![Aqua(4,4'-bipyridine-[kappa]N)bis(1,4-dioxo-1 ... - ScienceOpen](https://static.fdokumen.com/doc/165x107/63262349e491bcb36c0aa51f/aqua44-bipyridine-kappanbis14-dioxo-1-scienceopen.jpg)
Aqua(4,4'-bipyridine-[kappa]N)bis(1,4-dioxo-1 ... - ScienceOpen
Upload
independentCategory
view
1download
0
© The Author 2012. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
The resveratrol analog 4,4′-dihydroxy-trans-stilbene suppresses transformation in normal mouse fibroblasts and inhibits proliferation and invasion of human breast cancer cells
Cristina Maccario1,†, Monica Savio1,†, Daniela Ferraro1, Livia Bianchi1, Roberto Pizzala1,*, Luca Pretali2, Luca Forti3 and Lucia Anna Stivala1 1Dipartimento di Medicina Molecolare, 2 Dipartimento di Chimica, Università degli Studi di Pavia, Pavia, PV 27100, Italia and 3 Dipartimento di Chimica, Università di Modena e Reggio Emilia, Modena, MO 41100, Italia
*To whom correspondence should be addressed. Tel: +39 0382 986472; Fax: +39 0382 986893; Email: [email protected]
4,4′-dihydroxy-trans-stilbene (DHS) is a synthetic analog of res-veratrol, a phytoalexin known for its biological activities. We pre-viously demonstrated that DHS exerts an antiproliferative effect on normal human fibroblasts that is higher than that of the natu-ral parent molecule. No evidence regarding its role in human can-cer cell lines has been found thus far. In this study, we investigated the effects of DHS both on chemical-induced transformation of BALB/c 3T3 mouse fibroblasts and on the proliferation and inva-sion of human breast cancer MCF-7 cells. The results showed that DHS markedly suppresses the two-stage (3-methylcholanthrene plus 12-O-tetradecanoylphorbol-13-acetate) cell transfor-mation. Compared with resveratrol, DHS inhibited both anchorage-dependent and -independent MCF-7 growth more effi-ciently. In addition, a reduction in the number of cells in S-phase, characterized by a concomitant increase in the levels of p21 and p53 proteins, together with a strong inhibition of pRb protein phos-phorylation, was observed in DHS-treated cells. Furthermore, DHS effected a strong reduction in matrix metalloproteinase-2 and -9 activities, concomitantly with a marked inhibition of cell adhesion to the extracellular matrix components as well as inhibi-tion of cell migration and invasion. Importantly, modulation of the adhesion molecule E-cadherin was also found in DHS-treated cells. Taken together, these results demonstrate that the two 4,4′-hydroxyl groups on the stilbenic backbone make DHS a more active molecule than resveratrol in inhibiting neoplastic transfor-mation, cancer cell proliferation and invasion. In conclusion, this study suggests that DHS could be a promising anticancer agent.
Introduction
In recent years, many natural products have been investigated for their pharmacological potential, both in term of disease prevention and treat-ment. Resveratrol, i.e. 3,4′,5-trihydroxy-trans-stilbene (RSV), a major active component of the stilbene phytoalexins present in grapes, nuts and other plants, is one remarkable example (1). This molecule has received wide attention for its role in preventing or delaying several human patho-logical processes, including inflammation, cardiovascular diseases and cancer (2–10). In addition, due to its simple structure, chemical com-position and low molecular weight, RSV has multiple molecular targets, thereby accounting for its diverse biological effects as an antioxidant, cyclooxygenase inhibitor and an antiproliferative molecule (11–15). RSV was also shown to inhibit 12-O-tetradecanoylphorbol-13-acetate
(TPA)-induced tumorigenesis in a mouse skin cancer model and to suppress TPA-induced anchorage-independent neoplastic transform-ation in mouse epidermal JB6 P+ cells (8,16–18). In addition, it has been well documented that RSV blocks tumor development at differ-ent steps, including migration, invasion and matrix metalloproteinase (MMP) activity in several tumor cell types (19–21). We have previously demonstrated the relationship between the chemical structure and bio-logical activity of RSV, with the 4′-hydroxystyryl moiety being abso-lutely required for its antiproliferative activity and DNA polymerase inhibition (22). To enhance these effects, we modified the molecule by introducing two hydroxyl groups in the 4 and 4′ positions, resulting