
The role of HIV-1 DNA as an additional marker of HIV-1 infection
11
Current HIV Research, 2009, 7, 255-265 255 1570-162X/09 $55.00+.00 © 2009 Bentham Science Publishers Ltd. The Role of HIV-1 DNA as an Additional Marker of HIV-1 Infection Apostolos Beloukas *,1 , Dimitrios Paraskevis 1 , Mina Psichogiou 2 , Angelos Hatzakis *,1 1 Department of Hygiene, Epidemiology and Medical Statistics, Medical School, Athens University. Greece; 2 First Department of Propaedeutic Medicine, Medical School, University of Athens, Athens, Greece Abstract: After the infusion of HIV-1 virus into a host cell, RNA is reverse transcribed to dsDNA, which persists intra- cellular to the infected cell in a variety of forms. Numerous in-house assays have been developed for the quantification of the different cellular HIV-1 DNA forms; these implement conventional or real-time PCR methodology. In this review we discuss recent findings about the longitudinal monitoring of cell-associated HIV-1 DNA in naïve and pre-treated patients, as a marker for clinical progression, treatment initiation and long-term success of HAART. These findings underline the importance of monitoring HIV-1 DNA in clinical practice, in addition to HIV-RNA and CD4 + T Cell counts, for the better assessment of HIV-treatment and disease progression. The lack of a standardized real-time PCR assay is major impedi- ment to more wide-spread HIV-1 DNA monitoring. Keywords: HIV-1 DNA, Real-time PCR, Prognostic marker, Disease progression. INTRODUCTION The widespread use of Highly Active Antiretroviral Therapy (HAART) after 1996 had a profound effect on the prognosis of HIV infection [1-4]. HAART is effective in suppressing plasma HIV-1 RNA levels and restoring CD4 + T lymphocytes to levels where opportunistic infections are rare, but it cannot eliminate HIV-1 from the infected patients [5-31]. Indeed, despite powerful long-term viral suppression which may involve up to many years of uninterrupted sup- pressive therapy, there is evidence of ongoing viral replica- tion and the persistence of several forms of HIV-1 DNA in cellular reservoir and anatomical sanctuary sites. These viral reservoirs are one of the major obstacles in HIV-1 eradica- tion [9-12, 16-36]. In this review, we discuss numerous PCR-based methods developed for the quantification of cell-associated HIV-1 DNA. We also present recent findings about the prognostic value of HIV-1 DNA as a marker to disease progression, treatment initiation and prolonged efficacy of HAART. HIV-1 DNA AND THE VIRUS LIFE CYCLE Viruses depend on host’s cellular machinery to dis- seminate their genetic information. The human immunodefi- ciency virus (HIV), a prototype of the lentiviral subfamily of retroviruses, is one of the most efficient viruses in exploiting the host cell’s functions. The life cycle begins with the re- verse transcription of genomic viral RNA into linear uninte- grated HIV-1 DNA. Completion of this reverse transcription step yields the so-called preintegration complex (PIC) which contains viral dsDNA (reverse transcription’s product), vi- rus-derived proteins including integrase (IN) and host pro- teins i.e., HMGa1 and BAF [37-41]. The PIC can be isolated from the cytoplasmic compartment of infected cells and is *Address correspondence to these authors at the Department of Hygiene, Epidemiology and Medical Statistics, Medical School, University of Athens, M. Asias 75, GR-115 27, Athens, Greece; Tel: (+30210) 7462090; Fax: (+30210) 7462190; E-mail: [email protected]; Tel: (+30210) 7486382; Fax: (+30210) 7486382; E-mail: [email protected] capable of integrating its dsDNA into the target’s host DNA genome to establish a stable provirus [41-44]. Although the linear dsDNA molecule is the precursor to the provirus, an infection of the target cells generates a num- ber of non-integrated DNA species [45-49]. These several unintegrated HIV-1 DNA forms, linear or circularized, arise when viral dsDNA fails to integrate the host genome. For instance, (i) the ends of the linear DNA may be joined to form a 2-LTR circle; (ii) homologous recombination be- tween the two LTRs in a 2-LTR circle yields circles with a single LTR (1-LTR); (iii) linear unintegrated HIV-1 DNA form (Fig. 1). Depending on the site of integration and sub- sequent recombination a series of complex DNA structures may result, as well [50]. Non-integrated forms represent the largest fraction of HIV-1 DNA in the nucleus. Several stud- ies have suggested that 2-LTR circular HIV-1 DNA forms might used as a marker of recent cell infection on the basis of their observed lability in vitro and in vivo [51-54]. This, conflicts with other studies, which reported that are highly stable [54-56]. These unintegrated viral DNA forms have been shown to be incapable of directing a productive infec- tion but seem to be responsible for residual viral replication [34, 36, 43, 47, 49]. However, it remains controversial whether these unintegrated viral DNA forms play some role in the HIV replication cycle [57-60]. Therefore, except pro- viral DNA (dsHIV-1 DNA integrated in the host’s genome), several unintegrated forms persist in intracellular HIV reser- voirs. ASSAYS DEVELOPED FOR HIV-1 DNA QUANTIFI- CATION Several assays have been developed for the quantification of cell-associated HIV-1 DNA based on endpoint and real- time PCR methodology. The latter have shown improved accuracy and reproducibility and a wider range of linear quantification since the concentration of the target is esti- mated during the linear phase rather than at the endpoint (plateau phase) of the PCR that is used in conventional end- point PCR methods. In this paragraph we review the major
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The role of HIV-1 DNA as an additional marker of HIV-1 infection

Current HIV Research, 2009, 7, 255-265 255
1570-162X/09 $55.00+.00 © 2009 Bentham Science Publishers Ltd.
The Role of HIV-1 DNA as an Additional Marker of HIV-1 Infection
Apostolos Beloukas*,1
, Dimitrios Paraskevis1, Mina Psichogiou
2, Angelos Hatzakis
*,1
1Department of Hygiene, Epidemiology and Medical Statistics, Medical School, Athens University. Greece;
2First
Department of Propaedeutic Medicine, Medical School, University of Athens, Athens, Greece
Abstract: After the infusion of HIV-1 virus into a host cell, RNA is reverse transcribed to dsDNA, which persists intra-
cellular to the infected cell in a variety of forms. Numerous in-house assays have been developed for the quantification of
the different cellular HIV-1 DNA forms; these implement conventional or real-time PCR methodology. In this review we
discuss recent findings about the longitudinal monitoring of cell-associated HIV-1 DNA in naïve and pre-treated patients,
as a marker for clinical progression, treatment initiation and long-term success of HAART. These findings underline the
importance of monitoring HIV-1 DNA in clinical practice, in addition to HIV-RNA and CD4+ T Cell counts, for the better
assessment of HIV-treatment and disease progression. The lack of a standardized real-time PCR assay is major impedi-
ment to more wide-spread HIV-1 DNA monitoring.
Keywords: HIV-1 DNA, Real-time PCR, Prognostic marker, Disease progression.
INTRODUCTION
The widespread use of Highly Active Antiretroviral Therapy (HAART) after 1996 had a profound effect on the prognosis of HIV infection [1-4]. HAART is effective in suppressing plasma HIV-1 RNA levels and restoring CD4
+ T
lymphocytes to levels where opportunistic infections are rare, but it cannot eliminate HIV-1 from the infected patients [5-31]. Indeed, despite powerful long-term viral suppression which may involve up to many years of uninterrupted sup-pressive therapy, there is evidence of ongoing viral replica-tion and the persistence of several forms of HIV-1 DNA in cellular reservoir and anatomical sanctuary sites. These viral reservoirs are one of the major obstacles in HIV-1 eradica-tion [9-12, 16-36].
In this review, we discuss numerous PCR-based methods developed for the quantification of cell-associated HIV-1 DNA. We also present recent findings about the prognostic value of HIV-1 DNA as a marker to disease progression, treatment initiation and prolonged efficacy of HAART.
HIV-1 DNA AND THE VIRUS LIFE CYCLE
Viruses depend on host’s cellular machinery to dis-seminate their genetic information. The human immunodefi-ciency virus (HIV), a prototype of the lentiviral subfamily of retroviruses, is one of the most efficient viruses in exploiting the host cell’s functions. The life cycle begins with the re-verse transcription of genomic viral RNA into linear uninte-grated HIV-1 DNA. Completion of this reverse transcription step yields the so-called preintegration complex (PIC) which contains viral dsDNA (reverse transcription’s product), vi-rus-derived proteins including integrase (IN) and host pro-teins i.e., HMGa1 and BAF [37-41]. The PIC can be isolated from the cytoplasmic compartment of infected cells and is
*Address correspondence to these authors at the Department of Hygiene,
Epidemiology and Medical Statistics, Medical School, University of Athens,
M. Asias 75, GR-115 27, Athens, Greece; Tel: (+30210) 7462090; Fax:
(+30210) 7462190; E-mail: [email protected]; Tel: (+30210) 7486382;
Fax: (+30210) 7486382; E-mail: [email protected]
capable of integrating its dsDNA into the target’s host DNA genome to establish a stable provirus [41-44].
Although the linear dsDNA molecule is the precursor to the provirus, an infection of the target cells generates a num-ber of non-integrated DNA species [45-49]. These several unintegrated HIV-1 DNA forms, linear or circularized, arise when viral dsDNA fails to integrate the host genome. For instance, (i) the ends of the linear DNA may be joined to form a 2-LTR circle; (ii) homologous recombination be-tween the two LTRs in a 2-LTR circle yields circles with a single LTR (1-LTR); (iii) linear unintegrated HIV-1 DNA form (Fig. 1). Depending on the site of integration and sub-sequent recombination a series of complex DNA structures may result, as well [50]. Non-integrated forms represent the largest fraction of HIV-1 DNA in the nucleus. Several stud-ies have suggested that 2-LTR circular HIV-1 DNA forms might used as a marker of recent cell infection on the basis of their observed lability in vitro and in vivo [51-54]. This, conflicts with other studies, which reported that are highly stable [54-56]. These unintegrated viral DNA forms have been shown to be incapable of directing a productive infec-tion but seem to be responsible for residual viral replication [34, 36, 43, 47, 49]. However, it remains controversial whether these unintegrated viral DNA forms play some role in the HIV replication cycle [57-60]. Therefore, except pro-viral DNA (dsHIV-1 DNA integrated in the host’s genome), several unintegrated forms persist in intracellular HIV reser-voirs.
