
Coherent Spontaneous Activity Identifies a Hippocampal-Parietal Memory Network

A
Fitrtn©
K
1
rcsapcit“psrs“
v
0d
Neuropsychologia 44 (2006) 2668–2684
The role of parietal cortex in visuomotor control:What have we learned from neuroimaging?
Jody C. Culham ∗, Cristiana Cavina-Pratesi, Anthony SinghalDepartment of Psychology, Social Science Centre, University of Western Ontario, London Ont., Canada N6A 5C2
Received 8 May 2005; received in revised form 7 October 2005; accepted 3 November 2005Available online 9 December 2005
bstract
Research from macaque neurophysiology and human neuropsychology has implicated the parietal cortex in the sensory control of action.unctional neuroimaging has been very valuable in localizing and characterizing specific regions of the human brain involved in visuomotor actions
nvolving different effectors, such as the eyes, head, arms and hands. Here, we review the areas discovered by human neuroimaging, includinghe putative functional equivalents of the following macaque regions: parietal eye fields (PEF), ventral intraparietal (VIP) area, parietal reach
egion (PRR) and the anterior intraparietal (AIP) area. We discuss the challenges of studying realistic movements in the imaging environment,he lateralization of visuomotor function, caveats involved in proposing interspecies homologies and the limitations and future directions foreuroimaging studies of visuomotor control.2005 Elsevier Ltd. All rights reserved.
raspin
ibmcmbhirspotcp2
eywords: Functional neuroimaging; Visuomotor functions; Eye movement; G
. Introduction
Research from macaque neurophysiology and human neu-opsychology has suggested that the parietal cortex plays aritical role in numerous cognitive functions, particularly in theensory control of action. In the traditional view of Ungerleidernd Mishkin (1982), parietal cortex forms part of a dorsal visualathway that encodes spatial location, in contrast to temporalortex, which forms part of a ventral visual pathway that isnvolved in object recognition. That is, they suggested that thewo pathways were involved in coding “where” an object is andwhat” an object is, respectively. Goodale and Milner (1992)rovided an alternate interpretation of the two visual streams byuggesting that the dorsal pathway, from visual cortex to supe-ior parietal cortex, was involved not in spatial processing pere, but in the visual guidance of action, specifically determining
how” to interact with an object.The posterior parietal cortex (PPC) is situated between theisual cortex in the occipital lobe and the somatosensory cortex
∗ Corresponding author. Tel.: +1 519 661 2111x85643; fax: +1 519 661 3961.E-mail address: [email protected] (J.C. Culham).URL: http://defiant.ssc.uwo.ca/Jody web.
gtieara
028-3932/$ – see front matter © 2005 Elsevier Ltd. All rights reserved.oi:10.1016/j.neuropsychologia.2005.11.003
g; Reaching; Hemispheric difference
n the postcentral gyrus. Thus, it is well positioned to receiveoth visual and somatosensory input and to send output to pre-otor and motor areas in frontal cortex. PPC is historically
onsidered to be “association cortex”, which integrates infor-ation across modalities; however, as research techniques have
ecome more sophisticated, more definitive parietal functionsave been attributed to specific subregions. Anatomically, PPCs divided into the superior parietal lobule (SPL) and infe-ior parietal lobule (IPL) by the intraparietal sulcus (IPS), ashown in Fig. 1. On the medial aspect of the parietal lobe, therecuneus (PCu) is the region that is anterior to the parieto-ccipital sulcus, posterior to the paracentral lobule and superioro the subparietal sulcus, which separates it from retrosplenialortex below (Damasio, 2005). The parietal lobe in general com-rises approximately 25% of a cerebral hemisphere (Damasio,005).
Electrophysiological data from macaque monkeys has sug-ested that there is a mosaic of sensorimotor areas aroundhe IPS. Various divisions within the IPS have been involvedn the control of specific effectors (body parts), including the
yes (lateral intraparietal area, LIP), head (ventral intraparietalrea, VIP), arms (parietal reach region, PRR) and hand (ante-ior intraparietal area, AIP). Above the IPS, macaque area 5 ishigher order somatosensory region. Below the IPS, neurons
J.C. Culham et al. / Neuropsychol
Fig. 1. The posterior parietal cortex (PPC). Anatomical illustration of thepostero-lateral (a) and medial (b) views of the left hemisphere of the human brain(pial surface) of one subject. The white lines highlight the principal sulci: cen-tral sulcus (CS), postcentral sulcus (PCS), intraparietal sulcus (IPS), transverseoccipital sulcus (TOS), parieto-occipital sulcus (POS), the ascending ramus ofthe cingulate sulcus (arCingS) and the subparietal sulcus (sPS). Different colorsrepresent different anatomical subdivisions of the PPC: the postcentral gyrus(PCG), the superior parietal lobule (SPL), the precuneus (PCu) and the inferiorparietal lobule (IPL), which is divided into the supramarginal gyrus (SMG) andaat
iol&
teepatn
po
tnonmavtttnifi
utitammpthmFtidlsHftcrpa
2
2
afie1B&
ngular gyrus (AG). The SPL and PCu include both Brodmann areas 5 (BA5)nd 7 (BA7). Note, the PCG is part of the parietal lobe, but is not included inhe PPC.
n macaque areas 7a (posterior IPL) show largely visual andculomotor responses; whereas, neurons 7b (anterior IPL) showargely somatosensory responses (Andersen, Asanuma, Essick,
Siegel, 1990).Neuropsychological data has demonstrated many different
ypes of deficits in human patients with parietal damage. Pari-tal patients may have difficulties exploring space with theiryes (gaze apraxia), reaching to visual targets (optic ataxia), or
rogramming and sequencing skilled movements (apraxia). Inddition, patients may have other attentional problems, such ashe inability to notice more than one object at a time (simultanag-osia) or reduced awareness of contralesional objects (neglect),nfimt
ogia 44 (2006) 2668–2684 2669
articularly when ipsilesional objects are presented simultane-usly (extinction).
The advent of neuroimaging, particularly positron emissionomography (PET) in the 1980s and functional magnetic reso-ance imaging (fMRI) in the 1990s, has advanced our knowledgef cognitive function. This is largely because, unlike monkeyeurophysiology or human neuropsychology, it allows the nor-al intact human brain to be studied with high spatial resolution
nd moderate temporal resolution. However, the advances pro-ided by neuroimaging have been slower for parietal cortexhan for other regions of cortex. This is largely due to theechnical challenges of studying visuomotor functions, givenhe many constraints of the imaging environment. Nevertheless,umerous labs have been solving these problems and advanc-ng the field, so the time is appropriate for a review of thesendings.
Here, we review the contributions of neuroimaging to thenderstanding of the role of parietal cortex in visuomotor con-rol. We consider a number of crucial visuomotor actions involv-ng different effectors. Specifically, we consider eye movements,he multimodal encoding of locations near the head, reachingnd pointing movements with the arm and finger and graspingovements that require preshaping of the hand. For each type ofovement, we begin with a brief review about what was known
reviously from the two main techniques that have investigatedhe neural substrates of actions: macaque neurophysiology anduman neuropsychology. Each section then follows with a sum-ary of the human neuroimaging results and their implications.ig. 2 and Table 1 highlight the anatomical locations and stereo-
axic coordinates of activation from key visuomotor neuroimag-ng studies (and one study of neurological lesions). Although weiscuss human areas in terms of their likely functional equiva-ence to the known macaque areas, as we will discuss in a laterection, arguments for homologies must be made with caution.ere, we will use an “h” prefix to denote the putative human
unctional equivalent of the respective macaque subregion inhe parietal lobe. We conclude with a general discussion of theontributions of neuroimaging to the understanding of the neu-al substrates underlying visuomotor control, particularly in thearietal cortex. We will also discuss the remaining challengesnd promising future directions.
. Eye movements
.1. Macaque neurophysiological research
Neurons in macaque LIP are activated when a monkey plansn abrupt eye movement, a saccade, to a location in the receptiveeld and also when the monkey attends to that location withoutxecuting a saccade (Colby, 1998; Colby, Duhamel, & Goldberg,996). LIP codes space in retinotopic coordinates (Andersen,racewell, Barash, Gnadt, & Fogassi, 1990; Andersen, Essick,Siegel, 1985; Cohen & Andersen, 2002), and interestingly,
eurons in LIP show spatial updating. That is, the receptiveelds of LIP neurons change in anticipation of an upcoming eyeovement (Duhamel, Colby, & Goldberg, 1992). This is thought
o be critical in the maintenance of a stable percept despite

2670 J.C. Culham et al. / Neuropsychologia 44 (2006) 2668–2684
Fig. 2. Human “functional equivalence” of monkey areas in IPS. Summary of activation and lesion sites identified by key studies in visuomotor control, listed inTable 1, shown for the left hemisphere from postero-lateral (a), superior (b) and medial (c) views. Foci were based on reported Talairach coordinates transformed tos le subW e a ms
fp
2
mem
ls1
2
urface locations on one subject’s pial surface. When studies provided both singhen possible, foci are shown in the three different brain views in order to provid
ulci, as described in Fig. 1.
requent changes of eye position resulting from several saccadeser second.
.2. Human neuropsychological research
Patients with bilateral parietal damage may suffer eye move-ent problems, such as gaze paralysis, difficulty keeping the
yes fixed on a target, difficulties with smooth pursuit eyeovements or vergence and accommodation. Even unilateral
hi
ject and averaged group coordinates, we only report the averaged coordinates.ore detailed description of each location. The white lines highlight the principal
esions can cause a slowing of eye movements in the contrale-ional direction (Pierrot-Deseilligny, Rivaud, Gaymard, & Agid,991).
.3. Human neuroimaging research
Eye movements are the visuomotor function in humans thatave been studied in the greatest detail using both neuroimag-ng and transcranial magnetic stimulation (TMS). The wealth

J.C. Culham et al. / Neuropsychologia 44 (2006) 2668–2684 2671
Table 1Talairach coordinates for the fMRI activation and the lesion foci shown in Fig. 2
Anatomical location Study Stereotaxic coordinates (Talaraich space)
X Y Z
hPEF (1) Petit and Haxby (1999) −30 −58 45(2) Sereno et al. (2001) −32 −68 46(3) Medendorp et al. (2003) −19 −58 47
hVIP (4) Bremmer et al. (2001) −40 −40 42
PCu (5) Astafiev et al. (2003) a −7 −79 32(6) Connolly et al. (2003) −1 −74 38(x) Karnath and Perenin (2005) b −6 −64 48
POJ (8) Prado et al. (2005) −16 −74 44SOG (x) Karnath and Perenin (2005) b −31 −79 32SPL (x) Karnath and Perenin (2005) b −29 −65 52
mIPS (7) Prado et al. (2005) −22 −52 66(9) Grefkes et al. (2004) a −25 −52 43
hAIP (10) Binkofski et al. (1998) −45 −35 43(11) Culham et al. (2003) −38 −48 52(12) Frey et al. (2005) −40 −33 43
h cIPS (13) Shikata et al. (2001) a −15 −69 46(14) James et al. (2002) −17 −72 36(15) Shikata et al. (2003) a −9 −76 43
a icalm
acros
ooamsbarMh
vCataa(&tfIat2ts2pt
sip
3
3
od1stm(mets1utdev
Talairach coordinates have been translated from the Montreal Neurolognispace.shtml).b Talairach coordinates from Karnath and Perenin (2005) have been averaged
f human neuroimaging studies on eye movements is basedn the rich eye movement literature using other techniquesnd is facilitated by the relative ease of studying eye move-ents compared to other visuomotor functions. Eye movement
tudies simply require projection screens, goggles or LEDs com-ined with MR-compatible eye trackers, which are now widelyvailable. This literature, including neuroimaging results, hasecently been reviewed in detail elsewhere (Pierrot-Deseilligny,
ilea, & Muri, 2004) so we provide only a brief summaryere.
Within the parietal cortex, saccadic eye movements acti-ate the parietal eye fields (PEF) (Muri, Iba-Zizen, Derosier,abanis, & Pierrot-Deseiligny, 1996). The human PEF (hPEF)re thought to be homologous with macaque LIP and the twoerms (PEF and LIP) are often used interchangeably. The hPEFre located midway along the anterior–posterior axis of the IPSnd activation is typically found medial to the IPS, in the SPLMedendorp, Goltz, Vilis, & Crawford, 2003; Sereno, Pitzalis,
Martinez, 2001). Even in comparisons using the same task,he location of saccade-related activation within the IPS dif-ers between the two species, being in the lateral bank of thePS in macaques and medial to the IPS in humans (Koyama etl., 2004). Evidence for spatial updating has been observed inhe hPEF (Medendorp, Goltz, & Vilis, 2005; Medendorp et al.,003; Merriam, Genovese, & Colby, 2003), which contains aopographic representation in retinotopic coordinates for both
accadic eye movements and peripheral attention (Sereno et al.,001). Smooth pursuit eye movements activate a similar fronto-arietal network, including the hPEF (Petit & Haxby, 1999) andhe PCu (Berman et al., 1999). This network of areas is highlyucpM
Institute (MNI) template (http://www.mrc-cbu.cam.ac.uk/Imaging/Common/
s multiple slices in their Fig. 5a. SOG, superior occipital gyrus.
imilar to that for peripheral attention (Corbetta, 1998), a find-ng that is consistent with the fact that attention shifts invariablyrecede saccades.
