
The summary on non-reactivation cholinergic properties of oxime reactivators: the interaction with...
11
1 23 Archives of Toxicology ISSN 0340-5761 Arch Toxicol DOI 10.1007/s00204-012-0977-1 The summary on non-reactivation cholinergic properties of oxime reactivators: the interaction with muscarinic and nicotinic receptors O. Soukup, D. Jun, G. Tobin & K. Kuca
Transcript of The summary on non-reactivation cholinergic properties of oxime reactivators: the interaction with...

1 23
Archives of Toxicology ISSN 0340-5761 Arch ToxicolDOI 10.1007/s00204-012-0977-1
The summary on non-reactivationcholinergic properties of oxime reactivators:the interaction with muscarinic andnicotinic receptors
O. Soukup, D. Jun, G. Tobin & K. Kuca

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

MOLECULAR TOXICOLOGY
The summary on non-reactivation cholinergic properties of oximereactivators: the interaction with muscarinic and nicotinicreceptors
O. Soukup • D. Jun • G. Tobin • K. Kuca
Received: 31 March 2012 / Accepted: 7 November 2012
� Springer-Verlag Berlin Heidelberg 2012
Abstract Organophosphorus inhibitors (OP) of acetyl-
cholinesterase (AChE) represent a group of highly toxic
compounds. The treatment of OP intoxication is, however,
insufficiently ensured. Currently, two main categories of
drugs—anticholinergics and oxime reactivators— are
employed as antidotes. Oximes have been reported to act at
several levels of the cholinergic transmission, and among
the non-reactivation effects, the interaction with choliner-
gic receptors stands out. This review addresses issues
correlated with non-reactivating effects of oxime reacti-
vators with a special focus on the muscarinic and nicotinic
receptors, but involvement of other cholinergic structures
such as AChE and choline uptake carriers are discussed
too. It can be concluded that the oxime reactivators show a
variation in their antagonistic effect on the muscarinic and
nicotinic receptors, which is likely to be of significance in
the treatment of OP poisoning. In vitro data reported oxi-
mes to exert higher efficacy on the muscarinic M2 subtype
than on the AChE. However, this effect seemed to be
subtype specific since the antagonistic M3 effect was
lower. Also, and importantly, the antimuscarinic effect was
larger than that on nicotinic receptors. Even though atro-
pine showed a much higher muscarinic antagonism, it is
supposed that non-reactivation properties of oxime reacti-
vators play a significant role in the treatment of OP
poisoning.
Keywords Oxime reactivators � Organophosphate �Non-reactivation �Muscarinic receptor � Nicotinic receptor �Anticholinergics
Introduction
Organophosphorus inhibitors (organophosphates, OP) of
acetylcholinesterase (AChE) are widely used as pesticides,
and its potential use as chemical weapons is a matter of
high concern. These compounds bind to the AChE at the
serine hydroxyl group at its active site, and the binding
inhibits the enzyme’s physiological role in the organism—
to cleave the neuromediator acetylcholine (ACh) when
occurring in the synaptic clefts. Consequently, ACh accu-
mulates in the cholinergic synaptic junctions and the poi-
soning manifests as a cholinergic syndrome. In cases of
insufficient treatment, death is caused by paralysis of the
respiratory muscles and of the respiratory center (Marrs
1993; Bajgar 2004).
The treatment of the OP poisoning is ensured by two
main categories of drugs. (1) Anticholinergics, for exam-
ple, atropine, are able to antagonize the effects of excessive
ACh by a blockade of mAChRs. Other drugs, for example,
benactyzine or scopolamine might be used due to their
better penetration to CNS. (2) Compounds belonging to the
other group, reactivators of the AChE—oximes—are able
O. Soukup � D. Jun � K. Kuca (&)
Biomedical Research Center, University Hospital of Hradec
Kralove, Sokolska 581, 50005 Hradec Kralove, Czech Republic
e-mail: [email protected]
O. Soukup
Department of Public Health, University of Defence, Trebesska
1575, 50001 Hradec Kralove, Czech Republic
D. Jun � K. Kuca
Faculty of Military Health Sciences, Center of Advanced
Studies, University of Defence, Trebesska 1575, 50001 Hradec
Kralove, Czech Republic
G. Tobin
Department of Pharmacology, Sahlgrenska Academy, University
of Goteborg, Medicinaregatan 13, 405 30 Goteborg, Sweden
123
Arch Toxicol
DOI 10.1007/s00204-012-0977-1
Author's personal copy

to restore the physiological function of inhibited AChE. In
addition to them, diazepam or other benzodiazepines can
be used as anticonvulsants. Anticholinergics and reactiva-
tors are usually co-administered because of their syner-
gistic effect (Kassa 2002; Bajgar 2004). Today, HI-6 and
obidoxime are the most commonly used oxime reactivators
in the treatment of OP poisoning; however, the reactivators
differ in their efficacy against individual nerve agents and
no universal antidote has been developed yet. Moreover,
the problem of phenomenon named ‘‘aging’’ (a time-
dependent loss of AChE ability to be reactivated) is an
important issue, since the therapeutic intervention must be
applied within a limited period time. Owing to this fact,
new, versatile AChE reactivators are thus required, or
alternative treatment approaches have to be developed.
Such an alternative approach presumes other mecha-
nisms of action of oximes, which are not related to the
reactivation. Oximes have been reported to act at several
levels of the cholinergic transmission, including the syn-
thesis, release, inactivation and reuptake of the transmitter,
but the interaction with muscarinic (mAChRs) and nico-
tinic (nAChRs) receptors seems to be the most plausible
alternative of mechanism of action (Vanhelden et al. 1996;
Soukup et al. 2010b).
This review deals mainly with the interaction of oxime
reactivators with muscarinic and nicotinic receptors, but
some other cholinergic properties were considered as well.
Particular focus is addressed to differences in efficacy,
anticholinergic potency of individual reactivators and the
correlation between in vitro and in vivo effects. To start
with, a review of literature concerning these effects of
interest is listed below.
