
the role of ankyrin repeat and socs box containing protein 4 ...
88
THE ROLE OF ANKYRIN REPEAT AND SOCS BOX CONTAINING PROTEIN 4 (ASB4) IN TROPHOBLAST DIFFERENTIATION AND PSEUDOVASCULOGENESIS WH Davin Townley-Tilson A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Department of Cell and Developmental Biology. Chapel Hill 2014 Approved by: Cam Patterson Keith Burridge Douglas Cyr Kathleen Caron Eleni Tzima
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of the role of ankyrin repeat and socs box containing protein 4 ...

THE ROLE OF ANKYRIN REPEAT AND SOCS BOX CONTAINING PROTEIN 4 (ASB4)
IN TROPHOBLAST DIFFERENTIATION AND PSEUDOVASCULOGENESIS
WH Davin Townley-Tilson
A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in
partial fulfillment of the requirements for the degree of Doctor of Philosophy in the
Department of Cell and Developmental Biology.
Chapel Hill
2014
Approved by:
Cam Patterson
Keith Burridge
Douglas Cyr
Kathleen Caron
Eleni Tzima

ii
©2014
WH Davin Townley-Tilson
ALL RIGHTS RESERVED

iii
ABSTRACT
WH Davin Townley-Tilson: The Role Of Ankyrin Repeat And SOCS Box Containing
Protein 4 (ASB4) In Trophoblast Differentiation And Pseudovasculogenesis (Under the
direction of Cam Patterson)
Vascularization of the placenta is a critical developmental process that ensures fetal viability.
The vascular health of the placenta affects both maternal and fetal well being; however,
relatively little is known about the early stages of placental vascular development. The
ubiquitin ligase Ankyrin repeat, SOCS box-containing 4 (ASB4) promotes embryonic
vascular lineage commitment and is highly expressed early in placental development. The
transcriptional regulator Inhibitor of DNA binding 2 (ID2) negatively regulates trophoblast
differentiation during development and is a target of many ubiquitin ligases. Due to their
contrasting effects during differentiation, we investigated whether ASB4 mediates vascular
differentiation through its ligase activity on ID2 in the placenta. Placentas from Asb4-/-
mice
exhibited myriad vascular differentiation defects, including abnormal overexpression of ID2,
and pregnant Asb4-/-
mice phenocopied human pre-eclampsia. We determined that ASB4
directly interacted with ID2 in trophoblast cells, leading to ID2’s ubiquitination and
subsequent degradation. Further, ASB4 promoted placental cell differentiation and function,
and co-expression of a degradation-resistant Id2 mutant abolished these effects. Together,
these findings indicate that ASB4 regulates trophoblast cell differentiation into placental
vasculature through the degradation of ID2 and that loss of Asb4 in the developing placenta
contributes to placental disease.

iv
This work is dedicated to mother, who wasn’t able to see me complete my goal; my father,
who instilled the value of education in me; my brother, who has always been smarter than
me; and my wife, who makes me better every day.

v
ACKNOWLEDGEMENTS
I would firstly like to thank my advisor, Dr. Patterson, for giving me the opportunity
to work in his laboratory. His advice, wisdom, and mentorship have been invaluable
throughout my time working for him.
I would like to thank my committee for pushing me to work faster, harder, and
smarter. Your suggestions, availability, and direction have guided and molded this work; it
couldn’t have been completed without your help.
Finally, to my Patterson laboratory colleagues; specifically Laura Dyer, who helped
me more than I deserved; Yaxu Wu, Judy Wu, and Pamela Lockyer who made up the rest of
“Team ASB4”; and the rest of my many lab-mates, who made (almost) every day at the lab a
fun, learning experience.

vi
TABLE OF CONTENTS
LIST OF FIGURES ............................................................................................................... viii
LIST OF ABBREVIATIONS .................................................................................................. ix
CHAPTER 1: GENERAL INTRODUCTION ..........................................................................1
Vasculogenesis, pseudovasculogenesis, and trophoblast differentiation .......................1
ASB4 as a ubiquitin ligase .............................................................................................4
ID family of proteins ......................................................................................................8
Pre-eclampsia and pathologies of the placenta ............................................................11
Hypothesis....................................................................................................................14
CHAPTER 2: ASB4 PROMOTES TROPHOBLAST DIFFERENTIATION
THROUGH THE DEGRADATION OF ID2 ..........................................................................22
Introduction ..................................................................................................................22
Materials and Methods .................................................................................................23
Mouse generation, blood pressure, and proteinuria .........................................23
In situ hybridization, immunofluorescence, and immunohistochemistry ........24
Cell culture and immunoblotting .....................................................................25
In vitro ubiquitination assay.............................................................................26
Placental cell differentiation assays .................................................................27
Statistical analysis ...........................................................................................28
Results ..........................................................................................................................28
ASB4 is expressed in undifferentiated TB cells and is
required for placental differentiation ...............................................................28

vii
ASB4 negatively regulates ID2 expression through
polyubiquitination and proteasome dependant degradation ............................31
ASB4 mediates placental cell differentiation and function in vitro .................33
Asb4-/-
mice phenocopy human patients with pre-eclampsia ...........................36
Discussion ....................................................................................................................37
CHAPTER 3: GENERAL DISCUSSION ...............................................................................55
The role of ASB4 in vascular commitment and patterning .........................................55
ASB4 as an E3 ligase ...................................................................................................58
The role of ASB4 in placental pathologies and pre-eclampsia ....................................61
REFERENCES ........................................................................................................................65

viii
LIST OF FIGURES
Figure 1.1 Placental development and differentiation ............................................................16
Figure 1.2 ASB4 as an E3 ligase ............................................................................................18
Figure 1.3 ID proteins regulates bHLH-mediated transcription .............................................19
Figure 1.4 Vascular remodeling during placental development .............................................20
Figure 2.1 Asb4 expression in the developing placental vasculature......................................41
Figure 2.2 Asb-/-
placentas express markers of undifferentiated
vasculature and TB cells ........................................................................................43
Figure 2.3 ID2 expression increases in placentas that lack Asb4 ...........................................44
Figure 2.4 ASB4 negatively regulates ID2 expression through
polyubiquitination and associates with ID2 in JAR cells .....................................45
Figure 2.5 ASB4 promotes JAR cell-mediated endothelial apoptosis
and stabilization of endothelial cell networks .......................................................47
Figure 2.6 ASB4 promotes TB cell differentiation in vitro ....................................................48
Figure 2.7 Pregnant Asb4-/-
mice display symptoms of pre-eclampsia ...................................49
Figure S2.1 Diminished mature vasculature in Asb4-/-
placentas is not
due to increased apoptosis or abnormal proliferation ............................................50
Figure S2.2 Asb4 deletion induces vascular dysfunction and mislocalization
of blood vessels in the placenta ............................................................................51
Figure S2.3 N-terminally tagged ID2 is resistant to ASB4-mediated degradation .................52
Figure S2.4 ASB4 degrades ID2 in a proteasome-dependant manner,
and does not affect ID2 cellular location ............................................................53
Figure 3.1 Model of ASB4 function in trophoblast cells ........................................................64

ix
LIST OF ABBREVIATIONS
APC/C Anaphase promoting complex/cyclosome
ASB4 Ankyrin repeat and SOCS containing protein 4
bHLH Basic helix-loop-helix
cKit Kit oncogene, a/k/a stem cell growth factor receptor
CTB Cytotrophoblast
dbcAMP N(6),2'-O-dibutyryladenosine 3':5' cyclic monophosphate
D-box Destruction box motif
DR Degradation-resistant
DS Degradation-sensitive
E Embryonic days post fertilization
E1 Ubiquitin activating enzyme
E2 Ubiquitin conjugating enzyme
E3 Ubiquitin ligase
ELISA Enzyme-linked immunosorbent assay
EPO Erythropoetin
ES Embryonic stem (cell)
FIH Factor inhibitin HIF
GAPDH Glyceraldehyde 3-phosphate dehydrogenase
hCG Human chorionic gonadotropin
HELLP Hemolysis, elevated liver enzymes, low platelet count
HIF Hypoxia inducible factors
HLH Helix-loop-helix

x
HRP Horseradish peroxidase
ID2 Inhibitor of DNA binding 2
IUGR Intrauterine growth restriction
JAR Choriocarcinoma cell line
K Lysine
KDM1 Lysine (K)-specific demethylase 1
LL Lysine-less
MEK1 Mitogen-activated protein kinase kinase 1
MMP Matrix metalloproteinase
PECAM Platelet endothelial cell adhesion molecule
PlGF Placental growth factor
Rb Retinoblastoma gene
RING-finger Really interesting new gene domain that binds zinc cations
ROC1 A RING-finger E3 ligase encoded by Rbx
SCF Stem cell factor ligand
SCFR Mast/stem cell growth factor receptor, a/k/a cKit
sFlt1 Soluble-Fms-like tyrosine kinase 1
SOCS Suppressor of cytokine signaling
TB Trophoblast
TBSC Trophoblast stem cell
TGC Trophoblast giant cell
TUNEL Terminal deoxynucleotidyl transferase dUTP nick end labeling
VEGF Vascular endothelial growth factor

1
CHAPTER 1
GENERAL INTRODUCTION
Vasculogenesis, pseudovasculogenesis, and trophoblast differentiation1
Blood vessel formation is classically divided into two categories: vasculogenesis and
angiogenesis. Vasculogenesis is the formation of new blood vessels from the de novo
production of endothelial cells. Angiogenesis refers to the formation of new blood vessels via
extension or remodeling of existing blood vessels. Angiogenesis occurs throughout
development and in adulthood, whereas vasculogenesis is generally thought to occur during a
limited period early in development.
Vasculogenesis is further divided into two categories: extraembryonic, that occurs in
the yolk sac (which functions as the source of early blood cell formation in the
developmental blood circulatory system) and allantois, (which gives rise to the umbilical
vasculature) and embryonic, i.e. restricted to the embryo itself [1]. Extraembryonic blood
vessel formation precedes embryonic vasculogenesis and provides communication between
the fetal circulation and the yolk sac to facilitate the transfer of nutrients and blood gases to
the developing embryo, and ultimately gives rise to the placenta [2]. In mammals, blood
islands assembling within the mesodermal layer of the yolk sac are the first occurrence of
vasculogenesis. Blood islands are foci of hemangioblasts that differentiate in situ, forming a
1 A version of this work was previously published as Townley-Tilson WHD, Wu Y, Ferguson JE III, Patterson
C. The ubiquitin ligase ASB4 promotes trophoblast differentiation through the degradation of ID2. PLoS ONE
9(2): e89451. doi:10.1371/journal.pone.0089451

2
loose inner mass of hematopoietic precursors and an outer luminal layer of angioblasts.
Blood islands eventually coalesce into a functional vascular network that constitutes the
vitelline circulation, which is adapted to transfer nutrients from the yolk sac to the embryo
proper [2]. The chorion is the outermost layer of extraembryonic mesoderm and
trophectoderm, surrounding the embryo and all other components of the conceptus. The
chorionic villi, which arise from the chorion, are small projections that intercalate the uterine
tissue, maximizing the surface area in contact with maternal blood [3]. Extraembryonic
vasculogenesis in Eutherians (placental mammals) also supplies the allantois with primitive
vessels in preparation for chorion fusion and umbilical vessel formation, thus initiating the
vascular connection between the fetal and maternal placental tissues [4]. This allantois-
chorion fusion is what is responsible for the placenta proper, whereby the allantois and
umbilical vasculature joins the chorion, which has now intertwined with the maternal
vasculature, forming a network of vessels that will supply the fetus with nutrients and
oxygen, and carry away waste from the fetus into the maternal blood stream where it can be
eliminated [3]. This vascularization of the early placenta is crucial for the health and viability
of not only the fetus, but also the mother (Figure 1.1) [5-7].
Embryonic vasculogenesis (sometimes referred to as intraembryonic vasculogenesis)
occurs throughout most of the embryonic mesoderm. The endocardium and great vessels are
the first embryonic endothelial structures formed during development [2]. Parallel with heart
development, vasculogenesis is initiated within the aortic primordia, a collection of
mesoderm just lateral to the midline, to give rise to the dorsal aortae and the cardinal veins
[8]. As the heart enlarges, passive diffusion of nutrients and waste becomes limiting, and a
coronary vasculature is formed to supply the metabolically active heart tissue. Vascular

3
precursor cells and the pro-epicardium make contact with the developing heart tube and
quickly spread over the entire heart, giving rise to the capillaries, veins, and arteries of the
coronary vasculature, smooth muscle cells and pericytes [9]. Lastly, the capillary plexi of
endothelial cells are remodeled into a unidirectional circuit to allow proper blood circulation
throughout the embryo, forming the mature vasculature. To accomplish this vascular
specification, endothelial cells are specified into an arterial or venous fate; then, as blood
flow commences, vascular smooth muscle cells are recruited to the endothelium to provide
structural support and elasticity to the now mature vasculature [10].
The junction of embryonic, extraembryonic, and maternal vasculature is the placenta.
Though the placenta is the only transient organ in the body, it requires a great deal of
specification for its development, especially within the context of the placental vasculature,
during its short window of development to ensure fetal and maternal wellness. Placental
vasculogenesis is similar compared with that of embryonic and extraembryonic
vasculogenesis in that endothelial cells must differentiate from progenitor cells to form the
vasculature. However, the initial progenitor cell reservoir is different, and many of the
differentiating endothelial cell characteristics are unique to the placenta. These characteristics
include different cell adhesion molecules [11] and intracellular markers of differentiation
[12].
The development of the placental vasculature involves many coordinated steps,
beginning in early gestation. The outer layer of the blastocyst is a population of trophoblast
cells, or the trophectoderm, which will form the extraembryonic and placental components of
the conseptus unit. During human pregnancy, a population of undifferentiated multipotent
placental cells, termed cytotrophoblasts (CTBs), differentiate into villous and extravillous

4
trophoblasts that form and remodel the placental vasculature [13]. Villous trophoblasts have
endothelial cell functions in the chorionic villi and also fuse into syncytiotrophoblasts, which
are the epithelial covering of these villi that penetrate the uterus [14]. Extravillous
trophoblasts invade and migrate through the junctional zone of the placenta into the maternal
decidua, where they replace the endothelial cells that line the spiral arteries in a process
called “pseudovasculogenesis” [6]. These differentiating CTBs “switch” integrin expression
profiles from one expression pattern that allows for cell motility and extracellular matrix
degradation during migration and invasion to an endothelial-like integrin profile of
differentiated cells that form tight junctions in the arteries, creating high capacity, low
resistance blood vessels that allow for the exchange of blood gasses and nutrients from the
mother to the developing fetus [11,15,16]. Many of these same processes are conserved in
mice [17] in that cells originating from a trophoblast (TB) stem cell progenitor migrate and
invade the maternal arteries, but in mice, these are thought to derive from trophoblast giant
cell intermediaries, rather than cytotrophoblast lineages [18].
Prior work in this lab uncovered the ankyrin repeat, SOCS box-containing 4 (ASB4)
protein as a mediator of embryonic stem cell to endothelial differentiation [19]. Further,
ASB4 was found to be highly expressed in the extraembryonic vasculature including the
allantois, yolk sac, and most notably, the developing placenta [19]. Therefore, we decided to
investigate the role of ASB4 in placental vascular development.
ASB4 as a ubiquitin ligase
ASB4 is an oxygen-sensitive E3 ligase that is a member of the Suppressor of
Cytokine Signaling (SOCS) superfamily of proteins. All SOCS proteins share a constant C-
terminal SOCS box domain and variable N-terminal protein-protein interaction motifs [20].

5
The SOCS box binds to an elongin-B/elongin-C/cullin/ROC complex of proteins whose
function as components of a ubiquitin ligase complex has been well characterized [21]. The
N-terminal protein interaction motif binds to substrate protein(s) to optimally position them
for ubiquitination and subsequent degradation [22]. Thus, SOCS proteins are the substrate-
receptors for an ubiquitin ligase complex that controls steady-state levels of substrate
proteins. Because of this essential function, SOCS proteins like ASB4 are carefully regulated
both at the transcriptional and post-translational level to tightly control substrate protein
levels.
The central role of all E3 ligases is targeting substrate proteins for ubiquitination. The
well characterized ubiquitination process involves three enzymes that catalyze the activation
and transfer of the 7 kDa ubiquitin protein to the targeted substrate protein. Briefly, the E1
ubiquitin activating enzyme covalently binds to the ubiquitin protein via an ATP-dependent
process then transfers it to the E2 ubiquitin conjugating enzyme. From there, the ubiquitin
molecule is either transferred directly to the substrate protein, due to the resulting structural
proximity modulated by the E3 ligase (where the E3 ligase never directly interacts with
ubiquitin itself), or is step-wise transferred to the E3 ligase, and further conjugated to the
substrate protein (Figure 1.2) [23].
There are three forms of ubiquitination: mono-, multi- (or multi-mono-), and
polyubiquitination. Mono-ubiquitination is where a single ubiquitin moiety is conjugated to a
single ε-NH2 group of an internal lysine residue of the target substrate. Proteins can also
undergo multi-ubiquitination, whereby a single ubiquitin is conjugated to multiple internal
lysine residues on the target protein. Lastly, polyubiquitination occurs when multiple
(typically greater than three) ubiquitin proteins are attached to a single internal lysine residue

6
(or a free α- NH2 group of the N-terminal residue) on the target substrate. These different
ubiquitination reactions result in very different consequences for the substrate protein.
Typically, mono- and multi-ubiquitination are thought to be involved in nonproteolytic,
reversible, signaling events such as endocytosis, membrane trafficking, DNA repair, and
gene silencing [24]. Polyubiquitination, in contrast, is generally thought of as being
responsible for the proteasome-dependent degradation of a target protein. However, adding
yet another layer of complexity to the ubiquitination process, ubiquitin chains can link to one
of seven different lysine residues (K6, K11, K27, K29, K33, K48, and K63) within the
ubiquitin molecule, leading to different signaling outcomes. Lysine 48-linked ubiquitin
chains are by far the most widely studied and best characterized, and proteins with these K48
side chains are targeted for proteolysis [25]. K63-linked polyubiquitin chains instead lead to
protein interactions that are involved in endocytic trafficking, inflammatory response, protein
translation, and DNA repair [26]. The other five homotypic polyubiquitination chains (K6,
K11, K27, K29 and K33) have been observed in cells; however, their roles are still emerging
[27].
ASB4 is one of 18 proteins in the ASB family, which are part of the suppressors of
cytokine signaling (SOCS) super-family. ASB4 contains nine ankyrin repeats, seven of
which are highly conserved, and a C-terminal SOCS box [28]. Ankyrin repeats are common
33-residue motifs that mediate protein-protein interactions and are found in proteins with
functions ranging from development to transcription and cell cycle control [29]. Like other
members of the ASB family [30], ASB4 associates with cullin, elongin, and ROC/Rbx
RING-finger proteins (possibly because ASB4 lacks a RING-finger domain), which are all
part of the ubiquitin ligase complex [19]. There is little evidence indicating a central function

