
A new Haplobunodontidae (Mammalia, Artiodactyla) from the Eocene of Turkey

THE SOUTHWESTERN NATURALIST 49(2):229–238 JUNE 2004
LATE PLEISTOCENE MYLODONT SLOTH PARAMYLODON HARLANI(MAMMALIA: XENARTHRA) FROM ARIZONA
H. GREGORY MCDONALD,* LARRY D. AGENBROAD, AND CAROL MANGANARO HADEN
Geologic Resources Division, National Park Service, P.O. Box 25287, Denver, CO 80225 (HGM)Department of Geology and Quaternary Sciences Program, Northern Arizona University,
Flagstaff, AZ 86011 (LDA, CMH)*Correspondent: [email protected]
ABSTRACT The late Pleistocene ground sloth Paramylodon harlani was widely distributed acrossNorth America, but it is represented in Arizona by only 2 records. These include a nearly completeskeleton from the Richville Gravels near Springerville, Apache County, and a second partial skel-eton from Shonto, Navajo County. Both specimens are from reportedly lacustrine deposits andsuggest that the species was not adapted to xeric conditions. Thus, its distribution in the Southwestand potential for dispersal might have been determined by the presence of permanent watersources.
RESUMEN El perezoso terrestre Paramylodon harlani del Pleistoceno tardıo tenıa una distribu-cion amplia en America del Norte, pero su presencia en Arizona esta documentada con solo dosregistros. Estos incluyen un esqueleto casi completo de los Richville Gravels cerca de Springerville,condado de Apache, y un esqueleto parcial de Shonto, condado de Navajo. Ambos especimenes,segun se dice son de depositos lacustres e indican que la especie no estaba adaptada a condicionessecas. Entonces, su distribucion en el sudoeste de USA y su potencial para diseminar podıa habersido determinado por la presencia de fuentes permanentes de agua.
Currently, 3 types of ground sloth are knownfrom the late Pleistocene of Arizona. The bestknown is Nothrotheriops shastensis, representedby numerous specimens, primarily from cavesites, such as Rampart Cave in the Grand Can-yon (Martin et al., 1961). The other 2 species,Megalonyx jeffersonii and Paramylodon harlani,have not been as well documented for Arizona(Lindsay and Tessman, 1974) or for the south-western United States in general. Herein weprovide descriptions of records of P. harlanibased on partial skeletons from near Shonto,Navajo County, and Springerville, ApacheCounty, Arizona. These records expand ourknowledge of the distribution of this species inthe Pleistocene, and the context from whichthey were recovered provides insights into thepaleoecology of Harlan’s ground sloth.
LOCALITY DESCRIPTIONS AND GEOLOGY Greg-ory (1917) first described the geology of theShonto area (listed by him as Shato Springs),providing a faunal list based on identificationsmade by R. S. Lull that included Megalonyx, Ele-
phas, Bison?, and Equus. Alan Townsend, of In-scription House, Arizona, discovered the hu-merus of the Shonto Paramylodon reportedhere in 1982. A visit to the site in 1982 by crewsfrom the Geology Department of Northern Ar-izona University resulted in the recovery of ad-ditional parts of the skeleton. The Shonto Par-amylodon and associated fauna were recoveredfrom the side of a stabilized sand dune inShonto Creek Canyon (368349N, 1108399W),Navajo County, Arizona, at an elevation of1,905 m (Museum of Northern Arizona[MNA] Locality 186–1). The dune formed onthe leeward side of a bedrock island near thecenter of the canyon. The deposit containingthe sloth is a remnant of a deltaic deposit in-undated by a perennial lake formed by an im-poundment within the canyon during the latePleistocene. The sloth was found in situ in aclay-rich horizon of the dune, which might in-dicate a former shoreline of the lake. The ageof the Shonto fauna is 30,800 6 1,700 yr BP(GX-10493) based on a conventional carbon-14 date on a sample of bone from the sloth(Agenbroad and Downs, 1984).

230 vol. 49, no. 2The Southwestern Naturalist
FIG. 1 Skull of Paramylodon harlani from Sprin-gerville (MNA V1374). A) dorsal view; B) ventralview; C) left lateral view.
The Springerville fauna comes from sections2, 11, and 12, T10N, R28E, Lyman Lake Quad-rangle, 348169N and 1098209W, Apache County,Arizona (MNA Locality 184–1). The almostcomplete skeleton of Paramylodon harlani re-ported herein was recovered from gravels inthe southwestern portion of section 12 at anelevation of about 1,906 m. The Richville grav-els were deposited behind lava basalts, whichimpounded the Richville reach of the LittleColorado River. These basalts have been datedusing K/Ar at 1.67 6 0.09 mya (Laughlin etal., 1980).
