
The role of fish in the regulation of nutrient cycling in Lake Balaton, Hungary
9
Freshwater Biotogy {I9ii6) 16,411-414 The role offish in the regulation of nutrient cycling in Lake Balaton, Hungary ISTVAN TATRAI and VERA ISTVANOVICS Balaton Limnological Research Institute, Hungary SUMMARY. 1. In 1984 an enclosure experiment was carried out in order to elucidate the importance of nutrient supply from fish to phytoplankton during the critical summer period in eutrophic Lake Balaton. One enclosure was stocked with fish and the other left empty. 2. Two peaks were recorded in primary production reaching maxi- mum values in the fish enclosure. 3. In the enclosure with fish, inorganic nitrogen concentrations increased gradually till the end ofthe experiment, with ammonia making up the largest component. Lower dissolved phosphorus concentrations in the fish enclosure, with its higher primary production rate, may indicate an intense cycling of this nutrient. 4. The biological and chemical changes occurring in the enclosure suggest that bream removal can be an effective tool in water quality control for Lake Balaton. Introduction It has become apparent that selective preda- tion by planktivorous and bcnthivorous fish affects not only the species composition and biomass of their prey populations, but also influences the primary producers by modifying populations of filter feeding cladocerans (Andersson ('/(;/., 1^78; Henrikson cr«/.. 1980; Shapiro & Wright, 1984). Moreover, benth- ivorous fish may facilitate regeneration of nutrients from sediments by mechanical mixing processes (Keen & Gagliardi. 1981). They may also, by excretion and egestion. release nut- rients, primarily ammonia and orthophos- phate. available for phytoplankton (Brabrand. Faafeng & Nilssen. 1985). In recent years the importance of fish in the nutrient cycling of lakes has been emphasued. Correspondence: Dr I. Tatrai. Balaton Limnolo- gical Research InMitute, H-8237 Tihany. Hungary. Lamarra (1975) has reported that in the pre- sence of bottom feeding carp. Cyprintts carpio L.. phosphorus concentration increased in lake water leading to enhanced primary production. Faafeng & Nillsen (1981) found that even a low biomass of roach. Rutilus rutitus (L.). increased markedly the concentration of chlorophyll-rt. On the other hand. Kitchell, Koonze & Tennis (1975) and Nakashima & Leggett (1980) came to the conclusion that nutrient excretion by fish, primarily by percids, was not of decisive importance to the phyto- plankton. In our preliminary short-term enclo- sure experiment, bream, Abramis brama (L.). not only increased primary production but also caused changes in the phytoplankton composi- tion of Lake Balaton (Tatrai. Toth & Ponyi, 1984). In addition, fish decreased the abund- ance and size of zooplankton. Bream is the most abundant fish species in Lake Balaton. The biomass of bream of 3+ to 417
Transcript of The role of fish in the regulation of nutrient cycling in Lake Balaton, Hungary

Freshwater Biotogy {I9ii6) 16,411-414
The role offish in the regulation of nutrient cyclingin Lake Balaton, Hungary
ISTVAN TATRAI and VERA ISTVANOVICS Balaton Limnological ResearchInstitute, Hungary
SUMMARY. 1. In 1984 an enclosure experiment was carried out in
order to elucidate the importance of nutrient supply from fish to
phytoplankton during the critical summer period in eutrophic Lake
Balaton. One enclosure was stocked with fish and the other left empty.
2. Two peaks were recorded in primary production reaching maxi-
mum values in the fish enclosure.
3. In the enclosure with fish, inorganic nitrogen concentrations
increased gradually till the end ofthe experiment, with ammonia making
up the largest component. Lower dissolved phosphorus concentrations
in the fish enclosure, with its higher primary production rate, may
indicate an intense cycling of this nutrient.
4. The biological and chemical changes occurring in the enclosure
suggest that bream removal can be an effective tool in water quality
control for Lake Balaton.
