
The shallow water gobiid assemblage of the Venice Lagoon: abundance, seasonal variation and habitat...
20
The shallow water gobiid assemblage of the Venice Lagoon: abundance, seasonal variation and habitat partitioning S. M ALAVASI *, A. F RANCO , R. F IORIN , P. F RANZOI , P. T ORRICELLI AND D. M AINARDI Department of Environmental Sciences, University of Venezia, Campo della Celestia, Castello 2737/b, 30122 Venezia, Italy The gobiid assemblage of the Venice Lagoon shallow waters was investigated by means of a semi-quantitative standardized sampling (using a small beach seine), stratified into five main types of shallow subtidal habitats and conducted on a seasonal basis during 1 year. The degree of overlap in resource utilization among six coexisting goby species was assessed, along both the time axis, by analysing the seasonal variation in abundance and reproductive status (as revealed by the gonado-somatic index) and the habitat axis, by comparing species abundance across different habitat types and controlling for the effects of some abiotic factors. Smaller species, and especially the marbled goby Pomatoschistus marmoratus, dominated the local assemblage. Although the cycle of shallow water colonization and seasonal variation in total abundance were basically similar, species showed differences in timing of reproduction and recruitment, as well as in habitat preference. The larger species belonging to the genera Gobius and Zosterisessor tended to overlap their habitat use, being more abundant in seagrass habitats than in the unvegetated habitats, whereas the smaller species belonging to the genera Knipowitschia and Pomatoschistus avoided seagrasses, preferring in most cases mud flats and salt marsh creeks. Within these two groups of species some further slight differences in species habitat preference, relationship with abiotic factors and reproductive ecology could be detected. Results are discussed in the light of both ecological mechanisms underlying coexistence of closely related species and the current knowledge of the phylogeny of Mediterranean gobies. # 2005 The Fisheries Society of the British Isles Key words: abundance; gobies; habitat partitioning; shallow waters. INTRODUCTION Fish communities are usually composed of sub-sets of taxonomically related and ecologically similar species (‘guilds’), among which resources are finely parti- tioned (Wilkins & Myers, 1992; Van Snik Gray & Stauffer, 1999; Lombarte et al., 2000; Plattell & Potter, 2001; Gladfelter & Johnson, 2004). Phylogenetically related species often show large overlap in resource utilization along one or more ecological axes (habitat, food and time) and for this reason the comparative study of resource utilization in such sub-sets of species is particularly fruitful for *Author to whom correspondence should be addressed. Tel.: þ39 041 2347739; fax: þ39 041 5281494; email: [email protected] Journal of Fish Biology (2005) 67 (Supplement B), 146–165 doi:10.1111/j.1095-8649.2005.00919.x, available online at http://www.blackwell-synergy.com 146 # 2005 The Fisheries Society of the British Isles
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The shallow water gobiid assemblage of the Venice Lagoon: abundance, seasonal variation and habitat...

The shallow water gobiid assemblage of the Venice
Lagoon: abundance, seasonal variation and habitat
partitioning
S. MALAVASI*, A. FRANCO, R. FIORIN, P. FRANZOI,P. TORRICELLI AND D. MAINARDI
Department of Environmental Sciences, University of Venezia, Campo della Celestia,Castello 2737/b, 30122 Venezia, Italy
The gobiid assemblage of the Venice Lagoon shallow waters was investigated by means of a
semi-quantitative standardized sampling (using a small beach seine), stratified into five main
types of shallow subtidal habitats and conducted on a seasonal basis during 1 year. The
degree of overlap in resource utilization among six coexisting goby species was assessed, along
both the time axis, by analysing the seasonal variation in abundance and reproductive status
(as revealed by the gonado-somatic index) and the habitat axis, by comparing species
abundance across different habitat types and controlling for the effects of some abiotic
factors. Smaller species, and especially the marbled goby Pomatoschistus marmoratus,
dominated the local assemblage. Although the cycle of shallow water colonization and
seasonal variation in total abundance were basically similar, species showed differences in
timing of reproduction and recruitment, as well as in habitat preference. The larger species
belonging to the genera Gobius and Zosterisessor tended to overlap their habitat use, being
more abundant in seagrass habitats than in the unvegetated habitats, whereas the smaller
species belonging to the genera Knipowitschia and Pomatoschistus avoided seagrasses,
preferring in most cases mud flats and salt marsh creeks. Within these two groups of species
some further slight differences in species habitat preference, relationship with abiotic factors
and reproductive ecology could be detected. Results are discussed in the light of both
ecological mechanisms underlying coexistence of closely related species and the current
knowledge of the phylogeny of Mediterranean gobies. # 2005 The Fisheries Society of the British Isles
Key words: abundance; gobies; habitat partitioning; shallow waters.
INTRODUCTION
Fish communities are usually composed of sub-sets of taxonomically related andecologically similar species (‘guilds’), among which resources are finely parti-tioned (Wilkins & Myers, 1992; Van Snik Gray & Stauffer, 1999; Lombarteet al., 2000; Plattell & Potter, 2001; Gladfelter & Johnson, 2004). Phylogeneticallyrelated species often show large overlap in resource utilization along one or moreecological axes (habitat, food and time) and for this reason the comparativestudy of resource utilization in such sub-sets of species is particularly fruitful for
*Author to whom correspondence should be addressed. Tel.: þ39 041 2347739; fax: þ39 041 5281494;
email: [email protected]
Journal of Fish Biology (2005) 67 (Supplement B), 146–165
doi:10.1111/j.1095-8649.2005.00919.x,availableonlineathttp://www.blackwell-synergy.com
146# 2005TheFisheries Society of theBritish Isles

the understanding of the ecological mechanisms underlying coexistence(Tokeshi, 1999).Resource partitioning among animal species contributes to the reduction of
competition intensity and to the consequent enhancement of species coexistence(Tokeshi, 1999). In aquatic communities, the food axis has been considered tohave the highest relative importance in promoting resource partitioning,although habitat is placed first in general terms (Schoener, 1974; Ross, 1986).Within estuarine and coastal lagoon fish assemblages, sub-sets of taxonomicallyrelated species and ecological guilds are easily recognizable and, among them,gobies represent an important group both in terms of number of species andrelative abundance (Thorman & Wiederholm, 1983; Elliott & Dewailly, 1995;West & King, 1996; Mathieson et al., 2000; Malavasi et al., 2004). Gobies share asimilar morphology which, together with a number of other behavioural andphysiological adaptations, is the result of an evolutionary history of a benthic lifestyle (Miller, 1984). Gobiid assemblages have been studied in northern Europeanestuaries (Fonds, 1973; Zander, 1982; Zander & Berg, 1984; Wilkins & Myers,1992), in Mediterranean coastal lagoons (Costello, 1992; Arruda et al., 1993;Boucherau & Guelorget, 1997; Pampoulie et al., 2001) and sub-tropical andtropical estuaries (Gill & Potter, 1993; Syms & Jones, 2004) and patterns offood and habitat partitioning among species have been shown in many cases.The diversification of Mediterranean gobies has been probably driven by sub-
sequent vicariant events, such as the separation caused by the mid-Miocene isola-tion of the Paratethys basin from the Mediterranean Tethys Sea and the strongenvironmental changes to which the Mediterranean basin was subjected during theMessinian salinity crisis in the Holocene followed by subsequent re-flooding pro-cesses (McKay & Miller, 1991; Miller, 2004). This process has led to the evolutionof several species adapted to a wide spectrum of habitats, characterized by differentsalinity levels (from truly marine to brackish and freshwater habitats) and differentsubstratum types. Most of these species occur in the coastal lagoons, as theseenvironments are composed of a mixture of habitats, determined by several gra-dients in both abiotic and biotic factors (Elliott & Hemingway, 2002). These specieshave therefore undergone both isolation and differentiation in the past as aresponse to environmental changes, and then interactions and coexistence withinthe same transitional environments. According to Tokeshi (1999), the assessment ofthe degree of overlap in resource use has to be considered as an important step forthe understanding of community organization and patterns of species coexistence.Within this theoretical framework based on the concepts of niche segregation, thepresent study aims to assess the degree of resource overlap along the temporal andthe spatial axes among coexisting gobies living in the largest lagoon of theMediterranean Sea, the Venice Lagoon.Abundance and distribution of the goby species found in this environment were
recorded bymeans of a standardized semi-quantitative sampling (using a small beachseine), stratified into five main habitat categories and conducted on a seasonal basis.Patterns of temporal segregation and overlap among species (time axis) were
studied by comparing the seasonal trends in abundance and reproductive status(as revealed by the gonado-somatic index), whereas the habitat axis was studiedby comparing species abundance among different habitat types by controllingfor the effects of some abiotic factors. Further, patterns of interspecific spatial
SHALLOW WATER GOBIES OF THE VENICE LAGOON 147
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