ASSAYS DEVELOPED FOR HIV-1 DNA QUANTIFI-CATION
Several assays have been developed for the quantification of cell-associated HIV-1 DNA based on endpoint and real-time PCR methodology. The latter have shown improved accuracy and reproducibility and a wider range of linear quantification since the concentration of the target is esti-mated during the linear phase rather than at the endpoint (plateau phase) of the PCR that is used in conventional end-point PCR methods. In this paragraph we review the major

256 Current HIV Research, 2009, Vol. 7, No. 3 Beloukas et al.
features of the different assays with special emphasis to the most recently developed real-time PCR methods.
1) End-Point PCR Assays
Numerous conventional (endpoint) PCR-based assays for quantifying cell-associated HIV-1 DNA have been devel-oped [15, 49, 61-81]. All these assays, including externally controlled PCR [5, 61-63, 69, 71, 82] and internally con-trolled PCR [64, 68, 83], implementing competitive PCR methodology [72, 73, 75, 78, 80, 81] or not, require specific optimization. These methods are also prone to intralabora-tory or interlaboratory variations due to the lack of standard-ized reagents. Conventional PCR assays demand detection and quantification of amplified products after the reaction procedure. These assays that have been reported use a vari-ety of methods for detection and quantification of the PCR product. For example, amplicons are often quantified by ra-dioisotope detection (Southern hybridization) [64, 65, 70, 72,
80, 83-85], by enzyme-linked methods [49, 61, 67, 68, 79], or using Ethidium-Bromide [73, 81].
Quantification of PCR products in all these assays using conventional PCR is achieved by comparing the strength of the signal from the standards (external or/and internal) with known concentrations that are quantified at the same time and the unknown samples. Conventional PCR based methods have the disadvantage of the plateau phase that conventional PCR reaches. In order to avoid the plateau phase and main-tain exponential phase of amplification, most assays perform two sequential amplifications with a low number of repeated cycles in the first and the second PCR (nested-PCR) [15, 49, 70, 74], while other semi-quantitative assays, limited dilution of PCR products occurred after end-point [65, 74, 83].
Christopherson et al. reported a modification of the Am-plicor HIV-1 Monitor test (Roche Molecular Systems) for total HIV-1 DNA quantification. This assay based on an end-point PCR measurement assay is almost identical to the Am-
Fig. (1). Intracellular HIV-1 DNA forms. HIV-1 DNA into host’s cell nucleus persists in proviral form (integrated linear HIV-1 dsDNA) and
several unintegrated forms, linear or/and circulazied, such as 1-LTR and 2-LTR HIV-1 DNA, as well.

The Role of HIV-1 DNA Marker Current HIV Research, 2009, Vol. 7, No. 3 257
plicor HIV-1 Monitor assay for RNA (Roche Molecular Sys-tems) but differs in (i) the sample preparation method, (ii) the use of plasmid DNA rather than an RNA transcript as the quantitation standard and (iii) the normalization of the DNA load to total cellular input. In this format, HIV-1 DNA is expressed as copies per microgram of total DNA and con-verted to copies per 10
6 PBMC by using a standard conver-
sion formula. The sensitivity of this method has been re-ported to be 10 copies/reaction or 5 copies/10
6 PBMC and
has a five-log10 detection range in DNA standards [86].
Another similar assay was reported by Lillo et al., in which an internal standard HIV-1 DNA obtained in vitro from the 8E5 cell line that was used [87]. Interestingly, Jennings et al. proposed the use of the Amplicor HIV-1 DNA Test in whole blood instead of isolated cells, if blood samples had been maintained at 4-25
o C and were less than
10 days old, but the specimens should be used as soon as possible after collection [88].
2) Real-Time PCR Assays
Implementation of real-time PCR considerably improved the quantification of linear or circular forms of HIV-1 DNA compared to the end-point PCR methodology based on dif-ferent platforms and detection chemistry. However, there is not yet any universal way to report HIV-1 DNA values. Spe-cifically in most of the assays HIV-1 DNA is reported as DNA copies per 10
6 PBMCs [6, 15, 89-95], or alternatively
as HIV-DNA copy numbers per number of resting CD4+
T cells [96, 97]. According to the described methodology for the estimation of the number of cells, previously developed HIV-1 DNA real-time PCR assays have been divided into 2 categories: A) direct methods are assays that direct estimate the number of cells from which HIV-1 DNA is quantified; direct estimations are based on quantification, in addition to the HIV-1 cellular load, of a housekeeping gene existing in a standard stoichiometric ratio in human cells (reporter gene); B) indirect methods estimate the amount of cells by using standard colorimetric methods; these are subsequently con-verted to cell copy numbers by a conversion formula.
In summary, among all the previously described meth-ods, the sensitivity for HIV-1 DNA was 2-10 copies/reaction [30, 86, 89-92, 94, 95, 98, 99], while for a single assay the sensitivity of HIV-1 DNA was reported 50 HIV-1 DNA cop-ies/CD4
+ T-cells [97]. The linear range of HIV-1 DNA quan-
tification in clinical samples tested for eight methods ranged between 2.0-2.5 log10 values [86, 90, 92, 94, 95, 97, 98], except from three assays for which HIV-1 DNA was quanti-fied within 3.0-3.5 log10 [89, 91, 99]. Additionally, the per-centage of samples with detectable HIV-1 DNA was high (89-100%) expect from a lower value (64%) reported for a single assay [97]. Both direct and indirect methods used dif-ferent methodology for the estimation of the number of cells or chromosomal DNA used in the HIV-1 DNA quantifica-tion experiments. The characteristics of all previous assays are reviewed in detail in the following paragraphs and shown in Table 1.
A) Direct Methods
As mentioned previously, in direct methods the number of cells from which the quantified HIV-DNA was obtained,
was estimated by direct amplification of a reporter gene in parallel or in a multiplex setting.
In particular, Desire et al. developed an assay using TaqMan probes methodology targeting HIV-1 pol region, however the reporter gene (albumin) was quantified in a separate reaction. Moreover, to normalize variations due to differences in the starting volume of PBMCs, an internal standard was used as an endogenous reference. Both stan-dards were constructed by cloning after PCR amplification of the two targets: a) intron 12 of the human albumin (re-porter gene) and b) the HIV-1 pol gene obtained from the LAVBRU strain. The normalized value of the HIV-1 proviral load was expressed as the number of HIV-1 copies per 10
6
PBMCs (Table 1) [99].
Kostrikis et al. developed three separate assays imple-menting molecular beacons [92] methodoly for real-time detection of amplicons: one was for detection and quantifica-tion of PBMC-associated HIV-1 DNA forms that have un-dergone the second template switch (HIV-1 STS DNA) (this assay mainly detects a pool of all HIV-1 forms including unintegrated and integrated linear dsDNA viral genomes and 1- and 2-LTR circles), a 2-LTR-specific (Table 2) and CCR5-specific assays in order to express HIV-1 DNA to be expressed per 10
6 PBMC (according to experimental evi-
dence, there is a steady stoichiometric ratio of CCR5 copies per cell). The analytical sensitivity of the three assays have been found to be similar (Tables 1 and 2) [92].
Eriksson et al. reported the first multiplex TaqMan-based real-time PCR targeting HIV-1 pol gene and, also in parallel, the albumin housekeeping gene (in multiplex setting). By this way the quantified amount HIV-1 DNA can be directly expresses as DNA copies per number of cells (Table 1) [90].
Kabamba et al. reported an assay implementing SYBR green methodology for amplicons quantification. The esti-mated HIV-1 DNA levels were reported as HIV-1 DNA cop-ies per CD4
+ counts (b-globin reference gene was amplified
in parallel in order CD4+ cell sample to be quantified) (Table
1) [97].
Finally, Beloukas and Paraskevis et al. reported the first multiplex molecular beacon based real-time PCR assay that quantifies all HIV-1 DNA forms (ss and ds DNA forms, inte-grated and unintegrated, linear or circularized) (Fig. 1) and the number of PBMCs co-instantaneously using CCR5 as reporter gene for which there is a steady number of copies per cell as described previously [92]. Specifically, primers and molecular beacon for HIV-1 DNA amplification and detection were newly designed targeting the conserved region downstream to LTR and gag. Furthermore, the target recognition sequence for the CCR5_beacon did not contain the 32 deletion of ccr5 and therefore it can bind to both the wild-type ccr5 and mutant ccr5 32 alleles. Noteworthy is that although this assay quanti-fies HIV-1 DNA in parallel with the reference gene that is abundant compared to the viral DNA, the sensitivity of the multiplex method was at the same order as for the single Real-Time PCR assays (Table 1) [89].
B) Indirect Methods
Valentin et al. reported a real-time PCR assay for the quantification of circularized HIV-1 genomes containing two

