
mRNA-encoded HIV-1 Env trimer ferritin nanoparticles induce ...
42
Article mRNA-encoded HIV-1 Env trimer ferritin nanoparticles induce monoclonal antibodies that neutralize heterologous HIV-1 isolates in mice Graphical abstract Highlights d mRNA-expressed HIV-1 Envs are well folded with optimal stabilizing mutations d mRNA-expressed stabilized Envs show preferential bnAb binding d mRNA-LNP elicit autologous tier 2 neutralizing antibodies with key bnAb mutations d Induced monoclonal antibodies with key mutations neutralize heterologous viruses Authors Zekun Mu, Kevin Wiehe, Kevin O. Saunders, ..., Norbert Pardi, Drew Weissman, Barton F. Haynes Correspondence [email protected] (B.F.H.), [email protected] (Z.M.), [email protected] (D.W.) In brief mRNA vaccines are highly effective against COVID-19. Mu et al. demonstrate the use of mRNA to express HIV-1 Env trimers scaffolded on ferritin nanoparticles. mRNA vaccination in mice induced autologous tier 2 neutralizing antibodies and key functional mutations. Isolated monoclonal antibodies neutralized heterologous HIV-1 isolates. Mu et al., 2022, Cell Reports 38, 110514 March 15, 2022 ª 2022 The Authors. https://doi.org/10.1016/j.celrep.2022.110514 ll
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of mRNA-encoded HIV-1 Env trimer ferritin nanoparticles induce ...
Article
mRNA-encoded HIV-1 Env
trimer ferritinnanoparticles induce monoclonal antibodies thatneutralize heterologous HIV-1 isolates in miceGraphical abstract
Highlights
d mRNA-expressed HIV-1 Envs are well folded with optimal
stabilizing mutations
d mRNA-expressed stabilized Envs show preferential bnAb
binding
d mRNA-LNP elicit autologous tier 2 neutralizing antibodies
with key bnAb mutations
d Inducedmonoclonal antibodies with keymutations neutralize
heterologous viruses
Mu et al., 2022, Cell Reports 38, 110514March 15, 2022 ª 2022 The Authors.https://doi.org/10.1016/j.celrep.2022.110514
Authors
Zekun Mu, Kevin Wiehe,
Kevin O. Saunders, ..., Norbert Pardi,
Drew Weissman, Barton F. Haynes
[email protected] (B.F.H.),[email protected] (Z.M.),[email protected] (D.W.)
In brief
mRNA vaccines are highly effective
against COVID-19. Mu et al. demonstrate
the use of mRNA to express HIV-1 Env
trimers scaffolded on ferritin
nanoparticles. mRNA vaccination in mice
induced autologous tier 2 neutralizing
antibodies and key functional mutations.
Isolated monoclonal antibodies
neutralized heterologous HIV-1 isolates.
ll

OPEN ACCESS
llArticle
mRNA-encoded HIV-1 Env trimer ferritinnanoparticles induce monoclonal antibodiesthat neutralize heterologous HIV-1 isolates in miceZekunMu,1,2,* KevinWiehe,2,3 Kevin O. Saunders,1,2,4,5 Rory Henderson,2,3 DerekW. Cain,2 Robert Parks,2 DianaMartik,2
Katayoun Mansouri,2 Robert J. Edwards,2,3 Amanda Newman,2 Xiaozhi Lu,2 Shi-Mao Xia,2 Amanda Eaton,2
Mattia Bonsignori,2,3,10 David Montefiori,2,4 Qifeng Han,2,11 Sravani Venkatayogi,2 Tyler Evangelous,2 Yunfei Wang,2
Wes Rountree,2 Bette Korber,6 Kshitij Wagh,6 Ying Tam,7 Christopher Barbosa,7 S. Munir Alam,2 Wilton B. Williams,1,2,4
Ming Tian,8 Frederick W. Alt,9 Norbert Pardi,9 Drew Weissman,9,* and Barton F. Haynes1,2,3,12,*1Department of Immunology, Duke University School of Medicine, Durham, NC 27710, USA2Duke Human Vaccine Institute, Duke University School of Medicine, Durham, NC 27710, USA3Department of Medicine, Duke University School of Medicine, Durham, NC 27710, USA4Department of Surgery, Duke University School of Medicine, Durham, NC 27710, USA5Department of Molecular Genetics and Microbiology, Duke University School of Medicine, Durham, NC 27710, USA6Theoretical Biology and Biophysics, Los Alamos National Laboratory, Los Alamos, NM 87545, USA7Acuitas, Inc, Vancouver, Canada8Howard Hughes Medical Institute, Program in Cellular and Molecular Medicine, Boston Children’s Hospital, Department of Genetics,Harvard Medical School, Boston, MA 02115, USA9Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA10Present address: Translational Immunobiology Unit, Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases,National Institutes of Health, Bethesda, MD 20892, USA11Present address: Novo Nordisk Research Center China, Novo Nordisk A/S, Beijing 102206, China12Lead contact
*Correspondence: [email protected] (B.F.H.), [email protected] (Z.M.), [email protected] (D.W.)https://doi.org/10.1016/j.celrep.2022.110514
SUMMARY
The success of nucleoside-modified mRNAs in lipid nanoparticles (mRNA-LNP) as COVID-19 vaccines her-alded a new era of vaccine development. For HIV-1, multivalent envelope (Env) trimer protein nanoparticlesare superior immunogens compared with trimers alone for priming of broadly neutralizing antibody (bnAb) Bcell lineages. The successful expression of complex multivalent nanoparticle immunogens with mRNAs hasnot been demonstrated. Here, we show that mRNAs can encode antigenic Env trimers on ferritin nanopar-ticles that initiate bnAb precursor B cell expansion and induce serum autologous tier 2 neutralizing activityin bnAb precursor VH + VL knock-in mice. Next-generation sequencing demonstrates acquisition of criticalmutations, and monoclonal antibodies that neutralize heterologous HIV-1 isolates are isolated. Thus,mRNA-LNP can encode complex immunogens and may be of use in design of germline-targeting andsequential boosting immunogens for HIV-1 vaccine development.
INTRODUCTION
Nucleoside-modified mRNA COVID-19 vaccines encoding
SARS-CoV-2 trimeric spike protein have demonstrated the
robust nature of mRNA vaccines (Baden et al., 2020; Polack et
al., 2020; Sahin et al., 2020). In addition to success with clini-
cally-approved COVID-19 spike trimer vaccines, pre-clinical
success has been demonstrated with nucleoside-modified
mRNA encapsulated in lipid nanoparticle (mRNA-LNP) expres-
sion of Zika prM-E (Pardi et al., 2017), influenza hemagglutinin
(Pardi et al., 2018a, 2018b), and HIV-1 envelope (Env) in gp120
monomers or gp140 trimers (; Pardi et al., 2018a; Saunders
et al., 2021). However, recent studies have shown that protein
trimer nanoparticle (NP) multimers may be advantageous as im-
This is an open access article under the CC BY-N
munogens (Abbott et al., 2018; Havenar-Daughton et al., 2018;
Kato et al., 2020; Saunders et al., 2019; Tokatlian et al., 2019).
HIV-1 broadly neutralizing antibodies (bnAbs) may be disfa-
vored by the immune system due to their characteristics of
long heavy-chain complementarity-determining region 3
(HCDR3) loops and polyreactivity or autoreactivity that predis-
pose bnAbs to immune tolerance control (Havenar-Daughton
et al., 2018; Haynes et al., 2005, 2012, 2016, 2019; Huang
et al., 2020; Moody et al., 2016; Saunders et al., 2019;
Steichen et al., 2019; Zhang et al., 2016). Thus, the biology of
HIV-1 bnAbs has necessitated a strategy whereby the unmu-
tated common ancestor (UCA) or germline (GL) precursor of
bnAb B cell lineage is targeted with priming immunogens to
expand the bnAb precursor pool (Haynes et al., 2012, 2019;
Cell Reports 38, 110514, March 15, 2022 ª 2022 The Authors. 1C-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).

A
B
E
C D
Figure 1. Antigenicity of modified mRNA-encoded CH848 10.17DT gp160 with stabilizing mutations
(A) Stabilizing mutations tested in this study mapped onto structure of CH848 10.17DT SOSIP trimer (PDB ID: 6UM5). One protomer is shown in rainbow color,
and the other two protomers are shown in gray. Amino acid mutations are listed in boxes. Black fonts outside of boxes are mutations in v4.1, and blue fonts are
mutations in v5.2.8, in addition to v4.1 mutations. Residue 561C in v5.2.8 or redesigned HR1 region in UFOmutation is not shown due to lack of HR1 region in this
structure.
(B) Antigenicity of mRNA-expressed CH848 10.17DT, F14, and 113C-429GCG transmembrane gp160s measured by binding of bnAb PGT125 and DH270 UCA.
Data are shown as means ± standard error of the mean (SEM) of Phycoerythrin + (PE +) cell percentage among live cells from three independent experiments.
(C) eCD4-Ig binding reactivity to mRNA-expressed CH848 10.17DT, F14, and 113C-429GCG gp160s. Data shown are averages of mean fluorescent intensity
(MFI) from three independent experiments. Error bars, means ± SEM.
(legend continued on next page)
2 Cell Reports 38, 110514, March 15, 2022
Articlell
OPEN ACCESS

Articlell
OPEN ACCESS
Jardine et al., 2013; McGuire et al., 2013). Env immunogens de-
signed to select for key antibody mutations, administered in a
specific order, are postulated to be able to guide antibody affinity
maturation toward bnAb breadth and potency (Bonsignori et al.,
2016, 2017; Havenar-Daughton et al., 2018; Haynes et al., 2012,
2016, 2019; Huang et al., 2020; Saunders et al., 2019; Steichen
et al., 2019; Zhang et al., 2016). However, guiding bnAb develop-
ment is difficult, because HIV-1 bnAbs are enriched in improb-
able functional somatic mutations that are required for neutrali-
zation potency and breadth (Bonsignori et al., 2017; Wiehe
et al., 2018). Rare somatic mutations are due to lack of targeting
by the somatic mutation enzyme activation-induced cytidine
deaminase (AID). To promote bnAb development, Envs will
need to engage B cell receptors that have accumulated func-
tional improbable mutations, thereby selecting intermediate
bnAb B cell lineage members to proliferate and evolve further
(Bonsignori et al., 2017; Haynes et al., 2012; Wiehe et al.,
2018). Whereas bnAbs arise in �50% of HIV-1-infected individ-
uals (Hraber et al., 2014), to date, potent and durable bnAbs have
not been induced in humans by vaccination.
HIV-1 Env is metastable and can adopt open and closed con-
formations (Tran et al., 2012; Ward and Wilson, 2017). The Env
open conformation exposes non-neutralizing antibody (nnAb)
epitopes that can create competition for Env antigen between
nnAb and bnAb precursors (Havenar-Daughton et al., 2017;
Lee et al., 2021; McGuire et al., 2014). To address the problem
of Env trimer opening and the exposure of non-neutralizing epi-
topes,multiple strategies have been designed to stabilize Env tri-
mers in native-like conformations (de Taeye et al., 2015; Gue-
naga et al., 2015; Henderson et al., 2020; Kong et al., 2016).
However, whether stabilizing mutations for mRNA expression
of complex multimers will result in the desired antigenicity and
immunogenicity of trimer multimer NPs is not known.
We have previously demonstrated that a HIV-1 Env trimer,
CH848, with two glycosylation sites in the first variable region
(V1), eliminated (CH848 N133D N138T, CH848 10.17DT), conju-
gated to a ferritin nanoparticle that was superior to the Env trimer
alone as an immunogen, and was capable of initiating a V3-
glycan bnAb lineage with key improbable mutations in bnAb
UCA heavy- and light-chain variable regions (VH + VL) knock-in
(KI) mice (Saunders et al., 2019).
Here, we determined stabilizing mutations in the CH848
10.17DT immunogen for formulation as mRNA-LNP. We demon-
strated thatmodifiedmRNAEnv expression resulted in Env bind-
ing to bnAbs in the forms of transmembrane gp160s, soluble
gp140 SOSIP trimers, or SOSIP trimer-ferritin NPs encoded by
mRNA (trimer-ferritin NPs, hereafter ‘‘NPs’’). Moreover, we
demonstrated that immunization of bnAb UCA VH + VL KI mice
with mRNA-LNP encoding CH848 10.17DT gp160s or NPs initi-
ated a V3-glycan bnAb B cell lineage, selected for bnAb lineage
B cells with B cell receptors (BCRs) bearing functional improb-
(D) Binding reactivity of nnAbs 17b and 19b to mRNA-expressed CH848 10.17D
eCD4-Ig. Data shown are means ± SEM of PE + cell percentage among live cell
(E) Binding of Env base antibodies to mRNA-encoded CH848 10.17DT, F14, and
negative control, and 2G12 was used as positive control to show gp160 expressio
See also Figures S1 and S2 and Table S1.
able mutations and induced high serum titers of tier 2 V3-glycan
bnAb N332-dependent autologous neutralizing antibodies.
RESULTS
Antigenicity of modified mRNA-encoded CH84810.17DT gp160s with stabilizing mutationsWe studied ten stabilization designs in CH848 10.17DT Env for
expression as mRNAs (de Taeye et al., 2015; Guenaga et al.,
2015; Henderson et al., 2020; Kong et al., 2016; Tran et al.,
2012; Ward and Wilson, 2017; Zhang et al., 2018) (Figure 1A
and Table S1; see STAR methods).We first designed mRNAs
with stabilizing mutations encoding CH848 10.17DT Envs as
transmembrane gp160s (Table S1). All mRNA constructs ex-
pressed and showed robust V3-glycan bnAb binding (Figure S1).
In particular, CH848 10.17DT F14, 113C-429GCG, and 113C-
431GCG gp160s exhibited binding reactivity to mature V3-
glycan bnAb PGT125 and the DH270 UCA equal to that of
10.17DT gp160 without stabilizing mutations (Figure 1B).
In contrast, DS, Vt8, and F14/Vt8 mutations decreased
DH270 UCA binding to 10.17DT gp160s (Figure S1B).
To evaluate expression of CD4-induced (CD4i) Env epitopes,
we examined the susceptibility of each Env gp160 to CD4 trig-
gering in transfected 293-F cells. Engineered CD4-Ig (eCD4-Ig)
(Fellinger et al., 2019) bound to CH848 10.17DT gp160 lacking
stabilizing mutations in a dose-dependent manner, but binding
to 10.17DT F14 and 113C-429GCG gp160s was minimal (Fig-
ure 1C). Next, we assessed whether mutations could stabilize
10.17DT gp160s in prefusion conformations and prevent V3
loop or C-C chemokine receptor type 5 (CCR5) co-receptor
binding site exposure. mRNA-transfected 293-F cells were
either untreated or treated with 20 mg/mL of sCD4, eCD4-Ig, or
CD4-IgG2 (Allaway et al., 1995), and Env conformation was
determined by binding of CCR5 co-receptor binding site nnAb,
17b or distal V3 loop nnAb, 19b. In the absence of CD4 treat-
ment, Env gp160s lacked binding to 17b or 19b. After treatment
with sCD4, eCD4-Ig, or CD4-IgG2, 10.17DT gp160 without sta-
bilizing mutations exhibited increased binding to both nnAbs
17b and 19b. In contrast, stabilizing the Env gp160s with F14
or 113C-429GCG mutations completely prevented CD4-
induced exposure of 17b and 19b epitopes (Figures 1D and
S1C). F14 and 113C-429GCG mutations are outside of the 17b
and 19b binding site; thus, these mutations themselves do not
interfere with 17b or 19b binding (Henderson et al., 2020; Zhang
et al., 2018). In addition, if F14 mutations themselves impair 17b
or 19b binding, then similar effects would be observed for any
Env strains. Thus, we performed surface plasmon resonance
(SPR) analysis of 17b and 19b binding to 10.17DT F14 and
CH505 M5 G458Y F14 SOSIP proteins. Some increase to 19b
binding was observed with 10.17DT F14 SOSIP after sCD4 treat-
ment. In contrast, robust 19b binding to CH505 M5 G458Y F14
T, F14, and 113C-429GCG gp160s with or without treatment with sCD4 and
s from three independent experiments.
113C-429GCG gp160s. Anti-Influenza hemagglutinin mAb CH65 was used as
n. Histograms shown are representative from three independent experiments.
Cell Reports 38, 110514, March 15, 2022 3

A
B
D
E
F G
C
(legend on next page)
4 Cell Reports 38, 110514, March 15, 2022
Articlell
OPEN ACCESS

Articlell
OPEN ACCESS
SOSIP was observed with or without sCD4 (Figure S1E), demon-
strating that F14 mutations do not intrinsically impair 17b or 19b
binding. Additionally, anti-gp41 nnAb 7B2 against the immuno-
dominant epitope of gp41 (Pincus et al., 2003) showed low bind-
ing to mRNA-expressed 10.17DT Env gp160s, confirming mini-
mal exposure of this gp41 epitope (Figure S1C).
Next, we determined whether expressing Env trimers as trans-
membrane gp160s resulted in trimer base exposure (Turner
et al., 2021). Binding of three Env base antibodies, DH1029,
DH1312, and RM19R IgG were tested (Cottrell et al., 2020; Mar-
tin et al., 2020). Here, we report the negative-stain electron mi-
croscopy (NSEM) structures of DH1029 and DH1312 (Figure S2).
We did not observe binding of neither DH1029 nor DH1312 anti-
bodies and observed only minimal binding of RM19R IgG to
mRNA-expressed CH848 10.17DT, 10.17DT F14, or 113C-
429GCG gp160s (Figures 1E and S1D). Thus, 10.17DT F14 and
113C-429GCG gp160s were stabilized such that they preferen-
tially bound to bnAbs versus nnAbs, had minimal non-neutral-
izing epitope exposure after CD4 triggering, and limited expo-
sure of the trimer base.
CH848 10.17DT gp160 mRNA-LNP elicited autologoustier 2 neutralizing antibodies in vivo
On the basis of stability and antigenicity, we selected CH848
10.17DT F14 and 113C-429GCG gp160s to test their immunoge-
nicity in heterozygous V3-glycan bnAb DH270 UCA heavy- and
light-chain (VH+/–, VL
+/–) knock-in (DH270 UCA KI) mice (Saunders
et al., 2019). mRNAs encoding 10.17DT F14 or 113C-429GCG
gp160s were encapsulated in LNP for immunization (Figure 2A).
Mice immunized with 10.17DT F14 or 113C-429GCG gp160
mRNA-LNP developed serum-binding IgGs to 10.17DT trimer
and gp120 monomer (Figures 2B, S3A, and S3B). Three
10.17DT F14- and four 113C-429GCG gp160 mRNA-LNP-vacci-
nated mice had IgGs binding to the linear CH848 V3 peptide
epitope, indicating low levels of V3 loop exposure in vivo (Figures
2B and S3C). We observed non-detectable levels of binding to
gp41 in both groups of mice, suggesting that the gp41 on stabi-
lized 10.17DT gp160 Env was not immunogenic in vivo (Fig-
Figure 2. Immunogenicity of CH848 10.17DT gp160 mRNA-LNP in mice
(A) Immunization schema in DH270 UCA KI mice with CH848 10.17DT F14 and 1
(B)Week 5 serum IgG binding to CH848 10.17DT SOSIP trimer, 10.17D11 gp120,
under the curve (logAUC). Each dot represents an individual mouse (n = 6 each
difference was observed between two groups (p > 0.05).
(C) Measurement of serum IgG that block DH1029 binding to CH848 10.17DT SO
mice were used to show the induction of base-binding antibodies after SOSIP tri
blocking.
(D) Week 5 serum neutralizing antibody titers measured in TZM-bl reporting cells w
leukemia virus (MuLV) was used as negative control. Neutralization titers are repo
No significant statistical difference was observed between the two groups (p > 0
(E) Improbable G57R andR98Tmutation frequencies in DH270 VH KI gene after CH
10.17DT DS Sortase ferritin NP protein immunization and empty LNP immunizat
(F) CH848 10.17DT gp160 mRNA-LNP vaccination induced GC B cell responses
and 10.17DT-specific memory B cells among total memory B cells in splenoc
vaccinated DH270 UCA KI mice are shown. Each dot represents an individual m
(G) CH848 10.17DT gp160 mRNA-LNP vaccination induced GC Tfh cell respons
splenocytes after 10.17DT F14 or 113C-429GCGgp160mRNA-LNP vaccination in
individual mouse. Horizontal bar, mean. No significant statistical difference was o
Exact Wilcoxon Mann-Whitney U test.
See also Figures S3, S4, and S10.
ure S3D). As confirmation of no relevant exposure of the Env
base on 10.17DT F14 or 113C-429GCG gp160s, no DH1029
serum blocking activity was detected (Figure 2C). Both types of
stabilized Env gp160 mRNA-LNP immunization induced serum
binding IgGs to 10.17DT, 10.17DT F14 and 113C-429GCG
gp160s on the surface of transfected 293-F cells (Figure S3F).
Next, we asked whether CH848 10.17DT F14 and 113C-
429GCG gp160 mRNA-LNP immunization in DH270 UCA KI
mice elicited serumneutralizing antibodies. Neutralizing antibody
titers 1 week after the third immunization (week 5) were assessed
with a panel of seven pseudotyped HIV-1 strains sensitive to
either DH270 UCA or DH270 intermediate antibody 4 (DH270
IA4) (Bonsignori et al., 2017; Saunders et al., 2019) (Figures 2D
and S3G). CH848 10.17DT F14 and 113C-429GCG gp160
mRNA-LNP elicited autologous tier 2 neutralizing antibodies
against CH848 10.17DT pseudovirus, with ID50 geometric mean
titers (GMT) of 8103 and 8812, respectively. Neutralization was
dependent on V3-glycan at position N332 in all 10.17DT F14
and four out of six 10.17DT 113C-429GCG-vaccinated mice,
demonstrating the elicitation of bnAb V3-glycan autologous
neutralizing antibodies. Lower titers of neutralizing antibodies
against the V1-glycans restored CH848 10.17 virus were
observed. Strain-specific holes in glycan shield can elicit glycan
hole-targeted autologous tier 2 neutralizing antibodies (Bradley
et al., 2016; Crooks et al., 2015; McCoy et al., 2016). Computa-
tional prediction (Wagh et al., 2018) identified potential glycan
holes at positions 230 and 289 on CH848 10.17 Env (Figure S3E).
We constructed a glycan holes filled CH848 10.17DTmutant with
amino acid mutations D230N H289N, as well as P291S. Compa-
rable neutralization titers were observed against CH848 10.17DT
D230N H289N P291S virus, indicating that most neutralizing an-
tibodies elicitedby vaccinationwere not targeted to glycanholes.
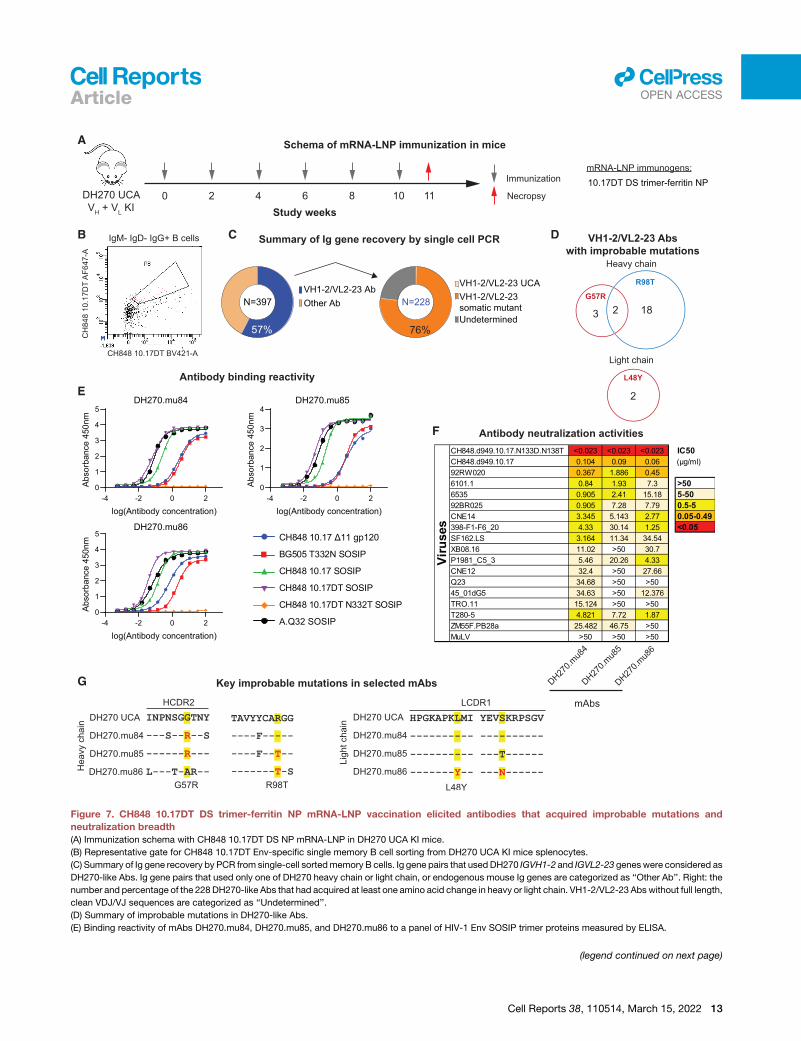
CH848 10.17DT gp160 mRNA-LNP selected for keyDH270 bnAb mutations and elicited germinal centerresponsesNext, splenocytes from 1 week after the third immunization
(week 5) were subjected to next-generation sequencing (NGS)
13C-429GCG gp160 mRNA-LNP.
and V3 loop peptidemeasured by ELISA. Data shown are log-transformed area
group). Error bar, mean ± standard deviation (SD). No significant statistical
SIP trimer. Sera from a group of 10.17DT DS SOSIP trimer protein immunized
mer immunization. Dotted horizontal line indicates background cut-off at 20%
ith a panel of autologous and heterologous tier 2 HIV-1 pseudoviruses. Murine
rted as ID50. Each dot signifies an individual mouse. Horizontal bar, ID50 GMT.
.05).
848 10.17DT gp160mRNA-LNP immunization. Frequencieswere compared to
ion. Each dot represents an individual mouse. Horizontal bar, median.
in spleens. Frequencies of 10.17DT-specific GC B cells among total GC B cells
ytes of CH848 10.17DT F14 or 10.17DT 113C-429GCG gp160 mRNA-LNP
ouse. Horizontal bar: mean. **P < 0.01.
es in spleen. Frequencies of total Tfh and GC Tfh cells among CD4+ T cells in
DH270UCAKImicewere assessed by flow cytometry. Each dot represents an
bserved between the two groups (p > 0.05). Significance was determined using
Cell Reports 38, 110514, March 15, 2022 5