. Multimodal coding of near space
.1. Macaque neurophysiological research
In macaques, area VIP is involved with multimodal encodingf the space and motion around the head. Neurons in VIP areirection-selective for motion (Colby, Duhamel, & Goldberg,993) and the area receives strong input from the highly motion-elective middle temporal (MT) area. VIP neurons respond wello optic flow patterns and may play a role in determining move-
ent direction in head-centered coordinates during self-motionZhang, Heuer, & Britten, 2004). Many VIP neurons respondost strongly to stimuli in near space close to the face (Colby
t al., 1993). Many neurons also respond whenever an incomingarget approaches a particular part of the face, regardless of thepecific trajectory (for a review, see Colby, 1998; Colby et al.,993). VIP can be driven by visual, tactile, or auditory stim-li (Bremmer, Schlack, Duhamel, Graf, & Fink, 2001). Indeed,he receptive fields of VIP neurons are often congruent acrossifferent modalities (Duhamel, Colby, & Goldberg, 1998). Forxample, a neuron with a visual receptive field in the upper leftisual quadrant might also have a tactile receptive field in the
pper left part of the face. VIP receives inputs from vestibularortex (Bremmer, Schlack, Shah et al., 2001) and projects toremotor area F4, which controls the neck muscles (Luppino,urata, Govoni, & Matelli, 1999). Electrical stimulation of VIP,
2 ychol
ashCVga
3
ammte(pPw&wF
3
breeMtcfTm
hfew2ostarptilrostp
4
4
wmPiB&a11p(2(
4
a1efssbdtesrffp
4
mspccPbmtoat
672 J.C. Culham et al. / Neurops
s well as air puffs to the face, lead to defensive movementsuch as head withdrawal, the raising of the arms to protect theead, grimacing and eye closure (Cooke & Graziano, 2003;ooke, Taylor, Moore, & Graziano, 2003). Various theories ofIP function have been suggested including multimodal inte-ration, determination of heading direction during locomotion,nd avoidance of obstacles and projectiles (Cooke et al., 2003).
.2. Human neuropsychological research
Given the importance of near space, where the body inter-cts with the environment (Previc, 1998), and the existence ofacaque brain regions that respond only to near targets, weight also expect human neuropsychological deficits specific
o peripersonal space. Indeed, patients with extinction may onlyxhibit the phenomenon when targets are presented in near spaceLadavas, 2002; Landis, 2000). Moreover, extinction in theseatients can occur between different modalities (Ladavas, diellegrino, Farne, & Zeloni, 1998) and can be modulated byhether the finger is near the location of the target (di PellegrinoFrassinetti, 2000). Interestingly, “near space” can be extendedhen the patient is given a tool to use (Berti & Frassinetti, 2000;arne & Ladavas, 2000; Ladavas, 2002).
.3. Human neuroimaging research
A human area functionally similar to VIP (hVIP) woulde expected to demonstrate motion selectivity, multimodalesponses for visual, tactile and auditory stimuli, and a pref-rence for stimuli in near or ultra-near space. At least six pari-tal areas are activated by visual motion (Sunaert, Van Hecke,archal, & Orban, 2000), so additional criteria are necessary
o narrow down the number of candidate regions. Bremmer andolleagues have suggested a possible candidate for hVIP in theundus of the human IPS (Bremmer, Schlack, Shah et al., 2001).his area was activated bilaterally by visual, tactile and auditoryotion, as would be expected from the neurophysiological data.Other studies investigating VIP-like response properties in
uman neuroimaging have had mixed results. Unpublished datarom our group found somewhat inconsistent activation in pari-tal cortex when visual stimulation, tactile stimulation, or bothas provided by a sphere looming toward the face (Goltz et al.,001). We also examined activation for looming and recedingbjects in near space versus far space (using stimuli equated forize and velocity, Quinlan, Goodale, & Culham, 2004). The con-rast of motion in near space versus far space did not yield anyctivation near hVIP, although it did show activation in the supe-ior parieto-occipital sulcus (as did simply verging the eyes on aoint in near space versus far space). One group has found thatasks in near space (versus far space) activate several regions,ncluding the left intraparietal sulcus (Weiss et al., 2000). Pre-iminary data from Huang and Sereno (2005) indicates that aegion of the postcentral sulcus responds to videos of looming
bjects and responds to tactile stimulation of the contralateralide of the face. In sum, although there are some suggestionshat a parietal area is activated by multimodal motion and has areference for near space, the results have been mixed.temt
ogia 44 (2006) 2668–2684
. Reaching and pointing actions
.1. Macaque neurophysiological research
The role of the posterior parietal cortex in reaching has beenell researched with single-unit recording studies in macaqueonkeys. This work has shown that various subdivisions of thePC contain cells that are selective for reaching movements,
ncluding the medial intraparietal area (MIP) (Johnson, Ferraina,ianchi, & Caminiti, 1996), area V6a (Fattori, Gamberini, Kutz,Galletti, 2001; Galletti, Fattori, Kutz, & Battaglini, 1997),
reas 7a and 7m (Battaglia-Mayer et al., 2000; Johnson et al.,996; MacKay, 1992) and area 5 (Ferraina et al., 2001; Kalaska,996). Some laboratories use functional properties to define aarietal reach region (PRR) that includes both MIP and V6aAndersen & Buneo, 2002; Buneo, Jarvis, Batista, & Andersen,002) and may also extend into the lateral bank of the IPSCalton, Dickinson, & Snyder, 2002).
.2. Human neuropsychological research
Patients with damage to parietal cortex can suffer from optictaxia, a deficit in visually guided hand movements (Balint,909). Misreaching occurs for actions directed to the periph-ral visual field, often only in the contralesional side, but notor targets in central vision. Patients may be able to accuratelyaccade to targets, indicating that the disorder is not merely aensory deficit, or to accurately reach to targets on their ownodies, indicating that the disorder is not merely a motor disor-er. In a classic paper, Perenin and Vighetto (1988) suggestedhat the common lesion site in patients with ataxia was in pari-tal cortex and included the IPS, often along with the SPL andometimes the IPL. Recently, Karnath and Perenin (2005) havee-examined the common focus in optic ataxia using new toolsor lesion analysis (Rorden & Karnath, 2004). They report threeoci, one in the PCu, one in the superior occipital gyrus near thearietal-occipito junction and one in the SPL.
.3. Human neuroimaging research
Numerous technical challenges make it difficult to study armovements with neuroimaging. As such, the topic has not been
tudied in nearly the same detail as eye movements. One keyroblem is the physical difficulty of reaching in the restrictedonfines of the imaging scanners. In addition, arm movementsan induce head movements, which are a problem in bothET and fMRI. With fMRI, the situation is even more difficultecause, even if the head remains perfectly still, the motion of aass as large as the arm can disrupt the magnetic field, leading
o severe artifacts (Culham, in press). For this reason, a varietyf tasks have been employed and there has been much confusionbout the terminology to describe them. Reaching per se refers tohe extension of the arm to touch a distant target. Pointing refers
o directing the index finger in the direction of the target withoutxtending the arm. Because of the moving mass-related artifacts,any fMRI studies have employed pointing rather than reachingasks. Even within “reaching” tasks, various studies have sub-

ychol
ji(a2tbtml
t(1K1svawj
rIvo
(oZspoiA
l2drPiaiG
s(r2daaolbe
hthgfi
5
cos
5
irstgc“
tiaLMadiSM&Mtpmdprc
rtgnpihneurons are selective for only one particular sub-phase of grasp-
J.C. Culham et al. / Neurops
ect reaching-to-point (e.g., reaching-to-touch an object with thendex finger, as in Kawashima et al., 1996), reaching-to-touche.g., reaching-to-touch an object imprecisely with the knuckles,s in Culham et al., 2003), or reaching-to-grasp (Chapman et al.,002). We and others hypothesize that these tasks may differ inheir demands; specifically, reaching-to-point tasks may requireoth preshaping of the hand and computation of object proper-ies (such as the centroid for large targets), and thus may involve
ore of the substrates for grasping (hAIP, discussed later) thaness precise reaching-to-touch tasks (Kawashima et al., 1996).
A number of early PET studies reported parietal activa-ion contralateral to the hand used for reaching-to-point actionsClower et al., 1996; Decety, Kawashima, Gulyas, & Roland,992; Grafton, Fagg, Woods, & Arbib, 1996; Inoue et al., 1998;awashima et al., 1996; Kertzman, Schwarz, Zeffiro, & Hallett,997). However, the PET results are often hard to interpret foreveral reasons. Often these studies have used contrasts betweenery different tasks (e.g., reaching versus passive fixation) thatctivate a large list of areas throughout the brain, sometimesith poor anatomical localization (due to smoothing, intersub-
ect averaging and/or poor structural images).fMRI studies of pointing (without a reach component) have
eported activation in both the medial IPS (mIPS) and the PCu.nterestingly, the results of human neuroimaging appear to con-erge well with the neuropsychological lesion overlap analysesf Karnath and Perenin (2005) (see Fig. 2 and Table 1).
One group using fMRI has suggested that a region in the mIPSjust posterior to the PCS) may be a human functional equivalentf macaque MIP, hMIP (Grefkes & Fink, 2005; Grefkes, Ritzl,illes, & Fink, 2004). They found activation in this region whenubjects used a joystick to move a cursor to a visual target (com-ared to a visuomotor control task). Although the task was notne of reaching or pointing, a similar paradigm has been usedn the macaque to demonstrate responses in MIP (Eskandar &ssad, 1999, 2002).The PCu focus has been suggested as hPRR, a putative homo-
ogue of the monkey PRR (Connolly, Andersen, & Goodale,003), although there are problems with this nomenclature (to beiscussed later). This region is activated during pointing prepa-ation (Connolly et al., 2003), as is macaque PRR, althoughCu activation for pointing execution is higher than for point-
ng preparation (Astafiev et al., 2003). In addition, the region isctivated more by anti-pointing (pointing to a location in the hor-zontal mirror plane of the target) than pro-pointing (Connolly,oodale, Desouza, Menon, & Vilis, 2000).A elegant new fMRI paper utilized a true reaching task and
uggested that mIPS and a region at the parieto-occipital junctionPOJ), slightly lateral to the PCu, may serve different roles ineaching (Prado, Clavagnier, Otzenberger, Scheiber, & Perenin,005). Prado and colleagues found that the mIPS was activateduring a reach-to-point task regardless of whether the targetppeared in central or peripheral vision. By contrast, the POJ wasctivated when the reach was made either to a peripheral target
r to a target that disappeared before a saccade was made to itsocation but not when the target remained visible and a saccaderought it into central vision. They propose that these resultsxplain neuropsychological findings of optic ataxic patients whoi(GR
ogia 44 (2006) 2668–2684 2673
ave deficits when reaching to peripheral or absent targets but noto visible targets following a saccade; presumably, such patientsave lesions to the parieto-occipital junction. These results sug-est that reach-related activation depends on whether gaze isxed or not.
. Grasping actions
A recent review paper covers a range of neurophysiologi-al, neuropsychological, behavioral and neuroimaging researchn grasping (Castiello, 2005). Nevertheless, we provide a briefummary here.
.1. Macaque neurophysiological research
The task of picking up an object typically requires a reach-ng movement to transport the hand to the object; in addition, itequires extraction of the visual features of the object (such ashape, size and orientation) in order to appropriately preshapehe hand prior to contact. These two aspects are the transport andrip components, respectively. Such movements should techni-ally be called “reach-to-grasp” actions, though the shorter termgrasping” is commonly employed.