258 Current HIV Research, 2009, Vol. 7, No. 3 Beloukas et al.
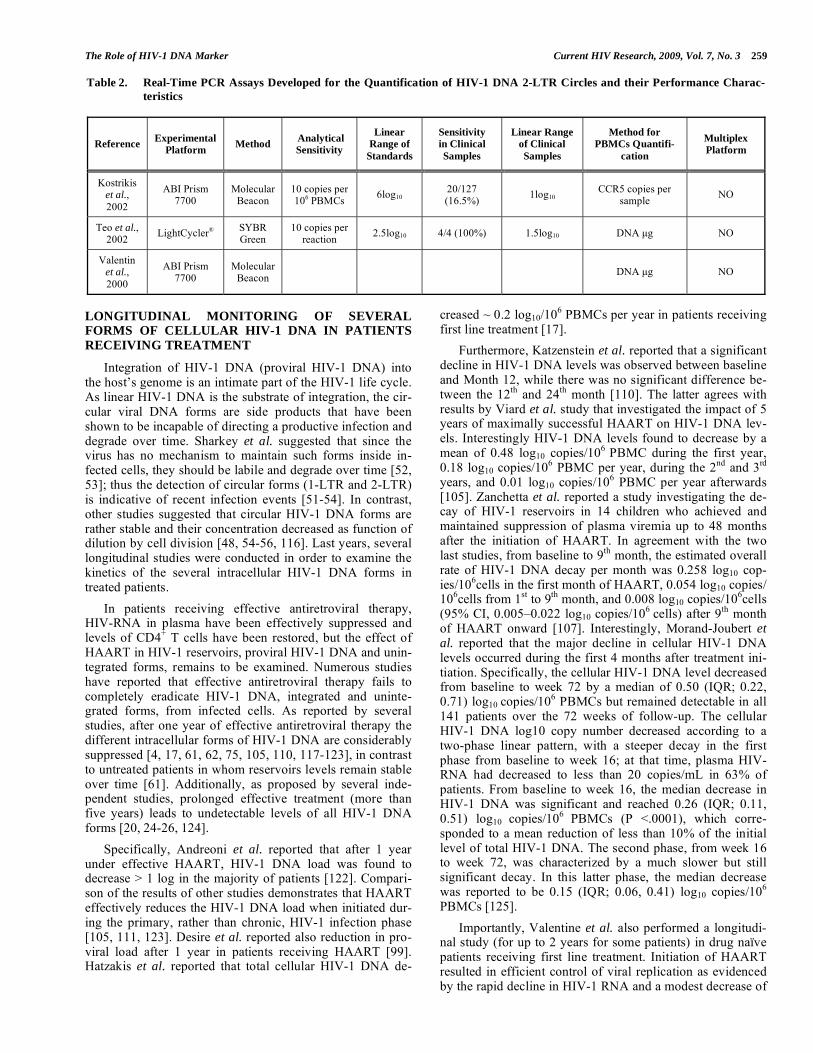
long-terminal repeats (2-LTR circle), targeting the LTR-U5 region. The 2-LTR copy number in the sample obtained by comparison with a standard curve derived from known DNA standards and was expressed as copies of 2-LTR copies per nanogram of cellular DNA (Table 2) [30]. This assay was the first one reported for real-time quantification of 2-LTR circularized intracellular HIV-1 DNA; however until now several others assays have been developed for the quantifica-tion of 2-LTR HIV-DNA [52, 54, 76, 85, 92, 100-103].
Yun et al. reported an assay targeted the terminal repeat region (LTR) of the HIV-1 genome and the assay reported by Zhao et al. targeted the HIV-1 gag gene [94, 95, 104]. Another TaqMan probe-based real-time PCR assay was re-ported by Viard et al., targeting the long terminal repeat (LTR) HIV-1 genome; the sensitivity was 2 copies per reac-tion (Table 1) [105]. This assay was modified by Samarti et al., in order to be implemented in LightCycler platform (Roche Molecular Biochemicals, Indianapolis, IN) with similar characteristics [25, 98]. In the latter assay, a fragment of human b-globine was used in order to verify DNA integ-rity, but the results were expressed as HIV-DNA per 10
6
PBMC since extracted DNA was obtained from dry pellets of 2 • 10
6 PBMCs.
Finally, two other methodologies were developed, using SYBR green for real-time PCR amplicons detection reported by Gibellini et al. and Casabianca et al., respectively (Table 1) [98, 106]. The performance characteristics of the two as-says reported to be similar (Table 1).
In summary in most of the previous assays, real-time PCR targeted partial gag [92, 95, 98, 105, 107], pol [90, 99, 106] or LTR regions [89, 94, 106, 108-110]. We should note that unless PCR targets the 2-LTR junction, any other PCR format priming within the HIV-1 genome will potentially amplify all different forms of intracellular HIV-1 DNA (Fig. 1). Most of the assays 2-LTR circular DNA [5, 30, 31, 36, 51-53, 56, 72, 76, 92, 93, 103, 106, 108, 109, 111, 112], have not implemented real-time methodology, except from assays reported by Kostrikis et al, Valentin et al. and Teo et al. (Table 2) [30, 92, 103]. In addition to the standard assays quantifying all different forms of intracellular HIV-1 DNA, attempts have been made towards specific detection of inte-grated viral DNA. The design was based on a PCR targeting one of the most abundant repeats (Alu elements) in human DNA; however these assays (Alu-HIV-1 DNA PCR assays) have not been used extensively for the assessment of the integrated HIV-1 DNA during the course of HIV-1 infection [49, 56, 77, 108, 113-115].
Table 1. Real-Time PCR Assays Developed for the Quantification of Total HIV-1 DNA and their Performance Characteristics
Reference Experimental
Platform Method
Analytical
Sensitivity
Linear
Range of
Standards
Sensitivity
in Clinical
Samples
Linear Range of
Clinical Samples
Method for
PBMCs Quanti-
fication
Multiplex
Platform
Beloukas, Paraskevis et al.,
(submitted)
LightCycler® 1.0 and 2.0
Molecular Beacon
12.5 copies per 106
PBMCs
6log10 115/115 (100%)
3log10 CCR5 copies per
sample YES
Casabianca et al., 2007
ABI Prism 7700 SYBR Green
2copies per reaction
(2copies per
15·104 PBMC)
6log10 72/73 (99%)
2log10 Flow cytometry NO
Kabamba-Mukadi et al., 2005
LightCycler® 1.0 SYBR Green
50 copies HIV-1 DNA
per CD4+
6log10 29/45 (64%)
2.5log10
Purified CD4+ Albumin - Quan-
tifying known dilutions of
8E5LAV cells
NO
Gibellini et al., 2004
LightCycler® 1.0 SYBR Green
5 copies per reaction
5log10 50/50
(100%) 3log10
Quantifying known dilutions of 8E5LAV cells
NO
Eriksson et al., 2003
ABI Prism 7700 TaqMan probes
4log10 8/8
(100%) 2log10 Albumin gene YES
Kostrikis et al., 2002
ABI Prism 7700 Molecular
Beacon 10 copies per 106 PBMCs
6log10 113/127 (89%)
2log10 CCR5 copies per
sample NO
Zhao et al., 2002 ABI Prism 7700 TaqMan probes
10 copies per reaction
>6log10 80/80
(100%) 2log10 DNA μg NO
Yun et al., 2002 ABI Prism 7700 TaqMan probes
5 copies per reaction
DNA μg NO
Desire et al., 2001 ABI Prism 7700 TaqMan probes
10 copies per reaction
6log10 36/37 (97%)
3.5log10 Internal standard
Albumin gene NO
Christopherson et al., 2000* Cobas TaqMan
Amplicor HIV-1
Monitor
10 copies per reaction
5log10 19/19
(100%) 2log10 Hoechst Dye NO
*Not a real-time PCR assay that is commercially available.
The Role of HIV-1 DNA Marker Current HIV Research, 2009, Vol. 7, No. 3 259
LONGITUDINAL MONITORING OF SEVERAL FORMS OF CELLULAR HIV-1 DNA IN PATIENTS
RECEIVING TREATMENT
Integration of HIV-1 DNA (proviral HIV-1 DNA) into the host’s genome is an intimate part of the HIV-1 life cycle. As linear HIV-1 DNA is the substrate of integration, the cir-cular viral DNA forms are side products that have been shown to be incapable of directing a productive infection and degrade over time. Sharkey et al. suggested that since the virus has no mechanism to maintain such forms inside in-fected cells, they should be labile and degrade over time [52, 53]; thus the detection of circular forms (1-LTR and 2-LTR) is indicative of recent infection events [51-54]. In contrast, other studies suggested that circular HIV-1 DNA forms are rather stable and their concentration decreased as function of dilution by cell division [48, 54-56, 116]. Last years, several longitudinal studies were conducted in order to examine the kinetics of the several intracellular HIV-1 DNA forms in treated patients.
In patients receiving effective antiretroviral therapy, HIV-RNA in plasma have been effectively suppressed and levels of CD4
+ T cells have been restored, but the effect of
HAART in HIV-1 reservoirs, proviral HIV-1 DNA and unin-tegrated forms, remains to be examined. Numerous studies have reported that effective antiretroviral therapy fails to completely eradicate HIV-1 DNA, integrated and uninte-grated forms, from infected cells. As reported by several studies, after one year of effective antiretroviral therapy the different intracellular forms of HIV-1 DNA are considerably suppressed [4, 17, 61, 62, 75, 105, 110, 117-123], in contrast to untreated patients in whom reservoirs levels remain stable over time [61]. Additionally, as proposed by several inde-pendent studies, prolonged effective treatment (more than five years) leads to undetectable levels of all HIV-1 DNA forms [20, 24-26, 124].
Specifically, Andreoni et al. reported that after 1 year under effective HAART, HIV-1 DNA load was found to decrease > 1 log in the majority of patients [122]. Compari-son of the results of other studies demonstrates that HAART effectively reduces the HIV-1 DNA load when initiated dur-ing the primary, rather than chronic, HIV-1 infection phase [105, 111, 123]. Desire et al. reported also reduction in pro-viral load after 1 year in patients receiving HAART [99]. Hatzakis et al. reported that total cellular HIV-1 DNA de-
creased ~ 0.2 log10/106 PBMCs per year in patients receiving
first line treatment [17].
Furthermore, Katzenstein et al. reported that a significant decline in HIV-1 DNA levels was observed between baseline and Month 12, while there was no significant difference be-tween the 12
th and 24
th month [110]. The latter agrees with
results by Viard et al. study that investigated the impact of 5 years of maximally successful HAART on HIV-1 DNA lev-els. Interestingly HIV-1 DNA levels found to decrease by a mean of 0.48 log10 copies/10
6 PBMC during the first year,
0.18 log10 copies/106 PBMC per year, during the 2
nd and 3
rd
years, and 0.01 log10 copies/106 PBMC per year afterwards
[105]. Zanchetta et al. reported a study investigating the de-cay of HIV-1 reservoirs in 14 children who achieved and maintained suppression of plasma viremia up to 48 months after the initiation of HAART. In agreement with the two last studies, from baseline to 9
th month, the estimated overall
rate of HIV-1 DNA decay per month was 0.258 log10 cop-ies/10
6cells in the first month of HAART, 0.054 log10 copies/
106cells from 1
st to 9
th month, and 0.008 log10 copies/10
6cells
(95% CI, 0.005–0.022 log10 copies/106
cells) after 9th
month of HAART onward [107]. Interestingly, Morand-Joubert et al. reported that the major decline in cellular HIV-1 DNA levels occurred during the first 4 months after treatment ini-tiation. Specifically, the cellular HIV-1 DNA level decreased from baseline to week 72 by a median of 0.50 (IQR; 0.22, 0.71) log10 copies/10
6 PBMCs but remained detectable in all
141 patients over the 72 weeks of follow-up. The cellular HIV-1 DNA log10 copy number decreased according to a two-phase linear pattern, with a steeper decay in the first phase from baseline to week 16; at that time, plasma HIV-RNA had decreased to less than 20 copies/mL in 63% of patients. From baseline to week 16, the median decrease in HIV-1 DNA was significant and reached 0.26 (IQR; 0.11, 0.51) log10 copies/10
6 PBMCs (P <.0001), which corre-
sponded to a mean reduction of less than 10% of the initial level of total HIV-1 DNA. The second phase, from week 16 to week 72, was characterized by a much slower but still significant decay. In this latter phase, the median decrease was reported to be 0.15 (IQR; 0.06, 0.41) log10 copies/10
6
PBMCs [125].