Pharmacological effect of oximes
The major mechanism of action of the antidotes is the
reactivation of inhibited AChE. However, other mecha-
nisms are probably involved. The data derived from tabun
or soman lethally intoxicated primates suggests that, due to
a protocol ensuring low or no reactivation, other mecha-
nisms, which lead to survival via the recovery of neuronal
transmission, must be involved (Hamilton and Lundy 1989;
Vanhelden et al. 1992). Similar life-saving results were
obtained in a rat model, where rapidly ‘‘aging’’ crotylsarin
was used, and administration of HI-6 caused the recovery
of neuronal transmission (Busker et al. 1991).
The exact pharmacological effects of oximes are still an
open question. Oximes probably play a role at various
levels of the cholinergic transmission. Thus, the oximes
may exert effects by interactions with pre and postsynaptic
receptors or with the reuptake of transmitters, in addition to
possible effects by affecting the synthesis and release of
ACh. Clement reported that oxime HS-6 inhibits the syn-
thesis of ACh, its release and the reuptake of choline in
chicken (Clement 1979). The release of ACh from pre-
synaptic fibers can be modulated by oximes, suggesting an
interaction with presynaptic muscarinic (Kloog and Soko-
lovsky 1985) and/or nAChRs (Fossier et al. 1990) or via
the interaction with the high-affinity choline uptake carrier
(HACU), which is a key regulatory system in the synthesis
of ACh (Kristofikova et al. 2003). In the latter case, the
interaction with presynaptic mAChRs was also suggested
(Zahniser and Doolen 2001). Controversially, while a
decrease in the release have been reported in the rat brain
(Kloog et al. 1986), an increase has been reported in the rat
striated muscle (Melchers and Vanderlaaken 1991). Similar
contradictive data are reported by several authors, which
seem to depend on different experimental conditions
(Oydvin et al. 2005). Notably, some compounds are
effective against OP poisoning even when the compound
lacks the oxime moiety (Inns and Leadbeater 1983; Loke
et al. 2002; Seeger et al. 2012).
Antinicotinic action
Bis-pyridinium compounds in general were reported to
cause a recovery of neuromuscular transmission mediated
by nicotinic receptors in OP poisoning (Wolthuis et al.
1981). The effect of oximes at the neuromuscular junctions
on nAChRs has been examined in a phrenic nerve-dia-
phragm model (Busker et al. 1991). They observed that
HI-6 was able to counteract the failure of neuromuscular
transmission caused by crotylsarin. Since HI-6 had a sim-
ilar effect as pure cholinergic receptor antagonists (galla-
mine, mecamylamine), which are not involved in AChE
reactivation, a nicotinic antagonism of this compound was
proposed. Furthermore, HI-6 has been shown to be able to
counteract crotylsarin-induced excessive release of 3H-
ACh from an isolated endplate (Vanhelden et al. 1996). On
the contrary, under different experimental conditions,
enhancement of the release was observed as well (Aas
1996; Oydvin et al. 2005). Furthermore, the pharmaco-
logical action against soman poisoning was also due to the
channel blocking activity of HI-6, obidoxime and similar
compounds (Tattersall 1993). However, contrasting to
these studies, single channel studies have also revealed that
HI-6 and pralidoxime increases the opening probability of
nAChRs that are activated by ACh (Alkondon et al. 1988).
nAChRs (and muscarinic as well) in the rat brain were
blocked by some bis-pyridinium compounds as judged by
displacement examinations with tritiated QNB and a-bun-
garotoxin (Broomfield et al. 1987). Ganglion-blocking
properties have been reported by Lundy et al. (Lundy and
Tremblay 1979). Recently, interaction of bys-pyridinium
compounds derived from SAD-128 with orthosteric
Arch Toxicol
123
Author's personal copy

binding side of human a7 nAChR has been showed Nies-
sen et al. (2011).
Antimuscarinic action
Concerning the antimuscarinic effects, binding studies of
some H-oximes have been performed on mice smooth
muscle tissue in vitro (Kuhnen-Clausen et al. 1983). The
inhibitory constants were determined to occur at the
micromolar range. However, observed nicotinic inhibitory
potency did not correlate with the efficacy against soman
in vivo (Kuhnen-Clausen et al. 1983). Functional musca-
rinic antagonism of some reactivators has been observed at
guinea pig (Kuhnen-Clausen 1970; Fusek and Patocka
1976; Kuhnen-Clausen et al. 1983). The mechanism of
binding to the mAChRs is still not fully understood, but an
allosteric manner has been suggested (Lee and Elfakahany
1991; Ellis and Seidenberg 1992; Tucek and Proska 1995;
Kostenis et al. 1996; Christopoulos et al. 1998). Moreover,
in vitro functional examination showed the oximes HGG-
12 and HGG-42 to be allosteric inhibitors (Kloog and
Sokolovsky 1985).
AntiAChE action
The stimulatory effect on cholinergic system by oximes,
which has been described by several authors, should be
more likely ascribed to a partial inhibition of AChE by the
oxime rather than to a direct agonism at muscarinic ACh
receptors (Reithmann et al. 1991). The antiAChE activity
of bis-pyridinium compounds has also been observed when
measured physiologically in the guinea pig ileum (Amitai
et al. 1980) or spectrophotometrically in vitro (Calic et al.
2006). Thus, it can be concluded that pharmacological
effects of oximes are complex and not yet well defined.
There is a hypothesis that oxime reactivators used in the
treatment of OP poisoning exert both antimuscarinic and
antinicotinic properties. However, many questions still
need to be answered. Are the non-reactivation properties of
oximes significant for the treatment of OP poisoning? Do
differences among reactivators in anticholinergic efficacy
exist? Why is there a difference between in vitro and
in vivo effects of the oximes? This review tries to sum-
maries findings that may put some light on these questions.
Our observations
Antimuscarinic and antiAChE findings
According to the literature and according to our observa-
tions, oxime reactivators have very complex mechanisms
of action and influence the cholinergic nervous systems at
various levels. An antagonistic effect on both muscarinic
and nicotinic receptors has been observed. However, cho-
linomimetic action of these drugs cannot be definitely
excluded. As mentioned above, an inhibitory effect on
AChE results in this type of action (Busker et al. 1991).