in the RSV analog, 4,4′-dihydroxy-trans-stilbene (DHS). Antioxidant and antiestrogenic activities of DHS have already been investigated. In particular, it has been demonstrated that DHS acts as an effective anti-oxidant against linoleic acid peroxidation and free-radical-induced per-oxidation of rat liver microsomes and of human low-density lipoprotein (23–25). More recently, some studies showed that this compound acts as a specific estrogen receptor (ER) ligand with a binding affinity much higher than that of other well-known estrogen receptor agonists (26,27). In our earlier study, we demonstrated that this molecule inhibited the cell proliferation of normal human fibroblasts with higher efficiency than RSV. This enhanced activity of DHS has been linked to the pres-ence of additional molecular targets in its mechanism of action (28). On the contrary, no evidence is available, till date, on the effects of DHS on chemical-induced cell transformation, as well as on the proliferation and invasion of human cancer cells. In this study, we have determined the efficiency of DHS as a potential anticancer agent by assessing its inhibitory potential against chemical-induced neoplastic transformation in BALB/c 3T3 mouse cells. We have subsequently investigated the ability of DHS to inhibit tumor cell proliferation and invasion by using human breast carcinoma MCF-7 cells. Finally, to explore the molecu-lar mechanism of DHS on in vitro cancer cell behavior, we evaluated its effect on the levels or activities of proteins closely related to pro-gression and invasion of cancer cells. We provide evidence that DHS is more efficient than RSV in suppressing cell transformation, as well as cancer cell proliferation and invasion. The mechanism underlying the anticancer activity of DHS appears to be related to the activation of the p53-dependent checkpoint, to the increasing levels of the E-cadherin adhesion molecule and to the reduction in MMP-2 and -9 activities.
Materials and methods
ReagentsRSV and all other chemicals of reagent grade were obtained from Sigma (St. Louis, MO), if not otherwise specified. DHS was synthesized by Perkin con-densation (29).
Cell cultures and treatmentsPrimary mouse epidermal BALB/c 3T3 clone A31-1-1 cells (ATCC, Manassas, VA) and human breast adenocarcinoma MCF-7 cells were cul-tured in Dulbecco’s modified Eagle’s medium and Eagle’s minimal essen-tial medium (Life Technologies Invitrogen, Grand Island, NY), respectively. Both culture media were supplemented with 10% fetal bovine serum (FBS), 100 IU/ml penicillin, 100 µg/ml streptomycin and 1 mM l-glutamine. The cells were treated with DHS or RSV at concentrations ranging from 1 to 90 µM for different incubation periods when approximately 70% conflu-ent. Both compounds were dissolved in dimethyl sulfoxide, without exceed-ing the 0.15% v/v final concentration for dimethyl sulfoxide in the culture media.
Cytotoxicity assaysCell growth was determined by the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay (30). Cell death was assessed by Trypan Blue staining. BALB/c 3T3 and MCF-7 cells, seeded at the density of 1 × 105
Abbreviations: BrdU, 5-bromo-2′-deoxyuridine; CDK, cyclin-dependent kinase; DHS, 4,4′-dihydroxy-trans-stilbene; FBS, fetal bovine serum; HUVEC, human umbilical vein endothelial cells; LSD, least significant difference; MCA, 3-methylcholanthrene; MMP, matrix metalloproteinase; MTT, [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide]; PBS, phosphate-buffered saline; RSV, resveratrol; SDS, sodium dodecyl sulphate; TPA, 1-2-Otetradecanoylphorbol-13-acetate.
† These authors contributed equally to this work.
Carcinogenesis vol.33 no.11 pp.2172–2180, 2012doi:10.1093/carcin/bgs244Advance Access publication July 25, 2012
2172
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

cells/ml in a 24-well plate, were cultured for 24 h, then treated for another 24 h with each compound at the following concentrations: 1, 2.5, 5, 7.5, 10, 12.5, 15, 30, 60 and 90 µM. After trypsinization, 50 µl of cell suspen-sion in phosphate-buffered saline (PBS) was mixed with 50 µl Trypan Blue and incubated for 3 min at room temperature. Unstained (viable) and stained (non-viable) cells were counted in a Bürker chamber, and the results were expressed as a percentage of non-viable versus the total number of counted cells.
