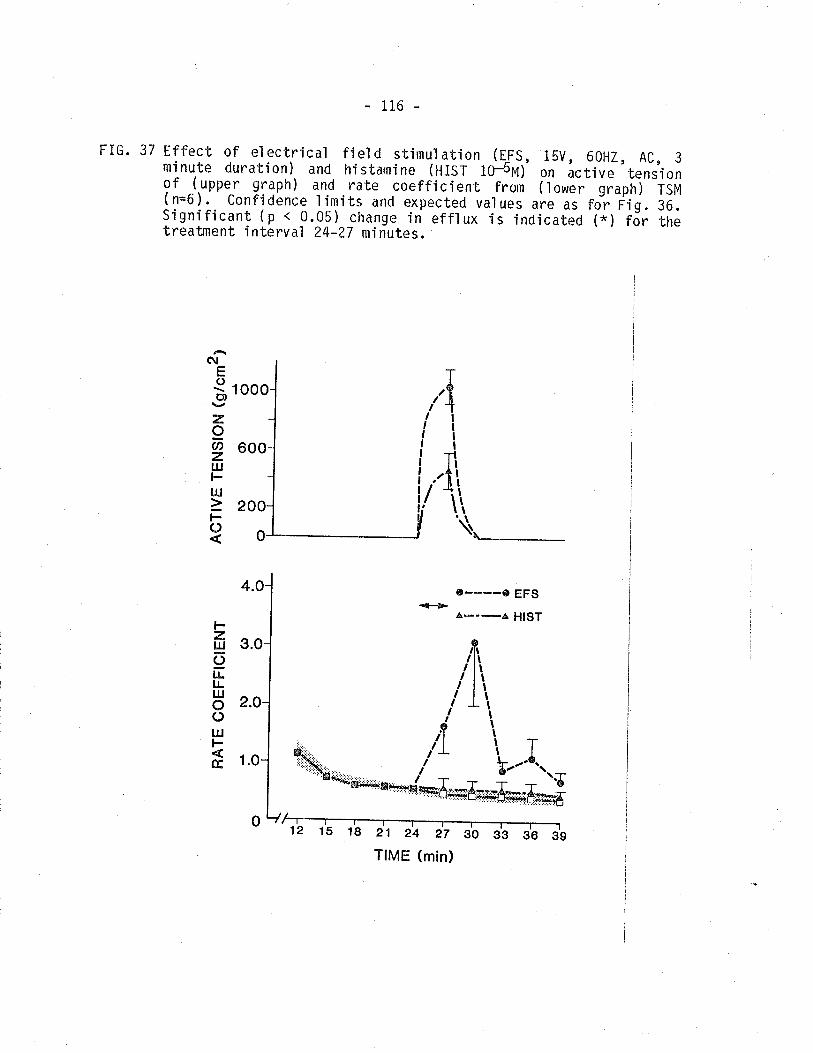
116
FIG. 37 Effect of electrical field stimulation (Efs, 15v, 60H2, AC, 3minute duraLion) and hÍstarnine (HIST tn5M)'on åctive tension9f _(.upper gfapr¡) and raüe coefficient from ilo*r- érupnl
-rõ¡¿
(n=6). Confidence limits and expected values are ãs ¡Jr Fig. 36.SignÍficant !p . _0.05) change in effl ux is indicate¿ 1-") rõr lrretreatment interval Zq-?7 minutes.
ñtEoo,
¿,oU)zUJ
UJ
o
F.zUJ
õlLILIJJoOu¡
G
100
60
2
3
2
1.0
IIII
\
4
12 15 1 21 24 27 30
TIMË (min)
6----6 EFS
aÈ--a HlsT
33 36 39
<þ
0
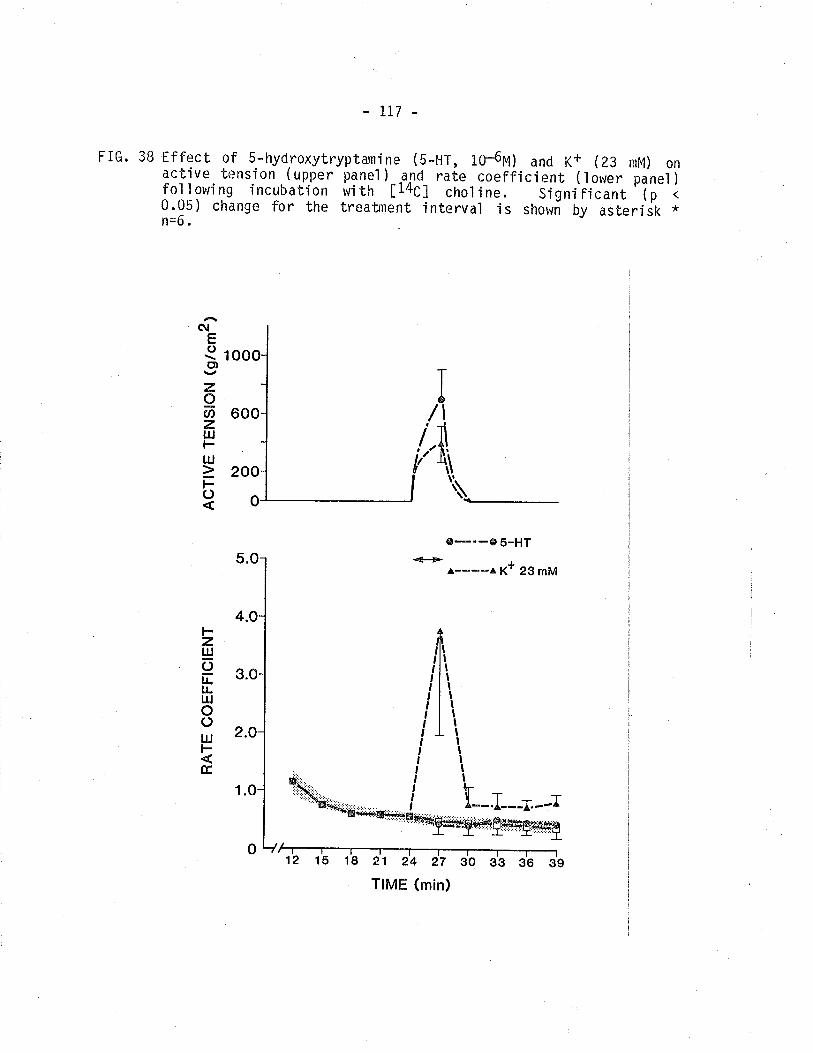
FIG. 38 Effect of 5.-hydroxytryptarnÍne (s-HT, to-{N) and ç+ (n mfil) onactive tensjon (up.per panel) ¡ld rate coefficient (lower punui)fo'llovríng íncubation with [14C] chol íne. Signi:ficJnt 'lþ- i0.05) change for the treatrnent interval is showñ by asterisk *n=6 .
LL7 -
I/l
€---6 s-HT
I
o01
NEog)
zIU)zUJþ[rJ
tro
5.0-
4.O-
3.0-
2.o)
,.1
.l
600
200
.@ra----aK'23mM
zulc)ËIJ.IJJoo[rJ
É,
IIII
21
I,lItIII,III,,
412 15 1
TIME (min)30 33
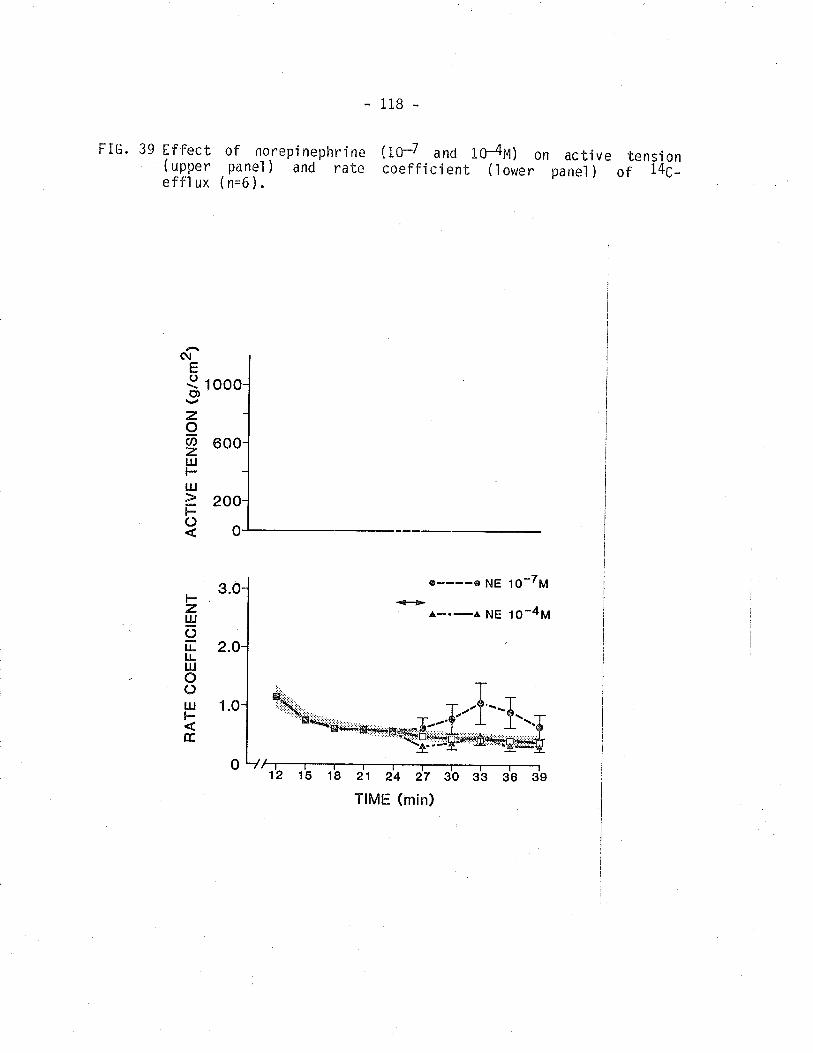
FIG.39 Effect( uppereffl ux
of norepi nephri nepanel ) and rate
(n=6).
118 -
(to-7 and 1ÉM) on activecoefficient (lov¡er panel )
tensionof 14c-
a
0-
0
100
NEoo)
zIU)¿-utt-TU
o
þ2uJou-TLtuoOtu
E
600
20
3.0-
2.O-
.@
6----@ ne lO-7¡¡
a-.8À rue lO-4U
1
18 2',1 4 30 33 36 39
TIME (min)
1
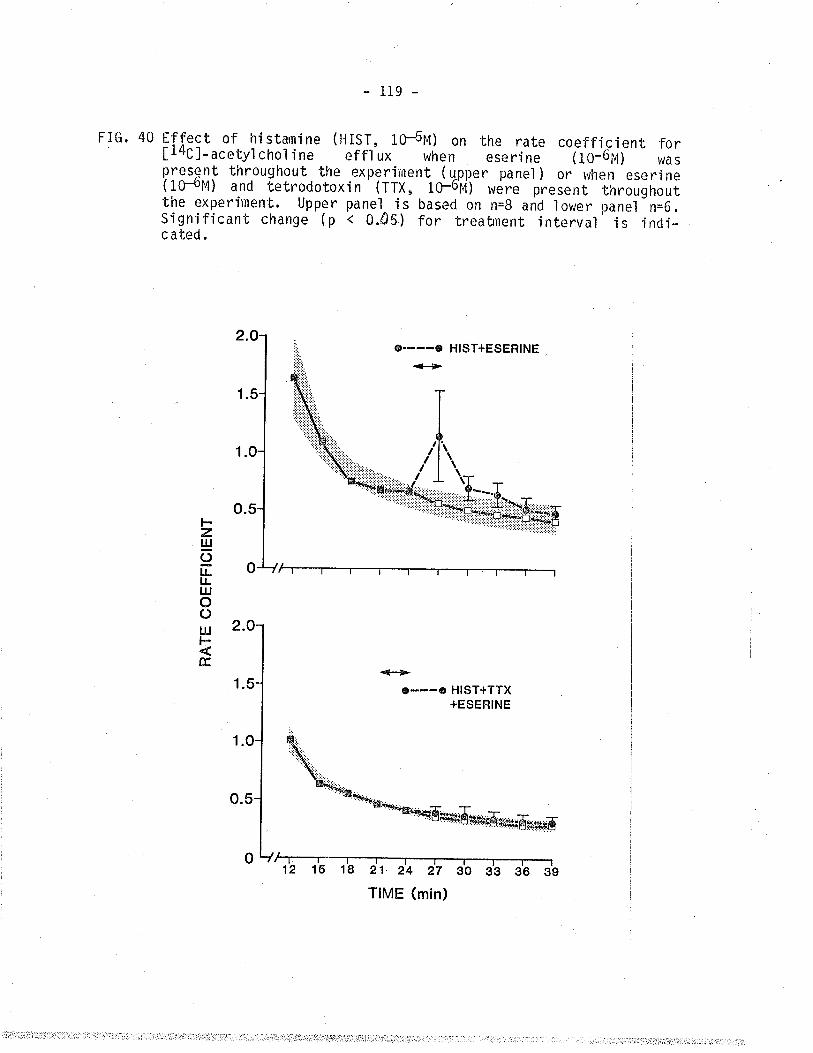
-119-
FIG.40 Effect of histarnine (HIST, 10-5M) on the rate coefficient fortl4cl-acerylchor ine erfr ux when äi.riiï ""iicijtìNl
was9f 9sÊ.lp thr.oughout the experirnent (çper panel ) or when eseri ne( l0{ M) and tetrodotoxi n (TTX, 1É14) were present throughoutthe experiment. Uppgr pane'l is based on n=B ari¿ lower panel n=6.Significant change (p < 0.CI5) for treatnrent interval is indi-c aied,
1
1
2.O
o
2
1.5
o
o
@.--.(Ð HIST+ESERINE.@
.{:-+
0
2uoTLTLUJooLUt-0c
1
@---@ HIST+TTX+ESERINE
24 30 36 39
TIME (min)
1 15 1

14c-effl ux after
120 -
5-HT) did not reach stati stical sÍgnificance (p <
0.05 ) .
41).
The action of TTX was to Ínhibit the response entireìy (fig.
In order to test for possib'le autoregu'lation of acetyì chol i ne
release in a resting state, both acetylcholine and atropine u/ere tested
in the presence of eserine and TTX. The results índicate that neÍther
was effective ín alteríng 14ç-.tflux (Fig. 42).
Pneviousìy (fig.39) a trend for norepinephrine (10-7M) to en-
hance 14C.effl ux was observed and thi s effect !'Jas enhanced, though
non-significantly (p > 0.05) by eserine (rig.43 upper pane'l). A higher
concentration of norepi nephrine (1ÉM) was observed to depress the
efflux of 14C in the presence of eserine; when eserine and TTX were
present throughout the experiment (Fig. 44), the effect was signÍficant
(p < 0.05). An attempt to demonstrate the relevance of this findíng by
using tyramine (tO-4u), an agent which releases norepinephrine from
adrenergíc nerves, was unsuccessful (Fig. 43, lower pane'l).
Since nerves are thought to release transmitter at a basal rate, two
antagonist,s of norepinephrine actions vvere used. The effect of phentola-
mine, an alpha-adrenergic antagonist was without effect on 14C over-
flow, whereas the beta-adrenoceptor antagonist propranoloì depressed the
effl ux of 14C signíficantìy in the presence of eserine and TTX,
(Fig" a5). This effect was similar to that observed for a high concentra-
tion (1ÉM) of norepinephrine (Fi9. 4a).
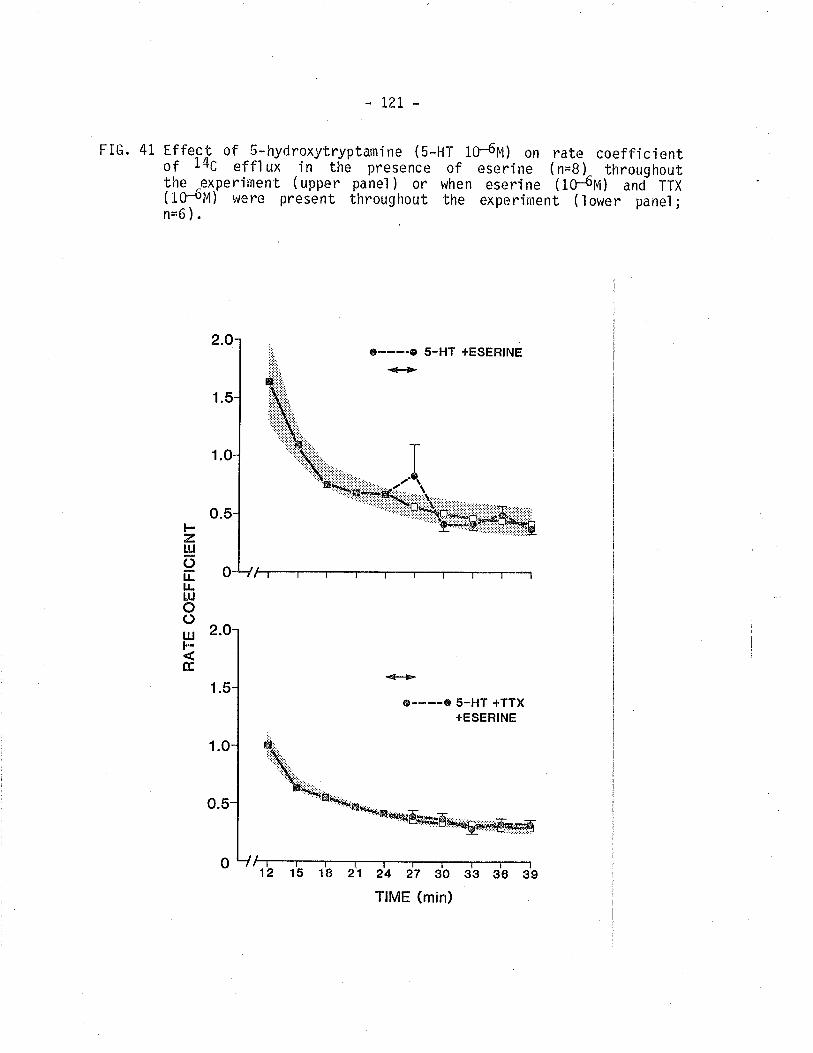
FIG.41 Effe-ct of 5-hydroxytryptamine (5-HT 1ÉM) on rate coefficientof 14c ef fl ui i n' tñå presence of eseri ne ( n=-a )- ihroughoutthe -experii¡ent ( upper panel ) or when eseri ne (1ÉM) an¿ ttX(10-oM) were present throughout the experÍinent (l ower pane'l ;n=6).
-121.
Ð----(Ð 5-HT +ESERINE<'+
.<Þ
€----e s-HT +TTX+ESERINE
1242730333639TIME (min)
zIUõILILLUoolUl--
E
2.O-
1.5-
1.0-
o
2.O-
1.5-
4
1.
0
o-
1 12 15 18
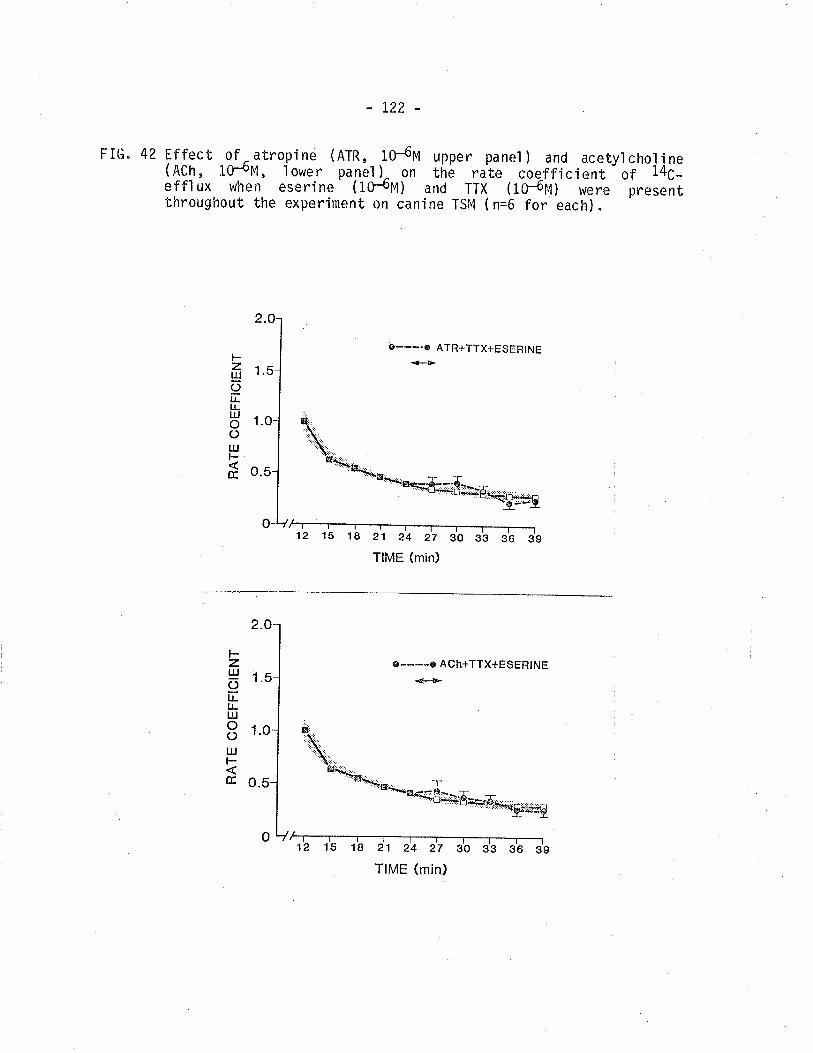
-L22-
FIG. 42 Effect o-f . atropine (ATR, to-6N upper panel ) and acetylchol ine(Rch, lrM, iower panel )^ on tire rate coefficient of 14c:effl ux when eserine (10-bM) and TTX. (tn6tq) viere presentthroughout the experiment on canine TSM ( n=6 for each).
0.5
FzgoLLLLtIJootrjF-
E
2
1.5
1.0
O---.@ ATR+TTX+ESERINE.e
12 15 18 21 24 27 30 33 36 39
TIME (min)
0
t--zTIJ
C)ËLLLUoo[rJl--
(Í.
2
1.5
1
e----@ ACh+TTX+ESERtNE.@
12427o
12 1
TIME (min)33 36
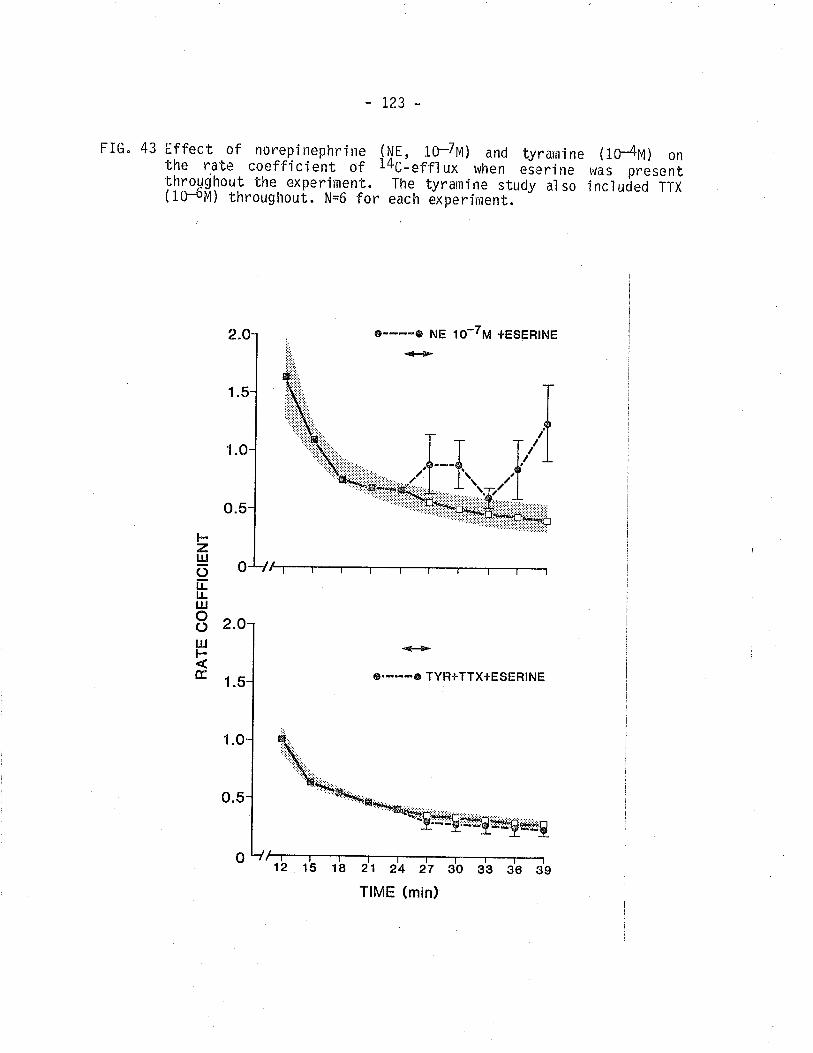
FIG.43 Ëffect of nore-pinephrine lryE, 1O-iM) and tyrarninethe rate coefficient of 14c-effrux when eierinelirlolgloyt the experiment. The tyrarnine study al so( lù{ M) throughout. N=6 for each óxperinrent.
123 -
@----@ rqe ro-7¡¿ +EsERINE.e
<+
@.-.-o TYR+TTX+ESERINE
12427
(1ÉM) onvvas presentincluded TTX
o
þ2tuõLLILTUo^o¿.IUþrr
1.
2
1.5
1.0
o.5
5
1
o
10 15 18
TIME (min)33 36 39

FIG.44 flffect of norepinephrine (NE l.ÉM) on the rate coefficient of14c-effl ux when eseri nã ' iro-¡-r'l)
"'wa"s' present throushout theexperr'ment (upper panel ) or when eserÍne (1É14) and TTX
t rtrum) were present throughout. N=6 for each study.
-124-
@---.e ¡¡e ro-4n¡+ESERtNE<"+
.sø---e ¡¡e lo-4 tr¡
+TTX+ESERINE
'--E124
2
1
1
o
0
t-zTU
õËltIJJootrJ
tr
2
1
1
o
1 1o
1
TIME (min)30 33639
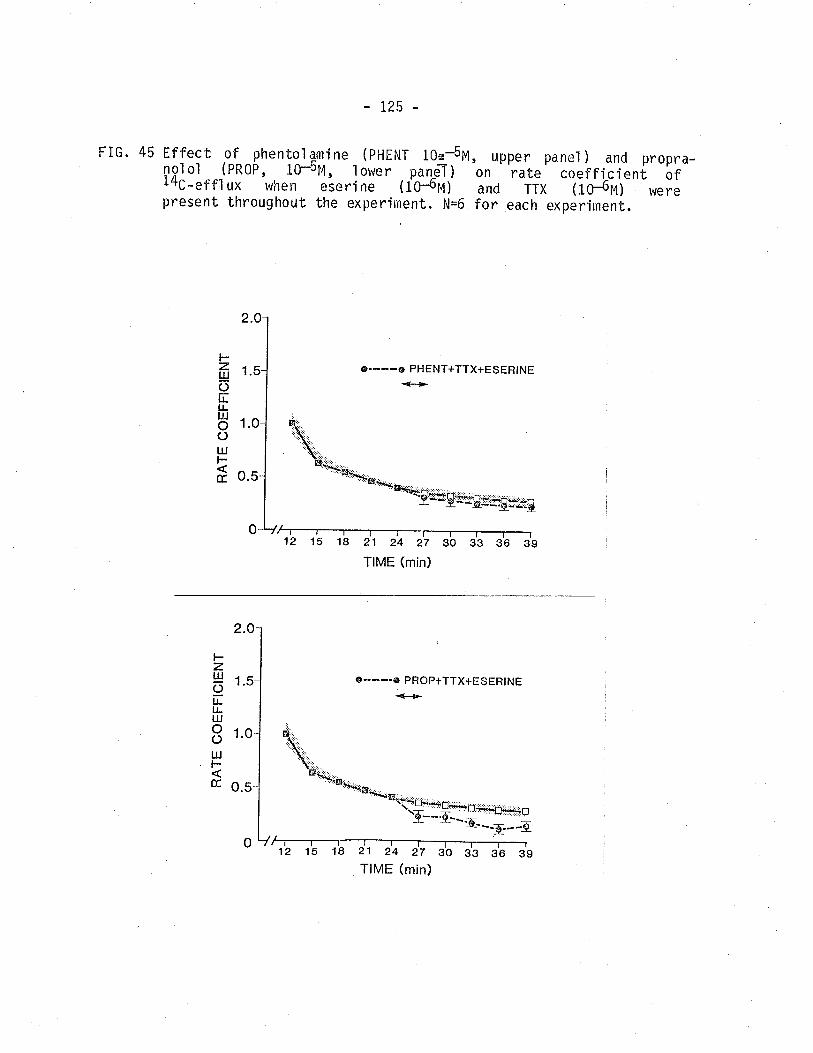
FIG. 45 Ef fect of ^phento^l ÊTi ne
_ (pHrruT to.--sN, upper panel ) and propra-
îPiol _-1PROP, 10-bM, 'lower parlel) on rate coeffícient ôrrîc-efft ux when eserÍne (lÉM) and rrx (lÉM) *.iupresent throughout the experirnent. N=6 for each experiment.
-L25-
@----@ PHENT+TTX+ESERtNE.+
2
l--zLrl
olJ.-ILtrJootlJl--
Ír
1
1
0.5
012 15 18 21 24 27 30 33 36
TIME (min)
F'zLU
oLLILIIJootuF.
0t
1
1
0.5
2.O
0
@----@ PRoP+TTX+ESERINE<-Þ
-g---Þ124
TIME (min)0 36