Ellman’s method, characterizing the potency of oxime re-
activators to inhibit AChE (Table 1) revealed donepezil (a
classical AChE inhibitor used in the treatment of Alzhei-
mer’s disease) to possess approximately 10,000 times
higher inhibition potency than HLo-7 and HI-6—the most
potent classical reactivators. However, in case of OP poi-
soning, the (repeated) dosage of HI-6 may result in
approximately 10 lM concentrations in the human body
(Kassa 2002), so the IC50 of 70 lM describing the potency
of HI-6 to inhibit AChE may cause a significant cholino-
mimetic effect at this compound.
However, cholinolytic effects are the most frequently
reported (sometimes both cholinolytic and cholinomimetic
effects were observed). Displacing experiments using (3H)-
QNB revealed an affinity of oximes to the mAChRs. The
affinity increases in following order HI-6 \ K203 &K027 \ Obidoxime (Table 2). However, the correlation
with a muscarinic standard antagonist—atropine—showed
an approximately threefold lower affinity of obidoxime, the
most potent compound tested (Soukup et al. 2011b). This
finding is in agreement with most other similar studies
(Kuhnen-Clausen et al. 1983). Binding to the mAChRs is a
prerequisite for the transduction of the signal through the
receptor, which, in the case of G protein coupled receptors
(GPCR), leads to the activation of the G protein and the
subsequent intracellular cascade. In the GPCR activation–
based experiment, the oximes antagonize the activation
induced by oxotremorine (muscarinic agonist; control). No
intrinsic activity of the oximes was observed, so the direct
cholinomimetic type of action was excluded (Soukup et al.
2010c). The inhibitory properties of the signal transduc-
tion increased in the following order: HI-6 \ K027
Table 1 Inhibitory potency of standard and newly in house-synthesized oxime reactivators
Compound Hlo-7 HI-6 Pralidoxime Trimedoxime Obidoxime Methoxime K113 K114 K112 K203 K027 Done pezil
IC50 (lM) 67 70 198 287 434 538 10 27 44 311 391 0.007
Error (± lM) 13 6 43 10 71 89 5 12 5 16 23 N/A
Values are expressed as IC50 ± error. IC50 for donepezil, classical AChE inhibitor was obtained from a literature (Ogura et al. 2000)
Arch Toxicol
123
Author's personal copy
\ K203 & Obidoxime. This roughly corresponds to the
binding affinities at least in that HI-6 has the smallest and
obidoxime has the largest pronounced effect. Similarly to
AChE inhibition, IC50 values in units or tens of micro-
moles, with regard to achievable concentrations in a human
body, may testify of an important antimuscarinic compo-
nent in the reactivator’s complex mechanism of action. The
correlation of anticholinesterase activity with the affinity to
the mAChRs was shown to be of inversely related (studied
for 4 reactivators (Soukup et al. 2010c). However, in
another report 7 of 8 OP inhibitors showed a direct cor-
relation between affinity to AChE and to the muscarinic
receptors (Ward et al. 1993). Therefore, it is hard to con-
clude whether a general relationship between AChE
inhibitors and their effect on mAChRs exists.
Organ bath and in vivo experiments
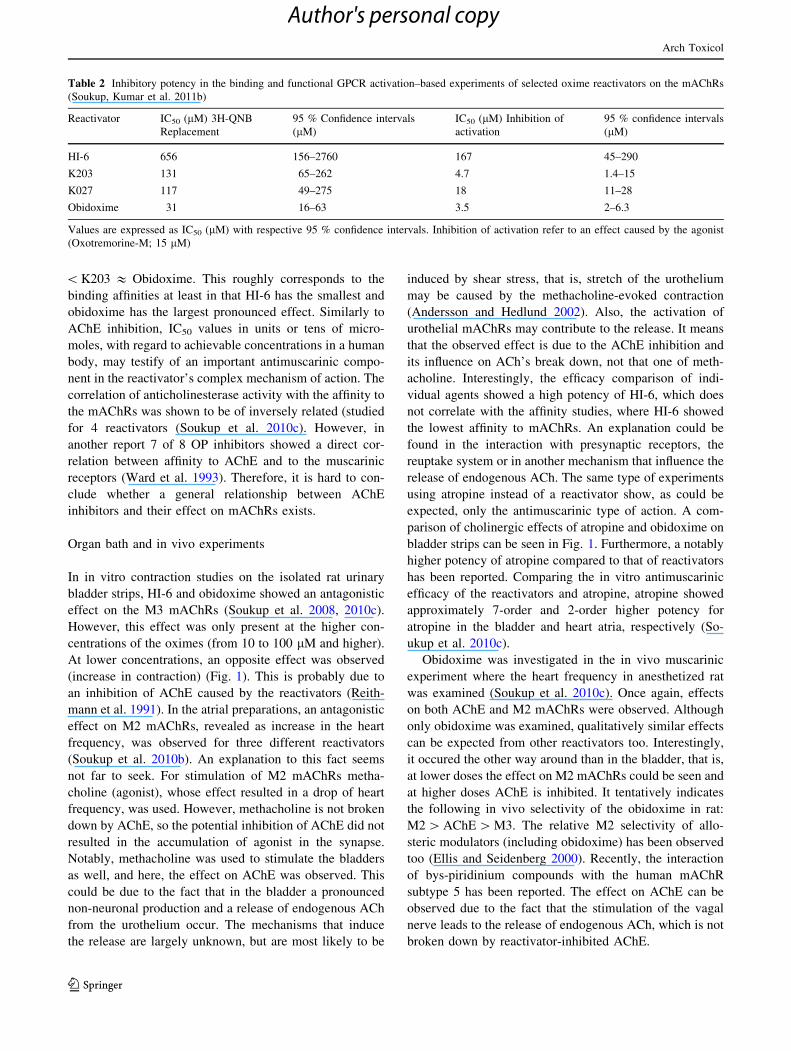
In in vitro contraction studies on the isolated rat urinary
bladder strips, HI-6 and obidoxime showed an antagonistic
effect on the M3 mAChRs (Soukup et al. 2008, 2010c).
However, this effect was only present at the higher con-
centrations of the oximes (from 10 to 100 lM and higher).