Introduction
It has become apparent that selective preda-tion by planktivorous and bcnthivorous fishaffects not only the species composition andbiomass of their prey populations, but alsoinfluences the primary producers by modifyingpopulations of filter feeding cladocerans(Andersson ('/(;/., 1^78; Henrikson cr«/.. 1980;Shapiro & Wright, 1984). Moreover, benth-ivorous fish may facilitate regeneration ofnutrients from sediments by mechanical mixingprocesses (Keen & Gagliardi. 1981). They mayalso, by excretion and egestion. release nut-rients, primarily ammonia and orthophos-phate. available for phytoplankton (Brabrand.Faafeng & Nilssen. 1985).
In recent years the importance of fish in thenutrient cycling of lakes has been emphasued.
Correspondence: Dr I. Tatrai. Balaton Limnolo-gical Research InMitute, H-8237 Tihany. Hungary.
Lamarra (1975) has reported that in the pre-sence of bottom feeding carp. Cyprintts carpioL.. phosphorus concentration increased in lakewater leading to enhanced primary production.Faafeng & Nillsen (1981) found that even alow biomass of roach. Rutilus rutitus (L.).increased markedly the concentration ofchlorophyll-rt. On the other hand. Kitchell,Koonze & Tennis (1975) and Nakashima &Leggett (1980) came to the conclusion thatnutrient excretion by fish, primarily by percids,was not of decisive importance to the phyto-plankton. In our preliminary short-term enclo-sure experiment, bream, Abramis brama (L.).not only increased primary production but alsocaused changes in the phytoplankton composi-tion of Lake Balaton (Tatrai. Toth & Ponyi,1984). In addition, fish decreased the abund-ance and size of zooplankton.
Bream is the most abundant fish species inLake Balaton. The biomass of bream of 3+ to
417
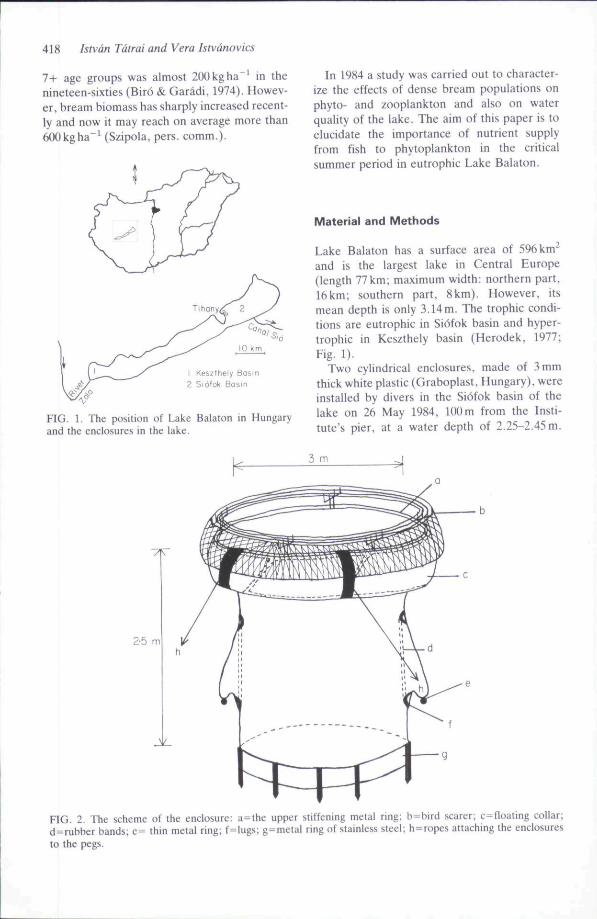
418 Istvdn Tatrai and Vera Istvdnovics
7+ age groups was almost 200 kg ha ' in thenineteen-sixties (Biro & Garadi, 1974). Howev-er, bream biomass has sharply increased recent-ly and now it may reach on average more than600kgha"' (Szipola, pers. comm.).
Keszthely2 Siofok Basin
FIG. 1. The position of Lake Balaton in Hungaryand the enclosures in the lake.
In 1984 a study was carried out to character-ize the effects of dense bream populations onphyto- and zooplankton and also on waterquality of the lake. The aim of this paper is toelucidate the importance of nutrient supplyfrom fish to phytoplankton in the criticalsummer period in eutrophic Lake Balaton.