7
of ASB4. However, areas of high energy consumption (e.g., testes, heart, and brain) in adult
mice have ASB4 ligase activity [31-33], reinforcing our hypothesis that ASB4 regulates
vascular development and differentiation [34,35]. Further, ASB4 is abundantly expressed in
the developing placenta and is highly upregulated during the differentiation of embryonic
stem (ES) cells into endothelial cell lineages [19]. In addition, Asb4 transcription decreases
when endothelial cells are challenged by laminar shear stress [36], highlighting the
importance of ASB4 in the vasculature.
Interestingly, many of the few reports describing ASB4 illustrate the epigenetic
regulation of Asb4, specifically as an imprinted gene [37-40]. In the case of Asb4, only the
maternal allele is expressed, while the paternal allele is silenced. Though genomic imprinting
has been found in all Eutheria, less than one percent of all genes are imprinted [41]. Though
not thoroughly understood, imprinting is thought as of a result of an evolving “disagreement”
between maternal and paternal genes over the allocation of maternal resources to offspring,
known as the Genetic Conflict Theory, which was originally identified by David Haig over
twenty years ago [42]. Briefly, this theory states that genes in offspring are predicted to
demand more resources from the mother than the mother is selected to provide, and the
optimal level of demand on maternal resources may differ for the two alleles depending on
the parent of origin. The genetic outcome of this parental antagonism predicts preferential
expression from one of the parental alleles [43,44].
Perhaps not surprisingly, most imprinted genes are highly expressed in embryonic,
fetal, and early post-natal stages of development. One site of high imprinting activity is in
trophoblast cells and is readily apparent due to the fact that many imprinted gene mouse
knockout models exhibit altered placental development and function [45], with many of these

8
imprinted genes showing conserved activity between mouse and human placental function
and growth [46]. In addition to morphological differences, recent evidence points toward the
role of imprinted genes mediating the cellular response to stressors or environmental cues
such as diet [47], alcohol [48], and superovulation [49]. Imprinted genes also mediate ion and
nutrient transport within the placenta [50,51] as well as growth and vascular function [52,53].
These functions illustrate the importance of imprinting as a key modulator of placental
development and outcome during gestation.
One of the key components to our investigation of ASB4 was the identification of a
target substrate protein. Though there are assays that can elucidate specific ligase-substrate
interactions (e.g. yeast-2 hybrid, co-immunoprecipitation, and the recently described
tripartite TUBE/2D-DIGE/MS assay [54]), all have major caveats. Thus, we resorted to a
candidate approach in our search for a substrate for ASB4. We isolated ID2, described
below, as a potential factor due to; its expression in the placenta during development, its role
in mediating vascular differentiation in the placenta, and its ability to be degraded by the
proteasome in ubiquitin-dependant manner.
ID family of proteins
The four members of the ubiquitously expressed family of Inhibitor of DNA binding
(ID) helix-loop-helix (HLH) proteins, ID1-ID4, function as dominant negative regulators of
basic HLH (bHLH) transcriptional regulators that mediate cell lineage commitment,
differentiation, proliferation, cell cycle control and senescence, metastasis, angiogenesis,
apoptosis, and maintaining cell ‘stemness’ [55-58]. Typical HLH proteins mediate homo-
and heterodimerization and contain a highly basic region adjacent to the HLH domain, which
facilitates transcriptional regulation via DNA binding to the canonical E- or N-box sequences

9
in target genes [59]. However, ID proteins lack this basic domain and instead function by
dimerization with transcription factors, namely members of the bHLH superfamily. The
resultant ID-bHLH heterodimer is thus unable to bind to DNA or mediate transcription
(Figure 1-3) [60]. Because bHLH proteins typically positively regulate differentiation though
DNA binding, ID proteins are also colloquially referred to as “inhibitors of differentiation.”
Further, there is significant, but not ubiquitous redundancy between the individual ID
proteins [61-64], which is not surprising given that the HLH domains of the ID proteins share
70-80% amino acid sequence identity [65].
ID proteins are tightly regulated by E3 ligases [66-68]. Unlike most ubiquitin
substrate proteins that are targeted for degradation, ID1 and ID2 are ubiquitinated on their
respective N-terminal residues [67,69]. To date, approximately a dozen other known proteins
undergo N-terminal ubiquitination [70], which differs from the N-end rule pathway. N-
terminal ubiquitination is when ubiquitin modification occurs on the free α- NH2 group of the
N-terminal residue of the substrate protein, with substrate recognition likely involving a
downstream or internal motif; in the case of ID proteins, this motif is a destructive box (D-
box) element [68]. Conversely, in the N-end rule pathway, the N-terminal residue of the
substrate protein serves at the recognition motif for modification, but the ubiquitin
modification itself takes place on an ε-NH2 group of an internal lysine residue [71]. While
the majority of proteins that are targeted for N-terminal ubiquitination contain internal lysine
residues, three (p14ARF
, HPV-58 E7, and p16INK4a
) are naturally occurring lysine-less proteins
[72,73]. However, while the internal lysine residues are not essential for the degradation of
these substrates of N-terminal ubiquitination, these internal residues may still play a
modulating role. That is, while lysine-less mutants of these substrate proteins are still

10
ubiquitinated and degraded, this reaction is slowed by two to three fold compared with that of
lysine-containing wild-type proteins [74,75].
ID2 is one of the better studied members of the ID protein family [76] and is involved
in vascular events, including angiogenesis [77] and tumor cell migration and invasion [78].
Further, ID2 is a tightly regulated mediator of placental development and vascular
differentiation [77,79,80]. Although ID2 is rapidly cleared via the proteasome [67], little is
known about the specific ubiquitin ligases that regulate its expression. To date, only the
anaphase-promoting complex/cyclosome-Cdh1 (APC/C(Cdh1)) has been identified as a
ubiquitin-mediated regulator of ID2 expression [68]. APC/C(Cdh1) restrains axonal growth
and controls axonal morphogenesis in post-mitotic neurons [81]. Conversely, ID2 mutants
that are resistant to APC/C(Cdh1) enhance axonal growth and overcome myelin inhibitory
signals to promote growth. However, through its proteolytic targeting of ID2, APC/C(Cdh1)
permits the accumulation of the Nogo receptor, a key transducer of myelin and axonal
inhibition mediated by bHLH transcriptional activation. Thus, APC/C(Cdh1) relieves the
ID2-mediated repression of bHLH transcription factors, which repress axonal growth, and is
vital for proper synaptic patterning [81]. Although these studies elegantly illuminate how E3
ligases control ID2 in post-mitotic cell morphogenesis and how ID2 interacts with bHLH
transcription factors in vivo, there is little evidence for other E3 ligases that regulate ID2
expression.
However, for ID2 to be a bona fide target of ASB4 in the placenta, the transgenic
over-expression of Id2 in mice would have to show some placental vascular defects. Indeed,
in retinoblastoma (Rb) deficient mice, there is a concordant upregulation of Id2 expression.
These mice display die embryonically due to abnormal trophoblast proliferation and

11
placental dysplasia [82]. However, when Id2 is also deleted in these mice, this placental
phenotype is abrogated [83], indicating that overexpression of ID2 induces abnormal
placental development. Therefore, using ID2 as a known regulator of placental differentiation
and vascularization, we examined the placenta in the context of Asb4 expression for similar
pathologies.
Pre-eclampsia and pathologies of the placenta
The central function of the placenta is to provide for an efficient and robust exchange
of nutrients and oxygen from the maternal blood supply to the fetal blood and to eliminate
waste products from the fetal blood supply back to the maternal blood. During development,
the entire conceptus unit (the embryo, yolk sac, chorion, and allantois) must synchronize
blood vessel formation. That is, shortly after the initiation of blood flow within the embryo,
the placental vasculature must also be completely formed, including its connection to the
maternal uterine arteries [84]. In normal placentation, the maternal and fetal vessels don’t
intermingle and the blood supplies never mix; however, the vessels must be localized in such
close proximity and arrangement to ensure adequate and constant exchange of nutrients,
oxygen, and waste. Therefore, the placenta must coordinate rapid expansion and growth to
ensure the adequate flow of nutrients and oxygen to the growing fetus, with precise and
controlled morphogenesis and vascular patterning to guarantee that vessels are in the correct
location and are properly developed in anticipation of a high rate of blood flow. These vessel
patterning events require an immensely intricate degree of vascular networking and
remodeling from the fetal umbilical cord and chorion through the outer layers of the decidua
and myometrium to ensure the health of not only the developing fetus but also that of the

12
mother, who is also at risk for vascular diseases during blood vessel development (Figure 1-
4).
As maternal-fetal blood flow proceeds, the oxygen levels of the developing embryo
increase rapidly from hypoxia to relative normoxia, and vessels experience rapidly increasing
shear stresses. All cells must transduce these environmental signals into appropriate
developmental responses [85]. This sudden increase in hemodynamics and the increase in
blood gas oxygen levels act as environmental cues that influence additional endothelial
development [86]. That is, hypoxia typically serves as a cue for angiogenesis, recruiting new
blood vessels by secreting growth factors that act specifically on vascular cells, which leads
to the breakdown of the vessel wall and concomitant migration and proliferation of
endothelial cells towards the ischemic tissue. In endothelium, HIF (hypoxia-inducible
factors) transcription factors induces; transcription of erythropoetin (EPO) which stimulates
blood cell formation; transcription and secretion of VEGF and FGF which stimulate
endotheilial migration towards hypoxic tissue; transcription of Flk1 and Flt1, which are
VEGF receptors potentiated under hypoxic conditions; and transcription of myriad genes that
are responsible for the immediate response to hypoxia and induce anaerobic respiration,
allowing for energy production in the absence of oxygen-dependant oxidative
phosphorylation [87]. However, the factor inhibiting HIF (FIH) is an asparaginyl
hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor
[88] in an oxygen dependant manner [89]. Shear stress also mediates vascular remodeling
through the cytoskeletal remodeling, release of nitric oxide, activation of transcription
factors, and mediating growth factor expression [90]. ASB4 is a well know target of FIH
hydroxylation [19], and is also regulated by shear stress at the transcriptional level [36].

13
Thus, the importance of ASB4 as an oxygen and hemodynamic sensor becomes apparent in
the context of vascular remodeling at key developmental time points [19,36,87].
Not surprisingly, when this complicated vascularization process goes awry,
disorganization or malformation of the placenta can cause deleterious effects. Typically,
most placental pathologies are due to either defects in differentiation, migration, and invasion
or are vascular. Failure of the decidua and/or blood vessels to attach to or penetrate the
maternal myometrium can result in complications such as abruptio placentae (placental
separation from the uterus) and placenta accreta, increta, and percreta (the abnormally strong
and advancing penetration into the uterus). Vascular defects in the myometrium,
endometrium, and placenta result in myriad defects ranging from placenta praevia and
chorangiosis to decidual and fetal vasculopathy [85]. Perhaps the most common placental
pathology is pre-eclampsia, which affects roughly five percent of all pregnancies [91].
Pre-eclampsia is characterized as the sudden onset of maternal hypertension,
proteinuria, and edema and typically occurs during the third trimester of pregnancy [92].
There is currently no cure for pre-eclampsia, other than delivery, which often happens prior
to the 40-week term in humans. Currently, the only treatments to alleviate the symptoms of
pre-eclampsia are antihypertensives, typically magnesium sulfate, which lower blood
pressure and slow the heart rate [93]. If left untreated and in severe cases of pre-eclampsia,
pre-eclampsia may progress into eclampsia (i.e., seizures) either during gestation or after
delivery, resulting in a relatively high incidence of maternal and/or fetal death [94,95]. Other
syndromes, such as HELLP (Hemolysis, Elevated Liver enzymes, Low Platelet count) and
intrauterine growth restriction (IUGR) may also be associated with pre-eclampsia [96,97].
The resultant costs of pre-eclampsia and consequent morbidities, treatment, and life-long

14
effects are considerable, and the hospital treatment of hypertensive pregnancies alone is
approximately $3 billion in the US annually [98].
The underlying pathophysiology of pre-eclampsia is thought to be rooted in vascular
dysfunction [99]. Incomplete or dysmorphic maternal spiral artery remodeling, global
endothelial cell dysfunction, and the aberrant reduction in placental vasculature are all
hallmarks of pre-eclampsia.[100]. While probably multifactorial, this vascular insufficiency
may be due to aberrant early TB differentiation [101]. In both humans and mice, vascular
progenitor trophoblasts must differentiate, migrate, and invade to ensure proper
neovascularization and vascular remodeling [102]. Some factors, such as placental growth
factor (PlGF), vascular endothelial growth factor (VEGF), and soluble-Fms-like tyrosine
kinase 1 (sFlt1) mediate the anti-angiogenic response [100], while other factors, such as
human chorionic gonadotropin (hCG) and matrix metalloproteinases (MMPs), affect invasion
and migration [103]. However, little is known about the initial differentiation events that
ensure vascular lineage commitment from trophoblast stem cells.
Hypothesis
In our pursuit to elucidate factors that mediate early vascular development, we have
identified ASB4 as an understudied E3 ligase that could potentially regulate extraembryonic
vascular development. Likewise, we have identified ID2 as a potential candidate protein that
may be regulated by ASB4, as ID2 mediates critical factors of placental differentiation and
vascularization. However, many questions remain. Is Asb4 expressed in the trophoblast stem
cells that have committed to a vascular lineage? Does the deletion of Asb4 in vivo induce
changes in differentiation? Does ASB4 regulate ID2 directly, polyubiquitinating ID2 and
targeting it for proteasomal degredation? Do ASB4 and ID2 have functional consequences in

15
placental cells? Does the deletion of Asb4 in mice induce a placental pathology? In the
following chapter, we address our central hypothesis that ASB4 promotes trophoblast-to-
endothelium differentiation through the degradation of ID2 by using a combination of in
vitro and in vivo approaches.
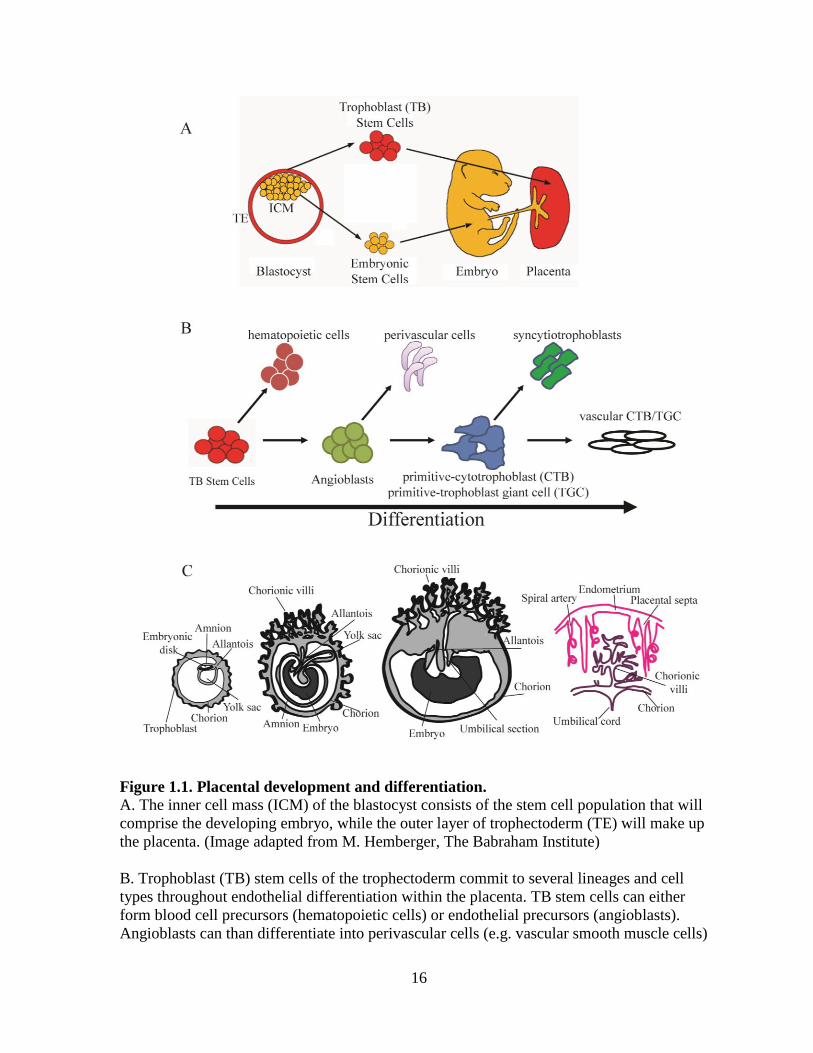
16
Figure 1.1. Placental development and differentiation.
A. The inner cell mass (ICM) of the blastocyst consists of the stem cell population that will
comprise the developing embryo, while the outer layer of trophectoderm (TE) will make up
the placenta. (Image adapted from M. Hemberger, The Babraham Institute)
B. Trophoblast (TB) stem cells of the trophectoderm commit to several lineages and cell
types throughout endothelial differentiation within the placenta. TB stem cells can either
form blood cell precursors (hematopoietic cells) or endothelial precursors (angioblasts).
Angioblasts can than differentiate into perivascular cells (e.g. vascular smooth muscle cells)

17
or primitive cytotrophoblasts (CTBs, in humans) or primitive trophoblast giant cells (TGCs,
in mice). These cells can then either form the epithelial syncytiotrophoblasts or the
endothelial-like vascular CTBs and vascular TGCs that will form the vascular system of the
placenta.
C. The placenta and corresponding fetal and maternal vasculature develops throughout
gestation. The outer trophoblast layer of the conceptus unit (left) develops into the chorionic
villi that circulates the fetal blood supply in close proximity to the maternal spiral arteries
(right). The allantois is the primitive vascular structure that eventually forms the umbilical
structure, while the yolk sac provides early circulatory function prior to the initiation of
blood flow in the placenta. The junction of the fetal vascular network (dark purple) and the
maternal arteries (pink) is highlighted on the right, demonstrating the compartmentalized, but
“intertwined” network of the placental vasculature. (Image adapted from Chaper 34 of [104])

18
Figure 1.2. ASB4 as an E3 ligase.
A. ASB4 is a 426 amino acid protein with three identifiable regions represented in ribbon
structure: an N-terminal variable domain (NTV), which functions in providing substrate
specificity; nine ankyrin repeats (denoted by AR1-AR9) that mediate protein-protein
interactions and protein folding; and a C-terminal suppressor of cytokine signaling (SOCS)
box, which binds to adaptor proteins of the ubiquitin ligase complex.
B. Like other SOCS proteins, ASB4 binds an elongin-B/elongin-C/cullin/ROC1 complex
through its SOCS box, which forms the components that mediate substrate protein
ubiquitination. In this reaction, ubiquitin (U) is transferred from the ubiquitin activating
enzyme (E1) to the ubiquitin conjugating enzyme (E2). Ubiquitin is either then transferred
directly to the substrate (as shown) or is transferred to the E3 ligase prior to the substrate.
This reaction repeats over several cycles, creating a polyubiquitin chain that targets the
substrate protein to the proteasome for degradation.