The first insight about neural substrates for the visual con-rol of hand movements came from electrophysiological studiesn macaque monkeys, showing “hand-manipulation” neurons inrea 7 of the IPL (Hyvarinen & Poranen, 1974; Mountcastle,ynch, Georgopoulos, Sakata, & Acuna, 1975). For example,ountcastle et al. (1975) observed parietal neurons that were
ctive when the monkey actively manipulated an object, but noturing passive stimulation of the arms or hand. Electrophys-ological (Murata, Gallese, Luppino, Kaseda, & Sakata, 2000;akata, Taira, Kusunoki, Murata, & Tanaka, 1997; Sakata, Taira,ine, & Murata, 1992; Taira, Mine, Georgopoulos, Murata,Sakata, 1990) and functional inactivation studies (Gallese,
urata, Kaseda, Niki, & Sakata, 1994) revealed that neurons inhe anterior intraparietal (AIP) sulcus play a key role in handreshaping for grasping. Neurons in AIP fire when the monkeyanipulates a particular object, with different neurons tuned to
ifferent objects (Murata et al., 2000) regardless of the object’sosition (Taira et al., 1990). Moreover, inactivation of AIP dis-upts the monkey’s ability to use vision for preshaping of theontralateral hand (Gallese et al., 1994).
We now know that subpopulations of neurons in area AIPespond to: (i) a grasping movement toward an object whenhe stimuli and the hand are visible (visuomotor neurons); (ii) arasping movement in the dark when the stimuli and the hand areot visible (motor dominant neurons) and (iii) the simple visualresentation of three-dimensional (3D) objects when no actions required (visual-dominant neurons). In addition, other studiesave shown that when grasping is executed in the light, some AIP
ng such as approaching, contact, or closure around the objectDebowy, Ghosh, Ro, & Gardner, 2001; Gardner, Debowy, Ro,hosh, and Babu, 2002; Gardner, Ro, Debowy, & Ghosh, 1999;o, Debowy, Ghosh, & Gardner, 2000).

2 ychol
5
in(deds
5
ihtaCFVvv(
i1AlKaa2oKrafw
tio2oMeswnAdSpwg&
ha
ghb(KsbIst&2
tiacetfO&So2oeItet1ertKbwcgrtuat
ri2o
674 J.C. Culham et al. / Neurops
.2. Human neuropsychological research
Optic ataxia can include deficits in grasping as well as reach-ng; the majority of these deficits relate to the transport compo-ents of reaching rather than the grip components of graspingJeannerod, Decety, & Michel, 1994). Binkofski et al. (1998)emonstrated that patients with lesions in the anterior intrapari-tal sulcus were more impaired at grasping than reaching. Theseeficits were far more pronounced for grasping with the contrale-ional hand than the ipsilesional hand.
.3. Human neuroimaging research
After almost a decade from the first grasping study in imag-ng (Grafton et al., 1996), consistent results suggest that theomologue of macaque AIP in humans is located at the junc-ion between the anterior portion of the intraparietal sulcusnd the inferior postcentral sulcus (Binkofski et al., 1998;ulham, Cavina Pratesi, & Goodale, 2004; Culham et al., 2003;aillenot, Sakata, Costes, Decety, & Jeannerod, 1997; Frey,inton, Norlund, & Grafton, 2005). This area is also acti-ated by reaching and appears to overlap with the IPS acti-ation reported during some pointing and reach-to-point tasksdescribed above).
Consistent with the range of responses in macaque AIP, hAIPs activated by (i) visually guided grasping (Binkofski et al.,998; Culham et al., 2003; Frey et al., 2005; Grezes, Tucker,rmony, Ellis, & Passingham, 2003); (ii) the act of hand manipu-
ation when vision is unavailable (Binkofski et al., 1999; Jancke,leinschmidt, Mirzazade, Shah, & Freund, 2001; Stoeckel et
l., 2003) and (iii) the presentation of visual stimuli withoutn action (Culham, Cavina Pratesi et al., 2004; Grezes et al.,003; Peuskens et al., 2004; Shikata et al., 2003, 2001). More-ver, two delayed grasping studies (Culham, 2003; Singhal,aufman, Valyear, & Culham, 2005) have shown that hAIP
esponds during both the stimulus presentation and the handction. In addition, activation in the vicinity of hAIP is strongeror object grasping than for arbitrary hand gestures associatedith objects (Toni, Rushworth, & Passingham, 2001).We have argued elsewhere that, although hAIP is fundamental
o the computation of grip during visually guided grasping, its not necessarily activated by tasks that require processing ofbject features for purely perceptual purposes (Culham et al.,003). hAIP is not activated by a contrast between 2D pictures ofbjects and their scrambled counterparts (Culham et al., 2003).oreover, a recent study from our lab (Culham, Cavina Pratesi
t al., 2004) used real, graspable, 3D objects that varied in size tohow that hAIP was activated during grasping (versus reaching)hen size information is necessary for hand preshaping, butot during a perceptual discrimination of size (versus pattern).lthough 2D pictures in general do not activate hAIP, they mayo so when the images have rich associations with hand actions.pecifically, 2D pictures of tools activate a region in the anterior
arietal cortex (Chao & Martin, 2000) that overlaps partiallyith hAIP (Culham, Valyear, & Stiglick, 2004). Tools lead toreater activation than other graspable objects (Creem-RegehrLee, 2005; Culham, Valyear et al., 2004), likely because they
Fesr
ogia 44 (2006) 2668–2684
ave much stronger affordances for a complex series of handctions.
Although we have emphasized the role of hAIP in visuallyuided grasping, it is likely also involved in haptic processing.AIP is activated for the precise and gentle grip force exertedetween the index and the thumb when no vision is availableEhrsson et al., 2000; Ehrsson, Fagergren, & Forssberg, 2001;uhtz-Buschbeck, Ehrsson, & Forssberg, 2001), suggesting a
pecific role of the motor-dominant neurons in the fine cali-ration of finger position and grip force during object lifting.n addition, hAIP has been activated in cross-modal matchingtudies in which the subjects had to transfer object informa-ion between visual and haptic domains (Grefkes, Weiss, Zilles,
Fink, 2002; Tanabe, Kato, Miyauchi, Hayashi, & Yanagida,005).
Functional imaging studies have focused on area hAIP, buthey have also shown that it does not work alone. Two stud-es from our group showed the existence of other parietal areasccompanying AIP activation when visually guided grasping isompared to visually guided reaching (Culham, Cavina Pratesit al., 2004; Culham et al., 2003). One area was in the horizon-al segment of IPS where similar activation has been reportedor 3D feature processing (Faillenot, Sunaert, Van Hecke, &rban, 2001; Shikata et al., 2003, 2001; Taira, Nose, Inoue,Tsutsui, 2001; Tsutsui, Sakata, Naganuma, & Taira, 2002).
ome have suggested that the region may be a homologuef macaque cIPS (Faillenot et al., 2001; Shikata et al., 2003,001; Taira et al., 2001; Tsutsui et al., 2002), which containsrientation-selective inputs and has projections to AIP (Sakatat al., 1997). Another possibility is that horizontal segment ofPS corresponds to human LIP (Sereno et al., 2001), which inhe macaque brain relays inputs from V3A to AIP (Nakamurat al., 2001). Sometimes, grasping also produces activation inhe PCu (Culham, Cavina Pratesi et al., 2004; Faillenot et al.,997), near the sites involved in pointing preparation (Astafievt al., 2003; Connolly et al., 2003). Indeed, recent physiologicalesearch suggests that area macaque V6A, which forms part ofhe PRR, is involved in grasping as well as reaching (Galletti,utz, Gamberini, Breveglieri, & Fattori, 2003). Despite the num-er of parietal regions activated by grasping (versus reaching),e believe that the area suggested as hAIP is the most likely
andidate for a functional equivalent to macaque AIP. This sug-estion comes from several sources: its necessity for accurateeaching in patient studies (Binkofski et al., 1998), the consis-ency of its activation across many protocols from many labssing visually guided actions (for a review, see Frey et al., 2005),nd the similarity of both the location and the response proper-ies to macaque AIP.
To address the issue of lateralization of reach- and grasp-elated activation, we conducted an fMRI experiment with reach-ng and grasping using the left and right hands (Culham et al.,001). Seven right-handed subjects performed either reachingr grasping to visual targets using a slow event-related design.
ull methods are described in greater detail elsewhere (Culhamt al., 2003). Due to the physical constraints of the arm brace,ubjects performed separate runs using either the left hand or theight hand. The results are shown in Fig. 3. Note that grasping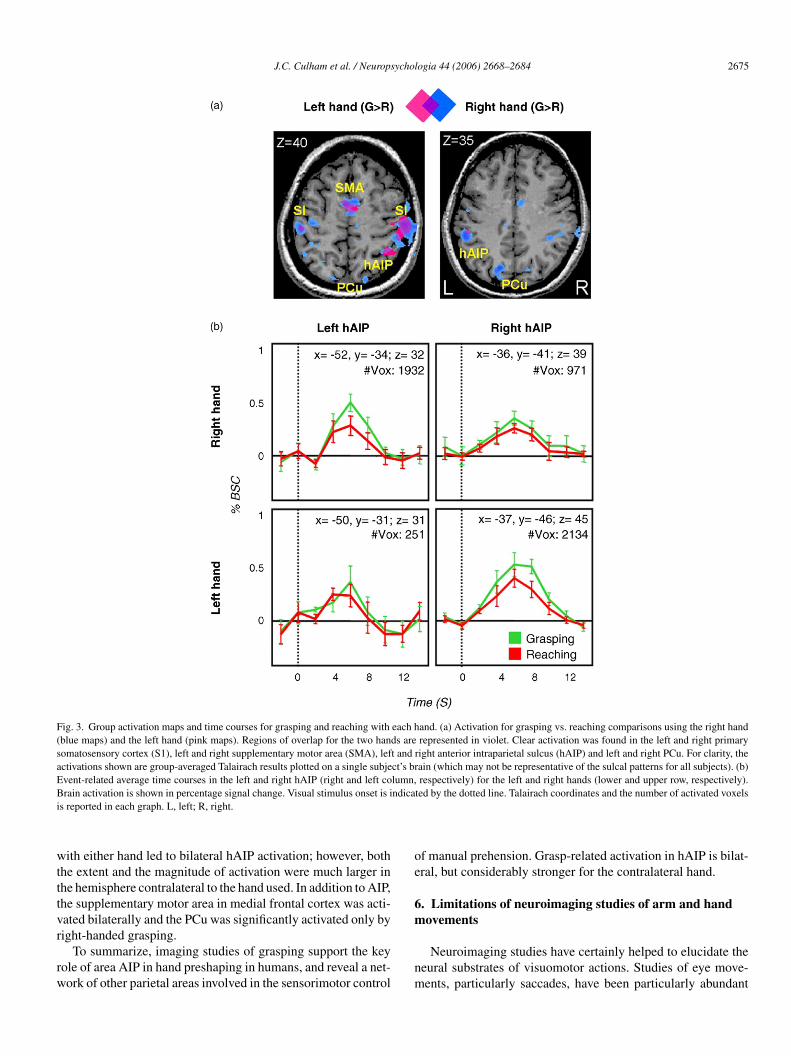
J.C. Culham et al. / Neuropsychologia 44 (2006) 2668–2684 2675
Fig. 3. Group activation maps and time courses for grasping and reaching with each hand. (a) Activation for grasping vs. reaching comparisons using the right hand(blue maps) and the left hand (pink maps). Regions of overlap for the two hands are represented in violet. Clear activation was found in the left and right primarysomatosensory cortex (S1), left and right supplementary motor area (SMA), left and right anterior intraparietal sulcus (hAIP) and left and right PCu. For clarity, theactivations shown are group-averaged Talairach results plotted on a single subject’s brain (which may not be representative of the sulcal patterns for all subjects). (b)Event-related average time courses in the left and right hAIP (right and left column, respectively) for the left and right hands (lower and upper row, respectively).B ndicai
wtttvr
rw
oe
6m
rain activation is shown in percentage signal change. Visual stimulus onset is is reported in each graph. L, left; R, right.
ith either hand led to bilateral hAIP activation; however, bothhe extent and the magnitude of activation were much larger inhe hemisphere contralateral to the hand used. In addition to AIP,he supplementary motor area in medial frontal cortex was acti-ated bilaterally and the PCu was significantly activated only by
ight-handed grasping.To summarize, imaging studies of grasping support the keyole of area AIP in hand preshaping in humans, and reveal a net-ork of other parietal areas involved in the sensorimotor control
nm
ted by the dotted line. Talairach coordinates and the number of activated voxels
f manual prehension. Grasp-related activation in hAIP is bilat-ral, but considerably stronger for the contralateral hand.