DESCRIPTIONS OF SPECIMENS The sloth fromSpringerville was reported briefly by Brady(1933), and the skeleton was mounted and dis-played at the Museum of Northern Arizona formany years. Its dismantling made it possible toobtain measurements provided in this paper.This paper expands upon the previous reportby McDonald et al. (1994) and provides moredetails.
The Springerville sloth consists of a reason-ably complete skeleton. Parts of the skeletonwere collected on separate occasions, resultingin separate catalog numbers. It is assumed thatonly a single adult is represented becausethere is no duplication of skeletal elements, allof the recovered material is from a similarstage of ontogenetic development, and allspecimens were collected in the same area.The catalog number MNA V1374 was assignedto the skull and jaw (Figs. 1, 3), atlas, rightscapula, right humerus, right ulna, left radius,pelvis, right femur, left tibia, left astragalus, leftcalcaneum, left third metatarsal, right fourthmetatarsal, vertebrae, and ribs. Other cata-loged elements of skeleton at the Museum ofNorthern Arizona include left third metacar-pal P1.1231, sesamoid P1.1235, stylohyalP1.1242, proximal phalanx from manusP1.1238 and P1.1232, left cuboid P1.1233, leftlunar P1.1240, right clavicle P1.1241, rightfourth metatarsal with phalanges, P1.1243, leftfourth metacarpal P1.1236, and unguals fromdigit 1 of manus P1.1228 and P1.1230.
The Shonto sloth, cataloged by the North-ern Arizona University Quaternary ScienceProgram (NAUQSP 12523), consists of theskull (Fig. 2) and lower jaw (Fig. 3), the cer-vical series, both humeri, and the left astraga-lus (MNA P1.1291, G2.6898).
Descriptions of individual bones of the skel-eton of P. harlani (Stock, 1925) were based ona large sample from Rancho La Brea, Los An-geles County, California. Both specimens fromArizona compare well with the Rancho La Breamaterial and can be assigned readily to P. har-lani. Because the measurements provided byStock (1925) were from isolated bones, and be-cause associated skeletons of P. harlani are un-common, we provide measurements of individ-ual bones in Appendix 1.
Stock (1925) noted the variability in thepresence of the caniniform in skulls of P. har-lani. Based on the sample of 50 skulls fromRancho La Brea, he reported that 21 individ-uals retained the caniniform on both sides, 14lacked the caniniform on both sides, 5 hadonly a right caniniform, 7 possessed only the
June 2004 231McDonald et al.—Late Pleistocene mylodont sloth from Arizona
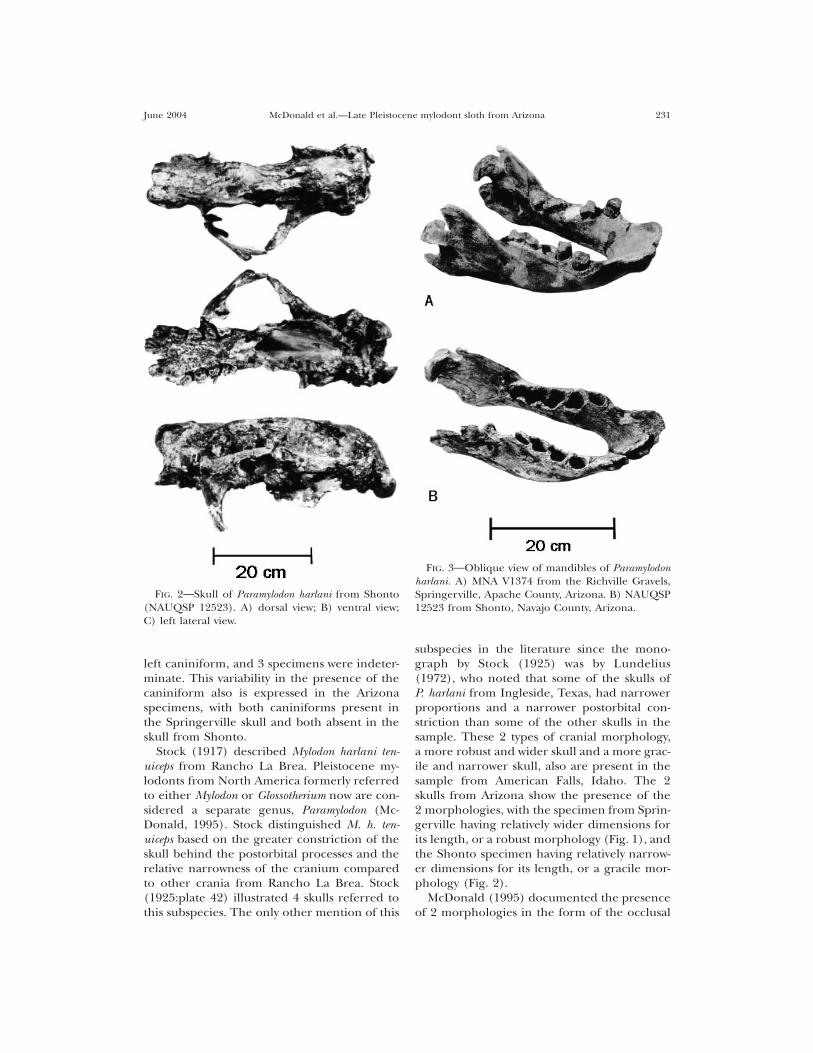
FIG. 2 Skull of Paramylodon harlani from Shonto(NAUQSP 12523). A) dorsal view; B) ventral view;C) left lateral view.
