
Morcegos do Parque Nacional do Iguaçu, Paraná (Chiroptera, Mammalia
Upload
xn--universit-poitiers-jwbCategory
view
4download
0
Zoological Journal of the Linnean Society, 2003, 138, 477 -493 . W i t h 4 figures
Juvénile cranial anatomy of Nimravidae (Mammalia, Carnivora): biological and phylogenetic implications
S T E P H A N E PEIGNÉ 1* and L O U I S D E BONIS^
^Laboratoire de Paléontologie, Muséum National d'Histoire Naturelle, 8 rue Buffon, 75005 Paris, France '^Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine, UMR 6046 CNRS, Université de Poitiers, 40 av. Recteur-Pineau, 86022 Poitiers Cedex, France.
Received June 2002; accepted for publication December 2002
The anatomy of the spectacular sabretoothed carnivorans bas been studied many times. Their behavioural and bio-mechanical imphcations are wel l l inown i n adults, but not i n juvénile individuals, which are poorly represented i n the fossil record. This work reports on a newly discovered Hoplophoneini (Nimravidae Nimravinae) of 5-6 months of âge, which represents the earliest and one of the most complète juvénile crania discovered. We discuss and compare the juvénile anatomy of Palaeogene nimravids relative to that of other mammals, especially other carnivorans inc luding the Miocène nimravids (i.e. Barbourofelinae). We also discuss the ontogenetic development of this ind i v idua l and other sabretoothed predators, which provides important data for reconstructing the biology of thèse extinct predators. Our study of the juvénile anatomy of the Hoplophoneini reveals an or ig inal association of basic-ran ia l features: the ectotympanic rotâtes away from the cran ium earlier than i n other mammals, which may be an addit ional autapomorphy dist inguishing the Palaeogene Nimrav idae from other Carnivora; w i th in Palaeogene N i m ravidae, at a s imi lar ontogenetic stage, there is a major différence i n development and max imum ventral extension of ossification of the caudal entotympanic; i n contrast to the Miocène n imrav id Barbourofelis, there is no strong rela-tionship between the éruption of the upper canines and the development of other sabretoothed features (especially the mandibular flange) i n the ind iv idua l described. Thèse results confirm the conclusions of récent studios which sug-gested that Palaeogene Nimravidae (Nimravinae) and Miocène Nimravidae (Barbourofelinae) are not closely related. Muscular reconstruction and biomechanical models also confirmed that juvénile indiv iduals were probably not able to k i l l or hunt any prey. They may have fed on carcasses, eating muscles by using the incisors, and associated w i th a pu l l ing or twist ing action of neck. © 2003 The L innean Society of London. Zoological Journal ofthe Linnean Society, 2003, 138, 477-493.
A D D I T I O N A L K E Y W O R D S : Hoplophonein i - muscu lar reconstruct ion - Oligocène - ontogeny - Palaeogene -phosphorites of Quercy - sabretoothed adaptat ion.
Reconstruct ing the behaviour and ecology of extinct an imais is a major goal i n palaeobiology. A m o n g extinct mammals , the sabretoothed predators have attracted m u c h attent ion from the scientific commu-ni ty because of the ir spectacular anatomy. Th is extrême adaptat ion bas appeared several t imes among m a m m a l i a n predators (Emerson & Radinsky, 1980) and, w i t h i n Carn i vo ra , the N imrav idae remains one of the most diverse famil les of sabretooths. Over
I N T R O D U C T I O N the course of the ir temporal range, from late Eocene to late Miocène (Bryant , 1996; M a r t i n , 1998), n imrav ids were among the most impressive predators i n the nor thern hémisphère. They displayed sabretoothed morphological features (e.g. elongated upper canines, deep flange on the mandible) wh i ch reached extrême proportions i n some taxa (e.g. Eusmilus and Barbourofelis). The history of Palaeogene N imrav idae , i.e. the N imrav inae , is by far the best documented, especially i n N o r t h Amer i c a where numerous remains have been found.
*Corresponding author. E -ma i l : [email protected]
The hun t ing and k i l l i n g behaviours of adult n imravids have been wide ly investigated. Most studies have dealt w i t h the funct ional analys is of the c ran ia l
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493 477

morphology and the use of sabre teeth (see review i n Bryan t & Russe l l , 1995). In contrast, juvénile mate-r i a l is poorly represented i n palaeontological collections and, as a resuit , a lack of knowledge of juvénile anatomy has h indered unders tand ing of the ontogenetic development of n imrav ids . Here we report on an exceptional juvénile spécimen of an early Oligocène n imrav id belonging to the Hoplophoneini . Th is is the earliest, as we l l as one of the most complète, juvénile n imr av i d c ran ia found. It sheds l ight on the overal l juvénile anatomy and ontogeny of the most impressive Palaeogene sabretoothed predators. It is also important , because, to date, no juvénile bas icran ia of the Hoplophonein i have been described. A s for many mammals , the juvénile anatomy i n Carn i vo ra is informative w i t h regard to phylogenetic re lat ion-ships. In part icular , the anatomy of the auditory région is especially important , since the re lat ionships of éléments m a k i n g up the hu i l a may be examined at an earl ier stage when they are not yet fused (Hunt , 1974, 1987).
Th is paper focuses on this question. In part icular , we describe the juvénile anatomy of th is new spécimen. We discuss différences i n c ran ia l (proportion, auditory région) and mand ibu la r morphology (mand ibu lar flange proport ion, size and or ientat ion of processes) wh i ch d is t ingu ish juvéniles from adults , and différences between anatomical features re lated to âge and those unre lated to âge that have systematic imp l i cations. We compare the séquence and t i m i n g of tooth éruption i n the Hoplophonein i i n order to détermine: (i) the s im i l a r i t y of the delayed éruption of the upper canine i n that group and i n the Barbourofe l inae, taxa wh ich developed an extrême sabretoothed adaptat ion, and (il) the conséquence of the delayed éruption of the upper canine for the éruption of other teeth and life history of th is ind i v idua l . We also describe the muscula r insert ions on the mandib le and c ran ium of Hoplophoneini , and address phylogenetical and biological (e.g. power of the bite, predatory capabi l i t ies and feed-ing behaviour) inferences of the juvénile anatomy i n n imrav ids .
M A T E R I A L A N D M E T H O D S
A B B R E V I A T I O N S A N D T E R M I N O L O G Y
Institutional A M N H , Depar tment of Vertébrale Paleontology, Amer i can Muséum of N a t u r a l History, New York; BSP , Bayer ische S taa tssammlung f i i r Palâontologie u n d historische Géologie, M u n i c h ; F : A M , F r i c k collection, Department of Vertébrale Paleontology, Amer i can Muséum of N a t u r a l History, New York; FSP , collections of palaeontology. Faculté des Sciences Fondamentales et Appliquées, Université de Poit iers,
Poit iers; L A C M , N a t u r a l H is to ry Muséum of Los Angeles County, Los Angeles; S M N S , Staat l iches Muséum fi ir Na tu rkunde Stut tgar t (Mammalogy) ; U C M P , Muséum of Paleontology, Un i ve r s i t y of Cal i for-n i a at Berkeley; U F , F l o r i da Muséum of N a t u r a l H i s tory, Un i ve r s i t y of F lo r ida , Gainesv i l l e ; U N S M , Nebraska State Muséum, Un i ve r s i t y of Nebraska , L inco ln ; Y P M - P U , Pr inceton Un ive rs i t y N a t u r a l H i s tory Muséum, collection now i n the Yale Peabody Muséum of N a t u r a l History, New Haven .
Measurements A i l measurements are i n millimètres. M L , mand ibu la r length, from anter ior t ip of dentary to posterior t ip of condyle; F H , height of the génial mand ibu la r flange, from vent ra l t ip of flange to anter ior border of postcanine diastema; L (for teeth), mesiodista l length; w, lab io l ingua l w id th ; ht, height of cusp.
Terminology B y convention, the dental notat ion is as follows: I l 12 13 C P3 P4 M l for upper teeth; i l 12 13 c p3 p4 m l for lower teeth. A 'd ' ind icates a deciduous tooth (e.g. dP4 for the upper deciduous fourth premolar). The te rm 'juvénile' corresponds here to ind iv idua ls w i th only deciduous teeth, i.e. s im i l a r i n âge to F S P - I T D 342. Indiv iduals w i t h some deciduous and some permanent teeth are here cal led 'subadult ' .
Materials The descript ion is ma in l y based on an exceptionally complète associated c ran ium and mandible (FSP- ITD 342) w i t h d I l -3 , dP3-dP4 on both maxi l lae , r ight d i l -dp4 and left broken dc-dp4 on the mandib le . Th is spécimen represents the f irst known n im rav i d w i th an associated c ran ium and mandib le from the Quercy fissures. It was recovered from Itardies (Phosphorites du Quercy, south-western France) several years ago dur-ing an excavation carr ied out by one of us (L.B.). Itardies is early Oligocène i n âge and the type local ity for the référence standard level M P 23 of the European biostrat igraphic scale (BiochroM'97, 1997: tables 2 and 3), wh i ch corresponds to the earl iest A r i ka r e ean (North Amer i can L a n d M a m m a l Age; M c K e n n a & Be l l , 1997: fig. 1).
