
The role of proprioception in the control of prehension movements: a kinematic study in a...
18
Exp Brain Res (1994) 99:483-500 Springer-Verlag 1994 M. Gentilucci I. Toni S. Chieffi G. Pavesi The role of proprioception in the control of prehension movements: a kinematic study in a peripherally deafferented patient and in normal subjects Received: 16 June 1993 / Accepted: 8 December 1993 Abstract In this study we investigated the role of pro- prioception in the control of prehension movements, with particular reference to the grasp component. Grasp and transport kinematics were studied in a peripherally deafferented patient and in five healthy subjects. Two experiments were carried out: the prehension experi- ment and the grasp perturbation experiment. In the pre- hension experiment both the patient and the control subjects were required to reach and grasp three objects of different size, located at three different distances, both with and without visual feedback. In the grasp perturba- tion experiment a mechanical perturbation was applied to the fingers during prehension movements, again exe- cuted with and without visual feedback. In the prehen- sion experiment temporal parameters of the patient's movements were generally slowed, with greater vari- ability on some measures. However, over the first phase of the movement the pattern of the patient's hand open- ing and transport acceleration, scaled to object size and distance, was the same as that of controls, both with and without visual feedback. On the contrary, during the final phase of the movement (the finger closure phase and deceleration) the patient's performance differed sig- nificantly from the controls. These phases were abnor- mally lengthened and frequent movement adjustments were observed. In the grasp perturbation experiment the patient was not able to compensate for the perturba- tions applied to the fingers, even with visual feedback. The data allowed us to investigate also the respective contribution of proprioception and of vision of the hand in the control of prehension. We compared prehension kinematics in two conditions: (a) with visual but no pro- M. Gentilucci ([N~I)- I. Toni - S. Chieffi Institute of Human Physiology, University of Parma, via Gramsci 14, 1-43100 Parma, Italy G. Pavesi Institute of Neurology, University of Parma, via del Quartiere 4, 1-43100 Parma, Italy prioceptive feedback (in the patient) and (b) with propri- oceptive but no visual feedback (in the controls). In both experiments proprioceptive control was more efficient than visual control. The results of this study are inter- preted in favour of the strict dependence of prehension control on proprioception. The first phase of the move- ment, however, can be appropriately planned and exe- cuted without the necessity of either proprioceptive or visual information about the hand. Key words Prehension Grasp Transport Proprioception Deafferented human Introduction It is known that loss or impairment of somaesthetic sen- sation produces severe disorders in motor control. However, the exact nature of the impairment is a matter of debate. Mott and Sherrington (1895) observed that monkeys failed to use their forelimbs after deafferenta- tion by section of the dorsal roots. They therefore pro- posed that somaesthetic information is necessary for movement initiation. This suggestion was questioned by Lashley (1917), who reported the case of a patient with a deafferented leg, following a gunshot injury of the spinal cord. The patient could extend and flex his knee to produce different amplitudes and velocities of leg movement without visual control. His impairment was only in compensating for mechanical perturbation of the moving leg. The problem of whether somaesthetic information is necessary for movement planning has been addressed more recently by several experimenters. It has been found, for example, that deafferented monkeys can reach for objects, provided that they have sufficient postoperative time to learn how to use their deafferent- ed limbs (Taub et al. 1975). Deafferented monkeys are also able to compensate for mechanical perturbations applied to the forelimb during extension-flexion move- ments executed without visual feedback (Polit and Bizzi
Transcript of The role of proprioception in the control of prehension movements: a kinematic study in a...

Exp Brain Res (1994) 99:483-500 �9 Springer-Verlag 1994
M. Gentilucci �9 I. Toni �9 S. Chieffi �9 G. Pavesi
The role of proprioception in the control of prehension movements: a kinematic study in a peripherally deafferented patient and in normal subjects
Received: 16 June 1993 / Accepted: 8 December 1993
Abstract In this study we investigated the role of pro- prioception in the control of prehension movements, with particular reference to the grasp component. Grasp and transport kinematics were studied in a peripherally deafferented patient and in five healthy subjects. Two experiments were carried out: the prehension experi- ment and the grasp perturbation experiment. In the pre- hension experiment both the patient and the control subjects were required to reach and grasp three objects of different size, located at three different distances, both with and without visual feedback. In the grasp perturba- tion experiment a mechanical perturbation was applied to the fingers during prehension movements, again exe- cuted with and without visual feedback. In the prehen- sion experiment temporal parameters of the patient's movements were generally slowed, with greater vari- ability on some measures. However, over the first phase of the movement the pattern of the patient's hand open- ing and transport acceleration, scaled to object size and distance, was the same as that of controls, both with and without visual feedback. On the contrary, during the final phase of the movement (the finger closure phase and deceleration) the patient's performance differed sig- nificantly from the controls. These phases were abnor- mally lengthened and frequent movement adjustments were observed. In the grasp perturbation experiment the patient was not able to compensate for the perturba- tions applied to the fingers, even with visual feedback. The data allowed us to investigate also the respective contribution of proprioception and of vision of the hand in the control of prehension. We compared prehension kinematics in two conditions: (a) with visual but no pro-
M. Gentilucci ([N~I) - I. Toni - S. Chieffi Institute of Human Physiology, University of Parma, via Gramsci 14, 1-43100 Parma, Italy
G. Pavesi Institute of Neurology, University of Parma, via del Quartiere 4, 1-43100 Parma, Italy
prioceptive feedback (in the patient) and (b) with propri- oceptive but no visual feedback (in the controls). In both experiments proprioceptive control was more efficient than visual control. The results of this study are inter- preted in favour of the strict dependence of prehension control on proprioception. The first phase of the move- ment, however, can be appropriately planned and exe- cuted without the necessity of either proprioceptive or visual information about the hand.
Key words Prehens ion �9 G r a s p �9 Transport Proprioception �9 Deafferented human
Introduction
It is known that loss or impairment of somaesthetic sen- sation produces severe disorders in motor control. However, the exact nature of the impairment is a matter of debate. Mott and Sherrington (1895) observed that monkeys failed to use their forelimbs after deafferenta- tion by section of the dorsal roots. They therefore pro- posed that somaesthetic information is necessary for movement initiation. This suggestion was questioned by Lashley (1917), who reported the case of a patient with a deafferented leg, following a gunshot injury of the spinal cord. The patient could extend and flex his knee to produce different amplitudes and velocities of leg movement without visual control. His impairment was only in compensating for mechanical perturbation of the moving leg.
The problem of whether somaesthetic information is necessary for movement planning has been addressed more recently by several experimenters. It has been found, for example, that deafferented monkeys can reach for objects, provided that they have sufficient postoperative time to learn how to use their deafferent- ed limbs (Taub et al. 1975). Deafferented monkeys are also able to compensate for mechanical perturbations applied to the forelimb during extension-flexion move- ments executed without visual feedback (Polit and Bizzi

484
1979). However, if the canonical position of the arm was changed, monkeys were no longer able to correct the effects of the perturbations. Polit and Bizzi (1979) proposed that either visual or proprioceptive knowl- edge of the body schema is necessary for movement planning.
The capacity to produce complex patterns and se- quences of finger movements without any feedback was observed in the deafferented patient described by Roth- well et al. (1982). However, the patient's performance became degraded if visual feedback was removed for several seconds. This result indicates that the motor program needs to be refreshed by either visual or ki- naesthetic information about its execution in order to remain efficient. Sanes et al. (1985) found that deaffer- ented patients were able to initiate flexion-extension arm movements of different amplitudes without visual feedback, but that the trajectories and the end points of the movement were inaccurate. Occasionally the pa- tients also moved initially in an incorrect direction.
These experiments were for the most part concerned with simple movements. The role of kinaesthetic infor- mation may be fundamental in the control of multijoint movements towards an external visual object, since with such movements the nervous system has to translate visual information about extrinsic object properties (spatial object position referred to the viewer) and/or intrinsic object properties (object size and shape) into a set of commands and controls specified in terms of mul- tijoint angular displacements (Soechting and Flanders 1989). Errors in movement direction and amplitude were observed in deafferented patients without visual feedback pointing to targets placed at different locations (Ghez et al. 1990). The errors in movement amplitude resulted primarily from overshooting the target and were present for those movement directions in which limb anisotropy was lower. The patients were not able to compensate for the greater peak accelerations during these movements. However, their performance im- proved if they could see their hands before the move- ment or if they had seen their hand trajectories on previ- ous trial. Consequently, the deficit was interpreted as being due to incorrect motor programming, rather than to inefficient control during the movement (Ghez et al. 1990).
The question of whether or not complex movements towards a visual target can be fully planned without any proprioceptive information has not been completely re- solved. In particular, it is not clear which kinematic parameters can still be accurately programmed after deafferentation. In the patients studied by Ghez et al. (1990) peak velocity and peak acceleration were scaled as a function of distance. However, the temporal pattern of the velocity and acceleration profiles was not report- ed. Secondly, complex movements executed in three-di- mensional (3D) space require more refined coordination among the several segments of the arm than do move- ments executed in the plane, such as those analysed by Ghez et al. (1990). Consequently, the time course of
these more complex movements may still depend on kinaesthetic information. Finally, the finding by Ghez et al. (1990) that peak velocity and peak acceleration were both scaled as a function of distance could have been due to the residual capacity of their patients to detect movements of their elbows and shoulders.
Prehension is a more complex movement than point- ing. It is composed of three contemporaneous motor components: transport, rotation and grasp (Arbib 1981). Kinematic studies (Jeannerod 1984) have shown that the transport and grasp components are planned by independent "visuo-motor channels". The visuo-mo- tor channel controlling transport computes extrinsic object properties, while the grasp channel computes in- trinsic object properties. The mechanism of coordina- tion between the two components is still a matter of debate (Jeannerod 1984; Jakobson and Goodale 1991; Paulignan et al. 1991 a,b; Gentilucci et al. 1992; Chieffi and Gentilucci 1993). Kinaesthesia may be necessary to select the appropriate parameters of the two compo- nents and to control their coordinated execution. A pre- vious study (Jeannerod et al. 1984) showed that the loss of somaesthesia produced severe impairments in both the grasp and the transport components. However, the anaesthesia in this case resulted from a cortical lesion located in the left parietal lobe. Besides the primary and secondary somatosensory cortices, the lesion probably included posteriorly area 5 and a large part of area 40. In addition, other cortical circuits may have been dis- connected from the strictly sensory areas. Thus, the deficit seen in this parietal patient probably resulted from an inability to transform information into motor commands rather than from the loss of somaesthetic information per se. Moreover, the motor impairments observed in the patient could have been due, as was pointed out by the authors (Jeannerod et al. 1984), to an exaggerated co-contraction of the agonistic and antago- nistic muscles. Muscle co-contraction seems to be a common finding in deafferented patients (see Luria 1973, Fig. 44; Rothwell et al. 1982).