Importantly, Valentine et al. also performed a longitudi-nal study (for up to 2 years for some patients) in drug naïve patients receiving first line treatment. Initiation of HAART resulted in efficient control of viral replication as evidenced by the rapid decline in HIV-1 RNA and a modest decrease of
Table 2. Real-Time PCR Assays Developed for the Quantification of HIV-1 DNA 2-LTR Circles and their Performance Charac-
teristics
Reference Experimental
Platform Method
Analytical
Sensitivity
Linear
Range of
Standards
Sensitivity
in Clinical
Samples
Linear Range
of Clinical
Samples
Method for
PBMCs Quantifi-
cation
Multiplex
Platform
Kostrikis et al.,
2002
ABI Prism 7700
Molecular Beacon
10 copies per 106 PBMCs
6log10 20/127 (16.5%)
1log10 CCR5 copies per
sample NO
Teo et al., 2002
LightCycler® SYBR Green
10 copies per reaction
2.5log10 4/4 (100%) 1.5log10 DNA μg NO
Valentin et al.,
2000
ABI Prism 7700
Molecular Beacon
DNA μg NO

260 Current HIV Research, 2009, Vol. 7, No. 3 Beloukas et al.
HIV-1 DNA copies in NK cells and total T lymphocytes as well; however HIV-1 was not eliminated from either total T cells or NK cells. The DNA decay slopes in T cells and NK cells, respectively, demonstrated significant differences; es-pecially in NK cells, HIV-1 DNA was detected at higher levels than total T cells. This difference may reflect a more stable reservoir in NK cells compared with total T cells. Al-ternatively, it may reflect changes in the kinetics of the dif-ferent cell populations during treatment [31].
Morlese et al. reported the first study for the longitudinal monitoring of 2-LTR HIV-1 DNA circular form in 60 drug-naïve patients initiating HAART. After starting antiretroviral therapy a decrease in the number of HIV-1 DNA 2-LTR cop-ies was observed in all patients enrolled in the study. Their results showed that there are two different groups of patients. In the first group, 2-LTR circles remained undetectable for many months given that the plasma HIV RNA also remained undetectable. In contrast, in the second group, 2-LTR circles were consistently detected and their presence was associated with the isolation of infectious virus from peripheral blood [51].
McDermott et al. reported on a longitudinal study moni-toring levels of total HIV-1 DNA and 2-LTR circles from 22 patients under HAART for a period of 8 years with sustained undetectable HIV RNA. As McDermott and colleagues men-tioned, 55% of the samples examined had detectable total DNA levels, even after 8 years. In contrast, although 2-LTR HIV-1 DNA was detectable in approximately 50% of the samples during the first 3 years, the percentage of positive samples dropped drastically to 6% at the end of the forthy year and 2-LTR HIV-1 DNA remained detectable in a small percentage of samples until the end of the study period. Overall, the longitudinal monitoring of total HIV-1 DNA and unintegrated 2-LTR circles in patients revealed a general decay for HIV-1 DNA levels to undetectable levels [112].
CLINICAL UTILITY OF HIV-1 DNA LEVELS
The persistence of HIV-1 DNA in viral reservoirs after effective HAART treatment provides the main obstacle to virus eradication, and suggests that even more-potent treat-ment will not be sufficient to clear the infection. There is accumulating evidence that HIV-1 DNA could be used in clinical practice as a marker of disease progression, assess-ment of HAART initiation and long-term success of treat-ment.
Prognosis of Disease Progression
Kostrikis et al. reported that the quantification of all HIV-1 DNA forms that have undergone Second Template Switch (STS) in PBMCs is an independent marker for dis-ease progression to clinical AIDS. The study was conducted using samples from 130 HIV-1-infected hemophiliacs with known HIV-1 seroconversion dates, followed up prospec-tively for more than 18 years after seroconversion. Particu-larly, STS HIV-1 DNA baseline levels were found to be moderately correlated with HIV-RNA levels (r=.38; P<.0001) and weakly correlated with CD4
+ T cell count (r=-
.27; P<.007). STS HIV-1 DNA levels were found to be higher in subjects that progressed to AIDS, in contrast with those who remained AIDS free. Interestingly, its concentra-
tion remained constant during the natural course of HIV-1 infection [92].
A study reported by Tierney et al. investigated the prog-nostic value of a baseline HIV-1 DNA level for disease pro-gression in 111 patients receiving HAART. The baseline HIV-1 DNA level was significantly associated with the base-line HIV-RNA level (r=.61; P<.01), but not with CD4
+ T-cell
count (r=-.14; P=.16). Kaplan-Meier estimates for disease progression indicated an association between higher (above median) baseline HIV-1 DNA level and more rapid disease progression (P<.01) [126].
Rouzioux et al. reported that baseline levels of HIV-1 DNA from 383 individuals during the first six months after primary infection was found to be a major predictive factor of progression to clinical AIDS, to a CD4
+ T cell count
<200/μl and to death. Especially, the predictive value of the baseline HIV-1 DNA level was found to be independent of the two other markers (CD4
+ T cell count, and HIV RNA
level). Specifically, patients with HIV-1 DNA levels 3log10 copies/10
6 PBMCs progressed to clinical AIDS more rapidly
than did those with HIV-1 DNA levels < 3log10 copies/106
PBMCs, independently of CD4+ T cell count and/or HIV-
RNA levels. Furthermore, baseline HIV-1 DNA levels dur-ing the plateau phase (between 6 and 24 months after sero-conversion) were found to be highly correlated with baseline HIV RNA levels (r=.69; P<.0001) and with CD4
+ T cell
counts (r=-.40; P<.0001), suggesting that a combination of these three markers could provide the most accurate estimate of the risk of disease progression [21].
Another study reported by Goujard et al. also demon-strated that the baseline HIV-1 DNA level was an independ-ent predictive marker for disease progression in 163 un-treated patients with primary infections. Particularly, patients with disease progression to AIDS had a significantly higher median baseline HIV-1 DNA level than those without pro-gression (3.3 vs 3.0 log10 copies/10
6 PBMCs; P<.003).
Moreover, Kaplan-Meier estimates revealed that a baseline HIV-1 DNA level >3.4 log10 copies/10
6 PBMCs (corre-
sponding to the 66th percentile value) was associated with a 62% risk of progression to AIDS in two years (95% CI, 44%–80%), compared with 13% (95% CI, 2%–23%) among patients with values <2.9 log10 copies/10
6 PBMCs (corre-
sponding to the 33rd percentile value) providing additional information at the time of acute infection [127].
Finally, Goedert et al. reported that a higher level of the HIV-1 2-LTR circulating form, which represents the number of PBMCs that are infected with actively replicating HIV-1, in conjunction with lower T cell receptor rearrangement ex-cision DNA circles (TRECs) levels were associated with increased risk of AIDS in non-treated patients [100]. The study was conducted in a cohort of 154 patients with known seroconversion dates who became infected between 1978 and 1985 and were followed up prospectively.
In conclusion, all these studies revealed that HIV-1 DNA levels at baseline are highly correlated with the rate of pro-gression to AIDS in patients with known or unknown sero-conversion dates, irrespective of the HIV-RNA level and CD4
+ T cell counts. Therefore, in addition to the latter mark-
ers, HIV-1 DNA can be utilized as an essential tool to pre-dict disease progression in HIV-infected patients.

The Role of HIV-1 DNA Marker Current HIV Research, 2009, Vol. 7, No. 3 261
Assessment of HAART Initiation
In HAART era, an issue of critical importance concerns the decision of the optimal time to initiate therapy [128, 129]. Although the effectiveness of HAART in delaying progression to AIDS or death has been convincingly demon-strated, prior to treatment initiation, a long-term treatment strategy plan should be designed, including considerations of the possibility of intolerance, long-term drug toxicity and treatment failure due to resistance [130-132]. Rouzioux et al. on behalf of the SEROCO Cohort study group investigated the potential value of HIV-1 DNA level, as an additional marker to define the best time to initiate treatment. In accor-dance with the guidelines currently used in North America and Europe, which are mainly based on CD4
+ T cell counts
and HIV-RNA levels, 271 previously untreated patients, at the plateau phase of HIV infection, were divided into three groups, as follows: Group 1, patients with CD4
+ T cell
counts <350 cells/μL (treatment or close follow-up recom-mended); Group 2, patients with CD4
+ T cell counts 350
cells/μL and serum HIV-RNA levels 15,000 copies/μL (treatment postponed); and Group 3, patients with CD4
+ T
cell counts 350 cells/μL and serum HIV-RNA levels <15,000 copies/μL (treatment not recommended). The risk of progression to clinical AIDS based on these two markers was estimated calculating Kaplan-Meier estimates of the probability of developing clinical AIDS at five years after infection and found to be 39% for patients in Group 1, 26% for patients in Group 2, and 11% for patients in Group 3. Additionally, the risk of progression to AIDS at five years after infection was high (>25%) for all patients with HIV DNA levels of 3 log10 copies/10
6 PBMCs, including pa-
tients from Group 3, who would not be treated on the basis of current guidelines [21]. Since HIV infection leads to AIDS through a continuous and progressive process, it is difficult to design treatment guidelines only on the basis of clear-cut values, such as 350 CD4
+ T-cells/μL or 100,000,
50,000, or 30,000 copies of HIV-RNA/μL in plasma. As Rouzioux et al. study suggests, it would be useful to com-bine all 3 markers when trying to obtain a better picture of the underlying dynamics of HIV disease for each patient, especially for decisions regarding treatment initiation [21].