association were investigated, adopting both a presence and absence and acorrelation approach. The current knowledge of the evolutionary history of theinvestigated species allowed the results to be discussed by taking into considera-tion the interplay between phylogeny and ecological processes, such as predationand competition, and their effects on the present distribution of these specieswithin the lagoon environment.
MATERIALS AND METHODS
STUDY SITE
The Venice Lagoon (Fig. 1) is situated in the northern Adriatic Sea and is the largestlagoon in the Mediterranean basin, with an area of c. 540 km2. It is a generally shallowwater area (average depth c. 1 m), being mostly composed of large shallow areasconnected by a network of channels. Deeper shipping canals are present near to thethree wide mouths (Lido, Malamocco and Chioggia) connecting the lagoon to the sea.
12° 12′ E
12° 12′ E
45° 24′ N45° 12′ N
45° 2
4′ N
45° 1
2′ N
12° 24′ E
12° 24′ E
12° 36′ E
12° 36′ E
Chioggia
Malamocco
VENICE
Lido
10° 00′ E 15° 00′ E
40° 0
0′ N
45° 0
0′ N
5 km
N
Adr
iatic
Se
a
FIG. 1. Sampling stations in the Venice Lagoon, divided by type of habitat (�, seagrass bed; , sparsely
vegetated habitat; , sand flat; &, mud flat;: , salt marsh creek). The two watersheds (~) dividing
the lagoon into the northern, central and southern sub-basins are shown.
148 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

Through them, tidal flows enter the lagoon, with tidal ranges of up to 1 m in height, thehighest in the Mediterranean Sea.High levels of spatial heterogeneity are present in the lagoon, because this system is
subjected to strong temporal and spatial changes in morphological and physico-chemicalvariables (Sacchi, 1985). Moreover, from a hydrological point of view, the lagoon area isdivided by two main watersheds into three sub-basins: northern, central and southern(Avanzi et al., 1979) (Fig. 1).Many types of habitat are present within the lagoon environment, such as seagrass
beds, sand and mud flats, and salt marshes. The seagrasses, mainly Zostera marina L. andCymodocea nodosa (Ucria) Asher (secondly Nanozostera noltii Hornem), consitute largemeadows especially in the central and southern sub-basins, whereas their retreat has beenrecently observed in comparison with 1990 in the northern basin (Rismondo et al., 2003).Seagrasses are mostly located in the proximity of the sea inlets and deeper canals, wheresediments contain higher percentages of sand. By contrast, habitats with muddy bottoms,such as mud flats and salt marshes, are found in the internal areas of the lagoon, fartherfrom the sea inlets, where some freshwater inflows are present.Seasonally, some macroalgae (mainly Ulva rigida C. Agardh) could be detected in the
lagoon, although they have almost completely disappeared from the lagoon as a result ofa declining trend begun in the 1990s (Sfriso et al., 2003).
SAMPLES AND DATA COLLECTION
The sampling was carried out on a seasonal basis, during the spring (April to May),summer (July to August) and autumn (October to November) of 2002, at 68 stationsdistributed in the shallow waters of the Venice Lagoon (Fig. 1). The sampling gear usedwas a 10 m long beach seine (mesh-size 2 mm), which was trawled over an area of 360 m2
(three replicate trawls, each one covering an area of 120 m2) at each station. The stationswere located throughout the northern, central and southern lagoon sub-basins and theywere allocated to five different main subtidal habitat categories (Fig. 1), previouslydetermined on the basis of preliminary surveys.The habitat types were identified on the basis of general morphological characteristics
and substratum type of the sampling stations, and were: (1) seagrass beds (SB; n ¼ 15);(2) sparsely vegetated habitats (SVh; n ¼ 16), where patches of seagrasses are present onbare sand-muddy bottoms, usually adjacent to seagrass beds; (3) sand flats (Sh; n ¼ 8);(4) mud flats (MFh; n ¼ 17); (5) salt marsh creeks (SMc; n ¼ 12).Gobies were identified to species, following Miller (1986). Fishes were sexed on the
basis of the shape of the uro-genital papilla (Miller, 1984) and the presence of secondarysexual characters was also noted. After being anaesthetized with an overdose of MS-222,fishes were preserved in 10% formalin solution and transported to the laboratory formorphometric analysis: standard length (LS, � 0�1 cm, using callipers) was measured foreach individual, whereas body mass (MT, � 0�01 g, using analytical balance) and gonadmass (MG, � 0�01 g, using analytical balance) were measured in random sub-samples (atleast 15 individuals), to calculate the gonado-somatic index (IG). WhereIG ¼ 100MGMT
�1. The number of individuals of each goby species at each site on eachsampling occasion was recorded and expressed as fish density, i.e. number of individualsper 100 m2.At each sampling station data on the main abiotic characteristics were collected. Water
temperature (�0�1� C) and salinity (�0�5) were measured before sampling, using a digitalthermometer and a temperature-compensated refractometer, respectively. Other environ-mental data were obtained for each sampling station in each season through the GISmapping (by kriging interpolation method) of some variables from the database resultingfrom the 2002 MELA1 survey, a monitoring project carried out during 2002 in the VeniceLagoon by Consorzio Venezia Nuova, on behalf of the Magistrato alle Acque, Venezia(Ministero delle Infrastrutture e dei Trasporti, 2003). These variables characterized theaverage features of the water column, such as dissolved oxygen (% of saturation) andturbidity (FTU), the bottom characteristics, such as percentages of clay and sand in the
SHALLOW WATER GOBIES OF THE VENICE LAGOON 149
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