Material and Methods
Lake Balaton has a surface area of 596 km^and is the largest lake in Central Europe(length 77km; maximum width: northern part,16km; southern part, 8km). However, itsmean depth is only 3.14m. The trophic condi-tions are eutrophic in Siofok basin and hyper-trophic in Keszthely basin (Herodek, 1977;Fig. 1).
Two cylindrical enclosures, made of 3 mmthick white plastic (Graboplast, Hungary), wereinstalled by divers in the Siofok basin of thelake on 26 May 1984, 100 m from the Insti-tute's pier, at a water depth of 2.25-2.45 m.
FIG. 2. The scheme of the enclosure: a^the upper stiffening melal ring; b=bird scaTer; c^floating collar;d=rubber bands; e= Ihin metal ring; f=lugs; g=metal ring of stainless steel; h = ropes attaching the enclosuresto the pegs.

The enclosures consisted of two sections. Theupper part consisted of five inflatable cylindric-al beach mattresses which were put into a sack.made of spun-rayon cloth, to form a circle(Fig. 2). The collars were fixed by ropes toconcrete metallic tubes driven into the sedi-ments. The bottom of the enclosure was made ofa stainless steel ring (15cm high and 1.5mmthick) to which plastic was folded by a metallicyoke. Eight spikes, also made of stainless steel,were screwed on to the metallic ring by whichenclosures were secured on the bottom. Birdswere prevented from perching on the top of theflotation collar by fishing line netting strungbetween iron projections. Unfortunately, in thefourth week of the experiment the bird scarer ofthe control enclosure was destroyed and itbecame heavily fertilized by duck's faeces. Thusthe results obtained in the control enclosureduring the last week of the experiment will notbe considered.
The enclosures. 3 m in diameter, were opento the sediment and to the atmosphere andcontained 16 m̂ of lake water. One enclosureserved as a control and was free of fish whilethe other was densely stocked with fifteenindividuals of bream of age groups 0-1- to 2-1-(average body weight 45.9±28.2g, range 7-105g) on the sixth day after installation. Sixfish died during the experiment, mostly duringthe first week. Biomass of dead fish was369.5 g. Dead fish were replaced by newlycaptured fish (biomass of 226.4g). The totalfish biomass amounted to 612 g, or 86 gm"'^.
During the experiment, lasting for 40 days,measurements and water samples for chemicalanalyses were taken at 1 ^ days intervals atthree stations: from the lake close to theenclosures, and from both enclosures. Watersamples were taken using a surface-mud tubesampler 12cm in diameter. Mixed, unfilteredwater samples were used for the determinationof total phosphorus (TP) and total nitrogen(TN) while total dissolved phosphorus (TDP)was determined from filtered samples afterdigestion with potassium persulphate accordingto standard analytical procedures (Mackereth,Heron & Tailing. 1978). Ammonia was deter-mined by reaction with hypochlorite and phe-nole to give indophenole using an autoanalyser(Contiflo, Labor MIM. Hungary). pH wasmeasured on an ammonia and pH meter(Radelkis, Hungary). The concentration of
Nutrient cycling by fish 419
dissolved oxygen was determined according toAhlgren & Ahlgren (1975) from samples takenat about 0.5 m depth. Secchi disc measure-ments were taken using a 25 cm black-and-white disc. Temperature and depth profiles oftemperature were measured with a thermistordigital thermometer (Balaton Limnological In-stitute, Hungary). Data on wind velocity wereobtained from the Meteorological Station atSiofok. Primary produetion was measured intriplicate samples at the temperature of thelake water with the ^'^C technique using Rack-Beta LKB (Sweden) liquid scintillation equip-ment.
Results
No significant differences were apparent indissolved oxygen concentrations in the enclo-sures and in the lake (Fig. 3). Temperaturedifferences between surface and bottom waterswere always less than TC. pH was lowest inthe fish enclosure in spite of the intensephotosynthesis, presumably due to enhanceddecomposition processes. Secchi disc trans-parency was similar at each station and wasapparently not affected much by fish, exceptduring the first and the last weeks of theexperiment. Between days 22 and 27 of theexperiment, transparency increased suddenly by1 m in both the enclosures and the lake, probablyas a result of the preceding four calm days whenwind speed remained below 3ms"'. A few dayslater this unusually high summer transparencywas reduced again by a storm. By the end of theexperiment, Secchi disc transparency was lowestin the enclosure with fish (Fig. 4).