-L26-
D ISCUSS ION
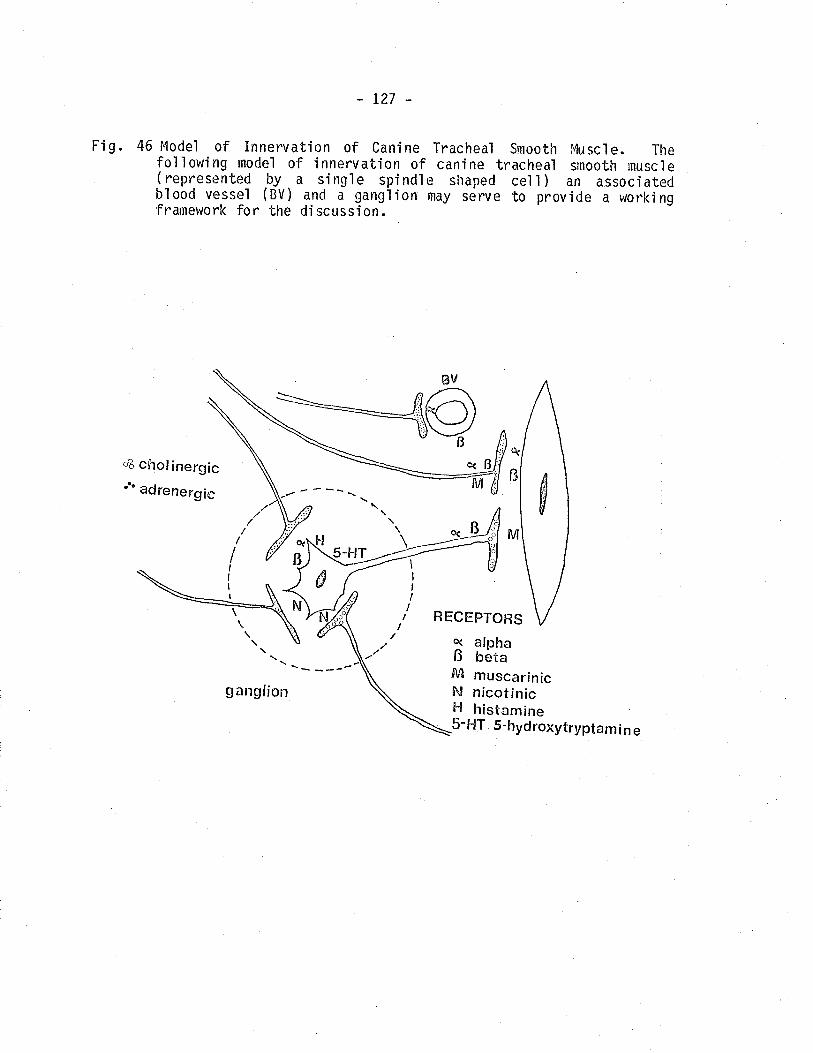
-127-
Fig. 46 Model of Innervation of Canine Tracheal Smooth Muscle. Thefoììowing tnodel of innervation of canine tracheal smooth muscle{ represented .by . a si ng'le spi ndl e shaped cel I ) an associ atedblood vessel (BV) and a ganglion may sdrve to provide a rvorkingframework for the discussion.
BV
coB chol inergic..'adrenerqic ß
VI
o<
M
ß
ß
N
-HTII,IIt
RECEPTORS
o< alphaß båtafL.l mt¡scarinicN nicotinic!-l histamine5-HT S-hydroxytryptamin e
ganglion

- 1.28 -
D ISCUSSION
A) Electrical Fieìd Stimulation and Isometric Tension Development
Electrical field stimulation of canine tracheal smooth muscle in
vitrg_ was used to demonstrate the stimulatory effect of nerves in the
preparat,ion. Previous s'L,udies on dog tracheal is in vitro (Russell, 1978)
ancl in vivo (Brown et. â1., 1980; Brovln, êt. â1., 1982) have shown the
parasympathetic system to be the mediator of contractile responses, while
the role of the adrenergíc system Ís controversial (Nadel, 1980). Our
studies indicate tha'E tFS stÍmulates cholinergic nerves, the response
(isometric tension) being enhanced by eserine and blocked by tetrodotoxin
which has a selective action on nerves (Bolton, L979; Narahashi, 1972),
or hyoscyamine which blocks muscarinic receptors. Simil ar resul ts !,Jere
obtained by Co'lebatch and Halmagyi (1963) and 0lsen, êt. al. (1965) who
demonstrated that atropi ne abol i shed i sometrÍc tension foì I owi ng EFS
stimulation, thereby indicating that muscle stÍmulation occurs via
postgangì Íonic vaga'l fibres.
I n vi vo studi es have shown a functiona'l adrenergic neural innerva-
tion of canine trachealÍs and suggested that the rnajor role was to pro-
mote relaxation (Russe'll, 1980; Brown e'ü. â1 ., 1980). Others (Beinfield
and Seifter, 1980) have demonstrated an alpha-adrenoceptor mediated con-
tractile response to low doses (¡ x 1É to 2,4 x 1ù€ mol/kg) of
exogenous norepinephrine administered intravenously in the absence of
beta-adrentrceptor b'lockade. However when Cabezas et. âl ., (1971 ) elect-
rically stimulated the sympathetic nerves in vagotomÍzed dogs no contrac-
tile activÍty was apparent and they interpreted these findings in terms

-129-of an absence of alpha-adrenoceptors in canine airways. gur studies ofTSM in vitro show a signifÍcant inhibition of the electricaìly stimulatedi sometric tension by the al pha-adrenoceptor antagoni st phentol amÍ ne.when a beta-adrenoceptor antagoni st propranol oì was used i sometríctension vltas reduced in all muscle strips. This unexpected decline may be
due to the anesthetic actÍon of high concentrations (> lù-sM) of pro-pranol ol . The addÍ tion of norepi nephri ne (1ÉM) was wi thout effect on
electrically stimulated tension development whereas a higher concentra-tion (1ÉM) non-signifÍcantly reduced tension in al I muscle strips.An attempt to block the relaxant effects of norepinephrine wÍth proprano-Ioì resulted in a 'Further reduction in isometrÍc tension. we attributethis unexpected decline to the possible anesthetic action of propranolol(1r5M) .
conclusions from these studies are that canine TSM is predominantlyi nnervated by chol inergic fibers, which when stimul ated e'lectrica.l.lymediate contractile responses by releasing acetylcholÍne which acts 0n
post-iunctional muscarÍnÍc receptors. AdrenergÍc fibers are al so stirnu-lated by EFS and the resurting rerease of norepinephrine may act on post-iunctional alpha- or beta-aclrenoceptors to med'iate contracti'le (phentola-mi ne sensi'bive)
i vely. I't i s
or relaxant (propranolor sensitive) responses, respect-
likely that an a]pha-adrenoceptor-meclÍated contraction,following treatment with hyoscyarnÍne, is not observed with EFS due to itsdependance on development of adequate active tone prior to stimulation.The small amount of EFS-induced tone remaÍning fo1'lowing hyoscyamÍne may
be due to stimulation of muscle directly or another, as yet unÍdentified,phentol ami ne-insensi tive stimul atory system.

130
B) Effect of Tone on Adrenoceptor Mediated Responses in Canine TSM invi tro
The present s'üudies investigated the effect of tonus on adrenocep-
tor-mediated responses Ín TSM. l^lhile controversy surrounds a functional
role of the adrenergíc system in hunran airways (Stone et. â1.,1973) itis generally agreed that beta-adrenoceptors have a functional role post-
junctionally in both human (Jack, L973; Fleisch, 1980) and canine airway
smooth rnuscle (Beinfield and Seifter, 1980; Bergen and Kroeger, L9791.
The existence of alpha-adrenoceptors mediating contractile responses is
less clear with some studies support,ing their presence (Beinfield and
Seifter, 1980; Kneussl and Rícharelson, 1978) and others finding no
evidence for their presence in airway srnooth muscle (Foster,1966; Stone
et. al., t973; Cabezas, êt. al., 1971). The reasons for this discrepancy
may be attrÍbuted to differences in the tonus of the respective muscles
at the time the a'lpha-adrenoceptor agonists were app'lied. Several recent
studíes (Bergen and Kroeger,19B0; Kneussl and Richardson,1978; 0hno et.
â1.,1981) have suggested that for canine TSM in vitro tone developed
prior to al pha-adrenoceptor stirnul ation f s requi red to demonstrate a
response,
Norepi nephri ne ( tO-B to 10-4M) was wi thout mechanical
effect in resting TSM in vitro even in the presence of propranolol.
Unlike the findings of Leff, et. â1., (1981) an Índependant effect of
al pha-adrenoceptor activat,ion v,ras not gradua'lly 'lost but was absent from
the earliest time at which the muscle was equilibrated (one hour) and
throughout the duration of the experiment, ( up to 6 hours) .
Since NE activates both alpha- and beta-adrenoceptors, the complex
responses seen are not surprising. The tone-dependence of the alpha-

-131-mediated component and the relative dorninance seen at various concentra-
tions of NE, however, is novel. The sarne muscles wltich had shown no
response to NE alone produced a contractile response to NE (1É to
1ÉM) after tension was augmented with ç+ (n mM). Further increases
in NE (1É and to-4u) produced a shift to relaxation (rlg. 5)" 'Ihe
contraction vlas mediated by a1 pha-adrenoceptors, relaxation by beta-
adrenoceptors as evidenced by the sensitivity to phentolamine and propra-
nolol respectÍvely. In considering the basis for this pattern of response
$re considered the possibility that the respective receptors might have
di fferent threshol ds for response. Figure 5, however, dernons'trates
similar thresholds for aìpha- or beta-adrenoceptor mediated responses in
the presence of propranolol and phentolomine, respectively. The finding
of Vermi ere and Vanhoutte Í979 ) that NE may Í nteract wi th chol i nergic
nerves via pre-junctional receptors provided another possible exp'lana-
tion. lllhile this interactíon is of importance it appears that the bi-
phasic response is independent of the cholínergÌc system since it is
unaffected by atropine (Fig. 6). lllith reference to the alpha-adrenoceptor
medÍated contraction its magnitude is striking as seen by the rise to
?23% above the initial pl ateau fo'll owi ng 6+ (22.4 mM) exposure in t'he
presence of NE 1É and propranolol (flg. 6). 0f special interest is
the fi ndi ng that whil e beta-adrenoceptor bl ockade greatly enhatlces
a'lpha-adrenoceptor-dediated contractions the antagonist is not required
to unmask the receptors as others have found for canine TSM iI vivo
(Beinfield and SÍefter,1980). When muscles were pretreated with
propranol ol al 1 concentrations of norepi nephri ne ì tO-€M produced
contract,ions which were antagonized by phentolamine (Fig" 7).

-r32-Our attention then turned to the functional role of these receptors.
The addÍ tion of tyrami ne (1É to 1ÉM) 'uo 23 mM K-stimul ated
muscles resulted in contractile responses which u/erq augrnented by pro-
prano'l o1 (10-5M) and dirni ni shed by phentoì amÍ ne (1Û-5M) " These
resul ts suggested the functional importance of the adrenergÍc nerves and
the respective alpha- and beta-adrenoceptor-mediated responses to their
neurotransrnitter release. Atropine did not alter the qualitative aspect
of thÍs response (Fig.8). Since the possibility of a direct effect of
tyramine should not be ignored, muscles tvere stored for 3 days at low
tentperature (4oC), a procedure known to destroy adrenergic nerves
(Hattori et. 41., L972); in these muscles the response to tyramÍne was
lost but the biphasic response to NE remained. These findings therefore
demonstrate the adrenergic neural component of canine TSM and the prob-
able innervation of alpha-adrenoceptors" Stimulation of nerves to air-
ways in vivo has dernonstrated a function role of adrenergic nerves in the
dog (Russell, l9B0; Suzuki, êt. al.,1976).
The nature of the alpha-adrenoceptor-meclíated component and biphasic
response was found to be Ínfluenced by the pre-existent tone in the
preparation, the use of adrenergic antagonists and the specific agonists
used to índuce tone. l^lith reference to the level of pre-existent tone a
reduction of both contractile and relaxant responses (as a % of the K+-
induced contraction) following addition of norepinephrine was observed in
48 mM K-stimulated muscles. The alpha-adrenoceptor mecliated response vrtas
demonstrated only in propranolol pretreated musc'les, while a reduced but
a'bropine-sensitive component of the total contraction was still evident
(Fig. e).

-133-Figure 10 demonstrates that a maximum alpha-adrenoceptor mediated
contraction is produced vrhen initial tone is deve'loped wilh ?2.8 mM K+.
This contractile response to NE was rnaximal at a concentration of 10-6N
and greatly augmented by the addition of propranolol (1ÉM). As basal
tension was increased by increasing concentrations of 6+ the alpha-
mecliated repsonse (g/cn?) declined and'lhe beta-adrenoceptor-mediated
relaxant response increased in absolute terms"
Al though the dose-response rel at,ion to NE (1É to 1ÉM) was
biphasic, the shift from aìpha- to beta-adrenoceptor-mediated responses
occurred at a lower concentration (tO{lr4 NE) when hÍstamine or acetyl-
cholÍne were used to initiate tone (Fig.11 and 13), than when K+ solu-
tion producing an equivalent amount of initial actíve tone was used. The
reason for this difference in a shift of the biphasic response is not
known "
The dependence of the al pha-adrenoceptor-medi ated contractil e
response on tone is further supported by the finding that hyoscyamine
whÍch eliminated tension stÍmu'lated by acetylcholine, also eliminated the
exísting alpha-adrenoceptor mediated contraction. ThÍs suggests that not
only is tone required to unmask an a'lpha-adrenoceptor response but that
tone i s requi red to rnai ntai n i t even when a substanti al a'l pha-adrenocep-
tor-mediated active tone is al ready present. l¡lhen histamine was used to
devel op tone initia'l'ly hyoscyamÍne al so reduced the adrenoceptor-stimu-
tated contraction to near initial tonus ìeve'ls, but the contraction was
not abolished as for acetylcholine. The fact that a'lpha-adrenoceptor
mediated contract,ions are not altered by atropine as vlas dernonstrated
earl Í er suggests that hyoscyami ne wl'rich has the same mode of action as

-134-atropine inhíbits a component of the histamine response. The proportion
of the remaining contraction wl'¡ich vlas produced by al pha-adrenoceptor
stimulation was not determíned.
Thus, these studies provide further evidence for an adrenergic
innervation of canine TSM and the functÍonal presence of both a'lpha and
beta adrenoceptors. l^lhile others have demonstrated that elevation of tone
may unmask a'l pha-adrenoceptor responses in human (Kneussl and Richardson,
1979) and dog TSM in vitro (Ohno et. ô1., 198i; Leff et. â1., 1982), the
relationship of the response to varying the active tone has not been
studied previously. The present work, as previously reported (Bergen and
Kroeger,1980), demonstrates a relationship of inÍtial active tone of the
muscle to the biphasÍc dose-response relation seen with Íncreasing NE
concentrations (1É to lÉM) . l¡Jhil e the role of the adrenergic
system in producing relaxant responses is widely accepted for canine air-
ways the contractile response has been questioned. llhether a contractile
response is observed is likely dependent on the resLing or active tone of
the preparation" Cabezas et. al. (L97L) were unable to demonstrate an
alpha-adrenoceptor mediated contractile response to electrical stimula-
tion of sympathetic nerves Ín vagotomized dogs treated with propranolol
in vivo. It is likely that active tone or resting tone following vagotomy
'Jl,as reduced and therefore a'l pha-adrenoceptor medi ated contractions were
not produced as we have seen for canine TSM in vítro.
C) Adrenoceptor Mediated Responses in Sensitized Canine TSM
Interest in the role of alpha-adrenoceptors mediating aírway narrow-
ing merits evaluatíon in the light of recent findings demonstrating an

-135-i ncreased contracti 1 e response of di seased hunran ai rways i n vi !r.o
(Kneussl and Richardson, 1978) and in asthmatics (Henderson et. â1.,
1979; Patel and Kerr, 1975) following stirnulation of alpha-adrenoceptors.
The role of adrenoceptors in airway hyperreactivity is of interest from
several perspectíves. Changes in the nunrber or activity of beta-adreno-
ceptors nray occur (Szentivanyi, 1968; Szentivanyi; L9791. Recently Barnes
et. al . (1980) dernonstrated an increase of a1 pha-adrenoceptor-agonist
binding in al1ergíc guinea pigs. Changes in basal tone were observed ín
a1 1 ergic dogs ( Antoni ssen et. ôl . , L979) and may al so contribute to
altered adrenoceptor responses in the light of the study dÍscussed above.
The present study examined the adrenrrceptor-mediated responses in dogs
sensilized to ovalbumin and cornpared the findings to those of a non-
sensÍ tized mongrel dog popul ation. The resul ts i ndicate simil ar
responses, both qualitatively and quantitative'ly for alpha-mediated con-
tractions and beta-mediated relaxant responses in sensítized and control
TSM. The biphasic response was unaltered for sensítized when compared to
control responses.
When resting muscle strÍps from sensitized dogs tvere exposed to
tyrarnine tO-4N they contracted while control strips did not respond
even in the presence of propranolol. Propranolo'l enhanced the contrac-
tile responses to tyramine and norepinephrine in sensitized rnuscle strÌps
and 'bhe a'lpha-adrenoceptor na'L,ure of the contractil e response hras shown
by Í ts sensi tÍvity to phento'l ami ne. In order to devel op a contractil e
response to norepinephrine, rnuscles from 0A sensÍtized dogs t,'tere pre-
treated with propranolol; tyramine-mediated contractíons, however, !úere
produced in unblocked resting sensitized TSM.

-136-I,'le conclude by att,ributing increased a1 pha-adrenoceptor-mediated
responsiveness of sensitized TSM to an increased restíng basal tone Ín
these muscles although we dÍd no't, measure the latter directly. In view of
the sirnilar responses to norepinephrine and tyratnine observed when tone
was increased, it is unlikely that the sensitization of dogs to ovalbumin
changes adrenoceptor status qualitative'ly, although these results do not
suggest that altered adrenoceptor status is improbable in other situa-
tions. The possibility that dÍsease states alter basal tone of airway
srnooth muscle requÍre further examination. The factors which may alter
resting tone and hence subsequent responses to adrenergic stimul ation or
cÍrculating catecholamines are not well understood. Cholinergic over-
activity may contribute to increased resting tone in vivo but this is an
un1 ikeìy exp lanation for data obtained in vitro. Resting tone may be due
to increased "leakiness" of the mernbrane to ca1cíum, and if this occurs
in sensitized canine TSM, we would expect increased responsiveness to
a1 pha-adrenoceptor stimul ation due to the dependance of the a1 pha-
mediated contraction on tone. The increased tone may also result from an
intracellular change wlrich regu'lates the contractile elements of TSM.
D) Actions of Histamine and Serotonin on Canine TSM
l4uch experimental interest has centered on aírway responsiveness to
hÍst,amine in normal and disease states such as asthma. Histamine plays a
major role in allergic asthma but the precise location of its action is
not known. Several sites of action are suggested: post junctional H1
receptors on srnooth muscle (Antonissen, et. â1.,1980; HimorÍ and Taisa,
1978; Nathan et. ô1.,1979); an irritant receptor in the epithe'lium of

-137-the airways (Kessler et. a1.,1973; Wassemlan, 1975; Yanta et. al.,1981)
and at an unspecified site interacting with the cholinergïc efferents to
airway srnooth muscle, perhaps in ganglia (Loring et. ô1., L97B). Less is
known of the role of serotonin but it also 1ike1y acts at several sites
to produce airway constriction. These include 1) receptors on airway
smooth muscle (llahn et. âl ., I97B; 0ffermeier and Ariens, 1966) tne
adrenergic or cholinergic system at the level of the ganglia (Hahn et.
â1 ., l97B; Dixon et. dl ., 1980; Sheller et. ô1 ., 1982) and 3) tl.re vagus
nerve to promote constrictor activity (Sheller et. al., 1982).
The present studies have focussed on the interaction of s-HT and
histamine on the cholinergic and adrenergic systems in canine TSM from
animals sensitized to ovalbumin, their lÍtter mates and a mongrel control
popul ation"
Hi stami ne stimul ates contractíon of TSM. The parti al (50"/") atro-
pine-sensitivity of thís response (Fig.23), suggests an actíon of hista-
mine on the cholinergic system to promote acetylcholÍne release. Besides
atropine sensitivity, several other of our studies support this view"
Eserine effects a hyoscyarnine-sensitive enhancement of the histamÍne-
induced contraction (Table 5). Other studies using histamine aerc¡sol
(Drazen and Austen,1975) or a specific antigen thought to release hista-
mine from mast cells (Gold, L975; Snow et. â1., L979) demonstrated a
marked inhibition of the constrictor response when atropine was admini-
stered prior to challenge. These studies suggested that histamine vlas
stirnulating a vagal reflex by acting on the irritant receptor (Mortola
et. al., 1975; Vidruk et. a1., 1976; Gold et. a1., L972). In the present
studies using TSM in vitro frorn which the mucosa has been removed, it is

-138-unlikely that either a reflex or irritant receptors remain" However the
ganglia present near or in the outer layer of smooth muscle may be intact
providing a possible site of histamine interaction with the cholinergic
efferents in vitro. Further evidence in the present studies demonstrated
dÍrectly a release of 14C- choline froln cholinergic nerves (rlg.40).
This measure of overflow of radioactivîty, which was Ínterpreted in terms
of neural release of intact acety'lcholine, was not sígnificantly elevated
in the absence of eserine (Fig.37) or the presence of tetrodotoxin (Fig.
40 lower panel). Thus it appears that histamine promotes the release of
acetylcholÍne from cholinergic nerves. The ínhibÍtory effect of TTX
suggests that hÍstatnÍne acts at the level of the ganglia to enhance ACh
efflux by increasing the action potentÍal frequency or that the stimula-
tory action of histamine on the nerve, at the ganglia or varicosities,
requires a TTX-sensitive influx of sodiunr ions. The finding that eserine
is required Ín order to measure an increase in overflow suggests that
neural elements efficíent1y take up the choline product of ACh hydroly-
sis, and that the effect is not as large as that seen following electri-
cal field stimulated overflow in the absence of eserine (Fig.37).
lli stamine induced contractions were al so partial ly inhil¡ited by
phentolarnÍne a1though this effect was smalIer than that following atro-
pine (Fig. 22). I¡lhil e this finding suggested that histamÍne may stimu-
I ate neural NE rel ease the overfl ow studi es usi ng 3H-l'¡f ¿tO not support
t,his possibility (Fig. 29).
The Burn-Rand hypothesis (Burn and Rand,1965) suggested a link of
cholinergic Ínfluence on adrenergic nerves and visa versa, based on the
close anatomical assocíatÍon seen for these two systems Ín the intestine.

-139-Manber and Gershon (1979) in fact demonstrated that such an interaction
occurs in intestÍnal smooth muscle. A similar close anatomical relation-
ship has been described for cholinergic and adrenergic nerves in airway
smooth rnuscìe, and it is conceivable that interaction between these
nerves could occur. Thus the ACh released by histamine stimulation of
cholinergíc nerves (flg.41) may promote norepinephrine release from
nearby adrenergic neurons and serve to exp'lain the phentolamine sensitiv-
ity of a histamÍne-mediated contraction (fig. 22). These studies índicate
a significant release of [3H]-norepinephrÍne stimulated by exogenously
applied acetylcholine. The norepinephrine thus released may bind to
postiunctional a'lpha, or beta2 receptors which could promote contrac-
tile or relaxant responses depending on the actíve tone of the muscle and
concentration of norepinephrine at receptor sites. Another possibility
Ís that any NË released rnay furbher promote ACh release forming a posi-
tive feedback 1oop. 0thers have suggested that as I'lE levels increase in
the space around the nerve they may inhibít ACh release and promote some
reciprocal inhibítion via this nrechanism fn canine trachealis (Vermiere
and Vanhoutte, L979l.. Our studies examined the actions of norepinephrine
at two concentrations (tC)_7 and lÉM) on ACh overfl ow. Norepi ne-
phri ne (1r7M) produced a smal I enhancement of the rel ease of 14C-
ACh overf'low, in the presence of eserÍne (Fig. 39,43). At, a hÍgher con-
centraLion, Nt (1ÉM) inhíbited choline overflow (ftg" 39). Additíon
of eserine dfd not qualitatively alter this effect (Fig. 44 upper panel)
but the further addÍtion of TTX resulted in a significant depression of
[14C]-acety'lcho'l i ne ef fl ux (f i g. 44 I ower panel ) . l,le have no expl an-
ation for this action of I'TX since the latter is present throughout the

-140-
experiment. The action of TTX on a lower concentration of NE was not
investigated. An attempt to release [3H]-norepinephrine with tyramine
was unsuccessful, likely due to the presence of cocaine which may inhibit
the uptake of tyramine (Fig. 43, lower pane'l). Therefore the effect of
tyramine (in the presence of TTX) on acetylcholine release measured
dÍrect effects of tyramine on cholinergic nerves rather than the indirect
effects produced by the release of norepinephrine. It was expected that
the release of endogenous norepinephrine rnight have a neuromoclulating
effect on acetylcholine release. The studies however do support a stimu-
latory ef'Fect of lower concentration and an inhibitory action of higher
concentrations (Ne lÉM) of norepinephrine on [14C]-acetylcholine
overflow. When propranolol was added an inhibition of ACh efflux was
observed (Fig.45 lower panel) suggesting that if cholinergic nerves have
both alpha- and beta-adrenoceptors prejunctional'ly it is ìikely that
alpha-stímulation inhibits ACh overflow at resting level s. Phentoìamitte,
which blocks alpha-adrenoceptors pre and postjunctionally should unmask
prejunctional beta'receptors and thus enhance acetylcholine overflow in
response to locaì'ly released norepinephrine or circulating catechola-
mines. The present studies dicl not demonstrate an effect of phentolamine
in the presence of TTX and eserine (Fig" a5). The results of figure 45
may be explained in tenns of a lack of endogenous norepÍnephrine reaching
cholÍnergic nerves when the preparation is unstimulated. Aìthough a low
basal efflux of norepinephrine is likely, as suggested by the steady
efflux of radioactivity in the present experirnents, this low level of
norepinephrine may be insufficient or too distant to act on cholinergic
nerves. Propranol ol may depress 14C-acetyl chol i ne overfl ow by a
nonspecific anesthetic action on nerves"

-141-These studies, including the actions of hístamine pre-iunctionally
on the chol i nergic system suggest that 1 ) hí stami ne increases basal
efflux of acetylcholine from cholinergic nerves by actÍng at a site on
the postgang'lionic efferent to the smooth muscle in canÍne trachea and 2)
acetylcholine increases the basal efflux of norepinephrine from postgang-
lionic adrenergic nerves and it is by this action that histamine contrac-
tions show partia'l inhibition by phentolamine. When an atropinized muscle
was stimulated with histamine the resulting tension was not signÍ'ticant'ly
altered by phentolamine, suggesting that the ACh action to pronote NE
efflux occurs at a muscarinic site which is blocked by atropine (fig.
23). When phentoìamine was added at the p'lateau of a histamine induced
contract,ion prior to atropine treatment a significant inhÌbition of iso-
metric tensÍon was observed (fig. 22). Histamine (lÉM) was also shown
to produce a further contraction in a fu'lly depo'larized muscle (127mM
K+) which had been pretreated with atropine, phentoìamine and proprano-
lol. This pharmacomechanícal coupling dernonstrates a direct action of
histamine on canine TSM, and has been shown to be mediated by HL,
receptors ( Antoni ssen et. al . , 1980 ).
A histamine-induced contraction is therefore cønposed of three
separate entities;1) postjunctÍonal H1 receptors on the airway smooth
muscl e cel I s 2l a stÍmul ation of acetyl chol i ne effl ux which has
postjunctíonal muscarinic effects and 3) an enhancement of Nt release (by
hÍstamÍne stimulated ACh overflow) which mediates contraction by alpha-
adrenoceptor stimulation" The latter Ís further supported by the finding
that phentolamine significantly reduced isometric tension developed by a
low concentration (lf€N) of ACh (Table B).