FIG. 3 Oblique view of mandibles of Paramylodonharlani. A) MNA V1374 from the Richville Gravels,Springerville, Apache County, Arizona. B) NAUQSP12523 from Shonto, Navajo County, Arizona.
left caniniform, and 3 specimens were indeter-minate. This variability in the presence of thecaniniform also is expressed in the Arizonaspecimens, with both caniniforms present inthe Springerville skull and both absent in theskull from Shonto.
Stock (1917) described Mylodon harlani ten-uiceps from Rancho La Brea. Pleistocene my-lodonts from North America formerly referredto either Mylodon or Glossotherium now are con-sidered a separate genus, Paramylodon (Mc-Donald, 1995). Stock distinguished M. h. ten-uiceps based on the greater constriction of theskull behind the postorbital processes and therelative narrowness of the cranium comparedto other crania from Rancho La Brea. Stock(1925:plate 42) illustrated 4 skulls referred tothis subspecies. The only other mention of this
subspecies in the literature since the mono-graph by Stock (1925) was by Lundelius(1972), who noted that some of the skulls ofP. harlani from Ingleside, Texas, had narrowerproportions and a narrower postorbital con-striction than some of the other skulls in thesample. These 2 types of cranial morphology,a more robust and wider skull and a more grac-ile and narrower skull, also are present in thesample from American Falls, Idaho. The 2skulls from Arizona show the presence of the2 morphologies, with the specimen from Sprin-gerville having relatively wider dimensions forits length, or a robust morphology (Fig. 1), andthe Shonto specimen having relatively narrow-er dimensions for its length, or a gracile mor-phology (Fig. 2).
McDonald (1995) documented the presenceof 2 morphologies in the form of the occlusal

232 vol. 49, no. 2The Southwestern Naturalist
FIG. 4—Scatter diagram for humeri of Paramylodonharlani showing relationship between maximumlength and latitude.
FIG. 5—Map showing a reconstruction of full-gla-cial vegetation in Arizona 17,000 to 23,000 years agofrom Martin and Mehringer (1965). White is desert,stippled is sagebrush/chaparral, hatched is pinon-juniper woodland, cross-hatched is yellow pine park-land, solid black is spruce, fir, pine forest, and thewhite area within the black is subalpine to alpinehabitat. Shonto and Springerville localities are as-sociated with areas interpreted as parkland habitat.
surface of the caniniform in P. harlani fromLeisey 1A, Florida, and suggested that these in-dicated sexual dimorphism in the species.These 2 occlusal morphologies also are presentin samples from Rancho La Brea and Ameri-can Falls, and in other samples of P. harlanirepresented by multiple specimens of the ca-niniform from a single locality. Based on thesamples mentioned above, and because the 2cranial and caniniform morphologies arefound together in faunas with multiple skulls,it is unlikely that they represent 2 subspeciesthat completely overlap in range. In contrastto Stock (1925), McDonald interpreted thepresence of the 2 morphologies to be indica-tive of sexual dimorphism in the species ratherthan subspecific variation. At this time, it is notpossible to equate a particular morphologywith a specific sex. Additional study and an in-dependent means of determining the genderof ground sloths are needed.
Utilizing the humerus as an indicator of size,the lengths of the 2 Arizona specimens are sim-ilar to those of individuals from Rancho LaBrea (Fig. 4), and both are smaller than indi-viduals from farther north in Idaho. Remainsof another sloth, Megalonyx jeffersonii, might bedemonstrating a Bergmann’s response, withthe average size of the animal changing withlatitude (McDonald et al., 2000). While thepresent sample for P. harlani is not robust, itdoes show a trend, with the average size of in-dividuals larger in northern populations.