In the present study the role of proprioception in the control of prehension movements was addressed in a peripherally deafferented patient. Among the different components of prehension, we were interested primarily in studying the grasp component (finger aperture and closure during reaching). This behaviour appears later during development than does reaching (Von Hofsten and Ronnqvist 1988). Electrophysiological studies on monkeys (Rizzolatti et al. 1988) have shown that 40% of premotor neurons that respond when an animal grasps an object during specific types of grasping are also acti- vated by somaesthetic stimulation of the fingers. Conse- quently, the planning and the execution of grasping movements may be dependent on proprioception. By using an experimental paradigm which permitted the exclusion of visual feedback with our deafferented pa- tient we aimed to establish which kinematic parameters of prehension can be correctly planned without the ben- efit of either visual or proprioceptive input. The second

aim of the experiment was to investigate the respective contributions made by vision of the hand and by propri- oception to the control of prehension. To this purpose, the kinematics of movements executed by the patient in the visual feedback condition were analysed and com- pared with those of the control subjects with and with- out visual feedback. An additional experiment was car- ried out in order to compare adjustment of grasp in response to a mechanical perturbation applied to the fingers, observed in both the patient and the control subjects (the grasp perturbation experiment).
Materials and methods Subjects
One patient (M.B., aged 27) and five neurologically normal sub- jects (four women and one man, aged 27-30) participated in the experament. The controls and the patient were right-handed.
The patient had developed severe, large-fibre sensory neuropa- thy, following primary Sj6gren's syndrome. Neuropathy affected both arms and lower extremities. She had complete loss of tactile sensation in both hands and forearms and of proprioceptive sen- sation in the fingers and wrist of both arms. She could detect movements of elbows and shoulders only at the extremes of range. Sensitivity to vibration at fingers, wrist and elbow was lost. Tem- perature and pain sensation were affected distally. Deep tendon reflexes were absent in all muscles of the upper extremities, but muscle strength was normal at clinical examination.
In everyday life the patient had great difficulty in using her hands with dexterity for eating, drinking and writing, especially without visual feedback. She was not able to use her hands to hold a glass, a pen or a cigarette for a long period of time. She had to look at them continuously, otherwise she dropped them. She was no longer able to ride her bicycle. However, individual digits could be activated independently and she could even touch her thumb with each finger in turn or outline figures of different shape or size in the air using her digits and wrist, also without visual feedback (for similar observations see Rothwell et al. 1982). How- ever, her performance deteriorated when she repeated these tasks over a longer period of time (more than 20 s).
Cervical and cortical evoked potentials were absent on stimu- lation of both median and ulnar nerves at wrist and elbow. They were absent on stimulation of the median nerves of the first, sec- ond and third finger and on stimulation of the ulnar nerve of the fifth finger. In Fig. 1 the results of median nerve stimulation at the elbow are shown. This figure was selected in order to give more global information on the loss of forearm somaesthesia. The sen- sory nerve action potentials were absent at the wrist and elbow of both arms. Electromyographic activity (EMG) recorded by con- centric needle electrodes of arm proximal and distal muscles was normal. The pattern of muscle contraction during fast index finger movements was characterised by the typical triphasic pattern of activity in agonistic and antagonistic muscles.
The patient participated in this study 2 years after the first symptoms of somaesthesia loss. She was tested in six experimental sessions. Each control subject was tested in two sessions. The interval between each session was 1 week. The patient and the control subjects were naive as to the purpose of the experiment.
Apparatus and procedure
The subjects sat in a dark room in front of a box which was open on the upper side and divided horizontally by a semireflecting mirror into two compartments. They positioned their head in a head-and-chin rest and looked at the stimuli through the mirror from the upper compartment. Their right hand rested below the
~ -Erb [ 5 as ~J 2 uV I
C7 - A ~ ? -Ai
485
C4' "Ca'
t _ C3 -Erb
Ca' -C4'
LEFT RIGHt
Fig. ! Patient's somatosensory evoked potentials during stimfila- tion of the median nerve at the elbow by surface electrodes. Cur- rent of the electrical stimulus was 15 mA, duration of the stimulus was 0.2 ms. A current well above motor threshold was selected in order to stimulate also small sensory fibres. Note the absence of any evoked response at the cervical spinal level (second trace from the top) and at the cortical level (third and fourth traces). The presence of a nerve action potential at the Erb point (top trace) was due to antidromic activation of motor fibres. This potential was absent after stimulation of the elbow median nerve at motor threshold. Similar results were obtained after stimulation of the digital nerves of the first, second and third fingers by ring elec- trodes (stimulus current, 25 mA, that is, 5 times the subjective sensory threshold). Each trace is the mean of the responses to 1000 stimulations
mirror on the base of the lower compartment (height from the floor, 75 cm) at a fixed position (hand starting position) along the subject's sagittal axis. The hand was in line with the forearm. The thumb and index fingers were held in pinch position and were distant 20 cm from the subjects' chest.
The objects to be reached to and grasped were translucent spheres that could have three sizes (3 cm diameter, small object; 4 cm, medium object; 5 cm, large object). One of the three objects was placed on the base of the lower compartment and was pre- sented along the subjects' sagittal plane at one of three locations, respectively 15 cm (near object position), 25 cm (central object position) and 35 cm (far object position) from the tips of the thumb and index fingers. Another translucent sphere of the same size, located at exactly the same distance from the tips of the thumb and index fingers with respect to the subject's sagittal axis, was fixed on a support placed in the upper compartment. The height of the support from the mirror was equal to that of the mirror from the plane of the lower compartment. Two lasers, one in the upper and one in the lower compartment were used to illuminate the upper or lower object, respectively. When the lower compartment was illuminated the object was seen through the mirror. When the lower compartment was darkened, illumination of the upper object produced a virtual image of the target object on the semireflecting mirror. Two experiments were carried out on the patient and on the controls: the prehension experiment and the grasp perturbation experiment.
Prehension experiment
Prehension movements were studied in two experimental condi- tions: visual feedback and no visual feedback. In the visual feed- back condition the lower compartment was illuminated and the subjects saw the target object, their hand and the base of the lower compartment both before and during the prehension movement.

486
In the no visual feedback condition the lower compartment was darkened before the trial and the subjects saw only the virtual image of the object.
In this experiment the patient was tested in five sessions and each control subject was tested in one session. In each session two blocks of trials were run, one in the visual feedback condition, the other in the no visual feedback condition. The order of presenta- tion of the two blocks was counterbalanced across sessions for the patient and across subjects for the controls. Prehension of the three objects located at the three distances (nine conditions) was studied both with and without visual feedback. Seven trials for each condition were presented to the subjects in randomised or- der.
Grasp perturbation experiment
In a separate session the patient and the control subjects were tested in the grasp perturbation experiment. In this experiment a spring could be attached to the second phalanx of the thumb and index fingers before the trial. The spring (stiffness 58.8 g/cm) start- ed to develop force when the finger aperture, measured on the palmar surface of digits during grasping, reached a value of ap- proximately 5.0 cm. The stimuli were the same as those of the prehension experiment. They were located at a distance of 20 cm from the starting position (central position). The perturbation was applied only during trials with the large object (diameter 5 cm). There were four conditions: unperturbed grasp of the small, medi- um, and large object, and perturbed grasp of the large object. Ten trials for each condition were randomly presented. Before each unperturbed trial a broken spring was placed between the sub- jects' fingers. By using this procedure the subjects did not know whether the perturbation would be applied during the trial. Pre- hension movements were studied in both visual feedback and no visual feedback conditions. Only the prehension movements di- rected to the large object were analysed for both unperturbed and perturbed trials.
In both experiments the subjects closed their eyes during the intertrial interval and opened them on the command "open", said by the experimenter. For both experiments, in the no visual feed- back condition the lower compartment remained illuminated for approximately 4 s. It was then darkened for 3 s before a tone, delivered by a PC, signalled the beginning of the trial. In the grasp perturbation experiment, when the lower compartment was illu- minated, the subjects could see only the tips of the thumb and index fingers, but not the spring. In both experiments the subjects were required to grasp the object by using the thumb and the index finger and to bring the object to the starting position. The instruction was to use a natural speed of movement, as in sponta- neous reaching.
Movement recording and data analysis
The movements of arm and hand were recorded using a 3D com- puterised movement recording system (ELITE system). Move- ment reconstruction and computation of the kinematic parame- ters are described elsewhere (Gentilucci et al. 1992).
In this study five markers were used. The first marker was placed on the styloid process of the radius, at the wrist; the second and the third markers were placed on the tips of the index finger and thumb, respectively; the fourth marker was placed on the object. The fifth marker was placed on the plane of the table along the subject's sagittal axis, 5 cm from the chest, in order to provide a reference point.
The markers on the subjects' wrists were used to analyse the transport component. The transport kinematic parameters mea- sured in this study were: transport time, peak velocity, time to peak velocity, peak deceleration, time to peak deceleration. Vari- ability of wrist trajectory was also calculated after normalisation in time (40 frames) of the wrist displacements along the subjects' sagittal, vertical and transverse axes. The standard deviation of
wrist positions along the three axes was taken into account for each frame, and the square root of the sum of their squares was calculated. Maximal variability of the wrist trajectory and vari- ability of the final wrist position were analysed in this study. The grasp component was studied by analysing the time course of the distance between the thumb and index finger. The grasp kinematic parameters measured in this study were: grasp time, maximal finger aperture, time to maximal finger aperture, variability of maximal finger aperture, peak velocity of finger opening, time to peak velocity of finger opening.
The mean distance between the two markers placed on the thumb and index fingers was calculated when the fingers were in pinch position in five separate trials at the end of each session. This value was subtracted from the grasp values recorded in that session, in order to obtain the true grasp parameters, that is, the time course of the distance between the palmar surfaces of the fingers.
Since the spatial error of the ELITE system, obtained by mea- suring the maximal variations of the recorded distance between two moving stimuli that have a fixed distance between them, is 0.4 mm, the transport and the grasp components were considered to start and stop in those frames in which the marker displace- ment was, respectively, greater and less than 0.4 mm. The frame corresponding to the beginning of the movement was selected only after verifying that the marker displacement increased in each of the 13 forthcoming frames. The frame corresponding to the end of the movement was chosen only after verifying that the marker displacement did not vary and/or reversed direction in the 10 successive frames.
