
The status of Mastodon angustidens pygmaeus Depéret, 1897 (Proboscidea, Mammalia): The contribution...
6
Original article The status of Mastodon angustidens pygmaeus Depe ´ ret, 1897 (Proboscidea, Mammalia): The contribution of X-ray tomography § Pascal Tassy a, *, Florent Goussard b , Miguel Garcia Sanz b a Muse ´um national d’histoire naturelle, de ´partement histoire de la Terre, UMR7207 CNRS-MNHN-UPMC CR2P, CP 38, 57, rue Cuvier, 75231 Paris cedex 05, France b Muse ´um national d’histoire naturelle, plateforme AST-RX (acce `s scientifique a ` la tomographie a ` rayons X), de ´partement syste ´matique et e ´volution, UMS 2700 CNRS-MNHN outils et me ´thodes de la syste ´matique inte ´grative, CP140, 57, rue Cuvier, 75231 Paris cedex 05, France 1. Introduction Today, a young palaeontologist and/or phylogenetist is first of all an internaut. If he/she is interested by the phylogeny of Proboscidea Illiger, 1811, he may search the Internet and find the address http://www.palaeos.org/Gomphotherioidea. There, he/she will find a peculiar phylogeny of the ‘‘Gomphotherioidea’’, including a taxon labelled ‘‘Gomphotherium pygmaeus (Depe ´ ret, 1897)’’. From various viewpoints (palaeontological, odontological, anatomical, evolutionary, and nomenclatural), the history of this problematic taxon and of its name is worth recalling with an explanation both frustrating and rational. The original name was conceived by Charles Depe ´ ret (1897: p. 519, pl. 19, figs. 1–3) as a trinom: ‘‘Mastodon angustidens Cuv. mut. asc. pygmaeus’’, based on a molar lacking the first lophid, and interpreted as an m3. The abbreviation ‘‘mut. asc.’’ means ‘‘mutation ascendante’’, that is ‘‘ascending mutation’’ (discussed below). The original hypodigm includes only the holotype which was found in Kabylie (Algeria) during a geologic excursion organized by the Socie ´te ´ ge ´ ologique de France in post-Paleogene outcrops, the ‘‘gre `s cartenniens a ` Clype ´ astres qui affleurent dans la tranche ´e de la route entre Chabet-el-Ameur et Isserville (Kabylie)’’. The molar looks like a germ of a bunolophodont m3 (Fig. 1) and Depe ´ ret pointed its peculiarities: small size (complete, the molar could have reached only 110 mm), large amount of coronal cement and ‘‘faible de ´doublement des mamelons principaux en mamelons secondaires’’ (Depe ´ ret, 1897: p. 519), i.e., weak development of mesoconelets (labial and lingual conelets on each side of the median sulcus). This molar has been interpreted in many ways since its identification as a gomphothere, a palaeomastodont, a choerolo- phodont, a supernumerary molar, and a nomen dubium. The taxonomic challenge introduced by this molar is that a thick coating of cement entirely occupies the interlophids (except the first, where the molar is broken). As a consequence, the organization of the central conules, which brings taxonomic information, cannot be seen since the central conules are hidden (Fig. 1). This is why numerous hypotheses have been erected, although the shape of the anterior pretrite central conule of the anteriorly-preserved lophid could have already disclosed some of them. Because the molar is a holotype, nobody dared to clean the interlophids. The non-destructive technique of X-ray tomography allows a virtual dissection and show for the first time the pattern of the crown and the shape of central conules (Figs. 2–5). Abbreviations: IM, Indian Museum (Calcutta, India); UCBCG, Universite ´ Claude Bernard Lyon 1, France (Collections de Ge ´ ologie); MNHN, Muse ´um national d’Histoire naturelle (Paris, France). 2. Material and methods The holotype of Mastodon angustidens pygmaeus belongs to the collection of geology of the University Claude Bernard Lyon 1, France (UCBCG), with number FSL 213759. Geobios 46 (2013) 329–334 A R T I C L E I N F O Article history: Received 24 September 2012 Accepted 28 January 2013 Available online 30 April 2013 Keywords: Proboscidea Gomphotherium pygmaeum Choerolophodon pygmaeus Molar X-ray tomography A B S T R A C T The status of Mastodon angustidens pygmaeus Depe ´ ret, 1897 is discussed on the basis of an examination of the holotype (a broken lower molar) with tomography using X-rays, and on a review of the previous taxonomic hypotheses. The conclusion is that the holotype is a possible supernumerary post-m3 molar that may be that of a tetralophodont gomphothere. ‘‘M. angustidens pygmaeus’’ is then a nomen dubium that should only be used in relation to the holotype. ß 2013 Elsevier Masson SAS. All rights reserved. § Corresponding editor: Gilles Escarguel. * Corresponding author. E-mail address: [email protected] (P. Tassy). Available online at www.sciencedirect.com 0016-6995/$ – see front matter ß 2013 Elsevier Masson SAS. All rights reserved. http://dx.doi.org/10.1016/j.geobios.2013.01.003
Transcript of The status of Mastodon angustidens pygmaeus Depéret, 1897 (Proboscidea, Mammalia): The contribution...

Geobios 46 (2013) 329–334
Original article
The status of Mastodon angustidens pygmaeus Deperet, 1897 (Proboscidea,Mammalia): The contribution of X-ray tomography§
Pascal Tassy a,*, Florent Goussard b, Miguel Garcia Sanz b
a Museum national d’histoire naturelle, departement histoire de la Terre, UMR7207 CNRS-MNHN-UPMC CR2P, CP 38, 57, rue Cuvier, 75231 Paris cedex 05, Franceb Museum national d’histoire naturelle, plateforme AST-RX (acces scientifique a la tomographie a rayons X), departement systematique et evolution, UMS 2700 CNRS-MNHN outils et
methodes de la systematique integrative, CP140, 57, rue Cuvier, 75231 Paris cedex 05, France
A R T I C L E I N F O
Article history:
Received 24 September 2012
Accepted 28 January 2013
Available online 30 April 2013
Keywords:
Proboscidea
Gomphotherium pygmaeum
Choerolophodon pygmaeus
Molar
X-ray tomography
A B S T R A C T
The status of Mastodon angustidens pygmaeus Deperet, 1897 is discussed on the basis of an examination
of the holotype (a broken lower molar) with tomography using X-rays, and on a review of the previous
taxonomic hypotheses. The conclusion is that the holotype is a possible supernumerary post-m3 molar
that may be that of a tetralophodont gomphothere. ‘‘M. angustidens pygmaeus’’ is then a nomen dubium
that should only be used in relation to the holotype.