For comparison, we studied addi t iona l mater ia l of juvénile Palaeogene n imrav ids from N o r t h Amer i can and European palaeontological collections. However, when a personal v is i t to a collection was not pract i -cal , avai lable i l lustrat ions and/or descriptions of spécimens from the l i terature were used. F i r s t , we restr icted our study to ind iv idua ls w i t h only deciduous teeth: no other juvénile c ran ia l mate r ia l has been
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

J U V E N I L E A N A T O M Y O F N I M R A V I D A E ( C A R N I V O R A ) 4 7 9
found i n Europe, and i n N o r t h Amer i ca only a few c ran ia l spécimens, associated or not w i t h mandib le , are avai lable. Those are assigned to Nimravus ( F : A M 99259), Dinictis ( F : A M 62063, F : A M 62103, F : A M 69350, U N S M 4051-81) and Eusmilus ( U C M P 123181). We d id not study U C M P 123181 but B r yan t (1984, 1988) described this spécimen, wh i ch is a very fragmentary juvénile skeleton, inc lud ing a few cran i a l remains w i th r ight dP3-dP4, e rupt ing M l , and a mandib le w i th roots of deciduous teeth. Several iso-lated juvénile mandibles w i t h deciduous dent i t ion add to the juvénile sample of Hoplophonein i {sensu Bryant , 1996). Two spécimens are assigned to Hoplo-phoneus primaevus (Leidy, 1851): A M N H 5337, holo-type of H. oreodontis (Cope, 1873), a jun io r synonym of H. primaevus (Bryant , 1996; M a r t i n , 1998) and A M N H 140510. On ly one subadult spécimen is referred to Eusmilus: A M N H 59573 (cast of L A C M 5465, r ight mand ibu la r fragment w i t h pa r t i a l m l ) assigned to Ekgmoiteptecela olsontau MacDona ld , 1963 by MacDona ld (1970), wh i ch may be a jun io r synonym of Eusmilus olsontau (see M a r t i n , 1998) or E. cerebralis (Cope, 1880) according to B r yan t (1984), see also discussion i n B r yan t (1996). Add i t i ona l cran i a l anatomical comparisons inc luded s l ight ly older ind iv idua ls wh i ch are fréquent i n N o r t h Amer i can collections, and especially those preserv ing deciduous upper canines associated w i t h permanent cheek teeth.
M E T H O D S
The discussion of any différences between the juvéni le and the adult n imrav ines and a detai led reconstruct ion are only possible w i th référence to a comparative sample of extant relat ives (Bryant & Seymour, 1990). A hypothesis of re lat ionship between extinct and extant taxa is indeed necessary for any phylogenetic inference, i.e. 'the transfer of known attr ibutes i n other taxa to the fossil taxon based on an assumed hypothesis of phylogenetic re lat ionships between the fossil taxon and two or more other taxa. ' (Bryant & Russe l l , 1995: 117). However, because the controversial re lat ionships of N imrav idae w i t h i n Carn i vo ra (see Bryant , 1991: fig. 3) give no support to a direct transfer of any one carn ivoran anatomy to n i m r a v i d anatomy, wh i ch group should be used remains cr i t i ca l i n the présent case. Nevertheless, anatomy consistent among extant carnivorans can be transferred to n imrav ids . A s a resuit , we used the muscu lar and osteological anatomy of several groups such as felids, canids and bears as a guide to infer unpreserved anatomical attr ibutes i n n imrav ids . Those taxa are not only members of the m a i n groups of extant Ca rn i vo ra (Aeluroidea, Cynoidea and Arcto idea, respectively).
they are also the most intensive ly studied carnivor a n taxa. In contrast, less work has been done on muscu lar attachments and anatomy i n other groups (i.e. v iverr ids, herpestids, most muste l ids, etc.). O u r own observations of the osteological anatomy i n extant carnivorans (collections i n F S P , S M N S ) add to the t rad i t i ona l and excellent comparative informat ion found i n the works of Dav is (1964), Barone (1967, 1986, 2000), Crouch (1969), Gaspard (1971) and Evans (1993), among others.
D E S C R I P T I O N O F F S P - I T D 342
C R A N I U M ( F I G S 1-3)
The c ran ium is la tera l ly compressed and badly frag-mented. The posterior part of left bas ic ran ium and occiput is miss ing . The length of the c ran ium from the anter ior t ip of the p remax i l l a to the foramen magnum notch is about 83 mm. Th is is about the same length as i n Leptailurus serval at the same dental âge (i.e. deciduous teeth only), a l though the proportions of the c ran ium from Itardies differ markedly. F S P - I T D 342 has a narrow and ta l l c r an ium s imi la r to that of adult n imrav ids , that might be a l i t t le exaggerated because of latéral compression. The ros t rum is long and par-t i cu lar ly narrow, wh i ch may par t ly resuit from the absence of a bulge on the surface of the max i l l a , latéral to the canine. Premaxi l lae are preserved, and the sutures are dist inct. The i r posterior port ion is narrow and extends far posteriorly between the nasa l and max i l l a . The premax i l l omax i l l a ry suture is s i tuated at the anterior-most port ion of the dC alveolus. Due to poor préservation, the posterior l im i t of the nasa l is not clearly d ist inguishable , but may be at, or s l ight ly posterior to, the maxi l lo f ronta l suture. The infra-orbi ta l foramen opens above the paracone of dP4; i t is large and l i t t le la tera l ly compressed. The latéral bridge is somewhat th in , as i n Nanosmilus (known only from the type, a c ran ium and associated mand i bles of a subadult w i th deciduous upper canines, U N S M 25505) and Eusmilus. The palate is narrow anter ior ly and broadens at the anterior end of dP3. The anter ior palat ine foramen is level w i th the para-style of dP3 ; the media l palat ine foramen level w i th the d is ta l marg in of dP4. Orbi ts are large, rounded and posteriorly l im i t ed by a strong postorbital process wh ich is as developed as i n the adult of Hoplophone in i . Bo th zygomatic arches are preserved, a l though latera l ly compressed due to crushing , and support a poorly developed dorsal juga l process. The suture between the lac rymal and juga l cannot be d is t in -guished. The foramen ovale and posterior opening of the a l isphenoid canal are closely associated and lie w i t h i n the same deep dépression, media l to the trans-versely elongated glenoid fossa. The postglenoid fora-
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

4 8 0 S. PEIGNÉ and L. D E B O N I S
F igu re 1. F S P - I T D 342, c r a n i u m i n (A) r i g i i t latéral view, a n d (B) left latéral view. Sca le = 1 cm.
men is very smal l , and located i n the base of the postglenoid process. The braincase is fragmented, but sagi t ta l and occipital crests are absent or not yet developed. In contrast, the sagit tal , and especial ly occipital , crests of juvénile spécimens of Dinictis ( F : A M 69350) are already developed at th is dental âge. There are no data for juvénile Hoplophoneus and Eusmilus.
Basicranium (Figs 2 and 3) The spécimen reported here is the youngest n i m r a v i d w i th a preserved bas ic ran ium. U p to now, the only publ ished descript ion of the bas i c ran ium of a young ind i v idua l is of a subadult spécimen w i t h permanent cheek teeth part ly or ful ly erupted (Hunt , 1987), thus represent ing a dist inct ly later dental stage t h a n that of the c ran ium from Itardies. Two juvénile bas icran ia have also been studied here: F : A M 99259 (east of Robertson Draw, Wyoming; latest Chadronian)
assigned to Nimravus brachyops (Cope, 1878) wh ich is as young as F S P - I T D 342, and F : A M 62063 (from Bar t l e t Ranch , nor th of Chadron , Dawes County, Nebraska ; Ore l l a Member, Brûle Formation) assigned to Dinictis felina Leidy, 1854; wh ich represents a later developmental stage (erupting permanent teeth). A l though the c ran ium from Itardies is somewhat fragmented, the r ight auditory région préserves not only the petrosal, but also the m a i n port ion of the ectotympanic and a few fragments of the caudal entotympanic , wh ich supply cr i t i ca l détails on n imrav id hu i l a structure.
The petrosal is the best preserved élément of the auditory région. Some hu i l a éléments are s t i l l i n place, and this obscures some of the posit ional re lat ionships of the petrosal to the surround ing bones. The shape of the vent ra l surface of the petrosal i n F S P - I T D 342 is very s im i l a r to that of Hoplophoneus and Eusmilus. F S P - I T D 342 has a round promontor i u m wh ich apparent ly lacks a ventra l process. A very
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477-493

J U V E N I L E A N A T O M Y O F N I M R A V I D A E ( C A R N I V O R A ) 4 8 1
Figure 2. F S P - I T D 342, c r a n i u m i n v e n t r a l view. 5cale = 1 cm.
i iscrete anteroposterior ridge is présent on the surface o f the petrosal. A l though i t is less marked than i n idu l t spécimens, th is may be the latéral l im i t of i t tachment of the osseous entotympanic, as i n other spécimens preserving th is région. The most dist inc-;ive feature of F S P - I T D 342 is the posterolateral extension of the promontor ium, wh i ch is i n life posi-;ion. In contrast to spécimens of Hoplophoneus, this îxtension is a near ly hor izonta l bony plat form w h i c h s ventra l ly recessed. The plat form may be broken i n ;he k n o w n spécimens of European Eusmilus; never-;heless there is a s im i l a r shal low dépression on the /entrai side of that p lat form i n Eusmilus bidentatus F i l ho l , 1872) ( M N H N - Q U 9521). In F S P - I T D 342, a l a r row latéral notch séparâtes the m a i n part of the j lat form from the promontor ium. Posterolateral ly i t widely contacts the masto id and exoccipital bones. Just dorsal ly and a l i t t le anter ior ly is the opening of ;he cochlear fenestra, wh i ch is or ientated posterolat-îrally and s l ight ly ventral ly. The anterolateral face of ;he petrosal is near ly ver t ica l and slopes dorsad. The aresence of the ectotympanic prevents a detai led iescr ipt ion of the tegmen tympan i . Moreover, the roof )f the tegmen is damaged and i ts placement, deeply nset i n the bas ic ran ium, precludes a detai led descrip
t ion of th is région. A s far as we can see, there are no marked différences w i t h the tegmen t ympan i of other Palaeogene n imrav ids .