Results
P r e h e n s i o n e x p e r i m e n t
Grasp component
Control subjects. R e p r e s e n t a t i v e e x a m p l e s of g r a s p m o v e m e n t s d i r e c t e d to the th ree ob jec t s p o s i t i o n e d at the th ree d i s t ances e x a m i n e d in th is s t u d y are s h o w n in Fig. 2. In the left c o l u m n m o v e m e n t s in the v isua l feed- b a c k c o n d i t i o n are s h o w n a n d in the r ight c o l u m n m o v e m e n t s in the no v i sua l f e e d b a c k c o n d i t i o n a re p re - sented. N o t e t ha t m a x i m a l f inger a p e r t u r e i nc r ea sed as a func t ion of ob j ec t size. M o r e o v e r , i t was l a rge r in the c o n d i t i o n of no v isua l feedback . T h e k i n e m a t i c p a r a m e - ters a v e r a g e d ac ross the five sub jec t s a re p r e s e n t e d in Table 1. T h e y were s u b m i t t e d to A N O V A s w h o s e m a i n fac tors were : c o n d i t i o n (visual f e e d b a c k vs no v isua l feedback) , ob jec t p o s i t i o n (near vs cen t ra l vs far ob jec t pos i t ion) , ob jec t size (smal l vs m e d i u m vs la rge object) . A s igni f icance c r i t e r ion of P < 0 . 0 1 was chosen in o r d e r to r educe the poss ib i l i t y of t ype I e r ro r s a s s o c i a t e d wi th the m a n y u n i v a r i a t e A N O V A s wh ich were ca r r i ed out. N e w m a n - K e u l s tes t was used as a p o s t h o c tes t in al l A N O V A s p e r f o r m e d in this s tudy.
Grasp time was s ign i f ican t ly l o n g e r in the no v isua l f e e d b a c k (649.1 ms) t h a n in the v isua l f e e d b a c k cond i - t i on (597.8 ms ; El, 4 = 147.7, P < 0.01). I t was s ign i f ican t ly affected a lso by ob jec t p o s i t i o n (f2, 8 = 257.2, P < 0.001 : n e a r ob jec t pos i t i on , 570.0 ms ; cen t ra l ob j ec t pos i t i on , 616.0 ms ; far ob jec t pos i t i on , 686.0 ms).
Maximal finger aperture i nc r ea sed wi th ob jec t size (F2,8= 1874.9, P < 0 . 0 0 0 1 : smal l object , 58.6 m m ; m e d i -

Fig. 2 Representative exam- ples of the time course of the grasp component recorded in one control subject (F.O.). The left column illustrates move- ments executed in the visual feedback condition (VF); the right column illustrates move- ments executed in the no visu- al feedback condition (NVF). In the top row movements di- rected to the small stimulus (S) are represented. In the mid- dle and bottom rows, respec- tively, movements directed to the medium (M) and large object (L) are represented. In each panel the curves N, C, and F show movements di- rected to the near, central and far object, respectively
I 0 0
80
6O
4O
2 0
VF
S
2 5 0 5 0 0 7 5 0
0 0
lOO F M
8O
40 ~ F I
20t~,/ r 2 5 0 5 0 0 7 5 0
I00
8O
6O
4 0
20
0 0
L
C
. / I I 2 5 0 5 0 0 7 5 0 m~
um object, 66.6 mm; large object, 74.4mm) and was greater in the no visual feedback condition, al though only a trend to significance was found (F,,4=13.9, P = 0 . 0 2 : 63 .3mm vs 69.7mm). Variability of maxi- mal finger aperture was not influenced by any of the three factors (F1.4=0.92, P = 0 . 3 9 ; F2,s=0.82, P = 0 . 4 8 ; F2,s=l.24, P=0.34) . Time to maximal finger aper- ture increased significantly with increasing distance (F2,g = 122.2, P <0.01:344.0 ms vs 380.0 ms vs 423.8 ms).
Peak velocity of finger opening was significantly af- fected by object size (F2, 8 = 235.7, P < 0.0001: small ob- ject, 302.5 ram/s; medium object, 337.4 ram/s; large ob- ject, 377.9 mm/s). The effect of condit ion approached s i g n i f i c a n c e ( E l , 4 ~ - - 1 5 . 7 , P = 0.016: visual feedback con-
487
1000 1250
10o - NVF
i s 8 0 -
~176 F 4 0
2 0 F
0 2 5 0 5 0 0 7 5 0 1000 1250
I000 1250
I00
80.
4o ~.. F
2oIs 0 r I ' F
0 2 5 0 5 0 0 7 5 0 1000 1250
I00
80
60
40
20
i J 0 i
I000 1250 0 I000 1250
L
250 500 750 m s
dition, 323.4mm/s; no visual feedback condition, 355.1 mm/s). Time to peak velocity of finger opening was only influenced by object distance (F2,8 = 11.0, P < 0.005: near object position, 187.6 ms; central object position, 210.7 ms; far object position, 226.1 ms).
The patient. Figure 3 shows representative examples of grasp movements in the visual feedback and no visual feedback conditions. As one can see, maximal finger aperture increased as a function of object size, including when the grasp was executed without visual feedback. The finger opening phase was generally similar to the controls (compare Fig. 2), al though the movement time was greater and occasionally bumps in the trajectory of

488
0
0
0
8
0
8 t~
0
0
0
o
~6
o
0
~.~ "~ m
~ 5z
.-=N
~ g
Z
o O ~ ~ ~ . eq..~-
oo~.. oo ~. ~ .
~ , o ~
o o ~
~ , oo oq.
�9 e q ~ - , q . c ~ ~ . ~ ,
~ ~ ~.~
O ~
finger opening were observed in the visual feedback condition. In contrast, the finger closure phase differed from that of the controls in the no visual feedback con- dition. It was characterised by frequent movement ad- justments. Note also the greater variability in the final distance between the tips of the thumb and index fin- gers. The object was not grasped with the tip, but with the second phalanx of the index finger. Sometimes the object slipped from the fingers at the end of grasping. Table 2 shows the grasp kinematics parameters aver- aged across the mean values obtained in the five experi- mental sessions. These data were submitted to ANOVAs whose factors were: condition, object posi- tion and object size, as for the controls.
Grasp time was significantly longer in the no visual feedback condition (1144.0ms) than in the visual feedback condition (936.0ms) (F1,4=33.0, P<0.005). Grasp time also increased with distance (F2,8=32.7, P<0.001:926.0 ms vs 1036.0 ms vs 1158.0 ms).
Maximal finger aperture increased with object size (F2, 8 = 234.9, P < 0.0001 : small object, 58.4 mm; medium object, 71.5 mm; large object, 79.0 mm) and was greater in the no visual feedback condition (F1,4=32.7, P < 0.005: 75.7 mm vs 63.7 mm). These two effects were similar to the controls. The effect of object position was also significant overall (F2, 8 = 9.6, P < 0.01). However, the difference was minimal and was significant only for the near position with respect to the other two positions (P<0.05: near position, 68.7mm; central position, 70.3 mm; far position, 70.1 mm). Variability of maximal finger aperture was not influenced by any of the three factors (F1,4 = 5.6, P =0.08; F2, 8 =5.6, P = 0.03; F2,8= 1.1, P=0.39). Time to maximal finger aperture was signifi- cantly affected only by object position (F2.8=39.1, P<0.0001: near position, 488.0ms; central position, 566.0 ms; far position, 656.0 ms).
Peak velocity of finger opening increased with object size (F1,4=96.4, P<0.0001:238.9 mm/s vs 275.7 mm/s vs 313.5 mm/s). It was not significantly influenced by condition (F1,4 = 1.7, P=0.27) although the mean peak velocity was slightly higher in the no visual feedback (285.0 mm/s) than in the visual feedback condition (267.4 mm/s). Time to peak velocity of finger opening in- creased significantly with object distance (F2,8= 10.3, P<0.01:244.1 ms vs 261.4 ms vs 277.3 ms).
Comparison between the grasp kinematics of the patient and the controls. Temporal parameters of the patient increased with respect to those of the controls. Mean grasp time of the patient was 1040.2 ms, whereas that of the controls was 623.2 ms. The mean value for the pa- tient lay outside the 99% t-confidence interval calculat- ed for the mean of the controls (623.2_+ 164.0ms). In addition, the mean grasp time of the patient in the visual feedback condition (935.3 ms) was greater than the mean grasp time of the controls in the no visual feed- back condition (649.0__ 166.0 ms). Further, the mean time to maximalfinger aperture of the patient (570.2 ms) lay outside the t-confidence interval estimated for the
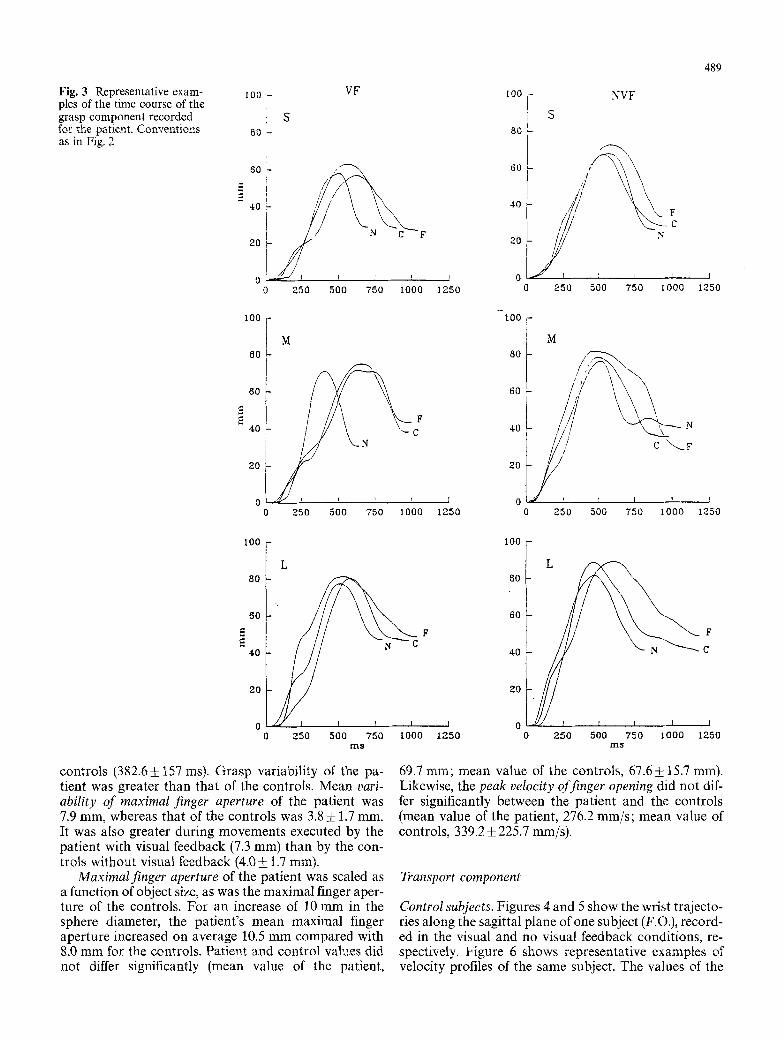
Fig. 3 Representative exam- 100 - VF pies of the time course of the grasp component recorded : S for the patient. Conventions t0 - as in Fig. 2
6 0 - i ,4
,oL N F
2O / / /
0 2 5 0 5 0 0 7 5 0 i000
1 ~ 1 7 6 M 80 L
4 0
20
0 J ' ! J 0 2 5 0 5 0 0 7 5 0 1000
i00
80
6O
4O
20
L
t~ C
2 5 0 5 0 0 "750 1 0 0 0 m s
controls (382.6!157 ms). Grasp variability of the pa- tient was greater than that of the controls, Mean vari- ability of maximal finger aperture of the patient was 7.9 mm, whereas that of the controls was 3.8 • 1.7 ram. It was also greater during movements executed by the patient with visual feedback (7.3 mm) than by the con- trols without visuM feedback (4.0 +_ 1.7 ram).