- t42
The action of 5-HT on caníne TSM is to induce contraction whích is
partly ( L0%) atropíne-sensitive (fig. 24). Shel ler et. âl ., (1982)
noted an enhancernent of vaga'l'ly stimul ated contractions in the presence
of 5-HT in canine airways Ín vivo and suggested an interaction between
5-llT and efferent cholínergÍc fibers to airway srnooth muscle. The Íso-
metric contractile response followÍng 5-HT was increased dramatically by
eserine, following which proprano'lo1 did not further enhance the contrac-
tion (Table 4). In fact all muscles showed a reduction Ín tension
fol 1 owi ng propranol oì admi ní stration a'l though the response r,las not
statistically significant. These results suggest a possible enhancing
actÍon of 5-HT on acetylcholine release frorn unstimulated (i.e. electri-
cal ly) nerves. Serotonin, however, v,/as wÍ thout signí'ticant ef fect on
14C-acetylcholÍne overflow normal'ly (nig. 38), in the presence of
eserine (Fig.41) or TTX (Fi9.41, lower pane'l). The finding that eserine
enhanced 5-HT-stimulated contractions rnay be due to a non-specific action
of eserine on the muscle in addition to íts inhibitíon of acetylcholine-
sterase. Table 7 documents an íncrease of tension development following
eserine admÍnistration in muscles already contracted by ACh. The obser-
vation that hyoscyamÍne eliminated the entire contraction argues strongly
against a nonspecific contractile ef'fect of eserine on tracheal smooth
muscle. Perhaps the inhibitory effect of atropine on 5-HT induced con-
tractions was due to an indirect effect through a S-HT-stimulated release
of norepinephrine acting on cholinergic nerves to release ACh. Thís would
be ana'lagous to the situation suggested prevÍous1y for the actÍon of
histamine on the chol inergic system except in reverse. The potent
inhibitory action of phentolamine on 5-hlT induced isometrÍc tension

-143-responses (fig. 25l. suggested a stimulatory action of S-HT on basal NE
release. However 5-HT was found to be without significant effect on
t3Hl-norepinephrine overflow. It Ís therefore concluded that phentola-
mine and perhaps atropÍne have nonspecific Ínhibitory actions on 5-HT
induced contractions perhaps by interferring with the binding of S-HT to
specific serotonergic receptors on tracheal smooth muscle. It is also
possible that,5-HT, which is somewhat sirnilar Ín structure to norepíne-
phrÍne, ffiâY interact wi th postjunctional a1 pha receptors which woul d
explain the potent inhibitory action of phentolamÍne on 5-HT contrac-
tions. The inhibitory actions of phentolamine as an antiserotonergÍc
compound have been demonstrated to occur in other systems (Garattini and
Samanin, 1978).
These studÍes are not inconsistent with an effect of 5-HT on neuro-
transmitter overflow since only non-stimulated nerve preparations were
studÍed. Ear'ly studies have demonstrated an excitatory effect of 5-HT on
smooth lnuscle directly and via Íncreasing the activi'üy of intramural
ganglion cells in isolated guinea pig ileum (Gaddum and PicarellÍ,19b7).
An excitatory actÍon of 5-HT is observed for gang'lion cells which have an
inhibÍtory action in the guinea pig stornach (eülUrtng and Gershon, 1968).
The present studies do not permit a predict,ion of the action of 5-HT on
the neural components of airway smooth muscle when action potentia'l fre-
quency is increased. Serotonin may enhance neurotransmitter release as
Ín the cholinergic nerves of the smalI intestine (Adarn-Vizi and Yizi,L97B) or inhibit rel ease as for adrenergic nerves to blood vessel s
( McGrath , L977 I .
The lack of a demonstrable increase of either t3Hl-norepinephrine

-144-or tl4Cl acetyl chol ine ef'fl ux fol I owi ng treatment wi th s-HT may be
due to the small amount of labelled transmitter in the nerve varicosity
in relatÍon to the unlabelled poo'l. Such an effect would decrease the
sensítivity of the technique and allow the demonstration of increased
transmitter overflow on'ly for those substances havíng a strong stimula-
tory action on nerves such as electrícal field stimulation or potassium
Índuced depolarization. The effects of these two stímulÍ were studied
and did indeed dramatical'ly increase t3Hl- norepi nephrine overfl ow
(Figs. 29 and 30) and tl4Cl- acety'lcholÍne overflow (figs " 37 and
3B), the latter in the absence of eserine. Histamine however, díd not
measureably stÍmul ate [14C]- acetylchol ine effl ux (fig. 37) in the
absence of eserine but when eserine r^ras added a significant stimulation
of efflux was ol¡served (nig. 40, upper panel). Although the efflux data
are not useful 'tor accurate quanti tative rneasurements of translnÍ tter
release, it is like1y that histamine stimulates a greater acetylcholine
release than does s-HT as iudged by the greater effect of both atropine
and eserílle on histamine Índuced contractions. Since hÍstamine stimulated
effl ux of [14C]-acetyl cho] ine was weak rel ative to el ectrical fiel d
stimulation or ç+ st,imulatetl efflux, it is not, surprising that the
effect of 5-HT was not found to be sígnificant.
The release of acetylcholÍne stímulated by hîgh-K+ is supported by
an increase of K+-Índuced contraction in the presence of eserine (Table
9) and a retluction in tension developed Ín the presence of atropine (Fig.
6). A reduction of tension, developed upon exposure to high-K+ and
eserine talas observed in the presence of phentolamine (Tabls 9), support-
i ng the rel evance of K+-stimul ated [3H]-norepinephrine overfl ow.

-145-Si nce the high K sol ution al so increased [14C]- acetyl chol ine over-
flow it is like1y that the ACh released also enhanced [3H]-norepine-
phrine efflux in addition to the direct effect of 6+ on adrenergÍc
nerves. The effect of propranolol on isornetrÍc tension developed Ín
response to potassium and eserine was negligibìe, probably due to the
high level of tone produced fol lowing eserine (table 10) (no'Ee our
finding above that the unmasked alpha-adrenoceptor response is very small
when tone is high, Table 1).
Isometric tensÍon deve'loped following exposure to 5-HT or histamíne
was similar when comparíng TSM froln 0A sensÍtized dogs to mongrel con-
trols. The effects of the antagonists phentolamÍne and a'fropÍne on these
i sometrÍc responses were al so sÍmi'lar (Figs. 26 and 27 , -lable 3 ). The
grea'ber transÍent inhÍbition by phentolamine of a histamine-induced con-
traction in sensitized tissue ís not explained (fig. 26). These studies
suggest that release of the neurotransmitters norepinephrine and acetyl-
choline by histarnine and 5-HT are similar to that of control tissues and
therefore labelled transmitter efflux studies on 0A sensitized TSM were
not carried out.
E) Autoregulation of TransmÍtter Overflow
Previous studies have suggested that prejunctÍonal receptors on both
adrenergic and cholinergïc nerves may regulate or modulate transmitter
rel ease (westfa'|1 , L977; Langer , L977; Ì,,leiner, LgTg). It has been shown
that adrenergic nerves possess prejunctiona'l alpha2-adrenoceptors which
when stímul ated inhÍbit stimul ated norepinephrine overfl ow and when
blocked by phentolamine or phenoxybenzamine enhance electríca1ly stimu-

-146-I ated norepi nephrine re] ease (Langer , L977). Dixon et. âl ., ( 1979)
suggested a feedback ÍnhibítÍon of norepinephrine on its olvn further
rel ease which was dependent on norepi nephrine concentratÍon in the
iunctional cleft and therefore on the action potentia'l frequency. At low
concentratÍons ( not known, but 1íkely th7¡4 or less) norepi nephrine may
enhance its release, an effect b'locked by propranolol (Langer, L9771
whil e at hÍgher concentrations i nhibi tÍon of norepi nephri ne effl ux
0ccurs.
The present studies have shown that for canine TSM phento'lamine
enhanced t3Hl-norepi nephrine overf'low and norepinephrine (1ÉM) al so
enhanced overflow (Figs. 31 and 32). }Jhile further studies vvere not
carrÍed out we conclude that aìpha-adrenoceptor blockade enhances nore-
pinephrine release as in other tissues even in the absence of eìectrical
stimulation. The action of norepinephrine to increase basal norepíne-
phrine release may occur by stimulation of prejunctional beta-adrenocep-
tors alt,hough the evidence at present is prelimÍnary. The fact, that
neuromodulation occurs ín adrenergÌc nerves in canine trachealis was pre-
viously not known and our present results serve as a starting point for
further investigatÍons designed to characteríze the receptor set
i nvol ved.
The present studies urere unable to dernonstrate an autoregulatory
effect of acetylcholine on basal tl4Cl- acetyìcholine efflux. The
addítion of atropine did not alter the response (fig" 421. Since both
experiments were carried out Ín the presence of TTX it, is possible that
acetyl chol í ne feedback on further acetyì chol i ne rel ease may requi re
action potential discharge.
All of the experiments on radioactive acety'lcholÍne or norepine-

-147-phríne efflux were intended to rleternrine whether several factors, especi-
ally hÍstarnÍne and 5-l{T, had effects 0n the nerve network in canine
trachealis at normal resting tone. Previous studies on other tÍssueshave exami ned the effects of these substances on el ec'bríca'l 'ly-or K+-
stÍmulated transmitter overflow. Further studies of thÍs nature are also
requÍred for canÍne and human airways. Changes in transrnitter release by
local factors (neurotranstnitters, prostaglandins, leukotrienes, pH,
hypoxia, etc.) nerve interactÍon, circulating substances (S-HT, catecho-
lamines) or therapeutíc agents may all influence the contractile state ofairway smooth muscle. 0n'ly when the nature of these interactions have
been elucÍdated can changes whÍch may occur in diseases such as asthina be
interpreted with confidence.

-148-REFERENCES
Adam-VÍzÍ, V. and Vizi, E.S. Direct evidence of acetylcholine reìeasingeffect of serotonin in the Auerbach plexus. J. Neural Transm.,4?:L27-t38, 1978.
Adcock, J.J. and Garland, L.G. A possible role for ìipoxygenase productsas regulators of airway smooth muscle reactivity. Br. J. Pharmac.69: 167-169,1980.
Adolphson, R.S., Abern, S.B. and Townley, R"G. Human and guinea pig'respiratory smooth muscle - DemonstratÍon of alpha adrenergic recep-
tors. J . A1 1 ergY 47: 110-1 LL,t97L.
Akasaka, K., Konno, K.Qno, Y., Mue, S., Abe, C., Kumagai, M. and Ise' T.A new electrode for e'lectromyographic study of bronchial smoothmuscle. Tohoku. J. exp. Med.117(1): 49-54,1975.
Akasaka, K" Konno, K.Ono, Y. Mue, S., Abe, C., Kumagair-Electromyographic study of bronchial smooth musclasthma. tóhoku ¡. Exp. Med. LL7: 55-59 ' L975.
M. and Ise, T.e in bronchial
Akcasu, A. The physiologic and phannacologic characteristicstracheal muscle. Arch. Int. Pharmacodyn. Ther. t22:1 959.
of the2QL-207,
.J. , Pol son, J.B" and SzentivanYi , A.s of tachyphylaxis to hístamine in canine
Adv. in Prostag'landin and Thromboxane
Anderson, W.H., Krzanowski, JProst,agl andi ns as medi atortracheal smooth muscle.Research 7: 995-1001, 1980.
Anderson, R., Nilsson, K., hlikberg, J. Johansson, S., Mohme-Lundholm' E"
and-Lundholm, L" Cyclic nucleotÍdes and the contraction of smooth
muscle. In Àdvances in Cyclic Nucleotide Research, Vol. 5., Editedby G.E. Drummond, P" Greengard and G.A. Robinson. New York, Raven
Press, 491-518, L975"
Antonissen, L.A., Mitchell, R.W", Kroeger, E.4., Kepron, |l|., Tse, K'S"and Stephené, N.L. Mêchanical alterat,íons of airway. smooth muscle
in a canÍne asthmatic model. J. Appl. Physiol.46(4): 681-687't979.
Antonissen, 1.A., Mitchell, R.lll", Kroeger, E"4., Kepron, W', Stephens'N.L. ánd AeÉgefl, J. Éistamine phairnaco'logy in_airway smo_oth musclef rom a cani ñe ino<tel of asthma " J . Ph armacol . Exp . Ther. 2L3z
150-155, 1980.
Austen, K.F and Orange, R.P. Bronchial asthma: The possible role of thecñemical mediatórs of immediate hypersensitÍvity Ín the pathogenesisof subacute chronic disease. Amer. Rev" Resp. Dis. LLZz 413,197 5"

-L49-
Barnes, P.J., Do'l1ery, C.T. and MacDermot, J. Increased pulmonarya1 pha-adrenergic and reduced beta-adrenergíc receptors inexperimenta'l asthma. Nature 285: 569, 1980.
Barnes, P., Fitzgeral d, G., Brown, M. and Dol'lery, C" Nocturnal asthmaand changes in circulating epínephrine, histamine and cortísol.Med. Intel 1 igence 303(5): 263-?67, 1980.
Beinfield, 1,l.H. and Seifter, J. Spontaneous mechanical activities of dogtracheal Ís muscle in vÍvo. J. Appl . Physiol.: Respirat. tnviron.Exerci se Physiol . T8f2T: 320-328, 1980.
Beinfield, W.H. and Seifter, J. Contraction of dog trachealis muscle invivo: role of alpha-adrenergic receptors. J. Appl. Physiol. 48(2):329-336,1980.
Bergen, J. and Kroeger, E.A. Responses of normal and Ímmunologíca'llysensitized canine tracheal smooth muscle to alpha adrenoceptor acti-vation. Proc. Can. Fed. Biol. Soc.22z L75, 1979.
Bergen, J"M. and Kroeger, E.A. Isometric tension responses to norepine-phrine in canine tracheal smooth muscle (Abstr). Fed, Proc, 39(3):LL75,1980.
Bernstein, I.L. Cromolyn sodÍum in the treatment of asthma: changingconcepts. J. Allergy Clin. Immunol. 68: 247-53, 1981.
Bi as, l,l. The genetic basÍ s of asthma In: Asthma: Phsyi o'logy,Immunopharmacology and Treatment, ed. by LichtensteÍn, L. and Austen,K.F., Acad. Press. 1973"
Bolton, T.B. MechanÍsms of action of transmÍtters and other substanceson smooth muscle. Physiol. Rev. 59(3): 606-718, L979.
Bolton, T.B" CholinergÍc mechanisms in smooth muscle. British MedicalBulletin 35(3): 275-283, 1979.
Born, G.V.R. S-Hydroxytryptamine receptors. In: Smooth Muscle, edÍtorsBu'lbring, E., Brading, 4.F., Jones, A.W" and Tomita, T.; Arnold,London,418-450,1970.
Bosen R" and Bose, D. Excitation - contraction couplingtracheal smooth muscle during metabolÍc dep'letion:rhythrnÍcity. Amer. J" Physíol " 233: C8-13, t977.
in multi unitinduction of
Boushey, H.A. Neural mechanisrns in asthma. In, Brain, Behaviour, andBodily Disease, editor l^leiner, H" Hofer, M.A. and Stunkard, 4.J.,Raven Press, New York, 27-44, 1981.
Boushey, H"A. Richardson, P.S" and l^liddicombe, J.G. Reflex effects oflaryngeal Írritation on the pattern of breathing and total 'lung
resistance. J. PhysÍo'l . (London), 224: 501-513, 1'97?.

-150-Boushey, H.4., Holtzman, M.J. Sheller, J.R. and Nadel, J.A. Bronchial
hyperreactÍvity. Amer. Rev. Respir. Dis. tZLl 389-413, 1980.
Bozl er, E. Conduction, automaticÍty and tonus of vi sceral muscles.Experientia 4: 2L3-2t8, 1948"
Brink, C., Grimaud, C. Guil 1ot, C. and Orehek, J. The interactionbetween indomethacin and contractÍle agents on human isolated airwaymuscle. Br. J. Pharmac. 69: 383-388, 1980.
Brown, J.K., Leff , 4.R., Frey, M.J., Reed, B.R. andPhysio'logical and pharmacologÍcal properties of caninemuscle in vivo" J. Appl. Physiol. 49(1): 84-94, 1980.
Go'ld, l,'l.M.tracheal i s
tn6A:
g33,
the va323-3
al
Brown, J.K., Shields, R. and Go'ld, l,l.M. Parasympathetic Ínnervation ofthe cervical trachealis muscle in 1ivÍng dogs. J. Appl Physiol.53(3): 6L7-625, 1982.
Burn, J"H. and Rand, M.J. Acetylcholine in adrenergic transmission.Ann. Rev. Pharmacol. 5: 163-182, 1965.
Burnstock, G. Structure of smooth muscle and its innervation. In SmoothMuscle. Edited by E. Bulbring, A.F. BradÍng, A.N" Jones and T.TomÍta" London, Edward Arnold, L-69, L970"
Burnstock, G. PurÍnergÍc nerves. Pharmacol. Rev. 24: 509-581, L972.
Brítish MedicalBurnstock, G. Autonomic innervation and transmission.Bulletin 35(3): 255-262, 1979.
Bu'lbring, E. and Gershon, M.D. Serotonin participationÍnhíbitory pathway to the stomach. Adv. Pharmacol.1968"
Busse, W.W., Bush, R"K" and Cooper, lrl. Granulocyte response in vitro toisoproterenol, histamine, and prostaglandin E, during trdãTmãñEîithbeta-adrenergic aerosols in asthma" Amer. Rev. Respir. Dis" L20:"377-384, L979.
Cabezas, G.4., Graf, P.D. and Nadel, J.A. Sympathetic versus parasym-pathetic nervous regulation of aÍrways ín dogs, J. Appl. Physiol.31(5): 651-655, t971"
Cambridge, D. , Davey, M.J. and Massingham, R"
antagonist of post-synaptic adrenoceptors"(Jan.), 514P, L977.
Prazosin, a selectiveProceed. of B.P.S.
Cameron, A.R. and Kirkpatrick, C.T. A study of excítatory neuromusculartransmition in the bovine trachea. J. Physíol. (Lond.) 2702733-45, L977.

-151-Car'lyle, R.F. The mode of action of neostígmine and physostÍgmine on the
guinea pig trachealÍs muscle. Br. J. Pharmacol. 2Iz 137'L49,1 963.
Castro de 1a Mata, R., Penna, M. and Aviado, D"M. Reversal ofsympathomimetic bronchodi 1 atÍon by dichl oroi soproterenol . J .Pharmacol. Expt. Therap. 1352 L97-203, L962.
Cavero0P
, I., Lefevre, F. and Roach, A.G. Differential effects of prazosinn the pre-and. postynaptic alpha-adrenoceptors in the rat and dog.roceed. of B.P.S. (Sept.), 469P, L977.
Cha
Cha
hd, N. Deroth, L. and Eyre, P. Pharmacology of Schultz-Dale reactionin canine lung strip in vitro: PossÍble npdel for allergic asthma.Br. J. Pharmac" 66: 511-516, L979.
hd, N., Dhawan,8.N., Srimal, R.C", Rahmani, N.H., Shukla, R.K" andAltura, B.M. Reactivity of trachea, bronchí and 'lung strips tohistamine and carbachoì in rhesus monkeys. J. Appl. Physiol. 49(4):729-734, 1980.
Chuang, D. and Costa, t. Evidence for internalization of the recognitÍonsite of beta-adrenergic receptors during receptor subsensitivityinduced by (-) - isoprotereno'|. Proc" Natl. Acad. Scí. USA 76(6lt3024-3028 , L979.
Church, M.K., Co1lier,dexamethasone andbronchoconstriction56-65, L97?.
H"0. and James, G.W. The i nhibÍ tion bydi sodi um cromoglycate of anaphy'l actic
in the rat. Br. J. Pharmac. Chemother.46z
Church, J. and Zsoter, T.T. Calcium antagonistic drugs. Mechanism ofaction. Can. J" Physiol. Pharmacol. 58: 254'264, 1980"
burnCo
E
, R. F. and Yamaguchi , T. Membraneindependent tension in the canine trachealxp. Ther. 20L: 276-284, 1977.
Coburn, R.F. Electromechanical coupl ing ín canineacetyl chol í ne contractÍons. Amer. J. Physiol .
1979.
potenti al -dependent andmuscle" J. Pharmacol .
tracheal is muscle:236(3): 177-184,
Coburn, R.F. and Tomita, T. Evidence for nonadrenergic inhÍbitory nervesin the guinea pig trachealÍs muscle. Amer. J. Physiol. 224:1072-1080, L973.
Colebatch, H.J"H. and Halmagyi, D"F.J. Effect of vagotomy and vaga'lstimulation on lung mechanics and circulation. J. Appl. Physiol.18: BBl-887, 1963.
Coleman, R.A. and Levy, G.P. Non-adrenergic inhibitory nervous pathwayin guinea pig trachea. Br. J. Pharmaco'l . 52: L67-L74, 1974-

-L52-
Coleridge, H.M. and Coleridge, J.C.G" Impulse activity in afferent vagalC-fibres wi th endi ngs i n the i ntrapulmonary aÍ rways of dogs.RespÍr. Physiol. 292 L25-L52, t977.
Conol'ly, M.E. Cyclic nucleotÍdes, beta receptors, and bronchial asthma.Adv. Cyclic Nucleotide Res. L2¡ 151-159, 1980.
Cookson, D.V. and Reed, C"E. A
isoproterenol in the normal andRespir. Dis. 88: 636-643, 1963.
compari son of theasthmatic subject.
effects ofAmer. Rev.
Cropp, G.J.A. The role of the parasympathetic system in the maintenanceof chronic airway obstruction Ín asthmatic children. Amer. Rev.Respir. Dis. lLZz 599-605, 1975.
Curry, J.J. Comparative action of acety'l-beta-methyl-choline and hista-mine on the respÍratory tract in normals, patients with hay fever,and subjects wÍth bronchial asthma" J. Clin" Invest. 26: 430-438,t947.
Dahlen, S.E., Hedquist, P. Hammarstrom, S. and Samuelsson, B. Leuko-tiíenes are potent constrictors of human bronchi. Nature 288(4):484-486, L980.
Daly, M. DeBurgh and Mount, L.E. The origin, course and nature of bron-chomotor fibers in the cervical sympathetic nerve of the cat. J.Physiol. 113: 43-62, 1951.
Daniel, E.E., Davis, C., Jones, T. and Kannan, M.S. Control of airwaysmooth muscle, In, Airway Reactivity: mechanisms and clinÍcalrelevance. EdÍtor F.E. Hargreave, Astra Pulmonary, Canada, 80-107,1 980.
Dea1, E.C., McFadden, JF. E.R., Ingram Jr. R.H., and Jaeger, J.J.Effects of atropine on potentiatÍon of exercÍse - induced broncho-spasm by cold air. J. Appl. Physiol. 45(2): 238'243, 1978"
DeKock, M.4., Nade1, J"4., Svi, S. Colebatch, H.J"H. and 0lsen, C"R. New
method for perfusÍng bronchial arteries: Histamine bronchoconstric-tion and apnea. J. Appl. Physiol. 21: 185-194, 1966.
Diamond, J. Role of cyclic nucleotides in control of smooth muscle con-traction" Adv. In" Cyclic Necleotide Research. 9: 327-340, 1978.

-153-
Dixon, M., Jackson, D.M. and Richards, I.M. The effect of a respiratorytract infection on histamÍne-induced changes in lung mechanics andirrÍtant receptor discharge in dogs" Amer. Rev. Respir. Dis" L20z843-848, L979.
Dixon, M., Jackson, D.M. and Richards, I.M. The effects of 5-hydroxy-tryptamine, histamine and acetylcholine on the reactivity 0l !!elung of 'bhe anesthetized dog. J. PhysÍol. London. 307: 85-96,1 980.
DÍxon, W.R., Mosimann, lll.F. and Weiner, N. The role of presynaptic feed-back mechanÍsms Ín regulatÍon of norepinephrine release by nervestimulation. J. Pharm. Exp. Ther. 209: L96-204, L979-
Douglas, J.S., Dennis, M.!'l", Ridgway, P. and Bouhays, A.
strÍctÍon in guinea pigs: interaction of histaminedrugs" J. Phannacol. Exp. Ther. 184: L69'L79, 1973.
tmpey, D.W., LaÍtinen, L"A.Mechanisms of bronchíalupper respiratory tract113: 131-139,1976.
Jacobs, L., Go1d,hyperreactivity ininfection. Amer.
Airway con-and autonomic
t,l.M. and Nadel , J.A.novmal subjects after
Rev. Respir. Diseases,
Doxey, J.C., Smith, C"F.C,, l^lallcer, J.M. Selectívity of blocking agents-for pre-and postsynaptic alpha-adrenoceptors. Br. J. Phannac. 60:91-96, L977.
Drazen, J.M. and Austen, K.F. Atropine modification of the pu'lmonaryeffects of chemical mediators in the guinea pig. J. Appl. Physiol.38(5): 834-838, 1975.
Drew, G.M. Pharmacologica'l characterizatÍon of presynaptic alpha-adreno-ceptors which regulate cholinergic activity in the guinea-pig i1eum.Proceed. of B.P.S. (Jan.), 513P, L977.
Dubey, M.P., Muscho11, E. and Pfeiffer, A. Muscarinic inhibition of-potassium-induced noradrenaline release and its dependence on thebalcium concentration" Nauyn-Schmiedebergs. Arch. Pharmacol . 291:1-15, L975.
Dubois, A.B. and Dautrebande, L. Acute effects 0f breathíng inert dustparticles and of carbachol aerosol on the mechanical characteristicsbt tfre lungs in man: changes in response after inha'ling sympatho-mímetic aeiosols. J. Clin. Invest. 372 L746-55' 1958.
Duckles, S.P. and Raport, R. Release of endogenous norepinephrine frorn ara6Ui t cerebral' artery. J . Pharmacol . Exp. Ther. ZLLz 2L9'?.24 ,L979.

Ene
-154-t^0, M.A. and Langer, S.Z. Inhibition by dopamine of 3H-noradrena-
lÍne release elicited by nerve stimulation in the isolated cat'snicitating membrane. Naunyn-Schrniedeberg's Arch. Phar¡nac " 289:L79-203,1975"
Everitt, B.J. and CaÍrncross, K.D. Adrenergic receptors in thegu'inea-pig trachea. J. Pharm. Pharmac . 2I: 97'L0?, 1969.
Eyre, p, The Pharmaco'logy of sheep tracheobronchial muscle: A relaxanteffect of histamÍne on the isolated bronchí. Br. J. Pharmacol. 36:409-4L7,1969.
Eyre, P. Histamine H2-receptors in the sheep bronchus and cat trachea:- the action of bur-ímamide. Br. J" Pharmac 48: 32I'323, L973.
Eyre, P. Pulmonary hístamíne H1 and H2 receptor studies. In AsthmaII: PhysÍology, Immunopharmacology and Treatment. ed. by K.F.Austen and L. Lichtenstein. p" 164, AcademÍc Press, Nelv York,L977 .
Feniuk, W., Humphrey, P.P.A. and Watts, A.D. Presynaptíc inhibítoryaction of 5-hydroxytryptamine ín dog Ísolated saphenous vein. Br.J. Pharmac . 67: 247-254, L979.
Ferrendel I i , J.A. , Ki nscherf, D.A. and Chang, M.M.of guanosine cyclic. 3'5'- monophosphatesystem: Effects of depol oarizi ng agents"445-454, L973.
Regu'l ati on of I evel sin the central nervous
Mol. Pharmacol , 9z
smooth muscle. InEdÍted by J.A. Nadel.
r exi stence ofhea Amer. J.
Fillenz, M. Innervation of pu'lmonary and bronchial blood vessels of thedog. J" Anat. 106: 449, 1970"
Fteisch, J.H. and Calkins, P.J. Comparison of drug-induced. responses _ofra6bit trachea and bronchus. J. Appl. Physiol. 41(1): 62'66,L97 6"
Fl ei sh, J . H. Pharmacol ogi c aspects of ai rwayPhysiology and Pharmacology of 'the Airways"Marcel Dekker, Inc. New York, l9I'2L6, 1980.
Fleisch, J.H., Maìong, H"M. and Brodie, B"B. EvÍdence foal pha-adrenergic receptors i n the mammal i an tracPhys i o'l . 2L8(21; 596-599 ' 1970.
Foldeso A. and Ha11, R.C" The effects of hÍstamine on responses of theral¡bi t ear artery to el ectrical stimul ation and to exogenousnoradrenal Í ne. Br. J . Pharmac . 67 z 329-335, L979.
Folkow, B. The hemodynamic consequences of adaptive structural changesof the resistance vessels in hypertension" Clin. Sci. 41: L't?"T97L.