Articlell
OPEN ACCESS
analysis. Both CH848 10.17DT F14 and 113C-429GCG gp160
mRNA-LNP selected the critical improbable G57R mutation in
the DH270 UCA VH KI gene that is necessary for the V3-glycan
bnAb B cell lineage to acquire heterologous neutralization
breadth (Bonsignori et al., 2017; Wiehe et al., 2018), with the me-
dians of mutation frequency at 5.4% and 3.4%, respectively.
Induced antibodies also acquired a second key improbable
VH R98T mutation. Frequencies of the improbable VH G57R
and the R98T mutations were comparable to those in a group
of DH270 UCA KI mice immunized with sortase-ligated
10.17DT DS Env ferritin NP protein (Figure 2E; p > 0.05, exact
Wilcoxon Mann-Whitney U test). Thus, in DH270 UCA KI mice,
10.17DT gp160 mRNA-LNP were immunogenic, induced potent
N332-dependent autologous tier 2 neutralizing antibodies, and
selected DH270 antibodies that acquired improbable mutations
required for acquisition of heterologous HIV-1 neutralization.
Splenocytes from week 5 were phenotyped for germinal cen-
ter (GC) responses by flow cytometry, using fluorophore-labeled
10.17DT SOSIP trimer tetramers to detect 10.17DT-specific B
cells (Figure S4). Both 10.17DT F14 and 113C-429GCG gp160
mRNA-LNP elicited 10.17DT-specific GC B cells and memory
B cells (Figure 2F). The average frequencies of 10.17DT-specific
GC B cells among total GC B cell population was 1.42% in the
10.17DT F14 gp160 mRNA-LNP group and 0.95% in the
10.17DT 113C-429GCG mRNA-LNP group. The 10.17DT F14
gp160 mRNA-LNP vaccinated group had higher frequencies of
10.17DT-specific memory B cells among total memory B cells
compared with the 10.17DT 113C-429GCG gp160 mRNA-LNP
vaccinated group (mean at 13.14% versus 3.88%, p < 0.01,
Exact Wilcoxon Mann-Whitney U test). Additionally, 10.17DT
gp160 mRNA-LNP elicited spleen Tfh cell and GC Tfh cell re-
sponses in both groups (Figures 2G and S4).
Antigenicity of modified mRNA-encoded CH84810.17DT SOSIP trimers with stabilizing mutationsThe antigenicity of mRNA-expressed Galanthus nivalis lectin
(GNL)-purified 10.17DT SOSIP trimers was measured by
enzyme-linked immunosorbent assay (ELISA) using a panel of
bnAbs and nnAbs (Figures 1A and 3A; Table S1). Each stabilized
construct encoded by mRNA efficiently bound to V3-glycan
bnAbs 2G12, PGT125, PGT128, and DH270 lineage Abs
DH270 UCA, DH270 IA4, and DH270.1. In particular, mRNA-en-
coded 10.17DT DS SOSIP trimers displayed greater binding
reactivity to bnAbs compared with other stabilizing mutations.
Consistent with our observations with 10.17DT gp160s, the Vt8
and F14/Vt8 mutations decreased DH270 UCA binding to
10.17DT SOSIP trimers. 10.17DT Vt8 and F14/Vt8 SOSIP trimers
also displayed lower binding to trimer-specific bnAb PGT151
compared with 10.17DT v4.1, DS, and F14 SOSIP trimers, sug-
gesting less native-like conformation of Envs. Little to non-
detectable binding to bnAbs was observed with v5.2.8 and
UFO mutations combined (v5.2.8 + UFO). All stabilized con-
structs tested presented low to non-detectable levels of binding
to most nnAbs, except for 10.17DT SOSIPv5.2.8, which dis-
played about binding to nnAbs 19b and F105 compared with
other stabilized Envs tested.
SPR analysis showed that sCD4 treatment of mRNA-ex-
pressed non-stabilized 10.17DT SOSIPv4.1 trimers increased
6 Cell Reports 38, 110514, March 15, 2022
binding of nnAb 17b. In contrast, 10.17DT DS, F14, and F14/
Vt8 showed low to non-detectable binding to 17b with or without
sCD4 treatment. Although 10.17DT Vt8, v5.2.8, and v5.2.8 +
UFO trimers exhibited increased binding to 17b after sCD4 treat-
ment, the binding was at a lower response level compared
with 10.17DT SOSIPv4.1. Similar trends were observed for 19b
binding (Figure 3B).
We next used size exclusion ultra-performance liquid chroma-
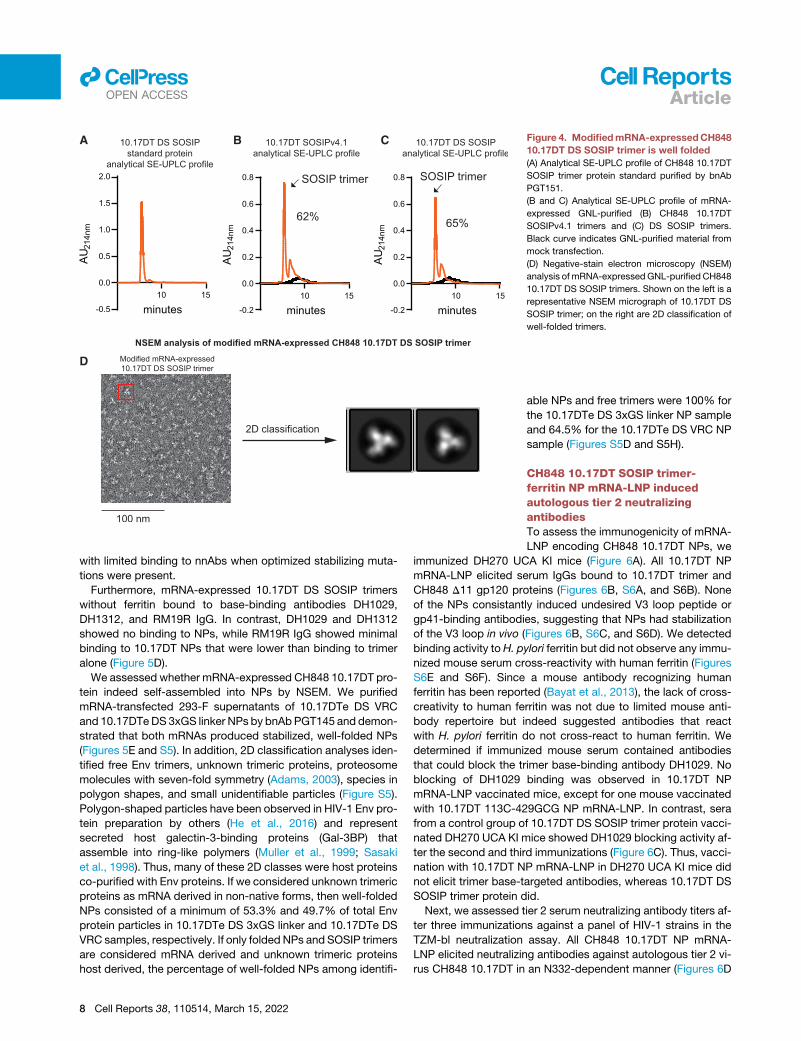
tography (SE-UPLC) to define the folding of mRNA-encoded
CH848 10.17DT SOSIP trimers. The analytical SE-UPLC profile
of bnAb PGT151-purified 10.17DT DS SOSIP trimer protein indi-
cated that a well-folded 10.17DT SOSIP trimer was separated
and eluted from the column, as shown in Figure 4A. GNL-purified
mRNA-expressed 10.17 DT v4.1 and DS SOSIP trimer samples
showed a dominant peak of trimer that was 62% and 65% of the
total peak, respectively (Figures 4B and 4C). NSEM analysis of
10.17DTDS trimer confirmed the expression ofwell-foldedSOSIP
trimers from mRNA-transfected 293-F supernatant (Figure 4D).
Antigenicity of CH848 10.17DT trimer-ferritin NPs withstabilizing mutations encoded by modified mRNAsCH848 10.17DT NPs were designed by gene fusion of the
10.17DT SOSIP trimer gene with Helicobacter pylori (H. pylori)
ferritin gene (FtnA; GenBank: NP_223316) and were tested for
expression, stability, and antigenicity (Figure 5A). Moreover, we
asked whether we could further improve the antigenicity and
immunogenicity of 10.17DT Env by filling the glycan holes at posi-
tions 230and289andby introducing169Kmutation,which is crit-
ical to Env interaction with V2-glycan bnAbs (Doria-Rose et al.,
2012; Lee et al., 2017; McLellan et al., 2011). This 10.17DT Env
trimer (CH848 10.17DTD230NH289NP291SE169K)was termed
‘‘enhanced CH848 10.17DT’’ (CH848 10.17DTe). Since the
linker sequence connecting ferritin and Env protein would affect
the expression and assembly of NPs, we tested 10.17DTe
DS NPs with two different linkers, the sequences of which
were GGGSGGGGSGLSK (termed ‘‘2xGS linker’’) and
GGGSGGGGSGGGGSGLSK (termed ‘‘3xGS linker’’). We also
designed another NP construct using the H. pylori ferritin with a
N19Qmutation,which removedapotential N-linkedglycosylation
site at position 19 and added a glycine and a serine to theC termi-
nus of the ferritin protein (‘‘VRC ferritin’’) (Kanekiyo et al., 2013).
All CH848 10.17DT and 10.17DTe NPs exhibited effective
binding to V3-glycan bnAbs tested (Figure 5B). 10.17DTe NPs
displayed robust binding to V2-glycan bnAbs PGT145, PG9,
and VRC26.25, while V3-glycan bnAb and DH270 lineage anti-
body bindings were comparable to 10.17DT with E169, demon-
strating that 10.17DTe Envs had improved antigenicity by filling
glycan holes and restoring V2-glycan bnAb binding without im-
pairing V3-glycan targeted antibody binding. In contrast,
10.17DT and 10.17DTeNPs showed low to non-detectable bind-
ing to nnAbs (Figure 5B). Specifically, none of the NPs showed
binding to 17b, and the binding to 19b was low, except for
10.17DT 113C-429GCG NPs, indicative of an exposed distal
V3 loop.
SPR analysis following sCD4 treatment showed no increased
binding of nnAb 17b to CH848 10.17DT and 10.17DTe NPs and
low levels of binding of nnAb 19b (Figure 5C). Thus, 10.17DTNPs
could be expressed with mRNAs and bound to bnAbs efficiently

A
B
Figure 3. Antigenicity of modified mRNA-encoded CH848 10.17DT SOSIP trimers with stabilizing mutations
(A) BnAb/bnAb precursor and nnAb binding reactivity to mRNA-expressed CH848 10.17DT SOSIP trimers with various stabilizing mutations measured by ELISA.
Data shown are means of logAUC from three independent experiments.
(B) SPR sensorgrams of nnAb 17b or 19b binding to mRNA-expressed CH848 10.17DT SOSIP trimers with or without sCD4 treatment.
See also Table S1.
Cell Reports 38, 110514, March 15, 2022 7
Articlell
OPEN ACCESS
A
D
B C Figure 4. ModifiedmRNA-expressedCH848
10.17DT DS SOSIP trimer is well folded
(A) Analytical SE-UPLC profile of CH848 10.17DT
SOSIP trimer protein standard purified by bnAb
PGT151.
(B and C) Analytical SE-UPLC profile of mRNA-
expressed GNL-purified (B) CH848 10.17DT
SOSIPv4.1 trimers and (C) DS SOSIP trimers.
Black curve indicates GNL-purified material from
mock transfection.
(D) Negative-stain electron microscopy (NSEM)
analysis of mRNA-expressedGNL-purified CH848
10.17DT DS SOSIP trimers. Shown on the left is a
representative NSEM micrograph of 10.17DT DS
SOSIP trimer; on the right are 2D classification of
well-folded trimers.
Articlell
OPEN ACCESS
with limited binding to nnAbs when optimized stabilizing muta-
tions were present.
Furthermore, mRNA-expressed 10.17DT DS SOSIP trimers
without ferritin bound to base-binding antibodies DH1029,
DH1312, and RM19R IgG. In contrast, DH1029 and DH1312
showed no binding to NPs, while RM19R IgG showed minimal
binding to 10.17DT NPs that were lower than binding to trimer
alone (Figure 5D).
We assessed whether mRNA-expressed CH848 10.17DT pro-
tein indeed self-assembled into NPs by NSEM. We purified
mRNA-transfected 293-F supernatants of 10.17DTe DS VRC
and 10.17DTeDS 3xGS linker NPs by bnAb PGT145 and demon-
strated that both mRNAs produced stabilized, well-folded NPs
(Figures 5E and S5). In addition, 2D classification analyses iden-
tified free Env trimers, unknown trimeric proteins, proteosome
molecules with seven-fold symmetry (Adams, 2003), species in
polygon shapes, and small unidentifiable particles (Figure S5).
Polygon-shaped particles have been observed in HIV-1 Env pro-
tein preparation by others (He et al., 2016) and represent
secreted host galectin-3-binding proteins (Gal-3BP) that
assemble into ring-like polymers (Muller et al., 1999; Sasaki
et al., 1998). Thus, many of these 2D classes were host proteins
co-purified with Env proteins. If we considered unknown trimeric
proteins as mRNA derived in non-native forms, then well-folded
NPs consisted of a minimum of 53.3% and 49.7% of total Env
protein particles in 10.17DTe DS 3xGS linker and 10.17DTe DS
VRC samples, respectively. If only folded NPs and SOSIP trimers
are considered mRNA derived and unknown trimeric proteins
host derived, the percentage of well-folded NPs among identifi-
8 Cell Reports 38, 110514, March 15, 2022
able NPs and free trimers were 100% for
the 10.17DTe DS 3xGS linker NP sample
and 64.5% for the 10.17DTe DS VRC NP
sample (Figures S5D and S5H).
CH848 10.17DT SOSIP trimer-ferritin NP mRNA-LNP inducedautologous tier 2 neutralizingantibodiesTo assess the immunogenicity of mRNA-
LNP encoding CH848 10.17DT NPs, we
immunized DH270 UCA KI mice (Figure 6A). All 10.17DT NP
mRNA-LNP elicited serum IgGs bound to 10.17DT trimer and
CH848 D11 gp120 proteins (Figures 6B, S6A, and S6B). None
of the NPs consistantly induced undesired V3 loop peptide or
gp41-binding antibodies, suggesting that NPs had stabilization
of the V3 loop in vivo (Figures 6B, S6C, and S6D). We detected
binding activity toH. pylori ferritin but did not observe any immu-
nized mouse serum cross-reactivity with human ferritin (Figures
S6E and S6F). Since a mouse antibody recognizing human
ferritin has been reported (Bayat et al., 2013), the lack of cross-
creativity to human ferritin was not due to limited mouse anti-
body repertoire but indeed suggested antibodies that react
with H. pylori ferritin do not cross-react to human ferritin. We
determined if immunized mouse serum contained antibodies
that could block the trimer base-binding antibody DH1029. No
blocking of DH1029 binding was observed in 10.17DT NP
mRNA-LNP vaccinated mice, except for one mouse vaccinated
with 10.17DT 113C-429GCG NP mRNA-LNP. In contrast, sera
from a control group of 10.17DT DS SOSIP trimer protein vacci-
nated DH270 UCA KI mice showed DH1029 blocking activity af-
ter the second and third immunizations (Figure 6C). Thus, vacci-
nation with 10.17DT NP mRNA-LNP in DH270 UCA KI mice did
not elicit trimer base-targeted antibodies, whereas 10.17DT DS
SOSIP trimer protein did.
Next, we assessed tier 2 serum neutralizing antibody titers af-
ter three immunizations against a panel of HIV-1 strains in the
TZM-bl neutralization assay. All CH848 10.17DT NP mRNA-
LNP elicited neutralizing antibodies against autologous tier 2 vi-
rus CH848 10.17DT in an N332-dependent manner (Figures 6D

A
B
C
D E
(legend on next page)
Cell Reports 38, 110514, March 15, 2022 9
Articlell
OPEN ACCESS