Cell transformation assayBALB/c 3T3 cells seeded in 25-cm2 flasks were treated with a sublethal dose (0.5 µg/ml) of the tumor initiator 3-methylcholanthrene (MCA) and subse-quently exposed to the tumor promoter TPA (100 ng/ml) for 2 weeks; the cells were then maintained in normal medium for another week. DHS (2.5 µM) or RSV (10 µM) was added in the culture medium 24 h before the MCA treatment, and the cells were incubated in these media for the following 72 h. The cells were then trypsinized and processed according to the manufacturer’s instruc-tions for the CytoSelect 96-well cell transformation assay (Cell Biolabs, San Diego, CA). The fluorescence proportional to the number and size of BALB/c 3T3 colonies was read using a 96-well fluorometer (Victor-1420 Multilabel readers, PerkinElmer, Waltham, MA) with a filter of 485/535 nm and 0.1 s of exposition time. The data were expressed in relative fluorescence units as a measure of DNA content.
Cell cycle analysisMCF-7 cells, seeded at a density of 1 × 106 in 75-cm2 flasks, were treated for 24 h with 10, 30, 60 and 90 µM DHS or RSV. For the 5-bromo-2′-deoxyuridine (BrdU) immunofluorescence assay, the cells were incubated with 30 µM BrdU during the last hour of treatment. All samples were then harvested, washed in PBS, fixed in 70% ethanol for at least 2 h at −20°C and processed as reported previously (28). Proteins involved in cell cycle progression were investigated by western blotting as described previously (28). Membranes were probed with anti-p21 (CP74 Neomarkers, Thermo Fisher Scientific, Kalamazoo, MI), anti-p53 (DAKO, Carpinteria, CA), anti-pRb (BD Biosciences, Sparks, MD), anti-phospho pRb (BD Biosciences) and anti-actin antibodies.
Clonogenic efficiency and soft agar assaysTwo hundred MCF-7 cells were seeded in 3-cm dishes and left to adhere for 24 h. DHS or RSV at concentrations of 1, 5, 15, 30 or 90 µM were added to the medium for a 24-h incubation. At the end of the treatment, the cells were washed with PBS, and 3 ml of complete medium was added to each dish before proceeding as described previously (28). To test anchorage-independent growth, MCF-7 cells were trypsinized and resuspended to obtain a density of 2 × 104 cells in 500 µl of Dulbecco’s modified Eagle’s medium containing 20% FBS. Increasing concentrations of DHS or RSV (10, 30, 60 and 90 µM) were added directly to the cell suspension together with 500 µl of 0.33% Bacto Agar (Difco Laboratories, Detroit, MI). The mixtures were poured quickly in culture cell dishes prepared in advance with 5 ml of 0.6% Bacto agar in Dulbecco’s modified Eagle’s medium containing 10% FBS and incubated at 37°C for 3 weeks. The colonies formed were photographed and counted using a ×20 magnitude inverted microscope (Leitz DM-IL, Leica, Wetzlar, DE).
Collagen type I adhesion assayA 96-well plate was coated with 50 µl/well of collagen type I diluted in PBS (10 µg/ml), incubated overnight at 4°C and then conditioned with 150 µl/well of 1% BSA in PBS for 30 min at room temperature to block non-specific sites. MCF-7, untreated or treated for 24 h with 10, 30, 60 or 90 µM DHS or RSV, were trypsinized, washed with PBS, plated into the coated wells (3.5 × 104 cells/well) and allowed to attach at 37°C for 55 min. Non-adherent cells were removed by washing with PBS, and the adherent cells were fixed in 96% etha-nol. After staining with 0.1% (w/v) crystal violet, the fixed cells were lysed in 0.2% Triton X-100, and the extracted dye was measured spectrophoto-metrically at 570 nm (Sunrise A-4082, TECAN, Männedorf, CH). E-cadherin protein levels were measured by western blotting using an anti-E-cadherin antibody (Abcam, Cambridge, MA).
Wound-healing assayFor cell motility assays, wound healing was performed using the Ibidi Culture-Insert (Madison, WI) according to the manufacturer’s instructions. Briefly, after trypsinization, 70 µl of MCF-7 cell suspension (5 × 105 cells) was applied to each culture-insert, allowing the cells to grow in the designated areas until a confluent layer was formed at 37°C. Thereafter, the culture-insert was removed, creating a cell-free gap of 500 µm. The cells were washed twice with PBS and treated with DHS or RSV (10, 30, 60 and 90 µM). Wound closure was monitored and photographed at 0, 24, 48, 72 and 96 h with an inverted light microscope equipped with a Canon A590 IS camera (Tokyo, JP). To quantify the migration area, pictures of the initial wounded monolayer were
compared with pictures of the wound area at different time points using the Wimasis web-based image analysis software (Wimasis, Munchen, Germany, http://www.wimasis.com).