Until the stocking of the enclosure with fish,no differences were observed in primary pro-duction between the enclosures and the lake.But after introduction of fish, phytoplanktonproduction increased to the greatest extent inthe fish enclosure (Fig. 5). Two peaks wererecorded in primary production at all stations:the first in the second week (maximum value55j«gCr'h"') and the second with a highermaximum value (65//gCr' h"') on the 28thday of the experiment.
Concentration of TN showed similar trendsat all stations. However, probably due to thestirring of bottom sediments, there was a

420 Istvdn Tatrai ami Vera Istvdnovics
8-8 -
j^ 8-6-a
6 2 -
2 0 -
18-
17-
I 10-
9-
e-
0
I .A.
Q.."o*.
m 18
I 'O-
? 0 -
^ • • < > • -
i8'5
r•I
nrf
-<<
lO 25 40Ooys
FIG. 3. Oxygen concentrations, temperature, tempcralure prolilc iind pH values in ihc enclosure with lish (•)withoiii lish (p) and in Lake Balaton (n). The arrnw indieales slocking of the enclosure wiih fish.
Doys
20 •50
20-1
4 0 -
6 0 -
E 80
100-
120-
10-18-
t0
FIG. 4. Secchi-dist transparency in the enclosures and in the lake and the wind velocity during the experiment(symbols as in Fig. 3).

Nutrient cycling by fish 421
6 0 -
_ 50-
4 0 -
30-
5 20E
10-
Days40
FIG. 5. Changes in primary production in ihe enclosures and in the lake (symbols as in Fig. 3).
striking increase in TN concentration in boththe lake and fish enclosure, exceeding1200//gl' ' by the last week of the experiment.Nitrogen fixation could ulso contribute to anincrease in TN. since the biomass <if nitrogenfixing algae increased markedly during ihal
period [Voros. pers. comm.) (Fig. 6). Untilthe supposed nitrogen fixation, inorganic nit-rogen constituted almost half of TN at allstations.
During the first week of the experiment theconccniration of inorganic nitrogen greatly
10 25 4 0
Days
FIG. ft. Concentrations of lolal nitrogen, ammonia and mineral nitrogen in the enclosures and in the lake(symbols as in FJg. ?>).

422 Istvdn Tdtrai and Vera Istvdnovics
exceeded the initial value (38OttgN! ') as aresult of intense mineralization processes (Fig.6). Thereafter the amount of inorganic nit-rogen decreased with increasing primary pro-duction and. later on, it did not change greatlyexcept in the fish enclosure. In the enclosurewith fish the concentration of inorganic nit-rogen, apart from small fluctuations, increasedgradually till the end of the experiment.Ammonia constituted the greatest part ofmineral nitrogen at all stations (Fig. 6).
TTie TP content of water increased suddenlyafter introduction of fish and. despite theincrease in phytoplankton production, its levelremained high thereafter. At the same time thepeak in the algal production coincided with TPvalues in the fish enclosures where its fluctuationwas more pronounced than in the controlenclosure and in the lake (Fig. 7). Consequently.TP values ranged between narrow limits (25-50/^gr ' ) in the fish-free enclosure and in thelake as well.
During the first half of the experiment TDPlevel was highest in Lake Balaton but fell tolower values in the fish enclosure. Lower TDPconcentrations in the fish enclosure with higher
primary production rate may indicate an in-tense cycling of this nutrient (Fig. 7). Surpri-singly, during the second half of the experi-ment. TDP gradually increased despite anincrease in algal production in the fish enclo-sure. This was probably due to intense decom-position of dead algal cells.
Discussion
Fish removal from lakes in most cases resultsin decreases of pl l , TN and TP (Andersson etat.. 1978: Henrjkson et ai, 1980). In ourinvestigation. pH decreased in the presence offish despite the fact that algal productionincreased. This could have resulted from anevolution of carbon dioxide as dead algal cellswere decomposed, indicating intense miner-alization processes in the fish enclosure.