superficial (0–15 cm) sediments of the lagoon, and the habitat characteristics, such asper cent area covered by salt marshes around each station (measured within 150 m fromthe station, on the basis of GIS maps provided by the Magistrato alle Acque, Venezia)and seagrass coverage score (semi-quantitative measure based on the percentage ofseagrass coverage, obtained as follows: 0, 0% coverage; 1, 0–5%; 2, 5–50%; 3, 50–75%; 4, 75–100%). The seasonal mean values and ranges of variation of the abovementioned abiotic variables are given in Table I.
DATA ANALYSIS
According to the observed IG and body size relationships, individuals were separatedinto ‘breeding’ and ‘non-breeding’ during spring and summer, when the spawning seasonof most species occurs, whereas during autumn fish were assumed to form a functionallyhomogeneous group. Non-breeding individuals coincide in most cases with the young-of-the-year, sexually indeterminate fishes. The effects of habitat type on the total abundanceof each goby species were tested, within each season, by means of Kruskal–WallisANOVA, and multiple comparisons between habitats were performed by means of theMann–Whitney U-test, as data did not meet the assumptions needed for the parametricANOVA. When the sample size was reasonably large, Kruskal–Wallis ANOVA was alsoperformed separately on breeding and non-breeding individuals, to test for differentialdistribution across habitats between these two groups of fish, to discriminate between thereproductive and the nursery and feeding function of the different habitats for eachspecies.
The effects of each environmental factor on goby abundance was tested by means ofSpearman correlation analysis (rs), using all the above mentioned variables. Since thesevariables also included some habitat characteristics, used to define the habitat categories,such as salt marsh and seagrass coverage, the real effects of abiotic factors was deter-mined by removing the effects of these two variables, using rs.
The combined effects of habitat structure and abiotic variables on seasonal speciesabundance were analysed by using the canonical correspondence analysis (CCA) (Manly,1994). The analysis was performed on fish abundance (log10 transformed), abiotic vari-ables and habitat structure data (both standardized to distributions with mean 0 andvariance 1), for each sampling season. Rare species were downweighted.
The degree of overlap among species in habitat utilization was studied by testing theseasonal interspecific association between each species and all the others, using bothpresence and absence data and abundance. The presence and absence data were analysedusing a w2 test (with Yates correction for small numbers of observations), which is basedon a contingency table of the frequencies of seasonal occurrence of the different species inthe sampling stations (Henderson, 2003). The degree of positive or negative associationbetween the species was considered at the 5% level of significancy. The densities of eachspecies in each season were then correlated with the densities of all the others by means ofa Spearman correlation test.
RESULTS
GENERAL PATTERNS
Seven goby species were recorded, three of relatively larger body size [grassgoby Zosterisessor ophiocephalus (Pallas), black goby Gobius niger L., and giantgoby Gobius cobitis (Pallas)] and four of smaller size [marbled gobyPomatoschistus marmoratus (Risso), sand goby Pomatoschistus minutus(Pallas), black spotted goby Pomatoschistus canestrinii (Ninni), and lagoongoby Knipowitschia panizzae (Verga)]. A total of 20 146 individuals was caughtin the lagoon. The most abundant species was P. marmoratus, which represented
150 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

TABLEI.
Seasonalmeanvalues
andranges
ofvariationofthemeasuredenvironmentalvariables
Spring
Summer
Autumn
Mean
Minim
um
Maxim
um
Mean
Minim
um
Maxim
um
Mean
Minim
um
Maxim
um
Tem
perature
(�C)
18�6
14�5
24�0
26�2
23�9
29�0
16�7
12�8
19�9
Salinity
28�9
8�0
36�0
27�2
12�0
32�0
29�0
2�0
36�0
Dissolved
oxygen
(per
centsaturation)
95�2
89�1
102�1
93�4
80�3
104�5
90�9
85�7
93�6
Turbidity(FTU)
19�2
9�1
39�2
17�0
6�9
44�4
14�5
4�1
47�5
Sand(%
)34�4
2�5
85�8
33�4
2�5
85�8
33�2
2�5
85�8
Clay(%
)17�0
4�2
44�2
17�2
4�2
44�2
17�0
4�2
44�2
Saltmarshes
(%)
11�3
0�0
74�0
10�6
0�0
74�0
11�2
0�0
74�0
Seagrasses
(scored)
1�1
0�0
4�0
1�1
0�0
4�0
1�0
0�0
4�0
SHALLOW WATER GOBIES OF THE VENICE LAGOON 151
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

c. 55% of the total catch (Table II). This species was followed, in terms ofrelative abundance, by K. panizzae (23%), whereas the larger gobies, Z. ophio-cephalus, G. niger and G. cobitis were less abundant (8, 0�9 and 0�3% of the totalcatch, respectively). Another large goby is present in the Venice Lagoon, therock goby Gobius paganellus L. (S. Malavasi, pers. obs.), but it was not caught inthis study, as this species is usually associated with rocky habitats (Miller, 1986),which were not suitable for the sampling technique used in the present study.The giant goby was rarely caught (<1%) for the same reasons, so that it has notbeen considered further in the analysis.Some species were recorded in each type of habitat throughout the year,
although with different abundance, such as the grass goby, the marbled gobyand the black goby, whereas some others were completely absent or onlysporadically present in given types of habitats: the black spotted goby, forexample, was never found in the vegetated habitats, neither sparsely vegetated,nor seagrass beds (Table II). The only species caught with variable but relativelyhigh abundance in each habitat and during each season was the marbled goby(Table II).
TIME AXIS
As shown in Fig. 2, the abundance of most species peaked in summer, whenthe sex ratio was strongly biased towards females. In the black spotted goby,abundance was still relatively high in autumn, whereas in the sand goby noindividuals were found in spring and abundance strongly decreased from sum-mer to autumn. As the season proceeded, the sex ratio tended to become closerto one for some species, like the grass goby and the black goby, but not for thesmall species belonging to the genera Knipowitschia and Pomatoschistus. Non-breeding individuals appeared in most cases in spring with low abundance,peaked during summer and had decreased by autumn.Temporal patterns of IG revealed that for all the species there was a peak in
spring, followed by a strong decrease in summer (Fig. 3). Within this generalpattern, some species, the black goby and the marbled goby, still showed arelatively high IG during summer, indicating a more extended breeding season.The sand goby never showed high IG values, demonstrating the non-reproduc-tive status of this species within the lagoon environment.
HABITAT AXIS
Effects of habitat and abiotic factorsThe two larger species, the black and the grass goby, were more abundant in
seagrass beds than in the rest of the habitats. Differences in the mean abundanceof breeding individuals across habitats were statistically significant during all theseasons for the grass goby (Kruskal–Wallis ANOVA: n ¼ 58 spring, n ¼ 67summer and n ¼ 59 autumn, P < 0�01), but only in spring for the black goby(Kruskal–Wallis ANOVA: n ¼ 58, P < 0�05). When seasonal data were pooled,however, black goby abundance was significantly higher in the seagrass beds andsparsely vegetated habitats than in the other habitats (Kruskal–Wallis ANOVA:n ¼ 184, P < 0�01). Abundance of non-breeding individuals did not significantly
152 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