19
Figure 1.3. ID proteins regulates bHLH-mediated transcription.
Cells are maintained in “State 1” by the helix-loop-helix (HLH) inhibitor of DNA binding
(ID) proteins, which lack the basic DNA binding region that is common to the basic HLH
transcription factor family members. The ubiquitously expressed basic helix-loop-helix
(bHLH) E protein transcription factors activate transcription (State 2) by binding to promoter
E boxes as E protein–E protein homodimers (not shown in figure) or as E protein–tissue-
specific bHLH factor heterodimers. Formation of ID protein–E protein dimers prevents E
proteins from forming DNA-binding transcriptionally active complexes. ID downregulation
is necessary to allow for cell entry to “State 2”. (Image reprinted with permission from [105])

20
Figure 1.4. Vascular remodeling during placental development.
A. Spiral arteries from the maternal myometrium are remodeled during placental
development, ensuring a high capacitance, low resistance blood flow to bathe the fetal
circulation. As cytotrophoblasts (CTBs, in humans) or trophoblast giant cells (TGCs, in
mice) differentiate, they migrate and invade from the fetal components of the placenta to the
decidua, and replace the endothelial cells of the spiral arteries. (Adapted from [106])
B. By the third trimester of normal placental development, invasive CTBs/TGCs of fetal
origin have transformed spiral arteries them from small-caliber resistance vessels to high-
caliber capacitance vessels capable of providing placental perfusion adequate to sustain the
growing fetus. During the process of vascular invasion, the CTBs/TGCs differentiate from an
epithelial phenotype to an endothelial phenotype, a process referred to as

21
"pseudovasculogenesis" (upper panel). In preeclampsia, CTBs/TGCs fail to adopt an invasive
endothelial phenotype. Instead, invasion of the spiral arteries is shallow and they remain
small caliber, resistance vessels (lower panel), which may result in the placental ischemia,
maternal hypertension, and fetal growth restriction. (Image reprinted with permission from
[107])

22
CHAPTER 2
ASB4 PROMOTES TROPHOBLAST DIFFERENTIATION THROUGH THE
DEGRADATION OF ID2
Introduction
Previous work in this laboratory demonstrated that ASB4 is an oxygen-sensitive E3
ligase that is abundantly expressed in the developing placenta and is highly upregulated
during the differentiation of embryonic stem (ES) cells into endothelial cell lineages [19].
Also, ASB4 associates with cullin, elongin, and ROC/Rbx RING-finger proteins (possibly
because ASB4 lacks a RING-finger domain), which are all part of the ubiquitin ligase
complex [19]. Based on the high expression levels of Asb4 in the developing placenta,
coinciding with the role of ASB4 in vascular differentiation [19], we reasoned that any
putative substrates would share expression patterns and function within in the developing
vasculature.
ID2, a part of the anti-differentiation ID protein family, is a tightly regulated mediator
of placental development and vascular differentiation [77,79,80]. Due to the spatial and
temporal overlap and the functional contrast between these two proteins, we hypothesized
that ASB4 negatively regulates placental endothelial differentiation via and degradation of
ID2.
In this Chapter, we investigated the role of ASB4 in TB cell differentiation and
function and identified ID2 as a substrate of ASB4s ubiquitin ligase activity. Placentas
isolated from Asb4-/-
mice exhibited vascular differentiation defects, dysmorphic placental

23
vessels, vascular dysfunction, and spontaneous abortion in a subset of fetuses. Using cell
culture models, we found that ASB4 directly interacted with ID2, leading to ID2’s
ubiquitination and subsequent degradation in JAR cells. Further, ASB4 promoted aspects of
placental cell differentiation and endothelial cell replacement and vessel stability. Co-
transfecting Asb4 with Id2 mutants that are resistant to proteasomal degradation abolished
these effects. Lastly, pregnant Asb4-/-
mice exhibited symptoms consistent with pre-
eclampsia, including proteinuria and hypertension.
Material and Methods
Mouse generation, blood pressure, and proteinuria
The Asb4-/-
mouse generation is described by Ferguson [87]. Briefly, exon 1 of Asb4 was
flanked by loxP excision sites in the pAMC vector. Positive recombinants were
electroporated into 129 SvEv ES cells and cultured with appropriate selection enzymes. ES
cells were then injected into C57Bl/6 blastocysts and implanted into pseudopregnant females.
The resultant chimera (Asb4flox/+
) was then mated with EIIa-cre mice to excise the loxP sites.
These mice were further bred to 129 SvEv wild-type mice to ensure germ-line transmission
of the deletion and to outbreed the cre allele, generating Asb4+/-
mice on the 129 SvEv
background.
Maternal blood pressure was measured in conscious, pregnant mice at the gestational
stage indicated using a CODA 8 tail-cuff monitor (Kent Scientific, Torrington, CT, USA).
Mice were habituated to the machine for one day prior to data collection and assayed for five
consecutive days. Urinary creatinine and albumin protein levels were measured using the
Creatinine Companion and Albuwell M Test kits, respectively (Exocell, Philadelphia, PA,
USA). Urine collection consisted of placing isolated mice in metabolic cages (a generous gift

24
from Dr. Nobuyo Maeda [University of North Carolina]) for 24 hours. Food and water were
provided ad libitum, and urine was collected in a microcentrifuge tube placed below the
mesh flooring. Particulate matters and solids were removed from the samples by benchtop
centrifugation, and urine was stored at -20°C until assayed. Placental disc invasion was
assessed in E17.5 placentas as described in Dokras et al. [108]. All experiments were
approved by the Institutional Animal Care and Use Committee of the University of North
Carolina at Chapel Hill.
In situ hybridization, immunofluorescence, and immunohistochemistry
In situ hybridization for Asb4 was performed by the UNC In Situ Hybridization Core
Facility on 16-μm thick cryosections of placental tissue harvested under RNase-free
conditions from E11.5 wild-type mice. To generate the Asb4 probe, a ~900 bp fragment of
Asb4 was TA-cloned into pCRII-TOPO using the primers 5’-
CTCCGAGGATGGACGGCATCACTGCCCCTATC-3’ and 5’-
CTCAGGCTGTGCAGCAGGACGC-3’. The fragment was excised using NotI and BamHI
restriction enzymes. Sense and anti-sense probes were generated by transcription with T7 and
Sp6 polymerase, respectively. Probes were digoxigenin-labeled prior to hybridization.
Immunofluorescence and immunohistochemistry were performed as described in Waldo et
al. [109] on placental tissue sections at the indicated embryonic day. Briefly, tissue was
harvested and either flash frozen or fixed in 4% paraformaldehyde overnight with subsequent
cryoprotection in 30% sucrose. Samples were embedded in OTC Compound (Sakura Finetek,
Torrance, CA, USA) and sectioned into 6-μm thick slices by the UNC Histology Research
Core Facility. Primary antibodies are as follows: antibody recognizing mouse ASB4 was
generated as in Ferguson [87]; c-kit (Cell Signaling Technology, Danvers, MA, USA);

25
PECAM (Becton-Dickinson, San Jose, CA, USA); cytokeratin-17, integrin alpha V, and
integrin beta 4 (Abcam); ID2 (Cell Signalling Technology); Von Willebrand factor (Dako,
Carpinteria, CA, USA); FITC-conjugated Dolichos biflorus agglutinin (DBA) (Sigma-
Aldrich, St. Louis, MO, USA); and phospho-histone 3 (Millipore, Billerica, MA, USA).
Alexa Fluor antibodies (Invitrogen, Grand Island, NY, USA) and ABC Elite kits and
diaminobenzidine (Vector Labs, Burlingame, CA, USA) were used to detect primary
antibodies. Apoptosis was quantified using the ApopTag In Situ Apoptosis detection kit
(Millipore). Hematoxylin and eosin staining was performed on fixed frozen sections by the
UNC Histology Core Facility. Tissues and cells were imaged on a Nikon E800 upright
fluorescent microscope, and ImageJ (http://rsbweb.nih.gov/ij/) was used for quantification
and intensity measurements.
Cell Culture and Immunoblotting
JAR choriocarcinoma cells were obtained from ATCC (Manassas, VA, USA) and
maintained in MEM supplemented with 10% FBS. HEK-293T/17 cells and 2H-11
endothelial cells were maintained in DMEM supplemented with 10% FBS. 2H-11 cells were
conditioned to constitutively ectopically express ASB4 by transfecting cells with p3xFLAG-
CMV10-Asb4, and stable clones were selected with G418 for 12 days. Transgene expression
was confirmed by anti-FLAG immunoblotting. A stable Asb4 knockdown cell line was
created in 2H-11 cells using mouse Asb4 shRNA lentiviral particles. Endogenous Asb4
expression was screened by reverse transcription-PCR. Transfection reactions were
performed using LTX and Plus reagent (Invitrogen) according to Wolfe [110]. pCMV2B-
Asb4 and p3xFLAG-CMV10-Asb4 were generated and used as described in Ferguson [87].
Id2-Sport6 and pCS2-Id2 mutants were generous gifts from Dr. Aaron Ciechanover

26
(Technion-Israel Institute of Technology). siRNA transfections were performed using X-
tremeGENE siRNA transfection reagent (Roche, Indianapolis, IN, USA) according to the
manufacturer’s instructions. The siAsb4 duplex sequence is as follows: 5’-
CCACAAUGCUACCAUCAA-3’ and 5’-AGUUGAUGGUAGCAUUG-3’, and siRNA was
synthesized and duplexed by Integrated DNA Technologies (Coralville, IA, USA). Cell lysis
reactions were performed in cell lysis buffer (50 mM Tris, pH 7.5, 150 mM NaCl, 1 mM
EDTA, 1 mM EGTA) containing 1% Triton (immunoblots) or 0.5% NP-40
(immunoprecipitations). Cell fractionation assays were performed using NE-PER Nuclear
and Cytoplasmic extraction kit (Thermo Scientific, Rockford, IL, USA) according to the
manufacturer’s protocol. Cycloheximide was used at 50 µM in DMSO.. Immunoprecipitation
reactions were lysed as described above and crosslinked with 2 mM DSP
(dithiobis[succinimidylpropionate]) (Thermo Scientific) for 2 hours at 4°C. Lysates were pre-
cleared with the appropriate species IgG and Protein A/G beads (Santa Cruz Biotechnology)
for 1 hour at 4°C and then incubated with either anti-c-myc or anti-FLAG affinity gel
(Sigma-Aldrich). Primary antibodies include ID2 (Cell Signaling Technology); FLAG-HRP,
myc-HRP, and GAPDH (Sigma-Aldrich); KDM1 and MEK1 (Abcam, Cambridge, MA,
USA); and HA-HRP (Roche). Proteins were detected using HRP-conjugated, species-
appropriate secondary antibodies (Sigma-Aldrich) and developed using TMA-6 reagents
(Lumigen, Southfield, MA, USA).
In vitro ubiquitination assay
Recombinant ID2 was generated using an ID2-GST fusion construct (pGEX-2T-Id2)
graciously supplied by Dr. Antonio Iavarone (Columbia University) in BL21(DE3)pLysS
chemicompetent cells (Agilent Technologies, Santa Clara, CA, USA). Recombinant ASB4

27
was generated by ectopically expressing p3xFLAG-CMV10-Asb4 in HEK-293T/17 cells. E1
(Ube1), E2 (UbcH5a), ATP, and ubiquitin were purchased from Boston Biochem
(Cambridge, MA, USA). The reaction buffer was as follows: 50 nM E1, 2.5 μM E2, 2.5 μM
ASB4, 5 μM ID2, 2.5 mM ATP, 50 mM Tris, pH 7.5, 50 mM KCl, 0.2 mM DTT, and 250
μM ubiquitin. Reactions were performed at 37°C for 1 hour.
Placental cell differentiation assays
Trophoblast stem cells (TBSCs) were isolated as previously described [111] from
E7.5 embryos. Cells were grown for 6-8 weeks under normal culture conditions to minimize
spontaneous differentiation. Cells were cultured for 72 hours without serum and then
visualized for the presence of trophoblast giant cells, a hallmark of TBSC differentiation in
culture. JAR cells were induced to secrete human chorionic gonadotropin (hCG) using
N(6),2'-O-dibutyryladenosine 3':5' cyclic monophosphate (dbcAMP, Sigma-Aldrich) at 1
mM for 24 hours as described in Hohn et al. [112]. hCG was measured using an ELISA
(DRG International, Mountainside, NJ, USA), and concentrations were normalized to cell
number. The JAR cell-mediated apoptosis of 2H-11 endothelial cells was evaluated based on
Chen et al. [113], using the TUNEL-based ApopTag staining kit (Millipore) as an index of
apoptosis. To assay endothelial network stability mediated by placental cells, we adapted the
JAR cell/endothelial tube association assay from Aldo et al. [114], using 2H-11 endothelial
cells as our vascular network substrate. JAR cells were transfected as indicated, then plated
upon 2H-11 cells that had formed tube-like networks. Total area of the JAR/2H-11 network
was measured using ImageJ and quantified.

28
Statistical analysis
Unless otherwise noted, statistical analysis for all quantification was performed using
a two-tailed, unpaired Student’s t-Test. p values are reported in the respective figure legends.
Results
ASB4 is expressed in undifferentiated TB cells and is required for placental
differentiation
Asb4 is localized to areas of high vascular activity and is highly expressed in the
developing placenta [19,87]. ID2 is also critical in the early development of the placenta,
including the maturation of the placental vasculature [77]. Therefore, we hypothesized that
ASB4 would be an important modulator of differentiation in the placental vasculature. As
shown in Figure 2.1A (4x magnification) and 2.1A’ (20x magnification), Asb4 mRNA was
only expressed in the labyrinth zone of E11.5 placentas. This zone exhibits high vascular
activity [115] and contains the reservoir of TB cells that cross the junctional zone into the
maternal decidua as they mature into functional endothelial-like cells [15,116]. This
observation supports our hypothesis that ASB4 is involved in early vascular differentiation
events in the placenta.
To test whether ASB4 is expressed in differentiating cells committing to a vascular
lineage, we examined placentas at E11.5, when stem cells undergo both self-renewal and
differentiation and the cells adopting a vascular lineage migrate from the stem cell niche
[117]. Wild-type placental tissue sections were co-labeled for ASB4 and markers of various
stages of TB-to-endothelial differentiation. ASB4 co-localized with markers of pluripotent,
cells, specifically in a subset of cells expressing the general stem cell marker c-kit (Figure
2.1B) [118]. At this stage in mouse placental development, invading and differentiating

29
trophoblasts express c-kit, but terminally differentiated trophoblast giant cells and
spongiotrohoplasts do not. Further, c-kit and its ligand SCF are implicated have been
implicated as being required for trophoblast spreading and implantation, due to their role in
trophoblast differentiation [119]. In addition, ASB4 also co-localized with a subset of
PECAM-positive cells (Figure 2.1B’), suggesting that ASB4 is involved with cells that are
differentiating into vascular lineages at this time point. Further supporting a specific role in
early vascularization, ASB4 did not co-localize with cytokeratin 17, which is a marker of
mature placental endothelial-like cells (Figure 2.1B”).
Because ASB4 co-localized with early markers of the endothelium, we hypothesized
that Asb4 deletion would lead to functional consequences later in development. Specifically,
if ASB4 promoted TB-to-endothelial cell differentiation, then placentas of Asb4-/-
mice
should have less mature endothelium than those of wild-type mice. Placentas from wild-type
and Asb4-/-
mice were examined at E17.5 and labeled for cytokeratin 17 (Cyto17) to visualize
differentiated, mature endothelial-like cells. In wild-type mice, there abundant Cyto17
expression in the lining of the vessels, while in Asb4-/-
mouse placentas there were fewer
mature Cyto17-positive cells (Figure 2.2A), and this decrease was not due to impaired
proliferation or increased apoptosis (Figure S2.1). During normal gestation, placental cells
undergoing TB-to-endothelial differentiation undergo integrin “switching” [120], in which
undifferentiated TB cells that adopt a vascular phenotype express beta 4 integrins. Later in
gestation, when blood vessels have differentiated, beta 4 integrins are turned off, and alpha V
integrin is highly expressed [117,121], consistent with what we see in our wild-type mice
(Figure 2.2B, top panels). Asb4-/-
placentas failed to express integrin alpha V, but maintained
beta 4 integrin expression late in gestation (Figure 2.2B, bottom panels), consistent with a

30
failure of the placenta to undergo integrin switching, indicating an immature and
undifferentiated placenta. Placental disc invasion, during which the fetal components of the
placenta extend and expand into the maternal decidual layers, was more shallow in Asb4-/-
placentas compared with wild-type mice (Figure 2.2C) further confirming that placental
development is compromised in Asb4-/-
mice in a manner that is consistent with abnormal
differentiation [108]. Additional observations indicate that the vasculature expressed markers
of injury and dysfunction in near-term (E17.5) Asb4-/-
placentas (Figure S2.2A) and were
mislocalized within the junctional zone rather than the stage-appropriate outer deciduas
(Figures S2.2B and S2.2C). These results indicate that differentiation defects in Asb4-/-
placentas may have deleterious effects that are observed into late gestation.
Because Asb4-/-
placentas showed signs of early differentiation defects and impaired
vascularization (Figures 2.2A, B), we examined ID2 expression due to its anti-differentiation
role in the placenta [79]. We hypothesized that ID2 expression would be increased in the
Asb4-/-
placenta due to ID2’s anti-differentiation role in the placenta, and thus might underlie
the observed differentiation defects in Asb4-/-
placentas. In wild-type mice, ID2 expression is
downregulated as TB cells differentiate. However, in whole placental cell lysates at E12.5,
placentas from Asb4-/-
mice have a ~2 fold increase in ID2 expression compared with
placentas from wild-type mice (Figure 2.3A), and this finding is confirmed by
immunofluorescence in E13.5 placentas (Figure 2.3B). These data indicate that a subset of
TB cell remain undifferentiated in Asb4-/-
placentas, and provide evidence that ASB4 may
mediate ID2 expression in the placenta.