Boyden chamber migration and invasion assayThe Boyden chamber (Neuroprobe, Gaithersburg, MD) was assem-bled by inserting collagen- (100 µg/ml) or Matrigel (200 µg/ml, BD Biosciences)-coated filters for determining cell migration or invasion, respec-tively. MCF-7 cells were treated with increasing concentrations of DHS or RSV (10, 30, 60 or 90 µM). Similar treatments, but in the presence of TPA (200 nM), were performed for the invasion assay to increase their potential invasiveness. After 24 h, the cells were trypsinized, resuspended (1 × 105 cells/ml) in serum-free medium, and 50 µl of the suspension was placed in each upper well of the Boyden chamber. Serum-free medium or medium containing 10% FBS was added to the bottom wells as a negative control or chemoattractant, respectively. After 4 h at 37°C, the filters were stained with a Diff-Quick staining kit (Medion Diagnostics, Düdingen, CH) and placed on a microscope slide. Non-migrated cells were wiped off from the filter with a cot-ton swab and mounted on a cover slip with dibutyl phthalate xylene -mounting medium (BDH, Poole, UK). Cell migration and invasion were evaluated by counting the migrated cells under a ×10 magnitude inverted light microscope.
Gelatin zymographyThe activity of MMP-2 and MMP-9 were measured by gelatin zymography as reported previously (31). Briefly, MCF-7 cells were treated with DHS or RSV (10, 30, 60 or 90 µM) in the presence of TPA (200 nM) as an MMP inducer. After 24 h, the conditioned culture medium was collected, centrifuged, mixed with equal amounts of sodium dodecyl sulphate (SDS) sample buffer and loaded on a 10% polyacrylamide gel containing 1 mg/ml gelatin as the protease substrate. After staining with Coomassie Blue R-250 (0.5%w/v) and subsequent destain-ing, the areas of protease activity appeared as horizontal white bands on a dark blue background wherever the proteases had digested the substrate. Gel images of the bands were acquired by a Canon Canoscan 9950F transparencies scanner, and densitometric analysis was performed using the public domain NIH Image program (developed at the United States National Institutes of Health and avail-able on the Internet at http://rsb.info.nih.gov/nih-image/) with the aid of bundled gel plotting macros.
Statistical analysisData are presented as means ± SD of at least three independent experiments. Because data were represented by multiple comparisons with control and planned different treatments, analysis of variance statistic followed by the Least Significant Difference (LSD) posthoc tests was carried out. *P values ≤ 0.05 and **P values ≤ 0.01 were considered to be statistically significant when compared with control cells; °P ≤ 0.05 and °°P ≤ 0.01 were considered to be statistically significant when comparing DHS-treated versus RSV-treated cells.
Results
Figure 1A shows the chemical structure of RSV and its synthetic analog, DHS.
DHS is cytotoxic in BALB/c 3T3 and MCF-7 cellsThe effects of DHS on the proliferation of BALB/c 3T3 and MCF-7 cells were determined using the MTT-based colorimetric assay after 24 h of treatment with increasing concentrations (in the 1–90 µM range) of DHS or RSV. In BALB/c 3T3 cells, RSV did not significantly affect cell growth or survival at concentrations up to 10 µM, whereas a dose-dependent inhibitory activity was detected at higher concentrations, with 80% of cell growth inhibition being observed at 90 µM of RSV. In DHS-treated cells, all concentrations caused significant growth inhibition, from the lowest (25% at 1 µM) to the highest concentration (52% at 90 µM) (Figure 1B). Trypan Blue staining was also performed to verify whether the observed cell growth inhibition was related to cell death. The results obtained confirmed a linear and dose-dependent increase of cell death after treatment with RSV concentrations in the range of 15–90 µM, resulting in 65% cell death at the maximum concentration, relative to the control. After DHS treatment, 33% of mortality was detected at 5 µM, and it plateaued at higher concentrations of DHS (Figure 1C). In MCF-7 cells, a similar growth inhibition trend was evident, with weak activity observed at a concentration as high as 60 µM for RSV and at 15 µM for DHS (Figure 1D). In
DHS inhibits cell transformation, proliferation and invasion
2173
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

particular, cell growth inhibitions of 30 and 40% were observed after treatment with 60 and 90 µM RSV, respectively. Compared with RSV, DHS appeared to be more potent, achieving 32 and 50% of growth inhibition at 15 and 90 µM, respectively. In contrast with mouse fibroblast data, no appreciable cell death was detected in MCF-7 cells in the Trypan Blue assay at the concentrations that affected MTT values (Figure 1E).