TABLEII.Mean�
S.D.fish
density
(number
ofindividuals100m�2)ofgobyspeciesin
each
seasonandhabitat
SB
SVh
Sh
MFh
SMc
Total
Zosterisessorophiocephalus
Spring
1�23�
1�23
0�67�
0�98
00�07�
0�16
0�15�
0�34
0�46�
0�87
Summer
11�93�
12�65
3�72�
4�74
1�88�
3�18
4�95�
13�02
2�66�
6�83
5�46�
10�09
Autumn
5�00�
7�88
0�44�
0�57
00�13�
0�50
0�38�
0�86
1�22�
3�97
Gobiusniger
Spring
0�74�
1�25
0�58�
0�67
0�05�
0�11
0�09�
0�29
0�08�
0�18
0�34�
0�72
Summer
0�59�
1�27
0�80�
2�22
0�07�
0�13
0�13�
0�30
0�02�
0�08
0�36�
1�23
Autumn
0�12�
0�22
0�26�
0�53
00�06�
0�22
00�10�
0�30
Gobiuscobitis
Spring
00
00
00
Summer
0�22�
0�60
01�22�
3�33
00
0�19�
1�18
Autumn
00�06�
0�16
0�12�
0�31
00
0�03�
0�13
Knipowitschia
panizzae
Spring
00�16�
0�32
00�80�
1�03
0�83�
1�07
0�40�
0�79
Summer
0�02�
0�07
0�78�
2�30
0�07�
0�13
19�64�
30�72
60�30�
75�82
15�97�
41�11
Autumn
0�05�
0�16
0�79�
1�56
05�78�
11�40
7�80�
14�02
3�12�
8�73
Pomatoschistuscanestrinii
Spring
00
00�31�
0�87
0�03�
0�08
0�09�
0�45
Summer
00
0�03�
0�10
3�32�
5�94
5�60�
15�61
1�85�
7�36
Autumn
00
04�87�
11�58
3�26�
9�81
1�85�
7�33
Pomatoschistusminutus
Spring
00
00
00
Summer
0�02�
0�07
12�61�
17�45
1�18�
2�27
12�48�
26�75
3�08�
6�12
6�69�
16�66
Autumn
0�02�
0�08
0�58�
0�74
0�04�
0�10
2�39�
6�20
0�08�
0�25
0�77�
3�22
Pomatoschistusmarm
oratus
Spring
0�14�
0�35
1�09�
1�55
1�53�
0�67
12�06�
24�70
15�83�
36�02
6�57�
20�53
Summer
0�07�
0�22
27�13�
43�64
39�20�
64�76
62�55�
81�08
31�71�
27�33
32�32�
55�23
Autumn
1�37�
2�15
5�87�
7�64
31�19�
45�40
10�31�
9�56
3�66�
5�59
8�68�
18�28
SB,seagrass
bed;SVh,sparselyvegetatedhabitat;Sh,sandflat;MFh,mudflat;SMc,
saltmarshcreek.
SHALLOW WATER GOBIES OF THE VENICE LAGOON 153
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

differ among habitats for the grass goby (Summer, Kruskal–Wallis ANOVA:n ¼ 67, P > 0�05), whereas that of the black goby was too low to perform anystatistical test.Regarding the small species (Pomatoschistus and Knipowitschia), abundance
was in each case much lower in seagrass beds than in the other habitats
0 0
10
20
30
40
50
60
0
10
20
30
40
50
60
70
80
90
0·0
0·2
0·4
0·6
0·8
1·0
1·2
1
2
3
4
5
6
7Spring Summer Autumn Spring Summer Autumn
Spring Summer Autumn Spring Summer Autumn
Spring Summer Autumn
Season
Spring Summer Autumn
0
2
4
6
8
10(a) (b)
(c) (d)
(e) (f)
Num
ber
of in
divi
dual
s pe
r 10
0 m
3
0
2
4
6
8
10
12
14
FIG. 2. Mean � S.E. abundance of the six goby species: (a) Zosterisessor ophiocephalus, (b) Gobius niger,
(c) Knipowitschia panizzae, (d) Pomatoschistus canestrinii, (e) Pomatoschistus marmoratus and (þ)Pomatoschistus minutus, in spring (n ¼ 58), summer (n ¼ 67) and autumn (n ¼ 59); individuals were
distinguished on the basis of the shape of urogenital papilla in females (&), males ( ) and
indeterminate (&).
154 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

(Table II). The abundance of adult (breeding) lagoon goby was significantlyhigher in salt marsh creeks and mud flats than in the other habitats in eachseason (Kruskall–Wallis ANOVA: n ¼ 58, P < 0�01 in spring, n ¼ 67,P < 0�001 in summer and n ¼ 59, P < 0�001 in autumn). In non-breedingindividuals this pattern was confirmed in summer (Kruskall–Wallis ANOVA:n ¼ 67, P < 0�001) but not in spring, when there was no statistically significantdifference among habitats (Kruskal–Wallis ANOVA: n ¼ 58, P > 0�05). Meanabundance of the sand goby was significantly higher in mud flats and sparsely
Spring Summer Autumn Spring Summer Autumn
Spring Summer Autumn Spring Summer Autumn
Spring Summer Autumn Spring Summer Autumn
0
2
4
6
8
10
12
14(a) (b)
(e) (f)
(c) (d)
0
2
4
6
8
10
12
14
I G
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
Season
FIG. 3. Seasonal mean � S.E. values of the gonado-somatic index of females (&) and males ( ) of the six
goby species: (a) Zosterisessor ophiocephalus (females ¼ 98, males ¼ 59) (b) Gobius niger
(females ¼ 48, males ¼ 24) (c) Knipowitschia panizzae (females ¼ 1082, males ¼ 43) (d)
Pomatoschistus canestrinii (females ¼ 224, males ¼ 106) (e) Pomatoschistus marmoratus
(females ¼ 1358, males ¼ 75) and (f) Pomatoschistus minutus (females ¼ 133, males ¼ 26).
SHALLOW WATER GOBIES OF THE VENICE LAGOON 155
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

