
Evidence for functionally important adenosine A2a receptors in the rat hippocampus

Epilepsy Research 65 (2005) 1–9
Research paper
The role of adenosine A1 receptors in the interaction betweenamygdala and entorhinal cortex of kindled rats
Mohammad Mohammad-Zadeh, Azam Amini, Javad Mirnajafi-Zadeh∗,Yaghoub Fathollahi
Department of Physiology, School of Medical Sciences, Tarbiat Modarres University, P.O. Box 14115-111, Tehran, Islamic Republic of Iran
Received 1 August 2004; received in revised form 18 February 2005; accepted 4 March 2005Available online 7 July 2005
Abstract
In this study the effect of adenosine A1 receptors of the entorhinal cortex (EC) and amygdala on kindled seizures wasinvestigated. Animals were kindled by daily electrical stimulation of amygdala (group 1) or EC (group 2). In the fully kindledanimals,N6-cyclohexyladenosine (CHA), a selective A1 receptor agonist, and 1,3-dimethyl-8-cyclopenthylxanthine (CPT), as seizurep roinjectiono , 15, 60a zure stage.A ateralm n ofC mals,i dledr dleds lah©
K
f
nd
elective A1 receptor antagonist, were microinjected bilaterally into the EC (group 1) or amygdala (group 2). Thearameters were measured at 5, 15, 60 and 120 min post injection. Obtained data showed that in group 1, intra-EC micf CHA at concentration of 10�M reduced amygdala- and, EC-afterdischarge duration and stage 5 seizure duration at 5nd 120 min post drug injection. It also increased the latency to stage 4 seizure but no alteration was observed in seit concentrations of 0.1 and 1�M, CHA reduced only EC-afterdischarge duration at 5 and 15 min post drug infusion. Bilicroinjection CPT at concentrations of 5 and 10�M into the EC did not alter seizure parameters. Intra-EC microinjectioPT (5�M), 5 min before CHA (10�M), blocked the anticonvulsant effects of CHA. On the other hand, in group 2 ani
ntra-amygdala CHA (10, 50 and 100�M) or CPT (5 and 10�M) had no significant effect on seizure parameters of EC-kinats. These results suggest that adenosine A1 receptors activation of the EC may have an inhibitory effect on amygdala-kineizures. But, despite of reciprocal interconnections between these two regions, activation of the A1 receptors of the amygdaas no effect on EC-kindled seizures.2005 Elsevier B.V. All rights reserved.
eywords: Seizure; Adenosine; Amygdala; Entorhinal cortex; Kindling
1. Introduction
Investigation of involvement of amygdala a
∗ Corresponding author. Tel.: +98 21 8011001;ax: +98 21 8006544.E-mail address: [email protected] (J. Mirnajafi-Zadeh).
entorhinal cortex (EC) in the epileptic process is ofclinical importance because EC (Heinemann et al.,1993) together with the amygdala (Bernasconi et al.,
0920-1211/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.eplepsyres.2005.03.012

2 M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9
2003) significantly contribute to epileptogenesis in thepatient with temporal lobe epilepsy. On the other hand,anatomical studies have shown evidence of reciprocalinterconnections between the EC and amygdala andindicated the presence of a monosynaptic pathway fromEC to the amygdala (Funahashi et al., 2000). Otherstudies show the presence of a population of amygdalacells whose axons project to EC (Beckstead, 1978).Therefore, it has been suggested that amygdala and ECcould be important relay points for seizure generationduring kindled seizures elicited from EC or amyg-dala stimulation (Squire and Zola-Morgan, 1991). Inthis regard, it may be postulated that neurotransmit-ters and/or neuromodulators that can affect the neuralactivity of the amygdala or EC, may alter the seizureselicited from the other region, but the precise role ofthese agents remains to be determined.