The anteroventral part of the ectotympanic i n F S P -ITD 342 is broken away, but the m a i n port ion of the élément is preserved. Breakage of the tympanic has produced an offset of the élément relat ive to the parasag i t ta l plane. A s a resuit , the anter ior crus is t i l t ed s l ight ly backward and not ful ly pressed against the squamosal , and the posterior crus is latera l ly dis-placed. Nevertheless, the posterior point of at tachment is ful ly vis ible. There is a near ly vert ica l ly orientated, shal low dépression located on the antero-in t e rna l edge of the masto id process, wh i ch suggests that the ectotympanic was probably orientated near ly vert ica l ly i n the l i v ing an ima l . The ectotympanic i n this juvénile resembles that of subadult and adult Palaeogene n imrav ids (i.e. Dinictis, Hoplophoneus, Nimravus and Eusmilus). It is a near ly vert ica l ly orientated crescent, not chambered l ike the ectotympanic i n most extant aeluroids. The vent ra l edge is about 0.9-1 m m i n thickness and has a s l ight ly rugose but no concave surface. There is no trace of breakage and therefore th is is probably the finished vent ra l marg in of the ectotympanic. The in te rna i surface of this élément is very s im i l a r to that ot Dinictis and Hoplophoneus. The cr is ta t ympan i is preserved as a t h i n bony r i m . Ven t ra l to the cr ista, the in te rna i surface is concave and about 5 m m i n m a x i m u m height. The anterior-most port ion of the ectotympanic préserves a sma l l crest wh i ch may correspond to the l im i t of the anterior extension of the cart i laginous entotympanic wh i ch forms the vent ra l floor of the bu l l a i n the N i m ravinae (Hunt , 1987). Th is is a t h i n crest 2.7 m m i n length and 1 m m i n height. It forms an angle of about 60° w i t h the long axis of the crescent. There is no contact between this crest and the cr ista tympani . Anterodorsa l ly is a t h i n plate of bone i n a s imi la r posit ion to that of the protubérant styl i form process présent i n other n imrav ids (Hunt , 1987: figs 3, 4 and 7; Joeckel et al., 2002: fig. 8). It is about 5 m m i n length but its height cannot be measured since i t is l i ke l y broken and not ful ly preserved on this spécimen. Dorsa l l y and media l ly to the posterior attachment of the ectotympanic to the c ran ium is a smal l pièce of bone wh i ch forms the anterior w a l l of the canal that carries the facial nerve as i n Dinictis (Hunt, 1987: fig. 3). A s H u n t described i n that genus, this sma l l fragment of bone is the dorsal port ion of the tympanohya l . The stylomastoid foramen is located posteriorly but lacks its media l part.
A sma l l fragment of a vert ica l ly orientated bony plate of about 1 m m i n thickness is présent anterome-d ia l l y i n the bu l l a . It is s l ight ly latera l ly concave. Accord ing to its posit ion, and i n comparison w i th other n im rav i d bul lae, there is l i t t le doubt that this
s) 2003 The L innean Society of London, Zoological Journal of the Linnean Society, 2003, 138, 477 -493

4 8 2 S. PEIGNÉ and L. D E B O N I S
F i gu re 3. F S P - I T D 342, c r a n i u m . Stereop l io tographs of t l i e v e n t r a l s ide the r i gh t aud i t o ry région, Abbrev i a t i ons : B O , bas iocc ip i ta l ; C F , cochlear fenestra ; CT, c r i s t a t y m p a n i c a ; E C , ec to tympan ic w i t h an te r i o r a n d poster ior c r u r a ; E O , exocc i p i t a l ; ET , f ragment of c auda l en to tympan ic ; F O , f o ramen ovale; G F , g leno id fossa; M P , mas t o i d process, P L E P , posterol a t e r a l ex tens ion of the pe t rosa l ; P N , pe t rosa l no tch s epa ra t ing the m a i n pa r t of the postero la tera l ex tens ion f rom the p r o m o n t o r i u m ; P O A C , poster ior open ing of the a l i spheno id c a n a l ; P P , pe t rosa l p r o m o n t o r i u m ; S Q , squamosa l ; T H , dorsa l f ragment of the t y m p a n o h y a l . S o l i d c irc les ind ica te the a rea of a t t a chment for brach io - and/or s te rnocepha l i cus ; the so l id t r i ang l e ind ica tes the s m a l l dépression pos te romed ia l l y to the mas to id .
fragment is part o f the ossified entotympanic. Because of its poor préservation, i t is not possible to d is t ingu ish the rostra l entotympanic as a discrète ossificat ion. The plate is 6 m m i n anteroposterior length and contacts anter ior ly the dorsomedial r i m of the ectotympanic , dorsal ly the petrosal, and medial ly, the
bas ic ran ium. The ventra l edge of the posterior-most port ion of th is élément has a smooth trough, as has been observed i n some other n imrav ines . As demon-strated by H u n t (1987), such an edge most probably represents the f inished marg in of the caudal entotympanic, wh i ch suggests the présence of an addit ional
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477-493

J U V E N I L E A N A T O M Y O F N I M E A V I D A E ( C A R N I V O R A ) 4 8 3
and dorsal ly developed cart i laginous entotympanic i n this spécimen.
The masto id process and glenoid fossa are not as dépendent as i n adults of Hoplophoneus and Eusmilus. The surface of the masto id région is a l i t t le vent ra l l y expanded and i ts ventra l surface displays a deep and rugose surface for a muscu lar at tachment. The squamosal port ion of the process is crest- l ike, prominent and more ventra l ly extended t h a n is the posterior (mastoid) part. O n the ventra l surface of the masto id there is a sma l l and rounded dépression posteromedial to the masto id wh ich is absent i n adults (solid tr iangle on F ig . 3). The smooth surface of th is fossa suggest that th is may not be a muscu lar attachment. The func-t ion of this fossa is unknown. Posterior to th is is the sma l l paroccipi ta l process, wh i ch is somewhat prominent, crest-l ike, and dorsoventral ly developed. The posterointernal port ion of the auditory région is damaged and, as a resuit , the petrobasi lar and posterior lacerate foramina cannot be d ist inguished.
M A N D I B L E ( T A B L E S 1 A N D 2, F I G . 4)
The ramus is about 70 m m i n length, shallow, and thick. Despite the young âge of th is ind i v idua l , the mand ibu la r flange is deep, extending about 24 m m from its ventral-most point to the anterior-most point of the postcanine diastema. Its proport ion relative to the mandib le length is thus greater than i n most n i m ravine spécimens assigned to Hoplophoneus (Table 2). A s i n most young ind iv idua ls , the anterior, latéral border of the flange is prominent. There are three menta l foramina, the largest l y ing beneath the anterior part of the postcanine diastema. The dorsal r i m ofthe mand ibu lar d iastema is t h i n and a l i t t le la tera l ly concave, as i n adult n imrav ines . A s i n n imrav ids such as Dinictis, Hoplophoneus and Eusmilus, there is a rugose knob on the l ingua l face of the mandible beneath the alveolar border a l i t t le posterior to dp4, wh ich is l i ke l y to be a muscu lar at tachment (see Discussion). The mand ibu la r foramen opens posteroventral ly and is
Table 1. C o m p a r a t i v e measurements of dec iduous tee th i n F S P - I T D 342. O t h e r gênera are represented by i n d i v i d u a l s w i t h on ly dec iduous tee th
Nimravus Hoplophoneus F S P - I T D 342 F : A M 99959 Binictis A M N H 5337
L w L w L w L w L w L w h t proto %tr igo L/w
L w L w L w L w L w h t p a r a L w 5.7
1.5 I. 9 2.4 2.5 2.1 3.4 4 .7* 2.2 7.45* 2 .55*
I I . 1* 3 .85* 8.1 9 0 . 1 * 2 .88*
3 2.8 3.3 2.8 3.7 2.4
11.8 4.7 7.3 5.25* 6.6
11.7 4.4 15.6 4.9
84 3.18
9.7 4.7 16.9* 7.8
6.9 10.8
5.3 3.5 10.15 (9.3-10.8; N = 6) 3.5 (3.2-3.9; AT =6 ) 13.55 (11.8-15.3; AT = 2) 10.8 4.6 (4 .2-5; = 2) 4.1
84.5 88.6 2.93 (2.8-3.06; AT = 2) 2.63
9 (8-10; AT = 2) 3.85 (3.8-3.9; AT = 2) 15.5 (14.4-16.3; AT = 3) 7.85 (7 .7-8; Af = 2)
7.7 11
*Le f t a n d r i g h t tee th are preserved ; for Dinictis, w h e n more t h a n one i n d i v i d u a l , range a n d sample size are ind i ca ted w i t h i n parenthèses. Excep t for the propor t i on of the t r i g on id of m l ( in per cent), a i l measurements are i n m m .
© 2003 The L i n n e a n Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

4 8 4 S. PEIGNÉ and L. D E B O N I S
Table 2. P r opo r t i on of the m a n d i b u l a r f lange i n n i m r a v i n e n i m r a v i d s
Juvéniles S u b a d u l t s - a d u l t s
N m range S D m range S D
F S P - I T D 342 1 34.3 — _ — _ _ Dinictis 2 22.09 22 .01-22 .17 0.11 13 27.66 24 .29-31 .23 1.8
Nanosmilus - - - - 1 34.90 - -U N S M 25505 H. primaevus - - - - 16 36.73 31 .09-39 .46 2.24
H. occidentalis - - - - 3 41.16 39 .71-43 .65 2.17
H. mentalis - - - - 2 42.41 40 .65-44 .18 2.49 H. dakotensis - - - - 1 45.43 - -Y P M - P U 11079 E u r o . Eusmilus — _ 1 53* —
BSP-Môhren31 N.A . Eusmilus - — _ 1 63* — _ F- .AM 69377
A i l measurements i n per cent: dep th of the f lange/length of the mand ib l e . N = sample s ize ; m = mean ; S D = one s t a n d a r d déviation. E u r o = E u r o p e a n ; N .A . = N o r t h A m e r i c a n . * E s t i m a t e d .
vi^ider t h a n that i n the adult . The anterior marg in of masseteric fossa is about 10 m m posterior to dp4; i t is deep but dorsal ly l im i ted by the réduction of the coro-noid process, and anterodorsal ly by a prominent crest wh i ch continues upward and backward to form the root of the coronoid process, as i n most species assigned to Hoplophoneus and Eusmilus (but not i n E. villebramarensis Peigné & B r u n e i , 2001). The anter ior border of the process is orientated backward , and its posterior r i m is near ly vert ica l . The mand ibu la r condyle is dépendent and extended m u c h below the alveolar border; as a resuit , i t is closely located to the angular process as is that of Eusmilus villebramarensis and E. sicarius S inc la i r & Jepsen (1927). The short angular process, ful ly preserved i n the r ight mandib le , displays a deep, la tera l ly expanded dépression as i n Hoplophoneus and Eusmilus.