At lower concentrations, an opposite effect was observed
(increase in contraction) (Fig. 1). This is probably due to
an inhibition of AChE caused by the reactivators (Reith-
mann et al. 1991). In the atrial preparations, an antagonistic
effect on M2 mAChRs, revealed as increase in the heart
frequency, was observed for three different reactivators
(Soukup et al. 2010b). An explanation to this fact seems
not far to seek. For stimulation of M2 mAChRs metha-
choline (agonist), whose effect resulted in a drop of heart
frequency, was used. However, methacholine is not broken
down by AChE, so the potential inhibition of AChE did not
resulted in the accumulation of agonist in the synapse.
Notably, methacholine was used to stimulate the bladders
as well, and here, the effect on AChE was observed. This
could be due to the fact that in the bladder a pronounced
non-neuronal production and a release of endogenous ACh
from the urothelium occur. The mechanisms that induce
the release are largely unknown, but are most likely to be
induced by shear stress, that is, stretch of the urothelium
may be caused by the methacholine-evoked contraction
(Andersson and Hedlund 2002). Also, the activation of
urothelial mAChRs may contribute to the release. It means
that the observed effect is due to the AChE inhibition and
its influence on ACh’s break down, not that one of meth-
acholine. Interestingly, the efficacy comparison of indi-
vidual agents showed a high potency of HI-6, which does
not correlate with the affinity studies, where HI-6 showed
the lowest affinity to mAChRs. An explanation could be
found in the interaction with presynaptic receptors, the
reuptake system or in another mechanism that influence the
release of endogenous ACh. The same type of experiments
using atropine instead of a reactivator show, as could be
expected, only the antimuscarinic type of action. A com-
parison of cholinergic effects of atropine and obidoxime on
bladder strips can be seen in Fig. 1. Furthermore, a notably
higher potency of atropine compared to that of reactivators
has been reported. Comparing the in vitro antimuscarinic
efficacy of the reactivators and atropine, atropine showed
approximately 7-order and 2-order higher potency for
atropine in the bladder and heart atria, respectively (So-
ukup et al. 2010c).
Obidoxime was investigated in the in vivo muscarinic
experiment where the heart frequency in anesthetized rat
was examined (Soukup et al. 2010c). Once again, effects
on both AChE and M2 mAChRs were observed. Although
only obidoxime was examined, qualitatively similar effects
can be expected from other reactivators too. Interestingly,
it occured the other way around than in the bladder, that is,
at lower doses the effect on M2 mAChRs could be seen and
at higher doses AChE is inhibited. It tentatively indicates
the following in vivo selectivity of the obidoxime in rat:
M2 [ AChE [ M3. The relative M2 selectivity of allo-
steric modulators (including obidoxime) has been observed
too (Ellis and Seidenberg 2000). Recently, the interaction
of bys-piridinium compounds with the human mAChR
subtype 5 has been reported. The effect on AChE can be
observed due to the fact that the stimulation of the vagal
nerve leads to the release of endogenous ACh, which is not
broken down by reactivator-inhibited AChE.
Table 2 Inhibitory potency in the binding and functional GPCR activation–based experiments of selected oxime reactivators on the mAChRs
(Soukup, Kumar et al. 2011b)
Reactivator IC50 (lM) 3H-QNB
Replacement
95 % Confidence intervals
(lM)
IC50 (lM) Inhibition of
activation
95 % confidence intervals
(lM)
HI-6 656 156–2760 167 45–290
K203 131 65–262 4.7 1.4–15
K027 117 49–275 18 11–28
Obidoxime 31 16–63 3.5 2–6.3
Values are expressed as IC50 (lM) with respective 95 % confidence intervals. Inhibition of activation refer to an effect caused by the agonist
(Oxotremorine-M; 15 lM)
Arch Toxicol
123
Author's personal copy
Antinicotinic findings: patch clamp and in vivo
The interaction of oximes with nAChRs has been investi-
gated too (Soukup et al. 2011a). In vivo observations
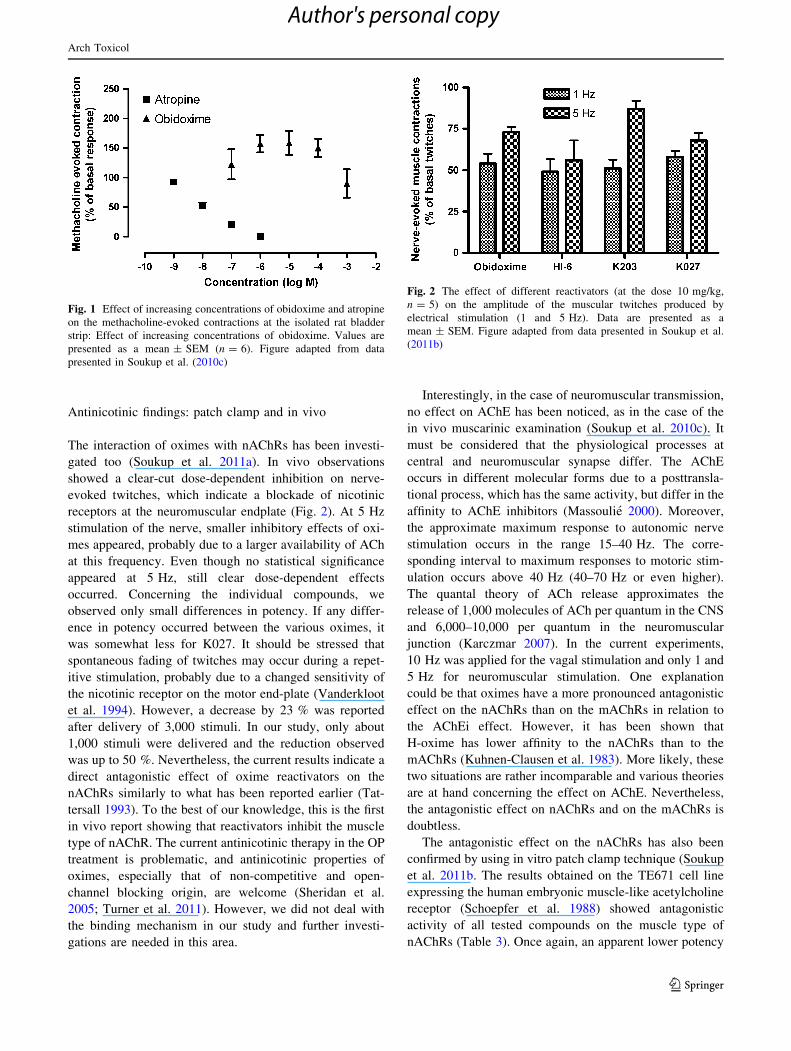
showed a clear-cut dose-dependent inhibition on nerve-
evoked twitches, which indicate a blockade of nicotinic
receptors at the neuromuscular endplate (Fig. 2). At 5 Hz
stimulation of the nerve, smaller inhibitory effects of oxi-
mes appeared, probably due to a larger availability of ACh
at this frequency. Even though no statistical significance
appeared at 5 Hz, still clear dose-dependent effects
occurred. Concerning the individual compounds, we
observed only small differences in potency. If any differ-
ence in potency occurred between the various oximes, it
was somewhat less for K027. It should be stressed that
spontaneous fading of twitches may occur during a repet-
itive stimulation, probably due to a changed sensitivity of
the nicotinic receptor on the motor end-plate (Vanderkloot
et al. 1994). However, a decrease by 23 % was reported
after delivery of 3,000 stimuli. In our study, only about
1,000 stimuli were delivered and the reduction observed
was up to 50 %. Nevertheless, the current results indicate a
direct antagonistic effect of oxime reactivators on the
nAChRs similarly to what has been reported earlier (Tat-
tersall 1993). To the best of our knowledge, this is the first
in vivo report showing that reactivators inhibit the muscle
type of nAChR. The current antinicotinic therapy in the OP
treatment is problematic, and antinicotinic properties of
oximes, especially that of non-competitive and open-
channel blocking origin, are welcome (Sheridan et al.