Among different neuromodulators, adenosine hasbeen shown to have anticonvulsant activity in vari-ous laboratory models of epilepsy including kindling(Barraco et al., 1984; Simonato et al., 1994) and thisactivity is mediated through activation of A1 receptors(Dunwiddie, 1999; Young and Dragunow, 1994; Zhanget al., 1994). The action of adenosine and its A1 recep-tors have been shown in the EC (Rivkees et al., 1995)and, amygdala (Davis et al., 1994). Therefore, accord-i andE aya EC.H eno-s resa fa sti-g eent ffectoc t,a tiveA dleds
2
2
ratsw ark
conditions with ad libitum access to food and water.The rats weighed 300–350 g at the time of surgery.Procedures involving animals and their care were con-ducted in accordance with the NIH guidelines for thecare and use of laboratory animals. All experimentswere carried out at the same time in the morning toavoid the bias of circadian rhythms.
2.2. Surgical and kindling procedure
For stereotaxic surgery, the animals were anes-thetized by ketamine (100 mg/kg, i.p.). Group 1 ani-mals were implanted with bipolar stimulating andmonopolar recording electrodes (twisted into a tripolarconfiguration) terminating in the basolateral amygdala(coordinates:A, −2.5 mm;L, 4.8 and 7.5 mm belowdura) of the right hemisphere. Two 22-gauge guidecannulae were also implanted in the right and left EC(coordinates:A, −6.7 mm;L, ±4.7 and 5.8 mm belowdura) (Paxinos and Watson, 1986). In group 2 animalsthe tripolar electrode was implanted in the right EC(coordinates as above) and two 22-gauge guide can-nulae in the right and left amygdala (coordinates asabove). In both groups, a monopolar recording elec-trode was attached to the guide cannula of the righthemisphere. Electrodes (stainless steel, Teflon-coated,1 u-l erec cor-t
sh-o p 1)o arew erei lsiu pre-v sw ityuw -ADd theo iono
2
I,N rti-
ng to anatomical connections between amygdalaC, activation of A1 receptors in these regions mffect kindled seizures elicited from amygdala orowever, there is not any report about the role of adine A1 receptors of EC on amygdala-kindled seizund vice versa (the role of adenosine A1 receptors omygdala on EC-kindled seizures). Thus, to inveate the role of adenosine in the interaction betw
hese two regions, in this study we examined the ef intra-EC and intra-amygdala microinjection ofN6-yclohexyladenosine, a selective A1 receptor agonisnd 1,3-dimethyl-8-cylclopenthylxanthine, a selec1 receptor antagonist, on amygdala and EC-kineizures.
. Materials and methods
.1. Animals
One hundred and fifteen male Sprague–Dawelyere individually housed under 12-h light/12-h d
27�m in diameter, AM-Systems, USA) were insated except at the tips. Two other electrodes wonnected to skull screws, placed above the leftical surface as earth and differential electrodes.
One week after surgery, afterdischarge (AD) threld was determined in basolateral amygdala (grour EC (group 2) by a 2 s, 60 Hz monophasic squave stimulus of 1 ms per wave. The stimulations w
nitially delivered at 10�A and then at 5-min intervancreasing stimulus intensity in increments of 10�Antil at least 5 s of ADs were recorded as describediously (Mirnajafi-Zadeh et al., 1999). Then, animalere stimulated daily at the AD threshold intensntil five consecutive stage 5 seizures (Racine, 1972)ere elicited. The recorded parameters were: ECuration (EC-ADD), amygdala-ADD, the latency tonset of bilateral forelimb clonus (S4L), the duratf stage 5 (S5D) and the seizure stage (SS).
.3. Drug administration
CHA (Sigma, Pool, Dorset, UK) and CPT (RBatick, MA 01760-2418, USA) were dissolved in a

M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9 3
ficial cerebrospinal fluid [ACSF (in mM): 114 NaCl,1.25 NaH2PO4, 2 MgSO4, 26 NaHCO3, 1 CaCl2 and10 glucose] and pH adjusted to 7.3–7.4 using 1NNaOH. The solutions were sterilized through micro-filters (0.2�m, Minisart, NML, Sartorius, Germany).Drugs were infused (1�l over 2 min) via a 27-gaugecannula, which was 1 mm below the tip of 22-gaugecannula. A different group of animals was used foreach of the drug doses employed and for the differenttime intervals after drug infusion. Most of the animalswere received only one dose of drug and stimulatedat one time. However, some animals received the sec-ond dose. In these cases, the time interval between twodoses was at least 2 weeks.