T E E T H ( T A B L E 1, F I G S 1, 2 A N D 4)
A i l upper deciduous incisors are preserved. They are canini form and curved. The dI2 is only s l ight ly larger than d l l , whi le dI3 is m u c h larger t h a n the others and also more la tera l ly compressed. The incisor arcade is strongly curved. A s i n juvéniles of other h igh ly sabretoothed n imrav ids (Bryant , 1988; see Discuss ion herein), the deciduous canine is probably not yet erupted. The dC alveolus is indeed not ful ly opened, and is about 7.4 m m mesiodista l ly and 2 m m lab io l in-gually. The dP3 is the f irst deciduous cheek tooth; i t is preserved on both sides and located about 8.5 m m posterior to the d is ta l end of the dC alveolus. It is three-rooted and blade-l ike; the l i ngua l root is level w i th the
elongated paracone and supports a low, sl ight bulge but not a cusp. The dP3 has a large, elongated para-style and a short metastyle. The carnass ia l notch is deep. The dP4 is t r i angu lar and three-rooted; i t is transversely narrow and the l i ngua l root does not support a protocone. The l ingua l ha l f is better developed t h a n i n Eusmilus cerebralis ( U C M P 123181; Bryant , 1984: fig. 2). L ingual ly , convex crests extend from the central cusp to the lab ia l corners. The lab ia l face is strongly concave. In some places, serrations are s t i l l évident on the teeth because they are l i t t le worn. Breakage of the d is ta l face of the alveolus exposes the l ingua l ha l f part o f the crown of M l ; i t cannot be dist inguished from that oî Eusmilus and Hoplophoneus.
Deciduous lower incisors and the canine are preserved on the r ight mandible . Incisors are canini form, more curved than the upper ones and the ir size increases from i l to 13. The 13 is more la tera l ly compressed t h a n i l or 12. Serrat ions are not v is ible. The deciduous canine may not be ful ly erupted since the base of the crown s t i l l l ies below the alveolar border; the t ip of the canine is broken but was probably ta l ler and longer than 13, but less curved and more lateral ly compressed. The inc isor -canine arcade is as curved as the upper incisor arcade. O n both hemimandib les , breakage of the l i ngua l port ion of r a m i exposed the mes ia l face of the crown of the permanent incisors (Fig. 4A, D). The postcanine d iastema is 12.5 m m . The first cheek tooth, dp3, is as t a l l as the paraconid of dp4; the m a i n cusp is t i l t ed s l ight ly posteriorly. There are two sma l l addi t ional cusps; the d is ta l one is s l ight ly larger than the mes ia l one. The dp4 has a complète tr igonid, where the paraconid and protoconid
© 2003 The L i n n e a n Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493
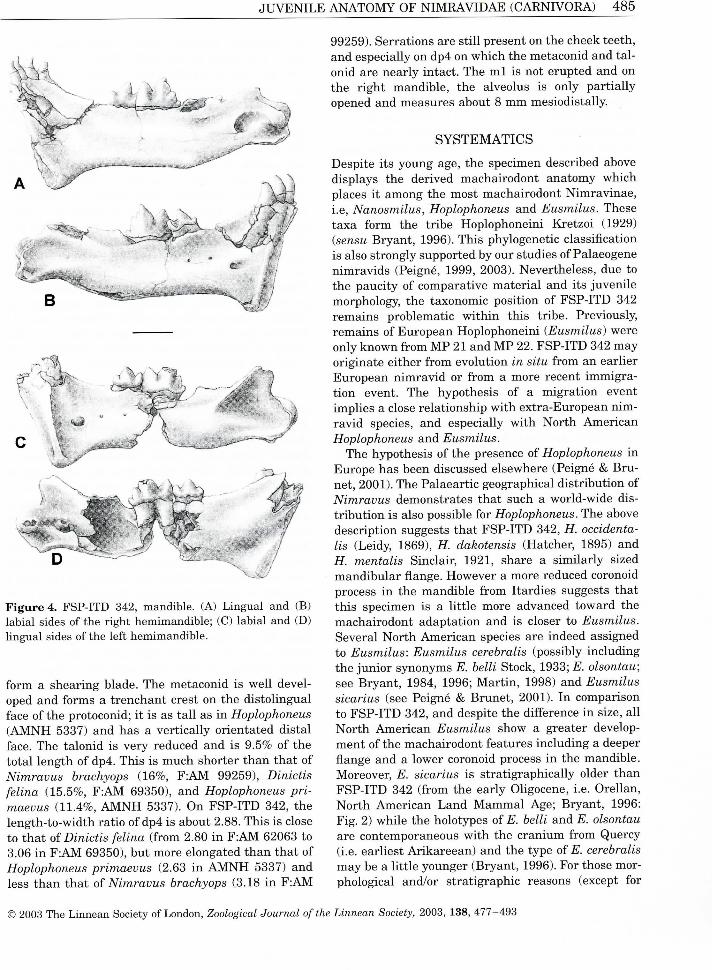
J U V E N I L E A N A T O M Y O F N I M R A V I D A E ( C A R N I V O R A ) 4 8 5
F i gu re 4. F S P - I T D 342, mand ib l e . (A) L i n g u a l a n d (B) l a b i a l sides of the r i g h t h e m i m a n d i b l e ; (C) l a b i a l a n d (D) l i n g u a l s ides of the left h e m i m a n d i b l e .
form a shear ing blade. The metaconid is we l l developed and forms a t renchant crest on the d is to l ingual face of the protoconid; i t is as t a l l as i n Hoplophoneus ( A M N H 5337) and has a vert ica l ly or ientated d is ta l face. The ta lon id is very reduced and is 9.5% of the tota l length of dp4. Th is is m u c h shorter t h a n that of Nimravus brachyops (16%, F : A M 99259), Dinictis felina (15.5%, F : A M 69350), and Hoplophoneus primaevus (11.4%, A M N H 5337). O n F S P - I T D 342, the length-to-width rat io of dp4 is about 2.88. Th is is close to that oï Dinictis felina (from 2.80 i n F : A M 62063 to 3.06 i n F : A M 69350), but more elongated than that of Hoplophoneus primaevus (2.63 i n A M N H 5337) and less t h a n that of Nimravus brachyops (3.18 i n F : A M
99259). Serrat ions are s t i l l présent on the cheek teeth, and especially on dp4 on wh i ch the metaconid and ta l onid are near ly intact. The m l is not erupted and on the r ight mandible , the alveolus is only par t ia l l y opened and measures about 8 m m mesiodistal ly.
S Y S T E M A T I C S
Despite its young âge, the spécimen described above displays the derived machairodont anatomy wh i ch places i t among the most machairodont N imrav inae , i.e, Nanosmilus, Hoplophoneus and Eusmilus. Thèse taxa form the tr ibe Hoplophonein i Kre t zo i (1929) {sensu B ryant , 1996). Th is phylogenetic classif ication is also strongly supported by our studies of Palaeogene n imrav ids (Peigné, 1999, 2003). Nevertheless, due to the paucity of comparative mate r ia l and its juvénile morphology, the taxonomic posit ion of F S P - I T D 342 remains problematic w i t h i n th is tr ibe. Previously, remains of European Hoplophonein i {Eusmilus) were only known from M P 21 and M P 22. F S P - I T D 342 may or ig inale e i ther from évolution in situ from an earl ier European n i mrav i d or from a more récent immigra t ion event. The hypothesis of a migrat ion event impl ies a close re lat ionship w i t h extra-European n imrav id species, and especially w i t h N o r t h Amer i can Hoplophoneus and Eusmilus.
The hypothesis of the présence of Hoplophoneus i n Europe has been discussed elsewhere (Peigné & B r u -net, 2001). The Palaeart ic geographical d is t r ibut ion of Nimravus demonstrates that such a world-wide dist r ibut ion is also possible for Hoplophoneus. The above descript ion suggests that F S P - I T D 342, H. occidentalis (Leidy, 1869), H. dakotensis (Hatcher, 1895) and H. mentalis S incla ir , 1921, share a s imi la r l y sized mand ibu la r flange. However a more reduced coronoid process i n the mandib le from Itardies suggests that th is spécimen is a l i t t le more advanced toward the machairodont adaptat ion and is doser to Eusmilus. Several N o r t h Amer i can species are indeed assigned to Eusmilus: Eusmilus cerebralis (possibly inc lud ing the jun io r synonyms E. belli Stock, 1933; £. olsontau; see Bryant , 1984, 1996; M a r t i n , 1998) and Eusmilus sicarius (see Peigné & B r u n e i , 2001). In comparison to F S P - I T D 342, and despite the différence i n size, a i l N o r t h Amer i can Eusmilus show a greater development of the machairodont features inc lud ing a deeper flange and a lower coronoid process i n the mandib le . Moreover, E. sicarius is s trat igraphica l ly older than F S P - I T D 342 (from the ear ly Ohgocene, i.e. Ore l lan , N o r t h Amer i can L a n d M a m m a l Age; Bryant , 1996: F i g . 2) whi le the holotypes of E. belli and E. olsontau are contemporaneous w i th the c ran ium from Quercy (i.e. earl iest Ar ikareean) and the type of E. cerebralis may be a l i t t le younger (Bryant , 1996). For those morphological and/or strat igraphie reasons (except for
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

4 8 6 S. PEIGNÉ and L. D E B O N I S
E. sicarius), F S P - I T D 342 cannot be a descendant of the Amer i can Eusmilus. One support for the hypothesis of a migra t ion event is that the European level M P 23 also marks the f irst appearance da tum for other m a m m a l i a n groups such as the Chal ico ther i -idae and Bachi ther i idae (Blondel , 2001) and the carnivorans Stenoplesictis and Stenogale. However, i t should also be noted that, i n contrast to those taxa, the Hoplophonein i were already présent i n Europe before M P 23. For th is reason, the hypothesis of an ancestor-descendant re lat ionship of F S P - I T D 342 to an endémie European n imrav i d species should be seriously considered. There are indeed two European Hoplophoneini : Eusmilus bidentatus, the type species of the genus, wh i ch is présent i n the ear ly Oligocène (European s tandard level M P 21), and Eusmilus villebramarensis, a recently described species (Peigné & B r u n e i , 2001) from V i l l eb ramar (Lot-et-Garonne, France) , the type local i ty of European level M P 22. Those species might belong to the same lineage and display a t rend toward greater development of machairodont features and larger size, since E. villebramarensis is about 35% larger than E. bidentatus. However the mate r ia l is fragmentary and this t rend remains to be confirmed. Some comparative data between F S P - I T D 342 and the European Hoplophone in i have been provided i n the descript ion and are briefly summar i zed here. Compared to Eusmilus villebramarensis, F S P - I T D 342 has a smal ler est imated adult body size, a less reduced, ta l ler coronoid process and a prominent crest located anterodorsal to the masseteric fossa. In contrast, F S P - I T D 342 shares a i l of thèse morphological features w i th Eusmilus bidentatus, wh ich also has a s im i l a r sized flange. The discussion above showed that i n re lat ion to the development of the machairodont adaptat ion, F S P -ITD 342 cannot be a descendant of E. villebramarensis. In contrast, th is spécimen shares many features w i th Eusmilus bidentatus, a l though F S P - I T D 342 is d is t inguished by some bas icran ia l features (e.g. the large plat form posterior to the promontor ium of the petrosal). Whether the c ran ium from Itardies belongs to th is species cannot therefore be determined, par-t i cu lar ly because of a lack of juvénile remains of E. bidentatus. A s a resuit , F S P - I T D 342 is assigned to c f Eusmilus bidentatus u n t i l the discovery of adult remains from localit ies of the M P 23 level.