31
ASB4 negatively regulates ID2 expression through polyubiquitination and
proteasome dependant degradation
Given our observation that ID2 was significantly upregulated in Asb4-/-
placentas, we
hypothesized that ID2 may be a substrate of ASB4s ubiquitin ligase activity. To test this
hypothesis, wild-type Id2 was co-transfected with Asb4 in JAR cells, and ID2 expression was
examined. JAR cells are a desirable cell type because they do not express endogenous ID2 or
ASB4, allowing us to modulate both proteins without endogenous protein interference.
Because ID2 is rapidly turned over by myriad other proteins, resulting in a very short half-
life (Figure S2.4B and [67]), we added a low dose of MG-132 to experiments in Figure 2.4A-
E to slow proteasomal degradation events and visualize ID2 protein expression. ASB4
degraded ID2 in a dose-dependent manner in Asb4 and Id2 co-transfected JAR cells (Figure
2.4A). To test whether ID2 expression increased in the absence of ASB4, we transfected Id2
into 2H-11 endothelial cells that stably overexpressed Asb4. ID2 expression increased when
co-transfected with increasing amounts of an siRNA duplex targeting Asb4 (siAsb4) (Figure
2.4B). To determine whether ASB4 binds directly to ID2, we performed co-
immunoprecipitation assays using co-transfected 3x-FLAG-tagged Asb4 and 6x-myc-tagged
Id2 in JAR cells. As shown in Figure 2.4C, ASB4 was detected when ID2 was
immunoprecipitated and, conversely, ID2 was detected when ASB4 was immunoprecipitated
(Figure 2.4D). Further, we observed a dose response of the ID2-ASB4 interaction in 2H-11
cells that had increasing amounts of Id2 transfected in (Figure 2.4E). Together, these data
show that in placental cells, ASB4 can mediate ID2 expression and that ASB4 and ID2
interact with each other.

32
Because ASB4 is an E3 ligase, we hypothesized that ASB4 regulates ID2 protein
levels by polyubiquitinating ID2 and targeting it for proteasomal degradation. To test this
hypothesis, we first co-transfected HA-tagged ubiquitin and myc-tagged Id2 in 2H-11 cells
that either express endogenous Asb4 or have constitutively knocked down Asb4 expression to
levels undetectable at either the transcript or protein level. We immunoprecipitated ID2 and
blotted for HA, expecting a ubiquitin “smear” if ID2 was modified by polyubiquitination. As
shown in Figure 2.4F, ID2 ubiquitination increased dramatically in cells that expressed Asb4,
compared to cells that do not express Asb4. We then tested whether ASB4 could directly
ubiquitinate ID2 by performing an in vitro ubiquitination assay. We combined recombinant
ID2 with ASB4 and the minimal components required for ubiquitination and saw that ASB4
ubiquitinated ID2 four-fold more than the reaction absent of ASB4 (Figure 2.4G, lane 6
versus lane 3). Importantly, these reactions were performed in the absence of Roc1/Rbx1, the
RING-finger protein that associates with ASB4, indicating that ASB4 does not require a
RING-finger protein for ubiquitination. Previous reports into the mechanism of ID2
ubiquitination have demonstrated that ID2 is only susceptible to N-terminal ubiquitination
[67]. To determine whether ID2’s N-terminus is sensitive to ASB4-mediated degradation, we
co-expressed ASB4 with ID2 mutants that lack all lysine residues (LL-ID2) or have 6x-myc
tags on either the N-terminus (degradation resistant, DR-ID2) or C-terminus (degradation
sensitive, DS-ID2) in JAR cells. Only the N-terminally tagged ID2 (DR-ID2) expression
level remained unchanged in the presence of ASB4 (Figure S2.3), indicating that ASB4
mediates ID2 degradation via N-terminal ubiquitination.
To confirm our hypothesis that ASB4 degrades ID2 in a proteasome-dependent
manner, we completely abolished proteasomal activity with a high dose of MG-132 to cells

33
ectopically expressing ID2 and either ASB4 or vector control and compared ID2 expression
to cells that were not treated with MG-132. In cells treated with DMSO, ASB4 expression
led to reduced expression of ID2 as in Figure 2.4A. While a high dose of MG-132 increased
total ID2 expression in the absence of ASB4, this increase was not diminished by the co-
expression of ASB4 in the presence of MG-132 indicating that ID2 is degraded via the
proteasome (Figure S2.4A). Further, when cells ectopically expressing ASB4 and ID2 were
treated with cycloheximide to block protein translation, the half-life of ID2 decreased in the
presence of ASB4 (Figure S2.4B). To ensure that the reduction in ID2 expression in the
soluble fraction assayed above was not caused by ASB4 inducing ID2 translocation to an
insoluble part of the cell, we performed a cell fractionation assay (Figure S2.4C). There was
no observable accumulation of ID2 in any of the cell fractions when co-expressed with
ASB4, suggesting that ID2 is not translocated to other insoluble fractions of the cell upon
treatment with ASB4.
ASB4 mediates placental cell differentiation and function in vitro
In culture, TB cells can induce endothelial turnover [113] and increase the stability of
endothelial cell networks [114], recapitulating the in vivo events that occur when TB cells are
differentiating into endothelial-like cells. Because Asb4-/-
placentas express markers of
undifferentiated TB cells (Figure 2.2), we hypothesized that ASB4 would increase the JAR
cell-meditated apoptosis of 2H-11 endothelial cells as well as the vascular stability of vessel-
like networks formed by 2H-11 cells in culture. We observed that JAR cells transfected with
ASB4 induced 2H-11 endothelial cells to apoptose 3-fold more than vector control cells. Co-
expression of wild-type ID2 did not attenuate apoptosis, but JAR cells co-expressing DR-ID2
and ASB4 resulted in significantly fewer TUNEL-positive 2H-11 cells compared with JAR

34
cells that were only transfected with ASB4. Of note, DR-ID2 and ASB4 co-expression
elevated apoptosis compared with vector control cells, but this was significantly less than
cells that did express ASB4 alone (Figure 2.5A, B). These data demonstrate that ASB4
promotes a functional vascular phenotype that recapitulates in vivo endothelial replacement
with differentiating TB cells and that ID2 represses this effect.
Previous reports demonstrated that endothelial cells induce TB migration in culture
and that TB cells stabilize these endothelial vascular networks [114], representing a model of
the in vivo events that occur during TB differentiation [15]. To examine whether ASB4 could
promote TB cell stabilization of endothelial cell networks, we measured the ability of JAR
cells transfected with Asb4 and Id2 to form stable vascular networks over time, using
branching 2H-11 tube-like structures as the strata upon which JAR cells could migrate to and
stabilize. In isolation, 2H-11 cells plated on Matrigel consistently form branching tube-like
structures within approximately four hours [122] but devolve into spheroid cluster of cells
within 16 hours (data not shown). However, the addition of trophoblast cells can stabilize
these networks for days and even weeks in culture [114]. Therefore, we hypothesized that
Asb4-transfected JAR cells would stabilize these 2H-11 cell networks for significantly longer
than vector-transfected JAR control cells. JAR cells expressing ASB4, either alone or co-
transfected with wild-type Id2, were able to maintain and stabilize the 2H-11 vascular
networks well past 16 hours, when vector control cell networks had destabilized (Figure
2.5C, D). Cells co-transfected with Asb4 and DR-Id2 quickly and significantly destabilized
2H-11 networks at 16 hours and were indistinguishable from control networks after 48 hours.
These results indicate that ASB4 promotes TB endothelial-like cell function in vitro and that

35
ASB4 mediates these effects by degrading ID2, since DR-ID2 attenuates this ASB4-mediated
effect in placental cells.
Because ASB4 mediates vascular differentiation in ES cells [19] and we have
demonstrated that ASB4 negatively regulated the anti-differentiation protein ID2 (Figure
2.4A, B), we hypothesized that ASB4 would mediate placental cell differentiation through
the regulation of ID2 and tested this hypothesis in vitro. First, TBSCs were isolated from the
extraembryonic ectoderm of early post-implantation (E7.5) wild-type and Asb4-/-
embryos
and cultured on a feeder layer of mitotically inactivated MEFs, which promote the long-term
maintenance and proliferation of undifferentiated stem cells [19]. Large-scale multipotent
differentiation is expected for the first several passages, so cultures were grown 6-8 weeks
prior to serum-withdrawal. Terminally differentiated cells were sub-cultured out, leaving
only the undifferentiated embryoid bodies of TBSCs. Although the factors required for
TBSC-to-endothelial transformation are not yet know, TBSCs readily differentiate into
trophoblast giant cells (TGCs) [111]. Based on previous work from this laboratory [123], we
used serum withdrawal to promote TBSC differentiation. Thus, we used the appearance of
TGCs as an index of TBSC differentiation. After serum withdrawal for 72 hours, we
visualized the isolated cells with bright-field microscopy. As shown in Figure 2.6A, wild-
type TBSCs largely differentiated into large, multinucleated TGCs, which were
morphologically very different from the small, clustered, undifferentiated TBSCs that form
embryoid bodies as seen in Asb4-/-
cells (right panel). Further, differentiated TGCs laid flat
on the culture dish, while undifferentiated embryoid bodies had raised edges and appeared
more convex on the culture dish, allowing for easy identification.

36
To determine whether ASB4’s influence on TB cell differentiation involves ID2, we
examined human chorionic gonadotropin (hCG) secretion, a well-established marker of
trophoblast differentiation [112], in JAR cells that ectopically express ASB4 and ID2. hCG
secretion was stimulated via the addition of dbcAMP to the growth medium following the
indicated transfection for 48 hours and was subsequently measured in the medium by ELISA.
ASB4 stimulated hCG production approximately 2 fold compared with the vector control.
Co-transfecting wild-type Id2 with ASB4 did not abolish hCG production, but co-
transfection of Asb4 and DR-Id2 prevented hCG stimulation (Figure 2.6B). Together with
data in Figure 2.5, these results illustrate that ASB4 promotes placental cell differentiation
and function in vitro, and that ID2 mutants resistant to ASB4-mediated degradation can
inhibit the differentiation and function of TB cells in vitro.
Asb4-/-
mice phenocopy human patients with pre-eclampsia
Because our data indicate that ASB4 mediates placental cell differentiation and
function (Figures 2.4 and 2.5), and that Asb4 deletion has negative consequences in the
placental vasculature throughout development (Figure 2.2 and Figure S2.2), we investigated
whether the placental abnormalities found in Asb4-/-
mouse placentas contributed to the
placenta-specific disease pre-eclampsia, whose pathogenesis may stem from abnormal
placental vascular development [7]. Asb4-/-
female mice produced significantly smaller litter
sizes compared with wild-type female mice (Figure 2.7B) due to spontaneous abortion mid-
gestation (Figure 2.7A). Similarly, Asb4+/-
breeding pairs produced non-Mendelian ratios of
pups that were significantly skewed toward higher numbers of wild-type animals at the
expense of Asb4-/-
pups (Figure 2.7C). When investigating the source of lethality in the Asb4-
/- pregnancies, we observed that fetal growth halted at approximately E10.5 to E11.5 in a

37
subset of Asb4-/-
embryos. These embryos lacked functioning placental vascularization
(Figure 2.7A, and data not shown), which may contribute to the abortion and fetal
reabsorption seen in Asb4-/-
embryos [5].
Because ID2 expression is elevated in trophoblast cells placentas of women with pre-
eclampsia [79] and Asb4-/-
mouse placentas (Figure 2.2D, E), combined with the vascular
defects observed in Asb4-/-
placentas (Figure 2.2A, Figure S2.2), we investigated whether our
Asb4-/-
mice shared traits with human patients with pre-eclampsia, which is widely believed
to be a disease of the placental vasculature [101]. Two hallmarks of pre-eclampsia are
maternal hypertension and proteinuria during late-stage pregnancy. Pregnant Asb4-/-
female
mice had increased blood pressure during late gestation (E14-term), as compared to both
gestationally age-matched wild-type mice and Asb4-/-
mice during the first week of gestation
(Figure 2.7D). Further, pregnant Asb4-/-
female mice had higher ratios of albumin:creatinine
protein in their urine during late stage pregnancy than wild-type mice (Figure 2.7E).
Together, these results suggest that Asb4-/-
mice phenocopy human pre-eclampsia and may
serve as a model for both early placental vascularization and human placental disease.
Discussion
Strict control over the vascular patterning of the placenta is critical for both maternal
and fetal survival [124]. Aberrant differentiation events early in development negatively
affect the later formation of the vasculature [15], but relatively little is known what drives
early differentiation events. Although none of the limited data that identify putative
substrates or functions of ASB4 support a central function for ASB4 in vivo
[19,33,36,40,125], prior work from this laboratory has shown that ASB4 is involved in early
vascular differentiation and is highly expressed in the developing placenta [19]. Therefore,

38
we utilized Asb4-/-
mice, in conjunction with placenta-derived cells, to determine the function
of ASB4 during placental vascular differentiation. Consistent with our previous work [19],
we found that ASB4 is largely localized to the early endothelium in the placenta. We also
found that Asb4 deletion induces the expression of markers of undifferentiation in the
placenta, including the anti-differentiation protein ID2. Based on this data, along with the
expression pattern of various markers of TB cells and endothelial differentiation in Asb4-/-
placentas, we determined that ASB4 is involved in the earlier stages of differentiation events,
and the consequences of Asb4 deletion persist into later stages of gestation resulting in
insufficient placental vascularization.
Due to the limited information found in the literature, identifying a substrate of
ASB4s ligase activity was central to this investigation. Taking a candidate approach, we
reasoned that any ASB4 substrate would have to share its narrow spatiotemporal expression
pattern, contribute to vascular phenotypes, and be involved in differentiation. We determined
that the ID family of proteins would fulfill these criteria [56]. The ID proteins (ID1 to ID4)
are known to mediate differentiation and cell cycle control, which impact cell functions such
as metastasis, angiogenesis, apoptosis, and maintaining stemness [55-58]. Within these
processes, there is significant, but not ubiquitous redundancy between the individual ID
proteins [61-64]. Further, ID proteins, and ID2 specifically, are known to be tightly regulated
by E3 ligases [66-68]. Therefore, ID2 was chosen for investigation in this study due to its
involvement in differentiation [55], vascular development [126], and placental maturation
[79]. Using cell culture and biochemical techniques, we determined that ASB4 can directly
negatively regulate ID2 expression.

39
Placental remodeling requires three unique vascular events for proper function: TBSC
differentiation, replacement of endothelium with trophoblast cells, and vascular stabilization
to form high capacity vessels [15]. Possibly because the JAR cell line was isolated from CTB
cells in choriocarcinomas, these cells can be induced to mimic in vivo cells under certain
conditions. We adapted several methods to assess cell differentiation and function in culture,
and whether ASB4 promoted these events through the inhibition of ID2. Although these
methods do not completely recapitulate in vivo events, they collectively indicate that ASB4
has a pro-vascular differentiation function in placental cells. By ectopically expressing ASB4
in the JAR cells, we were able to determine that ASB4 promotes all three aspects of placental
vascular remodeling. In addition, using isolated TBSCs, we were able to observe primary TB
cell differentiation in culture. TBSCs that lack Asb4 remained in undifferentiated embryoid
bodies, in contrast to wild-type TBSCs which differentiated into TGCs upon serum
withdrawal. Because exact markers of TBSC and endothelial differentiation are not well
defined in the placenta, future studies will be needed to more precisely address these
differentiation events.
Using Asb4-/-
mice as a model for ASB4 function in vivo, we explored the phenotypic
consequences of Asb4 deletion, focusing particularly on the early placenta. Although the
majority of Asb4-/-
embryos survived to term, all had placentas with varying degrees of
vascular dysfunction. Further, embryonic lethality occurred in a subset of Asb4-/-
embryos at
approximately E10.5 due to gross endothelial disruption in the placenta. Furthermore,
placental vascular dysfunction in Asb4-/-
placentas also had deleterious effects on pregnant
mice, phenocopying women with pre-eclampsia. Though extremely common, little is known
about the pathogenesis of pre-eclampsia [127] and to date there is no cure other than delivery

40
of the placenta. Both third trimester hypertension and proteinuria, hallmarks of pre-
eclampsia, were recapitulated in Asb4-/-
mothers. This disease state, in conjunction with the
differentiation defects in the placenta of Asb4-/-
mice, provides a unique model of early
vascular development. Ultimately, this model of pre-eclampsia and vascular dysfunction may
be used to investigate therapeutic strategies for treating pre-eclampsia and other diseases of
the placenta.

41
Figure 2.1. ASB4 is expressed in the developing placental vasculature.
A) Asb4 mRNA is expressed only in the labyrinth zone of developing placentas. In situ
hybridization was performed on E11.5 placental sections and imaged with bright field
microscopy. Wide-field (4x, A) and higher magnification (20x, A’) anti-sense (AS)-probed
sections illustrate Asb4 localized to the labyrinth zone. A sense probe was used as a negative
control (A”).
B) ASB4 is expressed in a subset of c-kit-positive and PECAM-positive cells but not mature
cytokeratin 17-expressing cells. E11.5 placental sections were probed with markers of stem
cells (c-kit, B), endothelial cells (PECAM, B’), and differentiated TB cells (cytokeratin 17,

42
B”) and fluorescently imaged at 20x magnification. These images were then merged to show
co-localization. ASB4 only co-localizes with cells expressing c-kit and PECAM (arrows) but
not cytokeratin 17 (filled arrows). There are also subsets of c-kit or PECAM positive cells
that ASB4 did not co-localize at this stage (filled arrows).