DISCUSSION Previous studies of the Shontofauna included those by Hay (1927), Saunders(1970), and Agenbroad and Mead (1989) on
Mammuthus, that of Agenbroad and Downs(1984) on Tapirus cf. T. merriami, and the fau-nal list by Lindsay and Tessman (1974). TheShonto fauna also contained Lepus, Peromyscus,Thomomys, a sciurid, Urocyon, Felis, Equus, Cer-vus, and Ovis. The fauna associated with theSpringerville Paramylodon in the Museum ofNorthern Arizona included Equus, Camelops, anantilocaprid, a cervid, Mammuthus, and Mega-lonyx jeffersonii.
Since the work of Stock (1925), P. harlani hasbeen considered a grazer in open country. Na-ples (1989) suggested that Paramylodon was ca-pable of using foods of lower nutritional valuethan other grazers and might better be consid-ered a browser-grazer rather than a grazeronly. Many of the taxa found with Paramylodonin Arizona and in other faunas included spe-cies of mammoth, horse, and camel, speciescommonly associated with open habitat. Eachof these groups also seems to have been labilein its food habits, including browse as well asgraze in the diet (Akersten et al., 1988; Dom-pierre and Churcher, 1996).
There are many physical and biological fac-tors that determine the distribution of a spe-

June 2004 233McDonald et al.—Late Pleistocene mylodont sloth from Arizona
cies. The identification of these limiting factorsis often difficult for extant species and mightbe nearly impossible for extinct ones. However,it is possible to make some reasonable infer-ences about the paleoecology of an extinct spe-cies based on a critical examination of its dis-tributional patterns.
Paramylodon is distributed widely across theUnited States, with numerous records west ofthe continental divide, except for the ColoradoPlateau, where it is relatively rare. Besides thespecimens from Arizona, the only other re-cords of the taxon from or immediately adja-cent to the Colorado Plateau are from SilverCreek, Summit County, Utah, based on skeletalmaterial (Miller, 1976) and hair recoveredfrom Bechan Cave (Davis et al., 1984; Mead etal., 1986). Elevation does not seem to havebeen a limiting factor. Paramylodon has beenfound at higher elevations, such as the SilverCreek fauna in Utah (1,952 m) and at the Mag-na Site in Saguache County, Colorado, (2,330m) east of the continental divide (McDonald,unpubl. data).
Remains of Paramylodon west of the conti-nental divide show that it was absent in areasnow considered deserts, although numerousrecords of another sloth, Nothrotheriops, areknown from these regions. McDonald (1993)listed 62 Rancholabrean localities in Californiafor P. harlani, noting that its distribution wasbiased towards the coastal lowlands and that itwas absent from the inland deserts, areas withnumerous faunas that include Nothrotheriops.Paramylodon and Nothrotheriops rarely are foundin the same fauna (McDonald, 1996), but theirdistributions were not totally exclusive. How-ever, in the few faunas that included both gen-era and a record of vegetation, such as RanchoLa Brea, the plants differed markedly fromthose preserved in the dung of Nothrotheriops incaves in the arid Southwest. These plant re-mains suggest a significant difference in thetypes of vegetation preferred by the 2 speciesof sloth, and that their co-occurrence was pos-sible only in a few areas with vegetation suit-able for both.
The Shonto and Springerville localities arein areas mapped (Fig. 5) as parkland with yel-low pine (Pinus ponderosa) during the full gla-cial (Martin and Mehrenger, 1965). Whereasthe vegetation map was extrapolated from anumber of widely distributed records, the area
of inferred vegetation for that part of Arizonacoincides with both localities with Paramylodon.This coincidence provides some support forour interpretation that the ecology of P. harlaniincluded a preference for open grassland orparkland habitat.
Paramylodon harlani is virtually absent fromall 4 of the current biologically-defined NorthAmerican deserts (Spaulding et al., 1983). Theonly exceptions are a few records along the pe-riphery of the Great Basin, including CarsonCity at the western edge of the basin near thefoothills of the Sierra Nevada Mountains, anda small area on the Snake River Plain (9 local-ities). The Carson City record is based primar-ily on tracks preserved in sediments associatedwith the shoreline of a pluvial lake, and theIdaho records are in close proximity to theSnake River. As suggested by the records fromArizona, the availability of a permanent watersource in southern Idaho and western Nevadaprovided the appropriate local environmentfor the species. Riparian vegetation associatedwith rivers and lakes would not only have beenimportant habitat, but also might have servedas corridors that permitted dispersal. Recordsof the lineage of P. harlani in Idaho begin inthe Blancan, about 2.5 mya, from sites associ-ated with Lake Idaho, which drained into Cal-ifornia at that time (Taylor, 1985). The riversystem that connected Lake Idaho with Cali-fornia was undoubtedly a dispersal route foraquatic species and also could have providedsuitable riparian habitat for terrestrial species.