. Limitations of neuroimaging studies of arm and handovements
Neuroimaging studies have certainly helped to elucidate theeural substrates of visuomotor actions. Studies of eye move-ents, particularly saccades, have been particularly abundant

2 ychol
ab
murnett“ipnwEkfsd2ng&tdetl
diva2mcaiivohutt
cotCahadrp
e“mas1(vot(Sttiw
7v
twlrtwtttlher
7
cich(Hmtrhmawca
676 J.C. Culham et al. / Neurops
nd fruitful. Studies of arm and hand movements, however, haveeen rather limited.
Because of the technical challenges of studying arm and handovements, many laboratories have taken to studying rather
nnatural tasks. As discussed earlier, rather than studying trueeaching, many fMRI studies have opted to study pointing (witho arm transport) instead. Pointing, however, may not be ancologically valid visuomotor task. In humans at least, pointingends to serve a communicative function rather than a visuomo-or function. For example, one might point to a cup to indicateThat is my cup”, but this is functionally quite different fromnteracting with the cup as one would to push it out of the way orick it up. Given that pointing is not a task monkeys do readily,europhysiological studies have not tested pointing nor shownhether it relies on the same neuronal substrates as reaching.ven when reaching per se is studied, in both humans and mon-eys, it often involves moving the hand between locations in therontoparallel plane (e.g., Andersen & Buneo, 2002) rather thantarting with the hand near the body and moving it outward inepth as typically happens in everyday actions (Galletti et al.,003). Because of the difficulty of studying real grasping in theeuroimaging environment, some papers have used pantomimedrasping (Shikata et al., 2003; Simon, Mangin, Cohen, Le Bihan,
Dehaene, 2002). In our lab, we have tried artificial graspingasks that involve imagery of grasping (Culham et al., 2001) orecisions about how objects could be grasped (Culham, Valyeart al., 2004); however, the results remain unpublished becausehe activations were weak, inconsistent across subjects, and faress robust than for real actions.
Viewing conditions should be another consideration inesigning tasks to study arm and hand actions with neuroimag-ng. Some have subjects perform actions on real objects whileiewing two-dimensional pictures of the same objects throughmirror (Grefkes et al., 2002; Grezes et al., 2003; Jancke et al.,001; Shikata et al., 2003). Other studies on “reaching” involveoving a cursor with a joystick (Lacquaniti et al., 1997). Opti-
al manipulations such as viewing the hand workspace throughprism (Clower et al., 1996) or mirror (Binkofski et al., 2003)
nduce additional activation in PPC (Clower et al., 1996), includ-ng hAIP (Binkofski et al., 2003). Thus, where possible, directiewing is preferable for the study of naturalistic actions, with-ut artificial transformations. Reassuringly, some areas, such asAIP appear to show similar properties in studies that did notse mirror-viewing viewing (Binkofski, personal communica-ion; Binkofski et al., 1998; Culham et al., 2003) as well as inhose that did (Frey, personal communication; Frey et al., 2005).
Although neuroimaging of arm and hand actions providehallenges, they are not insurmountable. Over many years inur lab, we have developed several novel solutions to optimizehe setup as much as possible (described in greater detail inulham et al., 2003). First, we tilt the subjects’ heads as muchs possible within a head coil to allow direct viewing of theand workspace slightly below eye fixation (without employing
mirror). In scanners that do not have corrections for spatialistortions, custom corrections may be required if the head isaised above magnet isocentre (Quinlan et al., 2005). Second, torovide a wide variety of objects in a small workspace, we
thad
ogia 44 (2006) 2668–2684
mploy a pneumatically driven rotating drum (known as thegrasparatus”) with objects mounted on the surface. Third, weinimize head motion by restricting the movement of the upper
rm using an arm brace. Fourth, and most importantly, we use alow event-related paradigm (Birn, Bandettini, Cox, & Shaker,999) to dissociate artifacts from head and arm movementswhich occur simultaneously with the action) and genuine acti-ation (which occurs with the characteristic hemodynamic lagf several seconds). Fifth, we are currently exploring equipmento collect kinematic data using a fiber optic tracking deviceShapeClaw, Measurand Inc., Fredericton, New Brunswick).ixth, we are also developing a setup where the magnet bed
ray is removed such that the body sits lower in the bore suchhat when the head is tilted, it will remain close to the magnetsocentre and the eyes can gaze directly at the hand workspaceithout eye strain.
. What have we learned from neuroimaging ofisuomotor control?
Of course, the greatest contribution of neuroimaging has beeno investigate the normal human brain, which is not possibleith invasive electrophysiological techniques or neuropsycho-
ogical studies. Functional MRI has provided human data at aelatively high spatial resolution and has allowed the identifica-ion of regions smaller than those typically damaged in patientsith strokes or lesions. As previously described, we have seen
hat neuroimaging has been useful at elucidating activation focihat, while somewhat variable, are beginning to show a consis-ent pattern across multiple studies from multiple labs. For ateast four areas (hPEF, hVIP, hPRR and hAIP), neuroimagingas suggested similarities between the layout of the human pari-tal cortex and that of the macaque monkey. What challengesemain?
.1. Lateralization of parietal function
A rich literature, largely based on behavioral and neuropsy-hological data, suggests a high degree of hemispheric special-zation within the parietal cortex. In general, the right parietalortex is specialized for spatial processing and attention, whichas often been described in neglect and extinction patientsHalligan, Fink, Marshall, & Vallar, 2003; Karnath, Ferber, &immelbach, 2001; Snyder & Chatterjee, 2004). Of course, theotor and somatosensory representations of the body, including
he hands is primarily contralateral (Kolb & Whishaw, 2003). Inight-handed subjects, many functions related to control of theands are also lateralized to left parietal cortex. In particular,any aspects of praxis (including the imitation, pantomiming
nd sequencing of gestures and tool usage) are compromisedith left parietal lesions (Leiguarda & Marsden, 2000), as are the
alculation of numerical quantities (Dehaene & Cohen, 1995),nd writing (Balasubramanian, 2005). However, it is important
o note that brain organization may differ dramatically based onandedness (Harshman, Hampson, & Berenbaum, 1983; Kim etl., 1993) and some functions such as praxis may be organizedifferently in males than females (Kimura, 1983).
ychol
cnbSmsitvv
tbpts2a
anvhtTdnbPwtaDCn
tis(dtorKecsrI
i(ihl
whirbGubf
sdaetahsbss2haramJap2mtpiiwtaBoeetig
7
gIi
J.C. Culham et al. / Neurops
In neuroimaging, the degree of lateralization has been lesslear and not always in concert with evidence from other tech-iques. The right parietal cortex specialization for attention haseen shown to be robust in many studies (e.g., Corbetta, Miezin,hulman, & Petersen, 1993; Gitelman et al., 1999), althoughore recent work suggests that there may be left hemisphere
pecialization for some attentional processes involving verbalnformation (Shulman, d’Avossa, Tansy, & Corbetta, 2002). Fur-hermore, the lateralization of visual processing, with the leftisual field being processed in the right hemisphere and viceersa, has been well established with fMRI.
In recent years, an increasing number of studies have shownhat topographically mapped visual areas can be found welleyond the classical areas (such as V1, V2, V3, V3a and V4),articularly when the stimuli are well suited to the properties ofhe region. In parietal cortex, hPEF shows a topographic repre-entation of the location of saccades and attention (Sereno et al.,001), and other parietal areas show a similar topography forttentional processes (Silver, Ress, & Heeger, 2005).
In terms of hand actions, reach- and grasp-related processingppears to be largely driven by the contralateral hemisphere, yeteuroimaging results are equivocal and often show bilateral acti-ation. Macaques with AIP inactivation (Gallese et al., 1994),uman patients with lesions to hAIP (Binkofski et al., 1998) orhe PCu (Karnath & Perenin, 2005), and normal humans withMS to PPC cortex (Desmurget et al., 1999) have all shown pre-ominantly contralateral deficits in reaching and grasping. Theeuroimaging literature is mixed. Some studies have reportedilateral hAIP activation (Binkofski et al., 1998; Culham, Cavinaratesi et al., 2004; Culham et al., 2003; Grefkes et al., 2002),hereas other studies have suggested only contralateral activa-
ion for reaching and grasping (Desmurget et al., 2001; Frey etl., 2005). However, with a few exceptions (Culham et al., 2001;eSouza et al., 2000; Kertzman et al., 1997; Medendorp, Goltz,rawford, & Vilis, 2005) neuroimaging studies have predomi-antly studied right-handed actions.
In addition to issues of an effect of the hand used for actions,here may also be an effect of the visual field in which the actions directed. Based on patient data, Perenin and Vighetto (1988)uggested that left hemisphere damage led to both a hand effecta greater deficit with the right hand) and a field effect (a greatereficit in the right field), whereas right hemisphere damage ledo a field effect only (a greater deficit in the left field). However,thers have found that a hand effect can also be observed withight hemisphere damage (Levine, Kaufman, & Mohr, 1978).ertzman et al. (1997) used PET to examine hand and field
ffects during a visually guided reaching task. Within parietalortex, they reported that the postcentral gyrus and SPL/IPShowed predominantly contralateral hand effects. Activation toeaching in one visual field was typically bilateral, except in thePL where a contralateral field preference was found.
Other studies have focused on the relationship between point-ng movements and eye position. An area in the rostral IPS
likely corresponding to hAIP) was most activated by point-ng with the contralateral hand to targets in the contralateralemifield (DeSouza et al., 2000). Even when the retinotopicocation of targets was held constant, the activation was highestco
f
ogia 44 (2006) 2668–2684 2677
hen the eyes were directed to the opposite field. Other studiesave suggested that other areas, including a retinotopic regionn the IPS (likely corresponding to the PEF) and a caudal IPSegion (in the vicinity of V3A and V7), also show preferences foroth the contralateral hand and contralateral field (Medendorp,oltz, Crawford et al., 2005). Furthermore, evidence for spatialpdating of target locations for pointing, as well as saccades, haseen reported in the hPEF, suggesting a gaze-centred coordinaterame (Medendorp et al., 2003).
It remains rather mysterious why neuroimaging showstronger bilateral activation for hand actions than would be pre-icted from other methods, particularly lesion studies. In ournalyses here (Fig. 3), we found that hAIP showed a bilat-ral response with higher activation over a larger extent whenhe contralateral hand was used. Given evidence that hAIPppears essential for control of the contra-but not ipsi-lateraland (Binkofski et al., 1998; Gallese et al., 1994), why do weee bilateral activation? One possibility is that there is cross-talketween corresponding areas within the brain (e.g., left hAIP istrongly interconnected with right hAIP). Given that the fMRIignal is thought to reflect synaptic inputs to a region (Logothetis,003), perhaps activation in ipsilateral hAIP arises from inter-emispheric inputs from the contralateral hAIP. If so, bilateralctivation may be observed even though only the contralateralegion is essential for the task. This could also explain whyctivation in somatosensory and motor paradigms tends to beore bilateral than may be expected (e.g., Jenkins, Jahanshahi,
ueptner, Passingham, & Brooks, 2000). However, one argumentgainst this possibility comes from the fact that tool namingaradigms yield robust activation in left PPC (Chao & Martin,000) including hAIP (Culham, Valyear et al., 2004), withoutuch activation in right PPC. A second possible explanation is
hat although subjects are instructed to use only one hand, theossibility to use either hand still exists. That is, when view-ng an object to be grasped, processing could occur bilaterallyn hAIP to evaluate the affordance of the object for graspingith each of the two hands. Even though only one hand is even-
ually selected (based on affordance or top down informationbout the rules of the task), both options may be considered.y this view, processing by both hemispheres would occur, butnly the contralateral hemisphere would be essential to the finalxecution of the action. This possibility could be addressed byxperiments manipulating the affordances of objects or the rela-ive utility of the two hands. In this view, field effects may arisen part because the location of an object in the contralateral fieldenerally affords an action with the contralateral hand.