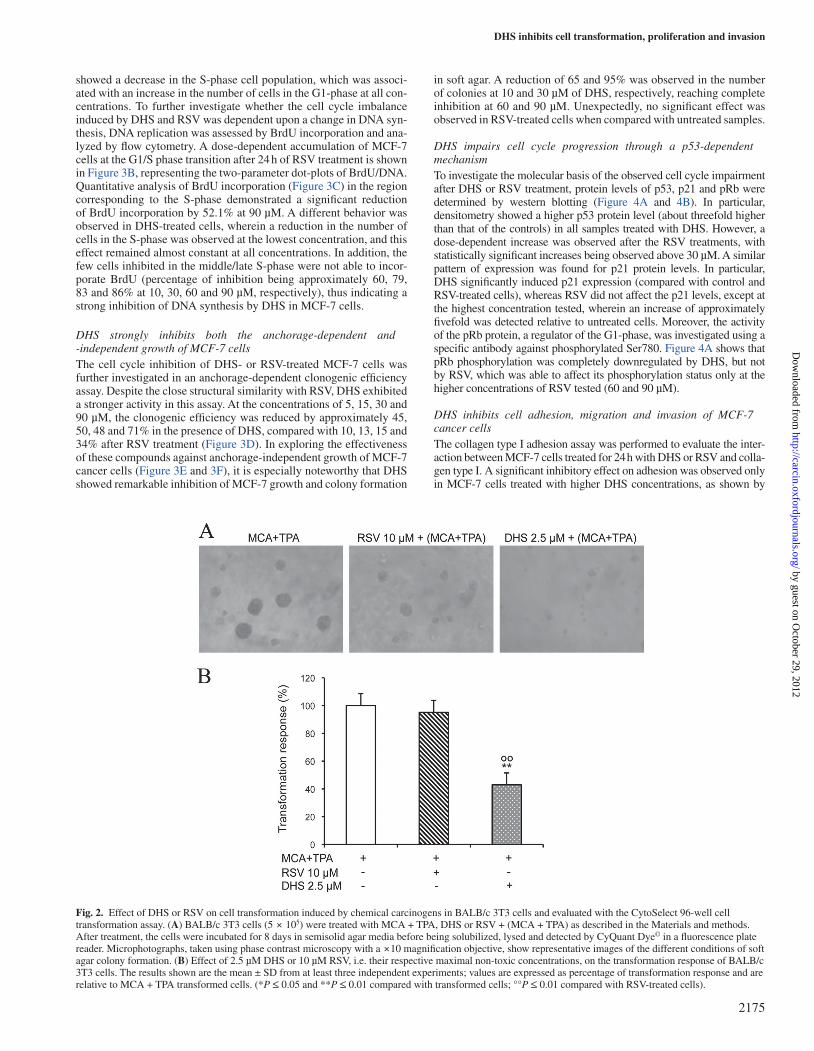
DHS inhibits chemical-induced cell transformation of mouse BALB/c 3T3 cellsIn vitro cell transformation was performed to check the ability of DHS or RSV to interfere with chemical-induced cell transformation in the process of tumor formation. The effects of the two compounds on the transformation response were evaluated by treating cells with 2.5 µM
DHS or 10 µM RSV (i.e. at their respective maximum non-toxic con-centrations) for 96 h. As shown in Figure 2A and 2B, the results dem-onstrated a statistically significant reduction (by approximately 50%) of the transformation response in the presence of DHS in comparison with both MCA + TPA (control) transformation and RSV treatment.