2005; Turner et al. 2011). However, we did not deal with
the binding mechanism in our study and further investi-
gations are needed in this area.
Interestingly, in the case of neuromuscular transmission,
no effect on AChE has been noticed, as in the case of the
in vivo muscarinic examination (Soukup et al. 2010c). It
must be considered that the physiological processes at
central and neuromuscular synapse differ. The AChE
occurs in different molecular forms due to a posttransla-
tional process, which has the same activity, but differ in the
affinity to AChE inhibitors (Massoulie 2000). Moreover,
the approximate maximum response to autonomic nerve
stimulation occurs in the range 15–40 Hz. The corre-
sponding interval to maximum responses to motoric stim-
ulation occurs above 40 Hz (40–70 Hz or even higher).
The quantal theory of ACh release approximates the
release of 1,000 molecules of ACh per quantum in the CNS
and 6,000–10,000 per quantum in the neuromuscular
junction (Karczmar 2007). In the current experiments,
10 Hz was applied for the vagal stimulation and only 1 and
5 Hz for neuromuscular stimulation. One explanation
could be that oximes have a more pronounced antagonistic
effect on the nAChRs than on the mAChRs in relation to
the AChEi effect. However, it has been shown that
H-oxime has lower affinity to the nAChRs than to the
mAChRs (Kuhnen-Clausen et al. 1983). More likely, these
two situations are rather incomparable and various theories
are at hand concerning the effect on AChE. Nevertheless,
the antagonistic effect on nAChRs and on the mAChRs is
doubtless.
The antagonistic effect on the nAChRs has also been
confirmed by using in vitro patch clamp technique (Soukup
et al. 2011b. The results obtained on the TE671 cell line
expressing the human embryonic muscle-like acetylcholine
receptor (Schoepfer et al. 1988) showed antagonistic
activity of all tested compounds on the muscle type of
nAChRs (Table 3). Once again, an apparent lower potency
Fig. 1 Effect of increasing concentrations of obidoxime and atropine
on the methacholine-evoked contractions at the isolated rat bladder
strip: Effect of increasing concentrations of obidoxime. Values are
presented as a mean ± SEM (n = 6). Figure adapted from data
presented in Soukup et al. (2010c)
Fig. 2 The effect of different reactivators (at the dose 10 mg/kg,
n = 5) on the amplitude of the muscular twitches produced by
electrical stimulation (1 and 5 Hz). Data are presented as a
mean ± SEM. Figure adapted from data presented in Soukup et al.
(2011b)
Arch Toxicol
123
Author's personal copy
of K027 was observed. Comparing this efficacy with
in vitro muscarinic examinations, the potency to affect the
mAChRs is approximately one order higher. This agrees
well with previous observations (Kuhnen-Clausen et al.
1983). In these experiments, antagonism on both the
embryonic and the adult form of the nAChRs was shown.
Interaction with HACU
As has been mentioned above, reactivators have a complex
mechanism of action. The presynaptic activity differs among
individual reactivators, and this effect influences the release
of ACh (Aas 1996; Oydvin et al. 2005). In another experi-
ment, the interaction of oximes with the HACU transport
system and their effect on membrane fluidity were examined
(Kristofikova et al. 2003; Soukup et al. 2010a, 2012). This
system is a key regulatory step (rate limiting) in the syn-
thesis of new ACh. We investigated standard reactivators
(obidoxime, HI-6, trimedoxime, methoxime and pralidox-
ime) and a new oxime—K112. In general, the results
showed that oxime reactivators are able to interact with this
carrier at low (10 lM; K112 and trimedoxime) or at high
concentrations (50 lM; HI-6, obidoxime and pralidoxime).
While methoxime did not significantly affect the carrier, an
influence on the fluidity of membrane was observed
(Table 4). It should be noticed that even though we observed
significant effects at micromolar concentrations, much lower
concentrations (picomolar or nanomolar) estimated in the
experiments in vitro will be relevant for the in vivo use
(Kristofikova et al. 1992, 2001; Kristofikova and Klaschka
1997). Due to this fact, prolonged administration would
need to be considered. However, a correlation between
HC-3 sensitive carriers and an impairment of cholinergic
neurotransmitter system was observed in various diseases
(e.g., Alzheimer’s), where the prolonged HACU inhibition is
connected with toxic detrimental effects. Therefore, a sig-
nificant inhibition of the carriers mediated by trimedoxime
and K112 could be a risk factor considering the prolonged
administration. On contrary, methoxime did not influence
the carriers, and its easy penetration into the membrane
could be an advantage since positively charged reactivators
hardly penetrate across the blood–brain barrier (Sakurada
et al. 2003). These results also show that reactivators are
able to modulate the release of ACh by the presynaptic
activity. Whether it is a result of direct interaction with
HACU, presynaptic cholinergic receptors or membrane
fluidity impairment is a matter of discussion; however, a
cooperation of M2 muscarinic autoreceptors and presynap-
tically localized HC-3 sensitive carriers through G proteins
has also been suggested (Zahniser and Doolen 2001).