.2. Caveats regarding suggested homologies
Neuroimaging has yielded suggestions of numerous homolo-ies between macaque and human brains (see Fig. 2 and Table 1).n some cases, such as AIP and PEF/LIP, there are numerous sim-larities between the regions in the two species and a moderate
ase can be made for a homology. In other areas, the suggestionf homologies is more tentative.The PRR is a source of particular confusion. In part, this arisesrom the neurophysiology literature, with various groups study-

2 ychol
iosamhe2ogAireHc
d(opihsaastbabtYtttrtnthostbst“taicfidci
ia2pc
raiitP&myit
bcFaoipaappitteh
idlSiparm1banrSe(w
678 J.C. Culham et al. / Neurops
ng MIP (Colby & Duhamel, 1991), V6A (Galletti et al., 2003)r a functionally defined PRR (Andersen & Buneo, 2002), whichubsumes MIP, V6A and sometimes adjacent cortex (Calton etl., 2002). Different groups have used neuroimaging data toake a variety of inconsistent interpretations. Specifically, it
as been proposed that mIPS corresponds to hMIP (Grefkest al., 2004), that PCu corresponds to hPRR (Connolly et al.,003), that mIPS corresponds to PRR (DeSouza et al., 2000),r that a larger region encompassing mIPS, PCu and POJ alto-ether form the hPRR (Astafiev et al., 2003; Prado et al., 2005).lthough the macaque PRR subsumes several adjacent anatom-
cal areas, the human brain contains several separate reach-elated foci (mIPS, PCu and POJ). It is possible that during thevolution of the human brain, the subregions of PRR diverged.owever, more comparative data is necessary to determine the
orrespondence.Certainly care is warranted in general while attempting to
raw parallels between regions of human and macaque brainsKrubitzer, 1995; Sereno & Tootell, 2005; Striedter, 2005). Asne reviewer put it, “It appears that getting spikes from someyramidal neurons in macaques, and BOLD from a few voxels,n roughly the same corner of the brain is sufficient to claimomology”. Clearly, such leaps are tenuous at best. First, inter-pecies comparisons typically involve differences in techniquess well as differences in species. Neurophysiological measuresre based on single neurons in a limited number of areas and areelected based on the ease of recording and subjective criteriahat can vary between laboratories. Neuroimaging studies areased on populations of neurons and the total response acrossvoxels containing millions of neurons depends on the num-
er of neurons tuned to a particular property, the profile of theuning function, and the vigour of the responses (Scannell &oung, 1999), as well as the dynamics of excitation and inhibi-
ion that are poorly understood at present. In addition, much ofhe temporal information about the response are lost because ofhe sluggishness and blurriness of the fMRI signal (which event-elated designs address, but only to a point). Second, althoughhe term “homology” tends to be used rather too loosely amongeuroimagers (ourselves included, Culham, 2003) to imply func-ional equivalencies between species, in evolutionary biology, aomology has a much stricter definition. Specifically, a homol-gy is a feature inherited from the common ancestor of twopecies. This is in contrast to a homoplasy, which is a similar fea-ure that evolved independently in two species. The distinctionetween homologies and homoplasies requires additional datauch as knowledge of which species in the evolutionary familyree share a common feature. Without such data, terms such asfunctionally equivalent area” may be more accurate, but evenhen, the degree of equivalence may be up for debate, and evenreas that are truly homologous may serve different functionsn different species. Even the term “area” requires some clarifi-ation. In its common use in neuroimaging, an area refers to aocus of functional activation, as does a “region”. In neurophys-
ology, however, the term area may have a much more restrictedefinition that includes multiple criteria, such as distinct cytoar-hitectonic properties and connectivity patterns. Despite thesessues, one has to start somewhere and it becomes very confus-tGr2
ogia 44 (2006) 2668–2684
ng to keep track of a literature where every lab has a differentcronym for each area. We and others (Culham & Kanwisher,001; Grefkes & Fink, 2005) have proposed tentative links torovide working hypotheses that remain to be examined morelosely in further experiments.
One trend that holds promise for elucidating interspecieselationships is the growing use of human cyto- and myelo-nd receptor architectonic mapping in both post mortem andn vivo brains (Eickhoff, Walters et al., 2005). Such stud-es have the potential to more objectively identify areas (inhe strict sense of the word) within the brain (Scheperjans,alomero-Gallagher, Grefkes, Schleicher, & Zilles, 2005; Zilles
Palomero-Gallagher, 2001). In addition, probabilistic areaaps have begun to be incorporated into neuroimaging anal-
sis software (Eickhoff, Stephan et al., 2005). Diffusion tensormaging and functional connectivity analyses may also aid inhe definition of area boundaries.
As arguments for functional equivalents accumulate, itecomes interesting to examine how the layout of areas on theortical sheet has changed with cortical expansion in humans.or example, the motion-selective middle temporal area (MTnd satellite areas; hMT+) has migrated inferiorly and posteri-rly and the frontal eye fields (hFEF) have migrated posteriorlyn humans with respect to macaques (Astafiev et al., 2003). Theroposed explanation is that the expansion of superior temporalnd prefrontal cortices has led to the displacement of pre-existingreas. It may be that displacement has also occurred in thearietal cortex, with the expansion of the regions in the IPLushing the areas involved in visuomotor control more mediallyn humans (Astafiev et al., 2003). The surface area of parietal cor-ex is 20× larger in humans relative to macaques (a ratio higherhan in temporal cortex, 9×, or occipital cortex, 2×, Van Essent al., 2001), indicating either that the areas within parietal cortexave become much larger or that new areas have been added.
One clear advantage of human neuroimaging over neurophys-ology is the ability to image functions that may be more/onlyeveloped in the human brain. For example, calculation andanguage functions have been localized to the human IPL (e.g.,imon et al., 2002). Interestingly, language-related activation
n Broca’s area overlaps with activation that occurs when peo-le observe others’ actions (Iacoboni et al., 1999; Rizzolatti etl., 1996). This has led to provocative theories regarding theelationship between “mirror neurons” in macaque ventral pre-otor cortex and the evolution of language (Rizzolatti & Arbib,
998). Recent evidence suggests that mirror responses can alsoe observed in parietal cortex of macaques (Fogassi et al., 2005)nd humans (Shmuelof & Zohary, 2005). The availability ofeuroimaging techniques has led to a plethora of papers on mir-or responses (for a review, see Rizzolatti & Craighero, 2004).ome very intriguing studies by Iriki and colleagues have usedlectrophysiology (Iriki, Tanaka, & Iwamura, 1996) and PETObayashi et al., 2001) in macaques to reveal parietal regions inhich coding of arm position can be modulated by the use of a
ool to extend the reach (for a review, see Maravita & Iriki, 2004).iven the greatly enhanced ability of humans to use tools, neu-
oimaging studies of tool use have proliferated (Johnson-Frey,004).

ychol
7
tarm
eiGCitavvgsiw
aaFV
nintstateothDwiMtrwNosv
iiteOS
Ehwoglick
nvRmmoMtp
mtcpfhpaabshtlna
A
cRrGPVeIgPJi
J.C. Culham et al. / Neurops
.3. Future directions
Although neuroimaging has already made significant con-ributions to understanding the neural substrates of visuomotorctions, advances in neuroimaging and the combination of neu-oimaging with other cognitive neuroscience techniques holduch promise.The development of more sophisticated designs in fMRI may
xtend the types of questions neuroimaging can ask. Specif-cally, fMR adaptation paradigms (Grill-Spector et al., 1999;rill-Spector & Malach, 2001) are being increasingly used.onventional fMRI cannot resolve activations smaller than an
maging voxel, which may contain millions of neurons; however,he logic of fMR adaptation can nevertheless allow inferencesbout the coding of specific subpopulations within a region (oroxel). For example, Shmuelof and Zohary (2005) presentedideos of a grasping hand and varied either the object to berasped or the grasp posture used. They found that hAIP waselective for both the grasp posture and the object to be grasped,n contrast to an object-selective area in the ventral stream, whichas selective to the object but not the posture.Although fMR adaptation may extend the questions neuroim-
gers can ask, thus far it is poorly understood and may reflectvariety of neural processes with different time scales (e.g.,
ang, Murray, Kersten, & He, 2005; Henson, 2003; Sawamura,ogels, & Orban, 2004).
Now that many basic functional areas have been defined witheuroimaging, its application to understanding neuropsycholog-cal cases is growing. We have already seen that anatomicaleuroimaging can be quite useful in narrowing down the lesionshat impair visuomotor functions (Karnath & Perenin, 2005). Inome cases, this approach has led to controversial suggestionshat challenge the historical interpretation of parietal function,s in the case of work indicating that neglect arises from lesionso superior temporal rather than inferior parietal cortex (Karnatht al., 2001). Functional neuroimaging has the added advantagef revealing spared regions that may subserve residual func-ions in neuropsychological patients. For example, our groupas demonstrated the utility of functional imaging with patientF (James, Culham, Humphrey, Milner, & Goodale, 2003),ho has impaired object recognition but spared object grasp-
ng (Goodale, Milner, Jakobson, & Carey, 1991). Goodale andilner’s (1992) two streams theory provided a coherent explana-
ion for the dissociation, but was vague about specific anatomicalegions. Functional neuroimaging identified the specific regionsithin the streams that are likely to account for the dissociation.amely, we found that DF had lesions corresponding to thebject-selective lateral occipital cortex in the ventral stream buthe nevertheless showed normal activation in hAIP for graspingersus reaching.
With the growing development of macaque fMRI, it is becom-ng increasingly common to run identical paradigms using fMRIn both humans and macaques, which should greatly facili-
ate interspecies comparisons (e.g., Denys et al., 2004; Koyamat al., 2004; Nakahara, Hayashi, Konishi, & Miyashita, 2002;rban, Van Essen, & Vanduffel, 2004; Vanduffel et al., 2001).avaki and colleagues (Gregoriou & Savaki, 2001, 2003; Raos,R
A
ogia 44 (2006) 2668–2684 2679
vangeliou, & Savaki, 2004; Savaki, Raos, & Dalezios, 1997)ave used autoradiography to examine glucose consumptionith high spatial resolution. They have described subregionsf parietal cortex which are activated by saccades, reaching,rasping and observation of grasping. Although neurophysio-ogical results led to hypotheses for neuroimaging experimentsn the early days of neuroimaging, the reverse is becoming moreommon and fMRI may soon be aiding neurophysiologists innowing where to plant their electrodes.
In addition to neuroimaging, TMS has been a useful tech-ique for investigating the causal role of brain regions inisuomotor control (Desmurget et al., 1999; Glover, Miall, &ushworth, 2005; Tunik, Frey, & Grafton, 2005). However,ost studies to date have used only rough anatomical land-arks to guide placement of the TMS coil. With the availability
f software using fMRI data to guide coil placement (Koski,olnar-Szakacs, & Iacoboni, 2005), it should be much easier to
arget known areas in individual subjects with TMS, such as theutative homologues in parietal cortex.
To summarize, over the last twenty years neuroimaging hasade a significant contribution to the understanding of the func-
ional role of the parietal lobes in visuomotor control. In thease of fMRI, the restricted confines of the magnet environmentrovide challenges for researchers wishing to undertake care-ully designed visuomotor experiments. Nevertheless, many labsave developed techniques and paradigms to overcome theseroblems, and we are now seeing a more consistent patterncross studies. Further challenges remain. In particular, therere difficulties associated with establishing clear homologiesetween the macaque and the human brain, and there are incon-istencies between imaging and neuropsychological studies ofemispheric specialization. Certainly the merging of differentechniques will be a promising approach to meeting these chal-enges and should include: TMS in conjunction with fMRI,euroimaging with patients, and more similarity between humannd monkey paradigms.
cknowledgements
The data presented on the laterality of grasping activation wasollected with the assistance of Stacey Danckert, Joe Gati andavi Menon, and was part of a larger project on imagined vs.
eal grasping done in collaboration with David Milner and Meloodale. The authors are grateful to Anthony Cate, Erik Chang,atrizia Fattori, Mary Ellen Large, Sukhvinder Obhi, Kennethalyear and three anonymous reviewers for their comments onarlier drafts. This work was funded by grants from the Canadiannstitutes for Health Research (operating grant to J.C.C. androup grant to the CIHR Group on Action and Perception), therovince of Ontario Premier’s Research Excellence Award (to.C.C.), and a training grant from the McDonnell-Pew Programn Cognitive Neuroscience (to J.C.C.).
eferences
ndersen, R. A., Asanuma, C., Essick, G., & Siegel, R. M. (1990). Cortico-cortical connections of anatomically and physiologically defined subdivi-

2 ychol
A
A
A
A
B
B
B
B
B
B
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
D
D
D
680 J.C. Culham et al. / Neurops
sions within the inferior parietal lobule. Journal of Comparative Neurology,296(1), 65–113.
ndersen, R. A., Bracewell, R. M., Barash, S., Gnadt, J. W., & Fogassi, L.(1990). Eye position effects on visual, memory, and saccade-related activ-ity in areas LIP and 7a of macaque. Journal of Neuroscience, 10, 1176–1196.
ndersen, R. A., & Buneo, C. A. (2002). Intentional maps in posterior parietalcortex. Annual Review of Neuroscience, 25, 189–220.
ndersen, R. A., Essick, G. K., & Siegel, R. M. (1985). Encoding of spatiallocation by posterior parietal neurons. Science, 230(4724), 456–458.
stafiev, S. V., Shulman, G. L., Stanley, C. M., Snyder, A. Z., Van Essen, D. C.,& Corbetta, M. (2003). Functional organization of human intraparietal andfrontal cortex for attending, looking, and pointing. Journal of Neuroscience,23(11), 4689–4699.