Articlell
OPEN ACCESS
and S6G). Comparable neutralizing titers against glycan holes-
filled CH848 10.17DT 230N 289N 291S virus were observed,
indicating that the antibody responses were not directed at
glycan holes but rather were targeted to the V3-glycan bnAb
site. 10.17DTe DS VRCNPmRNA-LNP vaccinatedmice showed
higher neutralizing titers to CH848 10.17DT 230N 289N 291S vi-
ruses compared with 10.17DT DS (p < 0.05, Exact Wilcoxon
Mann-Whitney U test) and 10.17DT 113C-429GCG (p < 0.01,
Exact Wilcoxon Mann-Whitney U test) NP mRNA-LNP vacci-
nated groups. Thus, 10.17DT NPs encoded as mRNA-LNP effi-
ciently elicited tier 2 autologous neutralizing antibodies that tar-
geted the N332-dependent V3 glycan bnAb site.
CH848 10.17DT SOSIP trimer-ferritin NP mRNA-LNPelicited germinal center responses and selected for keyDH270 bnAb mutationsEach of the five CH848 10.17DT NPs selected the improbable VH
G57R mutation in DH270 UCA KI gene, with the highest median
of mutation frequency at 3.2% observed in 10.17DT 113C-
429GCG NP vaccinated mice. Similarly, the R98T mutation in
the DH270 UCA VH KI gene was also selected in all groups by
mRNA-LNP (Figure 6E). Thus, mRNA-LNP encoded 10.17DT
NP immunizations in DH270 UCA KI mice efficiently elicited
key improbable mutations.
Each of the five CH848 10.17DT NP mRNA-LNP induced
10.17DT-specific GC B cells and memory B cells in spleens (Fig-
ure 6F). Tfh cells and GC Tfh cells were also observed in all
10.17DT NP vaccinated mice (Figure 6G). No significant differ-
ence was observed among the five immunization groups (p <
0.05, Exact WilcoxonMann-Whitney U test). Interestingly, empty
LNP immunizations also elicited Tfh cell responses, albeit at a
much lower frequency, consistent with previous observations
that mRNA-LNP have adjuvant effects that favor Tfh cell and
GC responses (Alameh et al., 2021; Pardi et al., 2018a).
CH848 10.17DT trimer-ferritin NP mRNA-LNPimmunization induced heterologous neutralizing mAbsthat acquired improbable mutationsTo further assess antibody responses elicited by CH848
10.17DT NP mRNA-LNP, we injected DH270 UCA KI mice intra-
dermally (i.d.) or intramuscularly (i.m.) with 10.17DT DS NP
mRNA-LNP for six immunizations and sorted 10.17DT-specific
single memory B cells on 96-well plates and amplified immuno-
globulin (Ig) heavy- and light-chain variable regions by PCR (Fig-
ures 7A, 7B, and S7).We cloned a total of 397 Ig heavy- and light-
chain pairs, 228 (57%) pairs of which used DH270 KI genes
IGVH1-2 and IGVL2-23. Among these 228 DH270-like anti-
bodies, 173 (76%) antibodies have acquired at least one amino
Figure 5. Antigenicity of CH848 10.17DT trimer-ferritin NPs with stabili
(A) Design of CH848 10.17DT SOSIP trimer-ferritin fusion NPs.
(B) BnAb/bnAb precursor and nnAb binding reactivity to mRNA-expressed CH8
shown are means of logAUC from three independent experiments.
(C) SPR sensorgrams of nnAb 17b or 19b binding to mRNA-expressed CH848 1
(D) Binding reactivity of Env base antibodies DH1029, DH1312, and RM19R IgG t
measured by ELISA. Data shown are means of logAUC from at least two indepe
(E) Representative NSEM images and 2D classifications of mRNA-expressed, PG
See also Figures S2 and S5 and Table S1.
10 Cell Reports 38, 110514, March 15, 2022
acid mutation (Figures 7C and S7B). We aligned all unique
VH1-2/VL2-23 Ig gene amino acid sequences with bnAb
DH270.6 and found that the Ig heavy-chain group accumulated
a total of 14 out of 19 (74%) DH270.6 probable mutations and
4 out of 8 (50%) DH270.6 improbable mutations (Figure S8).
Similarly, the Ig light-chain group accumulated 6 out of 9 (67%)
DH270.6 probable mutations and 4 out of 6 (67%) improbable
mutations (Figure S9). Specifically, 5 (2%) and 20 (9%) Ig heavy
chains acquired the DH270.6 bnAb G57R and the R98T improb-
able mutations, respectively, and 2 (1%) Ig light chains acquired
the DH270.6 bnAb L48Y improbable mutation. Two Ig heavy
chains had both G57R and R98T mutations (Figure 7D). Binding
reactivity of cloned antibodies was screened in ELISA (Table S3),
and antibodies with heterologous HIV-1 A.Q23 Env binding were
selected for further assessment. Among them, three mAbs
(DH270.mu84, DH270.mu85, and DH270.mu86) were identified
that showed strong binding to CH848 10.17DT, 10.17, and to
heterologous HIV-1 A.Q23 SOSIP trimers (Figure 7E). We as-
sessed neutralization of these three mAbs against a panel of
17 HIV-1 isolates that the first intermediate ancestor antibody
(DH270 IA4) in the DH270 lineage neutralizes (Figure 7F) (Saun-
ders et al., 2019). Each of the 3 mAbs neutralized autologous
tier 2 CH848 viruses and heterologous 92RW020 and 6101.1 vi-
ruses with titers comparable to DH270 IA4 (Saunders et al.,
2019). Antibody DH270.mu84 also neutralized each of the 13
other heterologous HIV-1 isolates tested. Antibodies
DH270.mu85 and DH270.mu86 neutralized 8 and 10 of 13 heter-
ologous isolates, respectively (Figure 7F). DH270.mu84 encoded
the VHG57R improbablemutation, DH270.mu86 encoded the VH
R98T improbable mutation, while DH270.mu85 had both the
G57R and R98T improbable mutations. Additionally,
DH270.mu86 acquired VL S27Y and VL S57N improbable muta-
tions (Figure 7G). One intriguing observation was that
DH270.mu84 makes a tyrosine mutation to the ‘‘invariant’’
cysteine that occurs before CDRL1 (C22Y). We hypothesized
that the C22Y mutation might enhance DH270.mu84 neutraliza-
tion potency or breadth due to increased antibody flexibility
(Klein et al., 2013). In summary, CH848 10.17DT DS NP
mRNA-LNP immunization induced heterologous tier 2 neutral-
izing DH270 antibodies with improbable mutations.
DISCUSSION
A major question addressed here is whether mRNA designs can
incorporate stabilizing mutations in trimers or NPs to optimize
immunogen expression and stability since the protein products
of mRNAs cannot be purified after mRNA-LNP injection
in vivo. In this study, we evaluated mutations that stabilize
zing mutations
48 10.17DT NPs with various stabilizing mutations measured by ELISA. Data
0.17DT NPs with or without sCD4 treatment.
o mRNA-expressed CH848 10.17DT DS SOSIP trimer or CH848 10.17DT NPs
ndent experiments.
T145-purified CH848 10.17DTe DS VRC and 10.17DTe DS 3xGS linker NPs.

A
B
D
E
F G
C
(legend on next page)
Cell Reports 38, 110514, March 15, 2022 11
Articlell
OPEN ACCESS

Articlell
OPEN ACCESS
mRNA-encoded Envs expressed as transmembrane gp160s,
soluble SOSIP trimers, or single-gene mRNA trimer-ferritin
NPs. For mRNAs encoding the V3-glycan germline-targeting
CH848 10.17DT Env (Saunders et al., 2019), we showed that
F14 mutations optimally stabilized transmembrane gp160s, the
DS mutations best stabilized SOSIP trimers and NPs expressed
from mRNA.
mRNA-LNP encoding both transmembrane gp160s and solu-
ble NPs induced high titers of autologous bnAb-targeted tier 2
neutralizing antibodies with groups of mutations present in
bnAb intermediate antibodies in DH270 UCAKI mice (Bonsignori
et al., 2017; Saunders et al., 2019). Moreover, we observed
accumulation of mature bnAb DH270.6 mutations in vaccine-
inducedmAbs, althoughmutations are distributed across all iso-
lated mAbs. The next goal is to have mutations concatenated on
the same bnAb B cell clonal lineage members. To achieve this,
prolonged GC responses or enhanced recruitment of memory
B cells back into GCs will need to be induced by vaccination
to allow bnAb lineage B cell BCRs to acquire functional mutation
required for full bnAb neutralization breadth.
Therefore, we have promising mRNA-LNP vaccine candidates
for priming V3-glycan bnAb DH270 B cell lineage in two different
Env forms: as a transmembrane Env gp160 and as an NP. Trans-
membrane gp160 and NP forms encoded by mRNA-LNPs
showed similar immunogenicity in DH270 UCA KI mice. High-
avidity protein nanoparticles are concentrated in B cell follicles
(Martin et al., 2020; Tokatlian et al., 2019) and activate and recruit
low-affinity B cells (Kato et al., 2020). It is intriguing that mRNA-
LNP-delivered Env gp160s induced serum neutralization titers
and the accumulation of functional improbable bnAb mutations
that were comparable to NPs (Figure S10). The success of using
immunogens in transmembrane form has been demonstrated by
two COVID-19 mRNA-LNP vaccines (Baden et al., 2020; Polack
et al., 2020). However, the mechanisms of mRNA-delivered Env
transmembrane protein in eliciting B cell responses is less clear.
Some studies suggest that B cell recognition of membrane-
associated antigens plays a role of concentrating antigens for
B cell extraction, processing, and presentation (Carrasco and
Batista, 2006).
Several studies published recently explored the use of mRNAs
for HIV-1 vaccines in various forms, such as self-amplifying
mRNA (Aldon et al., 2021; Melo et al., 2019) or mRNA-encoded
virus-like particles (Zhang et al., 2021). These studies highlighted
the versatility of mRNA vaccine platform for HIV-1 vaccine devel-
Figure 6. Immunogenicity of CH848 10.17DT trimer-ferritin NP mRNA-L
(A) Immunization schema with CH848 10.17DT NP mRNA-LNP in DH270 UCA K
(B) Serum IgG binding to CH848 10.17DT SOSIP trimer, 10.17 D11 gp120, and V3
rest of groups). Horizontal bar, mean. Error bar, SD.
(C) Measurement of serum IgG that block DH1029 binding to CH848 10.17DT SO
previous studies were included as positive control. This positive control is the sam
(D) Serum neutralizing titers after CH848 10.17DT SOSIP NP mRNA-LNP immuni
Neutralization titers are reported as ID50. Each dot signifies an individual mouse.
(E) Improbable G57R and R98Tmutation frequencies in DH270 VH KI gene after CH
CH848 10.17DT Sortase ferritin NP protein immunizations and empty LNP immu
(F) Frequencies of CH848 10.17DT-specific GC B cells and memory B cells in sp
(G) Frequencies of total Tfh cells and GC Tfh cells in spleen of 10.17DT NP mRNA
Exact Wilcoxon Mann-Whitney U test, without any p value adjustment for multip
See also Figures S4, S6, and S10.
12 Cell Reports 38, 110514, March 15, 2022
opment and stressed the need for careful studies to compare
and evaluate different mRNA technologies.
Eliciting bnAbs by vaccination has not been successful in hu-
mans. However, studies in HIV-1-infected individuals have
demonstrated that those who make bnAbs have higher levels
of T follicular helper (Tfh) cells (Locci et al., 2013; Moody et al.,
2016). Nucleoside-modified mRNA-LNP vaccines selectively
induce high levels of Tfh cells (Lederer et al., 2020; Pardi et al.,
2018a) and thus will be a key platform to test for bnAb lineage
initiation and selection of B cells with improbable functional mu-
tations that facilitate bnAb maturation. The CH848 10.17DTe DS
NP is currently in goodmanufacturing practice (GMP) production
to investigate the priming of such lineages in humans both as
mRNA-LNPs and as recombinant proteins.
In conclusion, we have demonstrated that single-chain
mRNAs can be designed to encode complex molecules and
that these immunogens are capable of selecting for difficult-to-
elicit improbable mutations critical for broad tier 2 virus neutral-
ization. The complex biology of HIV-1 bnAbs necessitates a vac-
cine strategy that utilizes a series of sequentially administered
Env immunogens that initially expand bnAb precursors and
then select for improbable mutations (Haynes et al., 2012,
2019; Saunders et al., 2019; Wiehe et al., 2018). Manufacturing
of complex nanoparticle protein immunogens in large scale is
faced with significant practical and funding challenges (Mu et
al., 2021). The use of mRNA-LNPs raises the possibility of mak-
ing such a complex immunization regimen both logistically
achievable and potentially cost effective.
Limitations of the studyThere are several limitations in this study. It is technically chal-
lenging to purify modified mRNA-encoded immunogens ex-
pressed in vivo and evaluate the stability directly. However, Pardi
et al. have compared the in vitro transfection of HEK293T cells
and dendritic cells with mRNA-LNPs and demonstrated mRNA
expression in each cell type to be at similar levels (Pardi et al.,
2015). Thus, the careful assessment of immune responses in an-
imal models for the induction of ‘‘off-target’’ antibody responses
targeting undesired epitopes on destabilized Env proteins is crit-
ical. Finally, the assessment of the immunogenicity of mRNA-
LNP in this study was performed in the DH270 UCA KI mouse
model. Although a valuable tool for testing HIV-1 vaccines, a
concern of this model is the relatively high frequency (�10%)
of DH270 UCA B cells among the naive B cell repertoire
NP in mice
I mice.
peptide. Each dot represents an individual mouse (n = 5 in Group 1; n = 6 in the
SIP trimer. 10.17DT DS SOSIP trimer protein vaccinated mice from one of our
e as that plotted in Figure 2C. Dotted horizontal line, 20% background cut-off.
zation measured in TZM-bl reporter cells. MuLV was used as negative control.
Horizontal bar, ID50 GMT.
848 10.17DTmRNA-LNP immunizations. Frequencies were compared to three
nizations. Horizontal bar, median.
lenocytes of 10.17DT NP mRNA-LNP vaccinated mice. Horizontal bar, mean.
-LNP vaccinated mice. Horizontal bar, mean. Significance was determined by
le comparison. *p < 0.05, **p < 0.01.
A
B
E
G
F
C D
Figure 7. CH848 10.17DT DS trimer-ferritin NP mRNA-LNP vaccination elicited antibodies that acquired improbable mutations and
neutralization breadth
(A) Immunization schema with CH848 10.17DT DS NP mRNA-LNP in DH270 UCA KI mice.
(B) Representative gate for CH848 10.17DT Env-specific single memory B cell sorting from DH270 UCA KI mice splenocytes.
(C) Summary of Ig gene recovery by PCR from single-cell sortedmemory B cells. Ig gene pairs that usedDH270 IGVH1-2 and IGVL2-23 geneswere considered as
DH270-like Abs. Ig gene pairs that used only one of DH270 heavy chain or light chain, or endogenous mouse Ig genes are categorized as ‘‘Other Ab’’. Right: the
number and percentage of the 228 DH270-like Abs that had acquired at least one amino acid change in heavy or light chain. VH1-2/VL2-23 Abs without full length,
clean VDJ/VJ sequences are categorized as ‘‘Undetermined’’.
(D) Summary of improbable mutations in DH270-like Abs.
(E) Binding reactivity of mAbs DH270.mu84, DH270.mu85, and DH270.mu86 to a panel of HIV-1 Env SOSIP trimer proteins measured by ELISA.
(legend continued on next page)
Cell Reports 38, 110514, March 15, 2022 13
Articlell
OPEN ACCESS

Articlell
OPEN ACCESS
(Saunders et al., 2019). Future studies in outbred animal models,
such as rhesusmacaques, will be important to further assess the
immunogenicity of mRNA-LNP vaccines.
STAR+METHODS
Detailed methods are provided in the online version of this paper
and include the following:
d KEY RESOURCES TABLE
d RESOURCE AVAILABILITY
B Lead contact
B Materials availability
B Data and code availability
d EXPERIMENTAL MODEL AND SUBJECT DETAILS
B Cell line
B Animals and immunizations
d METHOD DETAILS
B Design of CH848 10.17DT Envs with stabilizing muta-
tions
B Nucleoside-modified mRNA production
B Nucleoside-modified mRNA-LNP production
B Nucleoside-modified mRNA transfection in 293-F cell
line
B Evaluation of expression and folding of modified
mRNA-expressed CH848 10.17DT gp160s, SOSIP tri-
mers, and trimer-ferritin NPs
B Mouse serological analysis by ELISA
B HIV-1 pseudovirus neutralization assay
B Next-generation sequencing (NGS)
B Antibody sequence analysis
B Flow cytometric phenotyping of GC responses
B Isolation of CH848 10.17DT-specific neutralizing
monoclonal antibodies (mAbs)
d QUANTIFICATION AND STATISTICAL ANALYSIS
SUPPLEMENTAL INFORMATION
Supplemental information can be found online at https://doi.org/10.1016/j.
celrep.2022.110514.
ACKNOWLEDGMENTS
We thank Holly Zoeller for assistance with SE-UPLC assays; Cindy Bowman,
Grace Stevens, and Austin Harner for help with animal studies; Victoria Gee-
Lai and Maggie Barr for help with ELISA assays, and Matthew Gardner at
Emory University and Michael Farzan at Scripps Research Institute Florida
for providing eCD4-Ig. We acknowledge Andrew Ward and Christopher Cot-
trell at Scripps Research Institute and Marit van Gils at Academisch Medisch
Centrum Universiteit van Amsterdam for kindly sharing with us RM19R IgG
plasmid. We also thank Cynthia Nagel for project management. This project
was supported by NIH, NIAID, Division of AIDS Integrated Preclinical and Clin-
ical AIDS Vaccine Development Grant AI135902 and by NIAID, Division of
AIDS Consortia for HIV/AIDS Vaccine Development (CHAVD) Grant
UM1AI144371.
(F) DH270.mu84, DH270.mu85, and DH270.mu86 neutralization activity against a
50% of virus replication (IC50) in TZM-bl assay. MuLV was used as negative con
(G) Improbable mutations in mAbs DH270.mu84, DH270.mu85, and DH270.mu8
quences. Mutations are highlighted, and improbable mutations are shown in red
See also Figures S7–S9 and Table S3.
14 Cell Reports 38, 110514, March 15, 2022
AUTHOR CONTRIBUTIONS
Conceptualization, Z.M., K.O.S., D.W., and B.F.H.; Methodology, Z.M., R.P.,
K.O.S., and B.F.H.; Animal models, F.A., M.T.; Software, S.V. and K.J.W.;
Investigation, Z.M., K.J.W., K.O.S., R.H., D.W.C., R.P., D.M., A.E., B.K.,
K.W., K.M., R.J.E., A.N., X.L., S.X., M.B., D.M., Q.H., S.V., T.E., M.A.,
W.B.W., N.P., D.W., and B.F.H.; Statistical analysis, Y.W. and W.R.; Writing,
Z.M., K.O.S., K.J.W., and B.F.H.; Funding Acquisition, B.F.H.; Resources,
Y.T., C.B., N.P., D.W., and B.F.H.; Supervision, B.F.H.
DECLARATION OF INTERESTS
B.F.H., K.O.S., and K.W. have patent applications on some of the concepts
and immunogens discussed in this paper.
Received: August 7, 2021
Revised: January 9, 2022
Accepted: February 17, 2022
Published: March 15, 2022
REFERENCES
Abbott, R.K., Lee, J.H., Menis, S., Skog, P., Rossi, M., Ota, T., Kulp, D.W.,
Bhullar, D., Kalyuzhniy, O., Havenar-Daughton, C., et al. (2018). Precursor fre-
quency and affinity determine B cell competitive fitness in germinal centers,
tested with germline-targeting HIV vaccine immunogens. Immunity 48, 133–
146.e6.
Adams, J. (2003). The proteasome: structure, function, and role in the cell.
Cancer Treat Rev. 29, 3–9.
Alameh, M.G., Tombacz, I., Bettini, E., Lederer, K., Sittplangkoon, C.,Wilmore,
J.R., Gaudette, B.T., Soliman, O.Y., Pine, M., Hicks, P., et al. (2021). Lipid
nanoparticles enhance the efficacy of mRNA and protein subunit vaccines
by inducing robust T follicular helper cell and humoral responses. Immunity
54, 2877–2892.e7.
Aldon, Y., McKay, P.F., Moreno Herrero, J., Vogel, A.B., Levai, R., Maison-
nasse, P., Dereuddre-Bosquet, N., Haas, H., Fabian, K., Le Grand, R., et al.
(2021). Immunogenicity of stabilized HIV-1 Env trimers delivered by self-ampli-
fying mRNA. Mol. Ther. Nucleic Acids 25, 483–493.
Allaway, G.P., Davis-Bruno, K.L., Beaudry, G.A., Garcia, E.B., Wong, E.L.,
Ryder, A.M., Hasel, K.W., Gauduin, M.C., Koup, R.A., McDougal, J.S., et al.
(1995). Expression and characterization of CD4-IgG2, a novel heterotetramer
that neutralizes primary HIV type 1 isolates. AIDS Res. Hum. Retroviruses
11, 533–539.
Baden, L.R., El Sahly, H.M., Essink, B., Kotloff, K., Frey, S., Novak, R., Diemert,
D., Spector, S.A., Rouphael, N., Creech, C.B., et al. (2020). Efficacy and safety
of the mRNA-1273 SARS-CoV-2 vaccine. N. Engl. J. Med 384, 403–416.
Baiersdorfer, M., Boros, G., Muramatsu, H., Mahiny, A., Vlatkovic, I., Sahin, U.,
and Kariko, K. (2019). A facile method for the removal of dsRNA contaminant
from in vitro-transcribed mRNA. Mol. Ther. Nucleic Acids 15, 26–35.
Bayat, A.A., Yeganeh, O., Ghods, R., Zarnani, A.H., Ardekani, R.B., Mahmoudi,
A.R., Mahmoudian, J., Haghighat-Noutash, F., and Jeddi-Tehrani, M. (2013).
Production and characterization of a murine monoclonal antibody against hu-
man ferritin. Avicenna J. Med. Biotechnol. 5, 212–219.
Binley, J.M., Sanders, R.W., Master, A., Cayanan, C.S., Wiley, C.L., Schiffner,
L., Travis, B., Kuhmann, S., Burton, D.R., Hu, S.L., et al. (2002). Enhancing the
proteolytic maturation of human immunodeficiency virus type 1 envelope gly-
coproteins. J. Virol. 76, 2606–2616.
panel of 17 HIV-1 isolates. Data shown are antibody concentration that inhibit
trol.
6. Alignment of mAb heavy- and light-chain sequences with DH270 UCA se-
fonts.