Agenbroad and Downs (1984) argued thatriparian forests formed an interconnected net-work traversing the Pleistocene grasslands anddeserts of the Southwest. This network of ri-parian habitat would have provided links be-tween faunas of the Basin and Range Provinceto those of the Colorado Plateau, California,and elsewhere. Such a network might explainthe distribution of anomalous species, such asthe extinct tapir (Tapirus cf. T. merriami), incurrent semiarid and desert regions. Mc-Donald (1996) used a similar argument to ex-plain the distribution of another browsingground sloth, Megalonyx jeffersonii, in the South-west, as did Gillette et al. (1999) to infer themode of dispersal of Megalonyx into the Bon-neville Basin. Megalonyx is part of the Springer-ville fauna with Paramylodon.
The presence of tapir (Tapirus cf. T. merria-

234 vol. 49, no. 2The Southwestern Naturalist
mi) in the Shonto fauna (Agenbroad andDowns, 1984) indicates riparian habitat. Asnoted by Agenbroad and Downs (1984), thepollen-based environmental reconstruction forthe Southwest during the late Pleistocene sug-gests a broad expanse of grasslands with asso-ciated xeric oak-conifer woodlands in what arenow deserts (Martin and Mehringer, 1965).These woodlands would have been densestalong perennial streams and permanent water.The Springerville sloth was preserved in grav-els associated with the Little Colorado Riverand with a lake formed by an impoundmentresulting from lava flows. The Shonto speci-men also was associated with permanent water.Based on records from Arizona and elsewhere,it seems that the distribution of the species inxeric areas was controlled by the presence ofpermanent water. Although water seems to beone factor controlling the distribution of theanimal, it probably was not the single primarylimiting factor, because types of vegetationseem to have been equally critical. The GreatBasin contained numerous permanent pluviallakes during the Pleistocene (Morrison, 1965),yet Paramylodon was absent from the entire ba-sin except for the 2 areas on the periphery not-ed above. Its absence indicates that multiplefactors were needed to permit its survival in anarea. The distribution and potential dispersalof Harlan’s ground sloth in Arizona and Idahoseem to follow similar patterns, and these areasboth seem to have provided the necessary com-bination of water and appropriate vegetation.
We recognize that the size, extent, and typesof vegetation in these deserts have been dy-namic, changing throughout the Pleistocene(Spaulding et al., 1983; Van Devender, 1986),and that the present aridity in these areasmight have been less severe during the Pleis-tocene. Evidence indicates that during the latePleistocene the 4 present North American de-serts had less extreme temperatures, a greatervariety of vegetation, and more effective mois-ture. Yet, despite these less desert-like condi-tions, P. harlani is absent from these deserts,although its distribution west of the continen-tal divide extends all around them. In the fu-ture P. harlani might be recovered from withinthese deserts, but such records most likely willreflect specific restricted or localized condi-tions, similar to those just discussed, with both
a dispersal corridor and an area suitable forsupporting a local population.
Both Arizona records, along with the patternof distribution elsewhere, support the interpre-tation that P. harlani was not adapted to xericconditions. If Paramylodon was restricted to aparticular set of environmental conditions in-cluding specific types of vegetation, then itsdistribution might have been discontinuous,composed of numerous, small, isolated popu-lations in geographically restricted habitat.These habitats, particularly in the westernUnited States, might have been connected viaa tenuous network of narrow corridors alongwaterways. If our interpretation is correct, thenthese small, localized populations would havebeen particularly vulnerable to any changes inthe local habitat. Any explanation of the ex-tinction of Harlan’s ground sloth at the end ofthe Pleistocene needs to take into accountthese and other details of the ecology of thespecies.
Swetnam et al. (1999) documented the de-cline in the extent of grasslands in Arizonafrom 12,000 B.P. to present, and this declinewas paralleled by an increase in deserts. Thedecline of grassland, the presumed preferredhabitat of Paramylodon, and fluctuations in veg-etation no doubt influenced the distribution ofHarlan’s ground sloth and other fauna. Like-wise, any possibility of dispersal to appropriatehabitat could have been disrupted easily if Har-lan’s ground sloth depended on narrow corri-dors of vegetation along permanent waterways.Although it is always dangerous to argue fromnegative evidence, there are sufficient recordsfor the species that the gaps in its distributionin the western United States apparently arereal and not merely a sampling artifact. Assuch, the distributional records reflect specificattributes of the ecology of the species, withany exceptions to the general pattern reflect-ing suitable local conditions. As is often thecase, it might be these exceptions that provethe rule.