43
Figure 2.2. Asb4-/-
placentas express markers of undifferentiated vasculature and TB
cells.
A) Placentas lacking Asb4 have reduced cytokeratin 17 expression in near-term placentas.
E17.5 placental sections from wild-type and Asb4-/-
mice were labeled with cytokeratin 17
(cyto17), a marker of terminally differentiated endothelial-like TB cells. Blood vessels (BVs)
in Asb4-/-
placentas display reduced cytokeratin 17 labeling compared with BVs in wild-type
placentas.
B) Placentas from E15.5 wild-type and Asb4-/-
placentas were labeled for integrin alpha V, a
marker of mature, terminally differentiated TB cells and integrin beta 4, a marker of
immature, undifferentiated TB cells. Wild-type placentas express alpha V but not beta 4
integins. Cells in Asb4-/-
placentas retain integrin beta 4 expression and fail to express
integrin alpha V.
C) Placental disc invasion is reduced in Asb4-/-
mothers at E17.5, indicating restricted
trophoblast expansion. The ratio of the placental disc (P) to the total placenta (the sum of the
decidua (De) and the placental disc) is decreased in Asb4-/-
placentas compared to wild-type
placentas, indicating a defect in TB cell invasion and migration.
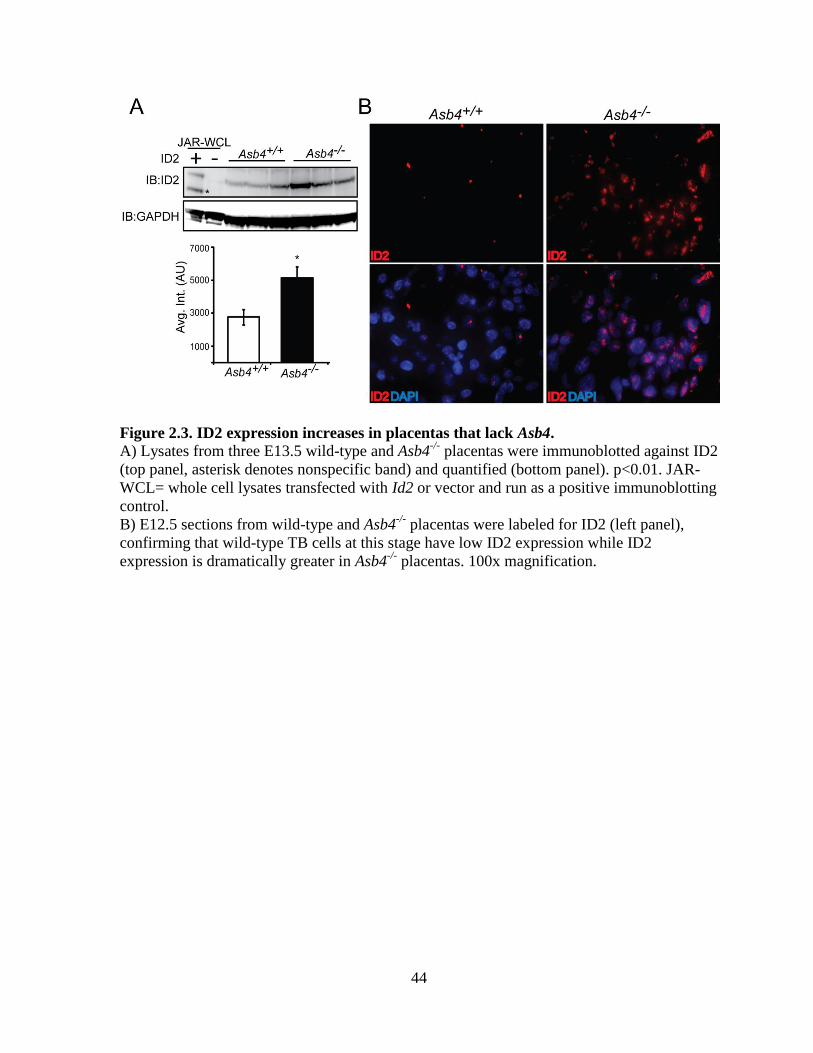
44
Figure 2.3. ID2 expression increases in placentas that lack Asb4.
A) Lysates from three E13.5 wild-type and Asb4-/-
placentas were immunoblotted against ID2
(top panel, asterisk denotes nonspecific band) and quantified (bottom panel). p<0.01. JAR-
WCL= whole cell lysates transfected with Id2 or vector and run as a positive immunoblotting
control.
B) E12.5 sections from wild-type and Asb4-/-
placentas were labeled for ID2 (left panel),
confirming that wild-type TB cells at this stage have low ID2 expression while ID2
expression is dramatically greater in Asb4-/-
placentas. 100x magnification.
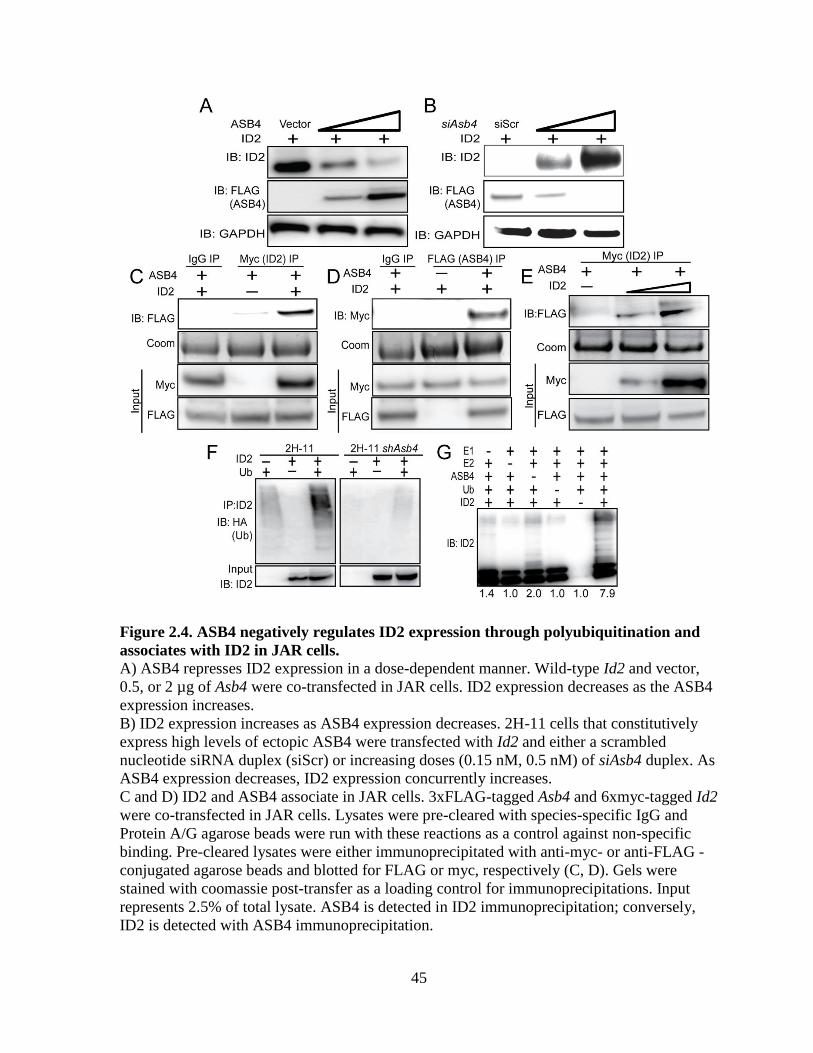
45
Figure 2.4. ASB4 negatively regulates ID2 expression through polyubiquitination and
associates with ID2 in JAR cells.
A) ASB4 represses ID2 expression in a dose-dependent manner. Wild-type Id2 and vector,
0.5, or 2 µg of Asb4 were co-transfected in JAR cells. ID2 expression decreases as the ASB4
expression increases.
B) ID2 expression increases as ASB4 expression decreases. 2H-11 cells that constitutively
express high levels of ectopic ASB4 were transfected with Id2 and either a scrambled
nucleotide siRNA duplex (siScr) or increasing doses (0.15 nM, 0.5 nM) of siAsb4 duplex. As
ASB4 expression decreases, ID2 expression concurrently increases.
C and D) ID2 and ASB4 associate in JAR cells. 3xFLAG-tagged Asb4 and 6xmyc-tagged Id2
were co-transfected in JAR cells. Lysates were pre-cleared with species-specific IgG and
Protein A/G agarose beads were run with these reactions as a control against non-specific
binding. Pre-cleared lysates were either immunoprecipitated with anti-myc- or anti-FLAG -
conjugated agarose beads and blotted for FLAG or myc, respectively (C, D). Gels were
stained with coomassie post-transfer as a loading control for immunoprecipitations. Input
represents 2.5% of total lysate. ASB4 is detected in ID2 immunoprecipitation; conversely,
ID2 is detected with ASB4 immunoprecipitation.

46
E) 2H-11 cells that stably express FLAG-tagged Asb4 were transfected with increasing
amounts of myc-tagged Id2. Cells were lysed and pre-cleared as in C and D, then
immunoprecipitated with anti-myc conjugated agarose beads and then blotted for FLAG.
FLAG expression increases in parallel with myc expression, indicating specific interaction
between ID2 and ASB4.
F) ID2 ubiquitination increases in cells with ASB4 expression. Wild-type Id2 and HA-tagged
ubiquitin were transfected into either 2H-11 cells that express endogenous Asb4 or 2H-11
cells that have Asb4 constitutively knocked down. ID2 was immunoprecipitated using anti-
ID2 and then blotted against HA. Reactions were blotted on the same membrane. Input
represents 2.5% of total lysate. Ubiquitination of ID2 increases in endothelial cells that
express ASB4 compared with cells that do not.
G) ASB4 directly ubiquitinates ID2 in vitro. Recombinant ID2 was incubated with
recombinant ASB4, and components of the reaction as indicated. Reactions were resolved on
SDS-PAGE gels and immunoblotted against ID2. ID2 is ubiquitinated approximately four-
fold more with ASB4 than without (lane 3). Quantification of ubiquitination is fold change
relative to lane 5 (without ID2).
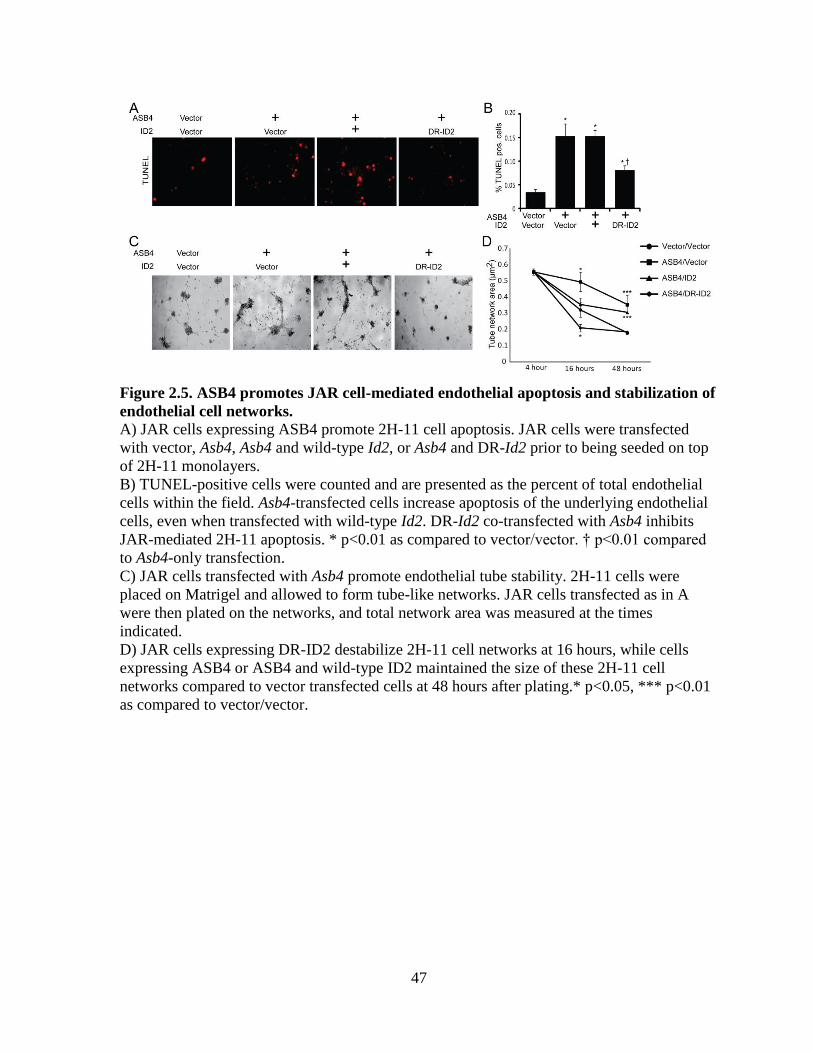
47
Figure 2.5. ASB4 promotes JAR cell-mediated endothelial apoptosis and stabilization of
endothelial cell networks.
A) JAR cells expressing ASB4 promote 2H-11 cell apoptosis. JAR cells were transfected
with vector, Asb4, Asb4 and wild-type Id2, or Asb4 and DR-Id2 prior to being seeded on top
of 2H-11 monolayers.
B) TUNEL-positive cells were counted and are presented as the percent of total endothelial
cells within the field. Asb4-transfected cells increase apoptosis of the underlying endothelial
cells, even when transfected with wild-type Id2. DR-Id2 co-transfected with Asb4 inhibits
JAR-mediated 2H-11 apoptosis. * p<0.01 as compared to vector/vector. † p<0.01 compared
to Asb4-only transfection.
C) JAR cells transfected with Asb4 promote endothelial tube stability. 2H-11 cells were
placed on Matrigel and allowed to form tube-like networks. JAR cells transfected as in A
were then plated on the networks, and total network area was measured at the times
indicated.
D) JAR cells expressing DR-ID2 destabilize 2H-11 cell networks at 16 hours, while cells
expressing ASB4 or ASB4 and wild-type ID2 maintained the size of these 2H-11 cell
networks compared to vector transfected cells at 48 hours after plating.* p<0.05, *** p<0.01
as compared to vector/vector.
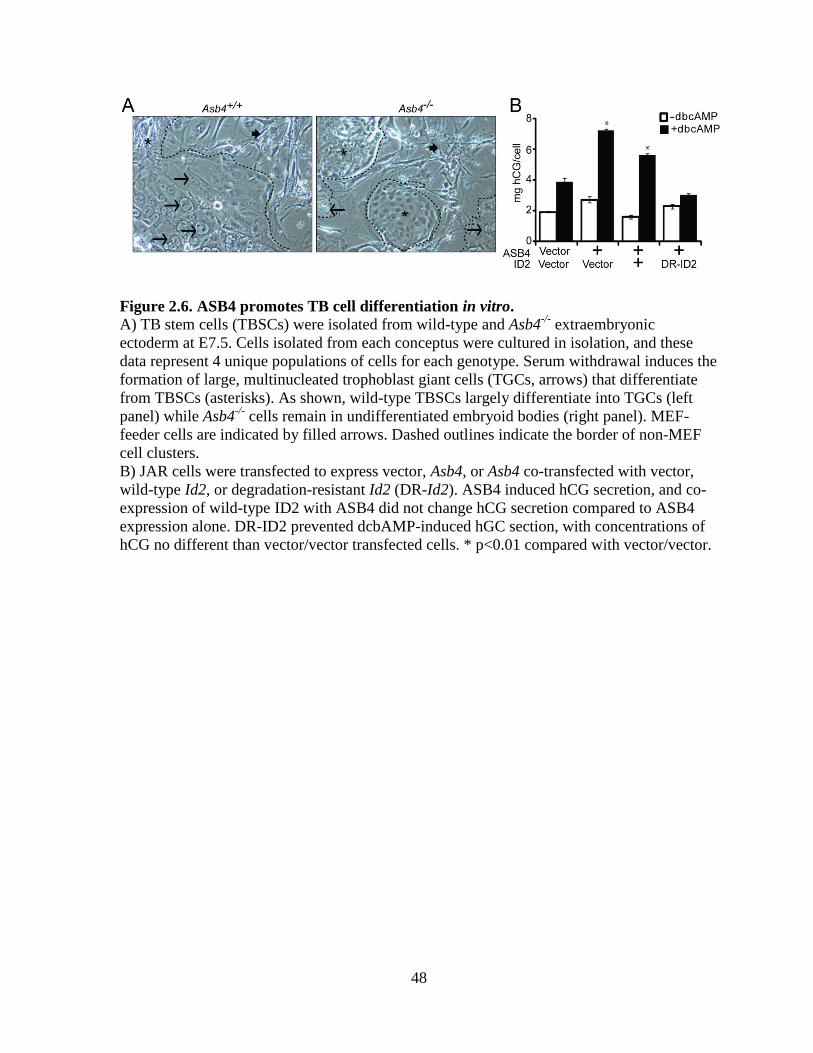
48
Figure 2.6. ASB4 promotes TB cell differentiation in vitro.
A) TB stem cells (TBSCs) were isolated from wild-type and Asb4-/-
extraembryonic
ectoderm at E7.5. Cells isolated from each conceptus were cultured in isolation, and these
data represent 4 unique populations of cells for each genotype. Serum withdrawal induces the
formation of large, multinucleated trophoblast giant cells (TGCs, arrows) that differentiate
from TBSCs (asterisks). As shown, wild-type TBSCs largely differentiate into TGCs (left
panel) while Asb4-/-
cells remain in undifferentiated embryoid bodies (right panel). MEF-
feeder cells are indicated by filled arrows. Dashed outlines indicate the border of non-MEF
cell clusters.
B) JAR cells were transfected to express vector, Asb4, or Asb4 co-transfected with vector,
wild-type Id2, or degradation-resistant Id2 (DR-Id2). ASB4 induced hCG secretion, and co-
expression of wild-type ID2 with ASB4 did not change hCG secretion compared to ASB4
expression alone. DR-ID2 prevented dcbAMP-induced hGC section, with concentrations of
hCG no different than vector/vector transfected cells. * p<0.01 compared with vector/vector.

49
Figure 2.7. Pregnant Asb4
-/- mice display symptoms of pre-eclampsia.
A) A subset of Asb4-/-
embryos dies in utero. Asb4-/-
littermates are shown at E12.5,
illustrating the lack of placental vasculature and dramatically reduced fetal growth in a subset
of Asb4-/-
embryos.
B) Quantification of the average litter size of wildtype mice compared to Asb4-/-
mice, taken
from more than 25 litters from each group.
C) Heterozygous breeding results in a lower than expected number of Asb4-/-
pups (p < 0.01,
Fisher’s exact test).
D) Pregnant Asb4-/-
mice have significantly elevated mean blood pressure in the third
trimester of pregnancy compared with both Asb4-/-
mice in the first week of pregnancy and
wild-type mice in the third week of pregnancy. * p<0.01.
E) Pregnant Asb4-/-
mice have significantly elevated urine-albumin:urine-creatinine in the
third trimester of pregnancy compared with both non-pregnant Asb4-/-
mice and wild-type
mice in the third week of pregnancy. * p<0.01.