Articlell
OPEN ACCESS
Bonsignori, M., Kreider, E.F., Fera, D., Meyerhoff, R.R., Bradley, T., Wiehe, K.,
Alam, S.M., Aussedat, B., Walkowicz, W.E., Hwang, K.K., et al. (2017). Staged
induction of HIV-1 glycan-dependent broadly neutralizing antibodies. Sci.
Transl. Med. 9, eaai7514.
Bonsignori, M., Zhou, T., Sheng, Z., Chen, L., Gao, F., Joyce,M.G., Ozorowski,
G., Chuang, G.Y., Schramm, C.A., Wiehe, K., et al. (2016). Maturation pathway
from germline to broad HIV-1 neutralizer of a CD4-mimic antibody. Cell 165,
449–463.
Bradley, T., Fera, D., Bhiman, J., Eslamizar, L., Lu, X., Anasti, K., Zhang, R., Su-
therland, L.L., Scearce, R.M., Bowman, C.M., et al. (2016). Structural con-
straints of vaccine-induced tier-2 autologous HIV neutralizing antibodies tar-
geting the receptor-binding site. Cell Rep. 14, 43–54.
Carrasco, Y.R., and Batista, F.D. (2006). B cell recognition of membrane-
bound antigen: an exquisite way of sensing ligands. Curr. Opin. Immunol.
18, 286–291.
Cottrell, C.A., van Schooten, J., Bowman, C.A., Yuan, M., Oyen, D., Shin, M.,
Morpurgo, R., van der Woude, P., van Breemen, M., Torres, J.L., et al. (2020).
Mapping the immunogenic landscape of near-native HIV-1 envelope trimers in
non-human primates. PLoS Pathog. 16, e1008753.
Crooks, E.T., Tong, T., Chakrabarti, B., Narayan, K., Georgiev, I.S., Menis, S.,
Huang, X., Kulp, D., Osawa, K., Muranaka, J., et al. (2015). Vaccine-elicited tier
2 HIV-1 neutralizing antibodies bind to quaternary epitopes involving glycan-
deficient patches proximal to the CD4 binding site. PLoS Pathog. 11,
e1004932.
de Taeye, S.W., Ozorowski, G., Torrents de la Pena, A., Guttman, M., Ju-
lien, J.P., van den Kerkhof, T.L., Burger, J.A., Pritchard, L.K., Pugach, P.,
Yasmeen, A., et al. (2015). Immunogenicity of stabilized HIV-1 envelope tri-
mers with reduced exposure of non-neutralizing epitopes. Cell 163, 1702–
1715.
Doria-Rose, N.A., Georgiev, I., O’Dell, S., Chuang, G.Y., Staupe, R.P., McLel-
lan, J.S., Gorman, J., Pancera, M., Bonsignori, M., Haynes, B.F., et al. (2012). A
short segment of the HIV-1 gp120 V1/V2 region is amajor determinant of resis-
tance to V1/V2 neutralizing antibodies. J. Virol. 86, 8319–8323.
Fellinger, C.H., Gardner, M.R., Weber, J.A., Alfant, B., Zhou, A.S., and Farzan,
M. (2019). eCD4-Ig limits HIV-1 escape more effectively than CD4-ig or a
broadly neutralizing antibody. J. Virol. 93, e00443.
Guenaga, J., Dubrovskaya, V., de Val, N., Sharma, S.K., Carrette, B., Ward,
A.B., and Wyatt, R.T. (2015). Structure-guided redesign increases the propen-
sity of HIV env to generate highly stable soluble trimers. J. Virol. 90, 2806–
2817.
Havenar-Daughton, C., Lee, J.H., and Crotty, S. (2017). Tfh cells and HIV
bnAbs, an immunodominance model of the HIV neutralizing antibody genera-
tion problem. Immunol. Rev. 275, 49–61.
Havenar-Daughton, C., Sarkar, A., Kulp, D.W., Toy, L., Hu, X., Deresa, I., Ka-
lyuzhniy, O., Kaushik, K., Upadhyay, A.A., Menis, S., et al. (2018). The human
naive B cell repertoire contains distinct subclasses for a germline-targeting
HIV-1 vaccine immunogen. Sci. Transl. Med. 10, eaat0381.
Haynes, B.F., Burton, D.R., and Mascola, J.R. (2019). Multiple roles for HIV
broadly neutralizing antibodies. Sci. Transl. Med. 11, eaaz2686.
Haynes, B.F., Fleming, J., St Clair, E.W., Katinger, H., Stiegler, G., Kunert, R.,
Robinson, J., Scearce, R.M., Plonk, K., Staats, H.F., et al. (2005). Cardiolipin
polyspecific autoreactivity in two broadly neutralizing HIV-1 antibodies. Sci-
ence 308, 1906–1908.
Haynes, B.F., Kelsoe, G., Harrison, S.C., and Kepler, T.B. (2012). B-cell-line-
age immunogen design in vaccine development with HIV-1 as a case study.
Nat. Biotechnol. 30, 423–433.
Haynes, B.F., Shaw, G.M., Korber, B., Kelsoe, G., Sodroski, J., Hahn, B.H.,
Borrow, P., and McMichael, A.J. (2016). HIV-host interactions: implications
for vaccine design. Cell Host Microbe 19, 292–303.
He, L., de Val, N., Morris, C.D., Vora, N., Thinnes, T.C., Kong, L., Azadnia, P.,
Sok, D., Zhou, B., Burton, D.R., et al. (2016). Presenting native-like trimeric
HIV-1 antigens with self-assembling nanoparticles. Nat. Commun. 7, 12041.
Henderson, R., Lu, M., Zhou, Y., Mu, Z., Parks, R., Han, Q., Hsu, A.L., Carter,
E., Blanchard, S.C., Edwards, R.J., et al. (2020). Disruption of the HIV-1 Enve-
lope allosteric network blocks CD4-induced rearrangements. Nat. Commun.
11, 520.
Hraber, P., Seaman, M.S., Bailer, R.T., Mascola, J.R., Montefiori, D.C., and
Korber, B.T. (2014). Prevalence of broadly neutralizing antibody responses
during chronic HIV-1 infection. AIDS 28, 163–169.
Huang, D., Abbott, R.K., Havenar-Daughton, C., Skog, P.D., Al-Kolla, R., Gro-
schel, B., Blane, T.R., Menis, S., Tran, J.T., Thinnes, T.C., et al. (2020). B cells
expressing authentic naive human VRC01-class BCRs can be recruited to
germinal centers and affinity mature in multiple independent mouse models.
Proc. Natl. Acad. Sci. U S A 117, 22920–22931.
Jardine, J., Julien, J.P., Menis, S., Ota, T., Kalyuzhniy, O., McGuire, A., Sok, D.,
Huang, P.S., MacPherson, S., Jones, M., et al. (2013). Rational HIV immu-
nogen design to target specific germline B cell receptors. Science 340,
711–716.
Jayaraman, M., Ansell, S.M., Mui, B.L., Tam, Y.K., Chen, J., Du, X., Butler, D.,
Eltepu, L., Matsuda, S., Narayanannair, J.K., et al. (2012). Maximizing the po-
tency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew.
Chem. Int. Ed. Engl. 51, 8529–8533.
Kanekiyo, M., Wei, C.J., Yassine, H.M., McTamney, P.M., Boyington, J.C.,
Whittle, J.R., Rao, S.S., Kong, W.P., Wang, L., and Nabel, G.J. (2013). Self-
assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1
antibodies. Nature 499, 102–106.
Kato, Y., Abbott, R.K., Freeman, B.L., Haupt, S., Groschel, B., Silva, M., Menis,
S., Irvine, D.J., Schief, W.R., and Crotty, S. (2020). Multifaceted effects of an-
tigen valency on B cell response composition and differentiation in vivo. Immu-
nity 53, 548–563.e8.
Klein, F., Diskin, R., Scheid, J.F., Gaebler, C., Mouquet, H., Georgiev, I.S., Pan-
cera, M., Zhou, T., Incesu, R.B., Fu, B.Z., et al. (2013). Somaticmutations of the
immunoglobulin framework are generally required for broad and potent HIV-1
neutralization. Cell 153, 126–138.
Kong, L., He, L., de Val, N., Vora, N., Morris, C.D., Azadnia, P., Sok, D., Zhou,
B., Burton, D.R., Ward, A.B., et al. (2016). Uncleaved prefusion-optimized
gp140 trimers derived from analysis of HIV-1 envelope metastability. Nat.
Commun. 7, 12040.
Kwon, Y.D., Pancera,M., Acharya, P., Georgiev, I.S., Crooks, E.T., Gorman, J.,
Joyce, M.G., Guttman, M., Ma, X., Narpala, S., et al. (2015). Crystal structure,
conformational fixation and entry-related interactions of mature ligand-free
HIV-1 Env. Nat. Struct. Mol. Biol. 22, 522–531.
Lederer, K., Castano, D., Gomez Atria, D., Oguin, T.H., 3rd,Wang, S., Manzoni,
T.B., Muramatsu, H., Hogan, M.J., Amanat, F., Cherubin, P., et al. (2020).
SARS-CoV-2 mRNA vaccines foster potent antigen-specific germinal center
responses associated with neutralizing antibody generation. Immunity 53,
1281–1295.e5.
Lee, J.H., Andrabi, R., Su, C.Y., Yasmeen, A., Julien, J.P., Kong, L., Wu, N.C.,
McBride, R., Sok, D., Pauthner, M., et al. (2017). A broadly neutralizing anti-
body targets the dynamic HIV envelope trimer apex via a long, rigidified, and
anionic beta-hairpin structure. Immunity 46, 690–702.
Lee, J.H., Hu, J.K., Georgeson, E., Nakao, C., Groschel, B., Dileepan, T., Jen-
kins, M.K., Seumois, G., Vijayanand, P., Schief, W.R., et al. (2021). Modulating
the quantity of HIV Env-specific CD4 T cell help promotes rare B cell responses
in germinal centers. J. Exp. Med. 218, e20201254.
Liao, H.X., Levesque, M.C., Nagel, A., Dixon, A., Zhang, R.J., Walter, E., Parks,
R., Whitesides, J., Marshall, D.J., Hwang, K.K., et al. (2009). High-throughput
isolation of immunoglobulin genes from single human B cells and expression
as monoclonal antibodies. J. Virol. Methods 158, 171–179.
Locci, M., Havenar-Daughton, C., Landais, E., Wu, J., Kroenke, M.A., Arle-
hamn, C.L., Su, L.F., Cubas, R., Davis, M.M., Sette, A., et al. (2013). Human
circulating PD-1+CXCR3-CXCR5+ memory Tfh cells are highly functional
and correlate with broadly neutralizing HIV antibody responses. Immunity
39, 758–769.
Cell Reports 38, 110514, March 15, 2022 15

Articlell
OPEN ACCESS
Maier, M.A., Jayaraman, M., Matsuda, S., Liu, J., Barros, S., Querbes, W.,
Tam, Y.K., Ansell, S.M., Kumar, V., Qin, J., et al. (2013). Biodegradable lipids
enabling rapidly eliminated lipid nanoparticles for systemic delivery of RNAi
therapeutics. Mol. Ther. 21, 1570–1578.
Martin, J.T., Cottrell, C.A., Antanasijevic, A., Carnathan, D.G., Cossette, B.J.,
Enemuo, C.A., Gebru, E.H., Choe, Y., Viviano, F., Fischinger, S., et al. (2020).
Targeting HIV Env immunogens to B cell follicles in nonhuman primates
through immune complex or protein nanoparticle formulations. NPJ Vaccin.
5, 72.
Mascola, J.R., D’Souza, P., Gilbert, P., Hahn, B.H., Haigwood, N.L., Morris, L.,
Petropoulos, C.J., Polonis, V.R., Sarzotti, M., andMontefiori, D.C. (2005). Rec-
ommendations for the design and use of standard virus panels to assess
neutralizing antibody responses elicited by candidate human immunodefi-
ciency virus type 1 vaccines. J. Virol. 79, 10103–10107.
McCoy, L.E., vanGils, M.J., Ozorowski, G., Messmer, T., Briney, B., Voss, J.E.,
Kulp, D.W., Macauley, M.S., Sok, D., Pauthner, M., et al. (2016). Holes in the
glycan shield of the native HIV envelope are a target of trimer-elicited neutral-
izing antibodies. Cell Rep. 16, 2327–2338.
McGuire, A.T., Dreyer, A.M., Carbonetti, S., Lippy, A., Glenn, J., Scheid, J.F.,
Mouquet, H., and Stamatatos, L. (2014). HIV antibodies. Antigen modification
regulates competition of broad and narrow neutralizing HIV antibodies. Sci-
ence 346, 1380–1383.
McGuire, A.T., Hoot, S., Dreyer, A.M., Lippy, A., Stuart, A., Cohen, K.W., Jar-
dine, J., Menis, S., Scheid, J.F., West, A.P., et al. (2013). Engineering HIV en-
velope protein to activate germline B cell receptors of broadly neutralizing anti-
CD4 binding site antibodies. J. Exp. Med. 210, 655–663.
McLellan, J.S., Pancera, M., Carrico, C., Gorman, J., Julien, J.P., Khayat, R.,
Louder, R., Pejchal, R., Sastry, M., Dai, K., et al. (2011). Structure of HIV-1
gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 480,
336–343.
Melo, M., Porter, E., Zhang, Y., Silva, M., Li, N., Dobosh, B., Liguori, A., Skog,
P., Landais, E., Menis, S., et al. (2019). Immunogenicity of RNA replicons en-
coding HIV env immunogens designed for self-assembly into nanoparticles.
Mol. Ther. 27, 2080–2090.
Moody, M.A., Pedroza-Pacheco, I., Vandergrift, N.A., Chui, C., Lloyd, K.E.,
Parks, R., Soderberg, K.A., Ogbe, A.T., Cohen, M.S., Liao, H.X., et al.
(2016). Immune perturbations in HIV-1-infected individuals who make broadly
neutralizing antibodies. Sci. Immunol. 1, aag0851.
Mu, Z., Haynes, B.F., and Cain, D.W. (2021). HIV mRNA vaccines-progress
and future paths. Vaccines 9, 134.
Muller, S.A., Sasaki, T., Bork, P., Wolpensinger, B., Schulthess, T., Timpl, R.,
Engel, A., and Engel, J. (1999). Domain organization of Mac-2 binding protein
and its oligomerization to linear and ring-like structures. J. Mol. Biol. 291,
801–813.
Pancera, M., Lai, Y.T., Bylund, T., Druz, A., Narpala, S., O’Dell, S., Schon, A.,
Bailer, R.T., Chuang, G.Y., Geng, H., et al. (2017). Crystal structures of trimeric
HIV envelope with entry inhibitors BMS-378806 and BMS-626529. Nat. Chem.
Biol. 13, 1115–1122.
Pardi, N., Hogan, M.J., Naradikian, M.S., Parkhouse, K., Cain, D.W., Jones, L.,
Moody, M.A., Verkerke, H.P., Myles, A., Willis, E., et al. (2018a). Nucleoside-
modified mRNA vaccines induce potent T follicular helper and germinal center
B cell responses. J. Exp. Med. 215, 1571–1588.
Pardi, N., Hogan, M.J., Pelc, R.S., Muramatsu, H., Andersen, H., DeMaso,
C.R., Dowd, K.A., Sutherland, L.L., Scearce, R.M., Parks, R., et al. (2017).
Zika virus protection by a single low-dose nucleoside-modified mRNA vacci-
nation. Nature 543, 248–251.
Pardi, N., Muramatsu, H., Weissman, D., and Kariko, K. (2013). In vitro tran-
scription of long RNA containing modified nucleosides. Methods Mol. Biol.
969, 29–42.
Pardi, N., Parkhouse, K., Kirkpatrick, E., McMahon, M., Zost, S.J., Mui, B.L.,
Tam, Y.K., Kariko, K., Barbosa, C.J., Madden, T.D., et al. (2018b). Nucleo-
side-modified mRNA immunization elicits influenza virus hemagglutinin
stalk-specific antibodies. Nat. Commun. 9, 3361.
16 Cell Reports 38, 110514, March 15, 2022
Pardi, N., Tuyishime, S., Muramatsu, H., Kariko, K., Mui, B.L., Tam, Y.K.,
Madden, T.D., Hope, M.J., and Weissman, D. (2015). Expression kinetics of
nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various
routes. J. Control. Release 217, 345–351.
Pincus, S.H., Fang, H., Wilkinson, R.A., Marcotte, T.K., Robinson, J.E., and Ol-
son,W.C. (2003). In vivo efficacy of anti-glycoprotein 41, but not anti-glycopro-
tein 120, immunotoxins in a mouse model of HIV infection. J. Immunol. 170,
2236–2241.
Polack, F.P., Thomas, S.J., Kitchin, N., Absalon, J., Gurtman, A., Lockhart, S.,
Perez, J.L., Perez Marc, G., Moreira, E.D., Zerbini, C., et al. (2020). Safety and
efficacy of the BNT162b2mRNA covid-19 vaccine. N. Engl. J.Med. 383, 2603–
2615.
Sahin, U., Muik, A., Derhovanessian, E., Vogler, I., Kranz, L.M., Vormehr, M.,
Baum, A., Pascal, K., Quandt, J., Maurus, D., et al. (2020). COVID-19 vaccine
BNT162b1 elicits human antibody and TH1 T cell responses. Nature 586,
594–599.
Sasaki, T., Brakebusch, C., Engel, J., and Timpl, R. (1998). Mac-2 binding pro-
tein is a cell-adhesive protein of the extracellular matrix which self-assembles
into ring-like structures and binds beta 1 integrins, collagens and fibronectin.
EMBO J. 17, 1606–1613.
Saunders, K.O., Pardi, N., Parks, R., Santra, S., Mu, Z., Sutherland, L.,
Scearce, R., Barr, M., Eaton, A., Hernandez, G., et al. (2021). Lipid nanopar-
ticle encapsulated nucleoside-modified mRNA vaccines elicit polyfunctional
HIV-1 antibodies comparable to proteins in nonhuman primates. NPJ Vaccin.
6, 50.
Saunders, K.O., Verkoczy, L.K., Jiang, C., Zhang, J., Parks, R., Chen, H.,
Housman,M., Bouton-Verville, H., Shen, X., Trama, A.M., et al. (2017). Vaccine
induction of heterologous tier 2 HIV-1 neutralizing antibodies in animal models.
Cell Rep. 21, 3681–3690.
Saunders, K.O., Wiehe, K., Tian, M., Acharya, P., Bradley, T., Alam, S.M., Go,
E.P., Scearce, R., Sutherland, L., Henderson, R., et al. (2019). Targeted selec-
tion of HIV-specific antibody mutations by engineering B cell maturation. Sci-
ence 366, eaay7199.
Steichen, J.M., Lin, Y.C., Havenar-Daughton, C., Pecetta, S., Ozorowski, G.,
Willis, J.R., Toy, L., Sok, D., Liguori, A., Kratochvil, S., et al. (2019). A general-
ized HIV vaccine design strategy for priming of broadly neutralizing antibody
responses. Science 366, eaax4380.
Tokatlian, T., Read, B.J., Jones, C.A., Kulp, D.W., Menis, S., Chang, J.Y.H.,
Steichen, J.M., Kumari, S., Allen, J.D., Dane, E.L., et al. (2019). Innate immune
recognition of glycans targets HIV nanoparticle immunogens to germinal cen-
ters. Science 363, 649–654.
Tran, E.E., Borgnia, M.J., Kuybeda, O., Schauder, D.M., Bartesaghi, A., Frank,
G.A., Sapiro, G., Milne, J.L., and Subramaniam, S. (2012). Structural mecha-
nism of trimeric HIV-1 envelope glycoprotein activation. PLoS Pathog. 8,
e1002797.
Turner, H.L., Andrabi, R., Cottrell, C.A., Richey, S.T., Song, G., Callaghan, S.,
Anzanello, F., Moyer, T.J., Abraham,W., Melo, M., et al. (2021). Disassembly of
HIV envelope glycoprotein trimer immunogens is driven by antibodies elicited
via immunization. Sci. Adv. 7, eabh2791.
Wagh, K., Kreider, E.F., Li, Y., Barbian, H.J., Learn, G.H., Giorgi, E., Hraber,
P.T., Decker, T.G., Smith, A.G., Gondim, M.V., et al. (2018). Completeness
of HIV-1 envelope glycan shield at transmission determines neutralization
breadth. Cell Rep. 25, 893–908.e7.
Ward, A.B., andWilson, I.A. (2017). The HIV-1 envelope glycoprotein structure:
nailing down a moving target. Immunol. Rev. 275, 21–32.
Wiehe, K., Bradley, T., Meyerhoff, R.R., Hart, C., Williams,W.B., Easterhoff, D.,
Faison, W.J., Kepler, T.B., Saunders, K.O., Alam, S.M., et al. (2018). Functional
relevance of improbable antibody mutations for HIV broadly neutralizing anti-
body development. Cell Host Microbe 23, 759–765.e6.
Williams, W.B., Meyerhoff, R.R., Edwards, R.J., Li, H., Manne, K., Nicely, N.I.,
Henderson, R., Zhou, Y., Janowska, K., Mansouri, K., et al. (2021). Fab-dimer-
ized glycan-reactive antibodies are a structural category of natural antibodies.
Cell 184, 2955–2972.e5.

Articlell
OPEN ACCESS
Zhang, P., Gorman, J., Geng, H., Liu, Q., Lin, Y., Tsybovsky, Y., Go, E.P., Dey,
B., Andine, T., Kwon, A., et al. (2018). Interdomain stabilization impairs CD4
binding and improves immunogenicity of the HIV-1 envelope trimer. Cell
Host Microbe 23, 832–844.e6.
Zhang, P., Narayanan, E., Liu, Q., Tsybovsky, Y., Boswell, K., Ding, S., Hu, Z.,
Follmann, D., Lin, Y., Miao, H., et al. (2021). A multiclade env-gag VLP mRNA
vaccine elicits tier-2 HIV-1-neutralizing antibodies and reduces the risk of het-
erologous SHIV infection in macaques. Nat. Med. 27, 2234–2245.
Zhang, R., Verkoczy, L., Wiehe, K., Munir Alam, S., Nicely, N.I., Santra, S.,
Bradley, T., Pemble, C.W.t., Zhang, J., Gao, F., et al. (2016). Initiation of im-
mune tolerance-controlled HIV gp41 neutralizing B cell lineages. Sci. Transl.
Med. 8, 336ra362.
Cell Reports 38, 110514, March 15, 2022 17

Articlell
OPEN ACCESS
STAR+METHODS
KEY RESOURCES TABLE
REAGENT or RESOURCE SOURCE IDENTIFIER
Antibodies
Goat F(ab’)2 Anti-Human IgG - (Fab’)2 (PE) Abcam Cat#ab98606; RRID:AB_10672217
FITC Rat Anti-Mouse IgG1, Clone A85-1 BD Biosciences Cat#553443; RRID:AB_394862
FITC Rat Anti-IgG2a/IgG2b, Clone R2-40 BD Biosciences Cat#553399; RRID:AB_394837
FITC Rat Anti-Mouse IgG3, Clone R40-82 BD Biosciences Cat#553403; RRID:AB_394840
PerCP/Cyanine5.5 anti-mouse
CD21/CD35 (CR2/CR1) antibody
Biolegend Cat#123416; RRID:AB_1595490
PE anti-MU/HU GL7 Antigen
(T/B Cell Act. Marker) antibody
Biolegend Cat#144608; RRID:AB_2562926
PE-CF594 Rat Anti-Mouse CD93
(Early B Lineage) Clone AA4.1
BD Biosciences Cat#563805; RRID:AB_2738431
PE/Cyanine5 anti-mouse TER-
119/Erythroid Cells antibody
Biolegend Cat#116210; RRID:AB_313711
PE-Cy7 Rat Anti-IgM, Clone R6-60.2 BD Biosciences Cat#552867; RRID:AB_394500
APC-R700 Rat Anti-Mouse CD19 Clone 1D3 BD Biosciences Cat#565473; RRID:AB_2739253
BV510 Rat Anti-Mouse IgD Clone 11-26C.1 BD Biosciences Cat#563110; RRID:AB_2737003
BV605 Hamster Anti-Mouse CD95
Clone Jo2
BD Biosciences Cat#740367; RRID:AB_2740099
BV650 Rat Anti-Mouse CD45R/B220
Clone RA3-6B2
BD Biosciences Cat#563893; RRID:AB_2738471
BV711 Rat Anti-Mouse CD138 Clone 281-2 BD Biosciences Cat#563193; RRID:AB_2631190
BV786 Rat Anti-Mouse CD23 Clone B3B4 BD Biosciences Cat#563988; RRID:AB_2738526
FITC Rat Anti-Mouse CD4 Clone RM4-5 BD Biosciences Cat#553047; RRID:AB_394583
PE Rat anti-Mouse CD25 Clone 7D4 BD Biosciences Cat#558642; RRID:AB_1645250
PE-CF594 Hamster Anti-Mouse CD279 (PD-1)
Clone J43
BD Biosciences Cat#562523; RRID:AB_2737634
PE-Cy7 Rat Anti-Mouse CD62L Clone MEL-14 BD Biosciences Cat#560516; RRID:AB_1645257
Biotin Rat Anti-Mouse CD185 (CXCR5)
Clone 2G8
BD Biosciences Cat#551960; RRID:AB_394301
APC-R700 Rat Anti-Mouse CD8a Clone 53-6.7 BD Biosciences Cat#564983; RRID:AB_2739032
BV421 Rat Anti-Mouse CD127 Clone SB/199 BD Biosciences Cat#566300; RRID:AB_2739672
BV510 Hamster Anti-Mouse CD3e
Clone 145-2C11
BD Biosciences Cat#563024; RRID:AB_2737959
BV570 anti-mouse/human CD11b
Antibody, Clone M1/70
Biolegend Cat#101233; RRID:AB_10896949
BV605 Rat Anti-Mouse CD90.2 Clone 53-2.1 BD Biosciences Cat#563008; RRID:AB_2665477
Brilliant Violet 650TM anti-mouse
NK-1.1 Antibody, Clone PK136
Biolegend Cat#108736; RRID:AB_2563159
BV711 Rat Anti-Mouse CD44
Clone IM7
BD Biosciences Cat#563971; RRID:AB_2738518
Goat polyclonal Secondary Antibody
to Rabbit IgG - H&L (HRP)
Abcam Cat# ab97080; RRID:AB_10679808
Mouse Anti-Monkey IgG-HRP SouthernBiotech Cat# 4700-05; RRID:AB_2796069
High Sensitivity Streptavidin-HRP Thermo Fisher Scientific N/A
Chemicals, peptides, and recombinant proteins
Galanthus Nivalis Lectin (GNL), Agarose bound Vector Laboratories Cat#AL-1243
Streptavidin, AF647 conjugate Thermo Fisher Scientific Cat#S21374; RRID:AB_2336066
(Continued on next page)
e1 Cell Reports 38, 110514, March 15, 2022