vegetated habitats than in the other habitats in each season (Kruskal–WallisANOVA: n ¼ 58, P < 0�001 in summer and n ¼ 59, P < 0�01 in autumn). Theabundance of the marbled goby (breeding individuals) was significantly lower inthe seagrass beds than in the rest of the habitats in each season (Kruskall–WallisANOVA: n ¼ 58 spring, n ¼ 67 summer, P < 0�001 and n ¼ 59, P < 0�01autumn), always peaking in the mud flats. Non-breeding individuals showedthe same patterns (Kruskall–Wallis ANOVA: n ¼ 58, P < 0�01 spring andn ¼ 67, P < 0�001 summer), although the most preferred habitat in summerwas in this case salt marsh creek. The black spotted goby was found only inthe mud flats and salt marsh habitats, so that no comparable statistical test wasperformed.Some correlations between mean abundance and abiotic factors were statisti-
cally significant (Table III), but for most species these relationships were notrecurrent throughout the seasons, with the exception of the black spotted gobyand the lagoon goby. The abundance of black spotted goby was positivelycorrelated with water turbidity, while it was negatively related with salinity,dissolved oxygen and percentage of sand in most seasons (Table III). Lagoongoby abundance was negatively correlated with salinity in each season(Table III).The relative importance of abiotic factors and habitat structure to the seasonal
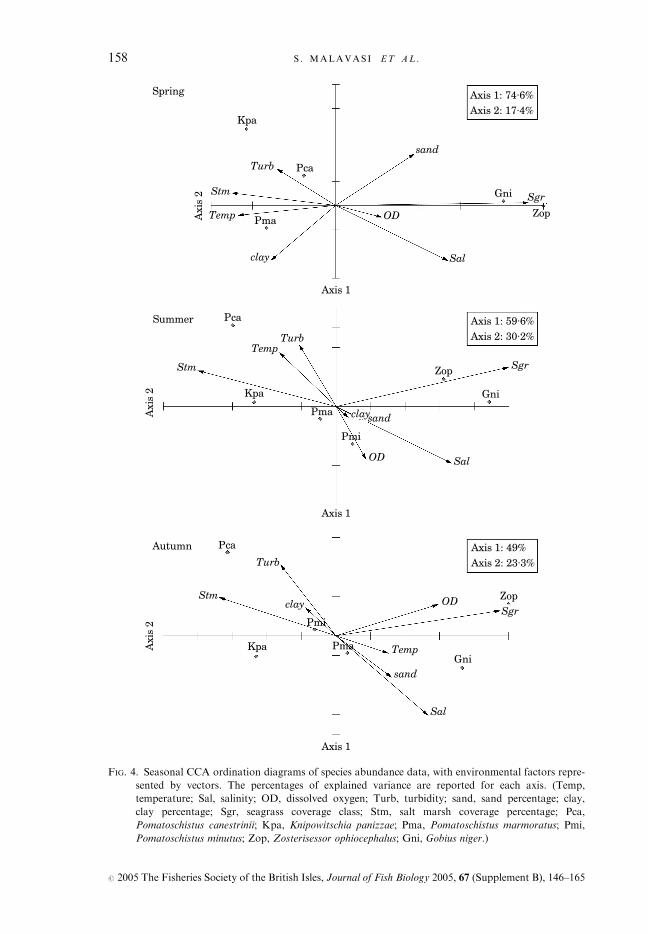
distribution of species abundance is represented in Fig. 4, as determined by CCAanalyses. Each seasonal analysis determined eight canonical axes, whichexplained a percentage of the total species variation of 42–45% in each season.Only axes 1 and 2, however, were plotted as they always accounted for morethan 69% of the variability explained by all canonical axes (Fig. 4).As shown in Fig. 4, two groups of species do not change their relative position
in the diagram throughout the seasons: the group constituted by the blackspotted goby P. canestrinii and the lagoon goby K. panizzae, on the left part ofthe diagram; and the group formed by the black goby G. niger and the grassgoby Z. ophiocephalus, on the opposite side of the diagram. These two groups ofspecies segregate mainly along axis 1, which is positively correlated with theseagrass coverage (the main effect, considering the relative length of the vector)along one direction, affecting the grass and the black goby abundance, and tosalt marsh coverage and turbidity along the opposite direction, positively affect-ing the lagoon goby and the black spotted goby abundance. By contrast, thesand goby and the marbled goby are located around the origin of the axes andtheir position tends to change across seasons. This means that their abundance isnot strongly correlated with any of the considered factors, probably dependingmore on seasonal components.
Interspecific associationsThe w2 test applied to the presence and absence data showed that during
autumn, the spatial distribution of the black goby was significantly and posi-tively associated with that of the grass goby, whereas during summer the occur-rences of the black spotted goby and the marbled goby were both positively andsignificantly associated to that of the lagoon goby. In turn, the distribution ofthe marbled goby was significantly and positively associated with that of thesand goby during summer. By contrast, significant and negative associations in
156 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

TABLEIII.
Seasonalsignificance
levelsoftotalandpartial(controlled
forhabitatfactors)Spearm
ancorrelationsbetweenspeciesabundance
andabioticfactors
Tem
perature
Salinity
Dissolved
Oxygen
Turbidity
Sand
Clay
Total
Partial
Total
Partial
Total
Partial
Total
Partial
Total
Partial
Total
Partial
Spring(n¼
58)
Zosterisessorophiocephalus
NS
NS
þþ
NS
NS
NS
NS
NS
NS
NS
NS
NS
Gobiusniger
NS
NS
þþ
NS
NS
NS
NS
NS
NS
NS
�NS
Knipowitschia
panizzae
NS
NS
���
��
NS
NS
NS
NS
NS
NS
NS
NS
Pomatoschistuscanestrinii
NS
NS
NS
NS
��
�þþ
þþ
�NS
NS
NS
Pomatoschistusmarm
oratusþþþ
NS
��
NS
NS
NS
NS
NS
��
NS
NS
Summer
(n¼
67)
Zosterisessorophiocephalus
NS
NS
NS
NS
NS
��
NS
þþþ
NS
NS
NS
NS
Gobiusniger
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
þKnipowitschia
panizzae
NS
NS
���
NS
NS
NS
NS
NS
NS
NS
NS
NS
Pomatoschistuscanestrinii
þþ
NS
���
����
�þþþ
NS
�NS
NS
NS
Pomatoschitusminutus
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
Pomatoschistusmarm
oratus
NS
NS
�NS
�NS
þNS
NS
NS
NS
NS
Autumn(n¼
59)
Zosterisessorophiocephalus
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
NS
Gobiusniger
NS
NS
NS
NS
NS
NS
��
NS
þNS
NS
NS
Knipowitschia
panizzae
NS
NS
��
NS
���
NS
NS
NS
NS
NS
NS
NS
Pomatoschistuscanestrinii
NS
NS
���
���
NS
NS
þþ
þþþ
�NS
NS
NS
Pomatoschitusminutus
NS
NS
NS
���
NS
NS
þþþþ
NS
NS
NS
NS
Pomatoschistusmarm
oratus
NS
NS
NS
NS
NS
þþ
NS
NS
NS
NS
NS
NS,notsignificant;þ
and�,P<
0�05
;þþ
and��,P<
0�01
;þþþ
and���
P<
0�00
1.
SHALLOW WATER GOBIES OF THE VENICE LAGOON 157
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165
Spring Axis 1: 74·6%
Axis 1
Axi
s 2
Axis 1
Axi
s 2
Axis 1
Axi
s 2
Axis 2: 17·4%
Axis 1: 59·6%Axis 2: 30·2%
Axis 1: 49%Axis 2: 23·3%
Summer
Autumn
Kpa
Kpa
Kpa
Pca
Pca
Pca
Turb
Turb
Turb
Stm
Stm
Stm
Temp
Temp
Temp
Pma
Pma
Pma
Pmi
Pmi
OD
OD
OD
Sal
Sal
Sal
Sgr
Sgr
Sgr
sand
sand
sand
clay
clay
clay
Gni
Gni
Gni
Zop
Zop
Zop
FIG. 4. Seasonal CCA ordination diagrams of species abundance data, with environmental factors repre-
sented by vectors. The percentages of explained variance are reported for each axis. (Temp,
temperature; Sal, salinity; OD, dissolved oxygen; Turb, turbidity; sand, sand percentage; clay,
clay percentage; Sgr, seagrass coverage class; Stm, salt marsh coverage percentage; Pca,
Pomatoschistus canestrinii; Kpa, Knipowitschia panizzae; Pma, Pomatoschistus marmoratus; Pmi,
Pomatoschistus minutus; Zop, Zosterisessor ophiocephalus; Gni, Gobius niger.)
158 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