Maximal finger aperture of the patient was scaled as a function of object size, as was the maximal finger aper- ture of the controls. For an increase of 10 mm in the sphere diameter, the patient's mean maximal finger aperture increased on average 10.5 mm compared with 8.0 mm for the controls. Patient and control values did not differ significantly (mean value of the patient,
4 8 9
lO0
80
60
40
20
I 0
1 2 5 0
NVF
S
\,
t
c
!
~50 500 7 5 0 [000 T
1250
tOO
80
60
40
20
I 0 1 2 5 0
I M
jk /// \ I /I \
/ / c ~_r
1
0 2 5 0 500 7 5 0 100O
!
1250
i00
80
60
40
30
I 0 1 2 5 0 0
L
F
C
2 5 0 5 0 0 7 5 0 1 0 0 0 1250 m s
69,7 mrn; mean value of the controls, 67.6! 15,7 mm). Likewise, the peak velocity of finger opening did not dif- fer significantly between the patient and the controls (mean value of the patient, 276.2 mm/s; mean value of controls, 339,2_+ 225.7 mm/s).
Transport component
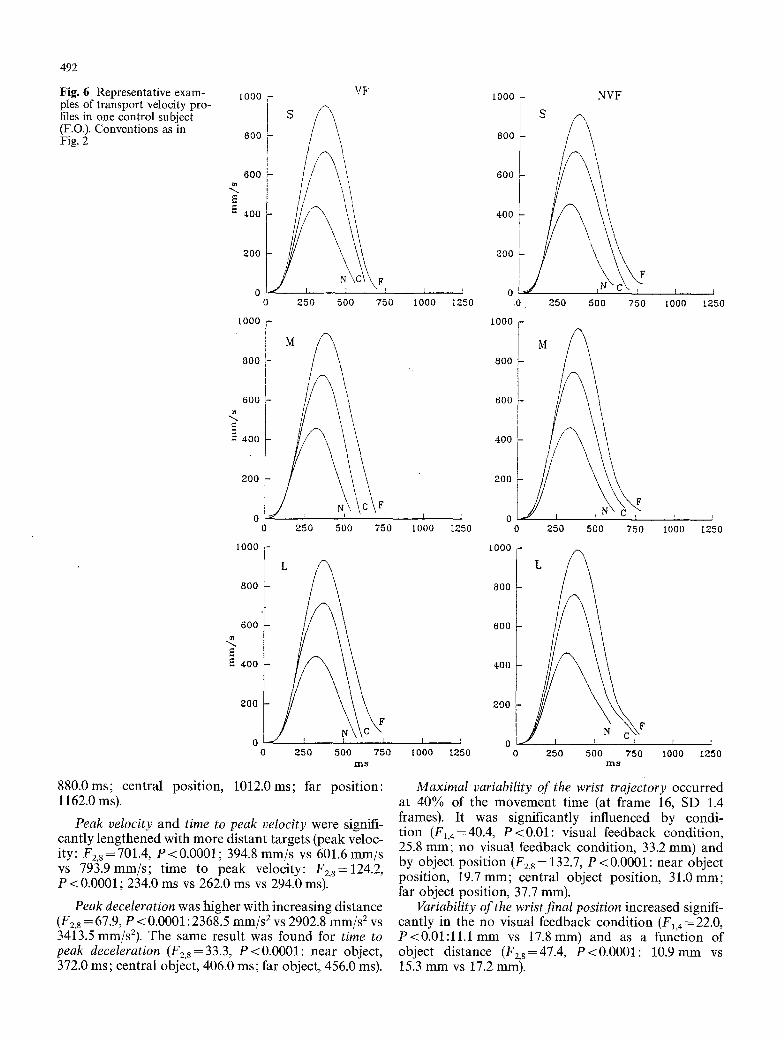
Control subjects. Figures 4 and 5 show the wrist trajecto- ries along the sagittal plane of one subject (F.O.), record- ed in the visual and no visual feedback conditions, re- spectively. Figure 6 shows representative examples of velocity profiles of the same subject. The values of the

4 9 0
r~
(3
P.
i
Z
"'~ tcq
r
N . ~
oo ~. ~ .~- ,
~ ' ~
t " q ~
t ' ~
r
r
t ' q ~
kinematic parameters averaged across the subjects are presented in Table 3. They were submitted to ANOVAs, as for the grasp kinematic parameters. The factors were: condition, object position, and object size.
Transport time increased with object position and was greater in the no visual feedback condition (object position: F2,8= 146.5, P<0.0001; 544.0 ms vs 620.8 ms vs 714.0 ms; condition: F1,4 = 108.3, P<0.001; 598.0 ms vs 654.0 ms).
Peak velocity and time to peak velocity were sig- nificantly influenced only by object position (peak ve- locity: F2,8=172.1, P<0.00001; near object position, 419.3ram/s; central object position, 695.3mm/s; far object position, 951.2mm/s; time to peak velocity: F2,s=63.9, P<0.0001; near object position, 246.0ms; central object position, 276.0 ms; far object position, 296.0 ms).
Peak deceleration and time to peak deceleration were both significantly greater with more distant object positions (peak deceleration: F2,8=34.5, P<0.001; 2372.3 mm/s 2 vs 3504.4 mm/s 2 vs 4534.5 mm/s2; time to peak deceleration: F2,8 = 130.6, P<0.0001; 368.0 ms vs 418.0 ms vs 436.0 ms).
Maximal variability of the wrist trajectories occurred at 50.0% of the movement time (at frame 20.1, SD 1.6 frames, after data normalisation in time). Variability in- creased with distance (F2,8 = 19.1, P < 0.0001 : 10.0 mm vs 14.6 mm vs 24.1 mm), but the increase was significant only for the far distance with respect to the other two distances (P < 0.05). The effect of condition was not significant (F~.4=4.1, P=0.11), although variability was on average greater in the no visual feedback than in the visual feedback condition (18.0 mm vs 14.5 mm).
Variability of the final wrist position increased signifi- cantly with distance (F2,8=15.3, P<0.005; 7.3 mm vs 8.5 mm vs 8.6 ram), although a significant difference was found only between the near object position and the other two positions (P < 0.05).
The patient. Figures 7 and 8 show the wrist trajectories along the sagittal plane of the patient recorded in the first experimental session, in the visual and no visual feedback conditions, respectively. Note the greater vari- ability of the patient trajectories than that of the con- trols (compare Figs. 4 and 5). In some trials the patient tended to overshoot the target, although she corrected the trajectory in the final phase of the movement, also without visual feedback. Figure 9 shows representative examples of velocity profiles. The velocity profiles show frequent reaccelerations during the deceleration phase, unlike the controls (compare Fig. 6). Table 4 shows the transport kinematic parameters averaged across the mean values obtained in the five experimental sessions. They were submitted to ANOVAs whose factors were: condition, object position, and object size.
Transport time was significantly affected by condi- tion (F1,4=30.6, P<0.01: visual feedback condition, 930.0 ms; no visual feedback condition, 1106.0 ms) and object position (F2.8=61.5, P<0.0001: near position,
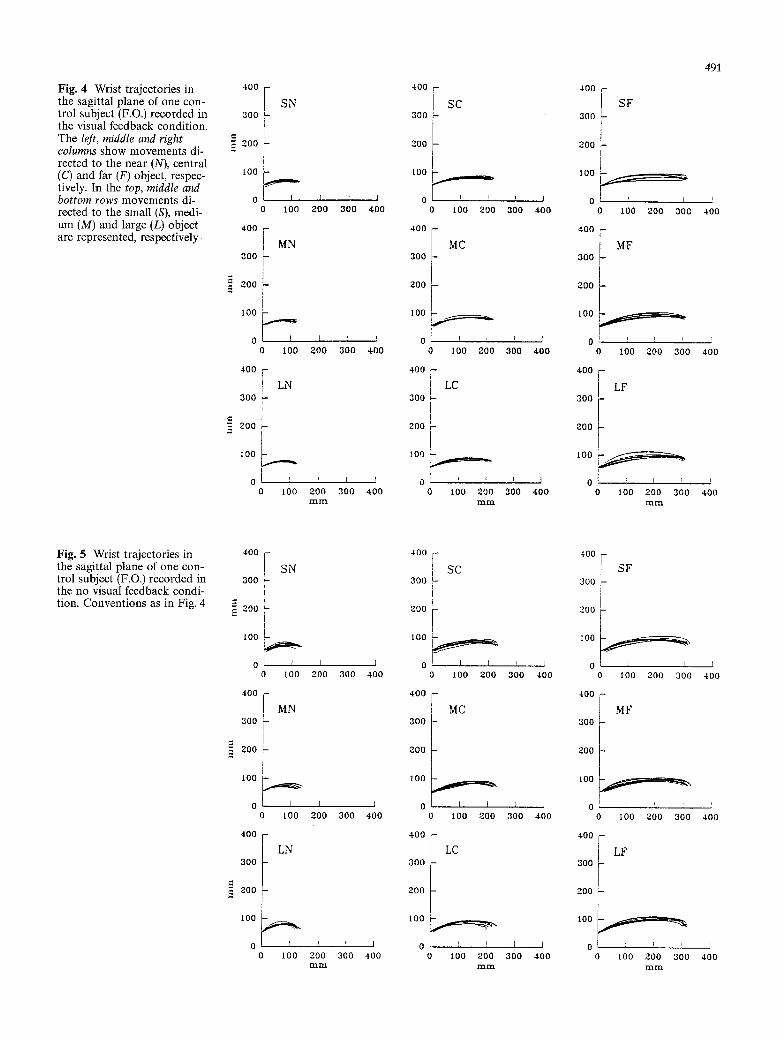
Fig. 4 Wrist trajectories in the sagittal plane of one con- trol subject (F.O.) recorded in the visual feedback condition. The left, middle and right columns show movements di- rected to the near (N), central (C) and far (F) object, respec- tively. In the top, middle and bottom rows movements di- rected to the small (S), medi- um (M) and large (L) object are represented, respectively
400 F SN
3 0 0 I " 200
tO0
0 I00
400 -
MN 300
Z00
I 0 0 -
0 0 100
400 F LN
300 i
200
i00 - ~
o[ : 0 I00
r
200
I 200
f
200 mm
I
300
300
I
300
I 400
I
400
!