Continued
REAGENT or RESOURCE SOURCE IDENTIFIER
m1-pseudouridine-50-triphosphate TriLink Cat#N-1081
CleanCap TriLink Cat#N-7413
HIV Env recombinant protein This paper N/A
Critical commercial assays
TransIT-mRNA Transfection Kit Mirus Cat#MIR2250
LIVE/DEAD Fixable Aqua Dead Cell Stain Kit Thermo Fisher Scientific Cat#L34957
SureBlue Reserve TMB 1-Component
Microwell Peroxidase Substrate
Seracare Cat#5120-0083
RNeasy Mini Kit Qiagen Cat#74104
SMARTer Mouse BCR IgG H/K/L Profiling Kit Takara Cat#634422
AMPure XP Beckman Coulter Cat#A63881
MiSeq Reagent Kit v3 (600 cycle) Illumina Cat#MS-102-3003
LIVE/DEADTM Fixable Near-IR Dead Cell Stain Kit Thermo Fisher Scientific Cat#L34975
Experimental models: Cell lines
Freestyle 293-F cell Thermo Fisher Scientific Cat#R79007
Human cell line TZM-bl NIH ARRRP Cat#8129
Experimental models: Organisms/strains
Heterozygous (VH+/-, VL+/-) DH270 UCA knock-in mouse Saunders et al., 2019 N/A
Recombinant DNA
See Table S2 for plasmids used for in vitro
transcription of modified mRNAs
This paper N/A
Software and algorithms
FACSDIVA BD Biosciences N/A
FlowJo version 10 BD Biosciences N/A
Prism version 9 GraphPad Software N/A
Articlell
OPEN ACCESS
RESOURCE AVAILABILITY
Lead contactFurther information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Barton F.
Haynes ([email protected]).
Materials availabilityNew plasmids generated in this paper will be shared by the lead contact upon request.
Data and code availabilityd Original data reported in this paper will be shared by the lead contact upon request.
d This paper did not generate original code.
d Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
EXPERIMENTAL MODEL AND SUBJECT DETAILS
Cell lineFreestyle 293-F cell line (Thermo Fisher Scientific, Cat# R79007) was purchased from Thermo Fisher and cultured in Freestyle 293
ExpressionMedium (Thermo Fisher Scientific, Cat# 12338-026). Cells weremaintained in 8%CO2 at 37�Cat a density between 0.33
106/mL to 33 106/mL. Mycoplasma test was performed when a new stock vial was thawed at Duke University Cell Culture Facility.
Animals and immunizationsThe DH270 UCA KI mice has been previously described (Saunders et al., 2019). For CH848 10.17DT gp160 mRNA-LNP immuniza-
tions, 8-12 weeks old mice were used. 12 DH270 UCA KI mice were split into two groups (N = 6 each group), each group had at least
one female mice. Mice were immunized intramuscularly (i.m.) with 20 mg of mRNA-LNP encoding CH848 10.17DT F14 gp160 and
Cell Reports 38, 110514, March 15, 2022 e2

Articlell
OPEN ACCESS
CH848 10.17DT 113C-429GCG gp160 every two weeks for three times. For CH848 10.17DT DS NP immunizations, 6-12 weeks old
mice were used, and each group had at least two female mice. Immunizations were performed similarly, except that the second and
the third immunizations were only one week apart. Control group mice were injected with 20 mg of empty LNP. Bleeding was per-
formed one week after each immunization. Necropsy was performed one week after the third immunization and blood, spleen,
and lymph nodes were collected. All mice were cared for in a facility accredited by the Association for Assessment and Accreditation
of Laboratory Animal Care International (AAALAC). All study protocol and all veterinarian procedures were approved by the Duke Uni-
versity Institutional Animal Care and Use Committee (IACUC).
METHOD DETAILS
Design of CH848 10.17DT Envs with stabilizing mutationsTen stabilization designs were tested in nucleoside-modified mRNA-encoded CH848 10.17DT Envs as transmembrane gp160s, sol-
uble SOSIP trimers, or trimer-ferritin NPs (Figures 1A and Table S1). The DS mutations introduce a disulfide bond in the closed Env
trimer and prevents CD4-triggered exposure of the CCR5 co-receptor binding site and the V3 loop (Kwon et al., 2015). The F14 mu-
tations are designed based on a structure of BG505 SOSIP trimer complexedwith BMS-626529, a small molecule that blocks soluble
CD4 (sCD4)-induced Env rearrangements (Pancera et al., 2017) and stabilize the SOSIP trimer by decoupling the allosteric confor-
mational changes triggered by CD4 binding (Henderson et al., 2020). Vt8 mutations stabilize the V3 loop in the prefusion, V1/V2-
coupled state (Henderson et al., 2020). The 113C-429GCG and 113C-431GCGmutations link the Env gp120 subunit inner and outer
domains through a neo-disulfide bond, resulting in prefusion stabilized Env trimer with impaired CD4 binding (Zhang et al., 2018). For
soluble gp140 trimer stabilization, we also tested SOSIPv4.1, v5.2.8 and uncleaved prefusion-optimized (UFO) mutations. Mutations
in v4.1 introduce hydrophobic amino acids to disfavor solvent exposure of the V3 loop and modify gp41 in the SOSIP.664 trimer,
which improve trimer formation and thermostability and decrease V3 loop exposure (de Taeye et al., 2015). The UFO design replaces
the bend between alpha helices in HR1 with a computationally designed linker and aims to minimize the metastability of HIV-1 gp140
trimer (Kong et al., 2016). Mutations in the v5.2.8 design are designed based upon v4.1 and combine an additional disulfide bond and
eight trimer-derived mutations that stabilize BG505 SOSIP trimers (Guenaga et al., 2015). Both v4.1 and v5.2.8 include an improved
hexa-arginine furin cleavage site R6 (Binley et al., 2002).
Nucleoside-modified mRNA productionModified mRNAs were produced by in vitro transcription using T7 RNA polymerase (Megascript, Ambion) on linearized plasmids en-
coding codon-optimized CH848 10.17DT gp160s, CH848 10.17DT SOSIP trimers, or CH848 10.17DT trimer-ferritin NPs. All the HIV-
1 modified mRNA constructs used in this study and their corresponding plasmids were listed in Table S2. One-methylpseudouridine
(m1J)-50-triphosphate (TriLink, Cat# N-1081), instead of UTP was used to produce nucleoside-modified mRNAs. Modified mRNAs
contain 101 nucleotide-long polyadenylation tails for optimized expression. Modified CH848 10.17DT SOSIPv4.1 trimer and CH848
10.17DT SOSIPv5.2.8 trimer mRNAs were capped using ScriptCap m7G capping system and ScriptCap 20-O-methyl-transferase kit
(ScriptCap, CellScript) (Pardi et al., 2013). Capping of all other in vitro transcribed mRNAs was performed co-transcriptionally using
the trinucleotide cap1 analog, CleanCap (TriLink, Cat# N-7413). All mRNAs were purified by cellulose purification, as described
(Baiersdorfer et al., 2019). All mRNAs were analyzed by agarose gel electrophoresis and were stored frozen at �20�C.
Nucleoside-modified mRNA-LNP productionNucleoside-modified mRNAs were encapsulated in LNP for mouse immunizations as previously described (Jayaraman et al., 2012;
Maier et al., 2013). Modified mRNAs in aqueous phase were rapidly mixed with a solution of lipids dissolved in ethanol. LNP formu-
lation contains ionizable cationic lipid (proprietary to Acuitas)/phosphatidylcholine/cholesterol/PEG-lipid. The cationic lipid and LNP
composition are described in US patent US10,221,127.
Nucleoside-modified mRNA transfection in 293-F cell line293-F cells were diluted to 0.7 3 106 cells/mL 24 h before transfection. On the next day, cells were diluted again to 13 106/mL and
seeded into tissue culture plates for transfection. 3 mg of mRNAs expressing gp160s were transfected into 6 mL of cells. For soluble
SOSIP trimers, 30 mL of cells were transfected with 12 mg of SOSIP-expressing mRNAs and 3 mg of Furin mRNAs. Transfection vol-
ume were doubled to 60 mL for trimer-ferritin NPs. TransIT-mRNA Transfection Kit (Mirus Cat# MIR2250) was used for mRNA trans-
fection following the manufacturer’s instructions. Transfected cells were cultured at 37�C with 8% CO2 and shaking at 120 rpm for
48 h (for gp160) or 72 h (for SOSIP trimers and trimer-ferritin NPs) before harvest.
Evaluation of expression and folding of modified mRNA-expressed CH848 10.17DT gp160s, SOSIP trimers, andtrimer-ferritin NPsThe expression and folding ofmodifiedmRNA-encodedCH848 10.17DT transmembrane gp160s, Soluble SOSIP trimers, and trimer-
ferritin NPs were defined as follows. For CH848 10.17DT transmembrane gp160s, flow cytometry was used to measure binding of
a panel of bnAbs and nnAbs. BnAb binding reactivity indicated successful expression of gp160 Envs on cell surface with desired
antigenicity. Binding of nnAbs 17b and 19b measured the ability of various stabilizing mutations to keep the gp160 Envs in prefusion
e3 Cell Reports 38, 110514, March 15, 2022

Articlell
OPEN ACCESS
conformation and to decrease the exposure of non-neutralizing epitopes CCR5 binding site and distal V3 loop. Additionally, 7B2
binding was used to measure the exposure of gp41.
For CH848 10.17DT soluble trimer, total Env formswere purified byGalanthus nivalis lectin (GNL) frommodifiedmRNA-transfected
293-F cell supernatant. A panel of nnAbs and bnAbs were used in ELISA to measure the expression of non-neutralizing and neutral-
izing epitopes. Binding of nnAbs 17b and 19b after CD4 triggering were measured by SPR. To assess the percent of trimeric Envs in
GNL-purified materials, size-exclusion ultra-performance liquid chromatography (SE-UPLC) analysis was performed with PGT151
affinity-purified CH848 10.17DT SOSIP trimer as a standard. Finally, Negative-stain Electron Microscopy (NSEM) analysis was per-
formed to confirm trimer formation in GNL-purified 293-F transfection supernatant.
Similar antigenicity measurement and SPR analysis was performed on GNL-purified 293-F cell supernatant transfected with modi-
fied mRNA expressing CH848 10.17DT trimer-ferritin NPs to evaluate the expression of well-folded Env trimers on the ferritin nano-
particle. Additionally, transfected 293-F supernatant were affinity purified with PGT145-conjugated beads, which exclude host
glycan proteins that may be purified by GNL. PGT145-purified materials were analyzed by NSEM to confirm the assembly of
CH848 10.17DT DS ferritin NPs.
Flow cytometry
Binding of bnAbs to CH848 10.17DT gp160s was performed by flow cytometry as previously described (Henderson et al., 2020; Sa-
unders et al., 2021). Briefly, modified mRNA-transfected 293-F cells were harvested 48 h after transfection and were washed once
with 1% BSA in PBS. Then, cells were incubated with 10 mg/mL of bnAbs in V-bottom 96-well plates for 30 min at 4�C. Cells were
then washed with 1% BSA in PBS and incubated with Goat F(ab’)2 Anti-Human IgG - (Fab’)2 (PE) (Abcam Cat# ab98606,
RRID:AB_10672217) for 30 min at 4�C in dark. Then, cells were washed once with PBS and dead cells were stained with LIVE/
DEAD Fixable Aqua Dead Cell Stain Kit (Invitrogen Cat# L34957, 1:1000 dilution in PBS) for 15 min at 4�C in dark, then washed twice
and re-suspended in 1%BSA in PBS. Flow cytometric data were acquired on a LSRII High-throughput system using FACSDIVA soft-
ware (BD Biosciences) and were analyzed with FlowJo software (FlowJo). The percentage of 293-F cells that were PE positive was
shown in the results.
Measurement of binding of nnAbs after CD4 treatment has been described previously (Henderson et al., 2020). Briefly, mRNA-
transfected 293-F cells were first incubated with 20 mg/mL of soluble CD4 (sCD4), eCD4-Ig or CD4-IgG2 for 10 min at 4�C. Cellswere washed once with 1% BSA in PBS and then incubated with 10 mg/mL of nnAbs 17b, 19b or 7B2 for 30 min at 4�C. Then, cellswere incubated with Goat F(ab’)2 Anti-Human IgG - (Fab’)2 (PE) and dead cells were stained with LIVE/DEAD Fixable Aqua Dead Cell
Stain Kit. Data acquisition and analysis were the same as described above.
Galanthus nivalis lectin purification of SOSIP trimers and trimer-ferritin NPs
293-F cells transfected with modified mRNAs expressing SOSIP trimers or trimer-ferritin NPs were harvested 72 h after transfection
and were centrifuged for 30 min at 3000 rpm to remove cells and debris. Supernatant were first filtered using a 0.22 mm vacuum filter
and were then concentrated by 50-fold using 10 kDaMWCO concentrators. Concentrated supernatant was incubated with 200 mL of
agarose boundGalanthus nivalis lectin (GNL) (Vector Laboratories Cat# AL-1243) with gentle rotation at 4�C overnight. The next day,
GNL agarose beads were washed with MES wash buffer (20 mM MES, 130 mM NaCl, 10 mM CaCl2 pH 7.0) for three times, and
SOSIP trimers or trimer-ferritin NPs were eluted by 500 mM Methyl alpha-D-mannopyranoside in MES wash buffer. The eluates
were then dialyzed to 10 mM Tris-HCl pH8 500 mM NaCl using 30kDa MWCO spin concentrators. GNL-purified SOSIP trimers or
trimer-ferritin NPs were snap-freezed and stored in �80�C.Enzyme-linked immunosorbent assay (ELISA)
Binding reactivity of modified mRNA-expressed CH848 10.17DT SOSIP trimers and trimer-ferritin NPs to bnAbs and nnAbs was
measured by enzyme-linked immunosorbent assay (ELISA). In brief, HIV-1 antibodies were coated onto 384-well assay plates in
0.1MSodium bicarbonate overnight at 4�C. GNL-purified SOSIP trimers or trimer-ferritin NPs with serial dilutions were then captured
on the plates. Next, poly-serum fromCH848 10.17DT-immunized rhesusmacaque was incubated for 1 h at room temperature. Then,
Mouse Anti-Monkey IgG-HRP (SouthernBiotech Cat# 4700-05, RRID:AB_2796069) was incubated for 1 h at room temperature and
plates were developed with SureBlue Reserve TMB 1-Component Microwell Peroxidase Substrate (Seracare Cat# 5120-0083) for
15 min and were stopped with 1% HCl solution. Absorbance at 450 nm were determined by SpectraMax Plus 384 microplate reader
(Molecular Devices) and log area-under-curve (log AUC) were calculated using Prism (Graphpad) and shown in figures.
The base binding antibody assay was performed similarly. Briefly, base binding antibody DH1029 was coated onto plates to cap-
ture samples. Then, a rabbit serum was incubated before detection with Goat polyclonal Secondary Antibody to Rabbit IgG - H&L
(HRP) (Abcam Cat# ab97080, RRID:AB_10679808). The plate development, data acquisition, and analysis were the same as
described above.
Size-exclusion ultra-performance liquid chromatography (SE-UPLC)
Size exclusion chromatography of modified mRNA-expressed GNL-purified CH848 10.17DT SOSIP trimers was performed using a
Waters Acquity H-Class Bio UPLC System with a Waters Acquity UPLC BEH SEC 450A, 2.5 mm, 4.63 150 mm column (Waters Cor-
poration). An isocratic elution with a mobile phase of 20 mM sodium phosphate 300 mM NaCl pH 7.4, and a flow rate of 0.2 mL/min,
was used for the analysis with a quaternary pump. Samples and protein standardsweremaintained at 5–8�C in the auto-sampler rack
prior to injection at a volume of 10 mL. Samples and protein standards with a concentration greater than 1.0 mg/mL were diluted to a
down to 1.0 mg/mL using Type 1 water. The column temperature was set to 30�C with detection at a wavelength of 214 nm using a
photodiode array detector.
Cell Reports 38, 110514, March 15, 2022 e4

Articlell
OPEN ACCESS
Negative-stain electron microscopy (NSEM)
Negative-stain electronmicroscopy (NESM) analysis of modifiedmRNA-expressed CH848 10.17DT SOSIP trimers and trimer-ferritin
NPs were performed as previously described (Saunders et al., 2017; Williams et al., 2021).
Surface plasmon resonance (SPR)
SPR analyses of modified mRNA-expressed SOSIP proteins incubated with and without sCD4 against distal V3 loop antibody 19b
and CCR5 binding site antibody 17b were obtained using the Biacore S200 instrument (Cytiva). Antibodies 19b and 17b were immo-
bilized onto a CM3 sensor chip to a level of 2000-4000RU. A negative control Influenza IgG1 antibody (CH65) was also immobilized
onto the sensor chip for reference subtraction. Modified mRNA-expressed GNL-purified CH848 10.17DT SOSIP trimers or trimer-
ferritin NPs were diluted down in HBS-N 1x running buffer to 0.5–2.0 mg and incubated with a 2-83 higher dose of soluble CD4
(4.4 mg) (Progenics Therapeutics). Proteins incubated with and without sCD4 were injected over the sensor chip surface using the
High performance injection type for 180s at 30 mL/min. The protein was then allowed to dissociate for 600s followed by sensor surface
regeneration of two 20 s injections of glycine pH 2.0 at a flow rate of 50 mL/min. Results were analyzed using the BIAevaluation Soft-
ware (Cytiva). Protein binding to the CH65 immobilized sensor surface as well as buffer binding were used for double reference sub-
traction to account for non-specific protein binding and signal drift.
Mouse serological analysis by ELISASerum IgG antigen binding assay
Serum IgG binding to HIV-1 antigens was measured by ELISA as previously described (Saunders et al., 2019).
DH1029 blocking assay
DH1029 blocking by vaccinated mouse sera was performed in ELISA. Briefly, 384-well assay plate were coated with 2 mg/mL
PGT145. Then, 0.125 mg/mL of CH848 10.17DT E169K SOSIP trimer were captured for 1 h at room temperature. Next, mouse
sera at 1:50 dilution or DH1029 mAb in serial dilution (2-fold dilution starting at 2 ug/mL) were incubated for 1 h. Next, biotinylated
DH1029 were added to the plate at 0.05 mg/mL for 1 h and binding were detected by High Sensitivity Streptavidin-HRP (Thermo
Fisher Scientific, Cat #21130). Plate development and data acquisition were the same as described above.
HIV-1 pseudovirus neutralization assayNeutralization assays were performed in TZM-bl reporter cells as described (Mascola et al., 2005). The analyses of possible glycan
holes onCH848 10.17DTEnvwas performed usingmethod described before (Wagh et al., 2018). Potential glycan holes were found at
amino acid positions 230 and 289. To restore glycosylation at these two sites, Asparagine (N) mutations at positions 230 and 289
(230N 289N) were made. In addition, a Serine (S) mutation at position 291 (291S) was made to complete the NXT/S glycosylation
sequence.
Next-generation sequencing (NGS)We performed next-generation sequencing (NGS) on mouse antibody heavy and light chain variable genes using an Illumina
sequencing platform. First, RNA was purified from splenocytes using a RNeasy Mini Kit (Qiagen, Cat# 74,104). Purified RNA was
quantified via Nanodrop (Thermo Fisher Scientific) and used to generate Illumina-ready heavy and light chain sequencing libraries
using the SMARTer Mouse BCR IgG H/K/L Profiling Kit (Takara, Cat# 634,422). Briefly, 1 mg of total purified RNA from splenocytes
was used for reverse transcription with Poly dT provided in the SMARTer Mouse BCR kit for cDNA synthesis. Heavy and light chain
geneswere then separately amplified using a 50 RACE approachwith reverse primers that anneal in themouse IgG constant region for
heavy chain genes and IgK for the light chain genes (SMARTer Mouse BCR IgG H/K/L Profiling Kit). The DH270 UCA KI mousemodel
has the light chain gene knocked into the kappa locus, therefore kappa primers provided in the SMARTer Mouse BCR kit were used
for light chain gene library preparation. 5 mL of cDNA was used for heavy and light chain gene amplification via two rounds of PCR;
PCR1 used 18 cycles and PCR2 used 12 cycles. During PCR2, Illumina adapters and indexeswere added. Illumina-ready sequencing
libraries were then purified and size-selected by AMPure XP (Beckman Coulter, Cat# A63881) using kit recommendations. The heavy
and light chain libraries per mouse were indexed separately, thus allowing us to deconvolute the mouse-specific sequences during
analysis. Libraries were quantified using QuBit Fluorometer (Thermo Fisher). Mice were pooled by groups for sequencing on the Il-
lumina MiSeq Reagent Kit v3 (600 cycle) (Illumina, Cat# MS-102-3003) using read lengths of 301/301 with 20% PhiX.
Antibody sequence analysisNGS data analysis and the analysis of improbable mutation frequencies was performed as described (Wiehe et al., 2018). Animals
with <1000 NGS reads likely reflect technical errors during NGS library preparation and thus were excluded from analysis.
Flow cytometric phenotyping of GC responsesFor immunophenotyping of murine B cells and Tfh cells, spleens from immunized mice one week after the third immunization were
processed into single-cell suspensions and treated with ACK lysis buffer to remove red blood cells. Splenocytes (2 3 106) were
suspended in 100 mL PBS/2% FBS. To detect antigen-specific B cells, fluorochrome-mAb conjugates and fluorochrome-conjugated
CH848 10.17DT Envs were prepared as a master mix at 23 concentration, then 100 mL of 23 master mix was added to an equal
volume of cells (Figure S4). Staining for T cell subsets was conducted in the same manner, with the additional step for detection
e5 Cell Reports 38, 110514, March 15, 2022