We would like to thank A. Townsend of Inscrip-tion House, Arizona for his discovery of the Shontospecimen and for reporting it to L. Agenbroad. M.Schaefer helped with the Spanish abstract. We wouldespecially like to thank W. Akersten and an anony-mous reviewer for comments and suggestions thathave improved the quality of our manuscript.

June 2004 235McDonald et al.—Late Pleistocene mylodont sloth from Arizona
LITERATURE CITED
AGENBROAD, L. D., AND W. R. DOWNS. 1984. A robusttapir from northern Arizona. Journal of the Ari-zona-Nevada Academy of Science 19:91–99.
AGENBROAD, L. D., AND J. I. MEAD. 1989. Quaternarygeochronology and distribution of Mammuthuson the Colorado Plateau. Geology 17:861–864.
AKERSTEN, W. A., T. M. FOPPE, AND G. T. JEFFERSON.1988. New source of dietary data for extinct her-bivores. Quaternary Research 30:92–97.
BRADY, L. F. 1933. A mounted skeleton of a groundsloth. Museum Notes, Museum of Northern Ari-zona 6:19–21.
DAVIS, O. K., L. D. AGENBROAD, P. S. MARTIN, AND J.I. MEAD. 1984. The Pleistocene dung blanket ofBechan Cave, Utah. Carnegie Museum of NaturalHistory Special Publication 8:267–282.
DOMPIERRE, H., AND C. S. CHURCHER. 1996. Premax-illary shape as an indicator of the diet of sevenextinct Late Cenozoic New World camels. Journalof Vertebrate Paleontology 16:141–148.
GILLETTE, D. D., H. G. MCDONALD, AND M. C. HAY-DEN. 1999. The first record of Jefferson’s groundsloth, Megalonyx jeffersonii, in Utah (Pleistocene,Rancholabrean Land Mammal Age). Utah Geo-logical Survey Miscellaneous Publication 99–1:509–521.
GREGORY, H. E. 1917. Geology of the Navajo Coun-try: a reconnaissance of parts of Arizona, NewMexico, and Utah. United States Geological Sur-vey Professional Paper 93.
HAY, O. P. 1927. The Pleistocene of the western re-gion of North American and its vertebrated ani-mals. Carnegie Institution of Washington Publi-cation 322B.
LAUGHLIN, A. W., P. E. DAMON, AND M. SHAFIQULLAH.1980. New K-Ar dates from the Springerville vol-canic field, central Jemez zone, Apache County,Arizona. Isochron/West Number 29:3–4.
LINDSAY, E. H., AND N. T. TESSMAN. 1974. Cenozoicvertebrate localities and faunas in Arizona. Jour-nal of the Arizona Academy of Science 9:3–24.
LUNDELIUS, E. L., JR. 1972. Fossil vertebrates fromthe late Pleistocene Ingleside fauna, San PatricioCounty, Texas. Bureau of Economic Geology Re-port of Investigations Number 77:1–74.
MARTIN, P. S., AND P. J. MEHRINGER, JR. 1965. Pleis-tocene pollen analysis and biogeography of theSouthwest. In: Wright, H. E., Jr., and D. G. Frey,editors. The Quaternary of the United States.Princeton University Press, Princeton, New Jer-sey. Pp. 433–451.
MARTIN, P. S., B. E. SABELS, AND D. SHUTLER, JR. 1961.Rampart Cave coprolite and ecology of the Shas-ta ground sloth. American Journal of Science259:102–127.
MCDONALD, H. G. 1993. Harlan’s ground sloth, Glos-sotherium harlani, from Pauba Valley, RiversideCounty, California. San Bernardino County Mu-seum Association Special Publication 93–1:101–103.
MCDONALD, H. G. 1995. Gravigrade xenarthransfrom the early Pleistocene Leisey Shell Pit 1A,Hillsborough County, Florida. Bulletin of theFlorida Museum of Natural History 37, Part 2:345–374.
MCDONALD, H. G. 1996. Biogeography and paleo-ecology of ground sloths in California, Arizonaand Nevada. San Bernardino County Museum As-sociation Quarterly 43:61–65.
MCDONALD, H. G., L. D. AGENBROAD, AND C. MAN-GANARO. 1994. Mylodont sloth in northern Ari-zona. Journal of Vertebrate Paleontology 14, Sup-plement:37A.
MCDONALD, H. G., C. R. HARINGTON, AND G. DE
IULIIS. 2000. The ground sloth, Megalonyx, fromPleistocene deposits of the Old Crow Basin, Yu-kon, Canada. Arctic 53:213–220.