2.4. Microinjection of CHA or CPT
In fully kindled rats, CHA (0.1, 1, 10�M in group 1and 10, 50, 100�M in group 2) or CPT (5 and 10�M)were microinjected into the EC or amygdala and 5, 15,60 and 120 min later, animals were stimulated at ADthreshold. In each case, 24 h prior to the experiment,animals received ACSF, were stimulated in the sameway and the results recorded as control values.
2.5. Microinjection of CHA with CPT
re-t sti-g andt mina
2
ce-d am-i posi-t ch asl thatp
2
nsS rva-t if-f fter
various doses of drug injections. Comparison of datafrom animals receiving CHA alone with those receiv-ing CHA plus CPT was carried out by Student’st-test.A P value less than 0.05 was considered to represent asignificant difference.
3. Results
All kindled rats responded with stable stage 5seizures in either a noninfusion condition or after ACSFinfusion. At the doses employed, CHA did not exert anynoticeable effect on behavioral or locomotor activity.Histological assessment indicated that the electrodesand the infusion cannulae were positioned in the amyg-dala and the EC (Fig. 1).
3.1. Effects of intra-EC CHA or CPT onamygdala-kindled seizures
The inhibitory effects of CHA on seizure parameterswere evident 5 min after drug infusion, and were stilldetectable after 120 min post infusion. All measuredparameters had returned to normal values (baseline) at24 h post drug administration (not illustrated). Differ-ent doses of CHA (0.1, 1 and 10�M) had not significante posti ledt rgesg mu-l ars ft
HAit tn fts thana Ar t offsi -i td afteri -
The effect of intra-EC or intra-amygdala CPT preatment upon administration of CHA was inveated. CPT was microinjected 5 min before CHA
he animals were stimulated at 5, 15, 60 and 120fter the agonist treatment.
.6. Histology
All the animals were sacrificed at the end of proure. Their brain were removed, sectioned and ex
ned under microscope for electrode and cannulaions and the presence of any tissue damage suesion. In case of any abnormality, the data fromarticular animal was not included in the results.
.7. Statistical analysis
Results obtained are expressed as the mea±.E.M. and accompanied by the number of obse
ions. A two-way ANOVA was done to compare derent groups of animals at different time points a
ffects on seizure stage at different time pointsnfusion. However, CHA administration into the ECo significant decrease of duration of afterdischaenerated from the amygdala after electrical sti
ation (Fig. 2). A two-way ANOVA of resultant datevealed significant effect of dose (P < 0.001) but noignificant effect of time (P = 0.28) or interaction oime× dose (P = 0.25).