Articlell
OPEN ACCESS
of biotinylated mAb with Streptavidin–APC. Cells were incubated at 4�C for 20 minutes, then washed with PBS. Cells were resus-
pended in 100 mL PBS containing Near-IR Live/Dead (Thermo Fisher Scientific) at 1:1000, and incubated at room temperature for
20 min. Cells were washed in PBS/2% FBS, then re-suspended in PBS/2% formaldehyde. Cells were analyzed on a BD LSRII
(BD Biosciences). Data were analyzed using FlowJo v10 (FlowJo).
Isolation of CH848 10.17DT-specific neutralizing monoclonal antibodies (mAbs)Antibody cloning, screening, and mAbs expression
Immunoglobulin (Ig) gene were cloned from sorted single B cells as previously described (Liao et al., 2009). Briefly, complementary
DNA (cDNA) of Ig genes were amplified by reverse-transcription with SuperScript III First-Strand Synthesis System (Thermo Fisher
Scientific, Cat# 18080051) using random hexamer oligonucleotides as primers. Ig gene cDNA was then used as template in nested
PCR for heavy and light chain gene amplification using AmpliTaq Gold 360 Master Mix (Thermo Fisher Scientific, Cat #4398881).
Mouse Ig-specific primers and DH270 variable region-specific primers were used to amplify mouse endogenous Ig genes and
DH270 KI Ig genes. Agarose gel electrophoresis was used to identify positive PCR amplification and Ig genes were recovered by
Sanger sequencing. Following sequencing, contigs of PCR amplicon sequences were assembled, and Ig genes were inferred
with human Ig gene library andmouse Ig gene library in Cloanalyst. PCR reactionswith successful Ig sequence recoverywere purified
using AMPure XP kit (Beckman Coulter, Cat# A63881). Purified PCR product was used for overlapping PCR to generate a linear anti-
body expression cassette. The expression cassette was transiently transfected with into 293i cells with ExpiFectamine 293 Trans-
fection Kit (Thermo Fisher Scientific, Cat# A14525). The supernatant was harvested 72 h after transfection and screened in ELISA
binding assays with a panel of protein of interests. The genes of selected heavy chains were synthesized with human IgG1 backbone
(GenScript). Kappa and lambda chains were synthesized similarly. To express mAbs plasmids were prepared for transient transfec-
tion using the Plasmid Plus Mega Kit (Qiagen, Cat #12981). Heavy and light chain plasmids were co-transfected into 293i cells using
ExpiFectamine 293 Transfection Kit for antibody production.
QUANTIFICATION AND STATISTICAL ANALYSIS
Exact Wilcoxon Mann-Whitney U tests were performed without any adjustment for multiple comparisons. Significant results were
indicated in figures and figure legends as: * P < 0.05; ** P< 0.01.
Cell Reports 38, 110514, March 15, 2022 e6

Cell Reports, Volume 38
Supplemental information
mRNA-encoded HIV-1 Env trimer ferritin
nanoparticles induce monoclonal antibodies
that neutralize heterologous HIV-1 isolates in mice
Zekun Mu, Kevin Wiehe, Kevin O. Saunders, Rory Henderson, Derek W. Cain, RobertParks, Diana Martik, Katayoun Mansouri, Robert J. Edwards, AmandaNewman, Xiaozhi Lu, Shi-Mao Xia, Amanda Eaton, Mattia Bonsignori, DavidMontefiori, Qifeng Han, Sravani Venkatayogi, Tyler Evangelous, Yunfei Wang, WesRountree, Bette Korber, Kshitij Wagh, Ying Tam, Christopher Barbosa, S. MunirAlam, Wilton B. Williams, Ming Tian, Frederick W. Alt, Norbert Pardi, DrewWeissman, and Barton F. Haynes

Table S1. Stabilizing mutations studied for modified mRNA-encoded CH848 10.17DT transmembrane gp160s, soluble SOSIP trimers, and trimer-ferritin nanoparticles. Related to Figure 1.
Env form Design Mutations Reference
Transmembrane DS 201C-433C Kwon et al., 2015
gp160s F14 68I, 204V, 208L, 255L Henderson et al., 2020
Vt8 203M, 300L, 302L, 320M, 422M Henderson et al., 2020
F14/Vt8 F14+Vt8 Henderson et al., 2020
113C-429GCG 113C-429C, 428G, 430G Zhang et al., 2018
113C-431GCG 113C-431C, 430G, 432G Zhang et al., 2018
Soluble SOSIP trimers v4.1
501C-605C, 559P, R6, ΔMPER, 535M, 543N/Q,
316W, 64K De Taeye et al., 2015
v5.2.8 V4.1, 66R, 73C-561C, L165, Q432, R429, K65, T106, E49,
D47, R500 Guenaga et al., 2015
DS 201C-433C Kwon et al., 2015
F14 68I, 204V, 208L, 255L Henderson et al., 2020
Vt8 203M, 300L, 302L, 320M, 422M Henderson et al., 2020
F14/Vt8 F14+Vt8 Henderson et al., 2020
UFO HR-1 re-designed, 501C-605C Kong et al., 2016
v5.2.8+UFO v5.2.8+UFO Guenaga et al., 2015; Kong et al., 2016
Multimeric trimer-ferritin NPs DS 201C-433C Kwon et al., 2015
113C-429GCG 113C-429C, 428G, 430G Zhang et al., 2018

Table S2. Plasmids used for in vitro transcription of each mRNA construct. Related to Figure 1.
mRNA Construct Plasmid
CH848 10.17DT gp160 ccap1-TEV-CH848-10.17-DTgp160-A101
CH848 10.17DT DS gp160 ccap1-TEV-CH848-10.17-DT-DS gp160-A101
CH848 10.17DT F14 gp160 ccap1-TEV-CH0848-10.17-DTgp160_F14-A101
CH848 10.17DT Vt8 gp160 ccap1-TEV-CH848-10.17-DTgp160-VT8-A101
CH848 10.17DT F14/Vt8 gp160 ccap1-TEV-CH0848-10.17-DTgp160_F14_VT8-A101
CH848 10.17DT 113C-429GCG gp160
ccap1-TEV-CH0848.d949.10.17gp160_N133DN138T_113C-429GCG-A101
CH848 10.17DT 113C-431GCG gp160
ccap1-TEV-CH0848.d949.10.17gp160_N133DN138T_113C-431GCG-A101
CH848 10.17DT SOSIPv4.1 trimer
cap1-TEV-CH848.3.D0949.10.17CHIM1.6R.SOSIP.664V4.1_N133DN138T-A101
CH848 10.17DT SOSIPv5.2.8 trimer
cap1-TEV-CH848.3.D0949.10.17CHIM.SOSIPV5.2.8_N133DN138T-A101
CH848 10.17DT F14 SOSIP trimer ccTEV-CH848-10.17-DT-F14 SOSIP-A101
CH848 10.17DT Vt8 SOSIP trimer ccTEV-CH848-10.17-DT-Vt8 SOSIP-A101
CH848 10.17DT F14/Vt8 SOSIP trimer ccTEV-CH848-10.17-DT-F14Vt8 SOSIP-A101
CH848 10.17DT 113C-429GCG SOSIP trimer
ccap1-TEV-CH848.3.10.17ch.SOSIP_DT_113C-429GCG-A101
CH848 10.17DT UFO-BG SOSIP trimer
ccap1-TEV-CH848.d949.10.17 N133D N138T SOSIP.UFO-BG-A101
CH848 10.17DT DS trimer-ferritin NP
ccap1-TEV-CH848.3.D0949.10.17chDS.SOSIP_N133DN138T-ferritin-A101
CH848 10.17DT 113C-429GCG trimer-ferritin NP
ccap1-TEV-CH848.10.17ch.SOSIP_DT_113C-429GCG-ferritin-A101
CH848 10.17DTe DS 3xGS linker trimer-ferritin NP
CH848.3.D0949.10.17_N113D_N138T_D230N_H289N_P291S_E169K_DS_SOSIP_LSK_ferritin_3xGSlinker
CH848 10.17DTe DS 2xGS linker trimer-ferritin NP
CH848.3.D0949.10.17_N113D_N138T_D230N_H289N_P291S_E169K_DS_SOSIP_LSK_ferritin_2xGSlinker
CH848 10.17DTe DS VRC trimer-ferritin NP
CH848.3.D0949.10.17_N133D_N138T_D230N_H289N_P291S_E169K_DS.SOSIP_VRCferritin

CH65 2G12 DH1029 DH1312 RM19R IgG10.17DT 74.90 3800.33 177.00 77.23 255.3310.17DT F14/Vt8 78.90 1346.00 152.33 69.63 217.0010.17DT DS 93.13 2569.33 165.00 83.50 270.3310.17DT Vt8 79.20 1417.00 161.33 74.73 351.0010.17DT F14 79.23 4204.00 173.00 76.17 281.6710.17DT 113C-429GCG 77.10 4200.33 156.67 76.50 255.0010.17DT 113C-431GCG 82.30 3572.67 156.33 73.90 273.67
D
CH65 17b 19b 7B2
No CD4 0.07 4.77 3.32 6.63+sCD4 0.07 43.93 26.77 8.29
+eCD4-Ig 0.07 72.77 69.83 26.67+CD4-IgG2 0.13 58.40 54.07 8.01
No CD4 0.36 0.68 13.68 21.53+sCD4 0.41 0.53 14.73 20.83
+eCD4-Ig 0.38 0.37 17.84 20.60+CD4-IgG2 0.39 0.53 22.74 19.60
No CD4 0.07 1.61 1.86 6.53+sCD4 0.04 1.95 2.06 6.17
+eCD4-Ig 0.05 3.86 4.19 5.44+CD4-IgG2 0.04 2.48 2.60 6.16
No CD4 0.70 0.86 2.41 11.52+sCD4 0.54 0.87 4.24 11.72
+eCD4-Ig 0.58 11.76 14.60 10.52+CD4-IgG2 0.60 11.78 13.76 11.29
No CD4 0.78 1.13 7.63 5.62+sCD4 0.24 0.40 7.11 6.46
+eCD4-Ig 0.24 0.53 9.56 5.70+CD4-IgG2 0.30 0.49 14.20 6.06
No CD4 0.07 1.88 2.26 7.65+sCD4 0.04 2.03 2.33 8.37
+eCD4-Ig 0.08 1.74 2.25 8.45+CD4-IgG2 0.06 2.11 2.23 10.10
No CD4 0.04 2.62 2.53 10.09+sCD4 0.07 2.67 2.50 9.59
+eCD4-Ig 0.09 2.52 2.40 6.32
+CD4-IgG2 0.14 2.43 2.49 10.02
10.17DT
10.17DT DS
10.17DT F14
10.17DT Vt8
10.17DT F14/Vt8
10.17DT 113C-429GCG
10.17DT 113C-431GCG
CH65 CH01 PG9 PGT125 DH270 UCA10.17DT 0.07 37.30 9.87 82.90 76.0710.17DT DS 0.12 25.33 15.61 64.97 39.6010.17DT F14 0.07 37.57 6.45 85.73 81.6310.17DT Vt8 0.09 13.31 1.65 86.13 34.7710.17DT F14/Vt8 0.09 8.48 1.15 82.50 35.4010.17DT 113C-429GCG 0.07 35.27 0.89 86.37 78.0310.17DT 113C-431GCG 0.04 18.33 0.90 85.87 77.07
A
B C
Singlets75.1
FSC-A
FSC-
HSingle Cells
91.3
SSC-A
SSC-
H
Live cells77.4
SSC-A
LIVE
/DEA
D-B
V510
PE+81.0
DH270 UCA-PE
SSC-
A
Figure S1. Antigenicity of modified mRNA-encoded CH848 10.17DT transmembrane gp160s. Related to Figure 1.(A) Gating strategy for flow cytometric analysis of antibody binding to modified mRNA-expressed CH848 10.17DT gp160s on 293-F cell surface. Figure shows representative gating of DH270 UCA binding to CH848 10.17DT gp160. (B) Binding of bnAbs to modified mRNA-expressed CH848 10.17DT transmembrane gp160s. Data shown were means of PE+ cell percentage among total live cells from three independent experiments.(C) Binding of nnAbs 17b, 19b, and 7B2 to CH848 10.17DT gp160 after CD4 triggering. Three forms of CD4: soluble CD4 (sCD4), eCD4-Ig, and CD4-IgG2 were used. Binding of Abs were detected by Goat anti-Human IgG F(ab’)2 secondary antibody labeled with PE. Data shown were means of PE+ cell percentage among total live cells from three independent experiment.(D) Binding of Env base antibodies to CH848 10.17DT gp160s. Data shown were averages of mean fluorescence intensity (MFI) from three independent experiment. Influenza hemagglutinin Ab CH65 and HIV bnAb 2G12 were used as negative control and positive control, respectively.(E) 17b and 19b binding to F14 stabilized CH848 10.17DT and CH505 M5 G458Y SOSIP trimers measured by SPR. The data demonstrated that F14 mutations do not intrinsically impair 17b or 19b binding.
E

A B C
300 Å
D90° 90°
30 Å
5,868particles
4,672particles
3,984particles
2,399particles
2,831particles
50 Å
300 Å
DH1029
E F G
H90° 90°
300 Å
30 Å
50 Å8,999
particles4,912
particles4,339
particles
300 Å
DH1312
I
J 90° 90°
30 Å
30 Å
DH1029 DH1312 RM19R

Figure S2. NSEM structures of SOSIP complex with DH1029 and DH1312, and comparison to RM19R. Related to Figures 1 and 5.(A) Raw micrograph of DH1029 Fab complexed with CH505M5chim.6R.SOSIP.664v4.1_G458Y/GnTI- Env trimer.(B) Representative 2D class averages of Fab-bound trimers.(C) 3D classification shows two classes with two Fabs bound (gray and yellow), one class with weaker density for one of the Fabs (cyan), and two junk particle classes (purple and pink). The number of particles is indicated below each class.(D) Final reconstruction from the combined particles from the first two classes in C.(E) Raw micrograph for DH1312 Fab complexed with CON-Schim.6R.DS.SOSIP.664_OPT_N130D_N135K_N138S_N141S/293F Env trimer.(F) Representative 2D class averages of Fab-bound trimers.(G) 3D classification shows one class with three Fabs bound (gray), one class with weaker density for one of the Fabs (yellow), and one junk particle classes (cyan). The number of particles is indicated below each class.(H) Final reconstruction from the combined particles from the first class in C.(I) Comparison of SOSIP complexes with DH1029, DH1312, and RM19R show side by side. For RM19R, the cryo-EM map (EMD 21227) has been low-pass filtered to 20 Å. (J) Superposition of DH1029, DH1312 and RM19R shown in three orthoganal views.

10.17DT F14 gp16010.17DT 113C-429GCG gp160
mRNA-LNP immunogens:
CH848 10.17DT trimer CH848 10.17DT Δ11 gp120 CH848 V3 peptide
gp41
A CB
D
10.17DT F14 gp160 mRNA-LNP 10.17DT 113C-429GCG gp160 mRNA-LNP
Mock10.17DT
10.17DT DS
10.17DT 113C-429GCG
101
102
103
104
Cell surface-expressed 10.17DT gp160 Envs
MFI
-1 5 -1 5 -1 5 -1 5
Mock10.17DT
10.17DT DS
10.17DT 113C-429GCG
101
102
103
104
Cell surface-expressed 10.17DT gp160 Envs
MFI
-1 5 -1 5 -1 5 -1 5Week
Week
F
G
MuLV 92RW020CH848.d949.10.17
6101CH848.d949.10.
17DT.N332TQ23
CH848.d949.10.17.DT
CH848.d949.10.17.DT.D230N.H289N.P291S
Immuniza�on
<200 <200 <200 <200 316 <200 3465 4016<200 <200 204 <200 37 <200 4771 6898<200 <200 <200 <200 <20 <200 5641 6797<200 <200 406 <200 56 <200 18342 24220<200 <200 584 <200 <20 <200 8550 11164<200 <200 745 <200 106 <200 19357 21662<200 <200 615 <200 29 <200 15519 13945<200 <200 424 <200 <20 <200 7191 7092<200 <200 480 <200 41 <200 6126 5534<200 <200 331 <200 <20 <200 5569 4540<200 <200 <200 <200 5767 <200 6130 7959<200 <200 <200 <200 23357 <200 20061 16434
ID50nt
<200160-999
1000-9999>10000
IC50 (1/dilu�on) in TZM-bl cells
CH848 10.17DT F14 gp160 mRNA-
LNP
CH848 10.17DT 113C-429GCG gp160 mRNA-
LNP
0 2 4 6
0
5
10
15
Study weeks
Log
AUC
0 2 4 6
0
5
10
15
Study weeks
Log
AUC
0 2 4 6
0
5
10
15
Study weeks
Log
AUC
0 2 4 6
0
5
10
15
Study weeks
Log
AUC
E

Figure S3. Serum antibody binding and neutralization activity after CH848 10.17DT gp160 mRNA-LNP immunizations in DH270 UCA KI mice. Related to Figure 2.(A-D) ELISA logAUCs of vaccinated mouse serum IgG binding to (A) CH848 10.17DT DS SOSIP trimer; (B) CH848 10.17 Δ11 gp120; (C) CH848 V3 loop peptide; (D) gp41 subunit.(E) Predicted possible glycan holes on CH848 10.17 Env. PNGL: potential N-linked glycosylation site.(F) Week 5 serum antibody binding to modified mRNA-expressed CH848 10.17DT, CH848 10.17DT DS, and CH848 10.17DT 113C-429GCG gp160s on 293-F cell surface. Each dot signifies an individual mouse (N = 6 each group). Data shown are MFI measured by flow cytometry.(G) Serum neutralizing titers after CH848 10.17DT gp160 mRNA-LNPs immunization. Serum neutralization titers one week after the third immunization in DH270 UCA KI mice were measured using TZM-bl reporter cells. Data shown were ID50s. Murine leukemia virus (MuLV) was used as negative control.

CH848DT-sp.GC B cells
CH848DT-sp.IgG+ Non-GC B cells
A
GC TFH
TFH
Antigen-specific B cell gating
Tfh cell gatingB
Figure S4. Flow cytometric gating strategies for antigen-specific B cells and Tfh cell populations. Related to Figures 2 and 6.(A) The gating strategy to identify CH848 10.17DT-specific germinal center B cells and memory B cells.(B) The gating strategy to identify T follicular helper (Tfh) and germinal center Tfh (GC Tfh) cells.

Figure S5. NSEM analysis of modified mRNA-expressed CH848 10.17DT DS trimer-ferritin NPs. Related to Figure 5.(A) Representative raw image of modified mRNA-expressed, bnAb PGT145-purified CH848 10.17DT DS 3xGS linker trimer-ferritin NPs.(B) Green circles indicate particles automatically picked by the software. From 60 images, a total of 12,365 particles were picked. (C) 2D class averages. Labels indicate tentative identification of particle types for trimer-ferritin nanoparticles (NP), apparent Env trimers (E), galectin-3 binding proteins (G), unknown trimeric proteins (T), and proteosomes (P). (D) Table indicating the number of classes, the number of particles in those classes, and the percentage of the total particles for each identified type.(E) Representative raw image of modified mRNA-expressed, bnAb PGT145-purified CH848 10.17DT DS VRC trimer-ferritin NPs.(F) Green circles indicate particles automatically picked by the software. From 60 images, a total of 12,365 particles were picked. (G) 2D class averages. Labels indicate tentative identification of particle types for trimer-ferritin nanoparticles (NP), apparent Env trimers (E), galectin-3 binding proteins (G), unknown trimeric proteins (T), and proteosomes (P). (H) Table indicating the number of classes, the number of particles in those classes, and the percentage of the total particles for each identified type.
Worst-casescenario
Best-casescenario
Worst-casescenario
Best-casescenario

10.17DT DS trimer-ferritin NP10.17DT 113C-429GCG trimer-ferritin NP10.17DT DS 3xGS linker trimer-ferritin NP
10.17DT DS 2xGS linker trimer-ferritin NP10.17DT DS VRC trimer-ferritin NPEmpty LNP
gp41
0 1 2 3 4 50
5
10
15
Study weeks
Log
AUC
CH848 V3 peptide
0 1 2 3 4 50
5
10
15
Study weeks
Log
AUC
CH848 10.17DT Δ11 gp120
0 1 2 3 4 50
5
10
15
Study weeks
Log
AUC
Human ferritin
Study weeks
0 1 2 3 4 50
5
10
15
Log
AUC
Helicobacter pylori Ferritin
0 1 2 3 4 50
5
10
15
Study weeks
Log
AUC
CH848 10.17DT trimer
0 1 2 3 4 50
5
10
15
Study weeks
Log
AUC
A CB
D FE
mRNA-LNP immunogens:
MuLV CH848.10.17 CH848.10.17.N332T CH848.10.17.DT CH848.10.17.DT.N332TCH848.10.17.DT.D230N.H
289N.P291S92RW020.2 6101.10
Immuniza�on
<160 <160 nt 4344 <160 5301 <160 170<160 <160 nt 7134 <160 5542 <160 <160<160 <160 nt 6034 <160 6018 172 285<160 <160 nt 4682 <160 3480 <160 185<160 194.5 <160 7362 <160 7803 <160 <160<160 <160 nt 4187 <160 5321 <160 197<160 <160 nt 1966 <160 1629 <160 <160<160 532 <160 15206 <160 20932 <160 177<160 <160 nt 4071 <160 5360 <160 168<160 532 <160 4396 <160 6340 <160 172<160 <160 <160 1519 <160 1936 <160 318<160 <160 nt 12110 <160 14835 <160 186<160 <160 nt 4549 <160 6242 <160 192<160 <160 nt 12339 <160 23328 <160 190<160 <160 nt 6308 <160 7913 <160 <160<160 <160 nt 3590 <160 5466 <160 219<160 <160 nt 1501 <160 1187 <160 <160<160 324.5 <160 10127 <160 16398 <160 <160<160 <160 nt 5041 <160 6892 <160 <160<160 <160 nt 6573 <160 12384 <160 185<160 <160 nt 4749 <160 5580 <160 163<160 <160 nt 5448 <160 9133 <160 225<160 <160 nt 9814 <160 15247 <160 160<160 415.5 <160 13412 <160 24893 <160 360<160 325 <160 14835 <160 18650 <160 492<160 467 <160 20052 <160 36649 <160 193<160 <160 nt 3117 <160 7575 <160 172<160 <160 nt 4399 <160 8302 <160 261<160 <160 nt <160 <160 <160 <160 <160<160 <160 nt <160 <160 <160 <160 <160<160 <160 nt <160 <160 <160 <160 261<160 <160 nt <160 <160 <160 <160 432<160 <160 nt <160 <160 <160 <160 225<160 <160 nt <160 <160 <160 <160 <160
ID50nt
<160160-999
1000-9999>10000
CH848 10.17DT DS 2xGS linker LSK ferri�n NP
mRNA-LNP
CH848 10.17DT DS VRC ferri�n NP mRNA-LNP
Empty LNP
ID50 (dilu�on) in TZM-bl cells
CH848 10.17DT DS ferri�n NP
mRNA-LNP
CH848 10.17DT 113C-429GCG
ferri�n NP mRNA-LNP
CH848 10.17DT DS 3xGS linker LSK ferri�n NP
mRNA-LNP
G

Figure S6. Serum IgG binding activity after CH848 10.17DT trimer-ferritin NP mRNA-LNP immunizations in DH270 UCA dKI mice. Related to Figure 6.(A-F) ELISA logAUCs of vaccinated mouse serum IgG binding to (A) CH848 10.17DT DS SOSIP trimer; (B) CH848 10.17 Δ11 gp120; (C) CH848 V3 loop peptide; (D) gp41 subunit; (E) Helicobacter pylori ferritin, and (F) human ferritin.(G) Serum neutralizing titers after CH848 10.17DT trimer-ferritin NP mRNA-LNP immunizations. Serum neutralization titers one week after the third immunization in DH270 UCA KI mice were measured using TZM-bl reporter cells. Data shown were ID50s. Murine leukemia virus (MuLV) was used as negative control.