-155-Foster, R.W. The nature of the adrenergic receptors of the trachea of
the guinea-pig. J. Pharm. Pharmac. 181 L-L?, 1966.
Foster, R.l,l. and 0'Donnel, S.R. Evidence that adrenergic nerves areresponsible for the actÍve uptake of noradrenalíne in the guínea-pigÍ sol ated trachea. Br. J " Pharmacol . 53: 109 , t975"
Fozard, J.R. and Muscho'11, E. Effects of several muscarinic agonists oncardi ac performance and the rel ease of noradrenal i ne fromsympathetic nerves of the perfused rabbit heart. Br. J. Phannacol.45: 61,6-629, L972.
Fozard, J.R. and Mwal uko, G.M.P. Mechani sm of the indÍ rectsympathomimetic effect of 5-hydroxytryptamine on the ísolated heartof the rabbit. Brit. J, Pharntacol. 57: L15-125, 1976.
Frigo, G.M., del Tacca, M., Lecchini, S. and Crema, A. Some observationson the intrinsic nervous mechanism ín Hirshsprung's disease. Gut1,4: 35-40, L973.
Gaddum, J.H. and Picarelli, 7.P. Two kinds of tryptamine receptor. Br.J. Pharmacol. L2: 323-328, L957.
Gandevia, B. Historical reveiw of the use of parasympatholytic agents inthe treatment of respiratory disorders. Postgraduate Med. J.51(7 ): 13-20 , L975.
GarattÍní, S. and Samanin, R. Drugs affecting serotonin: a survey. Irì,Physiological Regulation and Pharmacological Action. Vol. 2,SerotonÍn in Health and Disease. Editor Essman, W.8., SpectrumPub'l ÍcatÍons, Inc. , New York " 247 -293 , L978.
Gardinern P.J. and Collier, H.0"J. Receptors for E and F prostaglandinsin airways. Adv. Prostaglandin and Thromboxane Res. 7: L003-L008,1980.
Go1d, W.M. The role of the parasympathetic nervous system Ín airwaysdisease. Postgraduate Med. J. 51(Supp1 . 7lz 53'62, 1975.
Go1d, W.M., Kessler, G.F. and Yü, D.Y.C" Role of vagus nerves inexperimental asthma ín allergíc dogs. J" Appl. Physiol. 33(6):7L9-7?5, L972.
Go1d, W.M., Meyers, G.1., Dain, D"S. Mil ler' R.L. and Bourne, H"R.Changes in airway mast ce1ls and histamine caused by antigen aerosolin al lergic dogs" J. Appl . Physio'l . 43(3) z ?-7L-275, L977.
Golden, J.4., Nadel , J.A. and Boushey, H.A. Bronchial hyperirrítabi1 ityin hea'lthy subiects after exposure to ozone. Am. Rev" Respir. Dis.118: 287-?94, 1978.

-156,Goldstein,8.D., Pearson,8., Lodi, C. Buckìey, R.D. and Balchum, D.J.
The effect of ozone on mouse blood in vivo" Arch. Environ. Health.L6: 648-653, 1968.
Goldsmith, J.R. and Nadel, J.A. Experimental exposure of human subjectsto ozone. J. Air Pollution Control Assoc. 19: 329-330, 1969.
Green, M. and Widdicombe, J.G. The effects of ventilation of dogs withdifferent gas mixtures on airway calibre and lung mechanics. J.Physioì. (London), 186: 363-381, 1966.
Grieco, M.H. and Pierson, R.N. Jr. Mechanism of bronchoconstrictÍon dueto beta-adrenergic blockade. J. Al lergy Cl i n. Immunol . 48:L43-t52, t97L.
Gross, G.N., Souhrada, J.F. and Farr, R.S. The longterm treatment of anasthmatic patient using phenotolamine. Chest 66: 397-40L, L974.
HabÍb, M.P., Pare, P.D. and Engel, L"A. Variability of airway responsesto inhaled histamine in normal subjects. J. Appl. Physiol. 47(1):51-58 , 1,979.
Hahn, H.1., l,lilson, A.G. n Graft, P.D., Fischer, S. P. and Nadel , J.A.Interaction between serotonin and efferent, vagus nerves in doglungs. J. Appl. Physiol . 44:. L44, 1978.
Hajos, M.K. Clinical studies on the role of serotonin in bronchÍalasthma. Acta Allergy 17: 358-370, L962"
Hallman, H., Farnebo, 1., Hamberger, B. and Jonsson, G. A sensitivemethod for dtermi nation of pl asma catechol ami nes usi ng 1 i qui dchromatography with electrochemÍcal detection. Life Sci. 23:1049-1052, 1978"
Hattori, K., Kurahashi, K., Mori, J" and Shibata, S. The effect of coldstorage on the adrenergic mechanisms of intestinal smooth muscle"Br. J. Pharmacol. 46: 423-437, L972
Haynes, R"L., Ingram Jr. R.H. and McFadden, Jr. E.R. An assessment ofthe pulmonary response to exercise ín asthma and an analysis of thefactors influencing it. Am. Rev. Respírat,. Diseases 114:739-752, r976.
Hedquist, P. Further evidence that prostag'landÍns inhÍbit the release ofnoradrena'l i ne from adrenergic nerve termi nal s by restriction ofavailability of calciunr. Br. J" Pharmac. 58: 599-603, t976"
Hedquist, P. and Fredholm, B.B. Effects of adenosine on adrenergicneurotransmission prejunctiona'l inhÍbition and postiunctionalenhancement. Naunyn Schmi edeberg' s Arch . Pharmac . 293: 2L7 -?23,1976"

-L57-Henderson, W.R., Shelhamer, J.H., Reingo'ld, D.8., Smith, L.J., Evans, R.
and KaìÍner, M. Alpha adrenergic hyper-responsiveness in asthma.New Eng. J. Med. 300i 642, 1979.
HimorÍ, N. and Tai ra, N. Pharmacol ogÍca1 ana'lyi n muscul ature and vascul ature of the dogPharmacol . 64: 553-558, t978.
Hirshman, C.A. and Doulnes,i nfl uence of atropi neAppl. Physiol.50(4):
Hi rshrnan, C.A. , Ma1 'ley, A.
asthma: reactivity toJ. Appl . Physiol .:953-957, 1980.
sis of histamine receptorstrachea in situ. Br. J.
H. Basenii-Greyhound dog model of asthma:on antÌgen-induced bronchoconstriction" J.76t-765, 1981.
and Downes H. Basenii-Greyhound dog model ofascaris suum citric acid and methacholine.
i ron. Exerci se Physiol . 492sp ra nv
Hogg, J.C. Airways dimensions; changes in disease. Chest 80(6):789-79L,1981.
Hogg, J.C., Pare, P.D., Boucher, R.C. and Michoud, M.C" The pathoply!!9-logy of asthma" Canadian MedÍcal Assoc. Journal LZl: 409-414,t979.
Hol tzman, M.J. , Cunni ngham, J.H. , Shel 1er, J'R. , Irsigler, G.B. , Nadel 'J.A. and Boushey, H.A. Effect of ozone on bronchial reactivity in
atopic and non-atopic subiects. Amer. Rev. Respi r. Di s. 120:1059-1067, t979.
Hope, W., McCulloch, M.|lJ", Rand, M.J. and Story, D.F. Modulation of' noradrenergic transmission ín the rabbit ear artery by dopamÍne.Br. J. Pharmacol . 64 z 527'537 , 1978.
Hossain, S. and Heard, B.E. Hyperplasia of bronchial muscle in chronicbronchit,is. J. Pathol. 101: 171-184, 1970.
Hughes, J., Kosterlitz, H.lll" and Lesl ie, F.M. Effect of morphíne on- aãrenergic transmission in the mouse vas deferens. Assessment ofagonist and antagonist potencies of narcotic analgesÍcs. Br. J.Pharmac. 53: 371-381 , 1975.
Ishizaka, T. and Ishízaka, K" Bio'logy of immunoglobulin t. Prog.Allergy 19: 60'tZL, L975.
Iversen, L.L. Uptake mechanisms for neurotransmitter amines. Biochem.Pharmacol . 23t L927'L935, t974.
Jack, D" Select,Íve'ly acting beta-adrenoreceptor stirnulants in asthma"In Asthma: Physiology, Immunopharmaco'logy, and Treatment" Editedby K. F. Austen
- and L "-M. Li chtenstei n. New York , Academi c , 25L-266 ,
L973 "

-158-Jacobowi tz, D., Kent, K.M., Fleish, J.H. and Coope
study of catecholamÍne-containing elementsfrom the calf and the dog ìung' Proc. Soc.464-466, 1973.
r,in
E
T. Hi stofl uorescentcholÍnergíc ganglia
xp. Biol. Med. L44:
Kadowitz, P.J., Knight, D.S., Hibbs, R.G., Ellison, J.P., Joiner, P.dopam
in
D.,inethe
Brody, M.J" and Hyman, A.L. Influence of 5- and 6-hydroxyon adrenergic transmission and nerve terminal tnorphoìogycanine pulmonary vascular bed. Circ. Res. 39: 191, 1976.
Kaki uchi , S. , Ra'l 'l , T.l'1. and McIl wai n, H. The ef fect of el ectrical
stimulation upon the accumulation of adenosine-3'.5'-phosphate inisolated cerebral tissue. J. Neuochem. 16: 485-491,1969.
Kannan, M.S. and Daniel, E.E. FormatÍon of gap iunctíons by treatment invitro with potassium conductance bloc[eis" J. Cel 1. Biol. 7T33618, L978.
Karlin, J. The use of antihistamÍnes in asthma. Ann. Allergy 30:342-47, 1972.
Katz, B. and Miledi, R. Further study of the role of calcium in synaptictransmÍssion. J. Physiol. (Lond.l 207: 789-801, 1970.
Kay, 4.8., Stechschulte, D.J. and Austen, K.F. Eosinophi'l chemotacticfactor ín anaphylaxis. J. Exp. Med. 133: 602, L97L.
Kazimierczak, l^l. and Diamant, B. Mechanisms of histamine release inanaphylactic and anaphylactoid reactions. Prog. a1'lergy 242295-365, L978.
Kepron, W., James, J.M., Kirk, B. Sehon,4.H., and Tse, K.S. A canine- model for reagenic hypersensitivity and allergic bronchronstriction.J . A'l I ergy Am. Immunol . 59: 64-69 , 1977 .
Kessler, G"F., AustÍn, J.H.M., Graf, P.D., Gamsu, G" and Gold, I^l.M.
Airway constriction Ín experÍmental asthma in dogs: tantalumbronchographic studÍes. J. App1. Physiol.3S: 703-708' t973"
Killian, D. Cockcroft, D.W., Hargreave, F.E., and Dolovich, J" Factorsin allergen-induced asthma: relevance of the intensity of the air-ways ôl'lergic reaction and non-specific bronchial reactivity. Clin.A1 lergy. 6: 219-2?5, L976"
Kirkpatrick, C"T. Excitation and contractÍon in bovine tracheal smoothmuscle. J. Physiol. (Lond.) 244: 263'8L, 1975"
Kirpekar, S.M., Prat, J"C., and lllakade, A.R. Effect of calcium on there'l ationshi p between frequency of stimul atÍon and rel ease ofnoradrenal i ne frorn perfused spl een of the cat.Naunyn-Schmi edeberg' s Arch. Phannacol . 287 : 205-21'2, I975"

-159-Klock, L"E., Mi11er, T.D., Morrís, 4.H., L'latanabe, S. and Dickman, M. A
comparative study of i nhal ed atropi ne sul fate and i soproterenolhydrochloride in chronÍc bronchitis. Amer. Rev. Respir. Dis" LLZ:371-376, L975.
Kneussl, M.P. and Richardson, J.B" Alpha-adrenergic receptors in human
and canine tracheal and bronchÍal smooth muscle. J. Appl. Physiol.45(2): 307-311,1978.
Krell, R.D. and Chakrin, L.W. An in vÍtro model of canÍne immediate typehypersensitÍvity reactions" -T-nf,-Ãnchs. Al lergy Appl . Immun. 51:641-655, 1976.
Krel l, R.D. and ChakrÍn, L.l^1. Canine airway responses to acetylcholine,prostagl andi n F2a'l pha, hi stami ne and serotoni n after chronicantigen exposure. ¿. Allergy Clin" Immunol. 58: 664'675, 1976-
Krel I , R.D. and Chakrin, L.lll. Pharmacologic regulation of antigen-induced mediator from canine 1ung. Int. Archs. Allergy appl. Immun.56: 39-47, 1978.
Krell, R.D., Chakrin, L.W. and þlardell, J.R. Jr. The effect of choliner-gÍc agents on a canine model of a'llergic asthma. J. Allergy Clin.Immunol. 58: 19-30, 1976.
Kroeger, E. and Stephens, N.L. Effect of tetraethylammonium on tonic-aírway smooth muscle: initiation of phasic electrical activity.
Amer. J. Physíol . 228(21: 633-636, L975"
Laarson, S. and Svedemur, S. Bronchodilating effects and side effects ofbeta2 adrenoceptor stimul ants by di fferent modes ofadmiñístration. Amer. Rev. Respir. Di s. 116: 861-869 , 1977 .
Langer, S"Z. Presynaptic receptors and their 1e_gulation of transmitter- release. Br. J. Pharmac. 60: 4BL'497, 1'977.
Langer, S.2., Enero, M.4., Adler-GraschÍnsky, E., Dubocovich, M.L. and- Celuch, S"M. Presynaptic Regu'latory mechanisms for noradrenalinerelease by nerve stimulation. Proceeds. SymposÍum on CentralAction of Drugs in the Regulation of Blood Pressure. ed. Davies,D.S. and Reid; J"1., London: Pitman Medical, 133-151, L975"
Lee,1.Y., Bleecker, E.R. and Nadel, J"A. Effect of ozone on broncho-motor response to Ínhaled histarnine aerosol in dogs. J. Appl.Physiol. 432 626'63L, t977.
Leff, A.R. and Munoz, N.M. Cholinergíc and alpha adrenergic augmentationof histamine-Índuced contraction of canine aírway srnooth muscle" J.Pharmacol. Exp. Ther. 2l8z 582'587, 1981.

-160-Leff, 4.R", Munoz, N.M. and Schoulberg, N" Alpha-adrenergic contractile
properties of canine tracheal muscle in vítro. Fed. Proc. 41(4):1355 (Abstr.), 1982.
Leitch, A.G. Asthma-mechanisms and management. Clin" notes on Resp.Dis" zl(Ll: 3-9, t982.
Lewis, R.A. and Austen, K.F. Mediation of local homeostasis and inflama-tion by leukotrienes and other mast cel I dependant products.Nature, 293: 103-108, 1981.
Lichtenstein, L.M. and Gillespie, E. InhibitatÍon of histamÍne releaseis hÍstamine controlled by llZ receptor. Nature, 244: 287-?88,L973.
Lightbody, I.M. , Ingram, C.G. , Legge, J.S. and Johnston, R"N.Ipratropium bromide, salbutamol and prednisolone in bronchial asthmaand chronic bronchitís. Br. J. Dis. Chest 72: 181-186, L978.
Lindmar, R., Loffelholz, K. and Muscho'11, E. A muscarinic mechanisminhibit,ing the release of noradrenaline from perÍphera'l adrenergicnerve fibers by nicotÍnic agents. Br. J. Phamacol. Chemother. 322280-294, 1 968.
Long, J.P., Heintz, S., Cannon, J"G. and Kim, J. InhÍbition of the sym-pathetic nervous system by 5, 6-dihydroy-Z-dimethyl-amino tetralin(M-7), apomorphine and dopamine. J. Pharmacol. Exp" Ther. 192:336-342, L975.
Lorenz, R.R., Vanhoutte, P.M. and Shepherd, J.T. Interaction betweenneuronal amine uptake and prejunctional alpha-adrenergic receptoractivation in smooth muscle frorn canine blood vessels and spleen,Blood Vessels 16: 113-125, L979.
Loring, S"H. , Drazen, J.M., Snapper, J.R. and Ingram, R"H" , JF. Vaga'land aerosol histamÍne interactions on aÍrway response in dogs" J.Appl. Physiol. 452 40-44, 1978"
Lundholm, 1., Andersson, R" and Mohme - Lundholm, E" Role of cyclicnucleotides and calcium for smooth muscle function and netabolism.Pharmac. Rev., L976"
Mack'lem, P.T. and Engel , L.A. The physiological impl icatÍons of airwayssmooth muscle constrictÍon" Postgraduate Med. J. 51 (Suppl. 7)45-50, L975.
Macklem, P.T. and Mead, J" Factors detennining maxÍmum expiratory flowin dogs. J. Appl. Physío1. 25: 159, 1968"
Macklem, P.T., bloolcock, 4.J., Hoggn J.C., Nadel , J.A. and tdilson, N.J.Partitioning of pulmonary resistance in the dog. J. Appl. Physiol.26(6): 798-805, 1969.

Mann, S
1o3:
-161-
Ma'lo, P.E., LJasserman, M.A. and Griffin, R.L. Effects of intravenous and
aerosol nÍfedípÍne on prostaglandin F2alpha and histamine-inducedbronchoconstrÍction ín anesthetized dogs. J. Phannacol. Exp. Ther.22L: 410-415, L982.
Manber, L. and Gershon, M.D" A reciprocal adrenergic-cholinergicaxoaxonic synapse in the mammalían gut. Amer. J. Physio'1. 236(6):8738-8745, L979.
.P. The innervatÍon of mammalian bronchial smooth muscle: Thecalization of catecholamines and cholinesterases. Hístochem. J.
319-331, L97L"
Marshall, J.M. and Kroeger, E.A. Adrenergic influences on uterine smoothmuscle" PhÍ1. Trans. R. Soc. Land. B" 265: 135-148' 1973"
Mathe, 4.4., Astrom, A. and Persson, N.A. Some bronchoconstrictÍng and
brochodilating responses of human isolated bronchi: evidence forthe exi stence of a1 pha-adrenoceptors. J. Pharm. Pharmac . 23:905-910, L97L.
Mathe, 4.4., HedqvÍst, P. Effect of prostaglandins F2alpha and E2 onairway conductance in heal thy subjects and asthmatic patÌents.Amer. Rev. Respir. Dis. 111: 313-320,1975"
Mathe, A.A. , Hedqvi st, P. , Holmgren, A. and Svanborg, N. Bronchialhyperreactivity to prostagìandin F2alpha and hist,amÍne in patientswi th asthma. Br. Med. J. 1: 193-196 , L973.
McFadden, t.R., JF. An analysís of exercÍse as a stimulus for the pro-duction of airway obstruction. Lung 159: 3-LL, 1981.
McGrath, M.A. 5-Hydroxytryptamine and neurotransmitter release Ín caninebtood vessels. Cir. Res" 41: 428-435, L977-
McGrath, M.A. and Shepherd, J.T. Histamine and 5-hyroxytryptamineinhibitition of transmítter release mediated by H2' and
5-hydroxytrypt,amine receptors. Fed. Proc" 37z 195-198' 1978.
McNeÍ11, R.S. and Ingram, C.G. Effect of propranolo'l on ventilatoryfunction. Am. J. Cardiol. 18: 473-475, 1966'
McNeiì l, R.S., Nairn, J.R", Mi1 lar, J.S. and Ingram, C.G. Exerciseinduced asthma. Q.J. Med. 35: 55-67, 1966"
Mel I ow, A.M. Evidence of CaZ+ and Sr2+ i n transmi tter rel easefrom K+ depolarized nerve tenninals. Nature 282: 84-85, L979-

-L62-
Middendorf, }'l"F. and Russel'1, J"A. Innervation of aÍrway smooth musclein the baboon: evÍdence for a nonadrenergic ÍnhibÍtory system. J.Appl . Physio'1.: Respirat. Environ. ExercÍse Physiol. 48(6):947-956, 1980.
Mitchel1, H.l^I. Regu'latory mechanisms ín histamine-induced tone in swÍneisolated tracheal smooth muscle. Proceed. of B.P.S. 796P, 1981.
Morto'la, J., Sant'AmbrogÌo, G. and Clement, M.G. Local ization ofi rrÍ tant receptors i n the ai rways of the dog. RespÍ rationPhysiology 24: I07'LL4, 1975.
Nadel, J.A. Autonomic regulation of airway smooth muscle; Ín Physiologyand Pharmacology of the Airways, editor, J.A. Nadel, New York,2t7-257, 1980.
Nadel, J.A. Neurophysiologic aspects of asthma. In Asthma: PhysiologJImmunopharmacòlogy, and Treatment. Edited by K.F. Austen and L.M.LichtensteÍn. New York and London, Academic, 29-38, L973.
Nadel, J.4., Sa'lem, H., Tamplin, B. and Tokiwa. Mechanism of broncho-óonstriction during inhalation of sulfur dioxide" J. Appl.Physiol. 20: 165-167, 1965.
Nakanishi, H., Yoshida, H., Nakahata, N. and Suzuki, T. Effects ofprostagl andÍ ns on exci tatory transmi ssi on i n i sol ated cani ne
tractreãl muscle. Japan J. Pharmacol. 28: 883-889' 1978.
Narahashi, T. MechanÍsm of actíon of tetrodotoxin and saxitoxin on
excitable membranes. Fed. Proc. 31: LI24'1132' 1972.
than, R.4., Sega'll, N., Glover, G.C. and Schocket, A.L. The-effects ofHi and H2 antihístamines on histamine inhalation challenge-s__ina3thmatic -patients. Amer. Rev. Respir. Dis. L20z L25L-L258,L979 "
D" and Enero, M.A. Cocaine facilitates t,he effects of a'lpha-agon-sts and antagonists on adrenergic neurotransmi ssion, Gen.harmacol . L2: 255-259, 1981.
Na
roNig IiP
Norn, S., GeÍsler, A. Stahl Skova'l
'l ergÍ c hi stami ne rel ease.mast cel 1 s. Acta Al I ergol .
0ffermeier, J. and Ariens, E.J" Serotonin, I.action" Arch" Int. Pharmacodyn Ther. 164:
. and Klysner, R. Cyclicnfluence of methylxanthÍnes
183-191 , t977.
,PI
3?:
AMP
0nandrat
Norn, S. and Skov, P.S. PathophysiologÍc mechanisms in asthma.-cular and phârmacologíca1 aspects" Scand. J. Resp" Dis.
103) 25-32, L979.
Mol e-( Suppl .
Receptors involved in itsL92-2r5,1966.

0hno, Y., Watanabe, M. anda1 phaexci tatory responsepreparation rel ation25Lz 205-216, 1981.
Y. Mani festation of latentcani ne tracheal smooth muscl etone. Arch. i nt. Pharmacodyn.
-163-Kasuya,in the
to basal
0lsen, C"R., Colebatch, H.J.H., Mebel, P.E., Nadel, J.A. and Staub, N.C.Motor control of pulmonary airways studied by nerve s'timulation. J.Appl. Physiol. 202 ?02-208, 1965.
0lsen, C.R., Stevens, A.E. and McIlroy, M.Bbronchí durÍng muscular constriction"27-34, t967.
Ri gi dÍ tyJ. Appl .
of tracheaPhysiol.
and232
Orange, R.P. The formation and release of s'low-reacting substance ofanaphylaxis. Monogr. Allergy 12: 23L'240, 1977.
0range, R"P., Kaliner, M.4., Laraia, P.J. and Austen, K.F. Immunologicalrelease of hÍstamÍne and slow reacting substance from human 1ung.II. Influence of cellular levels of cyclic AMP. Fed. Proc.30:L725-L729, L97L.
Orehek, J., Gayrand, P., Grimaud, Ch. and Charpin, J.betaadrenergic blockade on bronchial sensitivityacetylcholine Ín normal subiects" J. Allergy Clin.164-169, r975.
Effect ofto i nhal ed
Immunol. 55:
tor30:
Qrehek, J., Gayrand, P., Smith,4.P., Grimaud, C. and Charpin, J. Airwayresponse to carbacol in normal and asthmatÍc subiects. Amer. Rev.Respir. DÍs. 115: 937-43, L977.
Pandya, K.H. Postnatal deveì opment changes i n adrenergic recepresponses of the dog trachea'l muscle. Arch. int. Pharmacodyn. 2
53-64, 1977.
Panzani, R. 5-Hydroxytryptamine (serotonin) in human bronchial asthma.Ann" A'llergy 20: 72L'732, L962.
Parker, C"D", Bilbo, R.E. and Reed, C.E. MethacholÍne aerosol as a testfor bronchial asthma. Arch. Intern. Med. 1152 45?'458, 1965"
Parker, C"l.l. and Smith, J.lrl. Alterations in cyclic adenosÍne monophos-phate metabolÍsm in human bronchial asthma" I. Leukocyte rqgpq!-siveness to beta-adrenergic agents" J" Clin. Invest. 522 48-59'1 973.
Patel, K.R. and Kerr, J.hJ. Alpha-receptor-blocking drug in bronchialasthma. Lancet L: 358-349 , 1.975"
Pat'Lerson, R. Laboratory model s of reaginic aì lergy. Prog. A1'lergy 13:322,1969.

Pepys, J. and Hutchcroft, B.J.díagnosis and analysis of829-859, L975.
provocation test,s in etÍologícAmer. Rev. Resp. Dis. LLZt
-164-Bronch i alasthma.
Permutt, S. Phys i o1 ogi ca'l changes i nAsthma. Edited by K.F. Austen a
Academic, L5-?7, 1973.
Pluchino, S. Direct and indirect effectstyrami ne on cat smooth muscles.Pharmacol . 2722 189-224,L972.
acute asthmatic attack. In.M. LichtensteÍn. New York,
eL
thnd
Pinckard, R.N., Halonen, M. and Meng, A.L. Preferential expression ofanti-bovine serum albumin IgE homocytotropic antibody synthesis and
anaphylactic sensitivity in the neonatal rabbit. J. Allergy Clin.Immunology 49: 301, L972.
Ploy-Song-Sang, V., Corbin, R"P. and Engel, L.A. Effects of intravenous- histaminé on lung mechanics Ín man after beta-blockade. J. Appl.Physiol. 442 690-695, L978"
of 5-hydroxytryptamine andNaunyn-Schrniedebergs Arch.
Popa, V. Bronchodil ating activity 9t -qn itt_!locker chl orpheniramine.' J . A1 lergy Cl i n. Immunol . 59: 54-63 , 1977 .
Powel1 , D.hl. and Tapper, E.J. Intestinal l ion t,ransport_:cñolinergicadrenergic interactions" In: MechanÍsms of IntestinalSecretíoñ, edíted by H.J. Binder, Kroc Foundation" New York: AlanHíss, L75-L92, L979.
Powell, J"R. Effects of hístamine on vascular sympathetic neuroeffectortransmission. J. Pharmacol. Exp. Ther.208: 360-365, L979.
Prosser, C.1., Burnstock, G. and Kahn, J. Conduction in smooth muscle:comparatt've structural properties" Amer. J. Physio'l . 199:545-55?., 1960"
Ribon, A. and Parikh, S. Drug-induced asthma: a revÍew" Annals ofA'llergy 44: 220'224' 1980.
Richardson, J.B. Nerve supp'ly to the'lungs" Amer. Rev. Respir. Dis"119: 785-802, 1979.
Richardsorìn J. and Beland, J" Nonadrenergíc inhíbít91V ne¡yous system inhuman aÍrways. J. Appl. Physiol. 41(5) z 764-77L' 1976"
Richardsorì, J. and Bouchard, T. Domonstration of a nonadrenergic inhibi-tory hervous system in the trachea of the guinea pig. J" A1'lergyClin" Immunol. 56: 473-480, L975"
Richardsoñ, J.B. and Ferguson, C"C. Neuromuscular structure and functíoni n the ai rways. Federation Proc. 38: 202-208, 1'979.