D I S C U S S I O N
J U V E N I L E V S . A D U L T A N A T O M Y : O N T O G E N Y , E R U P T I O N
S E Q U E N C E A N D P H Y L O G E N E T I C I M P L I C A T I O N S
The overal l proportions of the c ran ium and mand i ble i n juvéniles are considerably différent from those of adults. The c ran ium of the juvénile, w i th i ts longer
and narrower ros t rum, seems a l i t t le more progna-thous t h a n that of the adult . There is l i t t le doubt that the lack of c ran ia l crests and the poor development of bas ic ran ia l processes at th is early âge imp ly that muscu lar attachments to the c ran ium were poorly developed. The glenoid fossa and the masto id process are much less dépendent than i n the adult . The shape of the masto id process is also quite différent, especially the vent ra l surface wh i ch is deeply recessed w i th marked scarr ing i n the juvénile cran i u m . Th is contrasts w i t h adults of Eusmilus, Hoplophoneus and Nanosmilus, i n wh ich the ventra l surface is flat and s l ight ly concave, the posterior port ion being a l i t t le more dépendent. In contrast the postorbital processes and the infraorbi ta l foramina are as developed as i n adults . Several addi t ional features suggest a greater différence between the juvénile and adult , especially those regarding the auditory région and the mandib le . Whether those différences are only re lated to âge or are diagnostic enough to d is t ingu ish each species of Hoplophoneini requires a more detai led invest igat ion.
Auditory région The auditory région provides the most valuable set of characters to résolve inter- and sometimes in t ra -fami l i a l re lat ionships w i t h i n Carn ivora . The geometry of the petrosal, and the orientat ion, size and morphology of the ectotympanic and entotympanics have taxonomic impl icat ions. W i t h i n N imrav idae , detai led anatomica l studies of th is région identi f ied an anatomy dist inct ive to each genus (Neff, 1983; H u n t , 1987; Joeckel et al., 2002). However différences between juvénile and adul t n i m r a v i d auditory structure can only be interpreted w i t h i n the context of the bu l l a r ontogeny of extant carnivorans. W i t h i n Carn ivora , the ontogeny of the bu l l a has been par t i cu lar ly studied by H u n t (1974). Th i s work forms the m a i n anatomical basis for the fol lowing statements; nevertheless our observations of juvénile and adult ind iv idua ls of extant carnivorans add to this work. Three éléments of the auditory région, a i l preserved i n F S P - I T D 342, are the focus of the fol lowing discussion: the petrosal, the ectotympanic and the caudal entotympanic.
The petrosal is ossified very early i n development and shows no change i n i ts re lat ionship w i th other bu l l a r éléments or i n i ts geometry. In young ind iv iduals of Nimravus and Dinictis, the petrosal has a defi-nite geometry and placement: i t extends caudally, has a well-defined tegmen t ympan i and a ventra l ly rounded promontor ium, and it is deeply inset i n the auditory région (Joeckel et al., 2002; for Dinictis, see also Hun t , 1987: figs 2, 3 and 5). A definite geometry and placement are also probably the case of the petrosal of F S P - I T D 342. A s noted i n the description, the
© 2003 The L i n n e a n Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

dUVii . iMl jJi AiMAlUiVlï Uii JMiV lKAViJJAE ((JAKJNIVUKA) 4 » Y
petrosal i n th is i nd i v i dua l resembles both that of Hoplophoneus and especial ly that of Eusmilus. The m a i n différence w i t h both gênera regards the bony posterior p lat form extending from the promontor ium, wh ich is larger and forms a near ly hor izonta l plate instead of a s loping dorsad. Moreover, the narrow latéral notch wh i ch séparâtes the m a i n part of the platform from the promontor ium i n F S P - I T D 342 is absent i n both gênera.
A l though the ectotympanic is very s im i l a r i n each n imrav ine genus, the or ientat ion of the crescent re lat ive to the sagit tal plane may vary by a few degrees (Neff, 1983). In F S P - I T D 342, the tympanic is a l i t t le displaced, and hence does not have its or ig ina l orientat ion relat ive to the parasag i t ta l plane. However, the near ly ver t ica l or ientat ion of the élément reflects a na tu ra l posit ion. Th is is indeed the most surpr i s ing feature i n F S P - I T D 342, since the or ientat ion of the ectotympanic is s im i l a r to that of the adult . Accord ing to H u n t (1987: 19), the i n i t i a l or ientat ion o f the ectotympanic would have been as i n extant juvénile carnivorans and mammals i n gênerai (see also V a n der K laaw, 1931: 7, 16-18), i.e. i n a hor izonta l posit ion appl ied against the vent ra l surface of the petrosal promontor ium. However, the vert ica l or ientat ion of the ectotympanic at the early âge documented by F S P -ITD 342 is un usua l w i t h i n Carn i vo ra . Juvénile extant carnivorans at a s im i l a r stage d isplay an ectotympanic l y i ng i n a frontal plane or only s l ight ly rotated away from the s k u l l . The par t i cu lar ver t ica l or ientat ion of the plate- l ike ectotympanic i n the adul t n i m rav id was noticed by V a n der K l a a w (1931: 317); such a form and placement for the ectotympanic is cur-rent ly seen as a diagnostic feature of the fami ly (Neff, 1983; H u n t , 1987: table 1; Joeckel et al., 2002). However, young k i t tens such as F S P - I T D 342 (c£ Eusmilus) and F : A M 99259 (Nimravus) suggest that the rotat ion of the ectotympanic away from the s k u l l occurred marked ly ear l ier i n n i m r a v i d ontogeny t h a n i n other Carn i vo ra . The common bu l l a structure shared by a i l Palaeogene N imrav idae (Joeckel et al., 2002) suggests that they may have presented such an early rotat ion of the ectotympanic. Th is ontogenetical feature may therefore be an addi t iona l apomorphy wh ich dist inguishes the Palaeogene N imrav idae from other Carn ivora . In addit ion, the near ly ver t ica l or ientation of the ectotympanic suggests that the bu l l a may have been inf lated ventra l ly ear ly i n its ontogenetic development.
The concave finished ventra l marg in of the entotympanic fragment impl ies the présence of a n addi t iona l élément m a k i n g up the vent ra l floor of the bu l l a . A s i n other Palaeogene n imrav ids such as Dinictis, Hoplo-ohoneus and Nimravus (Hunt , 1987), a vent ra l hyaline extension of the caudal entotympanic was l ike ly présent i n F S P - I T D 342. Nevertheless, this juvénile
displays several important différences compared to other n imrav ids . The preserved port ion of the entotympanic is m u c h shal lower than i n other taxa, wh i ch are ma in l y represented by adults (i.e. Dinictis, Hoplophoneus and N o r t h Amer i can Nimravus and Eusmilus) i n wh ich the ventra l extension of tl ie entotympanic is level w i t h the vent ra l surface of the basioccipital . In F S P - I T D 342, the fragment of entotympanic extends ventra l ly about 2 m m from the petrosal surface, so that i ts vent ra l marg in is considerably dorsal to the surface of the basioccipital . A comparison w i t h spécimens of a s im i l a r âge is unfor-tunate ly l imi ted . On l y one spécimen, an associated c ran ium and mandib le assigned to Nimravus brachyops ( F : A M 99259), is avai lable and displays a th ick entotympanic wh i ch is ta l ler than that of F S P - I T D 342. The ventra l edge of th is élément i n F : A M 99259 may be a flnished marg in , representing the complète extent of ossif ication at th is stage (Neff, 1983). If we assume a s im i l a r t im ing the ontogenetic development w i t h i n N imrav inae , the very shal low entotympanic présent i n F S P - I T D 342 therefore suggests a strong différence i n development and m a x i m u m ventra l extent of ossification at this ontogenetic stage relat ive to Nimravus at least, and possibly w i t h other N o r t h Amer i can n imrav ines . No comparison is possible w i th European taxa since no cran ia l remains have the bu l l a éléments.
Mandible The génial mand ibu la r flange is we l l developed on F S P - I T D 342 (34.3% o f the total length o f the mand i ble). A study of a large sample of Dinictis and Hoplophoneus spécimens suggests that the development of the flange increases w i t h âge u n t i l a i l permanent cheek teeth are ful ly erupted. Because F S P - I T D 342 is a very young ind i v idua l , i ts adult flange is therefore expected to be m u c h larger. Table 2 provides the proport ion of the flange i n juvénile and subadult/adult spécimens i n some gênera. In F S P - I T D 342, this proport ion is greater than i n Dinictis, Nanosmilus and Hoplophoneus primaevus, whatever the âge of the ind i v idua l . In Dinictis, the size of the flange i n a juvéni le reaches about 80% of the size i n an adult (Table 2). A l though this proport ion is not based on a large sample and may vary depending on the genus, i t helps to infer the depth of the flange i n F S P - I T D 342 adults, wh i ch may therefore be about 43% of the total length o f the mandible . Th is is a s imi lar proportion to that of adul t spécimens of Hoplophoneus mentalis, H. occidentalis and H. dakotensis. In Eusmilus the flange is s l ight ly deeper, usua l l y more than 50% of the mand ibu la r length.