Total mutants Total mutants Total mutants48401 110 86 79 76 49 68 4548402 40 28 25 26 19 25 1848404 58 31 27 36 29 29 2348405 55 40 40 35 28 35 2848407 43 19 17 22 16 15 1348411 52 34 33 34 28 33 2948412 39 25 23 23 19 23 18
Total 397 263 244 252 188 228 174
VH1-2 # VL2-23 # VH1-2/VL2-23 #
1
2
Group Animal ID mAb #
A
B
Figure S7. Isolation of CH848 10.17DT-specific antibodies. Related to Figure 7.(A) Gating strategy used for single cell sort of CH848 10.17DT-specific memory B cells from vaccinated mice splenocytes. (B) Summary of single B cell sort and cloning of Ig genes by PCR. A total number of 617 B cells were sorted on 96-well plates. 397 pairs of heavy and light chains were successfully recovered by sequencing of PCR amplicon. Among these heavy and light chain pairs, 263 of heavy chains were DH270 VH1-2, 252 of light chains were DH170 VL2-23, and 228 pairs were both DH270 heavy chain VH1-2 and DH270 light chain VL2-23. 169 of 228 DH270-like antibodies have acquired at least one amino acid change. DH270.mu84 was isolated from animal 48402; DH270.mu85 was isolated from animal 48411; DH270.mu86 was isolated from animal 48412.
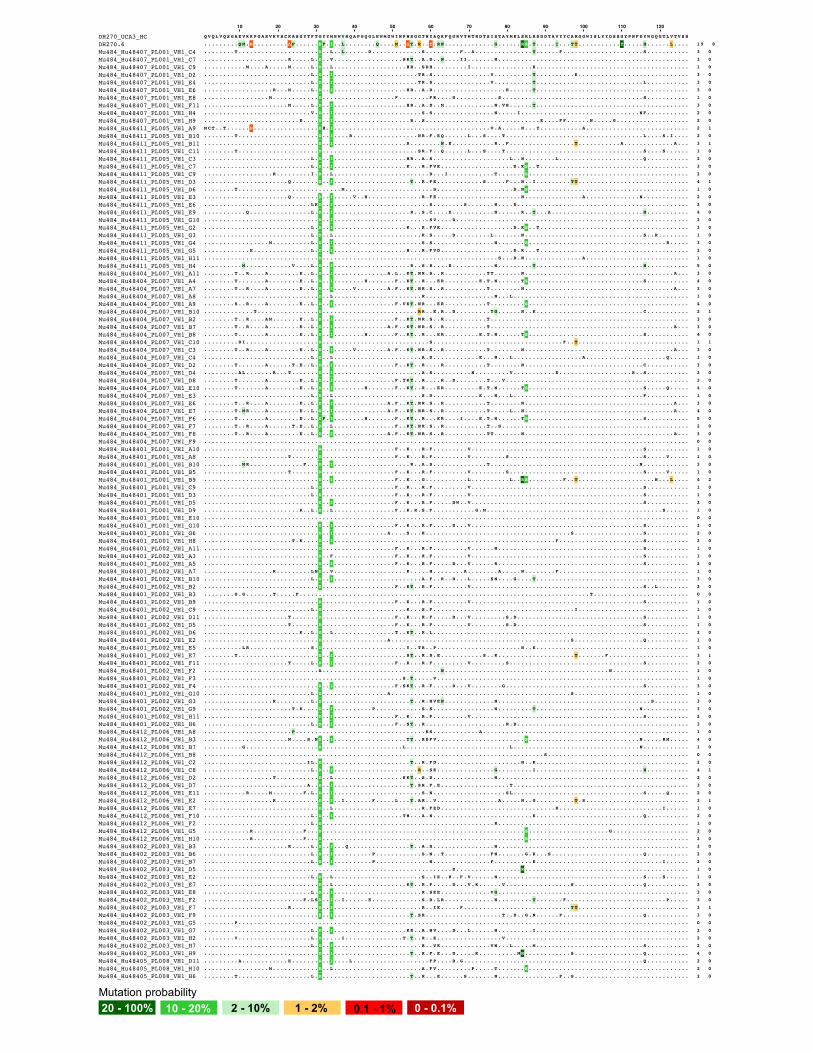
!"#$ %""&
Q V Q L V Q S G A E V K K P G A S V K V S C K A S G Y T F T G Y Y M H W V R Q A P G Q G L E W M G W I N P N S G G T N Y A Q K F Q G R V T M T R D T S I S T A Y M E L S R L R S D D T A V Y Y C A R G G W I S L Y Y D S S G Y P N F D Y W G Q G T L V T V S S
. . . . . . . . . Q M . N . . . . . . . . . A P . . . . . . D F . I . . L . . . . . . . . Q . . . . M . . Q T . R . . T . R N . . . . . . . . . . . . . G . . . . . . R S . T . . . . . I . . . T T . . . . . . . . . . . Y . . . . . H . . . . . . L . . . . 1 9 8
. . . . . . . . T . . . . . . . . . . . . . . . . . . . . . D . . L . . L . . . . . . D . . . . . . . . . . . . . S . . . . . . . . . F . . A . . . . . . . . . . . . . . . T . . . . . . F . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . R . . . . . L . D . . V . . . . . . . . . . . . . . . . . . S K T . . A . D . . N . . . . I I . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . M . . . . A . . . . . M . . . . . L . D . . L . . . . . . . . . . . . . . . . . . . H R . . S D H . . . . . . . . . I . . . . . . . . . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . T R . S . . . . . . . . . . . . . . . V . . . . . . . . . . T . . . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . T R . S . . . . . . . . . . . . . . . V . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . R . . . M . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . H R . . A . D . . . . . . . . . . . . . . . . . . . H . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . M . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . . . . . . . F E . . . . D . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . M . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . H R . . A . D . . M . . . . . . . . . . . . . N . V H . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . V . D . . I . . . . . . . . . . . . . . . . . . . . . . . S . S . . . . . . . . . . . . . . . . N . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N F . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . E . . . . D . . I . . . . . . . . . . . . . . . . . . . . R . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . E . . . . F F . . . . . . N . . . . . G . . . . . . . . . . . . . . . . . . . 2 0
M C T . . T . . . . . . N . . . . . . . . . . . . . . . . . D H . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V . A . . . . . N . . . Y . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 1
. . . . . . . . T . . . . . . . . . . . . . . . . . . . . . D . . I . . . . R . . . . . . . . . . . . . . . . . N R . F . E Q . . . . . . L . . . S . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . . S . I . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . S . . . . . . . . N . E . . . . . . . . . . . R . . F . . . . . . . . . . . . . . . . . T . . . . . . . . . . . A . . . . . . . . . . . . . A . . . 3 1
. . . . . . . . T . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . . . . . . S R . F . . Q . . . . . . L . . . S . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . S . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . H R . . A . S . . . . . . . . . . . . . . . . . . . . L . . N . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . K . . . R . F V E . . . . . . . . . . . . . . . . . . . D . K S . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . R . . . . . . . . . I . D . . L . . . . . . . . . . . . . . . . . . . . . . . . . D . . . I . . . . . . . . . . . . T . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . . T . . R . F E . . . . . . . . . . . . S . . . . . F . . . N . . I . . . . . . . . . T T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 1
. . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . M . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . D . N S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . D . . I . . . . . V . . H . . . . . . . . . . . . . . R . F E . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . N . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L K D . . I . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . S . . . . . . . N . . . . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . Q . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . R . . S . C . . . . E . . . . . . . . . . . N . . . . . . K . . T . . . A . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . 4 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . . . . . . . S V . . . . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . K . . . R . F V E . . . . . . . . . . . . . . . . . . . D . K S . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . L . . . . . . . . . . . . . . . . . . . . . . . K . S . . . . . D . . . . . . . . . L . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . R . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . M . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . . S . S . . . . . . . . . . . . . . . . N . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . R . . . . . 3 0
. . . . . . . . . . . . E . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . H . . . R . F V D . . . . . . . . . . . . . . . . . . . D . K . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . D . N . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . M . . . . . . . . . . . . V . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . R . . S . R . . . . E . . . . . . . . . . . N . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . 5 0
. . . . . . . . T . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . A . L . . K T . N R . S . . R . . . . . . . . . . . T T . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 3 0
. . . . . . . . T . . . . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . H . . . . . . . F . . K T . . R . . . E R . . . . . . . . . E . T . N . . . . . . T S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 4 0
. . . . . . . . T . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . V . . . . . . . . A . F . . K T . N R . S . . R . . . . . . . . . . . T . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . L . . . . . . . . . . . . . . . . . . . . . . . R . . . . . . . . . . . . . . . . . . N . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . S . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . . . F . V K T . N R . . . E R . . . . . . . . . . . T . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 0
. . . . . . . . . . . . . T . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . . . . . . R R . . E . R . . D . . . . . . . . . T G . . . . . . N . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . C . . . . . . . . . . . 2 1
. . . . . . . . T . . R . . . . A M . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . . . F . . K T . N R . S . . R . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . T . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . A . F . . K T . N R . S . . R . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 3 0
. . . . . . . . T . . . . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . H . . . . . . . F . . K T . . R . . . E R . . . . . . . . . E . T . N . . . . . . T S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 4 0
. . . . . . . . . D I . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . F . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 1
. . . . . . . . T . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . V . . . . . . . . A . F . . K T . N R . S . . R . . . . . . . . . . . T . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . L . . . . . . . . . . . . . . . . . . . . . . . A . D . . . . . . . . . . . . E . . . N . . . L . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . Q . . . . . 1 0
. . . . . . . . T . . . . . . . A . . . . . . T . E . . L . D . . I . . . . . . . . . . . . . . . . F . . K T . . R . . . . R . . . . . . . . . . . T . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C . . . . . . . . . . . 3 0
. . . . . . . . . A L . . . . . . . R . . . T . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . . . . . A . S . . . . . . . . . . N . . . . . . . . . V . . . . . . . . . . . E . . . . . . . . . . . . . . . . . . . S . . S . . . . . . . . . . . 2 0
. . . . . . . . T . . . . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . . . F . T K T . . R . . . . R . . D . . . . . . . . T . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . T . . . . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . H . . . . . . . F . . K T . . R . . . E R . . . . . . . . . E . T . N . . . . . . T S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . Q . . . . . 4 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . L . . . . . . . . . . . . . . . . . . . . . . . S . D . . . . . . . . . . . . E . . . N . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . F . . . . . . . . . . . 1 0
. . . . . . . . T . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . A . F . . K T . N R . S . . R . . . . . . . . . . . T . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 3 0
. . . . . . . . T . M R . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . A . F . . K T . N R . S . . R . . . . . . . . . . . T . . . . . L . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 4 0
. . . . . . . . T . . . . . . . A . . . . . . . . E . . L . D F . I . . . . . . . . H . . . . . . . F . . K T . . R . . . E R . . . . I . . . . E . T . N . . . . . . T S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 5 0
. . . . . . . . T . . R . . . . A . . . . . . T . E . . L . D . . L . . . . . . . . . . . . . . . . F . . K T . N R . S . . R . . . . . . . . . . . T . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . T . . R . . . . A . . . . . . . . E . . L . D . . I . . . . . . . . . . . . . . A . F . . K T . N R . S . . R . . . . . . . . . . . T T . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . T . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . V . . . . . 1 0
. . . . . . . . . . M R . . . . . . . . . . . . . . F . . . D . . I . . . . . . . . . . . . . . . . . . . . R . . A . S . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . T . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . V . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . F . . K . . . G . . . . . . . . . . . L . . . . . . . . . . L . . R S . . . . . . . . . F . . T . . . . . . . . . . . . . . . . . . . . H . . . L . . . . 4 2
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . D W . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . K . . L . D . . L . . . . . . . . . . . . . . . . F . . K . R . S . F . . . . . . . . . . . G . M . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . D . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . A . . . . S . . . R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . T . K . . . . D . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . F . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . F . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . D . . . V . . . . . . R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . R . . . . . . . . . L N D . . V . . . . . . . . . . . . . . . . . . . K . . . . . H . . . . . . . . A . . . . . . . . A . . . . . N . . . . . . . . F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . . A . F . . R . . D . . . L . . . . . S N . . . . G . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K T . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . L . . . . . . . . 2 0
. . . . . . . . G . G . . . . . . . T . . . . . F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . 0 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . . . . . . . . . . . . . . . . . . . . . K . . . S . F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . T . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . D . . . V . . . . . . . . . S . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . T . . . . . . . D . . . . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . S . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . K . . L . D . . L . . . . . . . . . . . . . . . . T . . K T . . R . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 1 0
. . . . . . . . . . L R . . . . . . . . . . . . . . . . S . D . . . . . . . . . . . . . . . . . . . . . . Y . . T R . . P . . . . . . . . . . . . . . . . . . . . . . N . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . T . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . S T . . R . S . E . . . . . . . . . . . S . . K . . . . . . . . . . . . . . . . . . . . T . . . . . . . F . . . . . . . . . . . . . . . . . . . . . 3 1
. . . . . . . . . . . . . . . . . . . . . . T . . . . . L . D . . I . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . T . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . F . S K T . . R . F . . . . . D . . . V . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . R . . . . . . . . . L . D . . . . . . . . . . . . . . . . . . . . . . . T . . R . H V E N . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . T . K . . . . D . . I . . . . . . . . . . P . . . . . . . . . . . . S . S . . . . . . . . . . . . . . . . N . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . F . . K . . . R . F . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . F . . S T . . R . . . . . . . . . . . . . . . . . . . . . N . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . P . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . K S . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . M . . . . S . N D . . I . . . . . . . . . . . . . . . . . . . T T . . R D F V . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . R M . . . . . 4 0
. . . . . . . . . . G . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . I L . D . . . . . . . . . . . . . . . . . . . . . . . T . . R . F D . . . . . . . . . . . . . . . . . . . . . . N . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . R . . S E . . . . . . . . . . . . . . . G . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . 4 1
. . . . . . . . . . . . . . . . . . T . . . . . . . . . . . D . . L . . . . . . . . . . . . . . . . . . S K T . . S . D . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . A . . D . . I . . . . . . . . . . . . . . . . . . . . T . S R . F . E . . . . . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . R . . . . . M . . . . . . . . F . L . D . . I . . . . . . . . . . . . . . . . . . . . . . . S . N . . . . . . . . . . . . . . . . . . . S L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . Q . . . . . 2 0
. . . . . . . . . . . . . . . . . . R . . . . . . . . . . . D . . I . . I . . . . . . . F . . . . . L . . . T . A R . . V . . . . . . . . . . . . . . . . A . . . . . N . . G . . . . . . . . . . T . S . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 1
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . L . . . . . . . . . . . . . . . . . . . . . . . R . F E D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . T H . . . A . N . . . . . . . . . . . . . . . . . . . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . R . . . . . . . . . . . . . F . . . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . R . . . . . . . . . . . . . F . . . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . R . . . . . L . D . . I . . . Q . . . . . . . . . . . . . . . . T . . A . S . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . P . . . . . . . . . . . . S . N . . T . . . . . . . . . . . . F N . . . . . . . G . K . . . S . . . . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . P . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . F . . . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . L . . . . . . . . . . . . . . . . . . . . . . . S . . I E . . R . . F . V . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . S . . . . . . 1 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . L . . . . . . . . . . . . . . . . . . . K T . . R . F . . . . . D . . . V . K . . . . . . V . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . . R . S E E . . . . . . . . . . . . . V G . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . . . . . F . L S D . . I . . I . . . . . . E . . . . . . . . . . . . . S . D . L R . . . . . . . . . . . . . N . . . . . . . . . T . . . . . . . F . . . . . . . . . . . . . . . . . . . . . . . . . . P . . . . . 3 0
. . . . . . . . . . . . . . . . . . . . . . R . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . . . . . R . . I E . . . . . F . . . . . . . . . . . . . . . . . . . . . . . . . . . . T T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 1
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . . T . S R . . . . . . . . . . . . . . . . . . . . T . . D . . G . N . . . . . . F . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 3 0
. . . . . . . . P . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . K R . . A . N V . . . . D . . . L . . . . . . N . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . V . . . . . . . . . . . . . . . . . . . L . D . . . . . I . . . . . . . . . . . . . . . T . T . . R . . E . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . D . . I . . . . . . . . . . . . . . . . . . . . . . . R . . V E . . . . . . . . . . . . . V N . . . L . . . . . M . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . I . . . . . . . . . . . . . . . . . . . . T . . R . F . E . . . D . . . . . K . . . . . . . . . . M R . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 4 0
. . . . . . . . . A . . . . . . . . . . . . E . . . . . . . D . . I . . . . L . . . . . . . . . . . . . . . . . . . . F P . . . . D . G . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . 2 0
. . . . . . . . . . . . . . . . . M . . . . . . . . . . . . D . . L . . . . . . . . . . . . . . . . . . . . . . . A . F V . . . . . . . . . P . . . . . T . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
. . . . . . . . T . . . . . . . . . . . . . . . . . . . L . D . . . . . . . . . . . . . . . . . . . . . . . T . . R . . . E . . . . . . S . . . . . . . N . . . . . . . . . . . . . . . . F . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0
10 20 30 40 50 60 70 80 90 100 110 120
Mu484_Hu48405_PL008_VH1_H6 Mu484_Hu48405_PL008_VH1_H10Mu484_Hu48405_PL008_VH1_D11Mu484_Hu48402_PL003_VH1_H9 Mu484_Hu48402_PL003_VH1_H7 Mu484_Hu48402_PL003_VH1_H2 Mu484_Hu48402_PL003_VH1_G7 Mu484_Hu48402_PL003_VH1_G5 Mu484_Hu48402_PL003_VH1_F9 Mu484_Hu48402_PL003_VH1_F7 Mu484_Hu48402_PL003_VH1_F2 Mu484_Hu48402_PL003_VH1_E8 Mu484_Hu48402_PL003_VH1_E7 Mu484_Hu48402_PL003_VH1_E2 Mu484_Hu48402_PL003_VH1_D5 Mu484_Hu48402_PL003_VH1_B7 Mu484_Hu48402_PL003_VH1_B6 Mu484_Hu48402_PL003_VH1_B3 Mu484_Hu48412_PL006_VH1_H10Mu484_Hu48412_PL006_VH1_G5 Mu484_Hu48412_PL006_VH1_F2 Mu484_Hu48412_PL006_VH1_F10Mu484_Hu48412_PL006_VH1_E7 Mu484_Hu48412_PL006_VH1_E2 Mu484_Hu48412_PL006_VH1_E11Mu484_Hu48412_PL006_VH1_D7 Mu484_Hu48412_PL006_VH1_D2 Mu484_Hu48412_PL006_VH1_C8 Mu484_Hu48412_PL006_VH1_C2 Mu484_Hu48412_PL006_VH1_B8 Mu484_Hu48412_PL006_VH1_B7 Mu484_Hu48412_PL006_VH1_B3 Mu484_Hu48412_PL006_VH1_A8 Mu484_Hu48401_PL002_VH1_H6 Mu484_Hu48401_PL002_VH1_H11Mu484_Hu48401_PL002_VH1_G9 Mu484_Hu48401_PL002_VH1_G3 Mu484_Hu48401_PL002_VH1_G10Mu484_Hu48401_PL002_VH1_F4 Mu484_Hu48401_PL002_VH1_F3 Mu484_Hu48401_PL002_VH1_F2 Mu484_Hu48401_PL002_VH1_F11Mu484_Hu48401_PL002_VH1_E7 Mu484_Hu48401_PL002_VH1_E5 Mu484_Hu48401_PL002_VH1_E2 Mu484_Hu48401_PL002_VH1_D6 Mu484_Hu48401_PL002_VH1_D5 Mu484_Hu48401_PL002_VH1_D11Mu484_Hu48401_PL002_VH1_C9 Mu484_Hu48401_PL002_VH1_B9 Mu484_Hu48401_PL002_VH1_B3 Mu484_Hu48401_PL002_VH1_B2 Mu484_Hu48401_PL002_VH1_B10Mu484_Hu48401_PL002_VH1_A7 Mu484_Hu48401_PL002_VH1_A5 Mu484_Hu48401_PL002_VH1_A3 Mu484_Hu48401_PL002_VH1_A11Mu484_Hu48401_PL001_VH1_H8 Mu484_Hu48401_PL001_VH1_G6 Mu484_Hu48401_PL001_VH1_G10Mu484_Hu48401_PL001_VH1_E10Mu484_Hu48401_PL001_VH1_D9 Mu484_Hu48401_PL001_VH1_D5 Mu484_Hu48401_PL001_VH1_D3 Mu484_Hu48401_PL001_VH1_C9 Mu484_Hu48401_PL001_VH1_B9 Mu484_Hu48401_PL001_VH1_B5 Mu484_Hu48401_PL001_VH1_B10Mu484_Hu48401_PL001_VH1_A8 Mu484_Hu48401_PL001_VH1_A10Mu484_Hu48404_PL007_VH1_F9 Mu484_Hu48404_PL007_VH1_F8 Mu484_Hu48404_PL007_VH1_F7 Mu484_Hu48404_PL007_VH1_F6 Mu484_Hu48404_PL007_VH1_E7 Mu484_Hu48404_PL007_VH1_E6 Mu484_Hu48404_PL007_VH1_E3 Mu484_Hu48404_PL007_VH1_E10Mu484_Hu48404_PL007_VH1_D8 Mu484_Hu48404_PL007_VH1_D4 Mu484_Hu48404_PL007_VH1_D2 Mu484_Hu48404_PL007_VH1_C4 Mu484_Hu48404_PL007_VH1_C3 Mu484_Hu48404_PL007_VH1_C10Mu484_Hu48404_PL007_VH1_B8 Mu484_Hu48404_PL007_VH1_B7 Mu484_Hu48404_PL007_VH1_B2 Mu484_Hu48404_PL007_VH1_B10Mu484_Hu48404_PL007_VH1_A9 Mu484_Hu48404_PL007_VH1_A8 Mu484_Hu48404_PL007_VH1_A7 Mu484_Hu48404_PL007_VH1_A4 Mu484_Hu48404_PL007_VH1_A11Mu484_Hu48411_PL005_VH1_H4 Mu484_Hu48411_PL005_VH1_H11Mu484_Hu48411_PL005_VH1_G5 Mu484_Hu48411_PL005_VH1_G4 Mu484_Hu48411_PL005_VH1_G3 Mu484_Hu48411_PL005_VH1_G2 Mu484_Hu48411_PL005_VH1_G10Mu484_Hu48411_PL005_VH1_E9 Mu484_Hu48411_PL005_VH1_E6 Mu484_Hu48411_PL005_VH1_E3 Mu484_Hu48411_PL005_VH1_D6 Mu484_Hu48411_PL005_VH1_D3 Mu484_Hu48411_PL005_VH1_C9 Mu484_Hu48411_PL005_VH1_C7 Mu484_Hu48411_PL005_VH1_C3 Mu484_Hu48411_PL005_VH1_C11Mu484_Hu48411_PL005_VH1_B11Mu484_Hu48411_PL005_VH1_B10Mu484_Hu48411_PL005_VH1_A9 Mu484_Hu48407_PL001_VH1_H9 Mu484_Hu48407_PL001_VH1_H4 Mu484_Hu48407_PL001_VH1_F11Mu484_Hu48407_PL001_VH1_E8 Mu484_Hu48407_PL001_VH1_E6 Mu484_Hu48407_PL001_VH1_E4 Mu484_Hu48407_PL001_VH1_D2 Mu484_Hu48407_PL001_VH1_C9 Mu484_Hu48407_PL001_VH1_C7 Mu484_Hu48407_PL001_VH1_C4 DH270.6 DH270_UCA3_HC
%"#$ !"& !#$ %"& %#$ !& "'%#$ %& "#$ "'%&!"#$#%&'()*&+$+%,%#-