-T65-
Richardson, J.B. and Ferguson, c.c. Morphology of the aÍrways;Physio'logy and Pharmacology of the Airways, edÍtor J.A. Nade1,York, L-30, 1980.
1nNew
Rinard, 4.G., Rubinfeld, 4.R., Brunton, L.L. and Mayer, S.E. Depressedcycl íc AMP I evel s Ín airway smooth muscle fro¡n asthmatic dogs.Proc. Natl . Acad. Sci . 76(3 ): L472-L476, L979.
Rodbard, s. and Kira, s. Lobar, airway, and pulmonary vascular effectsof serotonin. AngÍo1 ogy 23: 188-197, 1972.
Rubinfel d, A.R. and Pain, M.C.F. Rel ationship betweenreactivity, airway cal iber, and severi'fy of asthma.Resp. Di s. 115: 381-387 , 1.977 .
bronchi alAmer. Rev.
Ruffifl, R.E., Dolovich, M.8., h'lo'lff, R.K. and Newhouse, M.T. The effectsof preferential deposition of histamine in the human airway. Amer.Rev. Respir. Dis. tI7: 485-492,1978.
Russe11, J.A. Responses of isolated canine airways to electricstimul ation and acetyl chol Íne. J. Appl . Physí01 . 45(5 ): 690-698,t978.
Russell, J.A. and Bartlett, S. AdrenergÍc neurotransmíssion in airways:Ínhibition by acetylcholine. J. Appl. Physiol. 51(2): 376-383,1981.
Russell, J.A. and Lai-Fook, S.J. Reflex bronchoconstrÍction induced bycapsaicin in the dog. J. Appl " Physiol . 47(5 ): 96L-967 , 1979.
Sampson, S.R. and Vidruk, E.H. Properties of irritant receptors incanÍne lung. Respiration Physiology 25: 9-22, 1975.
Sax, R.D" and lnlest,fall, T.C. Alterations in prejunctional adrenoceptorfunctÍon in guínea-pig and rat was deferens after chronic ganglionicblockade. J. Pharmacol. Exp. Ther. zLgz 2L-26, 198L.
tz, G. Hard¡nan, J.G., Schul tz, K. Baird, C"E. and Sutherland, E.l,rl.The importance of calcium ions for the regulations of guanosine3':5'-cyclic monophosphate levels. Proc" Natl. Acad. Scí. USA, 70:3889-3893 , L973.
Russell, J.A. NoradrenergicJ. Appl . Physiol . 48(1 ):
SeverÍnghaus, ,J.W" and Stup'lel , M.fol 1 owi ng atropi ne in man andblockade and hypothermia in dogs"1955.
inhibitory innervatÍon of canine airways.t6-22, 1980.
hulSc
Respi ratoryatropi ne,
J. Appl .
dead space increasevagal or gangnl ionic
Physio'l . 8: 81-87,

Silva
- .l66 -
Shanes, A.M. and Bianchi, C.P. The distribution and kinetics of releaseof radiocalcium ín tendon and skeletal rnuscle. J. Gen. Physiol. 422L1,23-LL37, 1959.
Sheller, J"R., Holtzman, M.J., Skoogh, B. and Nadel, J"A. Interaction ofserotonin with vagal-and ACh-induced bronchoconstriction in canínelungs. J. Appl . Physiol.: Respirat. Environ" Exercise Physioì.52(4): 964-966, L982.
, D.G" and Ross, G. Ultrastructural and flourescence hístochemicalstuclíes on the innervation of the tracheobronchial muscle of catsand cats treated with 6-hydroxydopamine. J. Ultrastruct. Res. 47:310-328, L974.
Simonsson, 8.G., Andersson, R., Bergh, N.0., Skoogh, B.-E. and Svedmyr,N. In vivo and in vitro studies of pharmacological effects ondi fferent receptors regul ati ng bronchi al tone i n man. In:Bronchitis III (f¿. by 0rÍe, N.G.M and van der Lende, R.) RoyalVangorcurn, Assen, 334, 1970.
Sirnonsson,8.G., Jacobs, F.M. and Nadel, J.A. Role of autonomic nervoussystern and the cough reflex in the increased responsiveness ofairways Ín patients with chronÍc obstructÍve airway disease. J"Cl in. Invest. 46¿ 1812-1818, L967.
Simonsson, B.G. and Svedmyr, N. BronchoconstrÍctor drugs. Pharmacol.Ther. B, 3: 239-303, L977.
Simonsson, 8.G., Svedmyr, N", Skoogh, B.-E., Andersson, R" andN.P. In vivo and in vitro studies on a'lpha-receptors inai rwaysT- E:Eenti ation ln-Tñ-bacterÍ al endotoxi n. Scand. J .DÍs. 53: 227-?36, L972.
Bergh,humanresp
"
Smaldone, G.C. and Bergofsky, t.H. Delineation of flow-limiting segmentand predicted airway resistance by moveable catheter. J. Appl.Physiol. 40: 943-952, L976"
Snapper, J.R., Drazen, J.M., Loring, S"H., Braasch, P.S, and Ingram,Jr. Vagal effects on histarnine, carbachol, and prostaglaFZal pha responsiveness i n the dog. J. Appl . Physiol . 4713-16 , 1"979.
R. H.ndi n(1):
S now, R.M., Miller, W"C., Blair,asthna. Annal s of Al'lergy
. and Ri ce, D " L. Inhal ed atropi ne i rt
286-?89, L979.
Pharmacol ogy22: 249-353,
H.T422
Somlyo, A
ofnI 970
.P. and Somlyo, A.V. Vascul ar s¡nooth rnuscle II.ormal and hypertensÍve vessels, Pharmacol. Rev.
Starke, K, Regulation of noradrenaline release by presynaptic receptorsystems. Rev. Physiol. Biochem. Pharmacol. 77: L-L24, L977.

-167-
Starke, K. Alpha-sympathomimetic inhibition of adrenergÍc and cholÍner-gic transmission in the rabbit heart. Naunyn-Schmiedeberg's Arch.Pharmacol . 274: 18-45, L972.
Starke, K., Endo, T. and Taube, H.D. Rel ative pre-and postsynapticpotencies of -adrenoceptor agonists in the rabbit pu'lmonary artery.Naunyn-Schmiedberg's Arch. Pharmacol. 29L: 55-78, L975"
Stephens, N.L. Airway smooth muscle. Lung Cells in Disease (ed. A.Bouhuys) , Biomedical Press, Ë1 sevier, l'lorth Hol I and, 113-138 , L976.
Stephens, N.L. Airway smooth rnuscle: bÍophysics, biochemistry andpharmacology, in Asthma:ment, (1.M. LÍchtenGEãln
Immuno harmacol and Treat-s nr e rs , c Pre35,
Force-vel oci ty Characteri s-J. Appl . Physí01 . 26(6lz
P sioloan-i-rä,
New York , L47-L67, L977.
St,ephens, N.L. and Kroeger, E. Effect of hypoxia on a'irh,ay srnooth musclemechanics and el ectrophysiol ogy. J. Appl . Physiol . 28(5 ):630-635,1970.
Stephens, N.L. and Kroeger, E.A. Ultrastruc'üures, biophysics, and bio-chernistry of airway smooth rnusc'le; in Physiology and Pharmacology oft,he Ai rways, edi tor J.A. Nadel , New Yorl<, 3t-L22, 1980.
Stephens, N.l-., Kroeger, E.A. and Kromer, V. Induction of a myogenicresponse in tonÍc airway smooth muscle by tetraethybammonium. Amer.J. Physíol. ?28(2): 6?8-632, L975"
Stephens, N.1., Kroeger, E. and Mehta, J.A.tícs of respiratory airway srnooth muscle"685-692, 1969.
Stephens, N.L., Mitchell, R.W., Antonissen,1"A., Kromer, V., Hanks,8.,Kroeger, E.A. and Kepron, I^l. Airway srnooth muscle: physicalproperties and metabol i sm. In" Airway Reactivity: mechanisms andc1Ínical relevance. Editor t.E. Hargreave, Astra Pulmonary, Canada,110-131,1980.
St,erling, G.8., Ho1st, P.E. and Nattel , J.A. Effect of C0 and ptl onbronchoconstrÍction caused by serotonin vs. acety'lcholine. J. Appl.Physiol. 32: 39-53, L972.
Stjarne, L. Inhibitory effect of prostaglandin E2 on noradrenalinesecretíon from sympathetÍc nerves as a function of external calcium.Prostagl andi ns, 3: i05-109 , L973.
Stone, D.J., Sarkar, T.K. and Ke1tz, H.
and inhÍbition on hu¡nan airways"624-627, L973.
Effect of adrenergic stimul ationJ. Appl . Physioì . 34(5 ):

- t6B -
Strauss, R.H., McFadden, E.R" JF., Ingram, R.H. Jr. and Jaeger J.J"Enhancement of exercise-induced asthma by cold air breathing. NewEngl . J . Med. 297: 743-747 , L977 .
Strauss, R"H., McFadden, E.R. Jr., Ingram, R.H. Jr., Deal, E.C. Jr. andJaeger J.J" Influence of heat and humidity on the airway obstruc-tion induced by exercise in asthma, J. Clin" Invest. 61: 433-440,L978.
Suzuki, H., Morita, K. and Kuriyama, H. Innervation and properties ofthe smooth rnuscle of the dog trachea. Jap. J. Physio'l . 262303-320, L976"
Szentivanyi, A. The conformational flexibility of adrenoceptors and theconstitutional basis of atopy trÍang'le, Sandoz Journal of MedicalScience, 1B(4 ) : 109, L979.
Szentivanyi, A. The beta adrenergic theory of the atopic abnormal ity inbronchi al asthma. J. Al 1 ergy 42: 203-232, 1968.
Takizawa, T. and Thurlbeck, I,rl"M. Muscle and mucous gland size in themaj or bronchi of pati ents wi th chronic bronchi tÍ s, asthrna andasthmatic bronchítis" Amer. Rev. Respir. Dis. 104: 331-336,T97L.
Tashki n, D"P. , Conol'ly, M.E. , Deutsch, R"E. , Huí , K.K. , Lettner, M.,Scarpace, P. and Abrass, I. Subsensitization of beta-adrenoceptorsin airways and lymphocytes of healthy and asthmatic subiects. Am.Rev. Resp. Dis. L25: 185-193, 1982.
Tattersfiel d, 4.E., Leaver, D.G. and Pridebeta-adrenergic blockade and stimulation onJ. Appl. Physiol. 35(5): 6L3-619, L973.
Tauc, L. Nonvesicular release of neurotransmitter.857-893, L98?.
N. B.normal
tffects ofhuman aÍ rways.
Physiol . Rev. 62(3 ):
Townley, R.G., Dennis, M. and I'Ekin, J.M. Comparative action ofacety'l beta-methacholine, histamine and pol1en antÍgens in subiectswith hay fever and patients with lrronchial asthma" J. A1ìergy 36:L2L-t37, 1965
Townley, R.G., McGrady, S. and Bewtra, A. The effect of beta-adrenergicblockade on bronchial sensitivi'ty to acetyl-beta-methachol ine innormal and al lergic rhinÍtís subjects. J. Al'lergy Cl in. Imrnunol .57: 358-366 , L976.
Tung, R.S" and Litchtenstein, L.M. Cyclic AIvIP agonÍst inhibitionincreases at low levels of hístamine release from human basophils"J. Pharmacol. Exp. Ther. 2L8z 642-646, 1981.

- 169 -
Varonier, H.S. and Panzani, R. The effect of inhalations of bradykíninon heal thy and atopi c ( asthrnatic ) chi I dren. Int. Arch. Aì I ergyAppl . Immunol . 34: ?-93-296, 1968.
Verhaeghe, R"H., Vanhoutte, P.M. and Shepherd, J.T. Inhibition ofsympathetÍc neurotransmissíon in canine blood vessels by adenosineand adenine nucleotides. Circ. Res. 402 208-2L5, L977.
Vermeire, P.A. and Vanhoutte, P.M. Inhibítory effects of catecholaminein Ísolated canÍne bronchÍal smooth muscle. J. Appl. Physio'1.46(4)z 787-791, L979.
Vidruk, E.H., Hahn, H.1., Nadel, J.A. and Sampson, S.R. Mechanism ofstimulation of fast-adaptíng receptors by histamine ín dog lung(Abstr). Fed. Proc. 35: 841, L976.
Vi ncent, N.J . , Knudson, R. , LeÍ th, D. E. , Mackl em,Factors influencing pulmonary resistance. J.236-243, 1970.
P. T.Appl .
and Mead, J.Physiol. 292
Vizi, E.S. and Kno11, J. The effects of sympat,hetic nerve stimulationand guanethidine on parasympathetic neuroeffector transnission; theinhibÍtion of acetylcholine release. J. Pharm. Pharmacol. 23:918-925, L97L.
Wanner, 4., Mezey, R
bronchospasm in9L7-9?.2, L979.
J., Reinhart, M.E.conscious sheep.
and Eyre, P. Antigen-inducedJ. Appl . Physiol . 47(5 ):
Wasserman, M.A. Bronchopu'lmonary responseshistamine and acetylcholine in the dog146-155, L975.
to prostagl andi n F2a1 pha,tur. J. Pharmac . 322
Wasserman, M.A. Bronchopulmonary pharmaco'logy ofendoperoxide anal ogs ín the dog. European103-114, L976.
some prostagl andinJ" Pharmacol. 36:
['leiner, N. Multiple factors regulating the release of norepinepheineconsequent to nerve stimulation" Fed Pro. 38: 2L93-2202, L979.
l^leíss, E.B. and Viswanath, S.G. Calcium hypersensítivi'ty in airwayssmooth muscle. Resp. 38: ?-66-?72, L979.
tleiss, S., Robb, G.P. and Ellis, L.B" The systemic effects of histaminein man" Arch. Intern Med. 49: 360-396, L932.
Jp
Ioe
r,ca
t'lgme
reagTh
., Van der Lugt, J.C., de Langen, C.D.J. and Mulder, A.H. 0n theacity of presynaptic alpha receptors to modulate norepinephrineease from sl ices of rat neocortex and the affi nity of somenists and antagonists for these receptors. J. Pharmacol. Exp.r. ZLIz 445-45L, L979.

-170-
l^les't,fa1l, T.C. Local Regul ation of adrenergic neurotransnrission.Physiol. RevÍews 57(4): 659-7?8, L977.
I^Jiddicombe, J.G. Regul ation of tracheobronchial smooth rnuscle. Physioì .Rev. 43: t-37, 1963.
WiddÍcombe, J.G. Action potentials inefferent fibers to the trachea andPhysiol. (London) 186: 56-88, 1966.
parasynpathetic andlungs of dogs and
sympatheticcats. J.
l^liddicombe, J.G. Reflex control of airb,ays srnooth muscle. PostgraduateMed" Journal 51 (7 ): 36-43, 1975.
t.liddicornbe, J.G. and Nadel, J.A. Aírway volume,work and force of breathÍng: theory. J863-868, 1963"
aírway resi stance,Appl . Physiol.
and18:
Ìdilliams, M.l{. Jr. [.lhy do the airways contaín smooth muscle? Lung 159:29t-293, 1981
"
Wilson, A.F. and Galant, S.P. Recent advances in the pathophysiology ofasthrna. þJest. J . Mecl. 120: 463-470 , L974.
Wilson, A.F. and McPhillips. Pharmacological control of asthma. Ann.Rev. Pharmacol . Toxicol . 18: 541-561, 1978.
Wood, C.1., Arnett, 0.D., Clarke, l^l.R., Tsai, B.S. and Lefkowitz, R.J.Subclassification of a'lpha-adrenergic receptors by dÍrect bindingstudÍes. Biochern. Pharmacol. 28: L277-LZBT, L979.
l^loolcock, A.J. , Mackl enr, P.T. , Hogg, J "C. and Wi1 son, N.J. Infl uence ofautonomic nervous system on aírway resistance and elastic recoil.J. Appl. Physiol. 26(6): 814-818, 1969.
Woolcock, A.J. Mack'lem, P.T., Hogg, J.C., Wí1son, N.J., Nadel , J.4.,Frank, N.R. and Brain, J. tffect of vagal stimulat,ion on centraland peripheral airways in dogs. J. Appl. Physiol . 2(t(6lz 806-813,I 969.
Yanta, M.4., Loring, S.H., Ingram, R.H. Jr. and Drazen, J.M.reflex bronchoconstriction induced by hístamÍne aerosolin dogs" J. Appl. Physiol. 50(4): 869-873, 1981.
Di rect andinhal ation
Yu, D.Y.C., Ga'lant, P. and Gold, t,l"M. Inhibition of antigen-inducedbronchoconstríction by at,ropine Ín asthrnatic patients, J" Appl.Physiol. 32: 823-8?8, L972.
Zimmernan, B.G. Actions of angiotensin on arlrenergic nerve endÍngs.Fed" Proc. 372 L99-202, L978.

Zuski
- 171
rì, E. , Mi tchel 'l , C.A. and lJouhuys, A. Interaction between ef fectsof beta blockade and cigarette srnoke on airways. J. Appl. physiol.36: 449-452, L974.

t72
AP PEND i X
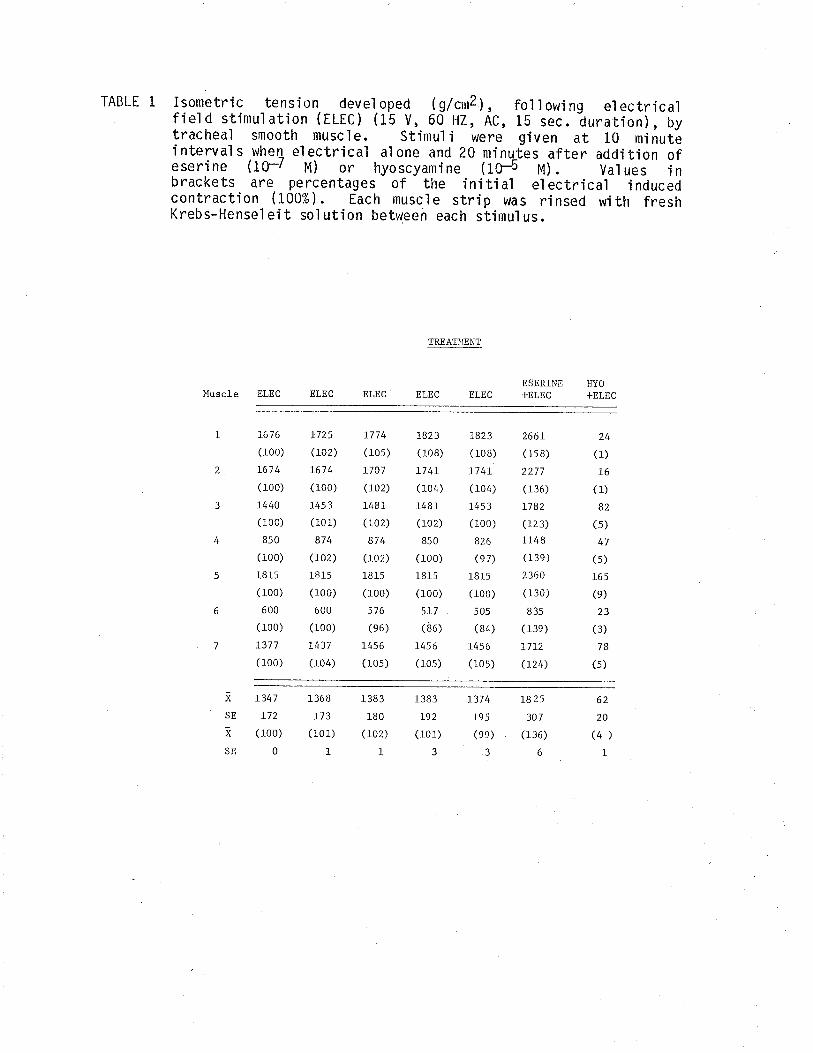
TABLE 1 Isometric tension. deve'l.oped (g/ctn?), fo'llorrring electricalfield stimulation (ELEc) (ts v, 60 HZ, Ac, t5 sec. ?uration), bytracheal smooth muscle. stirnul i were given at l0 miñuteintervals wheq electrical alone and 20 minuLes after additÍon ofeserine (to-7 M) or hyoscyamine (l15--ii. val ues inbrackets are percentages of the ini fial el ectrical inducedcontraction (100%). Each muscle strip was rinsed wÍth freshKrebs-Henseleit solution betvleeÀ each stimulus.
TREAT}ÍENT
Muscle ELEC ELEC ELEC ELEC ELECESERINE+ELEC
HYO
+ELEC
2
3
4
5
6
l676( 100)
L67 4
( r0o)
7440
( 1oo)
850
(100)
1815
( 100)
600
( 100)
L377
( 100)
r725
(r02)
L67 4
( 1oo)
r45 3
(101)
874
(Loz)
18 15
( r00)
600
(100)
L437
( 104)
r77 4
(10s)
r707
( 102)
148 I( 102)
874
( 102)
1815
( 100)
576
(e6 )
r456(10s)
1823
( roe;
17 4l( 104)
14 81
( 102)
850
( r00)
1815
( 100)
517
1 à6)
1456
(los)
r82 3
( 108)
17 41
( 104)
1453
( 100)
826
(e7)
18 15
(100)
505
( 84)
1456
( 10s)
266r
(1s8)
2277
( 136)
Lt82
(r23)1148
(t3e)
2360
( r30)
835
(139)
17l.2
(r24)
24
(1)
76
(1)
82
(s)
47
(s)
165
(e)
¿)
(3)
(s)7
SE
X
r347
172
( 100)
0
L37 4
195
( 9e)
3
62
20
(4)1
1368
173
( 101)
1
180
( 102)
I
1383
192
(101)
J
t825
307
(1 36)
6SE
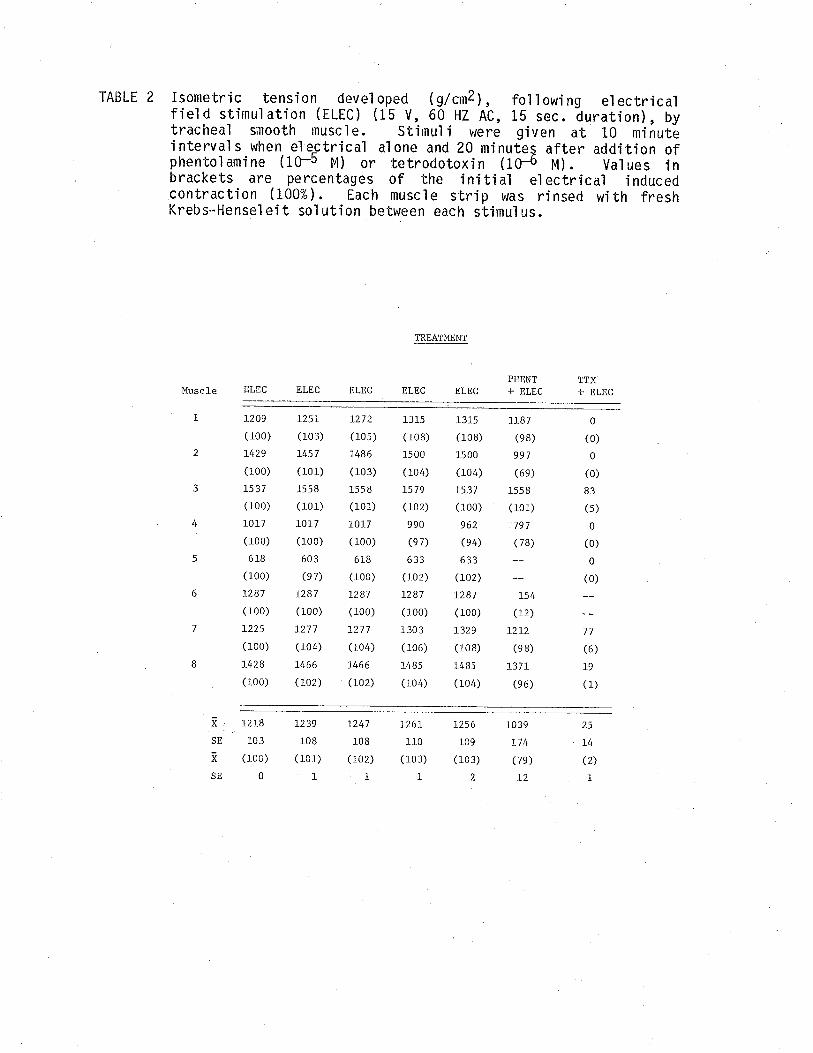
TABLE 2 Isometric tension. developed (g/cnZ\, following electricalf iel d stirnul ation (Elr_c) (ts v, 60 HZ Ac, l5 sec. ãuration), bytracheal smooth muscle. stimul i were gÍven at 10 miñuteÍntervals when gleJ_-trical alone and Z0 minutes after addition ofphentoì amÍ ne (to-5 M) or tetroàotoxi n tlo-0 il. vãl u.l inbrackets are percentages of the i ni ti al er ectrical i nducedcontractíon (100%). Each muscle strip was rinsed with freshKrebs-Hensel ei t sol ution bettveen each stimul us.
TREATMENT
Muscle ELEC ELEC ELEC ELEC ELECPHENT
+ ELECTTX+ ELEC
1209
( 100)
r429
( 100)
7s37
(1oo)
1017
( 100)
618
( 100)
1287
( 100)
r225
( r00)
l428( 100)
I25I(r03)
l.457
(r01)
1558
( 101)
TOIT
( 100)
603
(e7)
r287
( loo)
r277
(104)
].466
(102)
7212
(10s)
L486
( 103)
155 I( r0r)1017
( 100)
618
(1oo)
L287
(10o)
r277
(r04)
L466
'(ro2)
1315
(108)
15 00
( 104)
7579
(102)
990
(e7)
633
(102)
1281
( 1oo)
130 3
( 106)
1485
(104)
1315
(108)
15 00
(104)
1537
(100)
962
(e4)
633
( 102)
L281
( r00)
L329
(108)
14 85
( ro4)
118 7
(e8)
997
(6e)
155 I( 101)
101
( 78)
0
0
(0)2
J
4
5
6
7
(0)
83
(5)
0
(0)
0
(0)
(6)
19
(1)
154
(12)
12]-2
(eB)
r371
(e 6)
qF
ISE
12L8
103
(10o)
0
1,239
108
(101)
I
1247
108
(102)
1
1039
174
(te)12
T26I
110
(103)
1
L256
109
(103)
¿
L4
(2)
I
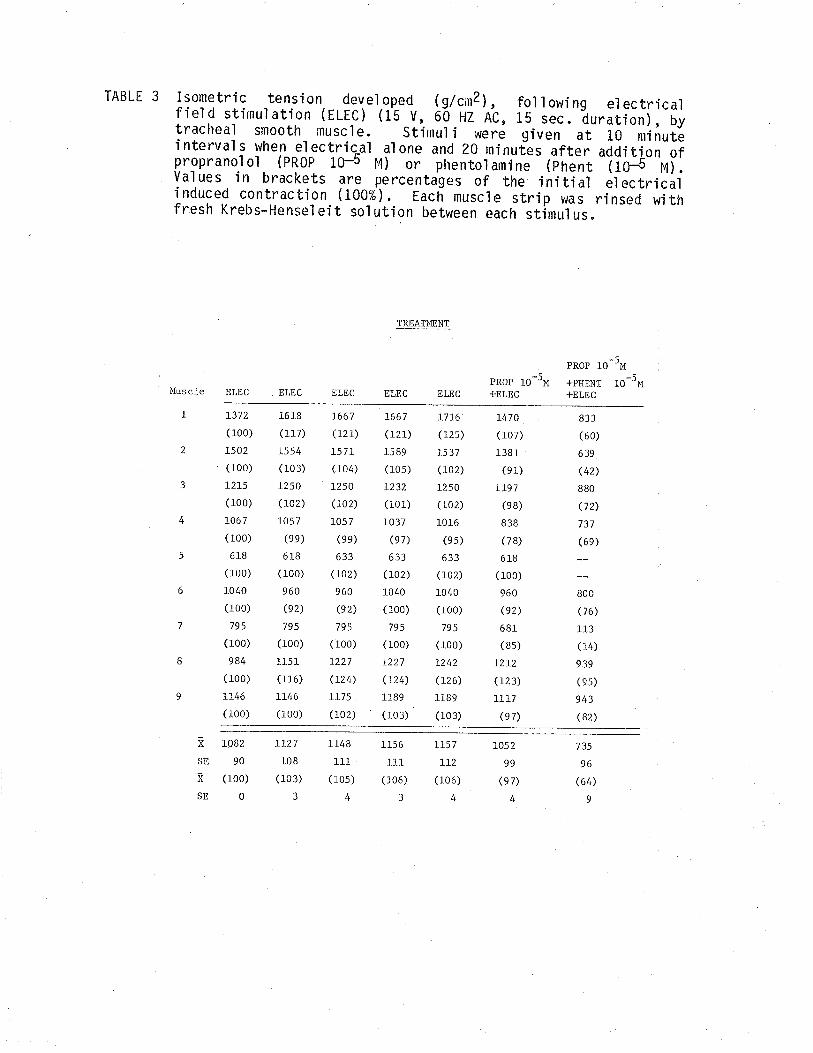
TABLT 3 Isometric tension,_ devel.op,ed (g/cm?) , foì louríng erectricarfiel,l stimulation (rlr-c) (t's v, oó Hz n-c, 1s sÀc. äuratióni, ovtracheal smooth muscre. stirnul Í weré given at 10 ninuteintervals when electric-al alone and z0 minutäs ìrt., addition órpr,opranolol (pnop 10-5 M) or phenrolarinJ- rËr,äît -iiË6""¡,li]
values Ín brackets gre percentages of the initial electriCalinduced contraction (L00y;). eacñ muscle strip yras rinsed withfresh Krebs-Henseleit solution between each stimulus.
TREATMDNT
Muscle ELEC . ELEC ELEC ELEC ELECPROP 10+ELEC
p*or 1o-5ot5r +pHENT to-5lt
+ELEC
I r372( 10o)
1502
(100)
1,215
( 100)
1067
( loo)
6L8
( 100)
1040
( 100)
795
(100)
984
( 1oo)
IT46
( 10o)
16 18
( 117)
rs54(103)
12 50
( 102)
105 7
(ee)
618
( r00)
960
(e2)
795
( 100)
115 1
(116)
r746
(100)
7667
(121)
I57 I(104)
r250(702)
105 7
( ee)
633
( 102)
960
(e2)
795
( 1oo)
r227
(L24)
L175
( 102)
1667
(12L)
15 89
( 105)
L232
(10r)
IO37
(e7)
633
( 102)
1040
( roo)
795
( ro0)
7227
(724)
1189
( ro3)
t7 76
(l2s)
1537
(Lo2)
1250
(102)
r016
(e 5)
633
( 102)
1040
(100)
795
( 100)
T242
(r26)
1189
(103)
7470
(107)
138 I(e1)
LLg7
(e 8)
838
(78)
618
( 100)
960
(92¡
681
(8s)
72T2
(123)
11 17
(e7)
833
( 60)
639
(42)
880
(7 2)
( 6e)
2
4
5
7
8
6 800
(7 6)
113
( 14)
939
( es)
943
( 82)
9
X 1082
90
(1oo)
0
LLz7
108
( 103)
3
1148
111
(tOs)
4
1156
111
( 106)
3
115 7
1_12
(106)
4
r05 2
99
(tt¡4
735
96
( 64)
9
SE
X
SE
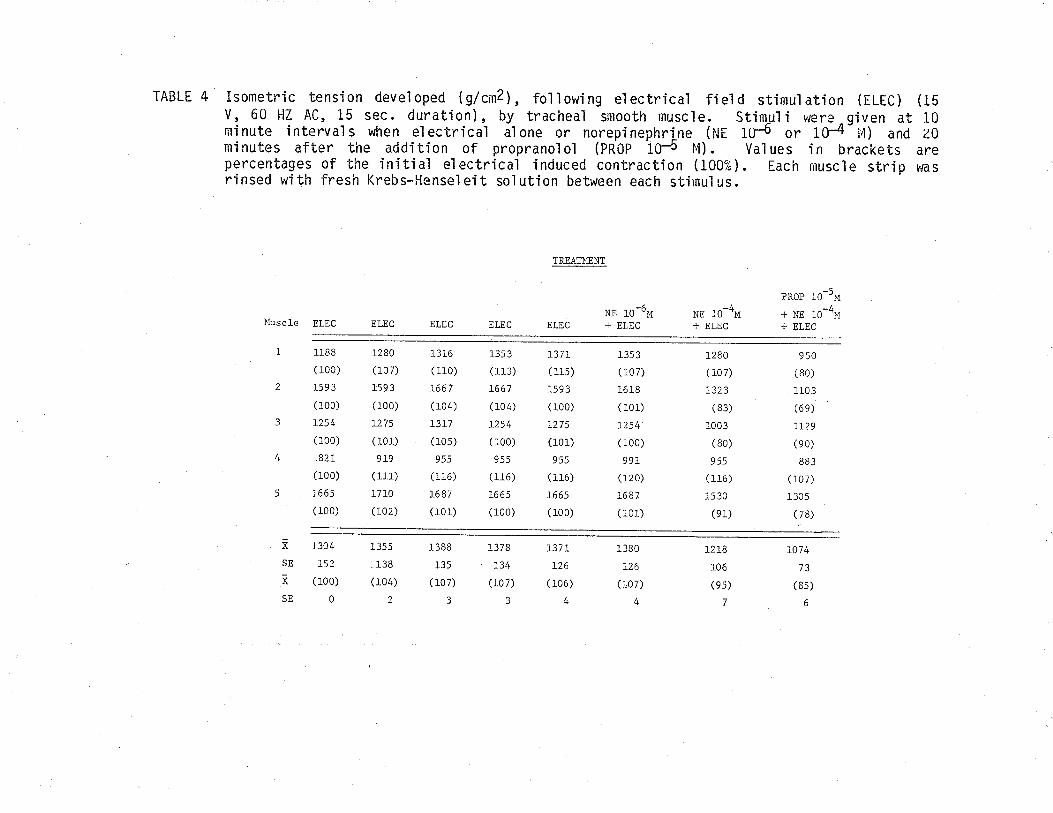
TABLE 4 Isometric tension devel oped (g/crn21 , fo1 lowÍng el ectrical fiel d stÍmul ation (ELtC)V, 60 HZ AC, 15 sec. duration), by tracheal snooth muscle. Stim-uli were siven atminute Íntervals when electrical alone or norepinephrlne (i'¡f 1É or 1É-M) andminutes after the additÍon of propranolol (pnOp if M). Values in bracketspercentages of the initial electrícal induced contraction (100%). Each muscle striprinsed v¡ith fresh Krebs-Henseleit solution between each stimulus.
(1510z0
arel{as
TREAT}IENT
-a -t,NE 10 "M NE t0 '¡l+ ELEC
PROP IO
+NE10+ ELEC
-JM
-1,'MIfuscle ELEC ELEC ELEC ELEC ELEC + ELEC
f
1188
(1oo)
1593
(100)
r254
( r0o)
821
( 100)
16 6s
( 100)
1280
(107)
ls93( 100)
r275
( 101)
919
( 111)
17l_0
(102)
1 355
138
( 104)
2
1316
( 110)
1667
( 104)
1317
( 10s)
95s
( 116)
1687
(101)
(1r3)
1667
(r04)
I254( 1oo)
955
( 116)
7665
( 100)
I37I( ll-s )
159 3
( loo)
r27 5
(101)
9s5
( 1"16)
1665
( 100)
135 3
(107)
1618
( 101)
I254( 100)
997
( 120)
1687
( 10r)
(107)
7323
( 83)
1003
( 80)
955
( 116)
l5 30
(9 r)
12 18
106
( es)
7
9s0
(80)
110 3
(6e)
7129
(eo)
883
( r07)
1305
( 78)
5
X
SE
X
cË
1304
L52
(100)
0
135
(107)
J
I37 T
L26
(106)
4
137 8
134
(107)
3
1380
t26( 107)
4
r07 4
/3
(8s)
6