Therefore, HIV-1 DNA levels could guide clinical prac-tice in order to define the best time to initiate treatment tai-lored to each patient, even in acute or chronic phase of HIV-1 infection.
Prediction for the Long-Term Success of Treatment
Hoen et al. on behalf of the QUEST Study group investi-gated several predictors of virological outcome in patients with a primary infection initiating quadruple antiretroviral therapy. Cell-associated HIV-1 DNA was found to be an independent predictor (multivariable model) of virological response (viral load 3 copies/ml) among 103 patients [118].
Hatzakis et al. assessed whether baseline cellular HIV-1 DNA levels in 51 HIV-1-infected individuals (prior to HAART initiation) was a predictive marker for the outcome of HAART and HIV-RNA rebound and/or sustained vi-rological response (VR). Cellular HIV-1 DNA baseline load was the only parameter that was significantly associated with viral rebound in patients showing an initial decline in HIV-
RNA levels. The median (IQR) baseline HIV-1 DNA load level was 1338 (1083–1374) and 91 (39–459) copies/10
6
PBMC in treated patients with virological rebound and sus-tained VR, respectively (P=.002) (median value of all esti-mated subjects was 297 copies/10
6 PBMC). Furthermore,
none of the initial responders with baseline HIV-1 DNA load below or equal to the median value (297 copies/10
6 PBMC)
experienced a subsequent virologic rebound, while among those with HIV-1 DNA load greater than 297 copies/10
6
PBMC, the cumulative probability for virological rebound by week 104, was 54.6% (P=.008). Cellular HIV-1 DNA baseline level was the only parameter associated with sus-tained VR, even after adjustment for the other virological, immunological and treatment factors in a multivariate analy-sis [17].
As have been reported, in HIV-infected patients success-fully treated with HAART, a low level of HIV-RNA persists in plasma at a steady state for years and varies among pa-tients [133-137]. The only independent predictor of this re-sidual viremia found to be pre-HAART levels of HIV-1 DNA [11, 12, 26, 136].
Levels of cell-associated HIV-1 DNA are a usefull marker with regard to treatment modifications in patients with discordant responses to HAART. Marchetti et al. inve-stigated several parameters in HIV-1 infected patients with discordant immune or virological responses to HAART. There were three seperate groups of patients: Immunological non-responders (INR; CD4
+ T cell count, 200 cells/ml;
HIV-RNA, 50 copies/ml), virological non-responders (VNR; CD4
+ T cell count, 200 cells/ml; HIV-RNA,
10,000 copies/ml) and full responders (FR; CD4+ T cell
count, 500 cells/ml; HIV-RNA, 50 copies/ml). Com-pared to VNR, FR presented a trend toward the lowest me-dian CD4
+ T cell HIV-1 DNA, within both memory cells
(FR, 90 copies/104 cells; VNR, 279 copies/10
4 cells) and
naive CD4+ T cells (FR, 35 copies/10
4 cells; VNR, 128 cop-
ies/104 cells; P >.05 in each pair-wise comparison). Despite
full HIV viremia suppression, INR displayed a consistent trend toward the highest median intracellular HIV-1 DNA levels in total CD4
+ T cell and, most importantly, within
both memory and naive cells (441 copies/104 and 161 cop-
ies/104 cells, respectively) versus either FR or VNR, (P >.05
in each pair-wise comparison; P =.09 for INR versus FR). Despite the low number of patients, these data suggest an enhanced rate of HIV infection within the whole CD4
+ T-
lymphocyte compartment in INR [138]. This finding strongly supports the hypothesis of ongoing residual low-level viremia, not detectable by standard assays in INR [96, 139].
Samarti et al. reported a study enrolled 62 patients to evaluate the total HIV-1 DNA quantification as a predictor of long-term success for Protease Inhibitor-sparing simpli-fied therapy, in order to overcome PI-associated side-effects (e.g., pills burden, lipid abnormalities, or clinical lipodystro-phy). Patients were categorized in two stratums; stratum A enrolled patients with cellular HIV-1 DNA levels above the median value (median value from all subjects estimated 226 copies per 10
6 PBMCs) (32, 51.6%) and stratum B with val-
ues below the median value (30, 48.4%). At month 18, 53 of 62 (85%) patients receiving the simplified regimen were considered to have virological success, 4 (6.4%) patients had

262 Current HIV Research, 2009, Vol. 7, No. 3 Beloukas et al.
virological failure, and 5 (8%) patients had viral blip. All but one patient with virological failure or blip had more than 226 HIV-1 DNA copies/10
6 PBMCs (P<.009). At the end of the
follow-up period, patients with virological success had sig-nificantly lower HIV-1 DNA levels with respect to patients with virological failure or blip (P<.001) [25].
Lafeuillade et al. reported a small study of 30 HIV-1 in-fected patients receiving HAART for up to 24 months, fol-lowed by 1-3 structured treatment interruptions (STIs). In this study the probability of viral control after STI, as esti-mated by Kaplan-Meier curves, was higher in patients with HIV-1 DNA levels less or equal than median values, at Month 24 [120].
Another study reported by Yerly et al. included 87 pa-tients receiving HAART without previous failure and having undetectable viremia for at least 6 months. HAART was in-terrupted for 2 weeks and restarted for 8 weeks (STI). After four repeated STIs, treatment was definitively interrupted at week 40. Virological failure was defined for 0-40 week pe-riod by HIV-RNA levels greater than 50 copies/ml after 8 weeks of re-treatment. In univariate analysis, virological failure was found to be significantly associated with pre-HAART viremia, pre-HAART CD4
+ T-cell count and provi-
ral DNA level at baseline, but not with the baseline CD4+ T
cell count (P=.48) [140].
CONCLUSIONS
Intracellular HIV-1 DNA provides a new marker for bet-ter monitoring of HIV-1 infections in addition to CD4
+ T cell
counts and HIV-RNA levels. Until now, a variety of assays have been used for the quantification of the HIV-1 DNA levels, implemented conventional or real-time PCR method-ology. Those based on real-time methodology have shown improved accuracy, reproducibility and a wider range of linear quantification. It is worth mentioning that there is not yet a universal format expressing the levels of cell-associated HIV-1 DNA, although most of the existing assays report HIV-1 DNA levels in copies per 10
6 PBMCs.
We reviewed recent findings about the potential conse-quences of cellular HIV-1 DNA levels as an independent prognostic marker for clinical progression of HIV-1 infected patients.
HIV-1 DNA can add important clinical information to HIV-RNA and CD4
+ T cell values, with regard to the opti-
mum time for treatment initiation. Specifically, HIV-1 DNA can be predictive marker of the risk for developing AIDS, so it facilitates decision making in patients not fulfilling current criteria for treatment (e.g. CD4
+ T cells 350 cells/μL and
HIV-RNA<15,000 copies/mL). Importantly, HIV-1 DNA testing before treatment initiation would add clinical value, in predicting long-term treatment outcomes. This is an im-portant advance for tailoring HIV-therapy. Additionally, HIV-1 DNA could guide treatment modification in patients with partial or incomplete immunological response to HAART; assessing each patient’s need to undergo treatment intensification due to ongoing viral replication despite unde-tectable levels of HIV-RNA. Similarly, HIV-1 DNA can be used to predict long-term success to simplification strategies such as Protease Inhibitor-sparing therapies. In conclusion, HIV-1 DNA testing is expected to be an important marker, in addition to those currently used, for optimizing therapy
initiation, predicting treatment outcome and tailoring treat-ment in patients with partial or complete response.
REFERENCES
[1] Mocroft A, Gill MJ, Davidson W, Phillips AN. Predictors of a viral response and subsequent virological treatment failure in patients
with HIV starting a protease inhibitor. AIDS 1998; 12: 2161-7. [2] Hogg RS, Yip B, Kully C, et al. Improved survival among HIV-
infected patients after initiation of triple-drug antiretroviral regi-mens. CMAJ 1999; 160: 659-65.
[3] Ledergerber B, Egger M, Opravil M, et al. Clinical progression and virological failure on highly active antiretroviral therapy in HIV-1
patients: a prospective cohort study. Swiss HIV Cohort Study. Lan-cet 1999; 353: 863-8.
[4] Egger M, May M, Chene G, et al. Prognosis of HIV-1-infected patients starting highly active antiretroviral therapy: a collaborative
analysis of prospective studies. Lancet 2002; 360: 119-29. [5] Furtado MR, Callaway DS, Phair JP, et al. Persistence of HIV-1
transcription in peripheral-blood mononuclear cells in patients re-ceiving potent antiretroviral therapy. N Engl J Med 1999; 340:
1614-22. [6] Chun TW, Stuyver L, Mizell SB, et al. Presence of an inducible
HIV-1 latent reservoir during highly active antiretroviral therapy. Proc Natl Acad Sci USA 1997; 94: 13193-7.
[7] Finzi D, Hermankova M, Pierson T, et al. Identification of a reser-voir for HIV-1 in patients on highly active antiretroviral therapy.
Science 1997; 278: 1295-300. [8] Zhang H, Dornadula G, Beumont M, et al. Human immunodefi-
ciency virus type 1 in the semen of men receiving highly active an-tiretroviral therapy. N Engl J Med 1998; 339: 1803-9.
[9] Chun TW, Justement JS, Moir S, et al. Decay of the HIV reservoir in patients receiving antiretroviral therapy for extended periods:
implications for eradication of virus. J Infect Dis 2007; 195: 1762-4.
[10] Chun TW, Nickle DC, Justement JS, et al. Persistence of HIV in Gut-Associated Lymphoid Tissue despite Long-Term Antiretrovi-
ral Therapy. J Infect Dis 2008; 197: 714-20. [11] Havlir DV, Strain MC, Clerici M, et al. Productive infection main-
tains a dynamic steady state of residual viremia in human immuno-deficiency virus type 1-infected persons treated with suppressive
antiretroviral therapy for five years. J Virol 2003; 77: 11212-9. [12] Havlir DV, Koelsch KK, Strain MC, et al. Predictors of residual
viremia in HIV-infected patients successfully treated with efavirenz and lamivudine plus either tenofovir or stavudine. J Infect Dis
2005; 191: 1164-8. [13] Chun TW, Fauci AS. Latent reservoirs of HIV: obstacles to the
eradication of virus. Proc Natl Acad Sci USA 1999; 96: 10958-61. [14] Chun TW, Engel D, Berrey MM, Shea T, Corey L, Fauci AS. Early
establishment of a pool of latently infected, resting CD4(+) T cells during primary HIV-1 infection. Proc Natl Acad Sci USA 1998;
95: 8869-73. [15] Chun TW, Carruth L, Finzi D, et al. Quantification of latent tissue
reservoirs and total body viral load in HIV-1 infection. Nature 1997; 387: 183-8.