400
400
I s c 300
l
200 t
lO0
0 0 I00 200
400 --
MC 300 -
300
10O
0 [ 0 I00 200
400 F
I LC I
300
[ 200
ZOO
0 t , !
0 I00 200 m m
i
3 0 0
q
300
t
300
I
400
I
400
I
400
400 r-
3oo ~ SF
200 L I
I
0 100 200 3 0 G
400
MF 3OO
200
I00
0 i t i
I00 200 300
400
LF 300
200 ~-
IO0
0 100 200 300 m m
491
i
400
i
400
i
400
Fig. 5 Wrist trajectories in the sagittal plane of one con- trol subject (F.O.) recorded in the no visual feedback condi- tion. Conventions as in Fig. 4
400
300
300 L
IO0
0 0 I 0 0
400 -
MN 3OO
2OO
I 0 0
0 R 0 I 0 0
4o0 f LN 3O0
200 --
100
0 0 I00
I ! I 200 300 400
[ ! I
200 300 400
r T i z o o 3 0 0 400 mFil
400
300
ZOO
I 0 0
0
400
300
300
I 0 0
0 0
400
300
300
I 0 0
0
F SC i ? i i
F
0 100 2 0 0
M C
F t
IO0 200
LC
' I i
I 0 0 200
l
300
]
400
r I 300 400
l I
300 400
4O0
300 I I
200
I00
0 0 I 00 200 300
4-OO
MF 300
200
I00
0 F ~ r I 0 0 20O 300
4oo I LF
300 F
200
tOO
0 0 100 200 300
miD_
r
400
I
400
!
,!-00
492
Fig. 6 Representative exam- ples of transport velocity pro- files in one control subject (F.O.). Conventions as in Fig. 2
<
I000 -
6O0
400
200
0 0 250 500
Io00.
800
0 ~ 0 250 500
I000
6OO <
4oo
800
600 <
400
200
Or-/ o
Vs
i•Fj 750
/c•F E 750
t I I 250 5OO 750
m s
1000 ~ NVF
0o! f, 600
400 [ ~
200
r ~ o f C ' - , T I
1000 1250 .0 . 250 500 750 IO00 1250
I 0 0 0 -
8 0 0
6OO
400 !
200 7~C D
0 250 500 750
I t lO00 s
1000
800
600
400
r i 1000 1250
I ' ~ F \ 250 500 750
m ~
200
I I 0 J [ T 1000 1250 0 1000 1250
880.0ms; central position, 1012.0ms; far position: 1162.0 ms).
Peak velocity and time to peak velocity were signifi- cantly lengthened with more distant targets (peak veloc- ity: F2,s=701.4, P<0.0001; 394.8 mm/s vs 601.6 mm/s vs 793.gmm/s; time to peak velocity: F2,s=124.2, P<0.0001; 234.0 ms vs 262.0 ms vs 294.0 ms).
Peak deceleration was higher with increasing distance (F2.8 = 67.9, P < 0.0001 : 2368.5 mm/s 2 vs 2902.8 mm/s 2 vs 3413.5 mm/s2). The same result was found for time to peak deceleration (F2,8=33.3, P<0.0001: near object, 372.0 ms; central object, 406.0 ms; far object, 456.0 ms).
Maximal variability of the wrist trajectory occurred at 40% of the movement time (at frame 16, SD 1.4 frames). It was significantly influenced by condi- tion (F,,4=40.4, P<0.01: visual feedback condition, 25.8 mm; no visual feedback condition, 33.2 mm) and by object position (F2,8= 132.7, P<0.0001: near object position, 19.7mm; central object position, 31.0mm; far object position, 37.7 mm).
Variability of the wrist final position increased signifi- cantly in the no visual feedback condition (F1,4 = 22.0, P < 0 . 0 1 : l l . l m m vs 17.Smm) and as a function of object distance (F2,8=47.4, P<0.0001: 10.9mm vs 15.3 mm vs 17.2 mm).

0 '.~
.~.
,Z3
~A
2~
O
O
c~
i
y,
tt'3 , ~
t t ~
eq~. o',q.
w.~- eq ~"
g
"m .
eel
~'-2.
m r
"e~d .~"
�9 o , ~ ~-~" . e ~ ' .
q ' 5 ~ ~ '-2. , ~ .
U ~
o ~
o ~ - ~ o ~ ~ ' ~ ~ ~
493
Comparison between the transport kinematics of the pa- tient and the controls. The main difference between the transport kinematics of the patient and the controls was the increase of trajectory variability observed in the pa- tient. Mean maximal variability of the wrist trajectory of the controls was 14.5 mm in the visual feedback condi- tion and 18.0 mm in the no visual feedback condition. Maximal variability of the patient trajectories was 25.8 mm in the visual feedback condition and 33.2 mm in the no visual feedback condition. The increase in vari- ability observed in the patient was significant. Mean variability value of the patient (29.5 mm) fell outside the 99% t-confidence interval estimated for the controls (16.2 + 7.2 mm). Note that movement control using only proprioception (trajectory variability of the controls in the no visual feedback condition, 18.0_+7.01 mm) was more efficient than using only vision (trajectory vari- ability of the patient in the visual feedback condition, 25.8 mm). Mean variability of the wrist final position was greater for the patient (14.5 ram) than for the controls (8.1 +2.0 mm). In addition, variability of the wrist final position was significantly greater for the patient in the visual feedback condition (11.1 mm) than for the con- trols in the no visual feedback condition (8.1 _ 2.4 mm).
Patient transport time was on average 1018 ms. It fell outside the 99% t-confidence interval calculated for the controls (626.0+ 168.0 ms). Transport time for the pa- tient in the visual feedback condition (930.4 ms) was greater than transport time for the controls in the no visual feedback condition (654.0+ 175.0 ms). However, the mean values of the other kinematic transport parameters for the patient were comparable with those of the controls (peak velocity: patient, 596.8 mm/s; con- trols, 688.5 _+ 241.9 mm/s; time to peak velocity: patient, 263.4 ms; controls, 273.0-t-86.4 ms; peak deceleration: patient, 2894.0 mm/s 2, controls, 3470.4_+ 2452.5 mm/s2; time to peak deceleration: patient, 411.4ms, controls, 407.4_+ 118 ms). The lengthening of transport time for the patient can be explained by the frequent reaccelera- tions (corrections) observed in the velocity profiles dur- ing the last phase of deceleration (low-velocity phase after peak deceleration).
Grasp perturbation experiment
Grasp component
Control subjects. Figure 10 (top row) shows representa- tive examples of the time course of the grasp during perturbed and unperturbed trials, in the visual and no visual feedback conditions, respectively. Note the ab- sence of grasp interruptions during the perturbed trials. Although the effects of the perturbation were corrected, as regards the final grasp aperture, finger opening did not reach the same maximum as in the unperturbed trials. The mean values of the grasp kinematic parame- ters are presented in Table 5. They were submitted to ANOVAs whose main factors were: condition (visual

494
Fig. 7 The patient's wrist tra- jectories in the sagittal plane recorded in the visual feed- back condition during the first experimental session. Conven- tions as in Fig. 4
300
400 -
SN
: 2 0 0
i00
, I I
0 100 2 0 0 3 0 0 400
400
300
~. 200
[ 0 0
0
400 --
SC 300 I
200
I 0 0 ~ ~
0 I I 0 10O 200 300 400
400
MN 300
200
I00
I I I ~ 0
400
300
200
I00
0
0 I 0 0 2 0 0 300 400
LN
t ~ T I
O I00 200 300 400 m m
MC
r ' I ] 0 I 0 0 200 300 4 0 0
40O
LC 30O
200 r"-
,ooL 0 ' ! ' [ [
0 I 0 0 200 300 400 m m
400
300
200
I 0 0
0
300
200
100
0
400
SF
1 I i 1
0 I 0 0 2 0 0 300 400
400
MF
P I F l
0 tO0 2 0 0 300 400
300
200
I00
0
LF
XO0 200 300 400 m m
Fig. 8 The patient's wrist tra- jectories in the sagittal plane recorded in the no visual feed- back condition during the first experimental session. Conven- tions as in Fig. 4
400 I SN
300
L 200
0 , ~ * I 0 100 200 300 400
400 -
300 -
2 0 0 -
iO0
0
MN
400
300
200
[00
0
I i [ ]
0 I 0 0 200 300 4-00
LN
l I ! I
0 I 0 0 200 300 400 m m
4 0 0 -
300 -
200
I 0 0
0
4 0 0 -
300
200
100
0
400
300
200
100
0
SC
I 0 0 200 300 400
MC
P ~ T I
I 0 0 200 :300 400
LC
T I T l 100 2 0 0 3 0 0 4 0 0
m m
400 -
SF O00 I
200
,oot 0 1
0 I 0 0 2 0 0 300 4-00
400
300
200
IO0
0
4-00
300
200
I.O0
0
MF
i ! , j
I 0 0 2 0 0 300 400
LF
i I i ]
I 0 0 2 0 0 300 400 m m
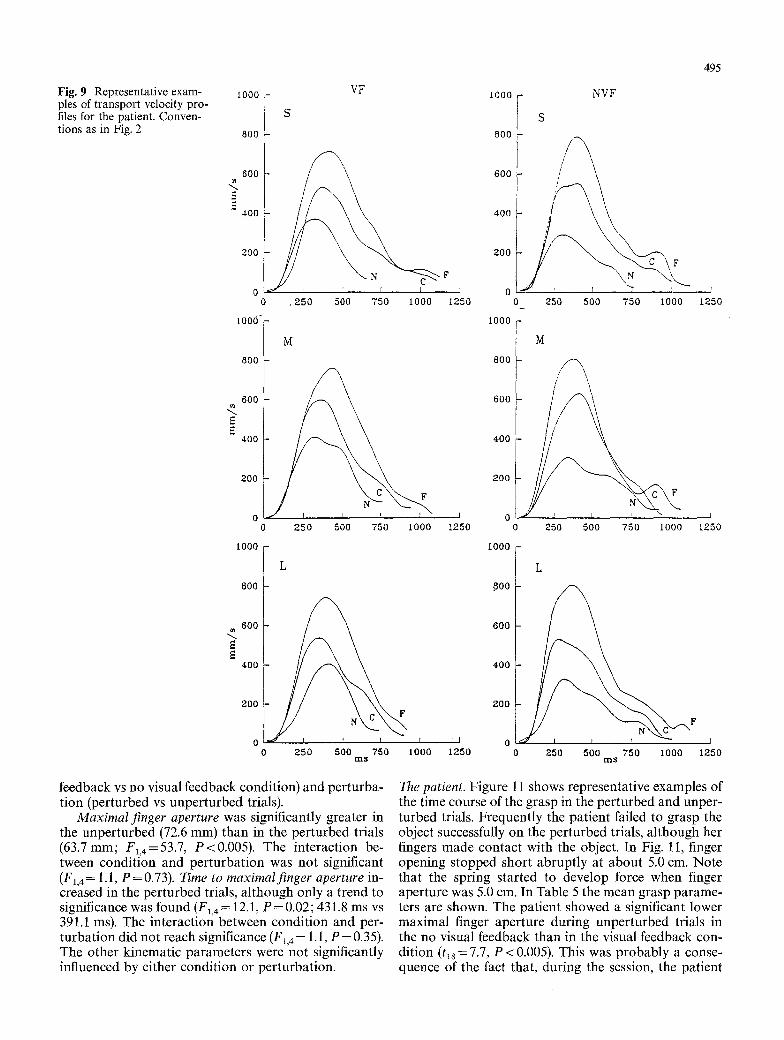
Fig. 9 Representative exam- ples of transport velocity pro- files for the patient. Conven- tions as in Fig. 2
[000
800
VF
S
600
40o
2OO
0 1 ~ r , 2 5 0 500 750 I 0 0 0
600 ~ / ~
,oot 0 250 500 750 1 0 0 0
1000
L
80O
. ~ 6 0 0
40O
20O F
0 t 250 500 750 I 0 0 0
m s
F F
1250
I
1250
I000
800
600
400
200
0 0
t 0 0 0
800
600
400
200
0 0
tO00
aoo
600
400
200
1 o 125o 0
NVF
S
L 250 500 750 1000
495
1250
S
.VI
/
! i f I
250 500 750 1000 1250
L
F
J
250 500 750 I 0 0 0 1250 m s
feedback vs no visual feedback condition) and perturba- tion (perturbed vs unperturbed trials).