the spatial distributions were found between the grass goby and the marbledgoby during all the seasons, and between the black goby and the black spottedgoby in the summer.The results of the correlations between species abundance were consistent with
those based on the presence and absence data. The abundance of the black gobywas highly significantly and positively correlated with that of the grass gobyduring all the seasons (spring n ¼ 58, rs ¼ 0�32, P < 0�05; summer n ¼ 67,rs ¼ 0�4, P < 0�001; autumn n ¼ 59, rs ¼ 0�33, P < 0�05), whereas the localabundance of the black spotted goby was positively and significantly relatedwith those of the lagoon goby during summer (n ¼ 67, rs ¼ 0�57, P < 0�001)and autumn (n ¼ 59, rs ¼ 0�27 P < 0�05). During summer, the abundance of themarbled goby was positively and significantly related with that of all the othersmall sized goby species (n ¼ 67, rs ¼ 0�48–0�60, P < 0�001). Abundance of themarbled goby was significantly and negatively correlated with that of the grassgoby during all the seasons (spring n ¼ 58, rs ¼ �0�44, P < 0�001; summern ¼ 67, rs ¼ �0�26, P < 0�05; autumn n ¼ 59, rs ¼ �0�33, P < 0�01), and anegative correlation was found also between the black goby and the blackspotted goby in summer (n ¼ 67, rs ¼ �0�3, P < 0�05).
DISCUSSION
The gobiid assemblage of the Venice Lagoon shallow waters is clearly domi-nated by the smaller sized species belonging to the genera Pomatoschistus andKnipowitschia, and especially by the marbled goby, which alone constitutes>50% of the total catches. A similar seasonal variation in total abundance isrecognizable for most species: colonization by reproductive individuals occursduring spring, whereas recruitment and total abundance peak during summer.The sex ratio tended to be biased towards females in all the species, at leastduring a given season. This is probably due to the sampling technique: drag netscannot easily catch the breeding males, which guard their eggs in nests (undershells or even in burrows in the case of the grass goby), according to Miller(1984). Thus, the temporal variation in the proportion of males in the catch mayindicate the occurrence and the duration of the breeding season. The temporalvariation in both the proportion of males in the population and in the mean IGsuggest that the breeding season of the marbled goby and the lagoon goby lastslonger than that of the black spotted goby, whereas the breeding season of theblack goby is shifted with respect to that of the grass goby. In the sand goby, theabsence of mature individuals during summer and autumn together with theirabsence in spring are consistent with the results of previous studies on themigratory behaviour and the marine spawning of this species, conducted inother Mediterranean environments (Bouchereau & Guelorget, 1997; Pampoulieet al., 1999). These results suggest that there is some degree of temporal segrega-tion (i.e. in timing of reproduction, ‘Time axis’; Tokeshi, 1999) among specieswithin this local goby assemblage.Although most species were ubiquitous, being present in all the habitat types,
contrasting patterns in species abundance among habitats could be detected.These results, together with the analysis of species association and spatial over-lap, suggest a certain level of spatial segregation within the goby assemblage.
SHALLOW WATER GOBIES OF THE VENICE LAGOON 159
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

This is consistent with the results found by other studies on estuarine gobyassemblages suggesting that within these fish guilds, spatial segregation couldplay an important role (Costello, 1992; Wilkins & Myers, 1992; Gill & Potter,1993). In particular, the results demonstrated the spatial association between thegrass and the black goby (the two larger sized species) and their preference forseagrass habitats (although this is more pronounced in the grass goby), whereasthe vegetated habitats seem to be avoided by the smaller species (Pomatoschistusand Knipowitschia), which are mostly associated with mud flats and salt marshcreeks. This suggests a certain degree of spatial segregation between the Gobiusand Zosterisessor group and the Pomatoschistus and Knipowitschia group, simi-lar to that reported by De Casabianca & Kiener (1969) for a Corsican lagoon,and to the results of Wiederholm (1987) for gobies of the Swedish coast. Inparticular, as revealed by the CCA analysis, the group of the larger speciesstrongly segregates from that formed by the black spotted goby and the lagoongoby. The former group is positively affected by the degree of seagrass coverage,while the latter is strongly associated with salt marsh environments and therelated high levels of water turbidity.According to Gill & Potter (1993), patterns of spatial segregation are likely to
reflect the life history characteristics of the species involved. In the case of thegoby assemblage of the Venice Lagoon, it appears that the spatial segregation isconsistent with the two contrasting goby life history features recognized byMiller (1984): the cryptobenthic, long lived, larger species (grass and blackgoby) and the epibenthic, short-lived, smaller species (Pomatoschistus andKnipowitschia). This segregation correlates strongly with body size which,according with Tokeshi (1999), can be considered as ‘one of the factors definingthe niche of a species’. Body size differentiation within a local assemblage ofclosely related species may therefore help in promoting coexistence and mitigat-ing competition. Within Mediterranean gobies, body size differentiation hasprobably arisen from common ancestors under contrasting environmental con-ditions, determining a major split between the Gobius complex and thePomatoschistus complex (Miller et al., 1994; Penzo et al., 1998; Sorice &Caputo, 1999). Once body size differences appear, they may promote speciescoexistence within a local assemblage through resource partitioning (Tokeshi,1999).Variation in habitat preference may originate from both phyletic heritage and
the influence of selective forces related to recent ecological processes, as has beenrecognized for all phenotypic traits (Miles & Dunham, 1993). In the case ofMediterranean gobies, the spatial segregation between the large-sized and thesmall-sized species seems to reflect the phylogeny of the group, with the pre-ference for vegetated and unvegetated habitats coinciding with habitat selectionof the two major clades; that is, the Pomatoschistus and Knipowitschia complex(sand goby group, Miller, 2004) and the Gobius and Zosterisessor complex.Under the phylogenetic constraint related to the splitting of the two major
clades and their main habitat preferences, however, other ecological processesshould act and contribute to the coexistence of species and to their presentdistribution in the lagoon environment.Within the clade of the large species, the above mentioned shift in timing
of breeding may promote the reduction of the potential for interspecific
160 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

competition, which would be expected to be high considering that the black gobyand the grass goby are closely related (Miller et al., 1994; Penzo et al., 1998;Sorice & Caputo, 1999).Within the group of the smaller species, the present distribution could be
determined not only by their intrinsic habitat choice, but also by the competi-tive/predation pressure due to the presence of larger gobies in seagrasses.Seagrasses are traditionally considered to be optimal habitats in terms of
protection and refuge for small fishes (Rooker et al., 1998; Hindell et al.,2000): the results presented here seem to contradict this statement for the smallergobies of the Venice Lagoon, and this contradiction could be explained bydifferent ecological reasons. Some smaller gobies do not show morphologicaladaptations to seagrasses: the marbled and the sand goby, for examples, havedorsal eyes, set very close together (Edlund et al., 1980) and this stronglyindicates an epibenthic adaptation to open substrata. In addition, their coloura-tion is cryptic for a sandy-muddy substratum. By contrast, the lagoon and theblack spotted goby have dorsolateral eyes (Miller, 2004) and a completelydifferent colouration, which potentially could promote survival in structuredenvironments such as seagrasses.Contrasting habitat choice between the field and the laboratory has been
found by Wiederholm (1987) in Pomatoschistus microps (Krøyer), and thiscontrast was considered to be due to the competitive and predatory effects ofthe presence of the black goby in seagrass habitats. Similarly, the high abun-dance of the grass goby in seagrass habitats in the Venice Lagoon may constrainthe distribution of the smaller species. Smaller gobies are sometimes found instomach contents of the grass goby (F. Riccato, pers. obs.) indicating thepotential of predation pressure by this species on the smaller goby species.A clear sub-group constituted by the two species P. canestrinii and K. panizzae
is recognizable, within the small species, in terms of habitat preference andspatial association, as also clearly represented in the CCA diagrams. These twospecies are spatially associated within the Venice Lagoon and this associationcould be partly due to their similar ecological requirements in terms of bothhabitat type and abiotic factors. The analysis of the influence of abiotic factorson goby abundance showed that these two species tend to select internal areaswith lower salinity (8–25) and higher turbidity. A recurrent significant relation-ship between these abiotic factors and abundance is shown especially by theblack spotted goby, which appears to be the most specialized and least abundantgoby in the Venetian assemblage, confirming the results of other authors(Gandolfi et al., 1982). These two species, however, probably strongly reducethe potential for interspecific competition through different nest site choice(Gandolfi et al., 1991), timing of reproduction and feeding tactics (Maccagnaniet al., 1985). By contrast, the other two smaller sized gobies, the sand goby andthe marbled goby, seem to be less associated with a particular habitat type andless affected by the given set of abiotic factors because the distribution of theirabundance is also subjected to some degree of seasonal change, as indicated bythe results of the multivariate ordination. This means that these two species aremore generalist, in terms of habitat selection, than the black spotted goby andthe lagoon goby. In turn, these two species tend to differ slightly in terms ofhabitat choice, with the sand goby being more selective for sparsely vegetated
SHALLOW WATER GOBIES OF THE VENICE LAGOON 161
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