� 2013 Elsevier Masson SAS. All rights reserved.
Available online at
www.sciencedirect.com
1. Introduction
Today, a young palaeontologist and/or phylogenetist is first ofall an internaut. If he/she is interested by the phylogeny ofProboscidea Illiger, 1811, he may search the Internet and find theaddress http://www.palaeos.org/Gomphotherioidea. There, he/shewill find a peculiar phylogeny of the ‘‘Gomphotherioidea’’,including a taxon labelled ‘‘Gomphotherium pygmaeus (Deperet,1897)’’. From various viewpoints (palaeontological, odontological,anatomical, evolutionary, and nomenclatural), the history of thisproblematic taxon and of its name is worth recalling with anexplanation both frustrating and rational.
The original name was conceived by Charles Deperet (1897: p.519, pl. 19, figs. 1–3) as a trinom: ‘‘Mastodon angustidens Cuv. mut.asc. pygmaeus’’, based on a molar lacking the first lophid, andinterpreted as an m3. The abbreviation ‘‘mut. asc.’’ means ‘‘mutationascendante’’, that is ‘‘ascending mutation’’ (discussed below). Theoriginal hypodigm includes only the holotype which was found inKabylie (Algeria) during a geologic excursion organized by theSociete geologique de France in post-Paleogene outcrops, the ‘‘grescartenniens a Clypeastres qui affleurent dans la tranchee de la routeentre Chabet-el-Ameur et Isserville (Kabylie)’’. The molar looks like agerm of a bunolophodont m3 (Fig. 1) and Deperet pointed itspeculiarities: small size (complete, the molar could have reached
§ Corresponding editor: Gilles Escarguel.
* Corresponding author.
E-mail address: [email protected] (P. Tassy).
0016-6995/$ – see front matter � 2013 Elsevier Masson SAS. All rights reserved.
http://dx.doi.org/10.1016/j.geobios.2013.01.003
only 110 mm), large amount of coronal cement and ‘‘faiblededoublement des mamelons principaux en mamelons secondaires’’(Deperet, 1897: p. 519), i.e., weak development of mesoconelets(labial and lingual conelets on each side of the median sulcus).
This molar has been interpreted in many ways since itsidentification as a gomphothere, a palaeomastodont, a choerolo-phodont, a supernumerary molar, and a nomen dubium. Thetaxonomic challenge introduced by this molar is that a thickcoating of cement entirely occupies the interlophids (except thefirst, where the molar is broken). As a consequence, theorganization of the central conules, which brings taxonomicinformation, cannot be seen since the central conules are hidden(Fig. 1). This is why numerous hypotheses have been erected,although the shape of the anterior pretrite central conule of theanteriorly-preserved lophid could have already disclosed some ofthem. Because the molar is a holotype, nobody dared to clean theinterlophids. The non-destructive technique of X-ray tomographyallows a virtual dissection and show for the first time the pattern ofthe crown and the shape of central conules (Figs. 2–5).
Abbreviations: IM, Indian Museum (Calcutta, India); UCBCG,Universite Claude Bernard Lyon 1, France (Collections de Geologie);MNHN, Museum national d’Histoire naturelle (Paris, France).
2. Material and methods
The holotype of Mastodon angustidens pygmaeus belongs to thecollection of geology of the University Claude Bernard Lyon 1,France (UCBCG), with number FSL 213759.
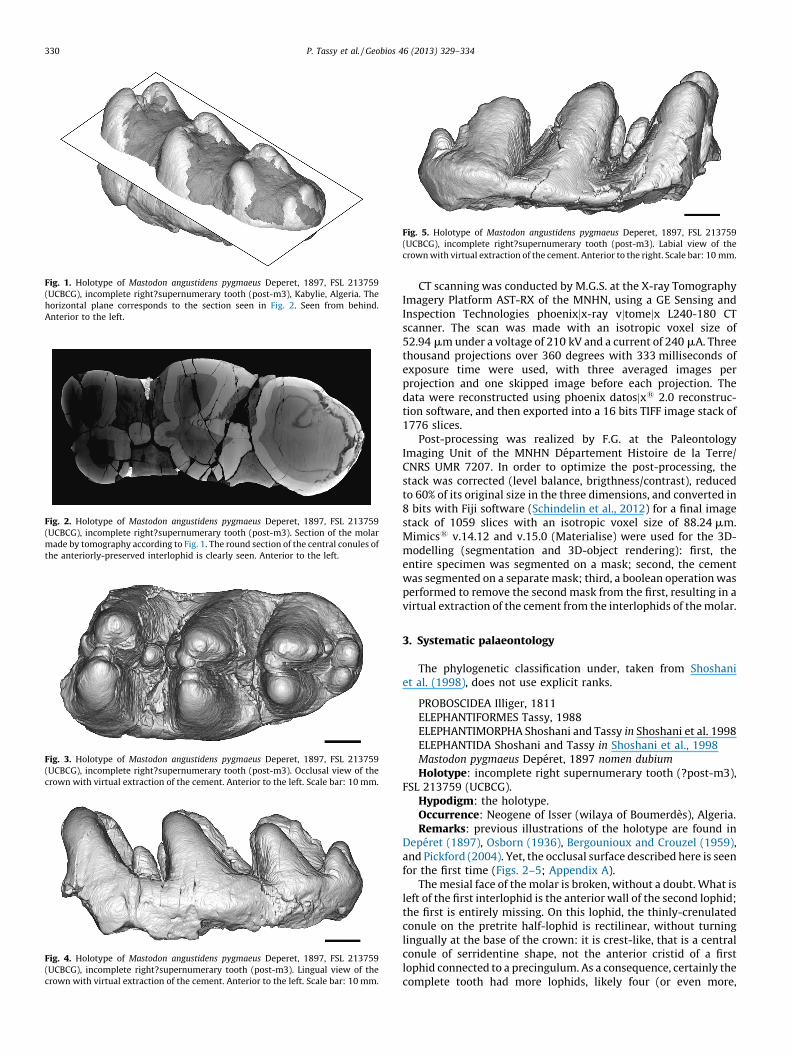
Fig. 1. Holotype of Mastodon angustidens pygmaeus Deperet, 1897, FSL 213759
(UCBCG), incomplete right?supernumerary tooth (post-m3), Kabylie, Algeria. The
horizontal plane corresponds to the section seen in Fig. 2. Seen from behind.