EC-ADD was also significantly reduced post Cnfusion at doses of 0.1, 1 and 10�M (Fig. 3). Here,here was a significant effect of dose (P < 0.001) buo significant effect of time (P = 0.6) or interaction o
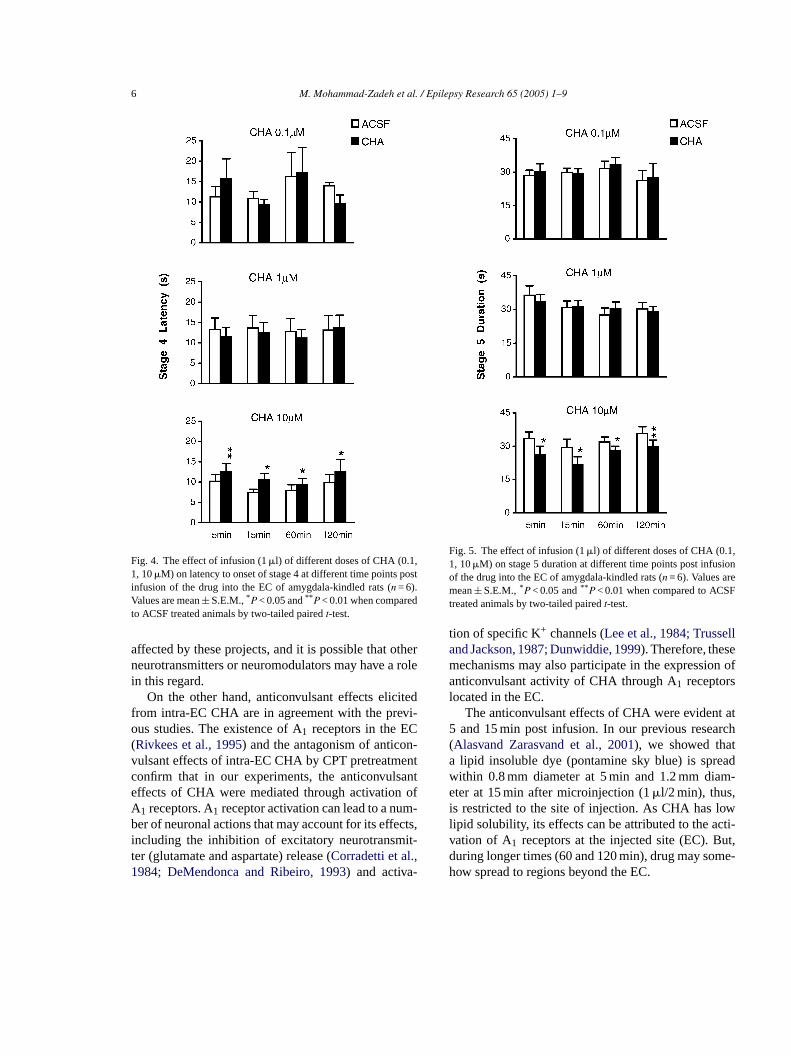
ime× dose (P = 0.61). The comparison ofFigs. 2 and 3hows that the ADs in the EC were more sensitivemygdala ADs. Likewise, intra-EC infusion of CHesulted in the prolongation of latency to the onseorelimb clonus (S4L;Fig. 4). The two-way ANOVAhowed a significant effect of dose (P < 0.0001) andnteraction of time× dose (P < 0.0001) but no signifcant effect of time (P = 0.36). In addition, differenoses of CHA reduced stage 5 seizure duration
ntra-EC administration (Fig. 5) and there were sig

4 M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9
Fig. 1. (a) A typical photomicrograph of a coronal section through the injection site in the left and right amygdala (left) and entorhinal cortex(right). Arrows show the site of dye injection into the amygdala or entorhinal cortex. (b) Schematic drawings of coronal plane through theamygdala (left) and entorhinal cortex (right) has been adapted from the atlas of Paxinos and Watson. Solid dots reveal the location of the cannulatips.
nificant effect of dose (P < 0.001) but no significanteffect of time (P = 0.49) or interaction of time× dose(P = 0.95).
Intra-EC injection of CPT did not lead to any sig-nificant changes in seizure parameters. However, asshown inFig. 6, pretreatment (5 min) of animals withCPT (5�M) significantly attenuated the CHA (10�M)inhibitory effect on seizure activity.
In Figs. 2–5, data showed at each time and each doseof drug are from the same group of rats.
3.2. Effects of intra-amygdala CHA or CPT onEC-kindled seizures
Intra-amygdala microinjection of different doses ofCHA (10, 50 and 100�M) or CPT (5 and 10�M)had no significant effects on seizure parameters (ADD,S4L, S5D and seizure stages) of EC-kindled rats.
4. Discussion
Results of the present study showed that microin-jection of CHA into the EC can exert an antiepilep-tic effect on seizures elicited from the amygdala;but, intra-amygdala CHA has no antiepileptic effectson EC-kindled seizures. CHA is a selective adeno-sine A1 receptor agonist (Jacobson et al., 1992) andthere are several studies showing its anticonvulsanteffects in different models of epilepsy including kin-dling. In most of these studies, the amygdala has beenconsidered as the epileptic focus. It has been shownthat intraperitoneal (Pourgholami et al., 1997a,b;Dunwiddie, 1999), and in situ (Pourgholami et al.,1997a,b; Mirnajafi-Zadeh et al., 1999, 2000; AlasvandZarasvand et al., 2001) administration of adenosineanalogs have anticonvulsant effects on seizures elicitedfrom amygdala.