habitats and mud flats than the marbled goby, and to differ strongly in terms ofreproductive and migratory behaviour. The sand goby is a migrant species withmarine spawning and the marbled goby a truly resident species with springreproduction. Considering that these two species are closely related (Milleret al., 1994; Penzo et al., 1998; Sorice & Caputo, 1999), these differences maypromote their coexistence in the lagoon environment. Furthermore, in this casean effect of body size can also be recognized, because the sand goby is signifi-cantly larger than the marbled goby.The higher abundance of the marbled goby, compared with that of the lagoon
and the black spotted goby, could be explained by the high level of tolerance ofthis species in terms of abiotic factors, and the low level of habitat preference. Asshown by Pampoulie et al. (2001), changes in salinity and general environmentalconditions can strongly change the composition of the local goby assemblage. Inthe Venice Lagoon, the oligohaline areas are restricted whereas the areas undermarine influences are relatively extensive, given also the construction of large anddeep canals in the past (Ravera, 2000). This should favour the euryhaline speciesthat prefer salinity >20, like the marbled goby (Gandolfi et al., 1991), whilelimiting the more specialized, oligohaline species like the black spotted goby.Within species of both groups, there is a tendency in some cases for a less
pronounced habitat selection in juvenile fishes (non-breeding individuals) whichis expected due to the dispersal mechanisms of goby postlarval stages (Miller,1984). Reproduction cannot fully explain this pattern because during autumn(outside the breeding season), most species show a habitat preference oravoidance). Thus, reproductive and feeding (nursery) habitats seem not to besegregated within most of these species (apart from the evident exception of thesand goby). In general, most of spatial patterns seem to partly reflect what hasbeen defined by Tokeshi (1999) as the ‘generic spread’ that is the local assem-blages of closely related species are likely to contain species belonging to differ-ent genera, contributing to reduce the potential for competition. This seemsto happen, in the present case, for the two sub-groups of species composed ofK. panizzae and P. canestrinii, in the salt marshes, and G. niger and Z. ophiocephalus,in the seagrasses.These results suggest that both field studies and laboratory experiments are
needed to test hypotheses on habitat selection and inter-specific interactions inorder to better clarify the mechanisms promoting coexistence among these gobyspecies.
The present work was financed by the Co.Ri.La. Project: ‘‘Lo studio della biodiversitaper un equilibrio fra conservazione e sfruttamento in laguna di Venezia’’.
References
Arruda, L. M., Azevedo, J. N. & Neto, A. I. (1993). Abundance, age-structure andgrowth, and reproduction of gobies (Pisces, Gobiidae) in the Ria de AveiroLagoon (Portugal). Estuarine, Coastal and Shelf Science 37, 509–523.
Avanzi, C., Fossato, V., Gatto, P., Rabagliati, R., Rosa Salva, P. & Zitelli, A. (1979).Ripristino, conservazione ed uso dell’ecosistema lagunare veneziano, Venezia:Comune di Venezia.
162 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

Bouchereau, J.-L. & Guelorget, O. (1997). Comparison of three Gobiidae (Teleostei) lifehistory strategies over their geographical range. Oceanologica Acta 21, 503–517.
Costello, M. J. (1992). Abundance and spatial overlap of gobies (Gobiidae) in LoughHyne, Ireland. Environmental Biology of Fishes 33, 239–248.
De Casabianca, M.-L. & Kiener, A. (1969). Niches ecologiques des Gobius des etangssaumatres Corses. Internationale Vereinigung fur Theoretische and AngewandteLimnologie Verhandlungen 17, 665–668.
Edlund, A.-M., Sundmark, G. & Thorman, S. (1980). The identification ofPomatoschistus pictus, P. microps and P. minutus (Gobiidae, Pisces). Sarsia 65,239–242.
Elliott, M. & Dewailly, F. (1995). The structure and components of European estuarinefish assemblages. The Netherlands Journal of Aquatic Ecology 29, 397–417.
Elliott, M. &Hemingway, K. L. (2002). Fishes in Estuaries. Oxford: Blackwell Science Ltd.Fonds, M. (1973). Sand gobies in the Dutch Wadden Sea (Pomatoschistus, Gobiidae,
Pisces). Netherlands Journal of Sea Research 6, 417–478.Gandolfi, G., Torricelli, P. & Cau, A. (1982). Osservazioni sulla biologia del ghiozzetto
cenerino, Pomatoschistus canestrinii (Ninni) (Osteichthyes, Gobiidae). NovaThalassia 5, 97–123.
Gandolfi, G., Zerunian, S., Torricelli, P. & Marconato, A. (1991). Gobiidae. In I Pescidelle Acque Interne Italiane. Parte 3 – Riconoscimento, Biologia, Distribuzione eStatus delle Specie (Gandolfi, G., Zerunian, S., Torricelli, P. & Marconato, A.,eds), pp. 487–553. Roma: Ministero dell’ambiente – Servizio ConservazioneNatura, Unione Zoologica Italiana, Istituto Poligrafico e Zecca dello Stato.
Gill, H. S. & Potter, I. C. (1993). Spatial segregation amongst goby species within anAustralian estuary, with a comparison of the diets and salinity tolerance of the twomost abundant species. Marine Biology 117, 515–526.
Gladfelter, W. B. & Johnson, W. (2004). Feeding niche separation in a guild of tropicalReef fishes (Holocentridae). Ecology 64, 552–563.
Henderson, P. A. (2003). Practical Methods in Ecology. Oxford: Blackwell Publishing.Hindell, J. S., Jenkins, G. P. & Keough, M. J. (2000). Variability in abundance of fishes
associated with seagrass habitats in relation to diets of predatory fishes. MarineBiology 136, 725–737.
Lombarte, A., Recasens, L., Gonzalez, M. & Gild de Sola, L. (2000). Spatial segregationof two species of Mullidae (Mullus surmuletus and M. barbatus) in relation tohabitat. Marine Ecology Progress Series 206, 239–249.
Maccagnani, R., Carrieri, A., Franzoi, P. & Rossi R. (1985). Osservazioni sulla strutturadi popolazione ed il ruolo trofico di tre specie di gobidi (Knipowitschia panizzae,Pomatoschistus marmoratus, Pomatoschistus canestrinii) in un ambiente del Deltadel Po. Nova Thalassia 3, 373–378.
Malavasi, S., Fiorin, R., Franco,A., Franzoi, P., Granzotto, A., Riccato, F. &Mainardi, D.(2004). Fish assemblages of Venice Lagoon shallow waters: an analysis based onspecies, families and functional guilds. Journal of Marine Systems 51, 19–31.
Manly, B. F. J. (1994). Multivariate Statistical Methods. A Primer. 2nd edn. London:Chapman & Hall and CRC Press.
Mathieson, S., Cattrijsse, A., Costa, M. J., Drake, P., Elliott, M., Gardner, J. &Marchand, J. (2000). Fish assemblages of European tidal marshes: a comparisonbased on species, families and functional guilds. Marine Ecology Progress Series204, 225–242.
McKay, S. I. & Miller, P. J. (1991). Isozyme criteria in the testing of phyletic relation-ships between species of Gobius and related eastern Atlantic-Mediterranean genera(Teleostei: Gobiidae). Journal of Fish Biology 39 (Suppl. A), 291–299.
Miles, D. M. & Dunham, A. E. (1993). Historical perspectives in ecology and evolu-tionary biology: the use of phylogenetic comparative analyses. Annual Review ofEcology and Systematics 24, 587–619.
Miller, P. J. (1984). The tokology of gobioid fishes. In Fish Reproduction: Strategies andTactics (Potts, G. W. & Wootton, R. J., eds), pp. 119–153. London: AcademicPress.
SHALLOW WATER GOBIES OF THE VENICE LAGOON 163
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