alasubramanian, V. (2005). Dysgraphia in two forms of conduction aphasia.Brain and Cognition, 57(1), 8–15.
alint, R. (1909). Seelenhammung des ‘Schauens’, optische Ataxie, raumlicheStorungen des Aufmersamkeit. Monastchrift fur Psychiatrie und Neurologie,25, 51–81.
attaglia-Mayer, A., Ferraina, S., Mitsuda, T., Marconi, B., Genovesio, A., Ono-rati, P., et al. (2000). Early coding of reaching in the parietooccipital cortex.Journal of Neurophysiology, 83(4), 2374–2391.
erman, R. A., Colby, C. L., Genovese, C. R., Voyvodic, J. T., Luna, B., Thul-born, K. R., et al. (1999). Cortical networks subserving pursuit and saccadiceye movements in humans: An FMRI study. Human Brain Mapping, 8(4),209–225.
erti, A., & Frassinetti, F. (2000). When far becomes near: Remapping of spaceby tool use. Journal of Cognitive Neuroscience, 12(3), 415–420.
inkofski, F., Buccino, G., Stephan, K. M., Rizzolatti, G., Seitz, R. J., & Freund,H. J. (1999). A parieto-premotor network for object manipulation: Evi-dence from neuroimaging. Experimental Brain Research, 128(1–2), 210–213.
inkofski, F., Butler, A., Buccino, G., Heide, W., Fink, G., Freund, H. J., et al.(2003). Mirror apraxia affects the peripersonal mirror space. A combinedlesion and cerebral activation study. Experimental Brain Research, 153(2),210–219.
inkofski, F., Dohle, C., Posse, S., Stephan, K. M., Hefter, H., Seitz, R. J., etal. (1998). Human anterior intraparietal area subserves prehension: A com-bined lesion and functional MRI activation study. Neurology, 50(5), 1253–1259.
irn, R. M., Bandettini, P. A., Cox, R. W., & Shaker, R. (1999). Event-relatedfMRI of tasks involving brief motion. Human Brain Mapping, 7(2), 106–114.
remmer, F., Schlack, A., Duhamel, J. R., Graf, W., & Fink, G. R. (2001).Space coding in primate posterior parietal cortex. Neuroimage, 14, S46–S51.
remmer, F., Schlack, A., Shah, N. J., Zafiris, O., Kubischik, M., Hoffman, K.-P.,et al. (2001). Polymodal motion processing in posterior parietal and premotorcortex: A human fMRI study strongly implies equivalencies between humansand monkeys. Neuron, 29(1), 287–296.
uneo, C. A., Jarvis, M. R., Batista, A. P., & Andersen, R. A. (2002). Directvisuomotor transformations for reaching. Nature, 416(6881), 632–636.
alton, J. L., Dickinson, A. R., & Snyder, L. H. (2002). Non-spatial, motor-specific activation in posterior parietal cortex. Nature Neuroscience, 5(6),580–588.
astiello, U. (2005). The neuroscience of grasping. Nature Reviews Neuro-science, 6(9), 726–736.
hao, L. L., & Martin, A. (2000). Representation of manipulable man-madeobjects in the dorsal stream. Neuroimage, 12(4), 478–484.
hapman, H., Gavrilescu, M., Wang, H., Kean, M., Egan, G., & Castiello, U.(2002). Posterior parietal cortex control of reach-to-grasp movements inhumans. European Journal of Neuroscience, 15(12), 2037–2042.
lower, D. M., Hoffman, J. M., Votaw, J. R., Faber, T. L., Woods, R. P., &
Alexander, G. E. (1996). Role of posterior parietal cortex in the recalibrationof visually guided reaching. Nature, 383(6601), 618–621.ohen, Y. E., & Andersen, R. A. (2002). A common reference frame for move-ment plans in the posterior parietal cortex. Nature Reviews Neuroscience,3(7), 553–562.
D
D
ogia 44 (2006) 2668–2684
olby, C. L. (1998). Action-oriented spatial reference frames in cortex. Neuron,20(1), 15–24.
olby, C. L., & Duhamel, J. R. (1991). Heterogeneity of extrastriate visual areasand multiple parietal areas in the macaque monkey. Neuropsychologia, 29(6),517–537.
olby, C. L., Duhamel, J.-R., & Goldberg, M. E. (1993). Ventral intraparietal areaof the macaque: Anatomic location and visual response properties. Journalof Neurophysiology, 6(3), 902–914.
olby, C. L., Duhamel, J.-R., & Goldberg, M. E. (1996). Visual, presaccadic,and cognitive activation of single neurons in monkey lateral intraparietalarea. Journal of Neurophysiology, 76(5), 2841–2851.
onnolly, J. D., Andersen, R. A., & Goodale, M. A. (2003). FMRI evidence fora ‘parietal reach region’ in the human brain. Experimental Brain Research,153(2), 140–145.
onnolly, J. D., Goodale, M. A., Desouza, J. F., Menon, R. S., & Vilis, T. (2000).A comparison of frontoparietal fMRI activation during anti-saccades andanti-pointing. Journal of Neurophysiology, 84(3), 1645–1655.
ooke, D. F., & Graziano, M. S. (2003). Defensive movements evokedby air puff in monkeys. Journal of Neurophysiology, 90(5), 3317–3329.
ooke, D. F., Taylor, C. S., Moore, T., & Graziano, M. S. (2003). Complexmovements evoked by microstimulation of the ventral intraparietal area.Proceedings of the National Academy of Sciences (USA), 100(10), 6163–6168.
orbetta, M. (1998). Frontoparietal cortical networks for directing attentionand the eye to visual locations: Identical, independent, or overlapping neu-ral systems? Proceedings of the National Academy of Sciences (USA), 95,831–838.
orbetta, M., Miezin, F. M., Shulman, G. L., & Petersen, S. E. (1993). A PETstudy of visuospatial attention. Journal of Neuroscience, 13(3), 1202–1226.
reem-Regehr, S. H., & Lee, J. N. (2005). Neural representations of gras-pable objects: Are tools special? Cognitive Brain Research, 22, 457–469.
ulham, J. C. (2003). Human brain imaging reveals a parietal area specialized forgrasping. In N. Kanwisher & J. Duncan (Eds.), Attention and performanceXX: Functional brain imaging of human cognition. Oxford, U.K.: OxfordUniversity Press.
ulham, J. C. (in press). Functional neuroimaging: Experimental design andanalysis. In R. Cabeza & A. Kingstone (Eds.), Handbook of functional neu-roimaging of cognition. (2nd ed.). Cambridge MA: MIT Press.
ulham, J. C., Cavina Pratesi, C., & Goodale, M. A., 2004. Object size mattersfor grasping but NOT for perception in the human anterior intraparietalarea. Paper presented at the Society for Neuroscience, San Diego, CA.
ulham, J. C., Danckert, S. L., DeSouza, J. F., Gati, J. S., Menon, R. S., &Goodale, M. A. (2003). Visually guided grasping produces fMRI activationin dorsal but not ventral stream brain areas. Experimental Brain Research,153(2), 180–189.
ulham, J. C., & Kanwisher, N. G. (2001). Neuroimaging of cognitive functionsin human parietal cortex. Current Opinion in Neurobiology, 11(2), 157–163.
ulham, J. C., Valyear, K. F., & Stiglick, A. J., 2004. fMRI activation in grasp-related regions during naming of tools and other graspable objects. Posterpresented at the Vision Sciences Society, Sarasota, FL.
ulham, J. C., Woodward, S. L., Milner, A. D., Gati, J. S., Menon, R. S., &Goodale, M. A., 2001. Laterality of fMRI activation in AIP during graspingand imagined grasping of visual targets. Paper presented at the Society forNeuroscience, San Diego, CA.
amasio, H. (2005). Human brain anatomy in computerized images (2nd ed.).Oxford, New York: Oxford University Press.
ebowy, D. J., Ghosh, S., Ro, J. Y., & Gardner, E. P. (2001). Comparison ofneuronal firing rates in somatosensory and posterior parietal cortex duringprehension. Experimental Brain Research, 137(3–4), 269–291.
ecety, J., Kawashima, R., Gulyas, B., & Roland, P. E. (1992). Preparation forreaching: A PET study of the participating structures in the human brain.
Neuroreport, 3(9), 761–764.ehaene, S., & Cohen, L. (1995). Toward an anatomical and functional modelof number processing. Mathematical Cognition, 1, 83–120.
enys, K., Vanduffel, W., Fize, D., Nelissen, K., Peuskens, H., Van Essen, D.,et al. (2004). The processing of visual shape in the cerebral cortex of human

ychol
D
D
D
d
D
D
E
E
E
E
E
E
F
F
F
F
F
F
F
F
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
H
J.C. Culham et al. / Neurops
and nonhuman primates: A functional magnetic resonance imaging study.Journal of Neuroscience, 24(10), 2551–2565.
esmurget, M., Epstein, C. M., Turner, R. S., Prablanc, C., Alexander, G. E.,& Grafton, S. T. (1999). Role of the posterior parietal cortex in updatingreaching movements to a visual target. Nature Neuroscience, 2(6), 563–567.
esmurget, M., Grea, H., Grethe, J. S., Prablanc, C., Alexander, G. E., & Grafton,S. T. (2001). Functional anatomy of nonvisual feedback loops during reach-ing: A positron emission tomography study. Journal of Neuroscience, 21(8),2919–2928.
eSouza, J. F., Dukelow, S. P., Gati, J. S., Menon, R. S., Andersen, R. A.,& Vilis, T. (2000). Eye position signal modulates a human parietal pointingregion during memory-guided movements. Journal of Neuroscience, 20(15),5835–5840.
i Pellegrino, G., & Frassinetti, F. (2000). Direct evidence from parietal extinc-tion of enhancement of visual attention near a visible hand. Current Biology,10(22), 1475–1477.
uhamel, J. R., Colby, C. L., & Goldberg, M. E. (1992). The updating of therepresentation of visual space in parietal cortex by intended eye movements.Science, 255, 90–92.
uhamel, J. R., Colby, C. L., & Goldberg, M. E. (1998). Ventral intraparietal areaof the macaque: Congruent visual and somatic response properties. Journalof Neurophysiology, 79(1), 126–136.
hrsson, H. H., Fagergren, A., Jonsson, T., Westling, G., Johansson, R. S., &Forssberg, H. (2000). Cortical activity in precision- versus power-grip tasks:An fMRI study. Journal of Neurophysiology, 83(1), 528–536.
hrsson, H. H., Fagergren, E., & Forssberg, H. (2001). Differential fronto-parietal activation depending on force used in a precision grip task: AnfMRI study. Journal of Neurophysiology, 85(6), 2613–2623.
ickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C., Fink, G. R., Amunts,K., et al. (2005). A new SPM toolbox for combining probabilistic cytoar-chitectonic maps and functional imaging data. Neuroimage, 25(4), 1325–1335.
ickhoff, S. B., Walters, N. B., Schleicher, A., Kril, J., Egan, G. F., Zilles, K.,et al. (2005). High-resolution MRI reflects myeloarchitecture and cytoar-chitecture of human cerebral cortex. Human Brain Mapping, 24(3), 206–215.
skandar, E. N., & Assad, J. A. (1999). Dissociation of visual, motor and predic-tive signals in parietal cortex during visual guidance. Nature Neuroscience,2(1), 88–93.
skandar, E. N., & Assad, J. A. (2002). Distinct nature of directional signalsamong parietal cortical areas during visual guidance. Journal of Neurophys-iology, 88(4), 1777–1790.
aillenot, I., Sakata, H., Costes, N., Decety, J., & Jeannerod, M. (1997). Visualworking memory for shape and 3D-orientation: A PET study. Neuroreport,8(4), 859–862.
aillenot, I., Sunaert, S., Van Hecke, P., & Orban, G. A. (2001). Orientationdiscrimination of objects and gratings compared: An fMRI study. EuropeanJournal of Neuroscience, 13(3), 585–596.
ang, F., Murray, S. O., Kersten, D. J., & He, S. (2005). Orientation-tunedfMRI adaptation in human visual cortex. Journal of Neurohysiology, 94,4188–4195.
arne, A., & Ladavas, E. (2000). Dynamic size-change of hand peripersonalspace following tool use. Neuroreport, 11(8), 1645–1649.
attori, P., Gamberini, M., Kutz, D. F., & Galletti, C. (2001). ‘Arm-reaching’neurons in the parietal area V6A of the macaque monkey. European Journalof Neuroscience, 13(12), 2309–2313.
erraina, S., Battaglia-Mayer, A., Genovesio, A., Marconi, B., Onorati, P.,& Caminiti, R. (2001). Early coding of visuomanual coordination duringreaching in parietal area PEc. Journal of Neurophysiology, 85(1), 462–467.
ogassi, L., Ferrari, P. F., Gesierich, B., Rozzi, S., Chersi, F., & Rizzolatti, G.(2005). Parietal lobe: From action organization to intention understanding.