M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9 5
Fig. 2. The effect of infusion (1�l) of different doses of CHA (0.1, 1,10�M) on amygdala afterdischarge duration (A-ADD) at differenttime points post infusion of the drug into the EC of amygdala-kindledrats (n = 6). Values are mean± S.E.M.,* P < 0.05 when compared toACSF treated animals by two-tailed pairedt-test.
The main anticonvulsant effects of adenosine areexerted through A1 receptors. So, it can be suggestedthat the microinjection of selective A1 receptor agonistsinto the regions containing these receptors can also sup-press kindled seizures elicited from other brain regions.However, results of the present study showed that thisis not true in all situations.
The absence of anticonvulsant effects of intra-amygdala CHA on EC-kindling cannot be attributedto weak A1 receptors activity or low concentrationof these receptors in the amygdala as we previ-ously showed that intra-amygdala microinjection of2-chloroadenosine (a nonselective adenosine receptorsagonist) at the doses as low as 0.25 nM could sig-nificantly reduce amygdala ADD and other seizureparameters in amygdala-kindled rats (Pourgholamiet al., 1997a,b). However, in the present study the ADs
Fig. 3. The effect of infusion (1�l) of different doses of CHA (0.1, 1,10�M) on EC afterdischarge duration (EC-ADD) at different timepoints post infusion of the drug into the EC of amygdala kindledrats (n = 6). Values are mean± S.E.M.,* P < 0.05 when compared toACSF treated animals by two-tailed pairedt-test.
elicited from EC did not significantly change afterintra-amygdala injection of CHA, even at high con-centrations (up to 200�M). It is in consist with ourprevious study showed that intra-amygdala CHA (upto 1 mM) had no significant effect on hippocampal(CA1)-kindled seizure parameters with the exceptionof secondary ADs (Mirnajafi-Zadeh et al., 2000). Thus,despite the inhibitory action of adenosine analogs onamygdala neuronal activity, the degree of inhibition isnot sufficient to reduce the severity of seizures elicitedfrom EC or hippocampus.
Although amygdala A1 receptors activity did notexert any anticonvulsant effect on the EC-kindledseizures, it cannot be concluded that amygdala hasno role in EC-kindling. As mentioned before, some ofthe amygdala neurons have axonal projects to the EC(Beckstead, 1978). Thus, EC neural activity may be
6 M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9
Fig. 4. The effect of infusion (1�l) of different doses of CHA (0.1,1, 10�M) on latency to onset of stage 4 at different time points postinfusion of the drug into the EC of amygdala-kindled rats (n = 6).Values are mean± S.E.M.,* P < 0.05 and** P < 0.01 when comparedto ACSF treated animals by two-tailed pairedt-test.
affected by these projects, and it is possible that otherneurotransmitters or neuromodulators may have a rolein this regard.
On the other hand, anticonvulsant effects elicitedfrom intra-EC CHA are in agreement with the previ-ous studies. The existence of A1 receptors in the EC(Rivkees et al., 1995) and the antagonism of anticon-vulsant effects of intra-EC CHA by CPT pretreatmentconfirm that in our experiments, the anticonvulsanteffects of CHA were mediated through activation ofA1 receptors. A1 receptor activation can lead to a num-ber of neuronal actions that may account for its effects,including the inhibition of excitatory neurotransmit-ter (glutamate and aspartate) release (Corradetti et al.,1984; DeMendonca and Ribeiro, 1993) and activa-
Fig. 5. The effect of infusion (1�l) of different doses of CHA (0.1,1, 10�M) on stage 5 duration at different time points post infusionof the drug into the EC of amygdala-kindled rats (n = 6). Values aremean± S.E.M.,* P < 0.05 and** P < 0.01 when compared to ACSFtreated animals by two-tailed pairedt-test.
tion of specific K+ channels (Lee et al., 1984; Trusselland Jackson, 1987; Dunwiddie, 1999). Therefore, thesemechanisms may also participate in the expression ofanticonvulsant activity of CHA through A1 receptorslocated in the EC.