Concerning the antidotal efficacy of individual reactiva-
tors, it is well known that results from in vitro examinations
do not always correlate with in vivo findings (Kuhnen-
Clausen et al. 1983; Tattersall 1993).For example, HI-6 can
handle in vivo tabun poisoning (Hamilton and Lundy 2007),
whereas in vitro it exerts only a minute reactivation potency
(Kuca et al. 2007). In our study, we observed a similar
situation. We performed experiments employing commonly
used reactivators and some of our new, promising com-
pounds, and the efficacy differed for individual compounds
when various methods are applied. Trimedoxime and oxime
K112 exerted high potency to inhibit choline reuptake,
Table 3 The effect of selected reactivators on the nicotinic receptors
expressed by the TE671 cell line in the whole-cell mode patch clamp
(Soukup et al. 2011b)
Reactivator Inhibition (200 lM) (%) S.E.M (%)
Obidoxime 51 2
HI-6 45 1
K027 31 1
K203 50 1
Inhibition reflects the drop (in %) in a response caused by reactivators
at the concentration of 200 lM on the ACh 10-lM-evoked response
(0 % = no effect, 100 % = complete inhibition). n = 4 for obidox-
ime and HI-6, n = 3 for K027 and K203
Table 4 Effect of reactivators on the high-affinity choline uptake (HACU) system and membrane anisotropy
Reactivator (3H)-choline (% of control) 10 lM (3H)-HC-3 (% of control) 50 lM Scatchard analysis TMA-DPH (%) DPH (%)
HI-6 91.0 ± 28.1 71.4 ± 12.9* : Kd 101.4 ± 2.6 97.1 ± 2.4
Trimedoxime 26.7 ± 6.2*** 48.4 ± 13.8*** : Kd 102.8 ± 2.6* 99.5 ± 1.7
Methoxime 85.9 ± 46.7 97.3 ± 27.2 – 103.1 ± 2.7* 94.4 ± 2.1**
Obidoxime 69.4 ± 26.1 40.2 ± 23.3*** ; Bmax 102.5 ± 2.0* 97.1 ± 2.4
Pralidoxime 71.8 ± 11.7 49.5 ± 25.7*** ; Bmax 106.1 ± 0.3*** 103.2 ± 1.1
K112 68.4 ± 9.9* 64.7 ± 4.7** : Kd 96.5 ± 1.0** 97.5 ± 1.1
All values with asterisk(s) represent a statistical difference from the control. Scatchard analysis refers to (3H)-hemicholinium-3 experiments.
Anisotropy was estimated in the region of the hydrophilic heads of phospholipid bilayer (TMA-DPH) or in the hydrocarbon core (DPH). Kd
characterizes the affinity between the carrier and its competitive inhibitor, and Bmax represents maximal number of binding sites. Table adapted
from Soukup et al. (2012) and extended
Arch Toxicol
123
Author's personal copy

obidoxime showed high affinity to muscarinic receptors, all
tested oximes exerted similar antinicotinic effect, whereas
oxime K027 showed considerably lower potency etc.
Therefore, it is very hard to choose the best reactivator
according to our examinations, but this has not been the aim
for our work at the present stage of understanding. However,
the generally accepted broad spectrum usage of HI-6 should
be pointed out (Kassa 2002), despite of the fact that this
versatility is not fully understood. Similarly, we observed
that even though HI-6 was less effective in some in vitro
examinations, it showed considerably higher potency when
examined in the whole (isolated) organ or in vivo. Unfor-
tunately, we have no exact explanation of this phenomenon,
but tentatively, the interaction with presynaptic receptors
(Vanhelden et al. 1996), or with other, so far, unrevealed
cholinergic mechanisms, may be the cause.
Concluding comments
In conclusion, one part of the mechanism of action of the
oxime reactivators that shows a pronounced variation is
their antagonistic effect on muscarinic and nicotinic
receptors. This effect is likely to be of significance in the
treatment of OP poisoning, where the relative ACh excess
can be counteracted by the blocking properties of oximes
on cholinergic receptors. The comparison of the in vitro
data reported oximes to exert higher efficacy on the mus-
carinic M2 subtype than on the AChE. However, this effect
seemed to be subtype specific since the antagonistic M3
effect was lower (i.e., M2 [ AChE [ M3). Also, and
importantly, the antimuscarinic effect was larger than that
on nicotinic receptors. Albeit oxime reactivators show
much lower muscarinic antagonism than another drug used
in the treatment, the classical antidote atropine, it is sup-
posed that non-reactivation properties of oxime reactiva-
tors play a significant role in the treatment of OP
poisoning. Another important phenomenon may occur in
the treatment with reactivators—a dual effect on cholin-
ergic receptors and on AChE simultaneously. This property
is of course not in favor in the antidotal treatment, and it
should be stressed that this effect on AChE has not been
observed at the nicotinic transmission. Since overstimula-
tion of nicotinic receptors is considered to be the cause of
death in the OP poisoning, clear antinicotinic action is of
advantage in this matter.
Acknowledgments This work was supported by the Grant No.
NT12062 from the Ministry of Health, Czech Republic and the
Postdoctoral project No. CZ.1.07/2.3.00/30.0012.
Conflict of interest The authors declare that they have no conflict
of interest.
References
Aas P (1996) In vitro effects of toxogonin, HI-6 and HLo-7 on the
release of H-3 acetylcholine from peripheral cholinergic nerves
in rat airway smooth muscle. Eur J Pharmacol 301:59–66
Alkondon M, Rao KS, Albuquerque EX (1988) Acetylcholinesterase
reactivators modify the functional properties of the nicotinic
acetylcholine receptor ion channel. J Pharmacol Exp Ther 245:
543–556
Amitai G, Kloog Y, Balderman D, Sokolovsky M (1980) Interaction
of bis-pyridinium oximes with mouse-brain muscarinic receptor.