MEAD, J. I., L. D. AGENBROAD, O. K. DAVIS, AND P. S.MARTIN. 1986. Dung of Mammuthus in the aridSouthwest, North America. Quaternary Research25:121–127.
MILLER, W. E. 1976. Late Pleistocene vertebrates ofthe Silver Creek local fauna from north centralUtah. Great Basin Naturalist 36:387–424.
MORRISON, R. B. 1965. Quaternary geology of theGreat Basin. In: Wright, H. E., Jr. and D. G. Frey,editors. The Quaternary of the United States.Princeton University Press, Princeton, New Jer-sey. Pp. 265–285.
NAPLES, V. L. 1989. The feeding mechanism in thePleistocene ground sloth, Glossotherium. NaturalHistory Museum of Los Angeles County Contri-butions in Science 415:1–23.
SAUNDERS, J. J. 1970. The distribution and taxonomyof Mammuthus in Arizona. Unpublished M.S. the-sis, University of Arizona, Tucson.
SPAULDING, W. G., E. B. LEOPOLD, AND T. R. VAN DE-VENDER. 1983. Late Wisconsin paleoecology of theAmerican Southwest. In: Porter, S. C., editor. LateQuaternary environments of the United States,volume 1, the late Pleistocene. University of Min-nesota Press, Minneapolis. Pp. 259–293.
STOCK, C. 1917. Further observations on the skullstructure of mylodont sloths from Rancho LaBrea. University of California Publications, Bul-letin of the Department of Geology 10:165–178.
STOCK, C. 1925. Cenozoic gravigrade edentates ofwestern North America with special reference tothe Pleistocene Megalonychinae and Mylodonti-dae of Rancho La Brea. Carnegie Institution ofWashington Publication Number 331.
SWETNAM, T. W., C. D. ALLEN, AND J. L. BETANCOURT.1999. Applied historical ecology: using the past

236 vol. 49, no. 2The Southwestern Naturalist
APPENDIX 1—Measurements (mm) of the skeletons of Paramylodon harlani from Springerville, ApacheCounty, and Shonto, Navajo County, Arizona. Numbers in parentheses are approximate. Measurementsmarked with an ‘‘a’’ are taken from the alveoli.
MeasurementSpringervilleMNA V1374
ShontoNAUQSP 12523
Skull and upper dentition
Skull length anterior process of maxilla to occipital condyles 469.7 513.7Length of skull from anterior edge of caniniform to occipital condyles 440.0 500.0Distance from posterior edge of last molariform to occipital condyle 286.0 333.2Alveolar length from front of caniniform to back of last molariform 157.5 —Alveolar length from front of first molariform to back of last molari-
form 115.7 129.8Width of skull between lachrymal foraminaWidth of occiput across mastoid processesWidth across the occipital condylesLength of occipital condylesHeight of occiput
128.2191.8134.252.3
119.5
(93)(169)(136)
57.2129.4
Length of temporal fossa from post-orbital process to anterior edge oc-cipital crest
Anteroposterior length of caniniformMediolateral width of caniniformAnteroposterior lenght of first molariformMediolateral width of first molariformAnteroposterior length of second molariform along medial side
251.6
22.121.626.318.226.0
253.9
—————
Mediolateral width of second molariform along anterior edgeMediolateral width of second molariform along posterior edgeAnteroposterior length of third molariform along medial sideMediolateral width of third molariform along anterior edgeMediolateral width of third molariform along posterior edgeAnteroposterior length of fourth molariform
26.634.521.629.532.330.7
——————
Mediolateral width of anterior lobe of fourth molariformMediolateral width of posterior lobe of fourth molariform
25.416.6
——
Mandible and lower dentition
Mandible length anterior edge of symphysis to posterior edge of angu-lar process 399 (421)
Mandible length anterior edge of symphysis to posterior edge of con-dyle 378.4 (396)
Length from anterior edge of symphysis to anterior edge of first cheektooth 107.4 107.4
Length from anterior edge of symphysis to posterior edge of last cheektooth 240.3 257.2
Alveolar length of tooth rowDepth of mandible below the last molariformMediolateral width of mandibular spout
117.295.391.2
(115)103.4—
to manage for the future. Ecological Applications9:1189–1206.
TAYLOR, D. W. 1985. Evolution of freshwater drain-ages and molluscs in western North America. In:Smiley, C. J., editor. Late Cenozoic history of thePacific Northwest. Pacific Division of the Ameri-can Association for the Advancement of Science,San Francisco, California. Pp. 265–321.