The anticonvulsant effects of CHA were evident at5 and 15 min post infusion. In our previous research(Alasvand Zarasvand et al., 2001), we showed thata lipid insoluble dye (pontamine sky blue) is spreadwithin 0.8 mm diameter at 5 min and 1.2 mm diam-eter at 15 min after microinjection (1�l/2 min), thus,is restricted to the site of injection. As CHA has lowlipid solubility, its effects can be attributed to the acti-vation of A1 receptors at the injected site (EC). But,during longer times (60 and 120 min), drug may some-how spread to regions beyond the EC.

M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9 7
Fig. 6. Effect of CPT pretreatment on reduction effects of CHAon EC afterdischarge duration (EC-ADD), amygdala afterdischargeduration (A-ADD), and stage 5 duration (S5D) and on incrementeffects of CHA on stage 4 latency (S4L). CPT (5�M) microinjected5 min before CHA (10�M). Values are mean± S.E.M., * P < 0.05using Student’st-test.
On the other hand, no significant effect of timewas observed. Two possibilities may be account forthis observation: first, the CHA has low lipid solu-bility and hence its clearance from the brain is veryslow, and second, the CHA is an adenosine stable ana-logue with high enzymatic resistance (Jacobson et al.,1992). Therefore, the concentration of CHA in theEC may be considered relatively constant at 5, 15,60 and 120 min post-drug infusion, and this is why
the drug effects are similar at different times post-administration.
It has been shown that EC cells, which receive inputsfrom CA1 cells of the hippocampus, form mono- oroligosynaptic contacts with the amygdala cells, thusproviding a link between the hippocampal CA1 cellsand the amygdala. Such a connection between thesetwo structures is of importance, because both are likelyto play a role in seizure initiation and propagation (Wanet al., 1998). However, the comparison of the effect ofintrahippocampal (CA1) with intra-EC microinjectionof CHA on amygdala-kindled seizures shows that inthe hippocampal CA1 region, lower concentration ofCHA (0.1�M) could produce a significant anticonvul-sant effect (Alasvand Zarasvand et al., 2001). It may besomehow related to high concentration of A1 receptorsin the hippocampus (Chen et al., 1992).
In the present study, intra-EC injection of CHA sig-nificantly reduced the duration of ADs recorded fromthe amygdala, in a dose-dependent manner. It meansthat inhibition of neural activity in the EC by CHAcan decrease the neural activity of the amygdala aswell. The S4L was also increased by CHA in a dose-dependent manner. This parameter is an index of gener-alized seizure initiation and its increase reveals that ECmay play a role in spreading of epileptic spikes froma yingo ign ofC in-j citedf rgesr ECn int
uso thati iono zures( ea ofta n-j andr jec-t ree
hasb tivity
mygdala to the other brain region(s) and/or relaf these spikes. Decrease of S5D is also another sHA anticonvulsant action. In addition, CHA micro
ected into the EC could reduce afterdischarges elirom the EC and the amygdala. The EC afterdischaeduction shows the inhibitory effect of CHA on theeuronal activity, which results from the reduction
he severity of amygdala-kindled seizures.In the case of the effect of CHA on the foc
f kindled seizures, our previous study showedn amygdala-kindled seizures, focal microinjectf adenosine agonist suppresses kindled seiPourgholami et al., 1997a,b). In addition, there arlso other studies about the direct inhibitory role
he adenosine agonists on the seizure focus (Rosennd Berman, 1987). Here we used bilateral microi
ection because of interconnections between leftight amygdala and EC. Thus, this manner of inion will inhibit the neural activity of the nucleus moffectively.
On the other hand, in contrast to our results, iteen shown that EC lesion can produce seizure ac

8 M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9
in vitro. Thus, it has been postulated that EC activityhas an inhibitory role in seizure propagation (Collinset al., 1983). However, this seizure activity can proba-bly be attributed to a sudden release of glutamate in thedentate gyrus, which occurs following the EC lesion(Deller and Frotscher, 1997).