TABLT 5 Isometric tensíon (g/çnZ),, norepi nephri ne (¡¡f 10-ö to
i n brackets are percentages(100%).
oped fol 1 owi ng addì tion ofM) and phento'l ami ne (iÉ M)the initial tension devel oped
devel1É
of
potassí urn (t<+
cumul ati v e'ly .fol I owi ng ¡1+
23 mM),Val ues
expo sure
Mus c1e
TRNATMENT
23 .¡t K* ,u to-8 M NE ro-7 ¡r *" to-6 M NE to-5 * u¡ ro-4 M pHENT t0-5 t507 57r 660 622 330 203 5r
(100) (1"12) (130) (L23) (6s) (40) (r0)854 880 920 998 880 657 34r
(100) (103) (108) (r17) (i03) (7t¡ (40)
163 163 169 136 109 87 7r(100) (r00) (103) (83) (67) (53) (43)
191 270 268 344 1s3 77 38
(100) (110) (140) (r80) (80) (40) (20)
4i6 456 473 529 489 244 33
(100) (110) (114) (L27) (1r8) (s9) (8)
232 250 267 285 :-78 98 54
(100) (108) (115) (123) (77) (42) (23)1195 1247 13i.8 1423 1423 1370 703(100) (104) (r10) (lre) (l-le) (11s) (se)
2
J
4
5
6
7
x 507
746
(r00)
0
539
151
(107)
2
JOa
L57
(ir7)
619
L70
(r25)
1I
508
782
(eo)q
390
180
(6 1)
10
183
96
çzo)
l
SE
iSE
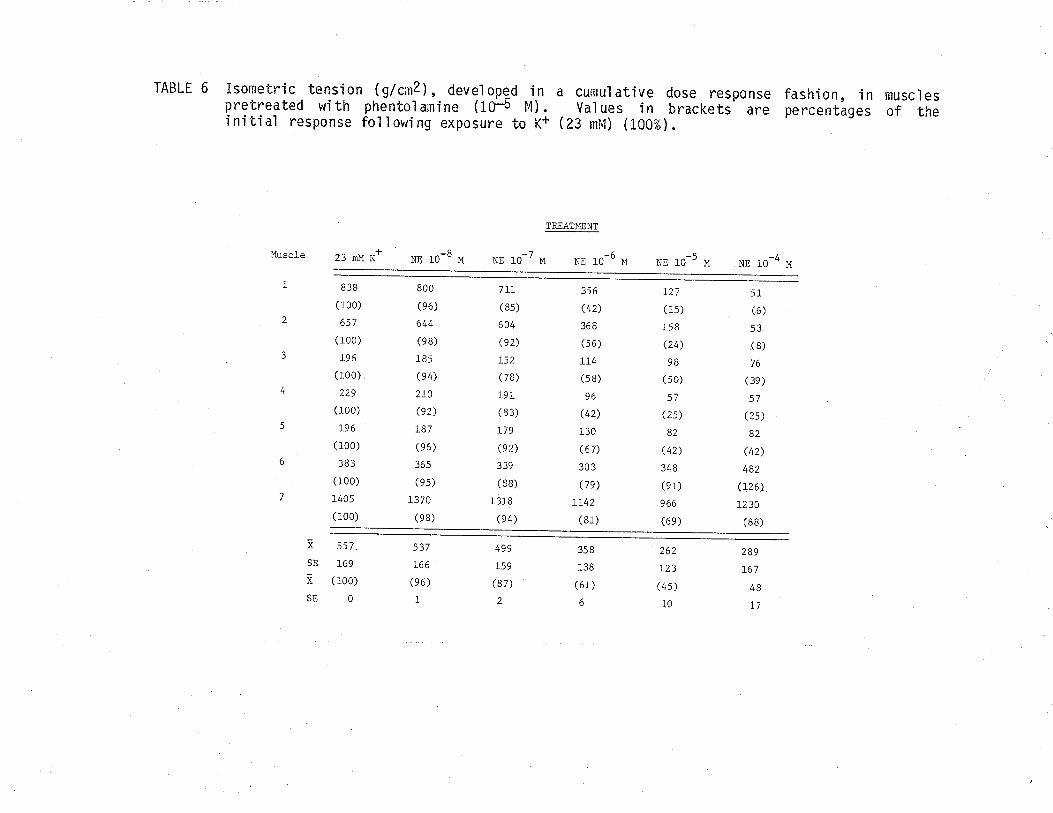
TABLE 6 Isometríc tensi on (g/ cnZ) , deve'l ope_d i.n a cumul ative dose responsepretreated wi th phento'l amine (1f M) . val ues in brackets arei ni ti al response fo'l I owi ng exposure to K+ ( 23 mt{) ( 1002" ) .
fashion, inpercentages
musc I esof the
TRNATMENT
t{uscle 23 r¡M x* uE t0-8 M NE -110M ton 10-6 M NE to-5 l,r "n
to-4 *1
2
3
4
6
7
)
838
( 100)
657
( 100)
796
( 100)
)ra( 1oo)
196
( 10o)
JOJ
( 1oo)
1405
( 100)
800
(e6)
644
(e 8)
185
(e4)
2L0
(e2)
10?
(e6)
365
(es)
13 70
(e8)
71L
(8s)
604
(e2)
752
( 78)
191
( 83)
779
(e2)
339
( 88)
(e 4)
3s6
(42)
368
(s6)
tr4(58)
96
(42)
130
(ot)303
(7e)
LT42
( 81)
127
( rs)
158
(24)
98
(so)
57
(2s)
Õ1
(42)
348
(er)
966
(6e )
51
(6)
53
(8)
76
(3e)
(25)
82
(42)
482
(726)
7230
(88)
x 557.
SE 169
Í (roo)
SEO
537
166
(e 6)
I
499
159
(87)
2
âco
1âO
( 61)
6
¿o¿
L23
(4s)
10
289
]'67
48
77
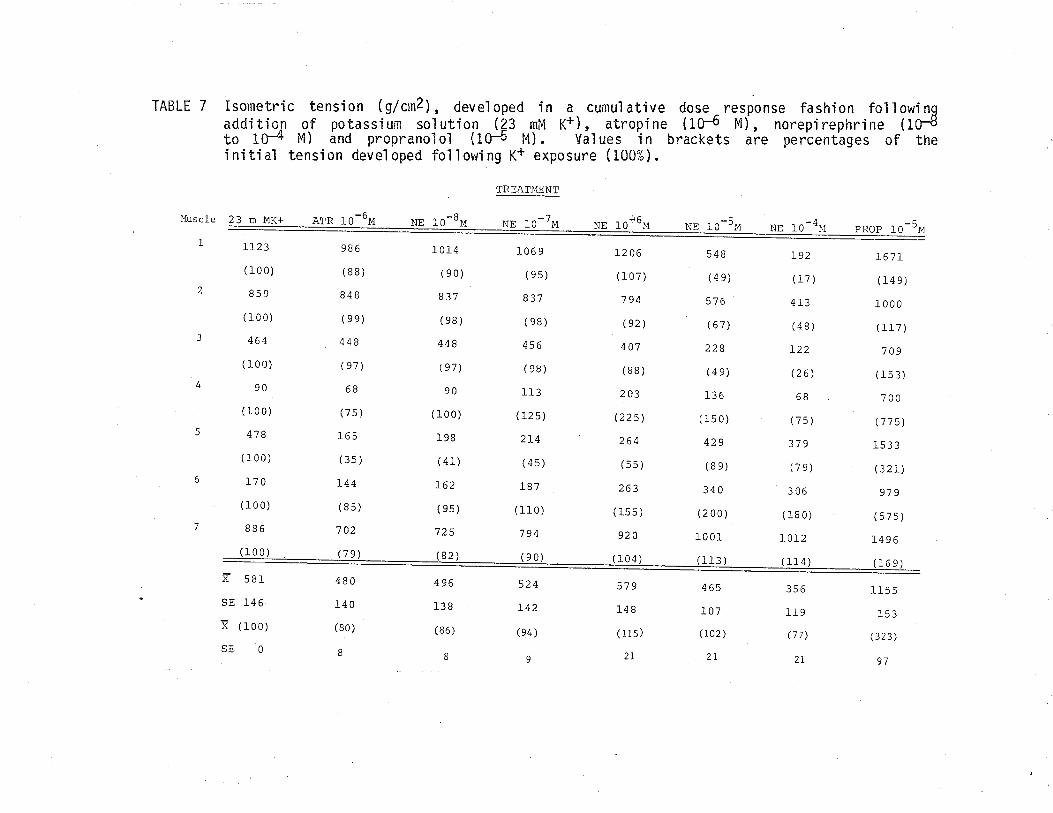
TABLE 7 isornetric tensÍon (g/cn2), developed in a cumulativeaddi tÍon of potassi um sol ution Q3 mM K+) , atropi neto 1É M) ånd pnopranol o1 (to-5 M) . Vál ues ìninitial tension developed frrllowing K+ exposure (100%).
dose ^ respo(10-0 M),
I owi ng(1N
bnackets are
nse fashion folnorepi rephri ne
pe rcentag es of the
Muscle 23 m MK+ -EatR ro-br,t nn lo-8M
TREATMENT
_aNE 10,M
106 9
(es)
837
( e8 )
456
(e8)
113
(125)
zL4
(4s)
I87
(110)
794
(90)
E' A
I42
(e4)
9
-M NE 10 'M t¡r 10 -lr PROP 10 .MNE 10
2
112 3
(100)
859
(100)
464
(100)
90 .
(100)
418
(100)
170
(100)
886
(100)
986
(88)
848
(ee)
448
(97 )
68
(7 s)
165
(3s)
I44
(8s)
702
(7 e)
48ô
140
(80 )
8
10 t4
(e0)
837
(e8)
448
(97 )
90
(100)
100
( 41)
I62
(es)
725
(82)
496
138
(86 )
8
1206
(107)
794
(e2)
407
(88)
) ñ2
(22s)
264
(ss)
aca
(lss)
920
(104)
548
(4e)
576
(67 )
228
(4e)
136
(ls0)
429
(89)
340
(200)
10 0I
( rt3
465
107
(i02)
2I
r92
(17 )
413
(48)
122
(26)
68
(7s)
379
(1e)
306
(r80)
I 012
16? I(r4e)
1000
( r_17 )
709
(1s3)
700
(775)
1533
(32L)
979
(s7s)
l-49 6
3
4
5
6
7
X- 58I
SE 146
I (roo)
SEO
R?O
148
(1rs)
2T
( 1r4 (169
1155
153
(323)
97
356
119
çtt)
2T
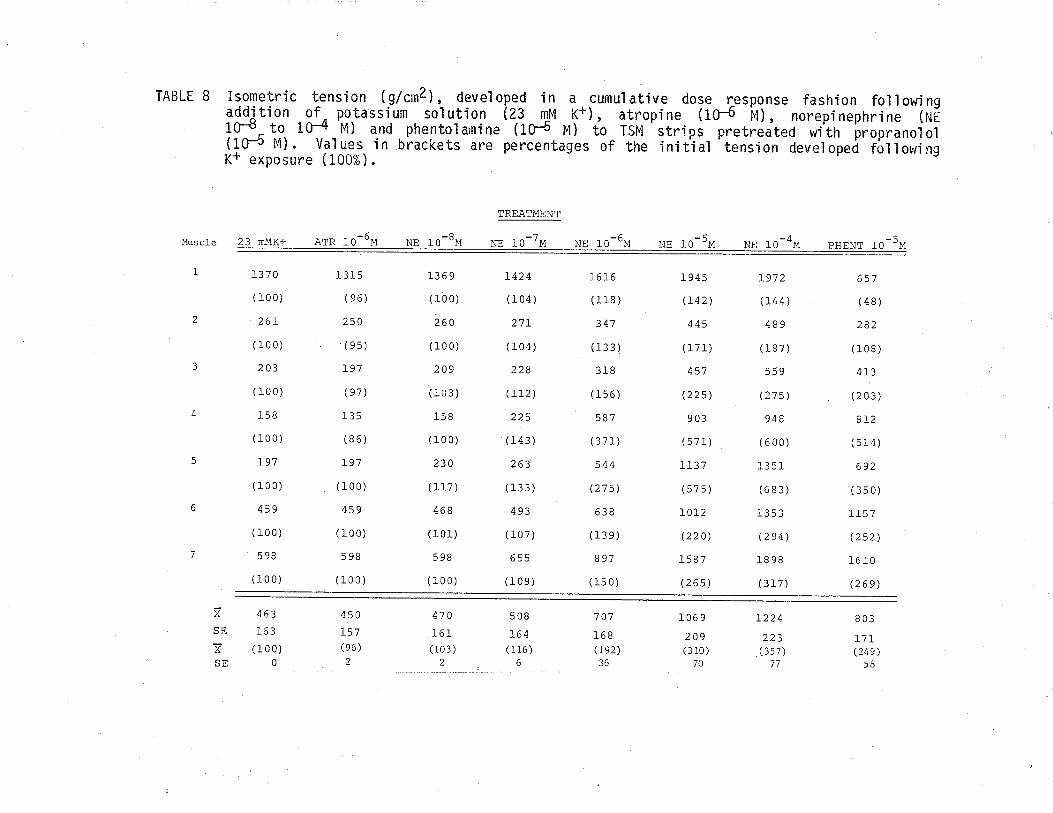
TABLE B Isornetric tension (g/c¡nZ) , devel oaddjtion of" potassi um sol utionlf _ to lft M) and phentol ami ne(1f l{). Values in brackets areK+ exposure (100%).
cumul ative dose Lesponse fashion fo1 I owi ng) , atropíne (1É it4) , norepi nephrine (NÉto TSM stríps pretreated with propranololof the inÍtial tension developed following
ped in a(Zl mM ç+(1É M)pe rcentages
TREATMENT
Muscle 23 mMK+ ATR 10 -6
13 15
(e6)
250
(es)
L97
(e7 )
135
(86)
197
(100)
4s9
(100)
598
(100)
450
157(e6)
2
1369
(10 o)
260
(100)
209
(103)
158
(100)
230
(117)
468
(r0l_)
598
(100)
41 0
161(ro3)
2
1616
(118)
(133)
318
(1s6)
s87
(371)
544
(27 5)
638
(r39)
897
(1s0)
I945
(L42)
445
( 171)
457
(225)
on?
(s7r)
LI37
(s75)
I012
(220)
1ÉO?
(265)
I97 2
(r_44)
489
(r87)
559
(27 5)
948
(600)
135r
(683)
1353
(2e4)
18 98
(317)
M -a -aNE 10 "M NF: 10 , M NE to-6¡,t ,0" ro-5r ue r0-4¡l pHENT lo-5r"r
2
1370
(100)
26l_
(100)
203
(100)
158
(100)
r97
(100)
459
(100)
598
(100)
r424
(104)
27r
(104)
228
(rl-2)
(r43)
263
(133)
Á oa
(107)
655
(r09)
657
(48)
282
(108)
413
(203)
8L2
(s14)
692
(3s0)
11s 7
(2s2)
161_0
(26e)
3
4
5
6
7
XqF
X
SE
463
163
(100)0
508
164(116)
6
707
168(re2)36
1069 1224 803
171(249)
58
209(3r0)
70
(357)77
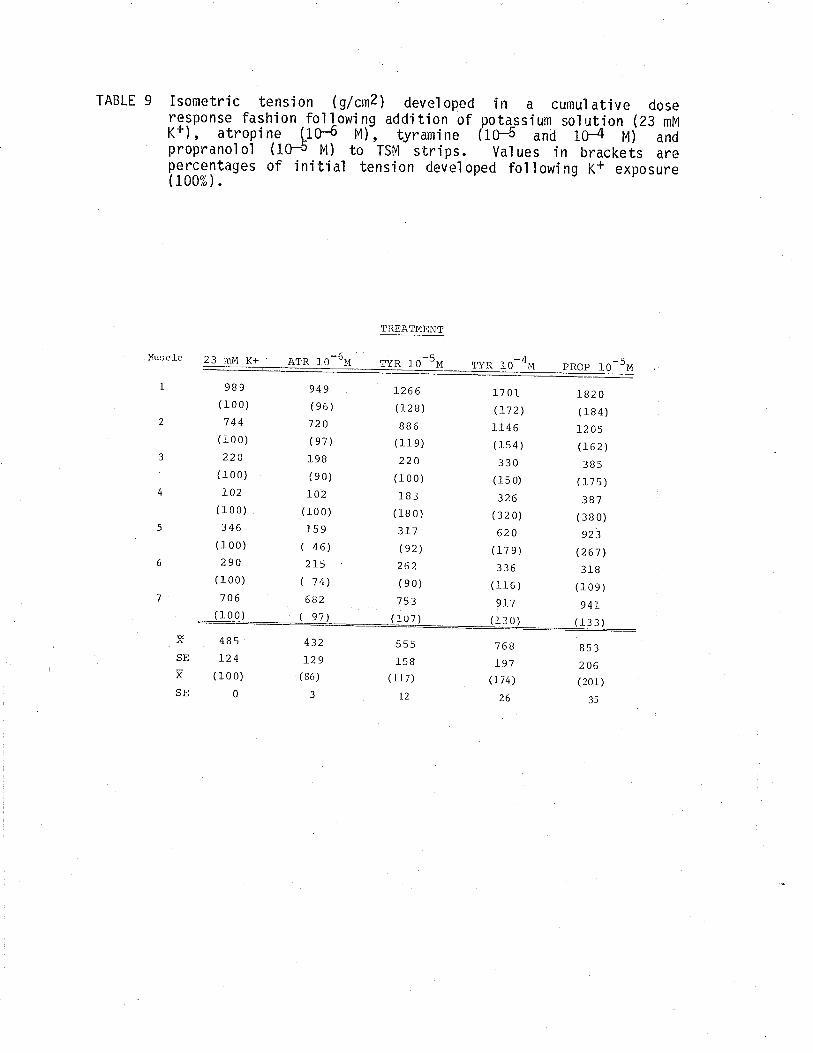
TABLE 9 Isometric tension (g/ctn?) devel oped Ín a cumul ative doserÇ:ponse fashion following addition of pot{ssiuin solution (23 mMK+) , atropi ne ßÉ M)-, tyrami ne (tO-S anO
- fF mi -
aÀ¿proprano] o] (10-5 M ) to TSM stri ps. val ues i n brackets arepercentages of initial tension developed following K+ exposurefi00%).
TREATMENT
Muscle 23 mM K+ ATR I o "¡l ryR r o-tl¿ TYR. t-0_Â=¡¿ pnop l-o-'trt
I 989(r00)744
(100)
220(100)
702(100)
346(100)
290(100)
706
100
949(e6)
720(e7)
198
(eo)
I02(100)
ls9( 46)
2Is( t+¡
1266(128)
886(1re)220
(100)
183(180)
377(e2)
262( e0)
17 01
(112)
l-14 6
(r54)330
(150)
326(320)
620(11 e)
336
(116)
9L7r-30)
182 0
(r84)12 05
(162)')oE
(17s)101
(380)
923(267 )
3 r-8
(10e)
94r(133
853
206(20r)
J)
2
3
4
5
6
7
xSE
X
432
729(86)
3
485
124(100)
0
682
97)753
(lu7
ttÉ
158(117)
I2
768
I97(t,7 4)
26
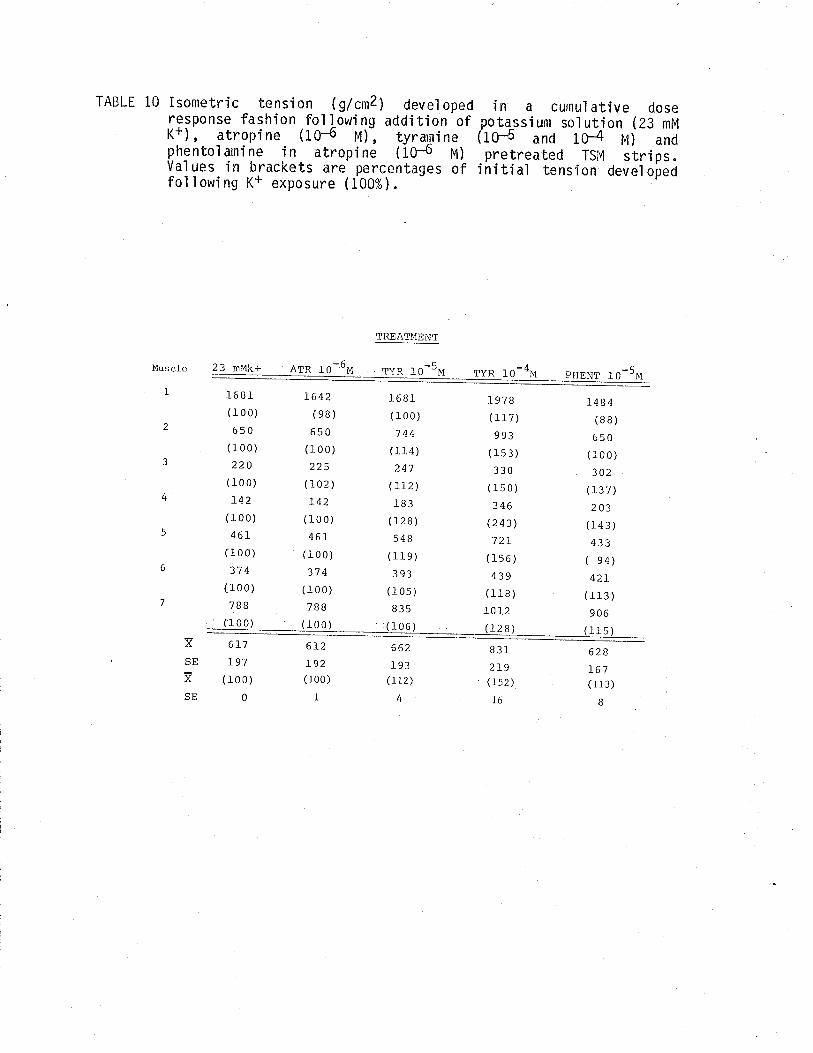
TABLT 10 Isometríc tension (g/ctn?) developedresponse fashion fo'l lovri ng addÍ tion ofK+), atropine (lù{ 14)-, tyrar¡inephento'l ami ne i n atropi ne (1f M)Val ues Ín brackets are percentages offollowing K+ exposure (f00%).
in a cumul ative dosepot{ssium solution (23 mM(1t-5 and 1É M) and
. pretreated TSM stri ps.initial tension develoþed
TREATMENT
Muscle 23 mMk+ arR l o-6¡.r -q,TYR IO 'M -ÂtyR t o =l¡ PHENT ro-Sot
2
4
5
16 81(100)
650(100)
220(100)
I42(100)46I
(100)
374(100)
788(100)
1642
(eB)
650(100)
aaÊ
(102)
142(100)
4 61_
(1oo)
374(100)
788
(roo)
16 81
(100)
744
(rr4)247
(112)
183
(128)
548
(tt9)? oa
(10s)
835
(106)
197 8
( 1r7)993
(1s3)
330(1s0)
346(243)
72r(156)
439(118)
I0 12
(l2B)
I48 4
(88)
6s0(100)
302(L37 )203
(r43)433
( e4)
42I(r13)906
6
7
( lrsX
ùE
cb
6I7L97
(100)
0
612
L92( ro0)
I
662
I93(rt2)
4
B3t2I9(r52)
L6
628
I67(113)
I