VAN DEVENDER, T. R. 1986. Pleistocene climates and
endemism in the Chihuahuan Desert flora. In:Barlow, J. C., A. M. Powell, and B. N. Timmer-mann, editors. Second Symposium on Resourcesof the Chihuahuan Desert Region. ChihuahuanDesert Research Institute, Alpine, Texas.
Submitted 16 August 2002. Accepted 9 June 2003.Associate Editor was Cheri A. Jones.

June 2004 237McDonald et al.—Late Pleistocene mylodont sloth from Arizona
APPENDIX 1—Continued
MeasurementSpringervilleMNA V1374
ShontoNAUQSP 12523
Anteroposterior length of first cheek toothMediolateral width of first caniniform
25.319.7
26.9a18.7a
Anteroposterior length of second molariform tooth along medial side 26.6 27.8aMediolateral width of second molariform along anterior sideMediolateral width of second molariform along posterior sideAnteroposterior length of third molariform along medial side
23.328.915.6
26.7a28.6a17.8a
Anteroposterior length of third molariform along lateral sideMediolateral width of third molariform along anterior sideMediolateral width of third molariform along posterior sideAnteroposterior length of fourth molariformMediolateral width of anterior lobe of fourth molariformMediolateral width of posterior lobe of fourth molariform
15.929.630.957.835.924.9
21.0a30.5a29.3a59.5a34.9a27.6a
Width of isthmus between anterior and posterior lobes of fourth molar-iform 9.4 7.3a
Atlas
Width across transverse processesDorsoventral heightMediolateral width of anterior articulationsMediolateral width of posterior articulationsAnteroposterior width of transverse process
224.183.7
135.9109.786.9
Scapula
Greatest length along scapular spineAnteroposterior length of glenoidMediolateral width of glenoidGreatest width of blade of scapula
320.1125.475.1
427.6
Humerus
Length from middle of head to middle of medial condyleMediolateral width of proximal endAnteroposterior length of proximal endWidth of shaft across deltoid crestMediolateral width of distal endMediolateral width of distal condylesAnteroposterior depth of lateral distal condyleAnteroposterior depth of medial distal condyle
444.8167.5—114.4247.9133.775.7
—
(445)189154.1131.8—133.882.778.5
Ulna
Greatest lengthLength of olecranon process
381.292.7
Length of shaft from anterior edge of radial articular facet to distalend (255)
Depth of ulna at coronoid processMediolateral width across humeral articulationAnteroposterior dimension of distal articulationMediolateral dimension of distal articulation
157.7121.856.555.8
Radius
Length from middle of proximal to middle of distal articular surfaceLength from middle to proximal articular surface to tip ofAnteroposterior dimension of proximal endMediolateral dimension of proximal endAnteroposterior dimension of distal endMediolateral dimension of distal end
229.9282.078.6
(59.5)112.989.9

238 vol. 49, no. 2The Southwestern Naturalist
APPENDIX 1—Continued
MeasurementSpringervilleMNA V1374
ShontoNAUQSP 12523
Femur
Length from head to distal condylesAnteroposterior diameter of headMediolateral diameter of headMediolateral width of proximal endMediolateral width of distal end of epicondylesMediolateral width of condylesMediolateral width of lateral condyleMediolateral width of medial condyle
514.3119.0126.4263.8227.8193.564.476.1
Patella
LengthMediolateral width of proximal endThickness through femoral articular surface
119.2121.163.5
Tibia
LengthMediolateral width of proximal endAnteroposterior width of proximal endMediolateral width of distal endAnteroposterior width of distal end
219.4179.1107.8140.2101.5
Astragalus
Greatest anteroposterior lengthHeight of fibular facetAnteroposterior length of lateral articular surface for tibiaMediolateral width of articular surface for tibia
132.473.5
117.991.9
146.476.7
123.797.4
Calcaneum
Greatest lengthMediolateral width of tuber calcisGreatest dorsoventral heightLength of tuber calcis along ventral sideLength of confluent cuboid-astragalar facetMediolateral width of astragalar facet
194.9113.4118.0137.7120.090.7
Third metatarsal
Greatest length from lateral edge to distal articular surface 75.5Length along medial side from proximal articular surface to middle of
distal articular surfaceMediolateral width of proximal endAnteroposterior length of distal endMediolateral width of distal end
55.257.858.037.5
Measurement MNA P1.1243
Fourth metatarsal
Greatest lengthMediolateral width of proximal endAnteroposterior width of proximal endAnteroposterior length of distal endMediolateral width of distal end
122.856.865.258.939.0
Copyright © 2022 FDOKUMEN




