[16] Chun TW, Nickle DC, Justement JS, et al. HIV-infected indivi-duals receiving effective antiviral therapy for extended periods of
time continually replenish their viral reservoir. J Clin Invest 2005; 115: 3250-5.
[17] Hatzakis AE, Touloumi G, Pantazis N, et al. Cellular HIV-1 DNA load predicts HIV-RNA rebound and the outcome of highly active
antiretroviral therapy. AIDS 2004; 18: 2261-7. [18] Hatzakis A, Touloumi G, Karanicolas R, et al. Effect of recent
thymic emigrants on progression of HIV-1 disease. Lancet 2000; 355: 599-604.
[19] Katzenstein TL, Oliveri RS, Benfield T, Eugen-Olsen J, Nielsen C, Gerstoft J. Cell-associated HIV DNA measured early during infec-
tion has prognostic value independent of serum HIV RNA measu-red concomitantly. Scand J Infect Dis 2002; 34: 529-33.
[20] Palmisano L, Giuliano M, Chiarotti F, et al. Modifications of HIV-1 DNA and Provirus-Infected Cells During 24 Months of Intermit-
tent Highly Active Antiretroviral Therapy. J Acquir Immune Defic Syndr 2008; 48: 68-71.
[21] Rouzioux C, Hubert JB, Burgard M, et al. Early levels of HIV-1 DNA in peripheral blood mononuclear cells are predictive of disea-

The Role of HIV-1 DNA Marker Current HIV Research, 2009, Vol. 7, No. 3 263
se progression independently of HIV-1 RNA levels and CD4+ T
cell counts. J Infect Dis 2005; 192: 46-55. [22] Re MC, Vitone F, Bon I, Schiavone P, Gibellini D. Meaning of
DNA detection during the follow-up of HIV-1 infected patients: a brief review. New Microbiol 2006; 29: 81-8.
[23] Re MC, Vitone F, Sighinolfi L, Schiavone P, Ghinelli F, Gibellini D. Different patterns of HIV-1 DNA after therapy discontinuation.
BMC Infect Dis 2005; 5: 69. [24] Sarmati L, Parisi SG, Nicastri E, et al. Association between cellular
human immunodeficiency virus DNA level and immunological pa-rameters in patients with undetectable plasma viremia level during
highly active antiretroviral therapy. J Clin Microbiol 2005; 43: 6183-5.
[25] Sarmati L, Parisi SG, Nicastri E, et al. Cellular HIV-1 DNA quanti-tation in patients during simplification therapy with protease inhibi-
tor-sparing regimens. J Med Virol 2007; 79: 880-6. [26] Palmisano L, Giuliano M, Nicastri E, et al. Residual viraemia in
subjects with chronic HIV infection and viral load < 50 copies/ml: the impact of highly active antiretroviral therapy. AIDS 2005; 19:
1843-7. [27] Belmonte L, Bare P, de Bracco MM, Ruibal-Ares BH. Reservoirs
of HIV replication after successful combined antiretroviral treatment. Curr Med Chem 2003; 10: 303-12.
[28] Avettand-Fenoel V, Mahlaoui N, Chaix ML, et al. Failure of bone marrow transplantation to eradicate HIV reservoir despite efficient
HAART. AIDS 2007; 21: 776-7. [29] Vitone F, Gibellini D, Schiavone P, Re MC. Quantitative DNA
proviral detection in HIV-1 patients treated with antiretroviral the-rapy. J Clin Virol 2005; 33: 194-200.
[30] Valentin A, Trivedi H, Lu W, Kostrikis LG, Pavlakis GN. CXCR4 mediates entry and productive infection of syncytia-inducing (X4)
HIV-1 strains in primary macrophages. Virology 2000; 269: 294-304.
[31] Valentin A, Rosati M, Patenaude DJ, et al. Persistent HIV-1 infec-tion of natural killer cells in patients receiving highly active antire-
troviral therapy. Proc Natl Acad Sci USA 2002; 99: 7015-20. [32] Hammer SM, Saag MS, Schechter M, et al. Treatment for adult
HIV infection: 2006 recommendations of the International AIDS Society--USA panel. Top HIV Med 2006; 14: 827-43.
[33] Bailey JR, Sedaghat AR, Kieffer T, et al. Residual human immu-nodeficiency virus type 1 viremia in some patients on antiretroviral
therapy is dominated by a small number of invariant clones rarely found in circulating CD4+ T cells. J Virol 2006; 80: 6441-57.
[34] Blankson JN, Persaud D, Siliciano RF. The challenge of viral re-servoirs in HIV-1 infection. Annu Rev Med 2002; 53: 557-93.
[35] Pierson T, McArthur J, Siliciano RF. Reservoirs for HIV-1: me-chanisms for viral persistence in the presence of antiviral immune
responses and antiretroviral therapy. Annu Rev Immunol 2000; 18: 665-708.
[36] Kelly J, Beddall MH, Yu D, Iyer SR, Marsh JW, Wu Y. Human macrophages support persistent transcription from unintegrated
HIV-1 DNA. Virology 2008; 372: 300-12. [37] Mansharamani M, Graham DR, Monie D, et al. Barrier-to-
autointegration factor BAF binds p55 Gag and matrix and is a host component of human immunodeficiency virus type 1 virions. J Vi-
rol 2003; 77: 13084-92. [38] Chen H, Engelman A. The barrier-to-autointegration protein is a
host factor for HIV type 1 integration. Proc Natl Acad Sci USA 1998; 95: 15270-4.
[39] Farnet CM, Bushman FD. HIV-1 cDNA integration: requirement of HMG I(Y) protein for function of preintegration complexes in
vitro. Cell 1997; 88: 483-92. [40] Lin CW, Engelman A. The barrier-to-autointegration factor is a
component of functional human immunodeficiency virus type 1 preintegration complexes. J Virol 2003; 77: 5030-6.
[41] Miller MD, Farnet CM, Bushman FD. Human immunodeficiency virus type 1 preintegration complexes: studies of organization and
composition. J Virol 1997; 71: 5382-90. [42] Fouchier RA, Malim MH. Nuclear import of human immunodefi-
ciency virus type-1 preintegration complexes. Adv Virus Res 1999; 52: 275-99.
[43] Bukrinsky MI, Sharova N, McDonald TL, Pushkarskaya T, Tarpley WG, Stevenson M. Association of integrase, matrix, and reverse
transcriptase antigens of human immunodeficiency virus type 1 with viral nucleic acids following acute infection. Proc Natl Acad
Sci USA 1993; 90: 6125-9.
[44] Karageorgos L, Li P, Burrell C. Characterization of HIV replication
complexes early after cell-to-cell infection. AIDS Res Hum Retro-viruses 1993; 9: 817-23.
[45] Robinson HL, Zinkus DM. Accumulation of human immunodefi-ciency virus type 1 DNA in T cells: results of multiple infection
events. J Virol 1990; 64: 4836-41. [46] Kim SY, Byrn R, Groopman J, Baltimore D. Temporal aspects of
DNA and RNA synthesis during human immunodeficiency virus infection: evidence for differential gene expression. J Virol 1989;
63: 3708-13. [47] Barbosa P, Charneau P, Dumey N, Clavel F. Kinetic analysis of
HIV-1 early replicative steps in a coculture system. AIDS Res Hum Retroviruses 1994; 10: 53-9.
[48] Butler SL, Hansen MS, Bushman FD. A quantitative assay for HIV DNA integration in vivo. Nat Med 2001; 7: 631-4.
[49] Vandegraaff N, Kumar R, Burrell CJ, Li P. Kinetics of human immunodeficiency virus type 1 (HIV) DNA integration in acutely
infected cells as determined using a novel assay for detection of in-tegrated HIV DNA. J Virol 2001; 75: 11253-60.
[50] Greene WC, Peterlin BM. Charting HIV's remarkable voyage through the cell: Basic science as a passport to future therapy. Nat
Med 2002; 8: 673-80. [51] Morlese J, Teo IA, Choi JW, Gazzard B, Shaunak S. Identification
of two mutually exclusive groups after long-term monitoring of HIV DNA 2-LTR circle copy number in patients on HAART.
AIDS 2003; 17: 679-83. [52] Sharkey ME, Teo I, Greenough T, et al. Persistence of episomal
HIV-1 infection intermediates in patients on highly active anti-retroviral therapy. Nat Med 2000; 6: 76-81.
[53] Sharkey M, Triques K, Kuritzkes DR, Stevenson M. In vivo evi-dence for instability of episomal human immunodeficiency virus
type 1 cDNA. J Virol 2005; 79: 5203-10. [54] Chavez HH, Tran TA, Dembele B, et al. Lack of evidence for
prolonged double-long terminal repeat episomal HIV DNA stabi-lity in vivo. J Acquir Immune Defic Syndr 2007; 45: 247-9.
[55] Pierson TC, Kieffer TL, Ruff CT, Buck C, Gange SJ, Siliciano RF. Intrinsic stability of episomal circles formed during human immu-
nodeficiency virus type 1 replication. J Virol 2002; 76: 4138-44. [56] Butler SL, Johnson EP, Bushman FD. Human immunodeficiency
virus cDNA metabolism: notable stability of two-long terminal re-peat circles. J Virol 2002; 76: 3739-47.
[57] Wu Y, Marsh JW. Selective transcription and modulation of resting T cell activity by preintegrated HIV DNA. Science 2001; 293:
1503-6. [58] Stevenson M, Stanwick TL, Dempsey MP, Lamonica CA. HIV-1
replication is controlled at the level of T cell activation and proviral integration. Embo J 1990; 9: 1551-60.
[59] Wu Y, Marsh JW. Early transcription from nonintegrated DNA in human immunodeficiency virus infection. J Virol 2003; 77: 10376-
82. [60] Wu Y. HIV-1 gene expression: lessons from provirus and non-
integrated DNA. Retrovirology 2004; 1: 13. [61] Montoya JG, Wood R, Katzenstein D, Holodny M, Merigan TC.