Maximal finger aperture was significantly greater in the unperturbed (72.6 mm) than in the perturbed trials (63.7mm; F~,4=53.7, P<0.005). The interaction be- tween condition and perturbation was not significant (F1, 4-= 1.1, P = 0.73). Time to maximalfinger aperture in- creased in the perturbed trials, although only a trend to significance was found (F1,4---- 12.1, P = 0.02; 431.8 ms vs 391.1 ms). The interaction between condition and per- turbation did not reach significance ( E l , 4 - - 1.1, P = 0.35). The other kinematic parameters were not significantly influenced by either condition or perturbation.
The patient. Figure 11 shows representative examples of the time course of the grasp in the perturbed and unper- turbed trials. Frequently the patient failed to grasp the object successfully on the perturbed trials, although her fingers made contact with the object. In Fig. 11, finger opening stopped short abruptly at about 5.0 cm. Note that the spring started to develop force when finger aperture was 5.0 cm. In Table 5 the mean grasp parame- ters are shown. The patient showed a significant lower maximal finger aperture during unperturbed trials in the no visual feedback than in the visual feedback con- dition (t18 = 7.7, P <0.005). This was probably a conse- quence of the fact that, during the session, the patient

496
o
o
. ~
z Z
O
(D
e 4 ~
eq~.. e z ~ O ,---~
o~
r
oO
c~
eqe4
"~''4
'q.~"
mm,~
O ~
~ t t 3
m . ~
,-06
t--,-A ~ "~',,d oq.~- t",l
t . - 4 ~
e q ~ : .
"~m~,~ ' ~
. ~ . ~ ,
e q ~ t",l ~
~ e 4
"~ e4
e n ~
eqe,i
eqr4
eq~/
o8
became aware of being unable to perceive and correct for perturbations. However, maximal finger aperture was significantly lower during perturbed trials than dur- ing unperturbed trials (t37 = 7.7, P <0.0001). No signifi- cant difference in maximal finger aperture was found when the perturbation was applied during movements executed either with or without visual feedback (tlv= 0.70, P = 0.50). On perturbed trials the mean value for the patient (51.0 mm) lay outside the 99% t-confi- dence interval measured for the control subjects (63.7 _+ 8.4 mm).
Transport component
Control subjects. Representative examples of velocity profiles are shown in Fig. 10. Note no difference be- tween perturbed and unperturbed trials. ANOVAs (fac- tors: condition and perturbation) were performed on the kinematic parameters whose mean values are pre- sented in Table 5. Neither factor significantly affected any kinematic parameter of transport.
The patient. Figure 11 shows representative examples of velocity profiles of the patient's hand transport during perturbed and unperturbed trials. Note that the trans- port time of the patient was greater than that of the controls, as shown in Table 5. However, the values of the other kinematic transport parameters were similar.
Discussion
The first aim of the present study was to determine which phases of prehension, and in particular which phases of the grasp component, can be appropriately controlled without proprioceptive information. For this purpose, the prehension kinematic parameters of a pe- ripherally deafferented patient were compared with those of healthy controls. Prehension movements exe- cuted with and without visual feedback were studied. An additional experiment was carried out in which we analysed the kinematic modifications of the prehension movement in response to a mechanical perturbation ap- plied to the fingers in both the patient and the controls.
The pattern of the patient's hand-shaping, during the finger-opening phase, was the same as that of the con- trols. This was also the case during movements executed without visual feedback. Maximal finger aperture and peak velocity of finger opening increased by the same amount in proportion to object size for the patient as for the controls. However, the patient's temporal parame- ters and variability of grasp were greater than those of the controls. An obvious explanation of these results is that in the absence of feedback control the movement variability of trajectories increased. Consequently, the patient adopted the strategy of moving more slowly in an attempt to compensate for this increase. Another possible explanation is that planning of the temporal
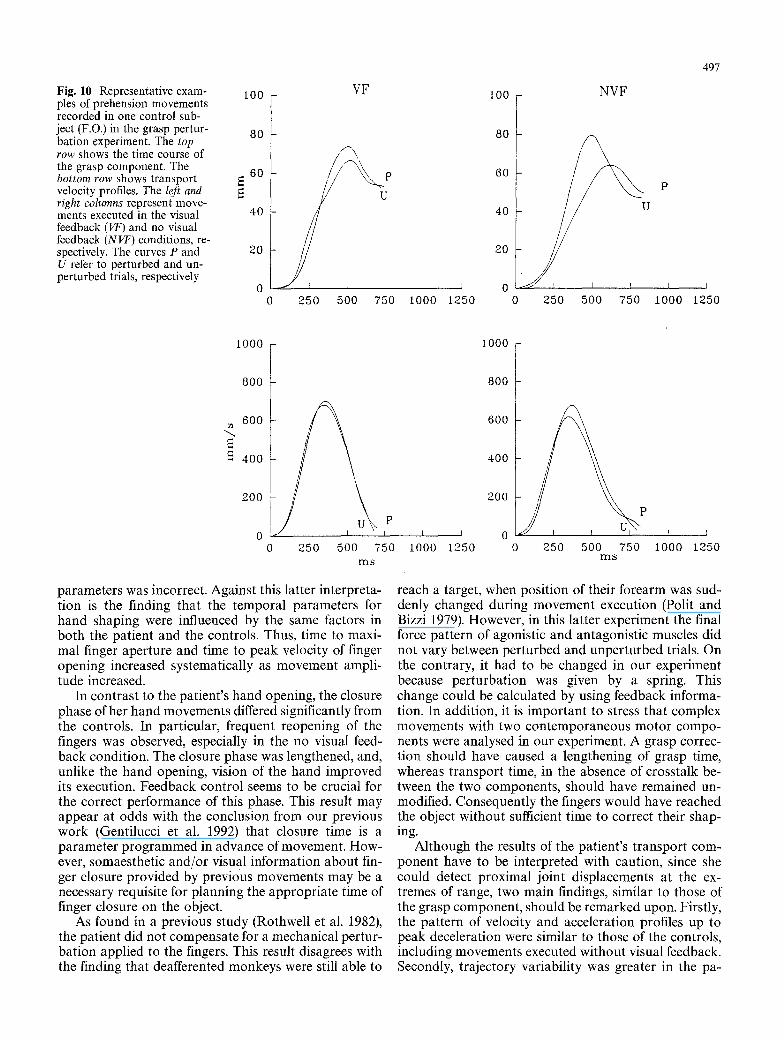
Fig. 10 Representative exam- ples of prehension movements recorded in one control sub- ject (F.O.) in the grasp pertur- bation experiment. The top row shows the time course of the grasp component. The bottom row shows transport velocity profiles. The left and right columns represent move- ments executed in the visual feedback (VF) and no visual feedback (NVF) conditions, re- spectively. The curves P and U refer to perturbed and un- perturbed trials, respectively
tO0
80
6O
40
20
VF
I
250 500 750
497
I 0 0 0
loo NVF
60 p
40
20
I I I p 0
1000 1250 0 250 500 750 i000 1250
I000
800
< =
800
600
4 0 0
200
0 250 500 75O m s
parameters was incorrect. Against this latter interpreta- tion is the finding that the temporal parameters for hand shaping were influenced by the same factors in both the patient and the controls. Thus, time to maxi- mal finger aperture and time to peak velocity of finger opening increased systematically as movement ampli- tude increased.
In contrast to the patient's hand opening, the closure phase of her hand movements differed significantly from the controls. In particular, frequent reopening of the fingers was observed, especially in the no visual feed- back condition. The closure phase was lengthened, and, unlike the hand opening, vision of the hand improved its execution. Feedback control seems to be crucial for the correct performance of this phase. This result may appear at odds with the conclusion from our previous work (Gentilucci et al. 1992) that closure time is a parameter programmed in advance of movement. How- ever, somaesthetic and/or visual information about fin- ger closure provided by previous movements may be a necessary requisite for planning the appropriate time of finger closure on the object.