Miller P. J. (1986). Gobiidae. In Fishes of the North-eastern Atlantic and theMediterranean (Whitehead, P. J. P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J.& Tortonese, E., eds), pp. 1019–1085. Paris: UNESCO.
Miller, P. J. (2004). The Freshwater Fishes of Europe, Vol. 8, part II. Germany: AulaVerlag.
Miller, P. J., Serventi, M., Soregaroli, D., Torricelli, P. & Gandolfi, G. (1994). Isozymegenetics and the phylogeny of Italian freshwater gobies (Teleostei: Gobioidei).Journal of Fish Biology 44, 439–451.
Ministero delle Infrastrutture e dei Trasporti (2003). Attivita di monitoraggio ambientaledella laguna di Venezia. Esecutivo del 1� stralcio triennale (2000–2003) MELA1.Risultati delle attivita di campo e laboratorio delle campagne VII-XXIX, Gennaio-Dicembre 2002 (Attivita A.1.). Venezia: Magistrato alle Acque-tramiteil suo con-cessionario consorzia Venezia Nuova.
Pampoulie, C., Rosecchi, E., Bouchereau, J. L. & Crivelli, A. J. (1999). Life history traitsof Pomatoschistus minutus in the Rhone Delta, France. Journal of Fish Biology 55,892–896. doi: 10.1006/jfbi.1999.1041
Pampoulie, C., Chauvelon, P., Rosecchi, E., Bouchereau, J.-L. & Crivelli, A. J. (2001).Environmental factors influencing the gobiid assemblage of a MediterraneanLagoon: empirical evidence from a long-term study. Hydrobiologia 445, 175–181.
Penzo, E., Gandolfi, G., Bargelloni, L., Colombo, L. & Patarnello, T. (1998). Messiniansalinity crisis and the origin of freshwater lifestyle in Western Mediterraneangobies. Molecular Biology and Evolution 15, 1472–1480.
Plattell, M. E. & Potter, I. C. (2001). Partitioning of food resources amongst 18 abundantbenthic carnivorous fish species in marine waters on the lower coast of Australia.Journal of Experimental Marine Biology and Ecology 261, 31–54.
Ravera, O. (2000). The lagoon of Venice: the result of both natural factors and humaninfluence. Journal of Limnology 59, 19–30.
Rismondo, A., Curiel, D., Scarton, F., Mion, D. & Caniglia, G. (2003). A new seagrassmap for the Venice Lagoon. In Proceedings of the Sixth International Conferenceon the Mediterranean Coastal Environment, MEDCOAST 03, 7–11 October 2003,Ravenna, Italy, Vol. 2 (Ozhan, E., ed.), pp. 843–852. Ankara: Middle EastTechnical University.
Rooker, J. R., Holt, G. J. & Holt, S. A. (1998). Vulnerability of newly settled red drum topredatory fish: is early life survival enhanced by seagrass meadows? MarineBiology 131, 145–151.
Ross, S. T. (1986). Resource partitioning in fish assemblages: a review of field studies.Copeia, 1986, 352–388.
Sacchi, C. R. (1985). Le sel de La Palice: reflexions sur le paralin mediterraneen.Memoires de Biologie Marine et Oceanographique 15, 71–89.
Schoener, T. W. (1974). Resource partitioning in ecological communities. Science 185,27–39.
Sfriso, A., Facca, C. & Ghetti, P. F. (2003). Temporal and spatial changes of macroalgaeand phytoplankton in a Mediterranean coastal area: the Venice lagoon as a casestudy. Marine Environmental Research 56, 617–636.
Sorice, M. & Caputo, V. (1999). Genetic variation in seven goby species (Perciformes:Gobiidae) assessed by electrophoresis and taxonomic inference. Marine Biology134, 327–333.
Syms, C. & Jones, G. (2004). Habitat structure, disturbance and the composition of sand-dwelling goby assemblage in a coral reef lagoon. Marine Ecology Progress Series268, 221–230.
Thorman, S. & Wiederholm A.-M. (1983). Seasonal occurence and food resource use ofan assemblage of nearshore fish species in the Bothnian Sea, Sweden. MarineEcology Progress Series 10, 223–229.
Tokeshi, M. (1999). Species Coexistence. Ecological and Evolutionary Perspectives.Oxford: Blackwell Science.
Van Snik Gray, E. & Stauffer, J. R. (1999). Comparative microhabitat use of ecologicallysimilar benthic fishes. Environmental Biology of Fishes 56, 443–453.
164 S . MALAVASI ET AL.
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165

West, R. J. & King, R. J. (1996). Marine, brackish, and freshwater fish communities inthe vegetated and bare shallows of an Australian coastal river. Estuaries 19, 31–41.
Wiederholm, A.-M. (1987). Habitat selection and interactions between three marine fishspecies (Gobiidae). Oikos 48, 28–32.
Wilkins, H. K. & Myers, A. A. (1992). Shelter utilization by Gobius cruentatus andThorogobius ephippiatus (Teleostei: Gobiidae). Journal of Fish Biology 43, 763–773.
Zander, C. D. (1982). Feeding ecology of littoral gobiid and blennioid fish of the Banyulsarea (Mediterranean Sea). I. Main food and trophic dimension of niche andecotope. Vie et Milieu 32, 1–10
Zander, C. D. & Berg J. (1984). Feeding ecology of littoral gobiid and blennioid fishes ofthe Banyuls area (Mediterranean Sea). II. Prey selection and size preference. Vie etMilieu 34, 149–157.
SHALLOW WATER GOBIES OF THE VENICE LAGOON 165
# 2005 The Fisheries Society of the British Isles, Journal of Fish Biology 2005, 67 (Supplement B), 146–165




