Science, 308(5722), 662–667.rey, S. H., Vinton, D., Norlund, R., & Grafton, S. T. (2005). Corticaltopography of human anterior intraparietal cortex active during visuallyguided grasping. Brain Research. Cognitive Brain Research, 23(2–3), 397–405.
H
H
ogia 44 (2006) 2668–2684 2681
allese, V., Murata, A., Kaseda, M., Niki, N., & Sakata, H. (1994). Deficit ofhand preshaping after muscimol injection in monkey parietal cortex. Neu-roreport, 5(12), 1525–1529.
alletti, C., Fattori, P., Kutz, D. F., & Battaglini, P. P. (1997). Arm movement-related neurons in the visual area V6A of the macaque superior parietallobule. European Journal of Neuroscience, 9(2), 410–413.
alletti, C., Kutz, D. F., Gamberini, M., Breveglieri, R., & Fattori, P. (2003).Role of the medial parieto-occipital cortex in the control of reachingand grasping movements. Experimental Brain Research, 153(2), 158–170.
ardner, E. P., Debowy, D. J., Ro, J. Y., Ghosh, S., & Babu, K. S. (2002). Sensorymonitoring of prehension in the parietal lobe: A study using digital video.Behavioural Brain Research, 135(1–2), 213–224.
ardner, E. P., Ro, J. Y., Debowy, D., & Ghosh, S. (1999). Facilitation of neuronalactivity in somatosensory and posterior parietal cortex during prehension.Experimental Brain Research, 127(4), 329–354.
itelman, D. R., Nobre, A. C., Parrish, T. B., LaBar, K. S., Kim, Y. H., Meyer, J.R., et al. (1999). A large-scale distributed network for covert spatial attention:Further anatomical delineation based on stringent behavioural and cognitivecontrols. Brain, 122(Pt 6), 1093–1106.
lover, S., Miall, R. C., & Rushworth, M. F. (2005). Parietal rTMS disrupts theinitiation but not the execution of on-line adjustments to a perturbation ofobject size. Journal of Cognitive Neuroscience, 17(1), 124–136.
oltz, H. C., Dukelow, S. P., De Souza, J. F. X., Culham, J. C., van den Berg,A. V., & Goosens, H. H. L., et al. (2001). A putative homologue of monkeyarea VIP in humans. Poster presented at the Society for Neuroscience, SanDiego, CA.
oodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perceptionand action. Trends in Neurosciences, 15(1), 20–25.
oodale, M. A., Milner, A. D., Jakobson, L. S., & Carey, D. P. (1991). A neuro-logical dissociation between perceiving objects and grasping them. Nature,349(6305), 154–156.
rafton, S. T., Fagg, A. H., Woods, R. P., & Arbib, M. A. (1996). Func-tional anatomy of pointing and grasping in humans. Cerebral Cortex, 6(2),226–237.
refkes, C., & Fink, G. R. (2005). The functional organization of the intraparietalsulcus in humans and monkeys. Journal of Anatomy, 207(1), 3–17.
refkes, C., Ritzl, A., Zilles, K., & Fink, G. R. (2004). Human medial intrapari-etal cortex subserves visuomotor coordinate transformation. Neuroimage,23(4), 1494–1506.
refkes, C., Weiss, P. H., Zilles, K., & Fink, G. R. (2002). Crossmodal pro-cessing of object features in human anterior intraparietal cortex: An fMRIstudy implies equivalencies between humans and monkeys. Neuron, 35(1),173–184.
regoriou, G. G., & Savaki, H. E. (2001). The intraparietal cortex: Subregionsinvolved in fixation, saccades, and in the visual and somatosensory guid-ance of reaching. Journal of Cerebral Blood Flow Metabolism, 21(6), 671–682.
regoriou, G. G., & Savaki, H. E. (2003). When vision guides movement: Afunctional imaging study of the monkey brain. Neuroimage, 19(3), 959–967.
rezes, J., Tucker, M., Armony, J., Ellis, R., & Passingham, R. E. (2003). Objectsautomatically potentiate action: An fMRI study of implicit processing. Euro-pean Journal of Neuroscience, 17(12), 2735–2740.
rill-Spector, K., Kushnir, T., Edelman, S., Avidan, G., Itzchak, Y., & Malach, R.(1999). Differential processing of objects under various viewing conditionsin the human lateral occipital complex. Neuron, 24(1), 187–203.
rill-Spector, K., & Malach, R. (2001). fMR-adaptation: A tool for studying thefunctional properties of human cortical neurons. Acta Psychologica (Ams-terdam), 107(1–3), 293–321.
alligan, P. W., Fink, G. R., Marshall, J. C., & Vallar, G. (2003). Spatial cog-nition: Evidence from visual neglect. Trends in Cognitive Sciences, 7(3),125–133.
arshman, R. A., Hampson, E., & Berenbaum, S. A. (1983). Individual differ-ences in cognitive abilities and brain organization. Part I: Sex and handednessdifferences in ability. Canadian Journal of Psychology, 37(1), 144–192.
enson, R. N. (2003). Neuroimaging studies of priming. Progress in Neurobi-ology, 70(1), 53–81.

2 ychol
H
H
I
I
I
J
J
J
J
J
J
J
K
K
K
K
K
K
K
K
K
K
K
K
L
L
L
L
L
L
L
L
M
M
M
M
M
M
M
M
M
N
N
O
682 J.C. Culham et al. / Neurops
uang, R.-S., & Sereno, M. (2005). Mapping the human homologue of theventral intraparietal area (VIP). Neuroimage, 26(Suppl.), S23.
yvarinen, J., & Poranen, A. (1974). Function of the parietal associative area 7as revealed from cellular discharges in alert monkeys. Brain, 97(4), 673–692.
acoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta, J. C., &Rizzolatti, G. (1999). Cortical mechanisms of human imitation. Science,286(5449), 2526–2528.
noue, K., Kawashima, R., Satoh, K., Kinomura, S., Goto, R., Koyama, M., etal. (1998). PET study of pointing with visual feedback of moving hands.Journal of Neurophysiology, 79(1), 117–125.
riki, A., Tanaka, M., & Iwamura, Y. (1996). Coding of modified body schemaduring tool use by macaque postcentral neurones. Neuroreport, 7(14),2325–2330.
ames, T., Humphrey, G., Gati, J., Menon, R., & Goodale, M. (2002). Differentialeffects of viewpoint on object-driven activation in dorsal and ventral streams.Neuron, 35(4), 793.
ames, T. W., Culham, J., Humphrey, G. K., Milner, A. D., & Goodale, M. A.(2003). Ventral occipital lesions impair object recognition but not object-directed grasping: An fMRI study. Brain, 126(Pt 11), 2463–2475.
ancke, L., Kleinschmidt, A., Mirzazade, S., Shah, N. J., & Freund, H. J. (2001).The role of the inferior parietal cortex in linking the tactile perceptionand manual construction of object shapes. Cerebral Cortex, 11(2), 114–121.
eannerod, M., Decety, J., & Michel, F. (1994). Impairment of grasping move-ments following a bilateral posterior parietal lesion. Neuropsychologia,32(4), 369–380.
enkins, I. H., Jahanshahi, M., Jueptner, M., Passingham, R. E., & Brooks, D. J.(2000). Self-initiated versus externally triggered movements. II. The effectof movement predictability on regional cerebral blood flow. Brain, 123(Pt6), 1216–1228.
ohnson, P. B., Ferraina, S., Bianchi, L., & Caminiti, R. (1996). Corticalnetworks for visual reaching: Physiological and anatomical organizationof frontal and parietal lobe arm regions. Cerebral Cortex, 6(2), 102–119.
ohnson-Frey, S. H. (2004). The neural bases of complex tool use in humans.Trends in Cognitive Sciences, 8(2), 71–78.
alaska, J. F. (1996). Parietal cortex area 5 and visuomotor behavior. CanadianJournal of Physiology and Pharmacology, 74, 483–498.
arnath, H. O., Ferber, S., & Himmelbach, M. (2001). Spatial awareness is afunction of the temporal not the posterior parietal lobe. Nature, 411(6840),950–953.
arnath, H. O., & Perenin, M. T. (2005). Cortical control of visually guidedreaching: Evidence from patients with optic ataxia. Cerebral Cortex, 15(10),1561–1569.
awashima, R., Naitoh, E., Matsumura, M., Itoh, H., Ono, S., Satoh, K., et al.(1996). Topographic representation in human intraparietal sulcus of reachingand saccade. Neuroreport, 7, 1253–1256.
ertzman, C., Schwarz, U., Zeffiro, T. A., & Hallett, M. (1997). The role ofposterior parietal cortex in visually guided reaching movements in humans.Experimental Brain Research, 114(1), 170–183.
im, S. G., Ashe, J., Hendrich, K., Ellermann, J. M., Merkle, H., Ugurbil,K., et al. (1993). Functional magnetic resonance imaging of motor cor-tex: Hemispheric asymmetry and handedness. Science, 261(5121), 615–617.
imura, D. (1983). Sex differences in cerebral organization for speech and praxicfunctions. Canadian Journal of Psychology, 37(1), 19–35.
olb, B., & Whishaw, I. Q. (2003). Fundamentals of human neuropsychology(5th ed.). New York: Worth Publishers.
oski, L., Molnar-Szakacs, I., & Iacoboni, M. (2005). Exploring the con-tributions of premotor and parietal cortex to spatial compatibility usingimage-guided TMS. Neuroimage, 24(2), 296–305.
oyama, M., Hasegawa, I., Osada, T., Adachi, Y., Nakahara, K., & Miyashita,Y. (2004). Functional magnetic resonance imaging of macaque monkeys
performing visually guided saccade tasks: Comparison of cortical eye fieldswith humans. Neuron, 41(5), 795–807.rubitzer, L. (1995). The organization of neocortex in mammals: Are speciesdifferences really so different? Trends in Neurosciences, 18(9), 408–417.
O
ogia 44 (2006) 2668–2684
uhtz-Buschbeck, J. P., Ehrsson, H. H., & Forssberg, H. (2001). Human brainactivity in the control of fine static precision grip forces: An fMRI study.European Journal of Neuroscience, 14(2), 382–390.
acquaniti, F., Perani, D., Guigon, E., Bettinardi, V., Carrozzo, M., Grassi, F., etal. (1997). Visuomotor transformations for reaching to memorized targets:A PET study. Neuroimage, 5, 129–146.
adavas, E. (2002). Functional and dynamic properties of visual peripersonalspace. Trends in Cognitive Sciences, 6(1), 17–22.
adavas, E., di Pellegrino, G., Farne, A., & Zeloni, G. (1998). Neuropsycho-logical evidence of an integrated visuotactile representation of periper-sonal space in humans. Journal of Cognitive Neuroscience, 10(5), 581–589.
andis, T. (2000). Disruption of space perception due to cortical lesions. SpatialVision, 13(2–3), 179–191.
eiguarda, R. C., & Marsden, C. D. (2000). Limb apraxias: Higher-order disor-ders of sensorimotor integration. Brain, 123(Pt 5), 860–879.
evine, D. N., Kaufman, K. J., & Mohr, J. P. (1978). Inaccurate reachingassociated with a superior parietal lobe tumor. Neurology, 28(6), 555–561.
ogothetis, N. K. (2003). The underpinnings of the BOLD functional mag-netic resonance imaging signal. Journal of Neuroscience, 23(10), 3963–3971.
uppino, G., Murata, A., Govoni, P., & Matelli, M. (1999). Largely segregatedparietofrontal connections linking rostral intraparietal cortex (areas AIP andVIP) and the ventral premotor cortex (areas F5 and F4). Experimental BrainResearch, 128(1–2), 181–187.
acKay, W. A. (1992). Properties of reach-related neuronal activity in corticalarea 7A. Journal of Neurophysiology, 67(5), 1335–1345.
aravita, A., & Iriki, A. (2004). Tools for the body (schema). Trends in CognitiveSciences, 8(2), 79–86.
edendorp, W. P., Goltz, H. C., Crawford, J. D., & Vilis, T. (2005). Inte-gration of target and effector information in human posterior parietal cor-tex for the planning of action. Journal of Neurophysiology, 93(2), 954–962.