As found in a previous study (Rothwell et al. 1982), the patient did not compensate for a mechanical pertur- bation applied to the fingers. This result disagrees with the finding that deafferented monkeys were still able to
600
400
200
P
250 500 750 m s
_ _ _ L _ _ J 0 ~ I
1000 1250 0 1000 1250
reach a target, when position of their forearm was sud- denly changed during movement execution (Polit and Bizzi 1979). However, in this latter experiment the final force pattern of agonistic and antagonistic muscles did not vary between perturbed and unperturbed trials. On the contrary, it had to be changed in our experiment because perturbation was given by a spring. This change could be calculated by using feedback informa- tion. In addition, it is important to stress that complex movements with two contemporaneous motor compo- nents were analysed in our experiment. A grasp correc- tion should have caused a lengthening of grasp time, whereas transport time, in the absence of crosstalk be- tween the two components, should have remained un- modified. Consequently the fingers would have reached the object without sufficient time to correct their shap- ing.
Although the results of the patient's transport com- ponent have to be interpreted with caution, since she could detect proximal joint displacements at the ex- tremes of range, two main findings, similar to those of the grasp component, should be remarked upon. Firstly, the pattern of velocity and acceleration profiles up to peak deceleration were similar to those of the controls, including movements executed without visual feedback. Secondly, trajectory variability was greater in the pa-
498
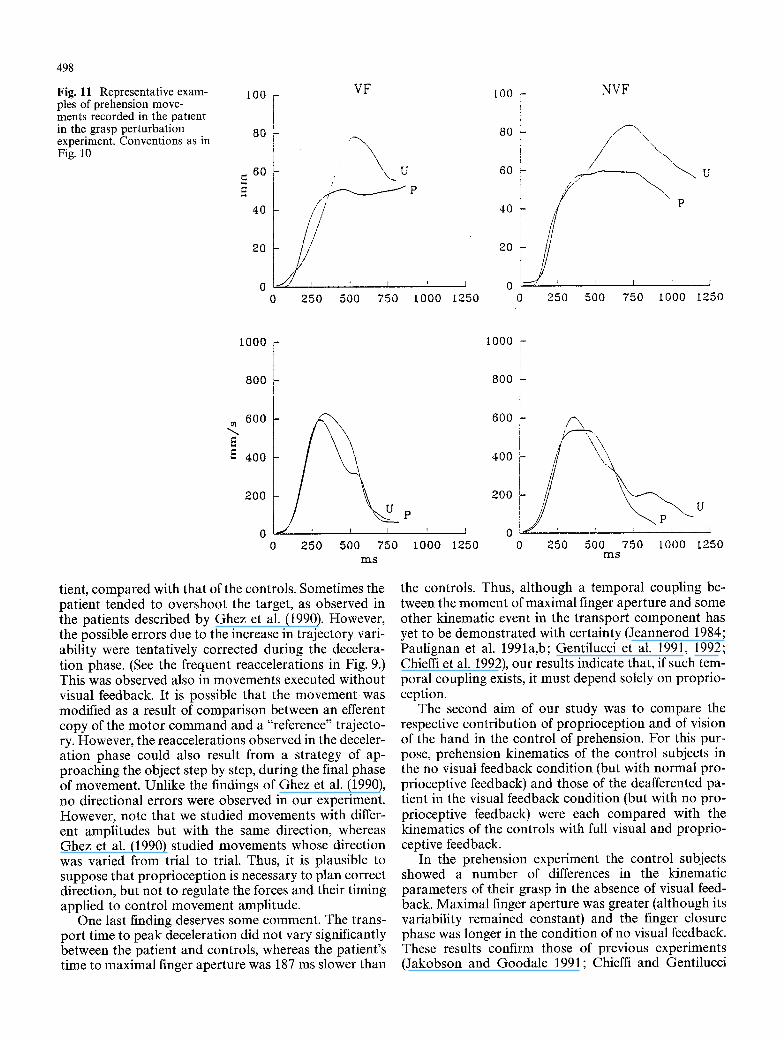
Fig. 11 Representative exam- ples of prehension move- ments recorded in the patient in the grasp perturbation experiment. Conventions as in Fig. 10
I00 -
80
. 6 o
4o
20
VF
L--
0 250 i I ' i
500 750 1000 1250
I 0 0 -
80 -
60
40
20
o o
NVF
\ \ \ /
250 500 750 1000
P
I
1250
1000
800
600
fi fi 400
200 U
o 250 500 750 m s
tient, compared with that of the controls. Sometimes the patient tended to overshoot the target, as observed in the patients described by Ghez et al. (1990). However, the possible errors due to the increase in trajectory vari- ability were tentatively corrected during the decelera- tion phase. (See the frequent reaccelerations in Fig. 9.) This was observed also in movements executed without visual feedback. It is possible that the movement was modified as a result of comparison between an efferent copy of the motor command and a "reference" trajecto- ry. However, the reaccelerations observed in the deceler- ation phase could also result from a strategy of ap- proaching the object step by step, during the final phase of movement. Unlike the findings of Ghez et al. (1990), no directional errors were observed in our experiment. However, note that we studied movements with differ- ent amplitudes but with the same direction, whereas Ghez et al. (1990) studied movements whose direction was varied from trial to trial. Thus, it is plausible to suppose that proprioception is necessary to plan correct direction, but not to regulate the forces and their timing applied to control movement amplitude.
One last finding deserves some comment. The trans- port time to peak deceleration did not vary significantly between the patient and controls, whereas the patient's time to maximal finger aperture was 187 ms slower than
I 0 0 0 -
P
1 0 0 0 1 2 5 0
800
600 t " \
U
0 I 0 250 500 750 1000 1250
m s
the controls. Thus, although a temporal coupling be- tween the moment of maximal finger aperture and some other kinematic event in the transport component has yet to be demonstrated with certainty (Jeannerod 1984; Paulignan et al. 1991a,b; Gentilucci et al. 1991, 1992; Chieffi et al. 1992), our results indicate that, if such tem- poral coupling exists, it must depend solely on proprio- ception.
The second aim of our study was to compare the respective contribution of proprioception and of vision of the hand in the control of prehension. For this pur- pose, prehension kinematics of the control subjects in the no visual feedback condition (but with normal pro- prioceptive feedback) and those of the deafferented pa- tient in the visual feedback condition (but with no pro- prioceptive feedback) were each compared with the kinematics of the controls with full visual and proprio- ceptive feedback.
In the prehension experiment the control subjects showed a number of differences in the kinematic parameters of their grasp in the absence of visual feed- back. Maximal finger aperture was greater (although its variability remained constant) and the finger closure phase was longer in the condition of no visual feedback. These results confirm those of previous experiments (Jakobson and Goodale 1991; Chieffi and Gentilucci
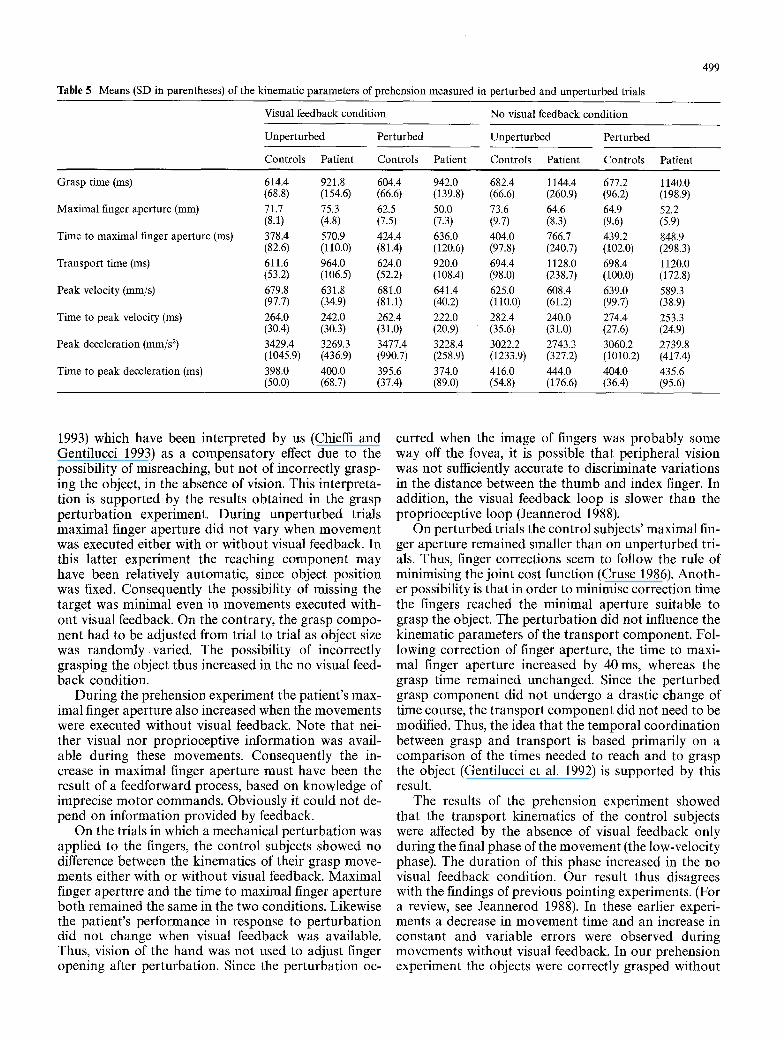
Table 5 Means (SD in parentheses) of the kinematic parameters of prehension measured in perturbed and unperturbed trials
499
Visual feedback condition No visual feedback condition
Unperturbed Perturbed Unperturbed Perturbed
Controls Patient Controls Patient Controls Patient Controls Patient
Grasp time (ms) 614.4 921.8 604.4 942.0 682.4 1144.4 (68.8) (154.6) (66.6) (139.8) (66.6) (260.9)
Maximal finger aperture (mm) 71.7 75.3 62.5 50.0 73.6 64.6 (8.1) (4.8) (7.5) (7.3) (9.7) (8.3)
Time to maximal finger aperture (ms) 378.4 570.9 424.4 636.0 404.0 766.7 (82.6) (110.0) (81.4) (120.6) (97.8) (240.7)
Transport time (ms) 611.6 964.0 624.0 920.0 694.4 1128.0 (53.2) (106.5) (52.2) (108.4) (98.0) (238.7)
Peak velocity (mm/s) 679.8 631.8 681.0 641.4 625.0 608.4 (97.7) (34.9) (81.1) (40.2) (110.0) (61.2)
Time to peak velocity (ms) 264.0 242.0 262.4 222.0 282.4 240.0 (30.4) (30.3) (31.0) (20.9) (35.6) (31.0)
Peak deceleration (mm/s 2) 3429.4 3269.3 3477.4 3228.4 3022.2 2743.3 (1045.9) (436.9) (990.7) (258.9) (1233.9) (327.2)
Time to peak deceleration (ms) 398.0 400.0 395.6 374.0 416.0 444.0 (50.0) (68.7) (37.4) (89.0) (54.8) (176.6)
677.2 1140.0 (96.2) (198.9) 64.9 52.2 (9.6) (5.9) 439.2 848.9 (102.0) (298.3) 698.4 1120.0 (100.0) (172.8) 639.0 589.3 (99.7) (38.9) 274.4 253.3 (27.6) (24.9) 3060.2 2739.8 (1010.2) (417.4) 404.0 435.6 (36.4) (95.6)
1993) which have been interpreted by us (Chieffi and Gentilucci 1993) as a compensatory effect due to the possibility of misreaching, but not of incorrectly grasp- ing the object, in the absence of vision. This interpreta- tion is supported by the results obtained in the grasp perturbation experiment. During unperturbed trials maximal finger aperture did not vary when movement was executed either with or without visual feedback. In this latter experiment the reaching component may have been relatively automatic, since object position was fixed. Consequently the possibility of missing the target was minimal even in movements executed with- out visual feedback. On the contrary, the grasp compo- nent had to be adjusted from trial to trial as object size was randomly varied. The possibility of incorrectly grasping the object thus increased in the no visual feed- back condition.