Anterior to the left.
Fig. 2. Holotype of Mastodon angustidens pygmaeus Deperet, 1897, FSL 213759
(UCBCG), incomplete right?supernumerary tooth (post-m3). Section of the molar
made by tomography according to Fig. 1. The round section of the central conules of
the anteriorly-preserved interlophid is clearly seen. Anterior to the left.
Fig. 3. Holotype of Mastodon angustidens pygmaeus Deperet, 1897, FSL 213759
(UCBCG), incomplete right?supernumerary tooth (post-m3). Occlusal view of the
crown with virtual extraction of the cement. Anterior to the left. Scale bar: 10 mm.
Fig. 4. Holotype of Mastodon angustidens pygmaeus Deperet, 1897, FSL 213759
(UCBCG), incomplete right?supernumerary tooth (post-m3). Lingual view of the
crown with virtual extraction of the cement. Anterior to the left. Scale bar: 10 mm.
Fig. 5. Holotype of Mastodon angustidens pygmaeus Deperet, 1897, FSL 213759
(UCBCG), incomplete right?supernumerary tooth (post-m3). Labial view of the
crown with virtual extraction of the cement. Anterior to the right. Scale bar: 10 mm.
P. Tassy et al. / Geobios 46 (2013) 329–334330
CT scanning was conducted by M.G.S. at the X-ray TomographyImagery Platform AST-RX of the MNHN, using a GE Sensing andInspection Technologies phoenixjx-ray vjtomejx L240-180 CTscanner. The scan was made with an isotropic voxel size of52.94 mm under a voltage of 210 kV and a current of 240 mA. Threethousand projections over 360 degrees with 333 milliseconds ofexposure time were used, with three averaged images perprojection and one skipped image before each projection. Thedata were reconstructed using phoenix datosjx1 2.0 reconstruc-tion software, and then exported into a 16 bits TIFF image stack of1776 slices.
Post-processing was realized by F.G. at the PaleontologyImaging Unit of the MNHN Departement Histoire de la Terre/CNRS UMR 7207. In order to optimize the post-processing, thestack was corrected (level balance, brigthness/contrast), reducedto 60% of its original size in the three dimensions, and converted in8 bits with Fiji software (Schindelin et al., 2012) for a final imagestack of 1059 slices with an isotropic voxel size of 88.24 mm.Mimics1 v.14.12 and v.15.0 (Materialise) were used for the 3D-modelling (segmentation and 3D-object rendering): first, theentire specimen was segmented on a mask; second, the cementwas segmented on a separate mask; third, a boolean operation wasperformed to remove the second mask from the first, resulting in avirtual extraction of the cement from the interlophids of the molar.
3. Systematic palaeontology
The phylogenetic classification under, taken from Shoshaniet al. (1998), does not use explicit ranks.
PROBOSCIDEA Illiger, 1811ELEPHANTIFORMES Tassy, 1988ELEPHANTIMORPHA Shoshani and Tassy in Shoshani et al. 1998ELEPHANTIDA Shoshani and Tassy in Shoshani et al., 1998Mastodon pygmaeus Deperet, 1897 nomen dubium
Holotype: incomplete right supernumerary tooth (?post-m3),FSL 213759 (UCBCG).
Hypodigm: the holotype.Occurrence: Neogene of Isser (wilaya of Boumerdes), Algeria.Remarks: previous illustrations of the holotype are found in
Deperet (1897), Osborn (1936), Bergounioux and Crouzel (1959),and Pickford (2004). Yet, the occlusal surface described here is seenfor the first time (Figs. 2–5; Appendix A).
The mesial face of the molar is broken, without a doubt. What isleft of the first interlophid is the anterior wall of the second lophid;the first is entirely missing. On this lophid, the thinly-crenulatedconule on the pretrite half-lophid is rectilinear, without turninglingually at the base of the crown: it is crest-like, that is a centralconule of serridentine shape, not the anterior cristid of a firstlophid connected to a precingulum. As a consequence, certainly thecomplete tooth had more lophids, likely four (or even more,

Fig. 6. Cf. Gomphotherium sp., right M3 of the palate MNHN 1961-5-12 referred by
Arambourg (1961) to ‘‘Trilophodon pygmaeus’’. Miocene of Gebel Zelten, Libya.
Occlusal view. Anterior to the right. Scale bar: 10 mm.
Photo. Ph. Loubry.
P. Tassy et al. / Geobios 46 (2013) 329–334 331
although it cannot be assessed); the preserved lophids areprobably the second, third and fourth ones. Each lophid isremarkable by the reduced size of the mesoconelets on each sideof the median sulcus, as already noticed by Deperet. On the lastlophid, the mesoconelet on the posttrite (lingual) side is extremelyreduced.
The preserved interlophids are wide and not closed by centralconules. Due to their weakness, the conules are entirely covered bythe cement. The conules, of moderate size, are cone-shaped at thebase of the anteriorly-preserved interlophid (second), and reducedto crests in the two posterior lophids (third and fourth). In thesecond interlophid the trefoil made by the central conules is of thebunodont type with a serridentine morphology. The central conuleon the posterior wall of the second lophid is made of a conicalcuspid connected to the lophid by a serridentine crest; the centralconule on the anterior wall of the third lophid is smaller butconical. In the third interlophid the central conules form two crestswithout conical isolated cuspid. The posterior crest of the lastlophid is reduced while a rectilinear thick serridentine crest runsfrom the top of the main cuspid (Figs. 3–5). The postcingulum ismade of two conical cuspids. The irregular shape of theserridentine crest of the last lophid is conspicuous in labial view(Fig. 5).