50
Figure S2.1. Diminished mature vasculature in Asb4-/-
placentas is not due to increased
apoptosis or abnormal proliferation.
E15.5 placental sections from wild-type and Asb4-/-
mice were evaluated for aberrant
proliferation or apoptosis using phospho-histone H3 (pH3) or TUNEL, respectively. In both
cases, no discernible differences were noted between genotypes. BV; blood vessel. Arrows
denote TUNEL-positive cells.

51
Figure S2.2. Asb4 deletion induces vascular dysfunction and mislocalization of blood
vessels in the placenta.
A) Near-term (E17.5) placental sections were harvested and labeled with von Willibrand
factor to measure thrombus response and DBA to determine uterine natural killer cell
response. Asb4-/-
placentas display elevated thrombus/thrombosis response (left panel)
compared with wild-type placentas, indicating damaged vasculature. Further, there is a
dramatic increase in activated uterine natural killer cells (right panel) in Asb4-/-
tissues,
indicating elevated macrophage and immune response, compared to wild-type tissue.
B) E17.5 placental sections were stained with hematoxylin and eosin and examined for gross
morphology. Blood vessels (arrows) were counted and classified based on their location in
the labyrinth (LZ), junctional (JZ), or decidual (DE) zones. Blood vessels in wild-type
placentas are seen at the edge of the deciduas in, whereas significantly more vessels in Asb4-/-
placentas are located in the junctional zone, at the expense of the decidual zone.
C) Quantification of placental vessels in each respective zone, indicating that vascular
invasion/migration is defective in the absence of Asb4. * p<0.01 compared to wild-type.
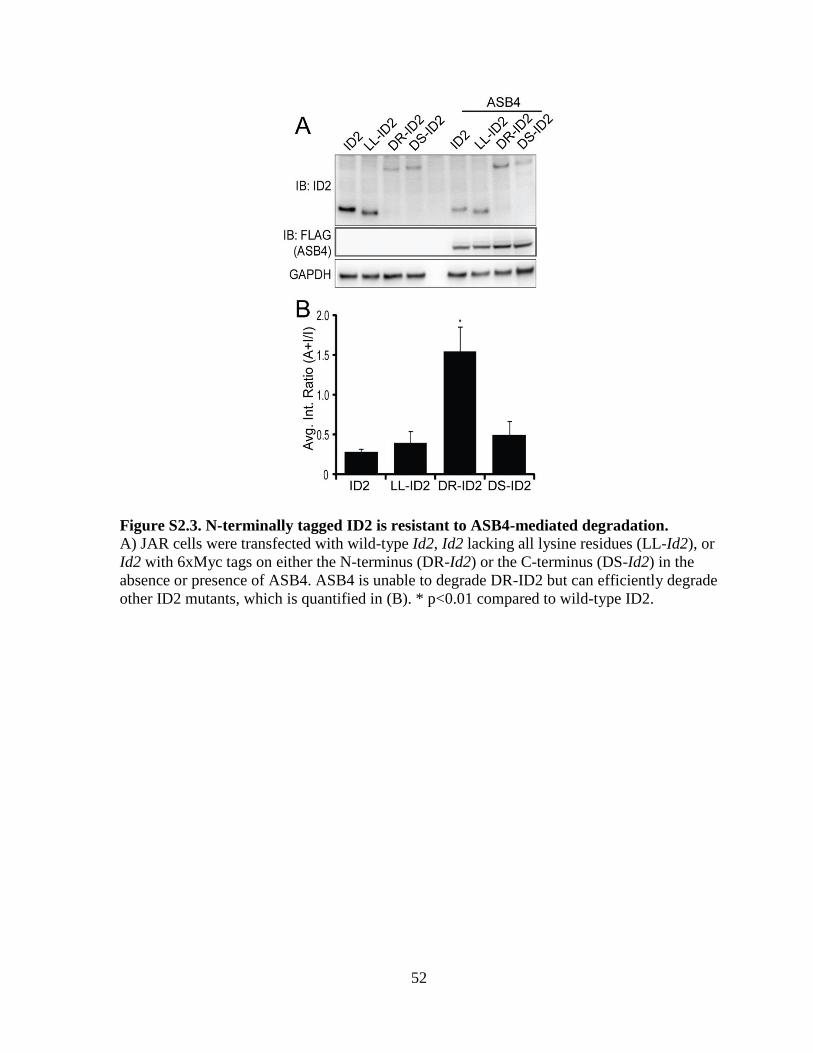
52
Figure S2.3. N-terminally tagged ID2 is resistant to ASB4-mediated degradation.
A) JAR cells were transfected with wild-type Id2, Id2 lacking all lysine residues (LL-Id2), or
Id2 with 6xMyc tags on either the N-terminus (DR-Id2) or the C-terminus (DS-Id2) in the
absence or presence of ASB4. ASB4 is unable to degrade DR-ID2 but can efficiently degrade
other ID2 mutants, which is quantified in (B). * p<0.01 compared to wild-type ID2.
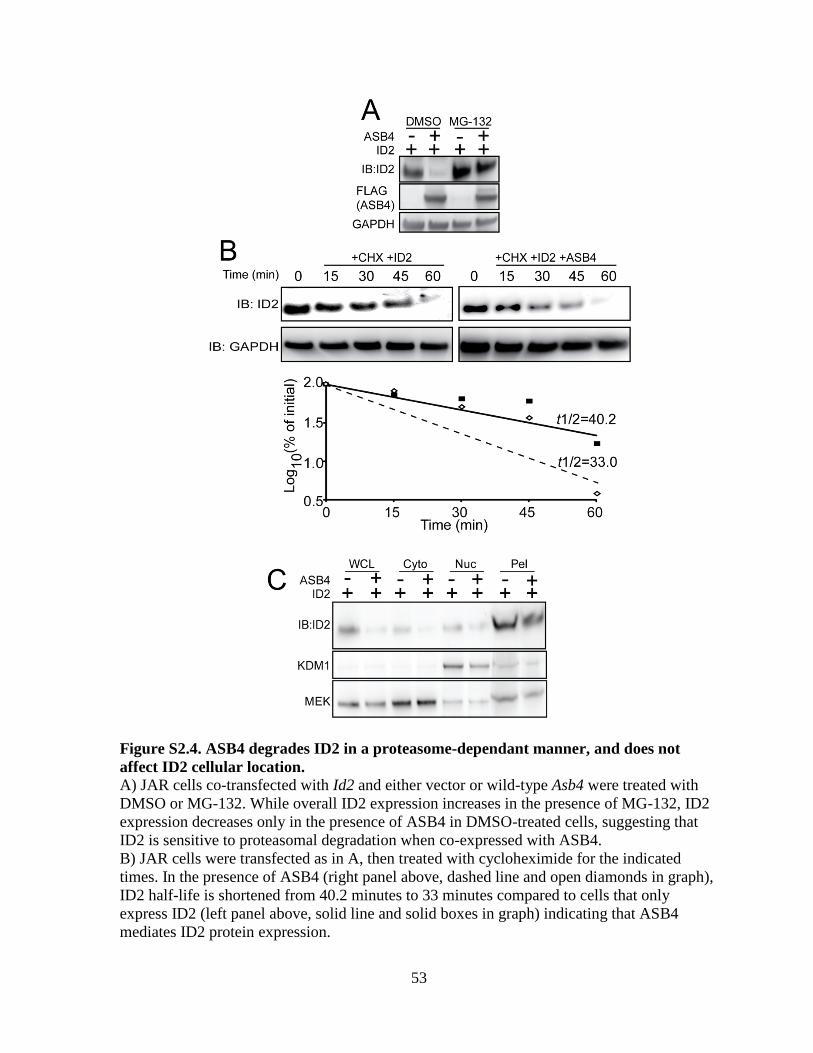
53
Figure S2.4. ASB4 degrades ID2 in a proteasome-dependant manner, and does not
affect ID2 cellular location.
A) JAR cells co-transfected with Id2 and either vector or wild-type Asb4 were treated with
DMSO or MG-132. While overall ID2 expression increases in the presence of MG-132, ID2
expression decreases only in the presence of ASB4 in DMSO-treated cells, suggesting that
ID2 is sensitive to proteasomal degradation when co-expressed with ASB4.
B) JAR cells were transfected as in A, then treated with cycloheximide for the indicated
times. In the presence of ASB4 (right panel above, dashed line and open diamonds in graph),
ID2 half-life is shortened from 40.2 minutes to 33 minutes compared to cells that only
express ID2 (left panel above, solid line and solid boxes in graph) indicating that ASB4
mediates ID2 protein expression.

54
C) ID2 sub-cellular localization is not altered in the presence of ASB4. JAR cells transfected
with Id2 and either vector or wild-type Asb4 were fractionated into the whole cell lysate
(WCL), cytoplasmic (Cyto), nuclear (Nuc), and Triton-insoluble pellet (Pel) fractions. In all
fractions, ID2 expression decreases in the presence of ASB4.

55
CHAPTER 3
GENERAL DISCUSSION
Placental development is a critical, but poorly understood process. Previous work
indicated that ASB4 mediated early vascular differentiation, and was localized to the
placenta. Thus I sought to fully characterize the role of ASB4 in placental vascularization
and trophoblast differentiation.
The experiments described herein attempt to fully characterize the role of ASB4
during placental vascular development by uncovering its expression, molecular function,
biologic consequences, and molecular regulation. Data generated here reveal that 1) Asb4 is
expressed in pluripotent trophoblast cells committed to the vasculature during a narrow time
window during differentiation; 2) ASB4, as a ubiquitination complex, degrades the HLH
protein ID2; and promotes differentiation of the vascular lineage through, in part, its
degradation of ID2; and 4) deletion of Asb4 in mice induces a placental vascular phenotype
and recapitulates pre-eclampsia in Asb4-/-
mothers (Chapter 2).
The role of ASB4 in vascular commitment and patterning
Previous investigations in this laboratory elucidated a role for ASB4 in the lineage
commitment step of pluripotent-to-endothelial differentiation [19]. Further, due to the high
expression pattern of Asb4 in “hotbeds” of vascular activity and vasculogenesis (e.g.
allantois, yolk sac, etc.) [19], we hypothesized that ASB4 would be responsible, in part, for
endothelial differentiation in these areas.

56
Earlier work from our laboratory on ASB4 function [19,36] led us to examine
vascular development in the placenta. Ferguson, et al [19] described how Asb4 expression
was restricted to a narrow spatiotemporal window in which the placenta was developing.
This study also elucidated that ASB4 functioned as an oxygen sensor, further implicating its
function in the placenta, where hemodynamic stresses and hypoxia/normoxia events are
required for proper vascularization. Likewise, Bode, et al. [36] described how laminar shear
stress repressed Asb4 expression. We therefore examined the gross morphological and
physiological consequences of globally deleting Asb4 expression in mice, which was
generated in [87].
Our initial investigation of Asb4-/-
mice established that there was embryonic lethality
in a subset of pups that was occurring at or about E9.5 (Figure 2.7A). This spontaneous
abortion appeared to occur in the absence of any gross fetal defect or pathology, but the
placentas of said pups resigned to intrauterine demise were dramatically less vascular or
developed (Figure 2.7A, lower conceptus) compared with littermates that appeared to be
morphologically “normal” (Figure 2.7A, upper conceptus). To begin understanding this
pathology, a histological examination was undertaken and revealed that ASB4 expression
only coincided with cells that were either pluripotent (Figure 2.1B) or cells that had
committed to the vascular lineage (Figure 2.1B’). Cells that had fully differentiated into
endothelium (Figure 2.1B”) and cells of different lineages (spongiotrophoblasts, trophoblast
giant cells, syncytiotrophoblasts, etc.) did not express Asb4.
Though there have been recent advances in understanding mouse placental
development, the field woefully lacks a definitive and authoritative consensus with regard to
markers of differentiating cells. Therefore, there are some caveats in trying to infer specific

57
cell types through the use of protein expression markers in the mouse placenta. To identify
pluripotent/stem cells in the context of the mouse placenta, we relied on the receptor c-Kit
(also known as CD117 or mast/stem cell growth factor receptor [SCFR]). c-Kit is a receptor
tyrosine kinase that binds to stem cell factor ligand and mediates proliferation,
differentiation, and serves as a marker of progenitor cells of mature endothelium [128].
However, c-kit expression is also found in trophoblasts that invade, migrate, and differentiate
into the TGC lineage [118,119], indicating that it may also serve as a marker undifferentiated
CTBs and TGCs, though this expression pattern has not been empirically determined.
Similarly, the use platelet endothelial cell adhesion molecule (PECAM) (also known as
CD31) and cytokeratin 17 for the determination of endothelial committed and mature
endothelial-like cells, respectively, was based on studies investigating their roles in human
placental tissue [129,130]. While we are confident that there are few or no discrepant
functions of these broad endothelial specific molecules between mice and humans, we must
acknowledge that there is a dearth of data that definitively describe these protein markers in
differentiating mouse trophoblasts.
These experiments illuminate the role of ASB4 in placental vascular determination
and differentiation. While we are limited by the scope of markers of endothelial
differentiation in the placenta, we are convinced that ASB4 plays an important role in early
placental vascular events, which correspond with earlier reports on the function of ASB4
[19]. However, to fully understand the role of ASB4, we asked how ASB4, as a ubiquitin
ligase, would function in the developing endothelium, and its context in the placenta.

58
ASB4 as an E3 ligase
Like all E3 ubiquitin ligase proteins, the functional ramifications of ASB4 are
dependent upon the function of the substrate protein(s) that it degrades. Multiple attempts to
identify binding partners of ASB4 have not yielded the identification of ASB4 ubiquitination
substrate proteins. Through the use of yeast 2-hybrid, the Patterson laboratory identified FIH
as an interaction partner of ASB4, though no putative substrate proteins were identified [19].
This result not only highlights the limitations of this assay, but also that of identifying
substrate proteins in general. The use of co-immunoprecipitation has also been successfully
utilized to identify co-factors of ASB4 [19], but protein turn-over, lack of specificity, and
difficulty in separating signal-to-noise all confound any results indicating a putative ASB4
target (data not shown and [87]).
Therefore, we took a candidate approach in our investigation into ASB4 substrates.
We knew that any substrate of ASB4 must be involved in early differentiation of the
vasculature, be expressed in CTBs in vivo, and have overexpression models that mimic Asb4
deletion. Literature searches and coinciding data (Figure 2.3B) indicated that members of the
ID family would fit these criteria. Specifically, studies of the retinoblastoma (Rb) protein in
mice indicate that Rb-deficient mice die embryonically due to abnormal trophoblast
proliferation and placental dysplasia [82]. This effect, however, is ameliorated by the
concomitant deletion of Id2 [83], signifying that Rb-mediated overexpression of ID2 induces
abnormal placental development. This Rb-induced ID2 overexpression also leads to the
formation of embryonal cancer, abnormal proliferation, and impaired angiogenesis, further
highlighting the importance of ID2 in early vascular and differentiation/proliferation events
[77].

59
Because ID2 mediates differentiation, invasion, and migration in CTBs [79], we
hypothesized that ASB4 normally regulates ID2 via its role as an E3 ligase. The role of ID2
in human placental differentiation has served as the crux for our further investigation in our
Asb4-/-
mice. Specifically, in cells driven to overexpress ID2, invasion was abrogated,
migration was abnormally enhanced, and these cells retained characteristics of
undifferentiated cells. Also of note, the abnormal maintenance of ID2 expression throughout
placental development was associated with placentas from pre-eclamptic patients. Taken
together, this report specifies the role of ID2 in the context of placental differentiation and
pre-eclampsia [79]. Other studies have also illuminated the importance of ID2 in the context
of proper placental development. Specifically, ID2 mediates transcriptional activity in the
placenta [80]; ID2 inhibits extravillous trophoblast-to-TGC differentiation [131]; and Id2 is
expressed and involved in trophoblast stem cell differentiation [132-134]. In conjunction
with the known expression profile of Asb4, the hypothesized function of ASB4, the
consequence of Asb4 deletion in mice, and our data demonstrating that ID2 and ASB4 have
reciprocal expression patterns (Figure 2.3B), we hypothesized that ID2 may be one substrate
of ASB4.
In undertaking our biochemical assays measuring the interaction of ASB4 and ID2,
we had to make several concessions. Foremost, we wanted to use the most representative cell
line but had to concede that no placental cell line endogenously expresses both Asb4 and Id2.
The CTB-derived JAR choriocarcinoma cell line best combined placental pertinence, ease of
manipulation, and assay-to-assay utility [135]. Because our JAR cells lack endogenous Asb4
expression, we also utilized the 2h-11 endothelial cell line for our experiments that required

60
endogenous Asb4 expression. Because these 2h-11 cells are not placental in origin, these
cells were limited to biochemical assays and assays that required endothelial cells [122].
ID2 has been shown to translocate from the nucleus to the cytoplasm during nervous
cell differentiation [136,137]. However, we observed a predominance of nuclear ID2
expression in vivo, as has been observed elsewhere [138], though nuclear and cytoplasmic
expression appears to be dependent on the differentiative and proliferative context of the
cells, such as in models of cancer [139,140]. We did not see any shifting of ID2 expression in
our JAR cell fractionation assay (Figure S2.4C), though we did not examine the expression
pattern during differentiation. Thus, while we show that ID2 expression decreases in the
presence of ASB4 in both the cytoplasm and nucleus, we did not explicitly demonstrate that
this was due to ubiquitination. That is, we cannot exclude the possibility that ASB4 may
somehow simply repress (or accentuate) translocation, which in turn may mediate ID2
stability. Similarly, we did not test whether ASB4 itself is able to translocation between the
cytoplasm and nucleus as is the case of another ID2 ubiquitin ligase, APC/C-Cdh1 [141].
These translocation events will have to be demonstrated in future studies to complete the
Pseudovasculogenesis and placental vascular remodeling occurs over time in a
complex three-dimensional space. Thus, to recapitulate these dynamic events, we adapted
various protocols to assay these complex and multifaceted in vivo events using in culture
techniques. By adapting various protocols, we were able to assay placental stem cell and
CTB differentiation [111,112], endothelial cell replacement by CTBs [113], and vascular
stability[114].
These assays and the resulting data are the best current techniques for addressing
complex questions regarding placental development; however, certain limitations are

61
obvious. Trophoblast stem cell and CTB differentiation assays either occur in isolation
(Figure 2.6A) or with exogenous pharmacological intervention (Figure 2.6B). Unlike true
endothelial differentiation [123], the endogenous factors required for trophoblast-to-
endothelial cell differentiation remain unknown. The models of CTB-mediated endothelial
cell apoptosis (Figure 2.5A) and vascular stability (Figure 2.5C) also occur in the absence of
exogenous factors, save for the components found in standard growth medium. However,
these assays represent the best current techniques available for recreating in vivo placental
defects and pathologies in cell culture.
The role of ASB4 in placental pathologies and pre-eclampsia
ASB4 is expressed in the developing placenta when the maturing vasculature is
exposed to dynamic changes in environmental cues and undergoes drastic morphologic
changes. Blood flow is initiated, which exerts hemodynamic stresses on the vasculature, and
maternal-fetal blood-gas exchange in the placenta is initiated, which drastically changes
embryonic oxygen tension from hypoxia to relative normoxia. Furthermore, the placental
vasculature undergoes extensive changes during this time, including differentiating and
making a vascular commitment, migration, and invasion. Links between these environmental
cues and morphologic changes are just beginning to be identified in both normal and
abnormal placental development, and ASB4 is spatiotemporally expressed and regulated in
such a way as to represent a potential direct link between differentiation cues and vascular
pattering responses.
The data presented in Chapter 2 suggest that ASB4 mediates the early differentiation
steps that promote trophoblast cells into the vascular lineage, but does not exclude the
possibility that ASB4 may also be involved in other vascular processes within the placenta.