The anatomy of the posterior part of the mandible has important taxonomic and biomechanical impl ica-
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

4 » S S. FElCiJNE and L . tSUJNlb
t ions i n Carn ivora . W i t h i n sabretoothed taxa, this région has modif ications associated w i t h the extrême development of sabers that produce a larger gape, inc lud ing a réduction i n the coronoid process, the lower ing of the condyle, and its latéral projection (Emerson & Radinsky, 1980). The coronoid process is somewhat developed i n F S P - I T D 342 but is antero-posteriorly short. Whether the morphology of the process is re lated to âge (i.e. is i t larger i n the juvéni le t h a n i n the adult?) or to phylogeny was tested through a study of extant (felids only) and extinct carnivorans (felids and n imrav ids ) at différent ontogenetic stages. In Leptailurus serval, Panthera pardus and Caracal caracal ( S M N S mammalogy collections), juvénile spécimens have a coronoid process that is generally, but not always, more backward ly orientated than i n adults . In contrast, th is process is not, or only sl ightly, lower than i n adults. Thèse observations are confirmed i n extinct felids, and especial ly i n machairodont ine felids such as 'Homotherium' sérum Cope, 1893 (Rawn-Schatzinger, 1983: figs 2, 3 and 6). The extensive sample of Smilo-don populator (= S. californicus Bovard , 1907; Ber ta , 1985) from Rancho L a B r e a is also informative i n this respect. M e r r i a m & Stock (1932: plate 4) figured 10 spécimens i l lus t ra t ing the ontogenetic growth of the mandible . The shape of the process differed according to âge, and is more backward ly t i l t ed i n juvéniles. However, the coronoid process i n juvénile and adult was s im i l a r i n height, compared to the mand ibu la r length. We observed a s im i l a r t rend i n the n imrav id Dinictis, but no juvénile spécimens are avai lable for Hoplophoneus and Eusmilus. D u r i n g the ontogeny of the barbourofeline n i m r a v i d Barbourofelis loveorum B a s k i n , 1981, the increas ing development of the flange was the major morphological change on the mandib le (Bryant , 1988), that marked ly differs from the s i tuat ion i n F S P - I T D 342. In contrast, juvénile spécimens of B. loveorum w i t h erupt ing m l ( U F 25054 and U F 24410; Bryant , 1988: figs 5A-B, C-D, respectively) have a coronoid process proport ional ly ta l ler and more rounded at the top than that of the adul t ( U F 24429; Bryant , 1988: fig. 6). Consequently there is a re lat ive ly good support for in ferr ing the présence of a coronoid process i n the adult wh i ch is as t a l l as that of the juvénile F S P - I T D 342, but s l ight ly less backward ly orientated. A s a resuit the coronoid process i n F S P - I T D 342 was probably ta l ler t h a n that of more derived n imrav ids such as Eusmilus sicarius and E. villebramarensis. That means F S P - I T D 342 trends less toward the machairodont adaptat ion t h a n those taxa, and more resembles Eusmilus bidentatus and Hoplophoneus dakotensis. The angular process of F S P - I T D 342 has a morphology w h i c h is s im i l a r to that of the adult .
Dentition and éruption séquence High l y derived machairodont taxa such as Barbourofelis and Eusmilus display a greatly delayed éruption of the deciduous upper canine (Bryant , 1988). The machairodont anatomy of F S P - I T D 342 suggests that a s imi la r delayed éruption of dC can be inferred. The ind i v idua l from Itardies represents an earl ier ontogenetic stage t h a n juvénile spécimens of Barbourofelis studied by B ryan t (1988). U F 36871 represents the earl iest stage known for B. loveorum (Bryant, 1988: figs 2 and 3); this is a max i l l a w i t h a ful ly erupted dP3 and a P4 wh i ch is most ly unerupted but vis ible pos-terodorsal ly to dP3. O n this spécimen, dC has not yet erupted and i ts t ip l ies 15 m m above the opening ofthe alveolus. The hypothesis of an unerupted dC i n F S P -ITD 342 is reinforced by the study of U C M P 123181 assigned to Eusmilus cerebralis wh i ch represents a s im i l a r developmental stage (Bryant , 1988: 303). However, the somewhat long diastema on the max i l l a and mandible , the latéral concavity of the diastema and the deep génial flange of F S P - I T D 342 d is t ingu ish th is spécimen from barbourofelines. I l is also important to note that, i n F S P - I T D 342 at least, and possibly i n juvéniles of other Hoplophoneini , the flange appears early dur ing the ontogeny and that i t is close to its adult size when the upper deciduous canines start to erupt. G i ven that at the same dental stage, F S P - I T D 342 has a more developed flange than i n species of Barbourofelis (especially Barbourofelis loveorum), we can assume that the very strong corrélation between the development of the flange and the érupt ion of the canines wh i ch appears i n Barbourofelis is absent i n Eusmilus. Th i s feature is an addi t ional and important différence between n imrav ines and barbourofelines, a l though it remains to be conflrmed i n juvéniles of Hoplophoneus and Nanosmilus.
Estimated body size and âge détermination The sma l l size of the c ran ium from Itardies is due to its young âge. Because body size sometimes discriminâtes taxa, a n est imat ion of the adult size for th is i nd i v i dua l may have taxonomic impl icat ions. Morpholog ica l measurements may help to estimate the adult body size of an ind i v idua l (Van Va lkenburgh, 1990). Some are related to âge: for example, sku l l and mand ibu la r length increase w i th âge. In contrast, denta l measurements are not related to âge. A comparison between deciduous teeth measurements may therefore provide a more accurate est imated body size for th is sma l l ind i v idua l . Table 1 shows that F S P - I T D 342 is the size of A M N H 5337, an ind i v idua l at a s imi la r dental âge (fully erupted dp4, but no permanent teeth) assigned to Hoplophoneus primaevus. Th is suggests F S P - I T D 342 may reach the adult body size of this species oï Hoplophoneus (i.e. L m l ~ 1 8 - 1 9 mm).
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477-493

JUVEJNiL l l i A INATUMY Ut i N l M K A V l U A J i ( .UAKlMVUKAj
The stage of dental development may be used to estimate the odontological âge of th is ind i v idua l . L i t tle is known about th is re lat ionship i n sabretoothed taxa and few studies provide comparative data. Thus the spécimen from Itardies is intermediate between stages III (deciduous cheek teeth not ful ly erupted) and IV (deciduous teeth ful ly erupted, incisors, canines and P4 erupting) defined by Rawn-Schatzinger (1983) for the machairodont ine fel id 'Homotherium' sérum. In comparison w i th several extant felids, this author gave an approximate équivalent âge of 3 -4 months for stage III, and 5-12 months for stage IV. However the 'Homotherium' sérum was larger than the est imated adult size of F S P - I T D 342 and s l ight ly larger than Panthera leo (Rawn-Schatzinger, 1983). G i v en that the t i m i n g of the éruption séquence is earl ier i n smal ler species than i n larger ones, the érupt ion séquence i n F S P - I T D 342 was s l ight ly ear l ier compared to a 'Homotherium' i nd i v i dua l at the same âge. A s a resuit the juvénile ind i v i dua l from Itardies may be est imated as be ing about 5-6 months old.
B E H A V I O U R A L A N D B I O M E C H A N I C A L I N F E R E N C E S
F R O M T H E J U V E N I L E A N A T O M Y I N N I M R A V I N A E
M a n y studies have dealt w i t h the hun t i ng and k i l l i n g behaviour of sabretoothed predators (see B r yan t & Russe l l , 1995; Turner & An ton , 1997). However, most of those concentrated on the adul t anatomy. Due to the nature of the fossil record, the juvénile anatomy of extinct taxa is poorly known and has therefore been rare ly studied. Few works have considered the érupt ion séquence of permanent and deciduous dent i t ion (e.g. Rawn-Schatzinger, 1983; Bryant , 1988) and i ts impl icat ions for the durat ion of parenta l care i n sabretoothed predators (e.g. Bryant , 1990). The spécimen described here displays extremely we l l developed sabretoothed features at an ear ly âge. F ina l l y , was th is juvénile able to k i l l prey? B r y a n t (1990) stated that the deciduous sabres of Barbourofelis were of suffi-cient size to have been used for k i l l i n g . The upper deciduous canines are not yet erupted i n F S P - I T D 342 and therefore th is i nd i v i dua l was probably s t i l l dépendent on adults and not able to part ic ipate i n hun t ing , a l though i t surely processed meat, as d id juvénile Barbourofelis (Bryant , 1990). In order to evaluate the bite strength of th is juvénile, we studied the skeletal attachments of i ts mast icatory apparatus.
Musc l e reconstruct ion helps us to better unders tand the biology of extinct organisms. However, as B r y a n t & Seymour (1990) pointed out, ' it requires assump-tions that have not been adequately tested'. The size of a muscle is ha rd to estimate, and the development of muscle attachment is often not a good indicator of the development and power output of the corresponding muscle (Bryant & Russe l l , 1995). For th is reason, we
ma in l y took the degree of scarr ing and the surface area of attachments as an approx imat ion of the size of the muscle.
M a n y authors argued that i n sabretoothed carnivorans the réduction of the posterior région of the mandib le (especially a low, sma l l coronoid process) is associated w i t h a poor leverage and/or reduced size of j aw adductors, especial ly the temporal is (see review i n B r y a n t & Russe l l , 1995). However, i n sabretoothed predators, th is is compensated for by a more posterior placement of the lower carnass ia l t h a n i n other carnivorans. Th is tooth is located 52% (back of condyle to carnass ia l notch/mandibular length) to 54% of the total mand ibu la r length i n a i l extant carn ivoran fami l les except Hyaenidae (Radinsky, 1981a,b; - 4 2 % i n Crocuta crocuta, pers. data). It is 46% i n adult Hoplophoneus and 42% i n Eusmilus sicarius (Bryant & Russe l l , 1995: table 7.1). The appl icat ion of Greave's (1983, 1985) model for carnass ia l funct ioning i n carnivorans by B r y a n t & Russe l l (1995) shows that the posterior location of the carnass ia l i n n imrav ids may compensate for a réduction i n leverage, but not for the reduced size of adductors. The leverage of the tempora l is i n adult n imrav ids was therefore s im i l a r to that of extant carnivorans, and felids and v iverr ids i n part icular.