Peripheral blood mononuclear cell human immunodeficiency virus type 1 proviral DNA quantification by polymerase chain reaction:
relationship to immunodeficiency and drug effect. J Clin Microbiol 1993; 31: 2692-6.
[62] Aoki S, Yarchoan R, Thomas RV, et al. Quantitative analysis of HIV-1 proviral DNA in peripheral blood mononuclear cells from
patients with AIDS or ARC: decrease of proviral DNA content fol-lowing treatment with 2',3'-dideoxyinosine (ddI). AIDS Res Hum
Retroviruses 1990; 6: 1331-9. [63] Bieniasz PD, Ariyoshi K, Bourelly MA, et al. Variable relationship
between proviral DNA load and infectious virus titre in the peri-pheral blood mononuclear cells of HIV-1-infected individuals.
AIDS 1993; 7: 803-6. [64] Gupta P, Ding M, Cottrill M, et al. Quantitation of human immu-
nodeficiency virus type 1 DNA and RNA by a novel internally con-trolled PCR assay. J Clin Microbiol 1995; 33: 1670-3.
[65] Izopet J, Tamalet C, Pasquier C, et al. Quantification of HIV-1 proviral DNA by a standardized colorimetric PCR-based assay. J
Med Virol 1998; 54: 54-9. [66] Katzenstein DA, Hammer SM, Hughes MD, et al. The relation of
virologic and immunologic markers to clinical outcomes after nu-cleoside therapy in HIV-infected adults with 200 to 500 CD4 cells

264 Current HIV Research, 2009, Vol. 7, No. 3 Beloukas et al.
per cubic millimeter. AIDS Clinical Trials Group Study 175 Viro-
logy Study Team. N Engl J Med 1996; 335: 1091-8. [67] Lin HJ, Haywood M, Hollinger FB. Application of a commercial
kit for detection of PCR products to quantification of human im-munodeficiency virus type 1 RNA and proviral DNA. J Clin Mi-
crobiol 1996; 34: 329-33. [68] Mallet F, Hebrard C, Livrozet JM, et al. Quantitation of human
immunodeficiency virus type 1 DNA by two PCR procedures cou-pled with enzyme-linked oligosorbent assay. J Clin Microbiol
1995; 33: 3201-8. [69] Sei S, Kleiner DE, Kopp JB, et al. Quantitative analysis of viral
burden in tissues from adults and children with symptomatic hu-man immunodeficiency virus type 1 infection assessed by polyme-
rase chain reaction. J Infect Dis 1994; 170: 325-33. [70] Vesanen M, Stevens CE, Taylor PE, Rubinstein P, Saksela K.
Stability in controlling viral replication identifies long-term non-progressors as a distinct subgroup among human immunodefi-
ciency virus type 1-infected persons. J Virol 1996; 70: 9035-40. [71] Yerly S, Kaiser L, Baumberger C, Hirschel B, Perrin LH. Early and
prolonged decrease of viremia in HIV-1-infected patients treated with didanosine. J Acquir Immune Defic Syndr Hum Retrovirol
1995; 8: 358-64. [72] Furtado MR, Kingsley LA, Wolinsky SM. Changes in the viral
mRNA expression pattern correlate with a rapid rate of CD4+ T-cell number decline in human immunodeficiency virus type 1-
infected individuals. J Virol 1995; 69: 2092-2100. [73] Menzo S, Bagnarelli P, Giacca M, Manzin A, Varaldo PE, Clemen-
ti M. Absolute quantitation of viremia in human immunodeficiency virus infection by competitive reverse transcription and polymerase
chain reaction. J Clin Microbiol 1992; 30: 1752-7. [74] Frenkel LM, Wang Y, Learn GH, et al. Multiple viral genetic anal-
yses detect low-level human immunodeficiency virus type 1 repli-cation during effective highly active antiretroviral therapy. J Virol
2003; 77: 5721-30. [75] Nygren M, Ronaghi M, Nyren P, Albert J, Lundeberg J. Quantifi-
cation of HIV-1 using multiple quantitative polymerase chain reac-tion standards and bioluminometric detection. Anal Biochem 2001;
288: 28-38. [76] Teo I, Veryard C, Barnes H, et al. Circular forms of unintegrated
human immunodeficiency virus type 1 DNA and high levels of vi-ral protein expression: association with dementia and multinuclea-
ted giant cells in the brains of patients with AIDS. J Virol 1997; 71: 2928-33.
[77] Vandegraaff N, Kumar R, Hocking H, et al. Specific inhibition of human immunodeficiency virus type 1 (HIV-1) integration in cell
culture: putative inhibitors of HIV-1 integrase. Antimicrob Agents Chemother 2001; 45: 2510-6.
[78] Debiaggi M, Zara F, Pistorio A, et al. Quantification of HIV-1 proviral DNA in patients with undetectable plasma viremia over
long-term highly active antiretroviral therapy. Int J Infect Dis 2000; 4: 187-93.
[79] McDermott JL, Giri AA, Martini I, et al. Level of human immuno-deficiency virus DNA in peripheral blood mononuclear cells corre-
lates with efficacy of antiretroviral therapy. J Clin Microbiol 1999; 37: 2361-5.
[80] Comandini UV, Sonnerborg A, Vahlne A, Yun Z. Quantification of HIV-1 proviral DNA from peripheral blood mononuclear cells u-
sing a high throughput four-competitor competitive PCR. J Virol Methods 1997; 69: 171-80.
[81] Comar M, Simonelli C, Zanussi S, et al. Dynamics of HIV-1 mRNA expression in patients with long-term nonprogressive HIV-
1 infection. J Clin Invest 1997; 100: 893-903. [82] Holodniy M, Katzenstein DA, Sengupta S, et al. Detection and
quantification of human immunodeficiency virus RNA in patient serum by use of the polymerase chain reaction. J Infect Dis 1991;
163: 862-6. [83] Gupta P, Kingsley L, Armstrong J, Ding M, Cottrill M, Rinaldo C.
Enhanced expression of human immunodeficiency virus type 1 cor-relates with development of AIDS. Virology 1993; 196: 586-95.
[84] Garbuglia AR, Salvi R, Di Caro A, et al. Peripheral lymphocytes of clinically non-progressor patients harbor inactive and uninducible
HIV proviruses. J Med Virol 1995; 46: 116-21. [85] Pauza CD, Trivedi P, McKechnie TS, Richman DD, Graziano FM.
2-LTR circular viral DNA as a marker for human immunodefi-ciency virus type 1 infection in vivo. Virology 1994; 205: 470-8.
[86] Christopherson C, Kidane Y, Conway B, Krowka J, Sheppard H,
Kwok S. PCR-Based assay to quantify human immunodeficiency virus type 1 DNA in peripheral blood mononuclear cells. J Clin
Microbiol 2000; 38: 630-4. [87] Lillo FB, Grasso MA, Lodini S, Bellotti MG, Colucci G. Few
modifications of the Cobas Amplicor HIV Monitor 1.5 test allow reliable quantitation of HIV-1 proviral load in peripheral blood
mononuclear cells. J Virol Methods 2004; 120: 201-5. [88] Jennings C, Danilovic A, Scianna S, Brambilla DJ, Bremer JW.
Stability of human immunodeficiency virus type 1 proviral DNA in whole-blood samples. J Clin Microbiol 2005; 43: 4249-50.
[89] Beloukas A, Paraskevis D, Haida C, Sypsa V, Hatzakis A. Develo-pment and Assessment of a multiplex Real-Time PCR Assay for
the Quantification of HIV-1 DNA (RTMP-HIV). J Clin Microbiol 2009; (submitted).
[90] Eriksson LE, Leitner T, Wahren B, Bostrom AC, Falk KI. A multi-plex real-time PCR for quantification of HIV-1 DNA and the hu-
man albumin gene in CD4+ cells. Apmis 2003; 111: 625-33. [91] Gibellini D, Monari P, Sassi M, Re MC. A rapid hemi-nested PCR
for HTLV-I detection. J Clin Virol 2003; 28: 341-3. [92] Kostrikis LG, Touloumi G, Karanicolas R, et al. Quantitation of
human immunodeficiency virus type 1 DNA forms with the second template switch in peripheral blood cells predicts disease progres-
sion independently of plasma RNA load. J Virol 2002; 76: 10099-108.
[93] Sharova N, Swingler C, Sharkey M, Stevenson M. Macrophages archive HIV-1 virions for dissemination in trans. EMBO J 2005;
24: 2481-9. [94] Yun Z, Fredriksson E, Sonnerborg A. Quantification of human
immunodeficiency virus type 1 proviral DNA by the TaqMan real-time PCR assay. J Clin Microbiol 2002; 40: 3883-4.
[95] Zhao Y, Yu M, Miller JW, et al. Quantification of human immuno-deficiency virus type 1 proviral DNA by using TaqMan techno-
logy. J Clin Microbiol 2002; 40: 675-8. [96] Zaunders JJ, Cunningham PH, Kelleher AD, et al. Potent antiretro-
viral therapy of primary human immunodeficiency virus type 1 (HIV-1) infection: partial normalization of T lymphocyte subsets
and limited reduction of HIV-1 DNA despite clearance of plasma viremia. J Infect Dis 1999; 180: 320-9.
[97] Kabamba-Mukadi B, Henrivaux P, Ruelle J, Delferriere N, Bodeus M, Goubau P. Human immunodeficiency virus type 1 (HIV-1) pro-
viral DNA load in purified CD4+ cells by LightCycler real-time PCR. BMC Infect Dis 2005; 5: 15.
[98] Casabianca A, Gori C, Orlandi C, Forbici F, Federico Perno C, Magnani M. Fast and sensitive quantitative detection of HIV DNA
in whole blood leucocytes by SYBR green I real-time PCR assay. Mol Cell Probes 2007; 21: 368-78.