Weak crests on the posterior side of posttrite half-lophids canbe seen. On the second lophid the crest is rectilinear; on the thirdlophid the crest has a peculiar orientation toward the pretrite half-lophid and not the base of the interlophid (Fig. 3). In lateral views,all lophids are anteriorly inclined, a trait of a lower molar (Figs. 4,5). In occlusal view (Fig. 3), the crown displays a torsion: themedian sulcus is slightly concave labially.
This lower molar is of the bunolophodont type, a morphologyshared by gomphotheres in general. Lophids and the trefoil in thefirst entirely preserved interlophid (the second) displays thecommon pattern of gomphotheres. The main peculiar features ofthis small molar are:
� the amount of cement and reduction of the mesoconelets,transversely narrow and lower than the main cusps, as describedby Deperet;� the orientation of the coarse posterior pretrite crest of the last
lophid which does not run toward the median sulcus, and;� the orientation of the thin posttrite crest on the posterior side of
the third lophid.
The association of these peculiarities is rather unusual.
4. Discussion
The description given in the previous section is a mean toconfirm and refute the various taxonomic hypotheses publishedsince the 19th century. The different characters used to assessthese hypotheses are discussed below, in the light of this newdescription. The discussion leads to the hypothesis that this molarfrom Kabylie is likely a supernumerary tooth.
4.1. Small size and the gomphothere hypothesis
Deperet (1897: p. 520) introduces a mutation in the lineagecalled Mastodon angustidens Cuvier, 1817 (today Gomphotherium
angustidens): ‘‘Mastodon angustidens Cuv. mut. asc. pygmaeus’’. Theconcept of mutation is used by Deperet (1907: p. 161) followingWaagen (1869: p. 187): a small morphological change inside aphyletic lineage, that is, in nomenclatural modern concepts, achronological subspecies. The trinom ‘‘Mastodon angustidens
pygmaeus’’ is an ascending mutation (‘‘mut. asc.’’) located deep
in time and looking for the later Mastodon angustidens stricto sensu.For Deperet, the small size of the molar is an evolutionarycharacter, in that case primitive, this tooth being a confirmation ofprevious Deperet’s (1897: p. 520) observations that early Miocenemolars (Orleanais sands) are smaller than middle Miocene molars(locality of Sansan). This concept – a pygmy subspecies – wasaccepted by Bergounioux and Crouzel (1959: p. 102) with adifferent trinom: Trilophodon olisiponensis pygmaeus. This taxo-nomic treatment is a contrivance to the priority rule of the Code ofzoological nomenclature (Ginsburg and Telles Antunes, 1966):Deperet’s taxon name is older than Trilophodon olisiponensis
Zbyszewski, 1949. This latter species is based on two smallbunolophodont m3s of trilophodont grade from Portugal (whichare devoid of a cement coating) and, according to Shoshani andTassy (1996: p. 370), T. olisiponensis is a junior synonym ofGomphotherium angustidens.
Arambourg (1961), followed (with various generic synonyms)by Savage (1971), Savage and Hamilton (1973), Coppens et al.(1978: p. 343), describes Deperet’s taxon in the Miocene of GebelZelten, Libya, on the basis of a small palate with M2-M3(MNHN 1961-5-12) called ‘‘Trilophodon pygmaeus Deperet’’ (sic.).During the years, due to the salty sediments of Zelten (and otherLibyan localities such as Dor el Talha), the bony part of the palatewas progressively destroyed despite various treatments (Harris,1971). Yet, the small M3s of this palate (length of rt.M3 = 112 mm), devoid of cement, show thick lophs, narrowinterlophs, and large mesoconelets (Fig. 6), so that occlusion with am3 like the molar from Isser is impossible. Most recently, thebinom ‘‘Gomphotherium pygmaeus’’ (sic.) was retained by Sanderset al. (2010: p. 197) for an African pigmy lineage of gomphotheres,excluding the Gebel Zelten teeth allocated to ‘‘Gomphotherium sp.indet’’.
Indeed size does not matter much. The phyletic evolution ofG. angustidens as seen by Deperet does not exist, while intraspecificvariation and sexual dimorphism exist. For instance, in France, inthe middle Miocene locality of En Pejouan in the vicinity ofSimorre, calibrated MN7 zone (Ginsburg et al., 1975; Pean et al.,1998; Ginsburg and Bulot, 2000), the length of m3s of females ofGomphotherium angustidens is only 70% of that of males (Tassy,1996: p. 98). This does not mean that small species cannot exist:this means that size alone is not a sufficient taxonomic criterion.
4.2. Small size and the Phiomia hypothesis
Osborn (1936), followed by Ennouchi in 1948 (cited by Pickford,2004: p. 430), and by Savage and Russell (1983: p. 162), allocatedthe Algerian molar to a palaeomastodontid species, Phiomia
pygmaeus. This species is then understood as a link betweenNeogene mastodont molars (the ‘‘Cartennien’’ being supposed to

Fig. 7. Holotype of Choerolophodon palaeindicus (Lydekker, 1884), A 426 (IM), right
m3 (Bugti, Baluchistan, Pakistan) — see also Lydekker (1884: Plate 4, Figure 3:
‘‘Mastodon (Trilophodon) angustidens, Cuv. var. palaeindicus’’). Occlusal view.
Anterior to the right. Scale bar: 10 mm.
Photo. P. Tassy.
P. Tassy et al. / Geobios 46 (2013) 329–334332
be early Miocene) and Paleogene molars of Phiomia from theFayum in Egypt, which are smaller. Osborn’s conclusions arestraightforward (Osborn, 1936: p. 247, fig. 187): the molar iscomplete, a little larger than that of the largest previously knownspecies of the genus (Phiomia osborni), with the same number oflophids (three) and a postcingulum made of two cusps, the summitof them not covered by cement. But the molar is definitelyincomplete and had four lophids (or more). Moreover, Osborn(1936: p. 247) also writes surprisingly that ‘‘in P. pygmaeus, as inP. osborni [. . .], the summits of the cones are compressed towardsthe central apices of the respective lophs’’. As said above thecompression is tied to the reduction of the mesoconelets, but thistrait is not seen in P. osborni as well as in any species of the genusPhiomia, as shown by Andrews (1906), Matsumoto (1924), Osborn(1936), Tobien (1978), and Sanders et al. (2004). One can alsonotice that the longitudinal torsion of the crown is often seen inNeogene molars, never in Palaeogene ones.