Figure S8. CH848 10.17DT trimer-ferritin NP mRNA-LNP immunization elicited antibodies with heavy chain somatic mutations shared with DH270.6. Related to Figure 7.Heavy chain amino acids of isolated VH1-2/VL2-23 DH270-like mAbs aligned with DH270 UCA and DH270.6. Dots represent amino acids that are in common with DH270 UCA. Mutations shared with DH270.6 are highlighted in colors according to mutation probability. The numbers to the right of each row are total numbers of probable mutations and improbable mutations observed, respectively.
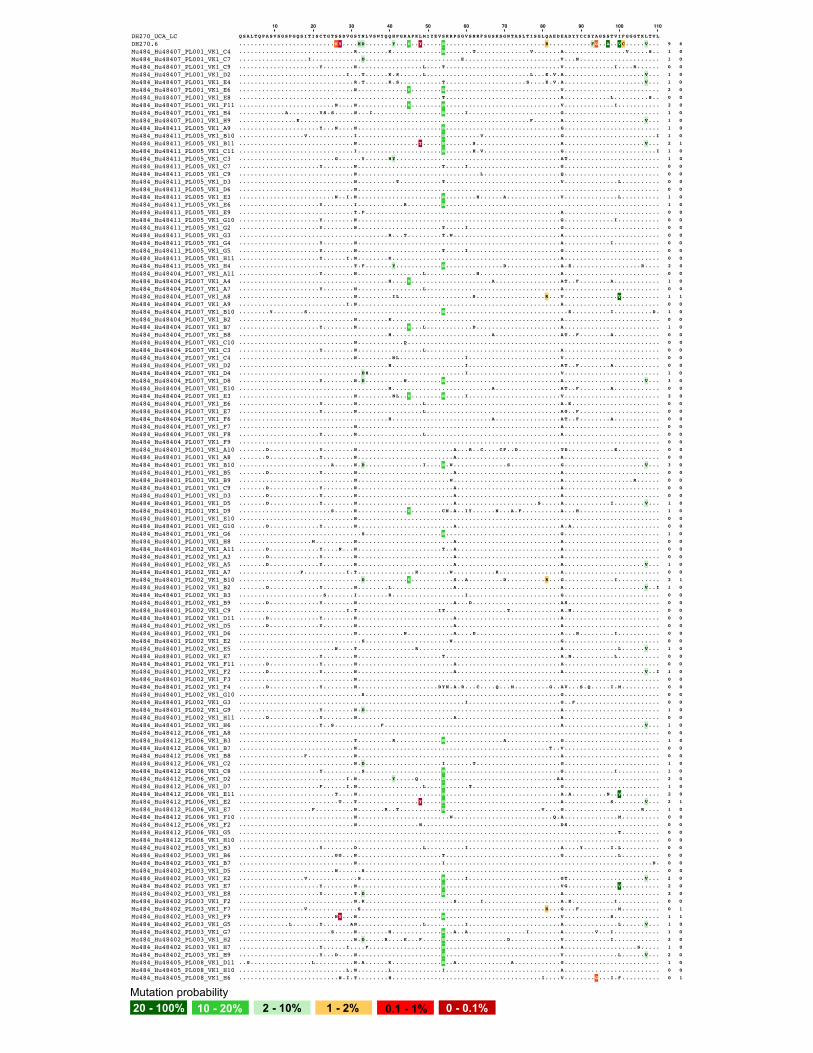
Q S A L T Q P A S V S G S P G Q S I T I S C T G T S S D V G S Y N L V S W Y Q Q H P G K A P K L M I Y E V S K R P S G V S N R F S G S K S G N T A S L T I S G L Q A E D E A D Y Y C C S Y A G S S T V I F G G G T K L T V L . . . . . . . . . . . . . . . . . . . . . . . . . K Y . . . . H D . . . . . . . Y . . . V . . Y . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . R . . . . . . . . . . . F G . . A . . V C . . . . . V . . . 9 6. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . R . . . . . . . . K . . . . . . . . . . . . . N . . . . . . . T . . . . . . . . . . . . . . V . . . . . . . A . . . . . . . . . . . . . . . . V . . . . . S . . 1 0. . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . .. . . . E . . . . . . . . . . . . . . . . . . . . . . . . . V . . . N . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . L . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . I . . . . R . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . T . . . . . . K . S . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . E . V . A . . . . . . . . . . . . . . . . . . . . . V . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . R . T . . . . . . K . S . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . S . . . . E . V . A . . . . . . . . . . . . . . . . . . . . .V . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . V . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . 2 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . L . . . . . . . . . S . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . N . . . . N . . . . . . . . . . . . . V . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . I . . . . . . . . . . . 2 0. . . . . . . . . . . . A . . . . . . . . Y S . S . . . . . N . . . I . . . . . . .. . . . . . . . . . . N . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . E . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . F . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . N . . . . N . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . V . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . G . . . . . . . . .. . . . . . . . . . . . . . . I 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . Y . . . . . N . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . . 2 1. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . H . V . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . I 1 0. . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . Y . . . . . . H Y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A T . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . .N . . . . . . . . . . . . . . . . . . . . . . T . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . Q . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . T . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . . L . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . N . . I . N . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . H . . . . . . A . . . . . . . . . . . . . . V . . . . . . . . . . . . . . L . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . I . . . . . . . . . . . . R . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . .. . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . I . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . T . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . R . . . T . . . . . . . . . T . W . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . A . . . . . . . . . . . . I . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . T . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . I . N . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . F . . . . . . . Y . . . . . . . . . . . . N . . . . . . . . . . . . . . . D . . . . . . . . . . . . . . A . E . . . . . . . . . . . . . . . . . . R . . . . 2 0. . . . . .. . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . H . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . V . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . A T . . F . . . . . . . . A . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . I L . . . . . . . . . . . . . . . .. . . S . . . . . . . . . . . . . . . . . . R . . . V . . . . . . . . . . . . . . V . . . . . . . . . . 1 1. . . . . . . . . . . . . . . . . . . . . . . . . . . . I . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . Y . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . E . . . . . . . . . . I . . . . . . . . . . D . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . K . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . V . . . L . . . . . . . . . . . . D . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . A T . . F . . . . . . . . A . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . Q . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . .. . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . N L . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . A T . . F . . . . . . . . A . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D H . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . .. . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . D . . . . . . . . . . N . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . . 3 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . A T . . F . . . . . . . . A . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . N L . . V . . . . . . . . N . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . . . . . . . . . . . . . 2 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . .. . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . E . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A G . . F . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . H . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . A T . . F . . . . . . . . A . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A .. . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . R . . C . . . . C F . . D . . . . . . . . . . . T D . . . . . . . . . . . . E . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y. . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . A . . . . . N . D . . . . . . . . . . . . . . . I . . . . N . W . . . . . . . . . . . . . . S . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . V . . . 3 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . W . . . . . . . . . . . . . . . . . .. . . . . . . . . . A . . . . . . . . . . . . . . . . . . R . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . S . . . . . A . . . . . . . . . . . . I . . . . . . . . V . . . 1 0. . . . . . . . . .. . . . . . . . . . . . . . S . . . . . N . . . . . . . . . . . . . V . . . . . . . . C N . A . . I Y . . . . . . N . . . A . F . . . . . . . . . . A . . . H . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . A . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . N . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . M . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . N . . . N . . . . . . . . . . . . . . . . . . . . . . T . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . . 1 0. . . . . . . . . . . . . . . . P . . . . . . . . . . . I . T . . . . . . . . . . . . . . . R . . . . . . . . W . . . . . . . . . . . R . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . . . . . . . . . . . V . . . . . . . . . . . S . . A . . . . . . . . . D . . . . . . . . . . R . . . G . . . . . . . . . . . . . I . . . . . . . . . . . 2 1. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . L . . . . . . . . . .. . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . I 1 0. . . . . . . . . . . . . . . . . . . . . . S . . . . . . . I . . . . . . . . R . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . D . . . . . . . . . . . . . . . . . . . . . . . A S . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . I . T . . . . . . . . . . . . . . . . . . . . . I T . . . . . . . . . . . . . . . . T . . . . . . . . . . . . . A . N . . . . . . . . . . . . . . .. . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . N . . . . . . . . . . . . A . . . . D . . . . . . . . . . . . . . . . . . . . . . A . . . H . . . . . . . . . I . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . S . . . . .. . . . . . . . . . . . . . . . . W . . . . . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . N . . . . T . . . . . . . . . . . . . . . R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . L . . . . . . V . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . N . . . . . . . . . . . L . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . .. . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . I 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . D Y N . A . R . . . C . . . . Q . . . M . . . . . . . . . G . . A V . . . S . Q . . . . . I . M . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G . . F . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . D . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . .. . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . S . . . . . . . . . . . . F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . V . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . . . . . . . . R . . . . . . . . . . . . N . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . V . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . F . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . D . . . . . . . . . . . . . . . . . . . . I . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . I . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . I . N . . . . . . . . . Y . . . . . Q . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A A . . . . . . . . . . . . . . . . . . . . . . . . . 2 0. . . . . . . . . . . . . . . . . . . . . F . . . . . . I . N . . . . . . . . . . . . . . . . . L . . . . N . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . T . . . . N . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . A . . . . . . . . . N . . V . . . . . . . . . . 2 0. .. . . . . . . . . . . . . . . . . . . . . . . . T . . . T . . . . . . . . . . . . . . . . Y . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . S . . . . . . . . V . . . 2 1. . . . . . . . . . . . . . . . . . . F . . . . . . . . . . N . . . . . . . R . . T . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . V . . . . G . . . . . . . . . . . . . . . . . . . . R . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . W . . . . . . . . . . . . . . . . . . . . . . . . . . Q . A . . . . . . . . . . . . . . M . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . N . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D S . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . D . . . . . . . . . . . . . . . . . L . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . A . . . . Y . . . . . . . I . L . . . . . .. . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . G G . . . N . . . . . . . . . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . L . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . N . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . S . . . . . . . . . .. . . . . . . . . . . N . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . G T . . . . . . . . . . . . . . . . . . . . V . . . 2 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V G . . . . . . . . . . . . . V . . . . . . . . . . 2 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . . . T . D . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 2 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . K . . . . . . . . . . . . . . . . . . . . . . . S . . . . . . I . . . . . . . . . . . . . . . . . . . . A . E . . . . . . .. . . . I . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . V . . . . . . . . . . . . . S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . R . . . G . . . F . . . . . . . . . . M . . . . . . . . . . 0 1. . . . . . . . . . . . . . . . . . . . . . . . . N Y . . . N . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V . . . . . . . . . . . . S . . . . . . . . . . . . 1 1. . . . . . . . . . . . . L . . . . . . . Y . . . . . . . A N . . . . . . . . . . . . . . . . . L . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . L . . . . . . V . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . S . . . . .N . . . . . . . . H . . . . . . . . . . . . . N . . A . . A . . . . . . . . . . . . . . . I . . . . . . . . A . . . . . . . . V . . . I . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N . D . . . . . R . . . . E . . . F . . . . . N . . . . . . . . . . . . . . . . D . . . . . . . . . . . . . V . . . . . . . . . . . . I . . . . . . . . . . . . 2 0. . . . . . . . . . . . . . . . . . . . . Y . . . . . . I . . . . F . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . S . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . Y . . . D . . . . N . . . . . . . . . . . . . . . . . . . . . . N . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . V . . . . . . . . . . . . . . L . . . . . . V . . . 2 0. . G . . . . . . . . . . . . . . . . L . . . . . . . . . . N . A . . . . . . K . . . . . . . . . . . . . N . . A . . . . . . . . . . . . . . A . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . 1 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . L . N . . . . . . . . L . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . 0 0. . . . . . . . . . . . . . . . . . . . . . . . . . N . I . T . . . . . . . . H . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . V . . . . . . . . G . . . I . F . . . . . . . . . . 0 1
10 20 30 40 50 60 70 80 90 100 110
Mu484_Hu48405_PL008_VK1_H6 Mu484_Hu48405_PL008_VK1_H10Mu484_Hu48405_PL008_VK1_D11Mu484_Hu48402_PL003_VK1_H9 Mu484_Hu48402_PL003_VK1_H7 Mu484_Hu48402_PL003_VK1_H2 Mu484_Hu48402_PL003_VK1_G7 Mu484_Hu48402_PL003_VK1_G5 Mu484_Hu48402_PL003_VK1_F9 Mu484_Hu48402_PL003_VK1_F7 Mu484_Hu48402_PL003_VK1_F2 Mu484_Hu48402_PL003_VK1_E8 Mu484_Hu48402_PL003_VK1_E7 Mu484_Hu48402_PL003_VK1_E2 Mu484_Hu48402_PL003_VK1_D5 Mu484_Hu48402_PL003_VK1_B7 Mu484_Hu48402_PL003_VK1_B6 Mu484_Hu48402_PL003_VK1_B3 Mu484_Hu48412_PL006_VK1_H10Mu484_Hu48412_PL006_VK1_G5 Mu484_Hu48412_PL006_VK1_F2 Mu484_Hu48412_PL006_VK1_F10Mu484_Hu48412_PL006_VK1_E7 Mu484_Hu48412_PL006_VK1_E2 Mu484_Hu48412_PL006_VK1_E11Mu484_Hu48412_PL006_VK1_D7 Mu484_Hu48412_PL006_VK1_D2 Mu484_Hu48412_PL006_VK1_C8 Mu484_Hu48412_PL006_VK1_C2 Mu484_Hu48412_PL006_VK1_B8 Mu484_Hu48412_PL006_VK1_B7 Mu484_Hu48412_PL006_VK1_B3 Mu484_Hu48412_PL006_VK1_A8 Mu484_Hu48401_PL002_VK1_H6 Mu484_Hu48401_PL002_VK1_H11Mu484_Hu48401_PL002_VK1_G9 Mu484_Hu48401_PL002_VK1_G3 Mu484_Hu48401_PL002_VK1_G10Mu484_Hu48401_PL002_VK1_F4 Mu484_Hu48401_PL002_VK1_F3 Mu484_Hu48401_PL002_VK1_F2 Mu484_Hu48401_PL002_VK1_F11Mu484_Hu48401_PL002_VK1_E7 Mu484_Hu48401_PL002_VK1_E5 Mu484_Hu48401_PL002_VK1_E2 Mu484_Hu48401_PL002_VK1_D6 Mu484_Hu48401_PL002_VK1_D5 Mu484_Hu48401_PL002_VK1_D11Mu484_Hu48401_PL002_VK1_C9 Mu484_Hu48401_PL002_VK1_B9 Mu484_Hu48401_PL002_VK1_B3 Mu484_Hu48401_PL002_VK1_B2 Mu484_Hu48401_PL002_VK1_B10Mu484_Hu48401_PL002_VK1_A7 Mu484_Hu48401_PL002_VK1_A5 Mu484_Hu48401_PL002_VK1_A3 Mu484_Hu48401_PL002_VK1_A11Mu484_Hu48401_PL001_VK1_H8 Mu484_Hu48401_PL001_VK1_G6 Mu484_Hu48401_PL001_VK1_G10Mu484_Hu48401_PL001_VK1_E10Mu484_Hu48401_PL001_VK1_D9 Mu484_Hu48401_PL001_VK1_D5 Mu484_Hu48401_PL001_VK1_D3 Mu484_Hu48401_PL001_VK1_C9 Mu484_Hu48401_PL001_VK1_B9 Mu484_Hu48401_PL001_VK1_B5 Mu484_Hu48401_PL001_VK1_B10Mu484_Hu48401_PL001_VK1_A8 Mu484_Hu48401_PL001_VK1_A10Mu484_Hu48404_PL007_VK1_F9 Mu484_Hu48404_PL007_VK1_F8 Mu484_Hu48404_PL007_VK1_F7 Mu484_Hu48404_PL007_VK1_F6 Mu484_Hu48404_PL007_VK1_E7 Mu484_Hu48404_PL007_VK1_E6 Mu484_Hu48404_PL007_VK1_E3 Mu484_Hu48404_PL007_VK1_E10Mu484_Hu48404_PL007_VK1_D8 Mu484_Hu48404_PL007_VK1_D4 Mu484_Hu48404_PL007_VK1_D2 Mu484_Hu48404_PL007_VK1_C4 Mu484_Hu48404_PL007_VK1_C3 Mu484_Hu48404_PL007_VK1_C10Mu484_Hu48404_PL007_VK1_B8 Mu484_Hu48404_PL007_VK1_B7 Mu484_Hu48404_PL007_VK1_B2 Mu484_Hu48404_PL007_VK1_B10Mu484_Hu48404_PL007_VK1_A9 Mu484_Hu48404_PL007_VK1_A8 Mu484_Hu48404_PL007_VK1_A7 Mu484_Hu48404_PL007_VK1_A4 Mu484_Hu48404_PL007_VK1_A11Mu484_Hu48411_PL005_VK1_H4 Mu484_Hu48411_PL005_VK1_H11Mu484_Hu48411_PL005_VK1_G5 Mu484_Hu48411_PL005_VK1_G4 Mu484_Hu48411_PL005_VK1_G3 Mu484_Hu48411_PL005_VK1_G2 Mu484_Hu48411_PL005_VK1_G10Mu484_Hu48411_PL005_VK1_E9 Mu484_Hu48411_PL005_VK1_E6 Mu484_Hu48411_PL005_VK1_E3 Mu484_Hu48411_PL005_VK1_D6 Mu484_Hu48411_PL005_VK1_D3 Mu484_Hu48411_PL005_VK1_C9 Mu484_Hu48411_PL005_VK1_C7 Mu484_Hu48411_PL005_VK1_C3 Mu484_Hu48411_PL005_VK1_C11Mu484_Hu48411_PL005_VK1_B11Mu484_Hu48411_PL005_VK1_B10Mu484_Hu48411_PL005_VK1_A9 Mu484_Hu48407_PL001_VK1_H9 Mu484_Hu48407_PL001_VK1_H4 Mu484_Hu48407_PL001_VK1_F11Mu484_Hu48407_PL001_VK1_E8 Mu484_Hu48407_PL001_VK1_E6 Mu484_Hu48407_PL001_VK1_E4 Mu484_Hu48407_PL001_VK1_D2 Mu484_Hu48407_PL001_VK1_C9 Mu484_Hu48407_PL001_VK1_C7 Mu484_Hu48407_PL001_VK1_C4 DH270.6 DH270_UCA_LC
!"#$ %""& %"#$ !"& !#$ %"& %#$ !& "'%#$ %& "#$ "'%&!"#$#%&'()*&+$+%,%#-

Figure S9. CH848 10.17DT trimer-ferritin NP mRNA-LNP immunization elicited antibodies with light chain somatic mutations shared with DH270.6. Related to Figure 7.Light chain amino acids of isolated VH1-2/VL2-23 DH270-like mAbs aligned with DH270 UCA and DH270.6. Dots represent amino acids that are in common with DH270 UCA. Mutations shared with DH270.6 are highlighted in colors according to mutation probability. The numbers to the right of each row are total number of probable mutations and improbable mutations, respectively.

mRNA-LNP gp
160s
mRNA-LNP N
Ps10-1
100
101
102
G57R
Mut
atio
n fre
quen
cy %
mRNA-LNP gp
160s
mRNA-LNP N
Ps10-1
100
101
102
R98T
Mut
atio
n fre
quen
cy %
mRNA-LNP gp
160s
mRNA-LNP N
Ps103
104
105
CH848 10.17DT
IC50
(1/d
ilutio
n) in
TZM
-bl c
ells
CH848 10.17DTD230N.H289N.P291S
mRNA-LNP gp
160s
mRNA-LNP N
Ps103
104
105
IC50
(1/d
ilutio
n) in
TZM
-bl c
ells
Figure S10. Comparing mRNA-LNP-encoded CH848 10.17DT gp160s and mRNA-LNP-encoded SOSIP trimer-ferritin NPs. Related to Figures 2 and 6.(A) Comparison of mutation frequencies of DH270 heavy chain improbable G57R and R98T mutations between CH848 10.17DT transmembrane gp160 (blue circle) and CH848 10.17DT SOSIP trimer-ferritin NP (red square) mRNA-LNP vaccinations in DH270 UCA KI mice. (B) Comparison of serum neutralization titers of autologous tier 2 CH848 10.17DT HIV-1 strains between CH848 10.17DT transmembrane gp160 (blue circles) and CH848 10.17DT SOSIP trimer-ferritin NP (red squares) mRNA-LNP vaccinations in DH270 UCA KI mice. Bar, geometric mean.Exact Wilcoxon Mann-Whitney U test.
A
B
p = 0.3678 p = 0.1096
p = 0.0843 p = 0.5899




