Muscle attachments to the cranium A s i n other carnivorans, the development of the adduc-tor muscles i n F S P - I T D 342 is expected to be less developed than that of the adult , wh i ch is confirmed by the anatomy of the c ran ium. The m a i n point of attachment ofthe temporal is muscle on the c ran ium is located i n the temporal fossa. The lack of c ran ia l crests (especially sagittal) on F S P - I T D 342 suggests a re lat ive ly sma l l at tachment area for the temporal is . The posterior part of the c ran ium is an important région where numerous muscles involved i n the neck apparatus attach. The digastr ic muscle is the only muscle wh i ch belongs to the mast icatory apparatus wh i ch attaches on this région, on the paroccipital apophysis i n extant mamma l s (Turnbu l l , 1970; Barone, 2000). In the most special ized n imrav ids such as Hoplophoneus and Eusmilus, the posterior extension of the paroccipi t a l process is strongly reduced, wh ich results i n a réduction of the surface of the attachment for the digastr icus. Moreover, the attachment area is also moved dorsally, wh i ch may be an adaptat ion wh i ch préserves a potential stretch for the muscle, especially at a m a x i m u m gape when the distance between the insert ions of the muscle is shortest. The paroccipital process is crest-l ike i n the juvénile F S P - I T D 342. M a n y of the neck muscles attach on the posterior port ion of the c ran ium. A i l of them act for the mobi l i ty and stabi l i ty of the neck and, dur ing the k i l l i n g pro-
© 2003 The L innean Society of London, Zoological Journal of the Linnean Society, 2003, 138, 477 -493

s . r j j i U j iNJ i a n a L. uhj t s u i M S
cess for example, l im i t the head movement and pre-vent any damage to the sabres. The muscu lar arrangement of th is région is not described here in for at least two reasons. F i r s t , the juvénile spécimen F S P -ITD 342 was probably not capable of k i l l i n g or hun t i ng any prey and second, the location of the attachment for the numerous muscles wh i ch insert i n that part of the c ran ium is diff icult to identify. Such a reconstruct ion is par t i cu lar ly diff icult i n the case of n imrav ids wh ich have no extant relatives. A s a resuit , we have restr icted our descript ion to the deepest at tachment wh ich is located on the ventra l and latéral surface of the masto id process on F S P - I T D 342 (solid circles i n F i g . 3). A s s u m i n g that th is at tachment is homologous to that wh i ch occurs i n the same région of other carnivorans, this may be the insert ion for the brachio-cephalicus as described i n the great panda (Davis, 1964), or the insert ion of the brachiocephal icus and sternocephalicus wh i ch are very close i n Canis famil-iaris (Gaspard, 1971: fig. 3 0 ; Barone, 1986: 314, p l . 147). The function of thèse flexor muscles is roughly s imi lar : the sternocephalicus draws the head and neck to one side, and the brachiocephal icus b i la tera l ly fixes the neck (Evans, 1993). If correct, thèse muscles were par t i cu lar l y strong and large i n the juvénile. Wi thout a complète dent i t ion, especially a lack of sabres, F S P -ITD 342 would not have been capable of k i l l i n g any prey. However those muscles may have been involved i n the feeding behaviour of F S P - I T D 342, especial ly i n feeding on carcasses. There is a strong re lat ionship between the act ion of neck, the use of teeth and the type of food i n some extant carnivorans (Van Va lkenburgh, 1996). Thus , no action o f the neck is typ ica l for a bite us ing the cheek teeth, but the use of f rontal teeth is often associated w i th pu l l i ng or tw is t ing . Thèse actions are most often associated w i t h feeding on muscle and s k i n p lus associated connective t issue or muscle (Van Va lkenburgh , 1996). The development ofthe neck musculature on th is juvénile may therefore be re lated w i t h the strongly curved arcade of can in i form and curved incisors; the importance of the inc i sors i n feeding has also been demonstrat ing i n adults of sabretoothed species (B iknev ic ius , V a n Va lkenburgh & Walker, 1996).
The mandible A s im i l a r development of the coronoid process i n adults and juvéniles of Hoplophonein i suggests a s imi l a r surface for the attachment of the temporal is on the mandible . Th is muscle is the largest and most powerful muscle i n the mast icatory apparatus. In comparison to other carnivorans, n imrav ines have addi t iona l scarr ing on the med ia l face of the mandib le beneath the posterior alveolus of m l . Th i s rugose knob is présent i n F S P - I T D 342 (Fig. 4A) and is near ly as
well-developed as i n adults of Hoplophoneus, Eusmilus and Dinictis. Th is scar is rare ly présent i n Nimravus. In adults of Eusmilus and Hoplophoneus, i t is located below the ta lon id of m l ; i n Dinictis, i t is more posteriorly located and beneath m2. Th is scar is not l ike ly to be re lated to the temporal is wh i ch inse r t s in to the mandib le on the coronoid process. However, a part of th is muscle, the deep temporal is , does insert on the media l face of the process. Th is muscle may extend downward, to jus t above the dorsal border of the mandibular foramen (i.e. lower or close to the level of the alveol i o f the posterior molar) i n urs ids (Davis, 1964), procyonids (Gorniak, 1986), and the domestic cat (Hovelacque & Virenque, 1913b; T u r n b u l l , 1970), or i t may l ie dorsal to the posterior molar occlusal plane as i n canids (Gaspard, 1971). In the majority of mammals the temporal is does not insert on the hor izonta l port ion o f the mandib le (Hovelacque & Virenque, 1913b; Davis , 1964; T u r n b u l l , 1970; Gaspard , 1971). Moreover, the deep temporal is has mostly fleshy insert ions on the coronoid process (Turnbu l l , 1970; Gorn iak , 1986). In extant taxa its at tachment is usua l ly a rugose area associated i n some species, w i th a shal low fossa as i n Mêles but not w i t h a prominent scarr ing. F l eshy or direct attachments are often associated w i th a rugose area, and tendinous and aponeurotic attachments w i t h osteological features (Bryant & Eusse l l , 1995). Accord ing to B r yan t (1984: 165), who first described th is structure, this is 'probably the point of attachment for the pterygoideus muscle, wh i ch adds power to the bite and restrains the jaws, prevent ing desart iculat ion. ' G i v en that th is scarr ing has not been noticed i n any extant carnivorans and that i t may have conséquences for the muscle arrangement of the mast icatory apparatus, we more carefully addressed Bryant ' s hypothesis. Besides the temporal is , several other muscles wh i ch are most ly re lated to masticatory function insert on the med ia l face of the posterior ha l f of the mandible . We examined the pterygoideus muscles, wh i ch were singled out i n Bryant ' s hypothesis. The pterygoideus lateral is (= external pterygoideus) is a sma l l muscle wh i ch invar iab ly inserts media l ly and s l ight ly ventra l ly to the condylar head of the mandible i n mammals . Th i s has been we l l documented i n Car n ivora (Hovelacque & Virenque, 1913b; Dav is , 1964; Barone, 1967, 2000; T u r n b u l l , 1970; Gaspard , 1971; La i son & Lau t r ou , 1975; Lau t r ou & La ison , 1975; Gorn iak , 1986). Thèse authors also dissected and described the pterygoideus media l is (= in te rna i pterygoideus). The i r observations are consistent, and suggest a re lat ive ly constant mand ibu la r at tachment of this muscle w i t h i n carnivorans. The pterygoideus media l is is a large muscle wh i ch inserts ventra l ly and posteriorly to the mand ibu la r foramen at the posterior end of the angular process. In other mammals , the pterygoideus media l is may be a l i t t le more extended
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477-493

d U V E J N l L E AJNATUMÏ U F J N I M K A V I U A E (tJAKJNlVUKA) 4 9 1
in te r io r l y and dorsal ly toward the mand ibu la r foramen (see Equus, Odocoileus, Ovis i n T u r n b u l l , 1970: i gs 17B, 22C and 26B, respectively). Gaspard (1971: îg. 3) also noted addi t ional muscles wh i ch insert near ;he mand ibu la r foramen: the vascular l a m i n a and the nterpterygoideus aponeurosis, wh i ch have indeed Deen precisely described i n some mamma ls ( inc lud ing :he domestic cat and dog) by Hovelacque & Vi renque 1913a,b). W i t h i n carnivorans, the insert ion of thèse muscles is l inear and narrow, s tar t ing from the poste-dor border of the mandib le up to the mand ibu la r foramen. The vascular l a m i n a remains dorsal to the ^oramen, whereas the interpterygoideus aponeurosis les ventral ly. J u s t anter ior to the foramen, the apo-leuros is reaches the dorsomedial face of the posterior 3nd of the mylohoideus. Whi l e Bryant ' s hypothesis is lo t confirmed (no re lat ion w i t h the pterygoideus lat-îralis or medial is ) , the best candidate for th is u n u s u a l i t tachment remains the interpterygoideus aponeuro-3is of Hovelacque & Virenque. It inserts i n a région i^ery close to th is scar, at least i n dogs and cats. How-3ver i t is important to note that only a careful dissec-don of the interpterygoideus région provides some îvidence for the présence of those structures, wh i ch lave rare ly been mentioned or i l lus t ra ted i n Car -l i v o r a (e.g. Hovelacque & Virenque, 1913b; Gaspard , 1971). Moreover, despite the précise descript ion and l lustrat ions avai lable, we were not able to identify a précise muscu lar attachment area i n re lat ion to thèse muscles i n extant Carn i vo ra . U n t i l further anatomical nvestigations have been under taken, we only assume ;hat th is scar is re lated to the interpterygoideus région and that i t is unusua l i n Carn i vo ra . No part ic-i l a r mode of feeding is suggested here from the pres-iïice of th is feature.
^owerful bite and hunting capabilities [n contrast to the adult , shear ing occlusion i n the ;heek teeth occurs between dp4 and dP3 i n juvénile carnivorans. In F S P - I T D 342, dp4 is located at the midpoint of the total mand ibu la r length, as i n Dinictis i du l t (Bryant & Russe l l , 1995: table 7.1). Accord ing to jreave 's (1983, 1985) model , a more anterior posit ion 3f dp4 impl ies a more reduced bite force at dp4 i n F S P -[TD 342 than i n adults of Hoplophoneini . Bo th the Dosition of dp4 and the size of muscula ture results i n 1 weaker bite i n the juvénile, as was suspected from ;he development and topology of the muscles involved n mast icat ion.