4.3. Cement and the Choerolophodon hypothesis
The thickness of cement of FSL 213759 is rather peculiar. Amongnon-elephantine Elephantimorpha few species have molars with athick coating of cement. The amebelodontid Platybelodon grangeri isone of them but the cement does not cover entirely the crown andthe complex crown pattern has nothing to do with that of theAlgerian molar (Tobien et al., 1986). It is among the Choerolopho-dontidae – with two genera: the paraphyletic Afrochoerodon and themonophyletic Choerolophodon, as revised by Sanders et al. (2010: p.203) – that species with a distinctive quantity of cement on themolars are found, noticeably the more transformed, late Miocenespecies Choerolophodon pentelici and C. corrugatus.
On the basis of the presence of cement alone, Tobien (1973: p.215) allocates Deperet’s taxon to Choerolophodon. More precisely,Tobien writes that the type molar ‘‘indicates relationship toChoerolophodon’’, and further adds in the paragraph on ‘‘TheChoerolophodon/Synconolophus Group’’ that ‘‘from the MiddleMiocene of Djebel Zelten, Libya, Savage (1971: p. 222) mentions‘Mastodon pygmaeus’ which might indicate a similar, cementprovided, trilophodont taxon’’ (Tobien, 1973: p. 249). This view-point is again endorsed without formal taxonomic treatment byTobien (1976: p. 180).
Tobien’s choerolophodont hypothesis was resurrected byPickford (2004) who allocates new material (teeth and postcranialbones) from the middle Miocene of Kenya to Choerolophodon
pygmaeus and thinks to have reached the ‘‘resolution of a centuryold enigma’’. This small species (1.50 m at the shoulder accordingto Pickford, 2004: p. 459) is further described from Algeria to SouthAfrica (Pickford, 2005).
Yet, the top of the lophids of the Algerian molar shows reducedmesoconelets, narrow and less high than the main cusps, andpretrite conules with a ‘‘serridentine’’ morphotype: these traits arenot seen in Choerolophodon. What characterizes the choerolopho-dont pattern of the molars is the V-shaped loph(id)s due toanteriorly placed enlarged mesoconelets and central conules (theycan fuse), and their pillar-shape (Tassy, 1986: p. 66; Sanders, 2010:p. 203). This is already outlined in ‘‘Choerolophodon ‘kisumuensis
group’’’, the early Miocene stem group of the family. This is obviouson the small m3s of two species of this stem group: C. palaeindicus(Fig. 7) first described by Lydekker (1884: p. 19, pl. 4, fig. 3) as‘‘Mastodon (Trilophodon) angustidens, Cuv. var. palaeindicus’’, andC. kisumuensis first described under the trinom ‘‘Trilophodon
angustidens kisumuensis’’ by MacInnes (1942: p. 66, pl. 6, fig. 7)(Tassy, 1986: p. 67).
Pickford’s (2004) taxonomic revision was not endorsed bySanders et al. (2010) who do not see choerolophodont characterson the molars of C. pygmaeus sensu Pickford. Although it is
warranted that the holotype from Algeria does not displaychoerolophodont traits (the tomography does not allow furtherdebate), it is not the case for the molars from Ngenyin (Tugen Hills,Kenya) described by Pickford (2004). As a matter of fact, no m3described at Ngenyin (Pickford, 2004: pl. 4-5) matches withFSL 213759. The hypodigm of Gomphotherium pygmaeum sensu
Sanders et al. (2010) or Choerolophodon pygmaeus sensu Pickford(2004) is heterogeneous.
4.4. Hypothesis of an abnormal tooth
A somehow more radical solution is given by Roger et al. (1994:p. 266) who think that the species ‘‘Mastodon pygmaeus [. . .] fondeesur un type inadequat [. . .] est un nomen dubium’’, a conclusionendorsed by Shoshani and Tassy (1996: p. 370). This taxonomichypothesis is frustrating but it is the one supported by the presentanalysis. The molar FSL 213759 is somehow teratologic, and maybe a post-m3. As such, it can belong to any gomphothere oftrilophodont or tetralophodont grade (probably the latter, seebelow). Welcomme (1993: p. 60) describing a supernumerarytooth in situ in a skull of a gomphothere, thinks that FSL 213759 is asupernumerary tooth of Gomphotherium, and bases his taxonomicallocation on the hypothesis that only the precingulum is lackingand not an entire lophid, which is refuted by the present analysis —the entire first lophid is indeed missing.
4.4.1. Known supernumerary molars
The living African elephant is a model that can be used tounderstand the characteristics of a supernumerary molar. Indivi-duals of Loxodonta africana with a posterior supernumerary toothrepresent 0.8% to 10% relatively to the different populationsstudied by Laws et al. (1975) and Hanks (1979). Post-m3s ofL. africana are short, with less lamellae (up to three or four), andsometimes are like a ball covered by cement. In the fossil recordsupernumerary molars of Mammuthus primigenius follow the samerule, and can have a few less plates than m3s or can be very short(Van Essen, 2004). For Roth (1989: p. 176), the relatively highfrequency of supernumerary teeth in elephants is explained by thefact that elephants ‘‘lack precise occlusion, so supernumerary teethmay cause little functional impairment, especially if they occur latein the series or late in life’’ and develop ‘‘enhanced ability toaccommodate an extra tooth’’ due to the continual displacement.
In the palaeontological record, non-elephantid proboscideansupernumerary teeth are scarce. Yet, three cases exist without adoubt, including species of trilophodont and tetralophodontgrades:
� in Mammut americanum from the Pleistocene of Terre Coupee(now Terre Coupe) in Michigan (Warren, 1855: p. 83), a post-m3

P. Tassy et al. / Geobios 46 (2013) 329–334 333
(in situ in a right hemimandible behind the m3) has three lophidsand a strong postcingulum that can be held as an incipient fourthlophid (the m3 has four lophids and a postcingulum). The firstlophid is slightly worn, due to a probable contact with thepostcingulum of the M3;� in Gomphotherium hannibali from the early Miocene of Langue-
doc, a cranium with M2-M3 described by Welcomme (1993,1994), a post-M3 is seen on the left side (behind a normalprimitive M3 with three lophs and a postcingulum). It has twolophs and a reduced postcingulum. On the right side, the post-M3is still in the maxilla;� in Anancus arvernensis from the Pliocene of Saint-Laurent des
Arbres described by Viret (1962), the supernumerary tooth, in
situ behind the m3, has been dissected. It has five lophids (the lastone is reduced) when the m3 has seven lophids.