TABLE 11 Isometric ^
tensiol I g/cn?) devel oped in a cumul ative doseresponse fashion fol I owi ng addi tion' of^ potassi unr sol uiion dj-.4mM K+), norepineohrinã (ruE 1ñ-- -;;"'"iû_4'"'r), - andpropranol o] (t o-5 ) to TSM pr.i..utuo wí th atrãpi ne i r ro M ) .Val ues i n brackets are .perc.entages of i n í ti al teñsi on ievel opedfol I ouri ng K+ exposure (iOo"¿) .
TREATMENT
Muscle 17.4 nù,{K+ NE 10 -a"tq NE Lo-'M NE ro-6¡,r ña ro-5¡¿ NE ]. -Ào =l'r pRop lo-r¡.r
3
4
469(100)
2I3(100)
50
(-r00)
2L(r-00)
204(100)
I67(r00)186
480(102)
213(100)
50
(100)
2I(100)
265(r30)183
(r10)200
539(1ls)213
(r00)56
( 111)
2I(r-00)
286(140)
200(r20)264
(r42)
527
(t13)L42
( 67)
44
( 89)
2T
(100)
316
(r5s)250
(1s0)?\7
(re2)
304( os ¡
59
( 28)
39
( 78)
2I(100)
92
( 4s)I75
(105)
264
(L42\
2TL( 4s)
59
( 2B)
JJ
( 66)
2I(100)
6t( 30)
t00( 60)
242
r289(21 5)
Bt8(383)
I44(289)
256(1200)
108 2
(s3 0)
875(525)
479
5
6
7
(100 l0B ) ( 131 (2s8X 202
57
(e3)
t5
X
SE
SE
187
(r00)0
136
43
( 80).
15
225
65
(rl8)6
231
69
(124)
16
r0433
(66 )
T4
706
161
(4e4)
125
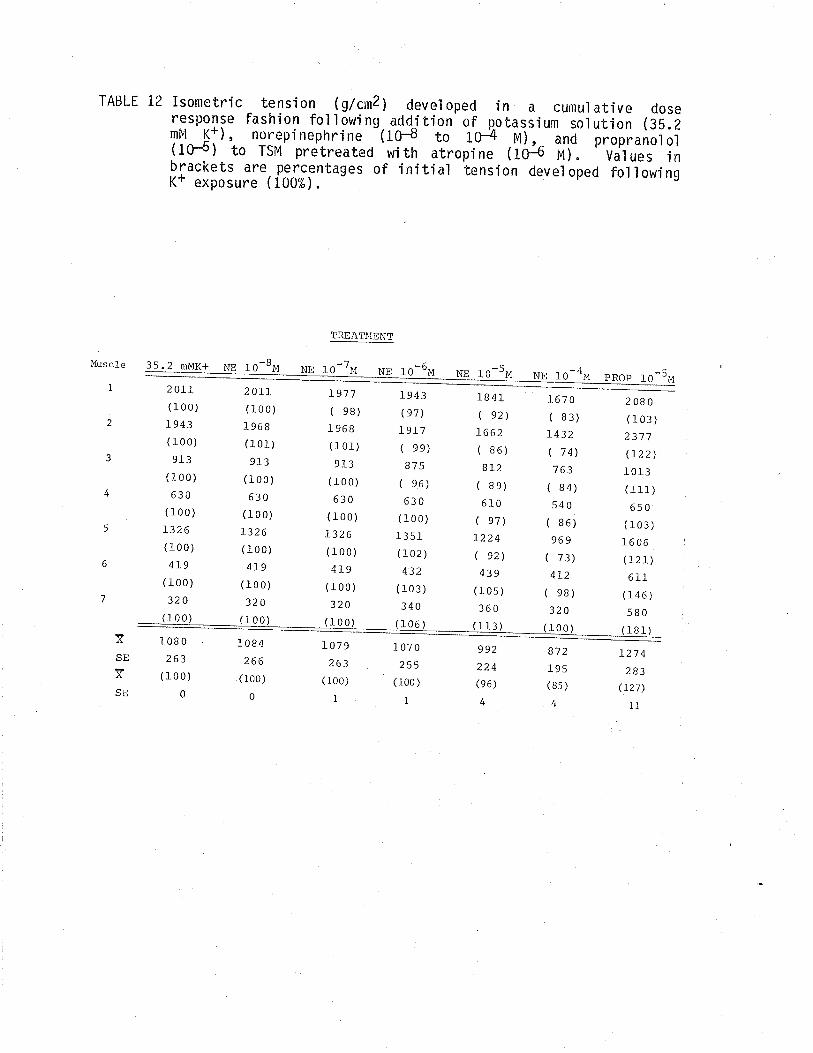
TABLE 12 Isometríc_ tension (g/cn?) developed in a cumulative doseresponse fashíon following. addÍtion of potassium solution (35.ãmM K+) ,. norepi nephrine -(1ù-B to th4 Ml ,-- ;rã"'ilopranot ot(rr-5) to TSM'prdtreared wirh arropìne (i0-6 üi" '''vuru* inbrackets are. _p^ercentages of initial tension devel oped fol l owi ngK+ exposure (iooz).
TREAT}lENT
Muscle 35. 2 m¡fK+ -oNE l0 "M Nrì -a10 'r4 Ne f 0-oM -cNE 10 -M -ÁNE 10 -M -ÊPRop l0 ',."r
a
2OIL(r0o)1943(r00)913
(100)
630(100)
1326(100)
4r9(100)
2OII(100)
196 B
( 101)
913
(100)
630(100)
1326(100)
479
(100)
32A
I97 7
( e8)
196I( 101)
913
(100)
630(100)
]-326
(100)
4I9(r00)320
1943(e] )
T9I7( ee)
( e6)
630(100)
13 51-
(102)
432
(103)
340
l-B4l_
( e2)1662
( B6)
B12
( Be)
610( e7)1224
( e2)
439
(105)
167 0
( 83)
I432( 74)
763
( 84)
540( B6)
969( 73)
4L2
( e8)
320
2 0B 0
(lo3)231 7
(122)
LO 13
(r1r)6s0
(103)
1606
(12r.)
6 l-1
(146)
580
4
5
6
7 320
100
xSE
F
108 0
263(100)
0
100
1084
266
(r00)
0
360
r13 )I00
1079
263( too)
I
I06
107 0
255(100)
I
992
224(e 6)
4
100
872
r9s(8s¡
4
181
I27 4
283(127)
11
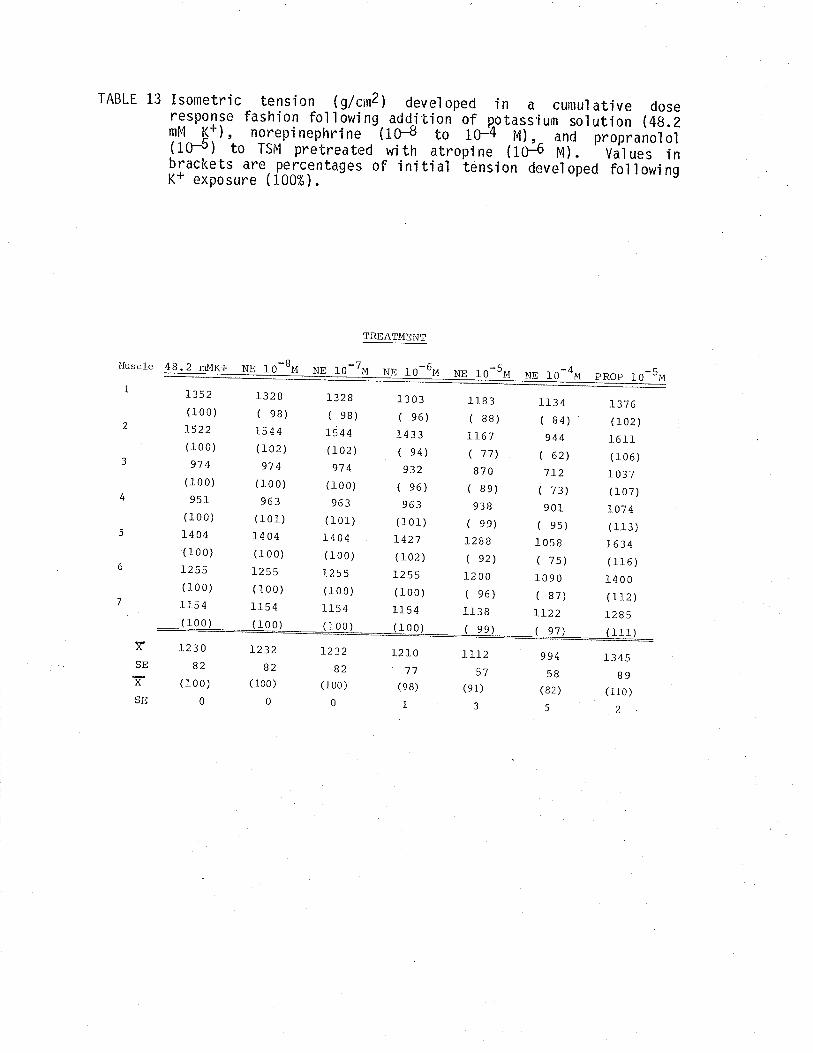
TABLE 13 Isometric _ tension (g/cin?) deve'loped in a cumul ative doseresponse fashion fol I owi ng. ad{i tion of p.otassium sol ution t+A.ãmM K+),. norepinephrine -(1É to 10-4 Ml,_- unã"'iiopranotot( ro-5 ) -to
TSilt' prdtr.àtud wi th uCropl n. (10-+ M ) . val ues i nbrackets are. percentages of initial tension developed fot towlngK+ exposure (IOO/"). -
TREAT}lENT
I'luscle 4B 2 il,rK+ me 1o-8t -aNE l0 't4 -âNE 10 -t"t -qNE l0 "Èt -ÁNE 10 -r.r -trpRop 10'lr1
2
4
I352(100)
I522(100)
974(r00)951
(100)
L404(l-00)
12 55
(100)
115 4
1328
( eB)
1544(102)
974
(100)
963
(10r)14 04
(r00)1255(100)
1t54
L328( eB).;,.IJ44
(102)
91 4
(r00)963
( 101)l-40 4
(100)
L255(100)
115 4
I3 03
( e6)
1433( e4)
932( e6)
963
( 101)I427(r02)1255(100)
LL54
118 3
( 88 )
LI67( 77)870
( 89)
938
( ee)
12BB
( e2)
12 00
( e6 )
113 8
1134( 84)
944( 62)?12
( 73)
901( es¡
105I( 7s)1090( 87)
I122
I37 6
(102)
1611
(r06)103 7
(r07)1074
(1r3)1634
(rÌ6)1400(r12)1285
)
6
lr
cÞ
100
123 0R)
(100)
0
100)
1232o1
( 1oo)
0
100
1232
B2
(100)
0
I00
L2IO
77(e8)
I
99
1112
57
(e t)3
97
994
58
(82)
5
1ll
1345
89(110)
2
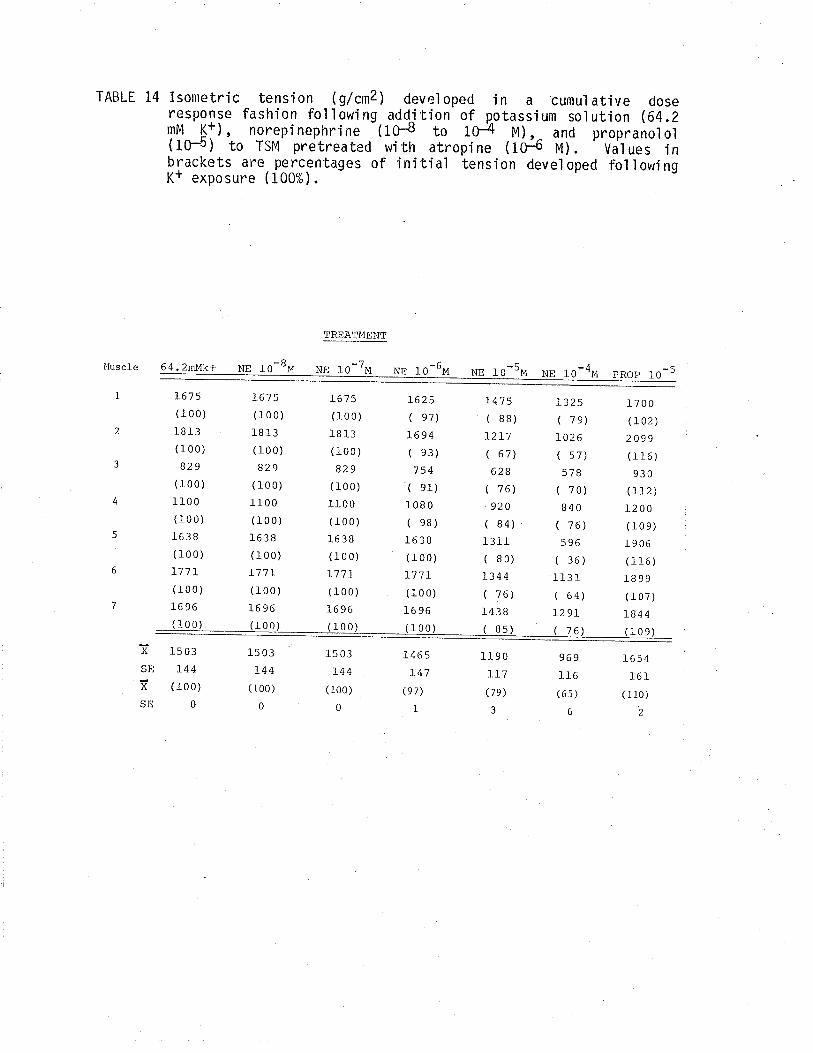
TABLE 14 IsFôt!
mM
(tbrç+
omespo
Kg,-5
ackEX
tric tension (g/cn?) devel oped in a cumul ative dosense fashion following add^ition of potassium solution (64.2+), norepinephrine (1É to lÉ M),- and ¡rropranolol) to TSM pretreated with atropine (10-6 M). 'Values
inets are. percentages of initial tension devel oped fo'll o'ri ngposure (L\O%).
TREATMENT
Þfuscfe 64.2mMk+ ne t0-8 -111 NE IO_a¡'r ue to " -qM NEIO-M -ANe ro '¡l pRop to-t
2
3
l_67 5
(100)
1B I3(100)
829(r00)110 0
( 100)1638
(r00)T77 I(100)
1696(100
L61 5
(ro0)18t3(r00)829
(100)
110 0
(100)1638
(100)
I]7L(r00)1696
L67 5
( 100)
1813
(100)
829(r00)1I0 0
(r00)T6 3B
(100)
T77L
(100)
]-696
I625( et)L69 4
( e3)
754( eI)1080( e8)
1638(100)
I7] I(100)
1696
147 5
( 8B)
12T7
( 67)
628( 76)920
( 84)
1311( I0)\344( 76)1438
L325( 7e)
L026( s7)
578( 70)
840
( 76)596
( 36 )
1l3t( 64)
l2 9l-
1700
(r02)2 099
(116)
930
(112)
1200(10e)
r906(1r6)1899
(r07)I844
4
5
6
7
(r00 (100 (100 ( 8s ( 76 (109
X
SE
XqF
t5 03
144(r00)
0
1503
144(100)
0
1s 03
144
( r0o)
0
l-465
I47(e 7)
I
119 0
IL7(7e)
3
969
1I6(6s)
6
1654
l- 61
(l to)
2
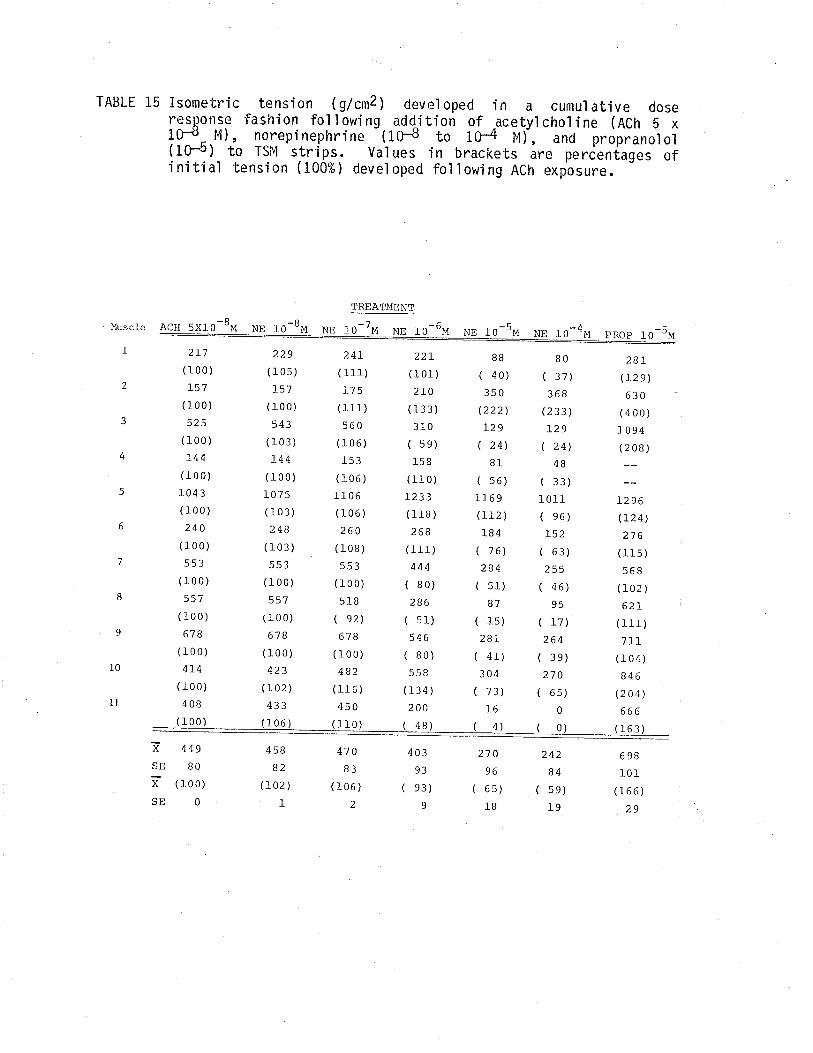
TABLT 15 Isometric tension (g/ctn?) deve]oped in a cumulative doseresponse 'fashion following addition of -acetylcholine (nCfr 5 x10-ö M)., 19repi nephrine - (1N to 1É t4i, and propranol o](to-5) io TSM'strìps- Values in brackets áre pàrJÁntages ofi ni ti al Lensi on ( 100% ) devel oped foì I owi ng ACh exposure.
TREATMENT
lluscle -oecn 5x1o "¡¡ l¡n to-8¡¿ ¡re ro-7¡¡ -^NE IO "M un 1o-5oo Nn to-4 M pnop r o '¡.rI 2L7
(r00)I57
(100)
525(100)
144(100)
I043(100)
240
(100)
553(100)
557
(100)
61 I(r00)414
(r00)408
(100
229
(10s)
I57(100)
543(103)
144(r00)I075(ro3)aÀo
(t03)5s3
(100)
5s7
(roo)678
(100)
423
(102)
433
(106)
24I( r11)L75
( rrl)560
(106)
153
(106)
1106(106)
260
(r08)553
(100)
518( e2)
618
(r00)482
(r16)450
22L( 101)
210(133)
310( 5e)
158
(r10)1233(118 )
¿oÕ
( 1r1)444
( 80)
286
( sr)546
( 80)ÊÉo
(134)
200
88
( 40)
350(222)
129( 24)
BI( s6)116 9
(112)
184
( 16)284( sl)
87
( ls)28T
( 4r)304
( 73)l6
BO
( 37)
368
(233)
129( 24)
48
( 33)
1 0rt( e6)
L52( 63)
255( 46)
95
( L7)
264
( 3e)11^
( 6s)
0
28L(12e)
630
(400)
1094
(208)
2
3
4
5
6
7
1296(L24)
276
(Lls)568
(r02 )
62I(111-)
7LI(104)
846
(20 4)
666
8
9
l0
1l
(110 (48 4)( (r630)
X
SE
X
ù¿
449
BO
(100)
0
458
ô¿
(102)
1
470
B3
(r06)2
403o?
( e3)
9
270
96
( 6s)
18
242
84
( se)
19
698
l0t(166)
29
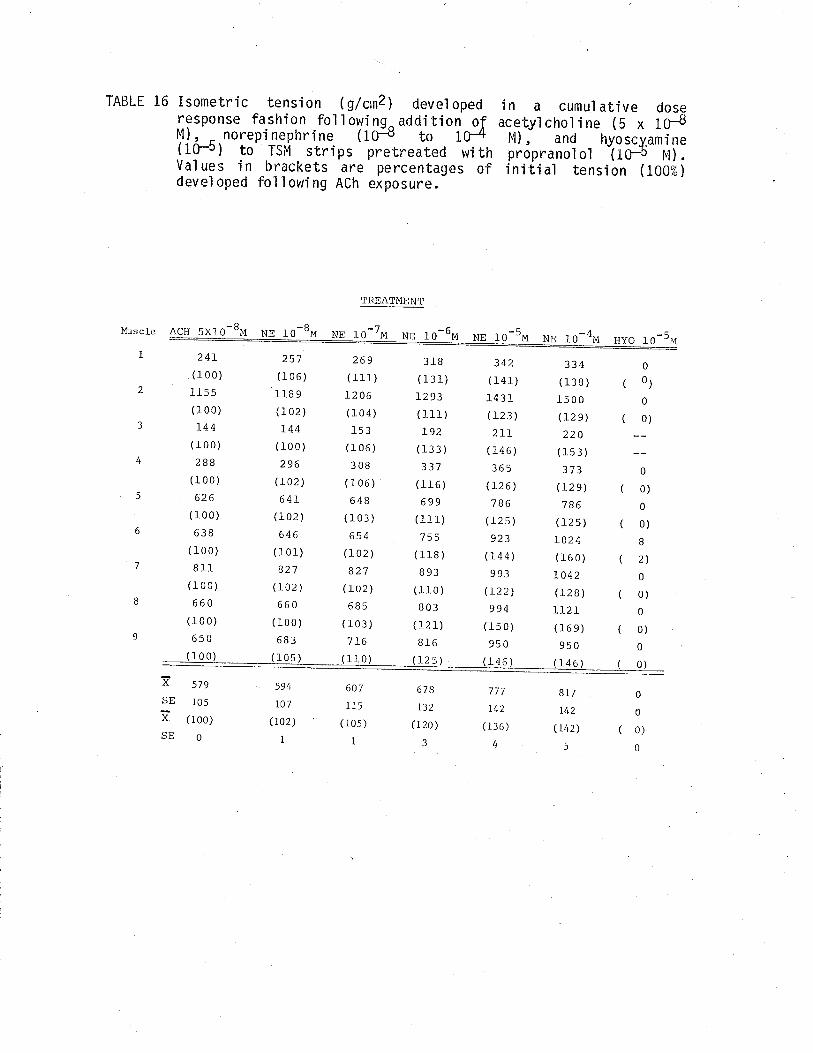
TABLE 16 Isometric tension (g/cn?) deve'lopedresponse fashion fol I orti ng^ addi tion of14), - norepinephrine (1É to LÉ( 1ù-þ ) to TSM stri ps pretreated rvi thVal ues i n brackets are percentages ofdevel oped fol 1 owÍ ng ACh exposure.
l"fuscle ÄCH 5X10 -8M un ro-8r, N¡ ro-7t
in a cumul ative doseacety'lcholÍne (S x 1f€M), and hyoscyamine
propranoì o1 (tO-s M) .i n ì ti al tensi on ( 100% )
Ne 1o-4r"1 Hyo
TREATI'ÍENT
-âNE 10 "M NE -t10 -r'{ 5l-0 M
24r(r00)l-15 s
(100)
l-44(100)
2BB
(100)
626
(100)
638(100)
BlI(100)
660
(r00)650
(100
257
(106)
118 9
(102)
L44(100)
296(r02)64L
(102)
646( 101)827
(r02)660
(r00)683
(10s)
269
(111)
1206(104)
153(106)
308
(106)
648(r03)654
(102)
o¿t
(Lo2)
685
(103)
716( 110)
3l- 8
( 131)L293(111)
l-92(133)
337(1r6)699
(111)
755(118)
893
(r10)803
(121)
o10
(l2s)
342
( r41)14 3l(123)
2\I(146)
36s(126)
786(125)
923(l-44)
993
(L22)
994(rs0)950
(146)
334(r38)1500(L2e)
220(1s3)
???
(129)
786
(12s)
L024(160)
L042(r2B)
I12I(16e)
950
( 0)
0
0)2
3
4
5
6
l
8
9
0
0)
0
0)
B
2)
0
0)
0
0)
0
0)r46)
X
ùE
QF
579
t05( ioo)
0
s94
107
(102)
I
607
115
( 10s)
I
678
132
(r20)
3
771
t42( 136)
4
8t7
142
(r42)
5
0
0
0
0
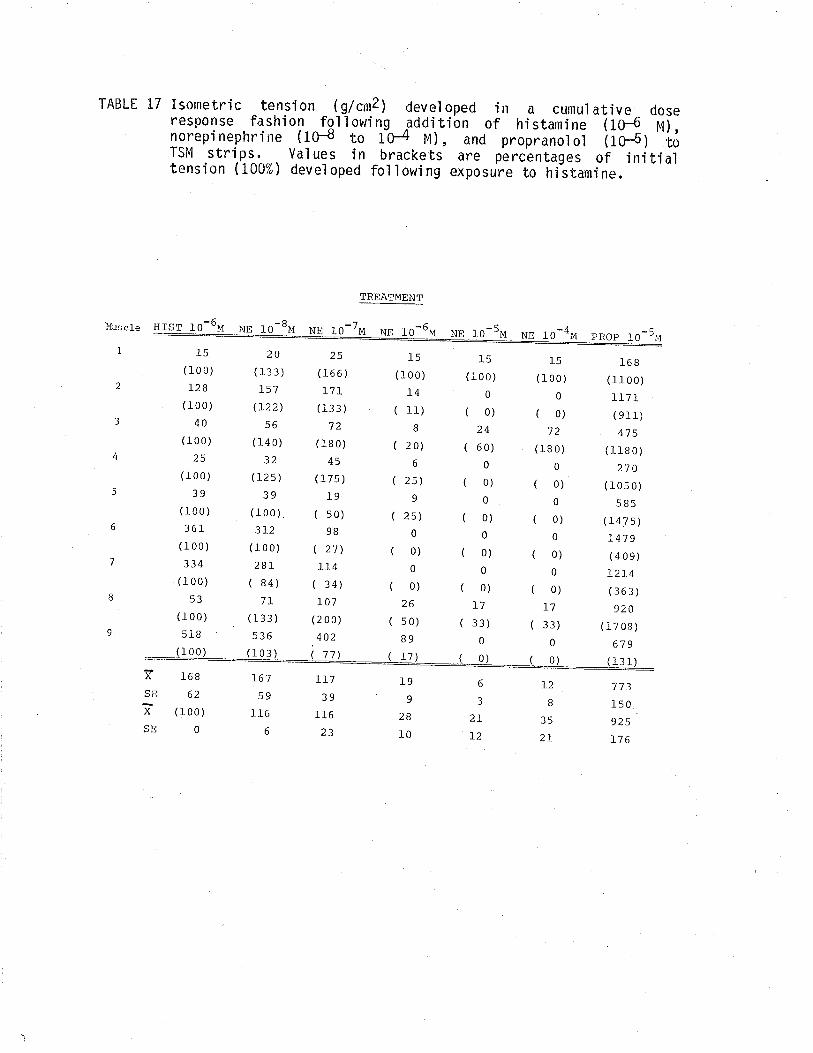
TABLE 17 Isornetric _tension (g/crn?) developed in a cumulative doseresp0nse fashion fg'llowing addition of histarnine (10-6 Mi.norepinephrine (tÉ to iry+ M), and propranotoi ilû-5) "iåTSM stri.ps. val ues in brackets are percentages of Ínitialtension (100%) deveroped foilowing exposure to histamÍne.
Muscle HIST l0 6 -a-M NE Io "M
TREATMENT
¡le ro-7u NE -Ê10 "M NEf o 'It NE PROP 10_À
10 =M 5,,!,1
2
3
4
t5(100)
I28(100)
40
(100)
25
(r00)39
(100)
361(r00)JJ+
(r00)E'
(r00)518
(1oo)
20
(133)
1s7(722)
56
(140)
)¿(12s)
39(100)
3]-2(100)
28L( 84)
7L(133)
536(103)
25
(166)
r7I(133)
72
(180)
45
(17s)19
( so)98
( 27)114
( 34)
I07(200)
,402( 77\
15
(100)
I4( 11)
o
( 2 0)
6
( 25)9
( 25)0
( 0)
t,
15
(1oo)n
(0)24
( 6o)
0
(0)0
( 0)
0
( 0)
0
( 0)
I7( 33)
0
( 0)
I5(100)
0
(0)1)
(180)
0
( 0)
U
( 0)
0
( 0)
0
( 0)
I1( 33)
0
( 0)
168(1100)
r 171( e1t)475
(1180)
270
(10s 0 )
585(r47 s)L47 9
(40e)
12I4(363)
920(1708)
679
5
6
7
8
9
0)
26
50)
89
]-7) (131X 168
62
(100)
0
I6759
1I66
19
9
2B
10
SE
Ä
CF
IT739
116
¿J
6
t
2L
I2
I2B
aÉ
2L
773
1s0
925
I'1 6
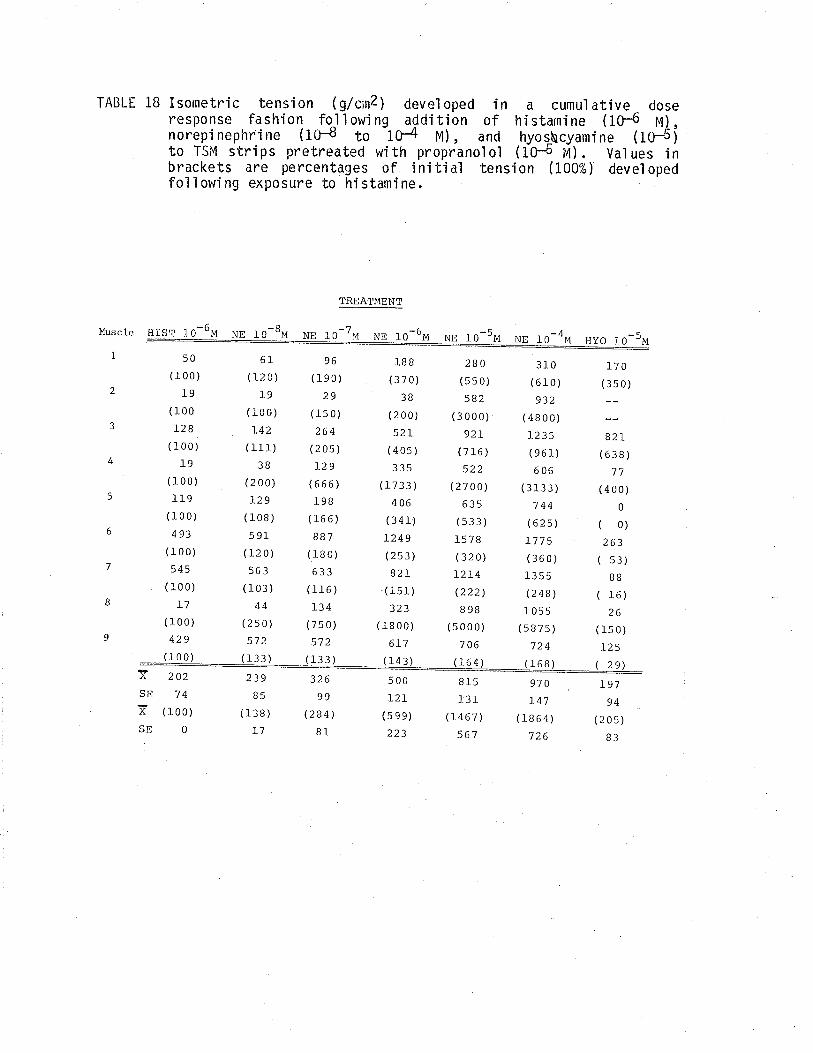
TABLE 18 Isometric tension (g/an?) devel oped Ín a cumul ative doseresponse fashion fo1 1 owi ng addi tion of hi stamine (1É M) .norepinephrine (1É to -iO-4
M) , and hyos_\cyamine (10-5ito TSM strips pretreated with propranolol (10-b M). Values inbrackets are percentages of initial tension ß00%) developedfollowing exposure to histamine.
TREAT}IENT
Muscle ursr 10-6¡¿ Ne ro-BÀ,r Ne ro-7¡¿ NE10"M NEto' -ÁM NE 10 =If Hyo 10 'MI 50
(l-00)
19
(100
l-28(100)
19
(loo)II9
(r00)493
(100)
545
(100)
I7(100)
429
6I(120)
I9(100)
I42( lrr)
?o
(200)
L29(108)
591
(120)
563(r03)
44
(2s0)
572
96
(190)
29
(1s0)
264
(20s)
129(666)
r9B(r66)887
(180)
633(116)
134
(750)
572
188
(370)
3B
(200)
52I(40s)
335
( r733 )
406(341)
1249(253)R)'l
(rs1)1a1
(1800)
6r7
280(5s0)
582(3000)
92L(7 r6t522
(27 0o)
635(s33)
]s78(320)12L4(222)
I9B(s000)
706
310(610)
932(4800)
L235(961)
606(3133)
744
(625)
r775(360)
1355
(2 4B)
105 5
(s87s)
724
L70(3s0)
82I(638)
(400)
0
( 0)
263( s3)
oó
( l6)26
(1s0)
L2s
2
3
4
5
6
7
8
9
Str
X
(100
202
74
(100)
0
(r33 (133 (143
500
I2I(5ee)
¿¿)
164
815
13r(L467 )
561
t68 29X
SE
??o
QC
(138)
T7
970
I47(r864)
726
I9794
(20s)
83
326
99
(284)
81