edendorp, W. P., Goltz, H. C., & Vilis, T. (2005). Remapping the rememberedtarget location for anti-saccades in human posterior parietal cortex. Journalof Neurohysiology, 94(1), 734–740.
edendorp, W. P., Goltz, H. C., Vilis, T., & Crawford, J. D. (2003). Gaze-centered updating of visual space in human parietal cortex. Journal ofNeuroscience, 23(15), 6209–6214.
erriam, E. P., Genovese, C. R., & Colby, C. L. (2003). Spatial updating inhuman parietal cortex. Neuron, 39(2), 361–373.
ountcastle, V. B., Lynch, J. C., Georgopoulos, A., Sakata, H., & Acuna, C.(1975). Posterior parietal association cortex of the monkey: Command func-tions for operations within extrapersonal space. Journal of Neurophysiology,38(4), 871–908.
urata, A., Gallese, V., Luppino, G., Kaseda, M., & Sakata, H. (2000). Selec-tivity for the shape, size, and orientation of objects for grasping in neuronsof monkey parietal area AIP. Journal of Neurophysiology, 83(5), 2580–2601.
uri, R. M., Iba-Zizen, M. T., Derosier, C., Cabanis, E. A., & Pierrot-Deseiligny,C. (1996). Location of the human posterior eye fields with functional mag-netic resonance imaging. Journal of Neurology, Neurosurgery and Psychia-try, 60, 445–448.
akahara, K., Hayashi, T., Konishi, S., & Miyashita, Y. (2002). FunctionalMRI of macaque monkeys performing a cognitive set-shifting task. Science,295(5559), 1532–1536.
akamura, H., Kuroda, T., Wakita, M., Kusunoki, M., Kato, A., Mikami, A., etal. (2001). From three-dimensional space vision to prehensile hand move-ments: The lateral intraparietal area links the area V3A and the anteriorintraparietal area in macaques. Journal of Neuroscience, 21(20), 8174–8187.
bayashi, S., Suhara, T., Kawabe, K., Okauchi, T., Maeda, J., Akine, Y., et al.
(2001). Functional brain mapping of monkey tool use. Neuroimage, 14(4),853–861.rban, G. A., Van Essen, D., & Vanduffel, W. (2004). Comparative mapping ofhigher visual areas in monkeys and humans. Trends in Cognitive Sciences,8(7), 315–324.

ychol
P
P
P
P
P
P
P
Q
Q
R
R
R
R
R
R
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
T
T
T
T
T
T
U
V
J.C. Culham et al. / Neurops
erenin, M. T., & Vighetto, A. (1988). Optic ataxia: A specific disruption invisuomotor mechanisms. I. Different aspects of the deficit in reaching forobjects. Brain, 111(Pt 3), 643–674.
etit, L., & Haxby, J. V. (1999). Functional anatomy of pursuit eye movementsin humans as revealed by fMRI. Journal of Neurophysiology, 82(1), 463–471.
euskens, H., Claeys, K. G., Todd, J. T., Norman, J. F., Van Hecke, P., & Orban,G. A. (2004). Attention to 3-D shape, 3-D motion, and texture in 3-D struc-ture from motion displays. Journal of Cognitive Neuroscience, 16(4), 665–682.
ierrot-Deseilligny, C., Milea, D., & Muri, R. M. (2004). Eye movementcontrol by the cerebral cortex. Current Opinion in Neurology, 17(1), 17–25.
ierrot-Deseilligny, C. H., Rivaud, S., Gaymard, B., & Agid, Y. (1991).Cortical control of reflexive visually-guided saccades. Brain, 114, 1473–1485.
rado, J., Clavagnier, S., Otzenberger, H., Scheiber, C., Kennedy, H., & Perenin,M. T. (2005). Two cortical systems for reaching in central and peripheralvision. Neuron, 48(5), 849–858.
revic, F. H. (1998). The neuropsychology of 3-D space. Psychological Bulletin,124(2), 123–164.
uinlan, D. J., Bell, I. E., Servos, P., Klassen, L. M., Gati, J. S., & Menon,R. S., et al. (2005). Gradient distortion correction enables raising andtilting the head for direct hand viewing during visuomotor fMRI exper-iments. Poster presented at the Organization for Human Brain Mapping,Toronto, ON.
uinlan, D. J., Goodale, M. A., & Culham, J. C., 2004. fMRI reveals a preferencefor near vs. far vergence in human superior parieto-occipital cortex. Posterpresented at the Society for Neuroscience, San Diego, CA.
aos, V., Evangeliou, M. N., & Savaki, H. E. (2004). Observation of action:Grasping with the mind’s hand. Neuroimage, 23(1), 193–201.
izzolatti, G., & Arbib, M. A. (1998). Language within our grasp. Trends inNeurosciences, 21(5), 188–194.
izzolatti, G., & Craighero, L. (2004). The mirror-neuron system. AnnualReview of Neuroscience, 27, 169–192.
izzolatti, G., Fadiga, L., Matelli, M., Bettinardi, V., Paulesu, E., Perani, D.,et al. (1996). Localization of grasp representations in humans by PET: 1.Observation versus execution. Experimental Brain Research, 111(2), 246–252.
o, J. Y., Debowy, D., Ghosh, S., & Gardner, E. P. (2000). Depression of neu-ronal firing rates in somatosensory and posterior parietal cortex during objectacquisition in a prehension task. Experimental Brain Research, 135(1),1–11.
orden, C., & Karnath, H.-O. (2004). Using human brain lesions to infer func-tion: A relic from a past era in the fMRI age? Nature Reviews Neuroscience,5(10), 812–819.
akata, H., Taira, M., Kusunoki, M., Murata, A., Tanaka, Y., & The TINS Lec-ture. (1997). The parietal association cortex in depth perception and visualcontrol of hand action. Trends in Neurosciences, 20(8), 350–357.
akata, H., Taira, M., Mine, S., & Murata, A. (1992). Hand-movement-related neurons of the posterior parietal cortex of the monkey: Their rolein the visual guidance of hand movements. In R. Caminiti, P. B. John-son, & Y. Burnod (Eds.), Control of arm movement in space: Neuro-physiological and computational approaches (pp. 185–198). Heidelberg:Springer.
avaki, H. E., Raos, V. C., & Dalezios, Y. (1997). Spatial cortical patterns ofmetabolic activity in monkeys performing a visually guided reaching taskwith one forelimb. Neuroscience, 76(4), 1007–1034.
awamura, H., Vogels, R., & Orban, G. A., 2004. Single cell investigation ofstimulus adaptation in monkey inferior temporal cortex. Paper presented atthe Society for Neuroscience, San Diego, California.
cannell, J. W., & Young, M. P. (1999). Neuronal population activity and func-tional imaging. Proceedings of the Royal Society of London, B, 266(1422),
875–881.cheperjans, F., Palomero-Gallagher, N., Grefkes, C., Schleicher, A., & Zilles,K. (2005). Transmitter receptors reveal segregation of cortical areas inthe human superior parietal cortex: Relations to visual and somatosensoryregions. Neuroimage, 28(2), 362–379.
V
ogia 44 (2006) 2668–2684 2683
ereno, M. I., Pitzalis, S., & Martinez, A. (2001). Mapping of contralateral spacein retinotopic coordinates by a parietal cortical area in humans. Science,294(5545), 1350–1354.
ereno, M. I., & Tootell, R. B. (2005). From monkeys to humans: What dowe now know about brain homologies? Current Opinion in Neurobiology,15(2), 135–144.
hikata, E., Hamzei, F., Glauche, V., Koch, M., Weiller, C., Binkofski, F., et al.(2003). Functional properties and interaction of the anterior and posteriorintraparietal areas in humans. European Journal of Neuroscience, 17(5),1105–1110.
hikata, E., Hamzel, F., Glauche, V., Knab, R., Dettmers, C., Weiller, C., etal. (2001). Surface orientation discrimination activates caudal and anteriorintraparietal sulcus in humans: An event-related fMRI study. Journal ofNeurophysiology, 85(3), 1309–1314.
hmuelof, L., & Zohary, E. (2005). Dissociation between ventral and dor-sal fMRI activation during object and action recognition. Neuron, 47(3),457–470.
hulman, G. L., d’Avossa, G., Tansy, A. P., & Corbetta, M. (2002). Twoattentional processes in the parietal lobe. Cerebral Cortex, 12(11), 1124–1131.
ilver, M. A., Ress, D., & Heeger, D. J. (2005). Topographic maps of visualspatial attention in human parietal cortex. Journal of Neurophysiology, 94,1348–1371.
imon, O., Mangin, J. F., Cohen, L., Le Bihan, D., & Dehaene, S. (2002).Topographical layout of hand, eye, calculation, and language-related areasin the human parietal lobe. Neuron, 33(3), 475–487.
inghal, A., Kaufman, L., Valyear, K., & Culham, J. C., 2005. fMRI reactivationof the human lateral occipital complex during delayed actions to rememberedobjects. Paper presented at the object perception and memory workshop,Toronto, ON.
nyder, J. J., & Chatterjee, A. (2004). Spatial-temporal anisometries followingright parietal damage. Neuropsychologia, 42(12), 1703–1708.
toeckel, M. C., Weder, B., Binkofski, F., Buccino, G., Shah, N. J., & Seitz,R. J. (2003). A fronto-parietal circuit for tactile object discrimination: Anevent-related fMRI study. Neuroimage, 19(3), 1103–1114.
triedter, G. F. (2005). Principles of brain evolution. Sunderland, Mass.: SinauerAssociates.
unaert, S., Van Hecke, P., Marchal, G., & Orban, G. A. (2000). Attention tospeed of motion, speed discrimination, and task difficulty: An fMRI study.Neuroimage, 11(6 Pt 1), 612–623.
aira, M., Mine, S., Georgopoulos, A. P., Murata, A., & Sakata, H. (1990).Parietal cortex neurons of the monkey related to the visual guidance of handmovement. Experimental Brain Research, 83(1), 29–36.
aira, M., Nose, I., Inoue, K., & Tsutsui, K. (2001). Cortical areas related toattention to 3D surface structures based on shading: An fMRI study. Neu-roimage, 14(5), 959–966.
anabe, H. C., Kato, M., Miyauchi, S., Hayashi, S., & Yanagida, T. (2005).The sensorimotor transformation of cross-modal spatial information in theanterior intraparietal sulcus as revealed by functional MRI. Cognitive BrainResearch, 22, 385–396.
oni, I., Rushworth, M. F., & Passingham, R. E. (2001). Neural correlates ofvisuomotor associations. Spatial rules compared with arbitrary rules. Exper-imental Brain Research, 141(3), 359–369.
sutsui, K., Sakata, H., Naganuma, T., & Taira, M. (2002). Neural correlatesfor perception of 3D surface orientation from texture gradient. Science,298(5592), 409–412.
unik, E., Frey, S. H., & Grafton, S. T. (2005). Virtual lesions of the ante-rior intraparietal area disrupt goal-dependent on-line adjustments of grasp.Nature Neuroscience, 8(4), 505–511.
ngerleider, L. G., & Mishkin, M. (1982). Two cortical visual systems. In D.J. Ingle, M. A. Goodale, & R. J. W. Mansfield (Eds.), Analysis of visualbehavior (pp. 549–586). Cambridge, MA: MIT Press.
an Essen, D. C., Lewis, J. W., Drury, H. A., Hadjikhani, N., Tootell, R.
B., Bakircioglu, M., et al. (2001). Mapping visual cortex in monkeys andhumans using surface-based atlases. Vision Research, 41(10–11), 1359–1378.anduffel, W., Fize, D., Mandeville, J. B., Nelissen, K., Van Hecke, P., Rosen,B. R., et al. (2001). Visual motion processing investigated using contrast

2 ychol
W
Z
684 J.C. Culham et al. / Neurops
agent-enhanced fMRI in awake behaving monkeys. Neuron, 32(4), 565–
577.eiss, P. H., Marshall, J. C., Wunderlich, G., Tellmann, L., Halligan, P.W., Freund, H. J., et al. (2000). Neural consequences of acting in nearversus far space: A physiological basis for clinical dissociations. Brain,123(Pt 12), 2531–2541.
Z
ogia 44 (2006) 2668–2684
hang, T., Heuer, H. W., & Britten, K. H. (2004). Parietal area VIP neuronal
responses to heading stimuli are encoded in head-centered coordinates. Neu-ron, 42(6), 993–1001.illes, K., & Palomero-Gallagher, N. (2001). Cyto-, myelo-, and recep-tor architectonics of the human parietal cortex. Neuroimage, 14, S8–S20.
Copyright © 2022 FDOKUMEN




