In conclusion, supernumerary molars have one less loph(id), oreven two or three, than M3/m3s, and are shorter. The pattern of theloph(id)s may also be different from the normal occlusal pattern ofthe M3/m3s.
4.4.2. The peculiar association of traits of FSL 213759
The tooth from Algeria is a fresh tooth, maybe a germ at thelatest stage of development with differentiating roots (broken onthe specimen). The development of the crown has finished, asshown by the thickness of the enamel, which is the same on theanterior and posterior sides. As a consequence, the strangecharacters associated on this molar, vast interlophids, smallmesoconelets, and thin lophids are not due to an early stage ofdevelopment. Moreover, normal gomphothere teeth, includinggerms, never have a thick coating of cement. Deperet (1897: p. 520)remarks that the important amount of cement could be due to theyoung individual age of the animal rather than a specific ormutation character. This cannot be followed: the presence ofcement on the crowns of proboscidean molars is associated to the‘‘horizontal’’ tooth displacement (Tassy, 1975). This mode of tootheruption is a character of elephantimorphs (all Neogene). Thedisplacement listed by Shoshani et al. (2006) in the Oligocenespecies from Eritrea, Eritreum melakeghebrekristrosi, is conjectural.The cement only begins to reach the interloph(id)s and the bottomof the cusp(id)s when the roots differentiate and the tooth erupts.It is rather thin, except for the taxa listed above (see Section 4.3).
The unique thick coating of cement of FSL 213759 is a first traitthat can be considered as abnormal. Moreover, the molar fromKabylie shows an association of several characters that canrepresent a derivation from the normal morphology of abunolophodont last lower molar. This association is:
� the cement is unusually thick;� the molar is small and narrow;� the interlophids are remarkably wide;� the main pretrite cusps of the two posterior preserved lophids
are compressed;� the mesoconelets on each of the median sulcus are reduced (low
and narrow), especially on the pretrite side;� the posterior pretrite crest of the posterior lophid is rectilinear,
slightly orientated labially and;� the posttrite crest of the third lophid has a labial orientation.
Each of these seven characters is not amazing in itself; hence,one could ask why these characters could not be used to diagnose asmall gomphothere species. The answer is that no other knownmolar shares the morphology displayed by the Algerian molar.Both hypodigms of Gomphotherium pygmaeum (sensu Sanderset al., 2010) and Choerolophodon pygmaeus (sensu Pickford, 2004,
2005) consist of molars, which do not exhibit this association.Then, the supposed intraspecific variation of such species appearsrather artificial.
It has long been known (Lartet, 1859: pl. 14, fig. 2F’) thatbunolophodont lower molars can exhibit reduced mesoconelets.But in that case it is due to an entirely different context — a globalcrowding, i.e.: 1) inflated main cusps; 2) fusion of highmesoconelet + central conule and/or main cusp; 3) both traits 1and 2 are associated to anteroposteriorly-inflated lophids andresulting narrow interlophids. The molar FSL 213759 shows areversed situation with narrow and low mesoconelets associatedto weak central conules and especially large interlophids. On theother hand, the central conules with a serridentine morphology ofFSL 213759 is a normal morphological variant of bunolophodontmolars (Tobien, 1972).
Based on the number of lophs and lophids of supernumerarymolars (see above), the molar from Algeria with four (or more)lophids could be a post-m3 of a species of tetralophodont grade,the m3s of which having five lophids or more. Although scarce andnot well known, non-elephantid tetralophodont taxa have beendescribed in the late Miocene of North Africa (Bergounioux andCrouzel, 1956; Gaziry, 1987; Geraads, 1989). For this reason thepost-Palaeogene ‘‘Cartennien’’ of former authors (Dalloni, 1917)may not be early Miocene (the type locality is already listed as‘‘?middle Miocene’’ by Sanders et al. (2010: p. 189)). The fact thatthis isolated discovery is that of a supernumerary tooth of atetralophodont gomphothere is rather surprising. Yet, historicalevents do not follow probability estimations.
5. Conclusion
The holotype of Mastodon angustidens pygmaeus is likely to bean abnormal lower molar – a teratological m3 or, more likely, asupernumerary molar (post-m3) – that is, in taxonomy, ‘‘a non-adequate type’’ as said by Roger et al. (1994). Anyway, to provedefinitely that it is a supernumerary tooth implies an observationin situ in a mandible, which will never be possible for FSL 213759.Yet, the doubt is sufficiently important to conclude that no speciesshould bear the binom Gomphotherium pygmaeum or Choerolo-
phodon pygmaeus. The opposite conclusion would be a taxonomicdiagnosis based on the association of strange characters of thismolar from Algeria. Hence, again, this position could only becorroborated by the discovery of an identical molar in a mandibleassociated to other molars, a situation which did not occur up tothe present. The odyssey of the ‘‘mutation pygmaeus’’ – oneexample among others – in relation to the contingency ofpalaeontology, is an illustration of what Eco (1990) labelled insemiotics ‘‘the limits of interpretation’’.
Actually, the description of the different characters ofFSL 213759 shows that this tooth is beyond a normal taxonomicinterpretation, the reason why we label it ‘‘incomplete right?supernumerary tooth (post-m3)’’. Because of this, we surmise thatour anatomical hypothesis, as all previous taxonomic hypotheses,will not necessarily reach a large consensus. As a consequence, thename coined by Deperet should apply only to the holotypeFSL 213759, which is another way to say that Mastodon angustidens
pygmaeus Deperet, 1897 is a nomen dubium.