TABLE 19 Isometric tensjon (g/on2) deveìoped in TSM from dogs (n = 10)sensitized to ovalburnin. Drugs tyramine (TYR 10-5 and 1Élil) and propranol ol (10-5 M) were added curnul at'ively.
TREATMENT
-5 -4Muscle TYR t0 t4 TYR IO M
36
0
0
2I66
107
0
0
0
aa¿z
CI¡
25
11
-5PROP IO ¡,I
9
0
0
0
o
0
0
0
0
0
1
2
J
4
5
6
7
I9
10
63
0
0
B6
99
r090
0
0
23
39
15
2
1

TABLT 20 Isometric tension (g/cn?)mongrel control dogs ( naddi tion of norepi nephri neresul ts were obtaÍned whenpretreated wi th proprano'lol
deve'l oped i n un stimul ated TSM from= . 10 ) , fol l owi ng -curnul ative dose(NE lf to 10+t M) . Simil armuscles in the same protocol were(10-5 M)
"
TREATMENT
Mus c1e
I
2
3
4
5
6
7
8
9
10
-R -'7 -^ -q -LNE TO "¡.I NE IO '¡4 NE 10 "}1 NE IO "M NE 10 'M
o
0
0
0
0
0
0
55
0
0
x0
9
U
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
U
0
0
0
0
0
0
0
0
0
0
U
0
0
0
0
0
60
000
0
SE
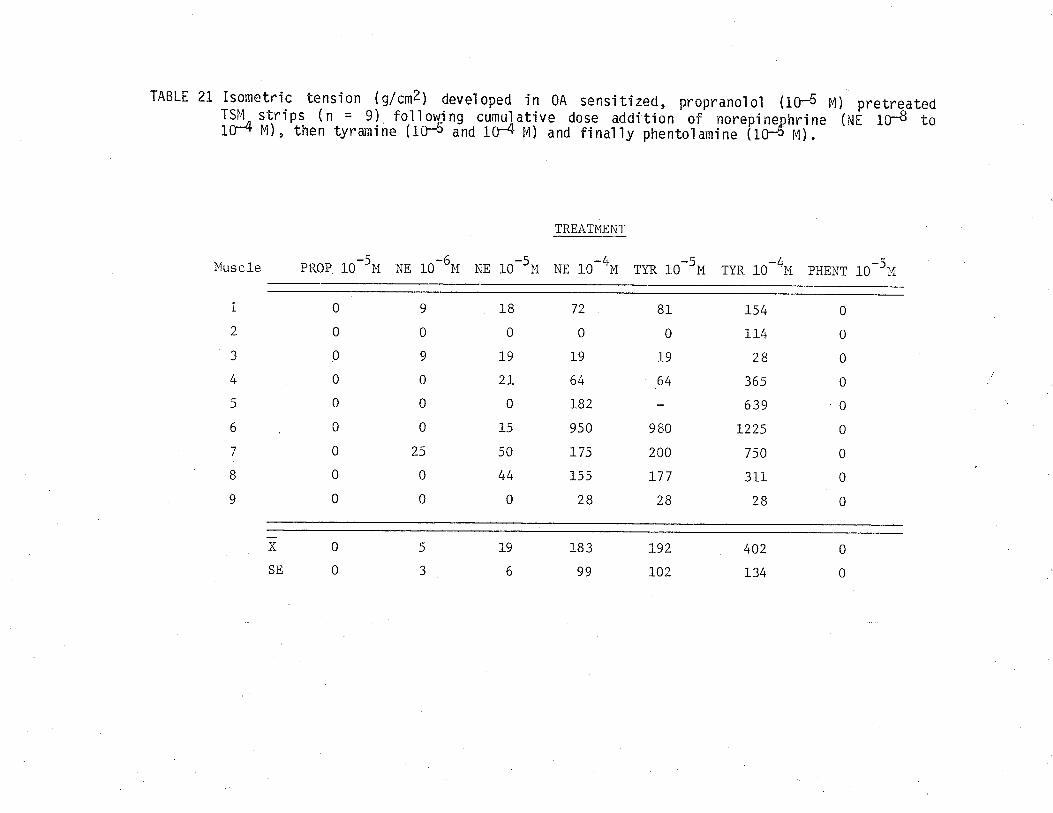
TABLE 21 Isometric t'ension (g/cm?) developed in 0A sensitized, propranolol (tO-s M) pretreatedTSl4 strips (n = 9) fo.l'lowing cumulative dose addition of norepinephrine (NÉ tO-8 toloa M) , 'then tyranr:ne (1É u"n¿ io-+-úi
';rd "riruiiv'ñr'ãlitrì.-iñå-iio-f Nl.
Muscle -qPROP 10 -M
0
0
_ANE IO "M -5
TREATMENT
72
0
19
64
LB2
9s0
L75
155
2B
183
99
NE 10 MNE 10 -M -qTYR 10 -M TYR l0 'M -5PHENT 10 M
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
U
I2
3
4
5
6
7
B
9
9
0
9
0
0
U
25
0
0
Iõ
0
I92L
0
15
50
44
0
l96
B1
0
L9
64
rs4
TL4
2B
365
639
L225
750
31r
2B
402
734
980
200
L77
2B
T 0
0
5
J
L92
L02SE
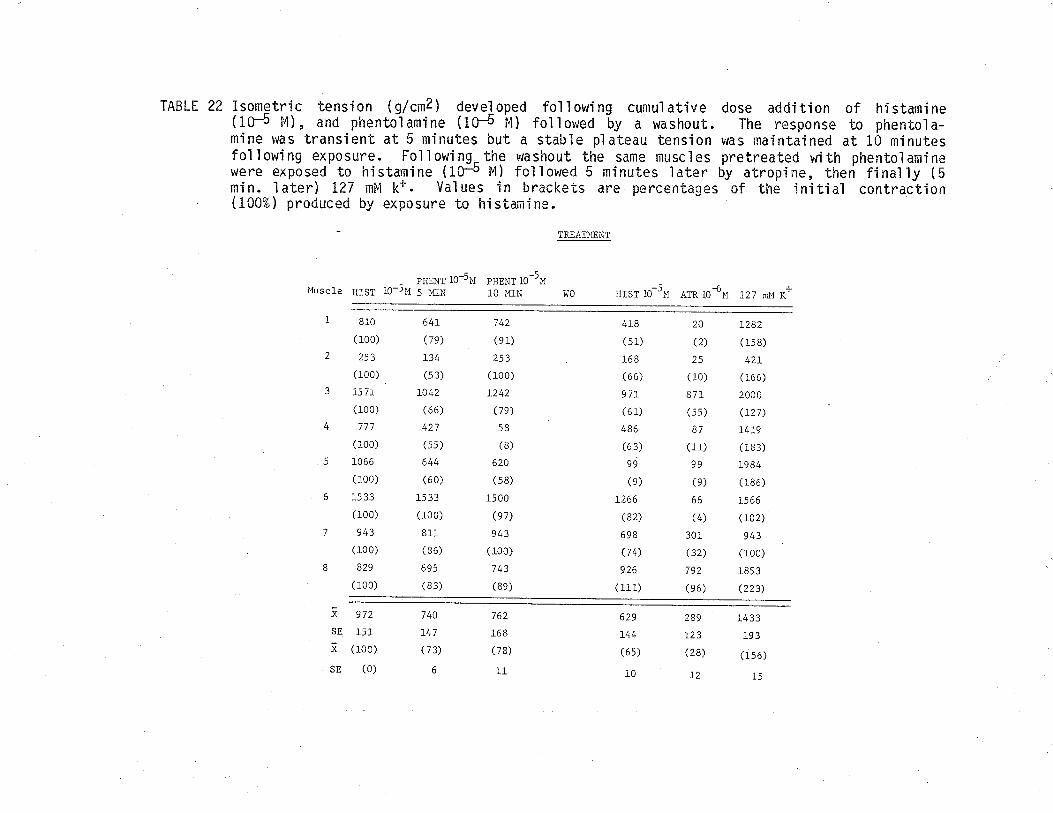
TABLE 22 Isomqtric tension (g/cin?) deve]oped fol'lowi ng cumul ative(1É 14), and phentoiamine (1É iq) followed -tV a washout.mine was transient at 5 minutes but a stable platear.l tensionfol I owi ng exposure. Fol I owi ng- the washout the same muscl eswere exposed to hi stalni ne (10_5 M ) fol I owerl 5 mÍ nutes I atermin. later) I27 mþl k+" Values in brackets are percentages(L09"/") produced by exposure to histamine.
--_qp¡1¡¡1 rU .M
nrsr 1O-5i't 5 ¡nu I^10 ursr 1o 5u or* 1o{" 127 nr¡r K*
dose addi tion of hi starni neThe response to phento'la-
was maintained at 10 minutespretreated wi t,h phentol ami neby atropine, then finally (Sof the initial contractÍon
r282
( ls8)42L
( 166)
2000
(rz7)
14L9
( 183)
19 84
(r86)
1566
(102)
943
( 100)
185 3
(223)
TRXAT}ÍENT
Èluscle-5PHENT 10 'M
10 MIN
742
(e1)
( 100)
(7e)
58
(8)
620
( s8)
15 00
(sz;
943
( t00)
/43
( 8e)
3
4
7
I
5
6
810
( 100)
¿5J
( 100)
T57t( loo)771
(1oo)
1066
( 10o)
1s 33
( 100)
943
( 100)
829
( 1oo)
64L
(7e)
134
(s 3)
r042
(66)
427
(ss )
644
(60)
15 33
( 100)
811
( 86)
695
(83)
4r8
(s1)
168
(66)o11
( 61)
486
(6 3)
gg
(e)
7266
(82)
698
(7 4)
oaa
(111)
20
(2)
t(
(10)
871
( ss)
87
( 1l)99
(e)
66
(4)
301
(32)
7 A',)
(e6)
xSE
iSE
972
151
(100)
(0)
740
147
(7 3)
6
762
168
( 78)
11
A)O
144
(65)
10
tao
rl3( 28)
12
L+JJ
r-9 3
( rs6)
i5
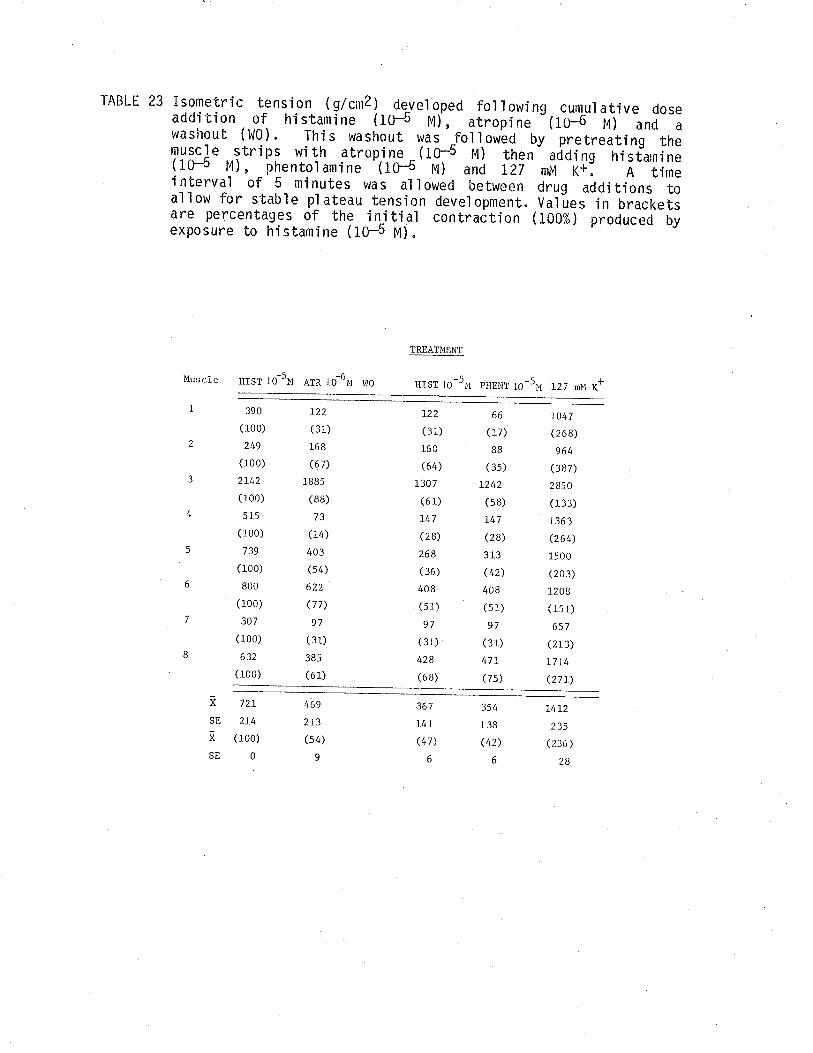
TABLE 23 Isometríc t^en1i9n (g/cn?)_ ^de.vel
oped fo.l.lowing cum-ul ative doseaddition of histamine (10-5 M), atropine (10-6- M) ãnJ--ãwashout (l.l0). This washout was -followä¿ by pretreating themuscle .strips with atrop'i_n_e.(10.5 M) then aå¿ing -histarnine
(1û-5 M), _þhentolamirié-iro-s'-ùl uüá n7 mM K+. A rírnei ttterval of 5 mi nutes was al I owed between drug addi.bions toallow for s'lable p]ateau tension development. Values Ín bracketsare percentages of the initial contraction (100%) produced byexposure to histainine (tf-5 Vi).
TREATI.IENT
Muscle Hrsr to 5u ¿,rR to 6lt
tr{o Htsr lo-5¡t "HE*,
to-5t, 127 o,¡l rc+
I
2
3
4
390
( 100)
)Lq
( 100)
2L42
(100)
515
(100)
739
( r00)
800
(100)
307
( loo)
632
( 10o)
122
( 3r)168
(67)
1885
( 88)
IJ
(14)
403
(54)
622
(77)
97
( 31)
385
(6r)
722
( 31)
160
(64)
r30 7
(6 r)747
(28)
268
( 36)
408
(s 1)
97
( 31)
428
(68)
66
( rz¡ÕÕ
(3s)
1242
(ss¡
L47
(28)
313
(42)
408
(s 1)
97
( 31)
47r(7s)
r047
(268)
964
(387)
2850
( 133)
136 3
(264)
1500
(203)
1208
(ls1)
657
( 213)
L7 14
(27 r)
5
6
7
8
X
SE
ç
SE
727
2L4
( 1oo)
0
361
14I(47)
6
469
2L3
(s 4)
9
3541 AO
(42)
6
14t2
235
(236)
28
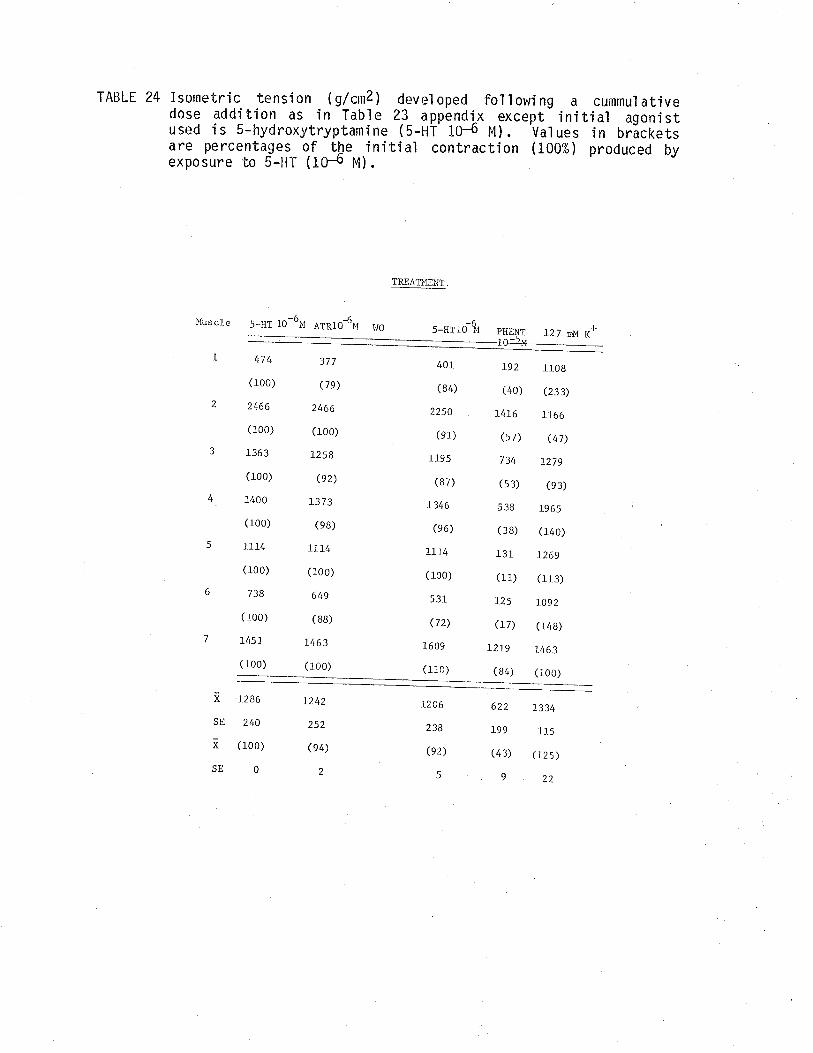
TABLE 24 Isornetric tension (glcn?) devel oped fol I owi ng a cummul ativedose addition as in Table 23 appendix except initial agonistused Ís 5-hydroxytryptamine (5-HT lÉ M). 'Values in brãcketsare percenta_ges o.{ tþe initial contraction (L00%) produced byexposure to 5-HT (l0á M).
TREATMENT
Ifus cle 5-trt to-6¡t ar*to{0, I{O s-nrto Tr 127 ûìM K'
2
474
( 100)
2466
( roo;
136 3
(100)
_r.400
(100)
1114
(100)
738
( loo)
145 1
( 100)
377
r 7qì
2466
(100)
7258
(ez)
I37 3
( e8)
11 14
( r00)
649
( 88)
7463
( r00)
401
( 84)
2250
(el)
119 5
(87)
7346
(e6 )
II74
( 10o)
JJA
(7 2)
1609
( 110)
192
(40)
14r6
(st¡
734
(s3)
538
(38)
131
( 11)
11c
(17)
T219
( 84)
1 108
(233)
1,L66
(qt)
r279
o3)
1965
(140)
r269
(lt3)
I092
(148)
1463
(100)
3
4
5
6
x 7286
SE 24O
I (roo)
SEO
1242
(e4)
2
\206
(e2)
5
622
199
( 43)
9
1 'ì'ì¿
115
(12s)
ta
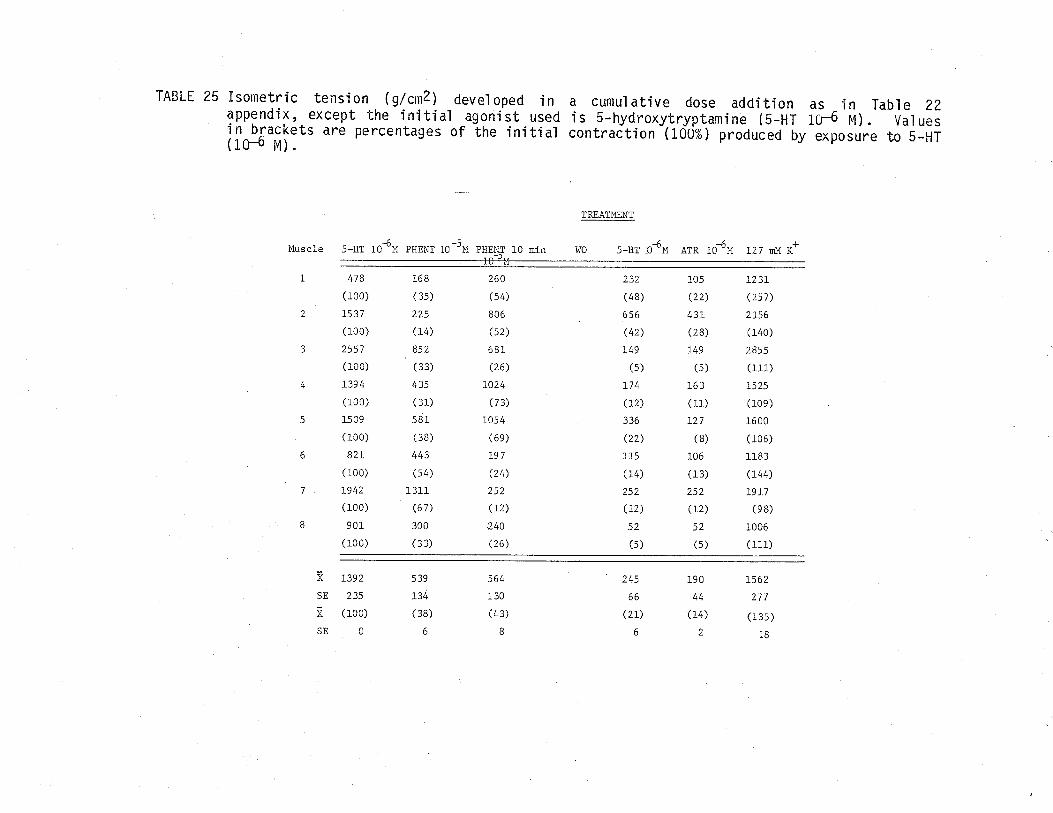
TABLE 25 I sornetric tensi on (g/ ctn?) devel oped i ngppendix, except the Ínitial agonist usedin brackets are percentages of tfie tnitial(1É M).
a cumul ative dose addi tÍon as i n Tabl e ??i s 5-hydroxytryptami ne (S-HT tCr6 N ) . Val uescontraction (1007,) produced by exposure to 5-HT
TREATMENT
Muscle 5-", to{* pusNr IO-5tt 10 nÍn I4rO 5-HT-4n-M -AL¿TR io "r.r 127 mM K'
2
4tó
( 100)
L537
( 100)
2557
( 100)
r39 4
( 100)
15 09
( 10o)
827
( 100)
1942
( 100)
901
(100)
x L392
SE 235
Í (:-oo)
SEO
168
(3s)
225
( 14)
852
( 33)
435
( 31)
581
( 38)
443
(s4)
1 311
(67)
300
(33)
260
(54)
806
(s2)
68r(26)
L024
(73)
105 4
( 6e)
797
(24)
252
( 12)
240
(26)
1237
(2s7)
2156
( 140)
2855
(11r)
1525
( r0e)
16 00
( r06)
1183
(r44)
1917
(e8)
1006
(11r)
1562111
(13s)
t8
232
( 48)
6s6
(42)
L49
(5)
r74(L2)
336
(22)
1r.5
( 14)
252
(tz¡52
(s)
105
(22)
qJI
(28)
r49
(s)
163
(u)127
(8)
106
(13)
252
(L2)
(s)
2
4
5
6
7
I
539
l,34
( 38)
6
564
l_ 30
(43)
o
66
( 21)
6
190
44
( 14)
2




