[99] Desire N, Dehee A, Schneider V, et al. Quantification of human immunodeficiency virus type 1 proviral load by a TaqMan real-
time PCR assay. J Clin Microbiol 2001; 39: 1303-10. [100] Goedert JJ, O'Brien TR, Hatzakis A, Kostrikis LG. T cell receptor
excision circles and HIV-1 2-LTR episomal DNA to predict AIDS in patients not receiving effective therapy. AIDS 2001; 15: 2245-
50. [101] Garbuglia M, Bianchi R, Verzini M, Giambanco I, Donato R. An-
nexin II2-p11(2) (calpactin I) stimulates the assembly of GFAP in a calcium- and pH-dependent manner. Biochem Biophys Res Com-
mun 1995; 208: 901-9. [102] Zazzi M, Romano L, Catucci M, et al. Evaluation of the presence
of 2-LTR HIV-1 unintegrated DNA as a simple molecular predictor of disease progression. J Med Virol 1997; 52: 20-5.
[103] Teo IA, Morlese J, Choi JW, Shaunak S. Reliable and reproducible LightCycler qPCR for HIV-1 DNA 2-LTR circles. J Immunol Me-
thods 2002; 270: 109-18. [104] Moroney SM, Heller LC, Widen RH. Evaluation of two TaqMan
PCR assays for the detection of HIV-1 proviral DNA in blood samples. J Microbiol Methods 2006; 65: 350-3.
[105] Viard JP, Burgard M, Hubert JB, et al. Impact of 5 years of maxi-mally successful highly active antiretroviral therapy on CD4 cell
count and HIV-1 DNA level. AIDS 2004; 18: 45-9. [106] Gibellini D, Vitone F, Schiavone P, Ponti C, La Placa M, Re MC.
Quantitative detection of human immunodeficiency virus type 1 (HIV-1) proviral DNA in peripheral blood mononuclear cells by
SYBR green real-time PCR technique. J Clin Virol 2004; 29: 282-9.

The Role of HIV-1 DNA Marker Current HIV Research, 2009, Vol. 7, No. 3 265
[107] Zanchetta M, Walker S, Burighel N, et al. Long-term decay of the
HIV-1 reservoir in HIV-1-infected children treated with highly ac-tive antiretroviral therapy. J Infect Dis 2006; 193: 1718-27.
[108] Carr JM, Cheney KM, Coolen C, et al. Development of methods for coordinate measurement of total cell-associated and integrated
human immunodeficiency virus type 1 (HIV-1) DNA forms in rou-tine clinical samples: levels are not associated with clinical parame-
ters, but low levels of integrated HIV-1 DNA may be prognostic for continued successful therapy. J Clin Microbiol 2007; 45: 1288-
97. [109] Gibellini D, Vitone F, Gori E, La Placa M, Re MC. Quantitative
detection of human immunodeficiency virus type 1 (HIV-1) viral load by SYBR green real-time RT-PCR technique in HIV-1 sero-
positive patients. J Virol Methods 2004; 115: 183-9. [110] Katzenstein TL, Ullum H, Roge BT, et al. Virological and immu-
nological profiles among patients with undetectable viral load fol-lowed prospectively for 24 months. HIV Med 2003; 4: 53-61.
[111] Ngo-Giang-Huong N, Deveau C, Da Silva I, et al. Proviral HIV-1 DNA in subjects followed since primary HIV-1 infection who sup-
press plasma viral load after one year of highly active antiretroviral therapy. AIDS 2001; 15: 665-73.
[112] McDermott JL, Martini I, Ferrari D, et al. Decay of human immu-nodeficiency virus type 1 unintegrated DNA containing two long
terminal repeats in infected individuals after 3 to 8 years of sustai-ned control of viremia. J Clin Microbiol 2005; 43: 5272-4.
[113] Brussel A, Delelis O, Sonigo P. Alu-LTR real-time nested PCR assay for quantifying integrated HIV-1 DNA. Methods Mol Biol
2005; 304: 139-54. [114] Kumar R, Vandegraaff N, Mundy L, Burrell CJ, Li P. Evaluation of
PCR-based methods for the quantitation of integrated HIV-1 DNA. J Virol Methods 2002; 105: 233-46.
[115] Yamamoto N, Tanaka C, Wu Y, et al. Analysis of human immuno-deficiency virus type 1 integration by using a specific, sensitive and
quantitative assay based on real-time polymerase chain reaction. Virus Genes 2006; 32: 105-13.
[116] Fischer M, Trkola A, Joos B, et al. Shifts in cell-associated HIV-1 RNA but not in episomal HIV-1 DNA correlate with new cycles of
HIV-1 infection in vivo. Antivir Ther 2003; 8: 97-104. [117] Clark AG, Holodniy M, Schwartz DH, Katzenstein DA, Merigan
TC. Decrease in HIV provirus in peripheral blood mononuclear cells during zidovudine and human rIL-2 administration. J Acquir
Immune Defic Syndr 1992; 5: 52-9. [118] Hoen B, Cooper DA, Lampe FC, et al. Predictors of virological
outcome and safety in primary HIV type 1-infected patients initia-ting quadruple antiretroviral therapy: QUEST GW PROB3005.
Clin Infect Dis 2007; 45: 381-90. [119] Lafeuillade A, Poggi C, Chadapaud S, Hittinger G, Khiri H, Halfon
P. Impact of immune interventions on proviral HIV-1 DNA decay in patients receiving highly active antiretroviral therapy. HIV Med
2001; 2: 189-94. [120] Lafeuillade A, Poggi C, Hittinger G, Counillon E, Emilie D. Pre-
dictors of plasma human immunodeficiency virus type 1 RNA control after discontinuation of highly active antiretroviral therapy
initiated at acute infection combined with structured treatment in-terruptions and immune-based therapies. J Infect Dis 2003; 188:
1426-32. [121] Torti C, Quiros-Roldan ME, Cologni G, et al. Plasma HIV load and
proviral DNA decreases after two standard antiretroviral regimens in HIV-positive patients naive to antiretrovirals. Curr HIV Res
2008; 6: 43-8. [122] Andreoni M, Parisi SG, Sarmati L, et al. Cellular proviral HIV-
DNA decline and viral isolation in naive subjects with <5000 co-pies/ml of HIV-RNA and >500 x 10(6)/l CD4 cells treated with
highly active antiretroviral therapy. AIDS 2000; 14: 23-9. [123] Burgard M, Izopet J, Dumon B, et al. HIV RNA and HIV DNA in
peripheral blood mononuclear cells are consistent markers for esti-
mating viral load in patients undergoing long-term potent
treatment. AIDS Res Hum Retroviruses 2000; 16: 1939-47. [124] Gibellini D, Borderi M, De Crignis E, et al. HIV-1 DNA load anal-
ysis in peripheral blood lymphocytes and monocytes from naive and HAART-treated individuals. J Infect 2008; 56: 219-25.
[125] Morand-Joubert L, Marcellin F, Launay O, et al. Contribution of cellular HIV-1 DNA quantification to the efficacy analysis of anti-
retroviral therapy: a randomized comparison of 2 regimens, inclu-ding 3 drugs from 2 or 3 classes (TRIANON, ANRS 081). J Acquir
Immune Defic Syndr 2005; 38: 268-76. [126] Tierney C, Lathey JL, Christopherson C, et al. Prognostic value of
baseline human immunodeficiency virus type 1 DNA measurement for disease progression in patients receiving nucleoside therapy. J
Infect Dis 2003; 187: 144-8. [127] Goujard C, Bonarek M, Meyer L, et al. CD4 cell count and HIV
DNA level are independent predictors of disease progression after primary HIV type 1 infection in untreated patients. Clin Infect Dis
2006; 42: 709-15. [128] Opravil M, Ledergerber B, Furrer H, et al. Clinical efficacy of
early initiation of HAART in patients with asymptomatic HIV in-fection and CD4 cell count > 350 x 10(6) /l. AIDS 2002; 16: 1371-
81. [129] Hogg RS, Yip B, Chan KJ, et al. Rates of disease progression by
baseline CD4 cell count and viral load after initiating triple-drug therapy. JAMA 2001; 286: 2568-77.
[130] Hammer SM, Squires KE, Hughes MD, et al. A controlled trial of two nucleoside analogues plus indinavir in persons with human
immunodeficiency virus infection and CD4 cell counts of 200 per cubic millimeter or less. AIDS Clinical Trials Group 320 Study
Team. N Engl J Med 1997; 337: 725-33. [131] Gulick RM. Current antiretroviral therapy: an overview. Qual Life
Res 1997; 6: 471-4. [132] Gulick RM, Mellors JW, Havlir D, et al. Treatment with indinavir,
zidovudine, and lamivudine in adults with human immunodefi-ciency virus infection and prior antiretroviral therapy. N Engl J
Med 1997; 337: 734-9. [133] Yerly S, Kaiser L, Perneger TV, et al. Time of initiation of antire-
troviral therapy: impact on HIV-1 viraemia. The Swiss HIV Cohort Study. AIDS 2000; 14: 243-9.
[134] Pomerantz RJ. Residual HIV-1 RNA in blood plasma of patients taking suppressive highly active antiretroviral therapy. Biomed
Pharmacother 2001; 55: 7-15. [135] Dornadula G, Zhang H, VanUitert B, et al. Residual HIV-1 RNA in
blood plasma of patients taking suppressive highly active antiretro-viral therapy. JAMA 1999; 282: 1627-32.
[136] Havlir DV, Bassett R, Levitan D, et al. Prevalence and predictive value of intermittent viremia with combination hiv therapy. JAMA
2001; 286: 171-9. [137] Palmer S, Wiegand AP, Maldarelli F, et al. New real-time reverse
transcriptase-initiated PCR assay with single-copy sensitivity for human immunodeficiency virus type 1 RNA in plasma. J Clin Mi-
crobiol 2003; 41: 4531-6. [138] Marchetti G, Gori A, Casabianca A, et al. Comparative analysis of
T-cell turnover and homeostatic parameters in HIV-infected pa-tients with discordant immune-virological responses to HAART.
AIDS 2006; 20: 1727-36. [139] Chun TW, Justement JS, Pandya P, et al. Relationship between the
size of the human immunodeficiency virus type 1 (HIV-1) reservoir in peripheral blood CD4+ T cells and CD4+:CD8+ T cell ratios in
aviremic HIV-1-infected individuals receiving long-term highly ac-tive antiretroviral therapy. J Infect Dis 2002; 185: 1672-6.
[140] Yerly S, Gunthard HF, Fagard C, et al. Proviral HIV-DNA predicts viral rebound and viral setpoint after structured treatment interrup-
tions. Aids 2004; 18: 1951-3.
Received: August 20, 2008 Accepted: March 17, 2009




