Biochem Pharmacol 29:483–488
Andersson KE, Hedlund P (2002) Pharmacologic perspective on the
physiology of the lower urinary tract. Urology 60:13–20
Bajgar J (2004) Organophosphates/nerve agent poisoning: mechanism
of action, diagnosis, prophylaxis, and treatment. Adv Clin Chem
38:151–216
Broomfield C, Dembure IJ, Cuculis J (1987) Binding of soman
antidotes to acetylcholine-receptors. Biochem Pharmacol 36:
1017–1022
Busker RW, Zijlstra JJ, Vanderwiel HJ, Melchers BPC, Vanhelden
HPM (1991) Organophosphate poisoning—a method to test
therapeutic effects of oximes other than acetylcholinesterase
reactivation in the rat. Toxicology 69:331–344
Calic M, Vrdoljak AL, Radic M, Jelic D, Jun D, Kuca K, Kovarik Z
(2006) In vitro and in vivo evaluation of pyridinium oximes:
mode of interaction with acetylcholinesterase, effect on tabun-
and soman-poisoned mice and their cytotoxicity. Toxicology
219:85–96
Christopoulos A, Lanzafame A, Mitchelson F (1998) Allosteric
interactions at muscarinic cholinoceptors. Clin Exp Pharmacol
Physiol 25:185–194
Clement JG (1979) Pharmacological actions of Hs-6, an oxime, on the
neuromuscular-junction. Eur J Pharmacol 53:135–141
Ellis J, Seidenberg M (1992) 2 Allosteric modulators interact at a
common site on cardiac muscarinic receptors. Mol Pharmacol
42:638–641
Ellis J, Seidenberg M (2000) Interactions of alcuronium, TMB-8, and
other allosteric ligands with muscarinic acetylcholine receptors:
studies with chimeric receptors. Mol Pharmacol 58:1451–1460
Fossier P, Baux G, Poulain B, Tauc L (1990) Receptor-mediated
presynaptic facilitation of quantal release of acetylcholine
induced by pralidoxime in aplysia. Cell Mol Neurobiol 10:
383–404
Fusek J, Patocka J (1976) Effects of some cholinesterase reactivators
on contractility of isolated guinea-pig heart atria. Acta Biol Med
Ger 35:657–661
Hamilton MG, Lundy PM (1989) HI-6 therapy of soman and tabun
poisoning in primates and rodents. Arch Toxicol 63:144–149
Hamilton MG, Lundy PM (2007) Medical countermeasures to
WMDs: defence research for civilian and military use. Toxicol-
ogy 233:8–12
Inns RH, Leadbeater L (1983) The efficacy of bispyridinium
derivatives in the treatment of organophosphonate poisoning in
the guinea-pig. J Pharm Pharmacol 35:427–433
Karczmar AG (2007) Cholinergic cells and pathways exploring the
vertebrate central cholinergic nervous system. Springer, New-
York, pp 49–54
Kassa J (2002) Review of oximes in the antidotal treatment of
poisoning by organophosphorus nerve agents. J Toxicol Clin
Toxicol 40:803–816
Kloog Y, Sokolovsky M (1985) Bisquaternary pyridinium oximes as
allosteric inhibitors of rat brain muscarinic receptors. Mol
Pharmacol 27:418–428
Arch Toxicol
123
Author's personal copy

Kloog Y, Galron R, Sokolovsky M (1986) Bisquaternary pyridinium
oximes as presynaptic agonists and postsynaptic antagonists of
muscarinic receptors. J Neurochem 46:767–772
Kostenis E, Cid HMB, Holzgrabe U, Mohr K (1996) Evidence for a
multiple binding mode of bispyridinium-type allosteric modula-
tors of muscarinic receptors. Eur J Pharmacol 314:385–392
Kristofikova Z, Klaschka J (1997) In vitro effect of ginkgo biloba
extract (EGb 761) on the activity of presynaptic cholinergic
nerve terminals in rat hippocampus. Dement Geriatr Cogn
Disord 8:43–48
Kristofikova Z, Benesova O, Tejkalova H (1992) Changes of high-
affinity choline uptake in the hippocampus of old rats after long-
term administration of two nootropic drugs (Tacrine and Ginkgo
biloba extract). Dement Geriatr Cogn Disord 3:304–307
Kristofikova Z, Tejkalova H, Klaschka J (2001) Amyloid beta peptide
1–40 and the function of rat hippocampal hemicholinium-3
sensitive choline carriers: effects of a proteolytic degradation
in vitro. Neurochem Res 26:203–212
Kristofikova Z, Platilova V, Klaschka J (2003) Age- and sex-
dependent effects of ethanol on hippocampal hemicholinium-3
sensitive choline carriers during postnatal development of rats.
Neurochem Res 28:397–405
Kuca K, Jun D, Bajgar J (2007) Currently used cholinesterase
reactivators against nerve agent intoxication: comparison of their
effectivity in vitro. Drug Chem Toxicol 30:31–40
Kuhnen-Clausen D (1970) Investigations on the parasympatholytic
effect of toxogonin on the guinea-pig isolated ileum. Eur J
Pharmacol 9:85–92
Kuhnen-Clausen D, Hagedorn I, Gross G, Bayer H, Hucho F (1983)
Interactions of bisquaternary pyridine salts (H-oximes) with
cholinergic receptors. Arch Toxicol 54:171–179
Lee NH, Elfakahany EE (1991) Allosteric antagonists of the muscarinic
acetylcholine-receptor. Biochem Pharmacol 42:199–205
Loke WK, Sim MK, Go ML (2002) O-Substituted derivatives of
pralidoxime: muscarinic properties and protection against soman
effects in rats. Eur J Pharmacol 442:279–287
Lundy PM, Tremblay KP (1979) Ganglion blocking properties of
some bispyridinium soman antagonists. Eur J Pharmacol 60:
47–53
Marrs TC (1993) Organophosphate poisoning. Pharmacol Ther
58:51–66
Massoulie J (2000) Molecular forms and anchoring of acetylcholin-
esterase. Cholinesterases and cholinesterase inhibitors. Martin
Dunitz Ltd, London, pp 81–94
Melchers BPC, Vanderlaaken AL (1991) On the mechanism of
spontaneous-recovery of neuromuscular-transmission after ace-
tylcholinesterase inhibition in the rat neuromuscular-junction.