In our experiments, CPT could not produce anychange in seizure parameters, which is consistent withour previous works (Mirnajafi-Zadeh et al., 2000;Alasvand Zarasvand et al., 2001). Thus, we cannotconsider any anticonvulsant role for the endogenousadenosine in the EC, although this role has been shownin many other brain areas in vivo (Dragunow, 1988;Dunwiddie, 1999) and in vitro (Kostopoulos et al.,1989).
Therefore, on the basis of obtained results, it maybe concluded that adenosine A1 receptors activation ofthe EC has an inhibitory effect on amygdala-kindledseizures. But, despite of reciprocal interconnectionsbetween these two regions, activation of the A1 recep-tors of the amygdala has no effect on EC-kindledseizures. It means that when amygdala is the epilep-tic focus, any changes in the activity of EC adenosinereceptors can affect the seizure severity. Further work iswarranted to determine the role of A1 receptors of otherbrain structures on different kind of kindled seizures.
R
A aliz-ofpus
B 984.kin-
B rea inadish
B der-em-m-
462–
C nousrs in8.
C onalorhi-
Corradetti, R., Conte, G., Moroni, F., Passani, M.B., Pepeu, G., 1984.Adenosine decreased aspartate and glutamate release from rathippocampal slices. Eur. J. Pharmacol. 104, 19–26.
Davis, M., Rainnie, D., Cassell, M., 1994. Neurotransmission in therat amygdala related to fear and anxiety. Trends Neurosci. 17,208–214.
DeMendonca, A., Ribeiro, J.A., 1993. Adenosine inhibits the NMDAreceptor mediated excitatory postsynaptic potentials in the hip-pocampus. Brain Res. 606, 351–356.
Deller, T., Frotscher, M., 1997. Lesion-induced plasticity of centralneurons: sprouting of single fibers in the rat hippocampus afterunilateral entorhinal cortex lesion. Prog. Neurobiol. 53, 687–727.
Dragunow, M., 1988. Purinergic mechanisms in epilepsy. Prog. Neu-robiol. 31, 85–108.
Dunwiddie, T.W., 1999. Adenosine and suppression of seizures. In:Delgado-Escueta, A.V., Wilson, W.A., Olsen, R.W., Porter, R.J.(Eds.), Jaspers Basic Mechanisms of the Epilepsy, vol. 79. Lip-pincott, Philadelphia, pp. 1001–1010.
Funahashi, M., Matsuo, R., Stewart, M., 2000. Propagation of syn-chronous burst discharges from entorhinal cortex to morpholog-ically and electrophysiologically identified neurons of rat lateralamygdala. Brain Res. 884, 104–115.
Heinemann, U., Zhang, C.L., Eder, C., 1993. Entorhinal cortex–hippocampal interactions in normal and epileptic temporal lobe.Hippocampus 3, 89–97.
Jacobson, K.A., van Galen, P.J., William, M., 1992. Adenosine recep-tors: pharmacology, structure activity relationships and therapeu-tic potential. J. Med. Chem. 35, 407–422.
Kostopoulos, G., Drapeau, C., Avoli, M., Olivier, A., Villemeure,J.G., 1989. Endogenous adenosine can reduce epileptiform activ-
eu-
L lsantsible
M 00.p-858,
M os-2-
x in
P ordi-
P 97a.ro-
259–
P liz-sine, 37–
R icaluro-
eferences
lasvand Zarasvand, M., Mirnajafi-Zadeh, J., Fathollahi, Y., Pvan, M.R., 2001. Anticonvulsant effect of bilateral injectionN6-cyclohexyladenosine into the CA1 region of the hippocamin amygdala-kindled rats. Epilepsy Res. 47, 141–149.
arraco, R.A., Swanson, T.H., Phillis, J.W., Berman, R.F., 1Anticonvulsant effect of adenosine analogues on amygdaloiddled seizures in rat. Neurosci. Lett. 46, 317–322.
eckstead, R.M., 1978. Afferent connections of the entorhinal athe rat demonstrated by retrograde cell-labeling with horserperoxidase. Brain Res. 152, 249–264.