Acknowledgements
The organizers of the OCIM colloquium held in Toulouse(Southern France; Nov. 15–16, 2011), especially F. Duranthon(Toulouse) and E. Maczek (Dijon) are deeply thanked: the decisionto write this paper was taken by P.T. due to discussions during theexcursion at Sansan (Gers) which was part of the colloquium. Abel

P. Tassy et al. / Geobios 46 (2013) 329–334334
Prieur (Universite Claude Bernard Lyon 1) forcefully cooperated insending cast and, last but not least, he and Emmanuel Robertallowed FSL 213759 to leave Lyon to be scanned in Paris. Manythanks to Philippe Loubry (MNHN) who took the picture of 1961-5-12, and numerized A426. The work was supported by UMR 7207CNRS-MNHN-UPMC (dir. P. Janvier). The quality of the manuscriptwas greatly improved by three incisive reviewers.
Appendix A. Supplementary data
Supplementary data (3-D model of the molar FSL 213759)associated with this article can be found, in the online version, athttp://dx.doi.org/10.1016/j.geobios.2013.01.003.
References
Andrews, C.W., 1906. A descriptive catalogue of the Tertiary Vertebrata of theFayum Egypt. British Museum (NH), London.
Arambourg, C., 1961. Note preliminaire sur quelques vertebres nouveaux duBurdigalien de Libye. Comptes rendus sommaires de la Societe geologiquede France 1961 (4) 107–108.
Bergounioux, F.M., Crouzel, F., 1956. Presence de Tetralophodon longirostris dans leVindobonien inferieur de Tunisie. Bulletin de la Societe Geologique de FranceSer. 6 6, 547–558.
Bergounioux, F.M., Crouzel, F., 1959. Nouvelles observations sur le petit Mastodontedu Cartennien de Kabylie. Comptes rendus sommaires de la Societe geologiquede France 1959 (5) 101–102.
Coppens, Y., Maglio, V.J., Madden, C.T., Beden, M., 1978. Proboscidea. In: Maglio,V.J., Cooke, H.B.S. (Eds.), Evolution of African Mammals. Harvard UniversityPress, Cambridge, pp. 336–367.
Dalloni, M., 1917. Contribution a l’etude des terrains miocenes de l’Algerie. LeCartennien des environs de Miliana. Bulletin de la Societe Geologique de France,Ser 4 17, 174–207.
Deperet, C., 1897. Decouverte du Mastodon angustidens dans l’etage Cartennien deKabylie. Bulletin de la Societe Geologique de France 25, 518–521.
Deperet, C., 1907. Les transformations du monde animal. Flammarion, Paris.Eco, U., 1990. I limiti dell’interpretazione. Bompiani, Milano.Gaziry, A., 1987. Remains of Proboscidea from the Early Pliocene of Sahabi, Libya. In:
Boaz, N.T., El-Arnauti, A., Gaziry, A.W., de Heinzelin, J., Boaz, D.D. (Eds.), Neo-gene Paleontology and geology of Sahabi. Alan R. Liss, New York, pp. 183–203.
Geraads, D., 1989. Vertebres fossiles du Miocene superieur du Djebel Krechem elArtsouna (Tunisie Centrale). Comparaisons biostratigraphiques. Geobios 22,277–801.
Ginsburg, L., Bulot, C., 2000. Le cadre stratigraphique du site de Sansan. In: Ginsburg,L. (Ed.), La faune miocene de Sansan et son environnement, 183. Memoires duMuseum national d’Histoire naturelle, pp. 39–67.
Ginsburg, L., Muizon, C., de Tassy, P., 1975. Decouverte d’un important gisement amastodontes dans le Miocene moyen de Simorre (Gers). Comptes rendus del’Academie des Sciences, Paris 280–D, 1547–1549.
Ginsburg, L., Telles Antunes, M., 1966. Considerations sur les mastodontes duBurdigalien de Lisbonne et des Sables de l’Orleanais (France). Revista daFaculdade de Ciencias de Lisboa Ser 2 14, 135–150.
Hanks, J., 1979. A Struggle for survival. The elephant problem. Country Life Books,London.
Harris, J.M., 1971. Chemical preparation of gypsum encrusted vertebrate material.Journal of Paleontology 45, 350.
Lartet, E., 1859. Sur la dentition des proboscidiens fossiles (Dinotherium, Masto-dontes et Elephants) et sur la distribution geographique et stratigraphique deleurs debris en Europe. Bulletin de la Societe Geologique de France Ser 2 16,469–516.
Laws, R.M., Parker, I.S.C., Johnstone, R.C.B., 1975. Elephants and their habitats. Theecology of elephants in North Bunyoro Uganda. Clarendon Press, Oxford.
Lydekker, R., 1884. Additional Siwalik Perissodactyla and Proboscidea. Memoirs ofthe Geological Survey of India. Palaeontologia Indica 10 (3) 1–34.
MacInnes, D.G., 1942. Miocene and Post-Miocene Proboscidea from East Africa.Transactions of the Zoological Society of London 25, 33–106.
Matsumoto, H., 1924. A revision of Palaeomastodon dividing it into two genera, andwith a description of two new species. Bulletin of the American Museum ofNatural History 50, 1–58.
Osborn, H.F., 1936. Proboscidea. Moeritherioidea, Deinotherioidea, Mastodontoi-dea, 1. The American Museum Press, New York.
Pean, S., Pathou-Mathis, M., Tassy, P., 1998. Agents climatiques et edaphiquesintervenant dans la mise en place d’un assemblage osseux : taphonomied’un gisement a mastodontes du Miocene (En Pejouan, Bassin d’Aquitaine,France). XIIIth Congress UISPP Proceedings, Forli 1996 6 (1), pp. 201–208.
Pickford, M., 2004. Partial dentition and skeleton of Choerolophodon pygmaeus(Deperet) from Ngenyin, Tugen Hills, Kenya: resolution of a century old enigma.
Zona Arqueologica: Miscelanea en Homenaje a Emiliano Aguirre. Paleontologia2, 429–463.
Pickford, M., 2005. Choerolophodon pygmaeus (Proboscidea: Mammalia) fromthe Middle Miocene of southern Africa. South African Journal of Science 101,175–177.