62
Preliminary data in this laboratory indicate that ASB4 may mediate other aspects of vascular
function, such as migration (via the Boyden Chamber and wound healing assay) and
angiogenesis (via endothelial tube formation and subcutaneous Matrigel/tumor injections).
While these data do not negate the effects we observe, they add to the possibility and
plausibility that ASB4 functions in different contexts and milieus throughout development
and adulthood.
Though many placental pathologies are not completely idiopathic, the etiology of
most remains cloudy at best. As discussed in Chapter 1, disorders of placental adherence or
invasion (e.g., abruption placentae, placenta accreta/increta/percreta) are often attributed to
general vascular or blood dysfunction in cases where no previous or obvious trauma has
occurred [142]. Disorders where the placenta blocks the ostium of uterus (i.e., external
opening of the cervix) like placenta previa are again attributed to either previous trauma, or
general vascular dysfunction/dysmorphia [143]. Lastly, common disorders such as pre-
eclampsia, IUGR, and HELLP syndromes are all attributed to impaired differentiation and
function of the early placental vasculature [96,97,101]. Almost all cases of dysfunctional,
dysmorphic, and developmentally impaired placentas share some underlying vascular
component to the etiology or consequence of the pathologies.
Though most medical technologies have made enormous strides in the past decades,
the diagnosis and treatment for placental pathologies has remained woefully inadequate. For
the vast majority of placental disorders in which comorbidities are present, early delivery
remains the only option. The effects and consequences on both the mother and baby can be
chronic and severe. Children delivered before term can suffer from broad respiratory
illnesses (asthma, bronchopulmonary dysplasia), neurological delay or disorders (cerebral

63
palsy, autism), and physical ailments (enterocolitis, hearing loss, retinopathy) [144]. The cost
to the mother can be just as great, resulting in complications from caesarean section,
hysterectomy, eclampsia, gestational diabetes, hypertension, and infertility [142,145,146].
Diagnostically, placental pathologies remain critically understudied, and can currently
only be diagnosed after they have developed, precluding early interventions Typically, only
the onset of sudden idiopathic and localized abdominal pain, edema, and dramatically
increased blood pressure are used as indices for placental pathology. Further investigation
using ultrasounds, elevated serum levels of alpha-fetoprotein or sFlt or Endoglin, and the
onset of proteinurea are the only tools commonly available for diagnosis, and even then, most
placental complications are little more than educated guesses. Early diagnostic biomarkers
are critically needed.
Perhaps confounding this research is the fact that many placental pathologies do not
present until late in gestation. Many patients appear asymptomatic until the third trimester, or
even delivery, where complications can present fatal consequences if they are not prepared
for. Our elucidation of ASB4’s function in early second trimester placental development may
set a precedent for the earlier detection of pre-eclampsia, which may allow early
interventional strategies that would prevent serious late-term complications. While the
genetic component and contribution of ASB4 in human placental pathologies remains to be
studied, there are many homologies between mouse and human placental development [147].
Thus, further studies that address the role of ASB4 in human placental pathologies will be of
critical importance for the health of future mothers and infants.
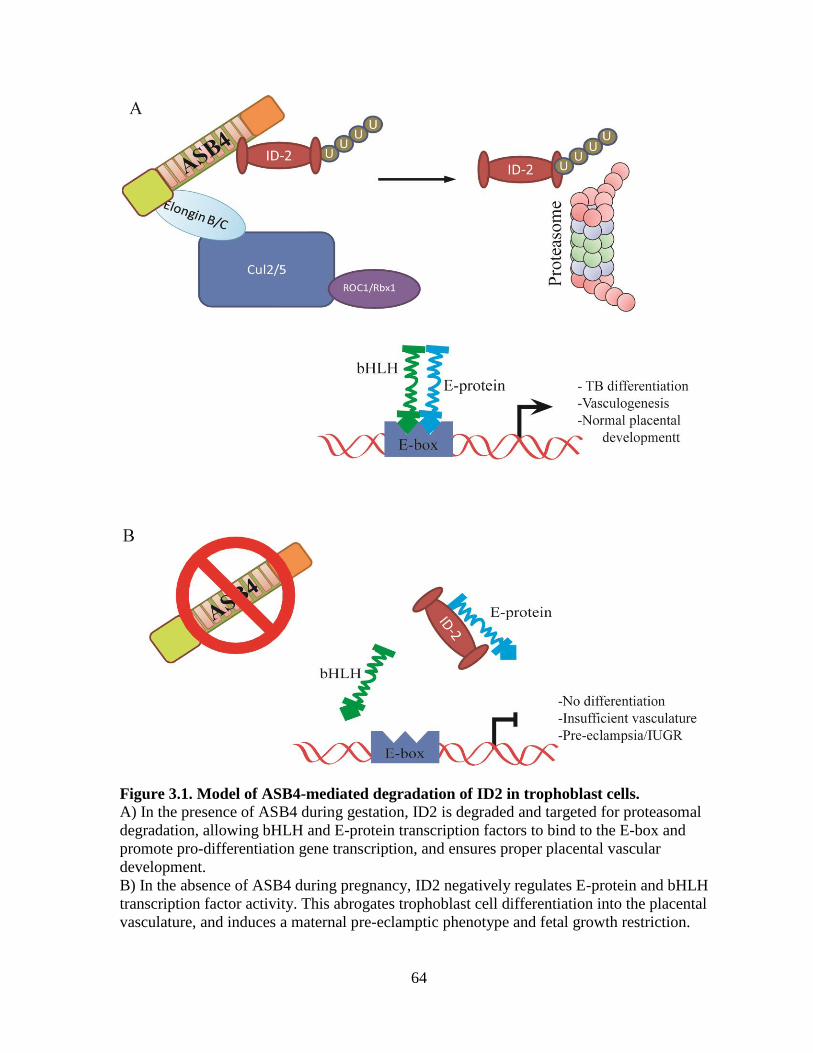
64
Figure 3.1. Model of ASB4-mediated degradation of ID2 in trophoblast cells.
A) In the presence of ASB4 during gestation, ID2 is degraded and targeted for proteasomal
degradation, allowing bHLH and E-protein transcription factors to bind to the E-box and
promote pro-differentiation gene transcription, and ensures proper placental vascular
development.
B) In the absence of ASB4 during pregnancy, ID2 negatively regulates E-protein and bHLH
transcription factor activity. This abrogates trophoblast cell differentiation into the placental
vasculature, and induces a maternal pre-eclamptic phenotype and fetal growth restriction.

65
REFERENCES
1. Ferguson JE, 3rd, Kelley RW, Patterson C (2005) Mechanisms of endothelial
differentiation in embryonic vasculogenesis. Arterioscler Thromb Vasc Biol 25:
2246-2254.
2. Drake CJ, Fleming PA (2000) Vasculogenesis in the day 6.5 to 9.5 mouse embryo. Blood
95: 1671-1679.
3. Huppertz B (2008) The anatomy of the normal placenta. J Clin Pathol 61: 1296-1302.
4. Downs KM (1998) The murine allantois. Curr Top Dev Biol 39: 1-33.
5. Conway SJ, Kruzynska-Frejtag A, Kneer PL, Machnicki M, Koushik SV (2003) What
cardiovascular defect does my prenatal mouse mutant have, and why? Genesis 35: 1-
21.
6. Kam EP, Gardner L, Loke YW, King A (1999) The role of trophoblast in the physiological
change in decidual spiral arteries. Hum Reprod 14: 2131-2138.
7. Myatt L, Webster RP (2009) Vascular biology of preeclampsia. J Thromb Haemost 7: 375-
384.
8. Dzierzak E (2003) Ontogenic emergence of definitive hematopoietic stem cells. Curr Opin
Hematol 10: 229-234.
9. Majesky MW (2004) Development of coronary vessels. Curr Top Dev Biol 62: 225-259.
10. Goldie LC, Nix MK, Hirschi KK (2008) Embryonic vasculogenesis and hematopoietic
specification. Organogenesis 4: 257-263.
11. Damsky CH, Fisher SJ (1998) Trophoblast pseudo-vasculogenesis: faking it with
endothelial adhesion receptors. Curr Opin Cell Biol 10: 660-666.
12. Ji L, Brkic J, Liu M, Fu G, Peng C, et al. (2013) Placental trophoblast cell differentiation:
physiological regulation and pathological relevance to preeclampsia. Mol Aspects
Med 34: 981-1023.

66
13. James JL, Stone PR, Chamley LW (2005) Cytotrophoblast differentiation in the first
trimester of pregnancy: evidence for separate progenitors of extravillous trophoblasts
and syncytiotrophoblast. Reproduction 130: 95-103.
14. Mayhew TM (2001) Villous trophoblast of human placenta: a coherent view of its
turnover, repair and contributions to villous development and maturation. Histol
Histopathol 16: 1213-1224.
15. Zhou Y, Fisher SJ, Janatpour M, Genbacev O, Dejana E, et al. (1997) Human
cytotrophoblasts adopt a vascular phenotype as they differentiate. A strategy for
successful endovascular invasion? J Clin Invest 99: 2139-2151.
16. Fisher SJ, Damsky CH (1993) Human cytotrophoblast invasion. Semin Cell Biol 4: 183-
188.
17. Adamson SL, Lu Y, Whiteley KJ, Holmyard D, Hemberger M, et al. (2002) Interactions
between trophoblast cells and the maternal and fetal circulation in the mouse
placenta. Dev Biol 250: 358-373.
18. Red-Horse K, Zhou Y, Genbacev O, Prakobphol A, Foulk R, et al. (2004) Trophoblast
differentiation during embryo implantation and formation of the maternal-fetal
interface. J Clin Invest 114: 744-754.
19. Ferguson JE, 3rd, Wu Y, Smith K, Charles P, Powers K, et al. (2007) ASB4 is a
hydroxylation substrate of FIH and promotes vascular differentiation via an oxygen-
dependent mechanism. Mol Cell Biol 27: 6407-6419.
20. Kile BT, Schulman BA, Alexander WS, Nicola NA, Martin HM, et al. (2002) The SOCS
box: a tale of destruction and degradation. Trends Biochem Sci 27: 235-241.
21. Kamura T, Sato S, Haque D, Liu L, Kaelin WG, Jr., et al. (1998) The Elongin BC
complex interacts with the conserved SOCS-box motif present in members of the
SOCS, ras, WD-40 repeat, and ankyrin repeat families. Genes Dev 12: 3872-3881.
22. Krebs DL, Hilton DJ (2000) SOCS: physiological suppressors of cytokine signaling. J
Cell Sci 113 ( Pt 16): 2813-2819.
23. Pickart CM (2001) Mechanisms underlying ubiquitination. Annu Rev Biochem 70: 503-
533.

67
24. Johnson ES (2002) Ubiquitin branches out. Nat Cell Biol 4: E295-298.
25. Hershko A, Ciechanover A (1998) The ubiquitin system. Annu Rev Biochem 67: 425-
479.
26. Pickart CM, Fushman D (2004) Polyubiquitin chains: polymeric protein signals. Curr
Opin Chem Biol 8: 610-616.
27. Komander D (2009) The emerging complexity of protein ubiquitination. Biochem Soc
Trans 37: 937-953.
28. Kile BT, Viney EM, Willson TA, Brodnicki TC, Cancilla MR, et al. (2000) Cloning and
characterization of the genes encoding the ankyrin repeat and SOCS box-containing
proteins Asb-1, Asb-2, Asb-3 and Asb-4. Gene 258: 31-41.
29. Mosavi LK, Cammett TJ, Desrosiers DC, Peng ZY (2004) The ankyrin repeat as
molecular architecture for protein recognition. Protein Sci 13: 1435-1448.
30. Kohroki J, Nishiyama T, Nakamura T, Masuho Y (2005) ASB proteins interact with
Cullin5 and Rbx2 to form E3 ubiquitin ligase complexes. FEBS Lett 579: 6796-6802.
31. Li JY, Chai B, Zhang W, Wu X, Zhang C, et al. (2011) Ankyrin repeat and SOCS box
containing protein 4 (Asb-4) colocalizes with insulin receptor substrate 4 (IRS4) in
the hypothalamic neurons and mediates IRS4 degradation. BMC Neurosci 12: 95.
32. Li JY, Chai BX, Zhang W, Wang H, Mulholland MW (2010) Expression of ankyrin
repeat and suppressor of cytokine signaling box protein 4 (Asb-4) in
proopiomelanocortin neurons of the arcuate nucleus of mice produces a hyperphagic,
lean phenotype. Endocrinology 151: 134-142.
33. Kim SK, Rhim SY, Lee MR, Kim JS, Kim HJ, et al. (2008) Stage-specific expression of
ankyrin and SOCS box protein-4 (Asb-4) during spermatogenesis. Mol Cells 25: 317-
321.
34. De Val S (2011) Key transcriptional regulators of early vascular development.
Arterioscler Thromb Vasc Biol 31: 1469-1475.

68
35. Luttun A, Carmeliet P (2003) De novo vasculogenesis in the heart. Cardiovasc Res 58:
378-389.
36. Bode M, Wu Y, Pi X, Lockyer P, Dechyapirom W, et al. (2011) Regulation of ankyrin
repeat and suppressor of cytokine signalling box protein 4 expression in the
immortalized murine endothelial cell lines MS1 and SVR: a role for tumour necrosis
factor alpha and oxygen. Cell Biochem Funct 29: 334-341.
37. Hu M, Zhao Z, Tuanmu LC, Wei H, Gao F, et al. (2014) Analysis of imprinted gene
expression and implantation in haploid androgenetic mouse embryos. Andrologia.
38. Bischoff SR, Tsai S, Hardison N, Motsinger-Reif AA, Freking BA, et al. (2009)
Characterization of conserved and nonconserved imprinted genes in swine. Biol
Reprod 81: 906-920.
39. Ono R, Shiura H, Aburatani H, Kohda T, Kaneko-Ishino T, et al. (2003) Identification of
a large novel imprinted gene cluster on mouse proximal chromosome 6. Genome Res
13: 1696-1705.
40. Mizuno Y, Sotomaru Y, Katsuzawa Y, Kono T, Meguro M, et al. (2002) Asb4, Ata3, and
Dcn are novel imprinted genes identified by high-throughput screening using RIKEN
cDNA microarray. Biochem Biophys Res Commun 290: 1499-1505.
41. Morison IM, Ramsay JP, Spencer HG (2005) A census of mammalian imprinting. Trends
Genet 21: 457-465.
42. Moore T, Haig D (1991) Genomic imprinting in mammalian development: a parental tug-
of-war. Trends Genet 7: 45-49.
43. Sandovici I, Hoelle K, Angiolini E, Constancia M (2012) Placental adaptations to the
maternal-fetal environment: implications for fetal growth and developmental
programming. Reprod Biomed Online 25: 68-89.
44. Wilkins JF, Haig D (2003) What good is genomic imprinting: the function of parent-
specific gene expression. Nat Rev Genet 4: 359-368.
45. Fowden AL, Coan PM, Angiolini E, Burton GJ, Constancia M (2011) Imprinted genes
and the epigenetic regulation of placental phenotype. Prog Biophys Mol Biol 106:
281-288.

69
46. Frost JM, Moore GE (2010) The importance of imprinting in the human placenta. PLoS
Genet 6: e1001015.
47. Coan PM, Vaughan OR, McCarthy J, Mactier C, Burton GJ, et al. (2011) Dietary
composition programmes placental phenotype in mice. J Physiol 589: 3659-3670.
48. Haycock PC, Ramsay M (2009) Exposure of mouse embryos to ethanol during
preimplantation development: effect on DNA methylation in the h19 imprinting
control region. Biol Reprod 81: 618-627.
49. Fortier AL, Lopes FL, Darricarrere N, Martel J, Trasler JM (2008) Superovulation alters
the expression of imprinted genes in the midgestation mouse placenta. Hum Mol
Genet 17: 1653-1665.
50. Coan PM, Fowden AL, Constancia M, Ferguson-Smith AC, Burton GJ, et al. (2008)
Disproportional effects of Igf2 knockout on placental morphology and diffusional
exchange characteristics in the mouse. J Physiol 586: 5023-5032.
51. Angiolini E, Fowden A, Coan P, Sandovici I, Smith P, et al. (2006) Regulation of
placental efficiency for nutrient transport by imprinted genes. Placenta 27 Suppl A:
S98-102.
52. Sekita Y, Wagatsuma H, Nakamura K, Ono R, Kagami M, et al. (2008) Role of
retrotransposon-derived imprinted gene, Rtl1, in the feto-maternal interface of mouse
placenta. Nat Genet 40: 243-248.
53. Coan PM, Angiolini E, Sandovici I, Burton GJ, Constancia M, et al. (2008) Adaptations
in placental nutrient transfer capacity to meet fetal growth demands depend on
placental size in mice. J Physiol 586: 4567-4576.
54. Rubel CE, Schisler JC, Hamlett ED, DeKroon RM, Gautel M, et al. (2013) Diggin' on
u(biquitin): a novel method for the identification of physiological E3 ubiquitin ligase
substrates. Cell Biochem Biophys 67: 127-138.
55. Lasorella A, Uo T, Iavarone A (2001) Id proteins at the cross-road of development and
cancer. Oncogene 20: 8326-8333.
56. Norton JD (2000) ID helix-loop-helix proteins in cell growth, differentiation and
tumorigenesis. J Cell Sci 113 ( Pt 22): 3897-3905.