Our hypothesis is that dense benthivorousfish populations may enhance eutrophicationprocesses even in a large shallow lake. Theenclosure results partly support this hypothesis.Both N and P were generally high in the
80-1
6 0 -
30-1
20-
10
Q
I'v'''
• — • —
cr
\ y
cr- • -
d'/
TDP
10 20
Days25 30 35 40
FIG. 7. Distributinn of total phosphorus and total dissolved phosphorus concentrations in the enclosures and inthe lakes (symbols as in Fig. 3).

presence of bream, despite a very high level ofproduction by phytoplankton. Moreover,according to the TN and TP contents of thewater column, phytoplankton was limited byboth N and P in the enclosure with fish, while Ndid not limit the algal production at all in the fishfree enclosure. This suggested that dense breampopulations might release nutrients in excess ofphytoplankton requirements.
Considerable amounts of nutrients, primari-ly ammonia, are released to the water by fishexcretion (Paulson. 1980). Thus, nitrogen ex-cretion of bream makes up 15% or l.2gm"-year"' of the total external load to LakeBalaton (Tatrai. 1984). Accordingly, breamwould exerete about 28mgm"- day"' ofammonia in the enclosure, which equals aloading rate of 12//gl"'. Oscillations in miner-al nitrogen concentrations can be accountedfor by the intense ammonia excretion by fish.Nitrogen load through excretion from benth-ivorous fish, even at moderate biomass. wasdecisive in the small eutrophic Lake Marion.Minnesota (().27gm"" year''; Shapiro et at..1982). In the deep eutrophic Lake Gjersj0cn.Norway, ammonia excretion of roach andbream caused an increase in algal biomass(Brabrand et al.. 1985).
Excretion rate of phosphorus by fish wascalculated using the data of Brabrand et at.(1985). On the basis of their work, breamwould be expected to excrete on average5.3mgFm~^ daily, which equals 15% of TDPand 3-8% of particuiate phosphorus in the fishenclosure. Lamarra (1975) showed that theinternal loading of P in shallow ponds in-creased in the presence of carp and that thechlorophyll concentration also increased,apparently as a result of this loading. Phos-phorus release by carp at a density of 50gm"'was quite similar to the excretion rate ofbream in the enclosure. However, phosphorusrelease by earp. at a density similar to what wehad in the enclosure (75gm''). was 40%higher than our calculated value. Benthivorousfish excretion of P in Lake Marion was asimportant as external sources supplying P tothe lake (88mg m'- year"'; Shapiro et at..1982). The study of Brabrand et al. (1985)indicates that large populations of omnivorousfish may be of importance in stimulating phyto-plankton growth in eutrophic lakes when theexternal supply of phosphorus is low.
Nutrient cycling by fish 423
Great summer variations observed in prim-ary production during the enclosure experi-ment in Lake Balaton, with equal or lowerednutrient levels, may be explained by internalnutrient sources from fish. This confirms thesuggestion of Shapiro et al. (1982) that man-agement of the fish population could be anefficient means to control primary producers inlakes. As the density of eyprinids increasedmarkedly during eutrophication in lakes inEurope, the activity of these and other plankti-vorous and benihivorous fish species probablycontributed to the eutrophication processes insuch lakes.
Fish removal as a means of biomanipulationhas led to improved water quality in manylakes and reservoirs (Shapiro & Wright. 1984;Benndorf c/a/., 1984). Bream removal is likelyto be an effective and eeonomic tool of waterquahty control even in large Lake Balaton.
Acknowledgment
We are grateful to Dr C. R. Townsend andtwo anonymous reviewers for their valuablecomments on the manuscript and for linguisticimprovements in the text. Our special thanksto Mr J- Zlinszky. G. Poor. B. Gyetvai and MrJ. Adamik for building the enclosures and toMrs B. Arszovszky for the drawing of figures.
References
Ahlgren 1 & Ahlgren G. (l^lfi) Methods of water-chemicat anatyses. Institute of Limnology, Up-psala.
Andersson G.. Berggrcn H.. C ronberg G. & GelinC. (1978) Effects of planklivorous and hcnthivor-ous fish on organisms and wLiicr chemistry ineutrophic lakes. Hydrobiotoj-ia. 59, 9-1?.