R E L A T I O N S H I P S O F N I I V I R A V I N A E T O
B A R B O U R O F E L I N A E
\n between the juvénile anatomy suggests ;hat the N imrav inae differ from the Barbourofel inae
i n the ontogeny of the auditory bu l l a and development of sabretoothed features. Thus , w i th in the N i m r a v i nae, the ectotympanic rotat ion away from the s k u l l appears marked ly ear l ier dur ing the ontogeny; w i t h i n the most sabretoothed n imrav ines , the Hoplophone in i , we d id not find a strong corrélation between the éruption of the upper canines and development of other sabretoothed features (e.g. mand ibu la r flange) wh i ch occurs i n the Barbourofel inae (especially Barbourofelis).
It is beyond the scope of th is paper to discuss the re lat ionships of the N imrav inae and Barbourofel inae i n depth. However, readers can f ind a careful discussion of th is topic i n two récent works report ing on dis-coveries of early barbourofelines (Morales et al., 2001; M . Mor lo , S. Peigné & D. Nage l , unpub l . data). We here provide the m a i n conclusions of thèse works. Thus , Mora les et al. (2001) reported on the discovery of barbourofelines from the early Miocène of A f r i ca and Spa in . Accord ing to thèse authors, the dental morphology of thèse spécimens suggests that the Miocène N imrav idae (i.e. Barbourofel inae) are not closely re lated to the Palaeogene N imrav idae (Nimravinae) , and that they should be assigned to the Fel idae. Mor lo et al. (unpubl . data) also reported on a new species of pr imi t i ve Barbourofel inae from the middle Miocène of Germany, and confirmed the dist inct ion between the two subfamil ies. However, on the basis of deep différences i n the auditory bu l l a between barbourofelines and felids, they propose to erect the Barbourofel inae to the fami ly rank.
ACKNOWLEDGEMENTS
We thank everyone who has part ic ipated i n the f ie ld campaigns i n Itardies, and to a i l ind iv idua ls who have given us access to collections i n the ir care: K . He iss ig (BSP) , R. M . H u n t J r . and G. Corner ( U N S M ) , M . C. M c K e n n a and R. H . Tedford ( A M N H ) , D. Môrike (SMNS ) , P. Tassy ( M N H N ) , M . A . Turner (YPM) and X . Wang ( L A C M , previously A M N H ) . We are deeply grateful to H . B ryan t and R. H u n t , who k ind l y agreed to review a f ina l draft of th is paper, for the ir constructive cr i t i c ism, and to two anonymous référées whose comments contr ibuted to improvements i n our paper. We are indebted to E . Stepleton and R. H u n t (Univers i ty of Nebraska) for m a k i n g and sending us a very nice cast of the c ran ium and mandible of Nanosmilus kurteni wh i ch was used i n comparison. We also thank S. Ri f faud (FSP) for drawing the figures and V. Bellefet ( F S P ) for photographs. A part of th is study was supported by grants from the Amer i can Muséum of N a t u r a l H is tory (collections study grant) and the Un i ve r s i t y of Po i t i e r s -CNRS U M R 6046 'Géobiologie, Biochronologie et Paléontologie humaine ' to S.R
3 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477 -493

4^2 S. F E I G N E and L. D E B O N I S
REFERENCES
Barone R. 1967. L a myologie du l ion (Panthera leo). Mammalia 31: 459-514.
Barone R. 1986. Anatomie comparée des mammifères domestiques. Vol . 1. Ostéologie. Par is : Vigot.
Barone R. 2000. Anatomie comparée des mammifères domestiques. Vol . 2. Arthrologie et myologie. Par is : Vigot.
Baskin JA . 1981. Barbourofelis (Nimravidae) and Nimravides (Felidae), w i th a description of two new species from the late Miocène of F lor ida . Journal of Mammalogy 62: 122-139.
Berta A. 1985. The status of Smilodon. No r th and South Amer ica . Contributions in Sciences of the Natural History Muséum, Los Angeles County 370: 1-15.
Biknevicius AR, Van Valkenburgh B, Walker J . 1996. Incisor size and shape: impl icat ions for feeding behaviors i n saber-toothed 'cats'. Journal of Vertébrale Paleontology 16: 510-521.
BiochroM'97.1997. Synthèses et tableaux de corrélations. In: Agu i l a r J P , Legendre S, M i chaux J , eds. Actes du congrès BiochroM'97. Mémoires et travaux de l'EPHE. Institut de Montpellier, 21: 769-305.
Blondel C. 2001. The Eocene-Oligocene ungulates from Western Europe and their environment. Palaeogeography, Palae-oclimatology, Palaeoecology 168: 125-139.
Bovard J F . 1907. Notes on Quaternary Fel idae from Califor-n ia . University of California Publication Bulletin, Geology 5: 155-170.
Bryant L J . 1984. Skeletons of the d iminut ive sabertooth Eusmilus from the Ar ikareean of South Dakota . In: Menge l R M , ed. Papers in vertébrale paleontology honoring Robert Warren Wilson. Carnegie Muséum of Natural History spécial publication 9: 161-170.
Bryant H N . 1988. Delayed éruption of the deciduous upper canine i n the sabretoothed carnivora Barbourofelis lovei (Carnivora, Nimravidae) . Journal of Vertébrale Paleontology 8: 295-306.
Bryant H N . 1990. Implications of the dental éruption séquence i n Barbourofelis (Carnivora, Nimravidae) for the function of upper canines and the durat ion of parenta l care i n sabertoothed carnivores. Journal ofZoology 222:585-590.
Bryant HN . 1991. Phylogenetic relat ionships and systematics of the N imrav idae (Carnivora). Journal of Mammalogy 72: 56-78.
Bryant HN . 1996. N imrav idae . In: Prothero DR, E m r y R J eds. The terrestrial eocene-oligocene transition in North America. New York: Cambridge Un ivers i t y Press, 4 5 3 -475.
Bryant HN, Russell AP . 1995. Carnass ia l functioning i n n imrav id and felid sabertooths: theoretical basis and robustness of inferences. In; Thomason J J , ed. Functional morphology in vertébrale paleontology. Cambridge, U K : Cambridge Un ivers i t y Press, 116-135.
Bryant HN , Seymour K L . 1990. Observations and comments on the re l iabi l i ty of muscle reconstruction i n fossil verte-brates. Journal of Morphology 206: 109-117.
Cope E D . 1873. Synopsis of new Vertebrata from the Tert iary of Colorado, obtained dur ing the summer of 1873. Annual
report of the Hayden Geological and Geographical Survey of the Territories 7: 1-19.
Cope E D . 1878. O n some characters of the Miocène fauna of Oregon. Proceedings of the American Philosophical Society 18: 63-78.
Cope E D . 1880. Notes on sabre-tooths. American Naturalist 14: 833-858.
Cope E D . 1893. A new Plistocene sabre-tooth. American Naturalist 27: 896-897.
Crouch J E . 1969. Text-atlas ofcat anatomy. Ph i lade lphia , PA: L ea & Febiger.
Davis DD. 1964. The giant panda. A morphological study of evolutionary mechanisms. Fieldiana: Zoology Memoirs 3: 1-339.
Emerson SB, Radinsky LB . 1980. Funct iona l analysis of sabertooth crania l morphology. Paleobiology 6: 295-312.
Evans H E . 1993. Miller's anatomy of the dog. Ph i lade lph ia , PA: W B Saunders Co.
Fi lho l H . 1872. Note relative à l a découverte dans les gisements de phosphate chaux du Lot d'un mammifère fossile nouveau. Bulletin de la Société Des Sciences Physique et Naturelle, Toulouse 1: 92-94.
Gaspard M. 1971. Anatomie comparative et fonctionnelle de l a musculature masticatrice chez les Carnivores. Mémoires Du Muséum National d'Histoire Naturelle, Paris A 63: 1-207.
Gorniak GC . 1986. Archi tecture of the masticatory apparatus in Eas t e rn raccoons (Procyon lotor lotor). American Journal of Anatomy 176: 333-351.
Greaves WS. 1983. A functional analysis of carnassia l bit-ing. Biological Journal of the Linnean Society 20: 3 5 3 -363.
Greaves WS. 1985. The generalized Carnivore jaw. Zoological Journal of the Linnean Society 85: 267-274.
Hatcher J B . 1895. Discovery, i n the Oligocène of South Dakota , of Eusmilus, a genus of saber-toothed cat new to No r th Amer ica . American Naturalist 29: 1091-1093.
Hovelacque A, Virenque M. 1913a. Les formations aponévrotiques de l a région ptérygo-maxillaire chez l 'Homme et chez quelques Mammifères. Journal de l'Anat-omie et de la Physiologie, Paris 49: 427-482.
Hovelacque A, Virenque M. 1913b. Les formations aponévrotiques de l a région ptérygo-maxillaire chez l 'Homme et chez quelques Mammifères. Journal de l'Anat-omie et de la Physiologie, Paris 49: 618-704.
Hunt R M J r . 1974. The auditory bu l la i n Carn ivora : an anatomical basis for reappraisal of Carnivore évolution. Journal of Morphology 143: 21-76.
Hunt R M J r . 1987. Evo lut ion of the aeluroid Carn ivora : sig-nificance of auditory structure i n the n imrav id cat Dinictis. American Muséum Novitates 2886: 1-74.
Joeckel R M , Peigné S, Hunt R M J r , Skolnick RI. 2002. The auditory région and nasal cavity of Oligocène N i m ravidae (Mammal ia : Carnivora) . Journal of Vertébrale Paleontology, 22: 830-847.
Kretzoi M . 1929. Mate r i a l i en zur phylogenetischen K lass i -fikation der Aeluroideen. Xth Congress International ofZoology, 1293-1355.
© 2003 The L innean Society of London, Zoological Journal ofthe Linnean Society, 2003, 138, 477-493
Copyright © 2022 FDOKUMEN




