During the prehension experiment the patient's max- imal finger aperture also increased when the movements were executed without visual feedback. Note that nei- ther visual nor proprioceptive information was avail- able during these movements. Consequently the in- crease in maximal finger aperture must have been the result of a feedforward process, based on knowledge of imprecise motor commands. Obviously it could not de- pend on information provided by feedback.
On the trials in which a mechanical perturbation was applied to the fingers, the control subjects showed no difference between the kinematics of their grasp move- ments either with or without visual feedback. Maximal finger aperture and the time to maximal finger aperture both remained the same in the two conditions. Likewise the patient's performance in response to perturbation did not change when visual feedback was available. Thus, vision of the hand was not used to adjust finger opening after perturbation. Since the perturbation oc-
curred when the image of fingers was probably some way off the fovea, it is possible that peripheral vision was not sufficiently accurate to discriminate variations in the distance between the thumb and index finger. In addition, the visual feedback loop is slower than the proprioceptive loop (Jeannerod 1988).
On perturbed trials the control subjects' maximal fin- ger aperture remained smaller than on unperturbed tri- als. Thus, finger corrections seem to follow the rule of minimising the joint cost function (Cruse 1986). Anoth- er possibility is that in order to minimise correction time the fingers reached the minimal aperture suitable to grasp the object. The perturbation did not influence the kinematic parameters of the transport component. Fol- lowing correction of finger aperture, the time to maxi- mal finger aperture increased by 40 ms, whereas the grasp time remained unchanged. Since the perturbed grasp component did not undergo a drastic change of time course, the transport component did not need to be modified. Thus, the idea that the temporal coordination between grasp and transport is based primarily on a comparison of the times needed to reach and to grasp the object (Gentilucci et al. 1992) is supported by this result.
The results of the prehension experiment showed that the transport kinematics of the control subjects were affected by the absence of visual feedback only during the final phase of the movement (the low-velocity phase). The duration of this phase increased in the no visual feedback condition. Our result thus disagrees with the findings of previous pointing experiments. (For a review, see Jeannerod 1988). In these earlier experi- ments a decrease in movement time and an increase in constant and variable errors were observed during movements without visual feedback. In our prehension experiment the objects were correctly grasped without

500
vision of the hand. In addition, variabil i ty of the final wrist posi t ion did not change between bo th the condi- tions of visual and no visual feedback. It is likely that visual feedback mainly intervenes in the control of the final phase of the m ovem en t when the image of bo th hand and object is near the fovea. When visual feedback informat ion is not available, a correct final hand posi- t ion can be achieved by decreasing hand velocity during this phase.
The kinematics of bo th the t ranspor t and the grasp componen t s differed between the condit ions of purely visual control of the m ovem en t (the pat ient 's perfor- mance in the visual feedback condition) and of purely propr iocept ive control of the m ovem en t (the control subjects' per formance in the no visual feedback condi- tion). Variability and t ime da ta of bo th the componen t s increased in the absence of propr iocept ive control, espe- cially in the final phase of the movement . Since the pro- prioceptive loop is faster than the visual loop (Jean- nerod 1988), it is likely that the slowed t ime course of movem e n t in the pat ient reflects an a t tempt to correct the effects of the noise inherent in m o t o r commands (Schmidt et al. 1979) by using the available visual feed- back. However , the increase in the t empora l parameters was greater than that accounted for in terms of the slow- er visual processing (approximate ly 50 ms slower than propr iocept ive processing, according to Jeannerod 1988). In addition, tactile sensation was completely lost in our patient. Tactile signals, when the fingers touch the object at the end of grasping, may contr ibute to the selection of the appropr ia te t empora l pa ramete rs of pre- hension.
Propr iocept ion thus appears to exert a more efficient control during whole m ovem en t than does vision of the hand. Unlike the initial phase of prehension, during exe- cution of the final phase of the m o v e m e n t bo th visual and propr iocept ive informat ion seem to be necessary. The necessity of sensory information, and in part icular of propr iocept ive information, accords with the hypoth- esis that the final phase of the m ovem en t is executed mainly on line (Woodsworth 1899). However , it is also possible that the final phase of a m o t o r p rog ram pro- gressively degrades over successive trials if it does not receive somaesthet ic informat ion abou t its execution.
Acknowledgements The authors are grateful to Dr. A. Allport for valuable comments on the manuscript. They thank Professors G. Rizzolatti and C.A. Umilta', and Dr. A. Negrotti for discussion of data. The work was supported by a research grant from the Human Frontier Science Program, and by grants CNR and MURST to M.G.
References Arbib MA (1981) Perceptual structures and distributed motor
control. In: Brooks VB (ed) Motor control. (Handbook of physiology, sect 1, The nervous system, vol II, part 2) William and Wilkins, Baltimore, pp 1449-1480
Chieffi S, Gentilucci M (1993) Coordination between the trans- port and the grasp components during prehension move- ments. Exp Brain Res 94:471-477
Chieffi S, Fogassi L, Gallese V, Gentilucci M (1992) Prehension movements directed to approaching objects: influence of stim- ulus velocity on the transport and the grasp components. Neu- ropsychologia 30:877-897
Cruse H (1986) Constraints for joint angle control of the human arm. Biol Cybern 54:125-132
Gentilucci M, Castiello U, Corradini ML, Scarpa M, Umilta' C, Rizzolatti G (1991) Influence of different types of grasping on the transport component of prehension movements. Neu- ropsychologia 29: 361-378
Gentilucci M, Chieffi S, Scarpa M, Castiello U (1992) Temporal coupling between transport and grasp components during pre- hension movements: effects of visual perturbation. Behav Brain Res 47:71-82
Ghez C, Gordon J, Ghilardi MF, Christakos CN, Cooper SE (1990) Roles of proprioceptive input in the programming of arm trajectory. Cold Spring Harb Symp Quant Biol 55:837- 847
Jakobson LS, Goodale MA (1991) Factors affecting higher-order movement planning: a kinematic analysis of human prehen- sion. Exp Brain Res 86:199-208
Jeannerod M (1984) The timing of natural prehension movements. J Mot Behav 16:235 254
Jeannerod M (1988) The neural and behavioural organization of goal-directed movements. Clarendon, Oxford
Jeannerod M, Michel F, Prablanc C (1984) The control of hand movements in a case of hemianaesthesia following a parietal lesion. Brain 107:899420
Lashley KS (1917) The accuracy of movement in the absence of excitation from the moving organ. Am J Physiol 43:169-194
Luria AR (1973) The working brain. An introduction to neu- ropsychology (English translation) Penguin Books, London
Mott FW, Sherrington CS (1895) Experiments upon the influence of sensory nerves upon movement and nutrition of the limbs. Proc R Soc Lond [Biol] 57:481-488
Paulignan Y, Mackenzie C, Marteniuk R, Jeannerod M (1991a) Selective perturbation of visual input during prehension movements. 1. The effects of changing object position. Exp Brain Res 83:415-451
Paulignan Y, Jeannerod M, Mackenzie C, Marteniuk R (1991b) Selective perturbation of visual input during prehension movements. 1. The effects of changing object size. Exp Brain Res 87:407-420
Polit A, Bizzi E (1979) Characteristics of the motor programs underlying arm movements in monkeys. J Neurophysiol 42:183-194
Rizzolatti G, Camarda R, Fogassi L, Gentilucci M, Luppino G, Matelli M (1988) Functional organization in inferior area 6 in the macaque monkey. II. Area F5 and the control of distal movements. Exp Brain Res 71:491 507
Rothwell JC, Taub MM, Day BL, Obeso JA, Thomas PK, Mard- sen CD (1982) Manual motor performance in a deafferented man. Brain 105:515-542
Sanes JN, Mauritz KH, Dalakas MC, Evarts EV (1985) Motor control in humans with large-fiber sensory neuropathy. Hum Neurobiol 4:101-114
Schmidt RA, Zelaznik H, Hawkins B, Frank JS, Quinn JT (1979) Motor output variability: a theory for rapid motor acts. Psy- chol Rev 86:415-451
Soechting JF, Flanders M (1989) Sensorimotor representations for pointing to targets in three-dimensional space. J Neuro- physiol 62:582-594
Taub E, Goldberg IA, Taub P (1975) Deafferentation in monkeys: pointing at a target without visual feedback. Exp Neurol 46:178-186
Von Hofsten C, Ronnqvist L (1988) Preparation for grasping an object: a development study. J Exp Psychol 4:610-621
Woodsworth RS (1899) The accuracy of voluntary movements. Psychol Res Monogr [Suppl] 3:1-114














![Peripherally administered [Nle4, D-Phe7]-MSH increases resting metabolic rate, while peripheral AgRP has no effect, in wild type C57BL/6 and ob/ob mice.](https://static.fdokumen.com/doc/165x107/631268023ed465f0570a36fc/peripherally-administered-nle4-d-phe7-msh-increases-resting-metabolic-rate-while.jpg)