Benndorf J.. Kneschke H.. Kossatz K. & Penz E-(19H4) Manipuliilion of Ihe pelagic food web bystoeking with predacious fishes. InternationaleRevue der Gesamten Hydrohiotogie. 69. 41)7-428.
Biro P. & Garadi P. (1974) Investigations on thegrowth and population structure of bream. Ahra-mis brama L.. al differenl areas of Lake Balalon.The assessment of mortality and production.Annates Instituti Biotogici Titiany. 41, i.^3-179.
Brabrand A.. Faafetig B.A. & Nilssen J.P. (1985)Fish and nutrienl dynamics in a deep, meso-trophic lake. Verhandtungen der Internationalen

424 Istvdn Tdtrat atid Vera Istvdnovics
Veri'inigung fur Theoretische und AngewandteLimnologie. 22. 2553-25M.
Faafctig B.A. & Nilssen J.P. (l'.18n A twenty-yearstudy of cuirophicalioti in a deep, softwaler laki?.Verhanilhmgeri iter Internationalen Vereinigung fiirTheoretisclie und AngeHanitte Limnologie. 21.412-424.
Henrikson L.. Nyman H.G.. Oscarson H.G. &Stenson J.A.E. (1980) Trophic changes withoutchanges in the esiernal nuiricnt loading. Ihtlro-biotogia. 68. 2ri(>-:63.
Herodek S. (1977) Recent results of phyloplanklonresearch in Lake BaUiton. Annates Instituti Bioio-gici Tihany. 44, IHI-19f>.
Keen W.H, & GaglJardi J. (I9SI) Effeel of brownbullheads on release of phosphorus in sedimentand waier systems. Progressive Fish-Cutturist, 43.183-185.
Kitchell J-F.. Koonze J.F & Tennis P S . (1975)Phosphorus flux through fishes. Verhandtungender hiternatinnaten Veretnigung fiir Theoretisctieund Angewandte Limnotogie. 19. 247S-24S4.
Lamarr:i V.A. (197:S) Digestive activities of carp as amajor contributor to ihe nutrient loading oflakes. Verhandlungen de.s hwrnatiimalen Vereini-gung fiir Theoretische und Angnvundte Limnolo-gie. 19. 2461-246S.
Maekercth F.J.A.. Heron J. & Tailing J.F. (197S)Freshwaler Biological Association, ScientificPublication 36.
Nakashima B.S. & Lcggelt W.C. (1980) The rote offishes in the regulation of phosphorus availabiliiyin lakes. Canadian Journal of Fisheries amiAqualic Sciences. 37, l.'i4IH549.
Paulson I...J. (198(1) Models for ammonia excretionfor brook trout. .Satvetinus fontinath. and rain-bow troul. .Satmo gairdneri. Canadian Journal ofFisheries anil Aquatic Sciences. 37, 1421-1425.
Shapiro J.. Forsberg B.. Lamarra V.. Lindmark G..Lynch M.. Smelizer E. & Zolo G. (1982) Experi-mcnls and experiences in biomanipulation: stu-dies of ways Io reduce algal abundance andcliEiiinate btuegreens. U.S. Envjronnienlal Pro-teciion Agency EPA-6(M)/.1-S2-1W6. Also Limno-logicai Research tenter Interim Report No. 19.
Shapiro J. & Wrighl D.J. (1984) Lake restoration bybiomanipulatinn: Round Lake. Minnesota, thefirst two years. Frvshwaier Biotogy. 14. .171-383.
Talrai 1. (I9S4| Contribution of bream. AbramLsbrama L., and chironomids to the nitrogen budgetof Lake Balaton. Ph.D. thesis. Tihany-I.odz.
Tatrai I.. Toth G.L. & Ponyi J.E. (19S4)Tlie role nffishes in Ihe process of euirophicalion. E:nclosureexperiment in Lake Balaton. BurgentandiescherForsfhungs Bericht. 51. 111-122.
iManuscripl accepted 30 August 1985)





