70
57. Zebedee Z, Hara E (2001) Id proteins in cell cycle control and cellular senescence.
Oncogene 20: 8317-8325.
58. Niola F, Zhao X, Singh D, Castano A, Sullivan R, et al. (2012) Id proteins synchronize
stemness and anchorage to the niche of neural stem cells. Nat Cell Biol 14: 477-487.
59. Massari ME, Murre C (2000) Helix-loop-helix proteins: regulators of transcription in
eucaryotic organisms. Mol Cell Biol 20: 429-440.
60. Benezra R, Davis RL, Lockshon D, Turner DL, Weintraub H (1990) The protein Id: a
negative regulator of helix-loop-helix DNA binding proteins. Cell 61: 49-59.
61. Langlands K, Yin X, Anand G, Prochownik EV (1997) Differential interactions of Id
proteins with basic-helix-loop-helix transcription factors. J Biol Chem 272: 19785-
19793.
62. Carmeliet P (1999) Developmental biology. Controlling the cellular brakes. Nature 401:
657-658.
63. Miyazono K, Miyazawa K (2002) Id: a target of BMP signaling. Sci STKE 2002: pe40.
64. Samanta J, Kessler JA (2004) Interactions between ID and OLIG proteins mediate the
inhibitory effects of BMP4 on oligodendroglial differentiation. Development 131:
4131-4142.
65. Bounpheng MA, Melnikova IN, Dimas JJ, Christy BA (1999) Identification of a novel
transcriptional activity of mammalian Id proteins. Nucleic Acids Res 27: 1740-1746.
66. Bounpheng MA, Dimas JJ, Dodds SG, Christy BA (1999) Degradation of Id proteins by
the ubiquitin-proteasome pathway. FASEB J 13: 2257-2264.
67. Fajerman I, Schwartz AL, Ciechanover A (2004) Degradation of the Id2 developmental
regulator: targeting via N-terminal ubiquitination. Biochem Biophys Res Commun
314: 505-512.
68. Lasorella A, Stegmuller J, Guardavaccaro D, Liu G, Carro MS, et al. (2006) Degradation
of Id2 by the anaphase-promoting complex couples cell cycle exit and axonal growth.
Nature 442: 471-474.

71
69. Trausch-Azar JS, Lingbeck J, Ciechanover A, Schwartz AL (2004) Ubiquitin-
Proteasome-mediated degradation of Id1 is modulated by MyoD. J Biol Chem 279:
32614-32619.
70. Ciechanover A (2005) N-terminal ubiquitination. Methods Mol Biol 301: 255-270.
71. Varshavsky A (1996) The N-end rule: functions, mysteries, uses. Proc Natl Acad Sci U S
A 93: 12142-12149.
72. Kuo ML, den Besten W, Sherr CJ (2004) N-Terminal polyubiquitination of the ARF
tumor suppressor, a natural lysine-less protein. Cell Cycle 3: 1367-1369.
73. Ben-Saadon R, Fajerman I, Ziv T, Hellman U, Schwartz AL, et al. (2004) The tumor
suppressor protein p16(INK4a) and the human papillomavirus oncoprotein-58 E7 are
naturally occurring lysine-less proteins that are degraded by the ubiquitin system.
Direct evidence for ubiquitination at the N-terminal residue. J Biol Chem 279: 41414-
41421.
74. Breitschopf K, Bengal E, Ziv T, Admon A, Ciechanover A (1998) A novel site for
ubiquitination: the N-terminal residue, and not internal lysines of MyoD, is essential
for conjugation and degradation of the protein. EMBO J 17: 5964-5973.
75. Reinstein E, Scheffner M, Oren M, Ciechanover A, Schwartz A (2000) Degradation of
the E7 human papillomavirus oncoprotein by the ubiquitin-proteasome system:
targeting via ubiquitination of the N-terminal residue. Oncogene 19: 5944-5950.
76. Wong MV, Palasingam P, Kolatkar PR (2012) Cloning, purification and preliminary X-
ray data analysis of the human ID2 homodimer. Acta Crystallogr Sect F Struct Biol
Cryst Commun 68: 1354-1358.
77. Lasorella A, Rothschild G, Yokota Y, Russell RG, Iavarone A (2005) Id2 mediates tumor
initiation, proliferation, and angiogenesis in Rb mutant mice. Mol Cell Biol 25: 3563-
3574.
78. Coma S, Amin DN, Shimizu A, Lasorella A, Iavarone A, et al. (2010) Id2 promotes
tumor cell migration and invasion through transcriptional repression of semaphorin
3F. Cancer Res 70: 3823-3832.

72
79. Janatpour MJ, McMaster MT, Genbacev O, Zhou Y, Dong J, et al. (2000) Id-2 regulates
critical aspects of human cytotrophoblast differentiation, invasion and migration.
Development 127: 549-558.
80. Liu YP, Burleigh D, Durning M, Hudson L, Chiu IM, et al. (2004) Id2 is a primary
partner for the E2-2 basic helix-loop-helix transcription factor in the human placenta.
Mol Cell Endocrinol 222: 83-91.
81. Konishi Y, Stegmuller J, Matsuda T, Bonni S, Bonni A (2004) Cdh1-APC controls
axonal growth and patterning in the mammalian brain. Science 303: 1026-1030.
82. Wu L, de Bruin A, Saavedra HI, Starovic M, Trimboli A, et al. (2003) Extra-embryonic
function of Rb is essential for embryonic development and viability. Nature 421: 942-
947.
83. Lasorella A, Noseda M, Beyna M, Yokota Y, Iavarone A (2000) Id2 is a retinoblastoma
protein target and mediates signalling by Myc oncoproteins. Nature 407: 592-598.
84. Reynolds LP, Borowicz PP, Caton JS, Vonnahme KA, Luther JS, et al. (2010)
Uteroplacental vascular development and placental function: an update. Int J Dev
Biol 54: 355-366.
85. Benirschke K (2012) Pathology of the human placenta. New York: Springer.
86. Eichmann A, Yuan L, Moyon D, Lenoble F, Pardanaud L, et al. (2005) Vascular
development: from precursor cells to branched arterial and venous networks. Int J
Dev Biol 49: 259-267.
87. Ferguson JE (2007) Investigating the Role of Ankyrin Repeat and SOCS Box Protein 4
(ASB4) During Vascular Development: The University of North Carolina at Chapel
Hill.
88. Lando D, Peet DJ, Gorman JJ, Whelan DA, Whitelaw ML, et al. (2002) FIH-1 is an
asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-
inducible factor. Genes Dev 16: 1466-1471.
89. Mahon PC, Hirota K, Semenza GL (2001) FIH-1: a novel protein that interacts with HIF-
1alpha and VHL to mediate repression of HIF-1 transcriptional activity. Genes Dev
15: 2675-2686.

73
90. Ballermann BJ, Dardik A, Eng E, Liu A (1998) Shear stress and the endothelium. Kidney
Int Suppl 67: S100-108.
91. Redman C, Walker I (1992) Pre-eclampsia : the facts : the hidden threat to pregnancy.
Oxford ; New York: Oxford University Press. x, 197 p. p.
92. Maynard SE, Min JY, Merchan J, Lim KH, Li J, et al. (2003) Excess placental soluble
fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction,
hypertension, and proteinuria in preeclampsia. J Clin Invest 111: 649-658.
93. American College of O, Gynecologists Committee on Obstetric Practice Society for
Maternal-Fetal M (2013) Committee Opinion No. 573: Magnesium sulfate use in
obstetrics. Obstet Gynecol 122: 727-728.
94. Bulletins--Obstetrics ACoP (2002) ACOG practice bulletin. Diagnosis and management
of preeclampsia and eclampsia. Number 33, January 2002. Obstet Gynecol 99: 159-
167.
95. Swain S, Ojha KN, Prakash A, Bhatia BD (1993) Maternal and perinatal mortality due to
eclampsia. Indian Pediatr 30: 771-773.
96. Haram K, Svendsen E, Abildgaard U (2009) The HELLP syndrome: clinical issues and
management. A Review. BMC Pregnancy Childbirth 9: 8.
97. Cerdeira AS, Karumanchi SA (2012) Angiogenic factors in preeclampsia and related
disorders. Cold Spring Harb Perspect Med 2.
98. Quality AfHRa (2005) Health Care Utilization Project Data Source.
99. Roberts JM, Taylor RN, Musci TJ, Rodgers GM, Hubel CA, et al. (1989) Preeclampsia:
an endothelial cell disorder. Am J Obstet Gynecol 161: 1200-1204.
100. Szpera-Gozdziewicz A, Breborowicz GH (2014) Endothelial dysfunction in the
pathogenesis of pre-eclampsia. Front Biosci (Landmark Ed) 19: 734-746.
101. Fisher SJ (2004) The placental problem: linking abnormal cytotrophoblast
differentiation to the maternal symptoms of preeclampsia. Reprod Biol Endocrinol 2:
53.

74
102. Pijnenborg R, Robertson WB, Brosens I, Dixon G (1981) Review article: trophoblast
invasion and the establishment of haemochorial placentation in man and laboratory
animals. Placenta 2: 71-91.
103. Chen JZ, Sheehan PM, Brennecke SP, Keogh RJ (2012) Vessel remodelling, pregnancy
hormones and extravillous trophoblast function. Mol Cell Endocrinol 349: 138-144.
104. Pansky B (1982) Review of Medical Embryology.
105. Perk J, Iavarone A, Benezra R (2005) Id family of helix-loop-helix proteins in cancer.
Nat Rev Cancer 5: 603-614.
106. Wang Y ZS (2010) Vascular Biology of the Placenta.
107. Karumanchi SA, Maynard SE, Stillman IE, Epstein FH, Sukhatme VP (2005)
Preeclampsia: a renal perspective. Kidney Int 67: 2101-2113.
108. Dokras A, Hoffmann DS, Eastvold JS, Kienzle MF, Gruman LM, et al. (2006) Severe
feto-placental abnormalities precede the onset of hypertension and proteinuria in a
mouse model of preeclampsia. Biol Reprod 75: 899-907.
109. Waldo KL, Kumiski D, Kirby ML (1996) Cardiac neural crest is essential for the
persistence rather than the formation of an arch artery. Dev Dyn 205: 281-292.
110. Wolfe M (2006) Culture and Transfection of Human Choriocarcinoma Cells. In: Soares
M, Hunt J, editors. Placenta and Trophoblast: Humana Press. pp. 229-239.
111. Himeno E, Tanaka S, Kunath T (2008) Isolation and manipulation of mouse trophoblast
stem cells. Curr Protoc Stem Cell Biol Chapter 1: Unit 1E 4.
112. Hohn HP, Linke M, Ugele B, Denker HW (1998) Differentiation markers and
invasiveness: discordant regulation in normal trophoblast and choriocarcinoma cells.
Exp Cell Res 244: 249-258.
113. Chen Q, Stone PR, McCowan LM, Chamley LW (2005) Interaction of Jar
choriocarcinoma cells with endothelial cell monolayers. Placenta 26: 617-625.

75
114. Aldo PB, Krikun G, Visintin I, Lockwood C, Romero R, et al. (2007) A novel three-
dimensional in vitro system to study trophoblast-endothelium cell interactions. Am J
Reprod Immunol 58: 98-110.
115. Muntener M, Hsu YC (1977) Development of trophoblast and placenta of the mouse. A
reinvestigation with regard to the in vitro culture of mouse trophoblast and placenta.
Acta Anat (Basel) 98: 241-252.
116. Uy GD, Downs KM, Gardner RL (2002) Inhibition of trophoblast stem cell potential in
chorionic ectoderm coincides with occlusion of the ectoplacental cavity in the mouse.
Development 129: 3913-3924.
117. Damsky CH, Fitzgerald ML, Fisher SJ (1992) Distribution patterns of extracellular
matrix components and adhesion receptors are intricately modulated during first
trimester cytotrophoblast differentiation along the invasive pathway, in vivo. J Clin
Invest 89: 210-222.
118. Horie K, Fujita J, Takakura K, Kanzaki H, Kaneko Y, et al. (1992) Expression of c-kit
protein during placental development. Biol Reprod 47: 614-620.
119. Mitsunari M, Harada T, Tanikawa M, Iwabe T, Taniguchi F, et al. (1999) The potential
role of stem cell factor and its receptor c-kit in the mouse blastocyst implantation.
Mol Hum Reprod 5: 874-879.
120. Merviel P, Challier JC, Carbillon L, Foidart JM, Uzan S (2001) The role of integrins in
human embryo implantation. Fetal Diagn Ther 16: 364-371.
121. Vatansever HS, Inan VS, Lacin S, Koyuncu F (2003) Immunolocalization of alphaV,
alpha3 and beta1 integrins in the human placenta with pre-eclampsia. Acta Histochem
105: 253-260.
122. Walter-Yohrling J, Morgenbesser S, Rouleau C, Bagley R, Callahan M, et al. (2004)
Murine endothelial cell lines as models of tumor endothelial cells. Clin Cancer Res
10: 2179-2189.
123. Moser M, Binder O, Wu Y, Aitsebaomo J, Ren R, et al. (2003) BMPER, a novel
endothelial cell precursor-derived protein, antagonizes bone morphogenetic protein
signaling and endothelial cell differentiation. Mol Cell Biol 23: 5664-5679.

76
124. Rossant J, Cross JC (2001) Placental development: lessons from mouse mutants. Nat
Rev Genet 2: 538-548.
125. Li JY, Chai BX, Zhang W, Liu YQ, Ammori JB, et al. (2007) Ankyrin repeat and SOCS
box containing protein 4 (Asb-4) interacts with GPS1 (CSN1) and inhibits c-Jun
NH2-terminal kinase activity. Cell Signal 19: 1185-1192.
126. Benezra R, Rafii S, Lyden D (2001) The Id proteins and angiogenesis. Oncogene 20:
8334-8341.
127. Mutter WP, Karumanchi SA (2008) Molecular mechanisms of preeclampsia. Microvasc
Res 75: 1-8.
128. Matsui J, Wakabayashi T, Asada M, Yoshimatsu K, Okada M (2004) Stem cell factor/c-
kit signaling promotes the survival, migration, and capillary tube formation of human
umbilical vein endothelial cells. J Biol Chem 279: 18600-18607.
129. Lyall F, Bulmer JN, Duffie E, Cousins F, Theriault A, et al. (2001) Human trophoblast
invasion and spiral artery transformation: the role of PECAM-1 in normal pregnancy,
preeclampsia, and fetal growth restriction. Am J Pathol 158: 1713-1721.
130. Proll J, Blaschitz A, Hartmann M, Thalhamer J, Dohr G (1997) Cytokeratin 17 as an
immunohistochemical marker for intramural cytotrophoblast in human first trimester
uteroplacental arteries. Cell Tissue Res 288: 335-343.
131. Quenby S, Mountfield S, Cartwright JE, Whitley GS, Chamley L, et al. (2005)
Antiphospholipid antibodies prevent extravillous trophoblast differentiation. Fertil
Steril 83: 691-698.
132. Xie Y, Abdallah ME, Awonuga AO, Slater JA, Puscheck EE, et al. (2010)
Benzo(a)pyrene causes PRKAA1/2-dependent ID2 loss in trophoblast stem cells. Mol
Reprod Dev 77: 533-539.
133. Zhong W, Xie Y, Abdallah M, Awonuga AO, Slater JA, et al. (2010) Cellular stress
causes reversible, PRKAA1/2-, and proteasome-dependent ID2 protein loss in
trophoblast stem cells. Reproduction 140: 921-930.

77
134. Marchand M, Horcajadas JA, Esteban FJ, McElroy SL, Fisher SJ, et al. (2011)
Transcriptomic signature of trophoblast differentiation in a human embryonic stem
cell model. Biol Reprod 84: 1258-1271.
135. Wolfe MW (2006) Culture and transfection of human choriocarcinoma cells. Methods
Mol Med 121: 229-239.
136. Lasorella A, Iavarone A (2006) The protein ENH is a cytoplasmic sequestration factor
for Id2 in normal and tumor cells from the nervous system. Proc Natl Acad Sci U S A
103: 4976-4981.
137. Wang S, Sdrulla A, Johnson JE, Yokota Y, Barres BA (2001) A role for the helix-loop-
helix protein Id2 in the control of oligodendrocyte development. Neuron 29: 603-614.
138. Kim NS, Kim HT, Kwon MC, Choi SW, Kim YY, et al. (2011) Survival and
differentiation of mammary epithelial cells in mammary gland development require
nuclear retention of Id2 due to RANK signaling. Mol Cell Biol 31: 4775-4788.
139. Rollin J, Blechet C, Regina S, Tenenhaus A, Guyetant S, et al. (2009) The intracellular
localization of ID2 expression has a predictive value in non small cell lung cancer.
PLoS One 4: e4158.
140. Stighall M, Manetopoulos C, Axelson H, Landberg G (2005) High ID2 protein
expression correlates with a favourable prognosis in patients with primary breast
cancer and reduces cellular invasiveness of breast cancer cells. Int J Cancer 115: 403-
411.
141. Almeida A, Bolanos JP, Moncada S (2010) E3 ubiquitin ligase APC/C-Cdh1 accounts
for the Warburg effect by linking glycolysis to cell proliferation. Proc Natl Acad Sci
U S A 107: 738-741.
142. Rao KP, Belogolovkin V, Yankowitz J, Spinnato JA, 2nd (2012) Abnormal
placentation: evidence-based diagnosis and management of placenta previa, placenta
accreta, and vasa previa. Obstet Gynecol Surv 67: 503-519.
143. Dashe JS, McIntire DD, Ramus RM, Santos-Ramos R, Twickler DM (2002) Persistence
of placenta previa according to gestational age at ultrasound detection. Obstet
Gynecol 99: 692-697.

78
144. Dimes Mo Long-term health effects of premature birth.
145. Wortman AC, Alexander JM (2013) Placenta accreta, increta, and percreta. Obstet
Gynecol Clin North Am 40: 137-154.
146. Norwitz ER (2006) Defective implantation and placentation: laying the blueprint for
pregnancy complications. Reprod Biomed Online 13: 591-599.
147. Krussel JS, Bielfeld P, Polan ML, Simon C (2003) Regulation of embryonic
implantation. Eur J Obstet Gynecol Reprod Biol 110 Suppl 1: S2-9.




