Brain Res 563:49–56
Niessen KV, Tattersall JEH, Timperley CM, Bird M, Green C, Seeger
T, Thiermann H, Worek F (2011) Interaction of bispyridinium
compounds with the orthosteric binding site of alpha-7 and
Torpedo californica nicotinic acetylcholine receptors (nAChRs).
Toxicol Lett 206:100–104
Ogura H, Kosasa T, Kuriya Y, Yamanishi Y (2000) Comparison of
inhibitory activities of donepezil and other cholinesterase
inhibitors on acetylcholinesterase and butyrylcholinesterase
in vitro. Methods Find Exp Clin Pharmacol 22:609–613
Oydvin OK, Tanso R, Aas P (2005) Pre-junctional effects of oximes
on H-3-acetylcholine release in rat hippocampal slices during
soman intoxication. Eur J Pharmacol 516:227–234
Reithmann C, Berger HJ, Hilf G, Werdan K (1991) The oxime Hgg-
12 as a muscarinic acetylcholine-receptor antagonist without
intrinsic activity in cardiac membranes. Arch Toxicol 65:
518–523
Sakurada K, Matsubara K, Shimizu K, Shiono H, Seto Y, Tsuge K,
Yoshino M, Sakai I, Mukoyama H, Takatori T (2003)
Pralidoxime iodide (2-PAM) penetrates across the blood-brain
barrier. Neurochem Res 28:1401–1407
Schoepfer R, Luther M, Lindstrom J (1988) The human medullo-
blastoma cell-line Te671 expresses a muscle-like acetylcholine-
receptor—cloning of the alpha-subunit cdna. FEBS Lett 226:
235–240
Seeger T, Eichhorn M, Lindner M, Niessen KV, Tattersall JEH,
Timperley CM, Bird M, Green AC, Thiermann H, Worek F
(2012) Restoration of soman-blocked neuromuscular transmis-
sion in human and rat muscle by the bispyridinium non-oxime
MB327 in vitro. Toxicology 294:80–84
Sheridan RD, Smith AP, Turner SR, Tattersall JEH (2005) Nicotinic
antagonists in the treatment of nerve agent intoxication. J R Soc
Med 98:114–115
Soukup O, Pohanka M, Tobin G, Jun D, Fusek J, Musilek K, Marek J,
Kassa J, Kuca K (2008) The effect of HI-6 on cholinesterases
and on the cholinergic system of the rat bladder. Neuro
Endocrinol Lett 29:759–762
Soukup O, Kristofikova Z, Proska J, Tobin G, Patocka J, Marek J, Jun
D, Fusek J, Ripova D, Kuca K (2010a) Novel acetylcholines-
terase reactivator K112 and its cholinergic properties. Biomed
Pharmacother 64:541–545
Soukup O, Tobin G, Kumar UK, Binder J, Proska J, Jun D, Fusek J,
Kuca K (2010b) Interaction of nerve agent antidotes with
cholinergic systems. Curr Med Chem 17:1708–1718
Soukup O, Tobin G, Kumar UK, Jun D, Fusek J, Kuca K (2010c)
Characterization of the anticholinergic properties of obidoxime;
functional examinations of the rat atria and the urinary bladder.
Toxicol Mech Methods 20:428–433
Soukup O, Krusek J, Kaniakova M, Kumar UK, Oz M, Jun D, Fusek
J, Kuca K, Tobin G (2011a) Oxime reactivators and their in vivo
and in vitro effects on nicotinic receptors. Physiol Res 60:
679–686
Soukup O, Kumar UK, Proska J, Bratova L, Adem A, Jun D, Fusek J,
Kuca K, Tobin G (2011b) The effect of oxime reactivators on
muscarinic receptors: functional and binding examinations.
Environ Toxicol Pharmacol 31:364–370
Soukup O, Kristofikova Z, Jun D, Tambor V, Ripova D, Kuca K
(2012) The interaction of standard oxime reactivators with
hemicholinium-3 sensitive choline carriers. Toxicol Lett 212:
315–319
Tattersall JE (1993) Ion channel blockade by oximes and recovery of
diaphragm muscle from soman poisoning in vitro. Br J
Pharmacol 108:1006–1015
Tucek S, Proska J (1995) Allosteric modulation of muscarinic
acetylcholine-receptors. Trends Pharmacol Sci 16:205–212
Turner SR, Chad JE, Price M, Timperley CM, Bird M, Green AC,
Tattersall JEH (2011) Protection against nerve agent poisoning
by a noncompetitive nicotinic antagonist. Toxicol Lett 206:
105–111
Vanderkloot W, Balezina OP, Molgo J, Naves LA (1994) The timing
of channel opening during miniature end-plate currents at the
frog and mouse neuromuscular-junctions—effects of Fasciculin-
2, other anti-cholinesterases and vesamicol. Pflugers Arch J
Physiol 428:114–126
Vanhelden HPM, Vanderwiel HJ, Delange J, Busker RW, Melchers
BPC, Wolthuis OL (1992) Therapeutic efficacy of Hi-6 in
soman-poisoned marmoset monkeys. Toxicol Appl Pharmacol
115:50–56
Vanhelden HP, Busker RW, Melchers BP, Bruijnzeel PL (1996)
Pharmacological effects of oximes: how relevant are they? Arch
Toxicol 70:779–786
Ward TR, Ferris DJ, Tilson HA, Mundy WR (1993) Correlation of the
anticholinesterase activity of a series of organophosphates with
Their ability to compete with agonist binding to muscarinic
receptors. Toxicol Appl Pharmacol 122:300–307
Arch Toxicol
123
Author's personal copy

Wolthuis O, Vanwersch RAP, Van Der Wiel HJ (1981) The efficacy
of some bis-pyridinium oximes as antidotes to soman in isolated
muscles of several species including man. Eur J Pharmacol
70:355–369
Zahniser NR, Doolen S (2001) Chronic and acute regulation of Na ?/
Cl-dependent neurotransmitter transporters: drugs, substrates,
presynaptic receptors, and signaling systems. Pharmacol Ther
92:21–55
Arch Toxicol
123
Author's personal copy




