Roger, J., Pickford, M., Thomas, H., Lapparent de Broin, F., de Tassy, P., Van Neer, W.,Bourdillon-de-Grissac, C., Al-Busaidi, S., 1994. Decouverte de vertebres fossilesdans le Miocene de la region du Huqf au Sultanat d’Oman. Annales de Paleon-tologie 80, 253–273.
Roth, V.L., 1989. Fabricational noise in elephant dentitions. Paleobiology 15, 165–179.
Sanders, W.J., Gheerbrant, E., Harris, J.M., Saegusa, H., Delmer, C., 2010. Proboscidea.In: Werdelin, L., Sanders, W.J. (Eds.), Cenozoic Mammals of Africa. University ofCalifornia Press, Berkeley, pp. 161–251.
Sanders, W.J., Kappelman, J., Rasmussen, D.T., 2004. New large-bodied mammalsfrom the late Oligocene site of Chilga Ethiopia. Acta Palaeontologica Polonica 49,365–392.
Savage, D.E., Russell, D.E., 1983. Mammalian palaeofaunas of the world. Addison-Wesley, Reading.
Savage, R.J.G., 1971. Review of the fossil mammals of Libya. In: Gray, C. (Ed.),Symposium of the geology of Libya. University of Libya, Tripoli, pp. 215–225.
Savage, R.J.G., Hamilton, W.R., 1973. Introduction to the Miocene mammal faunas ofGebel Zelten Libya. Bulletin of the British Museum (Natural History) Geology22, 515–527.
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T.,Preibisch, S., Rueden, C., Saalfeld, S., Schmid, B., Tinevez, J.-Y., White, D.J.,Hartenstein, V., Eliceiri, K., Tomancak, P., Cardona, A., 2012. Fiji: an open-sourceplatform for biological-image analysis. Nature Methods 9, 676–682.
Shoshani, J., Golenberg, E.M., Yang, H., 1998. Elephantidae phylogeny: morphologi-cal versus molecular results. Acta Theriologica Supplement 5, 89–122.
Shoshani, J., Tassy, P., 1996. Appendix C.1. In: Shoshani, J., Tassy, P. (Eds.), TheProboscidea. Evolution and Palaeoecology of Elephants and Their Relatives.Oxford University Press, Oxford.
Shoshani, J., Walter, R.C., Abraha, M., Brehe, S., Tassy, P., Sanders, W.J., Marchant,G.H., Libsekal, Y., Ghirmai, T., Zinner, D., 2006. A proboscidean from the lateOligocene of Eritrea, a « missing link » between early Elephantiformes andElephantimorpha, and biogeographic implications. Proceedings of the NationalAcademy of Sciences of the United States 103, 17296–17301.
Tassy, P., 1975. Valeur phylogenetique et taxonomique du cement coronaire chezles mastodontes miocenes (Proboscidea, Mammalia). Comptes rendus de l’Aca-demie des Sciences, Paris 280–D, 1463–1466.
Tassy, P., 1986. Nouveaux Elephantoidea (Mammalia) dans le Miocene du Kenya ;essai de reevaluation systematique. Cahiers de paleontologie – Travaux depaleontologie est-africaine. Editions du CNRS, Paris.
Tassy, P., 1996. Growth and sexual dimorphism among Miocene elephantoids: theexample of Gomphotherium angustidens. In: Shoshani, J., Tassy, P. (Eds.), TheProboscidea. Evolution and Palaeoecology of Elephants and Their Relatives.Oxford University Press, Oxford, pp. 92–100.
Tobien, H., 1972. Status of the genus Serridentinus Osborn 1923 (ProboscideaMammalia) and related forms. Mainzer geowissenschaftlichen Mitteilungen1, 143–191.
Tobien, H., 1973. On the evolution of mastodonts (Proboscidea Mammalia). Part 1:The bunodont trilophodont groups. Notizblatt des Hessischen Landesamtes furBodenforschung zu Wiesbaden 101, 202–276.
Tobien, H., 1976. Zur palaontologischen Geschichte der Mastodonten (ProboscideaMammalia). Mainzer geowissenschaftlichen Mitteilungen 5, 143–225.
Tobien, H., 1978. The structure of the mastodont molar (Proboscidea, Mammalia).Part 3: The Oligocene mastodont genera Palaeomastodon Phiomia and the Eo/Oligocene paenungulate Moeritherium. Mainzer geowissenschaftlichen Mittei-lungen 6, 177–208.
Tobien, H., Chen, G., Li, Y., 1986. Mastodonts (Proboscidea Mammalia) from the LateNeogene and Early Pleistocene of the People’s Republic of China. Mainzergeowissenschaftlichen Mitteilungen 15, 119–181.
Van Essen, H., 2004. A supernumerary tooth and an odontoma attributable toMammuthus primigenius (Blumenbach 1799) (Mammalia, Proboscidea) fromThe Netherlands, and various related finds. Acta zoologica cracoviensia 47,81–121.
Viret, J., 1962. Sur une mandibule d’Anancus arvernensis Croiz. et Job. a quatremolaires, du gisement de Saint-Laurent des Arbres (Gard) In: Problemes actuelsde paleontologie (Evolution des vertebres), Colloques Internationaux du Centrenational de la recherche scientifique N8104. Editions du CNRS, Paris, pp. 281–285.
Waagen, W., 1869. Die Formenreihe des Ammonites subradiatus. Versuch einerpalaontologischen Monographie. In: Benecke, E.W. (Ed.), Geognostisch –palaontologische Beitrage, Band 2, R. Oldenburg, Munchen, pp. 179–256.
Warren, J.C., 1855. Description of a skeleton of the Mastodon giganteus of NorthAmerica. John Wilson and Son, Boston.
Welcomme, J.L., 1993. Le proboscidien du Mazet et les faunes de vertebres con-tinentaux du Burdigalien de la Basse Vallee de l’Herault (Languedoc, Sud de laFrance). Diplome de l’Ecole Pratique des Hautes Etudes, Montpellier (unpub-lished).
Welcomme, J.L., 1994. Le plus ancien crane de proboscidien d’Europe, Gomphother-ium hannibali nov. sp. (Proboscidea, Mammalia) du Miocene inferieur du Lan-guedoc (France). Comptes rendus de l’Academie des sciences Ser 2 319, 135–140.




