ernasconi, N., Bernasconi, A., Caramanos, Z., Antel, S.B., Anmann, F., Arnold, D.L., 2003. Mesial temporal damage in tporal lobe epilepsy: a volumetric MRI study of the hippocapus, amygdala and parahippocampal region. Brain 126,469.
hen, Y., Graham, D.I., Stone, T.W., 1992. Release of endogeadenosine and its metabolic by the action of NMDA receptothe rat hippocampus in vivo. Br. J. Pharmacol. 106, 632–63
ollins, R.C., Tearse, R.G., Lothman, E.W., 1983. Functianatomy of limbic seizures: focal discharges from medial entnal cortex in rat. Brain Res. 280, 25–40.
ity in the human epileptogenic cortex maintained in vitro. Nrosci. Lett. 106, 119–124.
ee, K.S., Schubert, P., Heinemann, U., 1984. The anticonvuaction of adenosine: a postsynaptic dendritic action by a posendogenous anticonvulsant. Brain Res. 321, 160–164.
irnajafi-Zadeh, J., Fathollahi, Y., Pourgholami, M.H., 20Intraperitoneal and intraamygdalaN6-cyclohexyladenosine supress hippocampal kindled seizures in rats. Brain Res.48–54.
irnajafi-Zadeh, J., Pourgholami, M.H., Palizvan, M.R., Rtampour, M., Fallahi, M., 1999. Anticonvulsant action ofchloroadenosine injected focally into the perirhinal corteamygdaloid kindled rats. Epilepsy Res. 37, 37–43.
axinos, G., Watson, C., 1986. The Rat Brain in Stereotaxic Conates. Academic Press, New York.
ourgholami, M.H., Mirnajafi-Zadeh, J., Behzadi, J., 19Effect of intraperitoneal and intrahippocampal (CA1) 2-chloadenosine in amygdaloid kindled rats. Brain Res. 751,264.
ourgholami, M.H., Rostampour, M., Mirnajafi-Zadeh, J., Pavan, M.R., 1997b. Intraamygdala infusion of 2-chloroadenosuppresses amygdala kindled-seizures. Brain Res. 77542.
acine, R.J., 1972. Modification of seizure activity by electrstimulation. 2. Motor seizure. Electroencephalogr. Clin. Nephysiol. 32, 281–294.

M. Mohammad-Zadeh et al. / Epilepsy Research 65 (2005) 1–9 9
Rivkees, S.A., Price, S.L., Zhou, F.C., 1995. Immunohistochemicaldetection of A1 receptors in rat brain with emphasis on localiza-tion in the hippocampal formation, cerebral cortex, cerebellumand basal ganglia. Brain Res. 677, 193–203.
Rosen, J.B., Berman, R.F., 1987. Differential effects of adenosineanalogs on amygdala, hippocampus, and caudate nucleus kindledseizures. Epilepsia 28, 658–666.
Simonato, M.K., Varani, K., Muzzolini, A., Bianchi, C., Beani, L.,Borea, P.A., 1994. Adenosine A1 receptors in the rat brain in thekindling model of epilepsy. Eur. J. Pharmacol. 265, 121–124.
Squire, L.R., Zola-Morgan, S., 1991. The medial temporal system.Science 253, 1380–1386.
Trussell, L.O., Jackson, M.B., 1987. Dependence of an adenosine-activated potassium current on a GTP binding protein in mam-malian central neurons. J. Neurosci. 7, 3306–3316.
Wan, R.Q., Noguera, E.C., Weiss, S.R., 1998. Anticonvulsant effectsof intrahippocampal injection of TRH in amygdala kindled rats.Neuroreport 9, 677–682.
Young, D., Dragunow, M., 1994. Status epilepticus may be causedby loss of adenosine anticonvulsant mechanisms. Neuroscience58, 245–261.
Zhang, G., Franklin, P.H., Murray, T.F., 1994. Activation of adeno-sine A1 receptors underlies anticonvulsant effect of CGS 21680.Eur. J. Pharmacol. 255, 239–243.
Copyright © 2022 FDOKUMEN




