DHS decreases MCF-7 cell proliferationFor a better understanding of the effects of DHS or RSV on tumor cell proliferation, the distribution of MCF-7 cells in each phase of the cell cycle was analyzed by determining the DNA content by flow cytom-etry. As shown in Figure 3A, RSV-treated cells significantly accumu-lated in the S-phase, with a consequent reduction in the G1-phase only at the higher concentrations, as compared with the cell cycle distribu-tion of the control cells. Conversely, under the same conditions, DHS
Fig. 1. (A) Chemical structures of RSV and its synthetic analog, DHS. (B,D) The cell growth was determined by the MTT assay after 24 h of treatment with different concentrations of DHS or RSV (B: 1, 2.5, 5, 7.5, 10, 12.5, 15, 30, 60 and 90 µM in BALB/c 3T3 cells; D: 1, 2.5, 5, 7.5, 15, 30, 60 and 90 µM in MCF-7 cells). The reduction of MTT by active mitochondria, which results in a color change measured at 570 nm with a microplate reader, was used as an indicator of viable cell number. (C,E) Cell death was determined by the Trypan Blue staining. BALB/c 3T3 and MCF-7 cells were treated for 24 h with different concentrations of DHS or RSV (C: 1, 2.5, 5, 7.5, 10, 12.5, 15, 30, 60 and 90 µM in BALB/c 3T3 cells; E: 10, 30, 60 and 90 µM in MCF-7 cells). Unstained (viable) and stained (non-viable) cells were counted in a Bürker chamber, and the results were expressed as a percentage of non-viable versus the total number of counted cells. Data are the mean ± SD from at least three independent experiments; values are expressed as percentages and are relative to untreated control cells. B chart: **10–90 µM RSV; 1–90 µM DHS. D chart: *30 µM RSV, **60–90 µM RSV; **15–90 µM DHS. C chart: **15–90 µM RSV; **5–90 µM DHS. (*P ≤ 0.05 and **P ≤ 0.01 compared with control cells; °P ≤ 0.05 and °°P ≤ 0.01 compared with RSV-treated cells).
C.Maccario et al.
2174
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from
showed a decrease in the S-phase cell population, which was associ-ated with an increase in the number of cells in the G1-phase at all con-centrations. To further investigate whether the cell cycle imbalance induced by DHS and RSV was dependent upon a change in DNA syn-thesis, DNA replication was assessed by BrdU incorporation and ana-lyzed by flow cytometry. A dose-dependent accumulation of MCF-7 cells at the G1/S phase transition after 24 h of RSV treatment is shown in Figure 3B, representing the two-parameter dot-plots of BrdU/DNA. Quantitative analysis of BrdU incorporation (Figure 3C) in the region corresponding to the S-phase demonstrated a significant reduction of BrdU incorporation by 52.1% at 90 µM. A different behavior was observed in DHS-treated cells, wherein a reduction in the number of cells in the S-phase was observed at the lowest concentration, and this effect remained almost constant at all concentrations. In addition, the few cells inhibited in the middle/late S-phase were not able to incor-porate BrdU (percentage of inhibition being approximately 60, 79, 83 and 86% at 10, 30, 60 and 90 µM, respectively), thus indicating a strong inhibition of DNA synthesis by DHS in MCF-7 cells.
DHS strongly inhibits both the anchorage-dependent and -independent growth of MCF-7 cellsThe cell cycle inhibition of DHS- or RSV-treated MCF-7 cells was further investigated in an anchorage-dependent clonogenic efficiency assay. Despite the close structural similarity with RSV, DHS exhibited a stronger activity in this assay. At the concentrations of 5, 15, 30 and 90 µM, the clonogenic efficiency was reduced by approximately 45, 50, 48 and 71% in the presence of DHS, compared with 10, 13, 15 and 34% after RSV treatment (Figure 3D). In exploring the effectiveness of these compounds against anchorage-independent growth of MCF-7 cancer cells (Figure 3E and 3F), it is especially noteworthy that DHS showed remarkable inhibition of MCF-7 growth and colony formation
in soft agar. A reduction of 65 and 95% was observed in the number of colonies at 10 and 30 µM of DHS, respectively, reaching complete inhibition at 60 and 90 µM. Unexpectedly, no significant effect was observed in RSV-treated cells when compared with untreated samples.
DHS impairs cell cycle progression through a p53-dependent mechanismTo investigate the molecular basis of the observed cell cycle impairment after DHS or RSV treatment, protein levels of p53, p21 and pRb were determined by western blotting (Figure 4A and 4B). In particular, densitometry showed a higher p53 protein level (about threefold higher than that of the controls) in all samples treated with DHS. However, a dose-dependent increase was observed after the RSV treatments, with statistically significant increases being observed above 30 µM. A similar pattern of expression was found for p21 protein levels. In particular, DHS significantly induced p21 expression (compared with control and RSV-treated cells), whereas RSV did not affect the p21 levels, except at the highest concentration tested, wherein an increase of approximately fivefold was detected relative to untreated cells. Moreover, the activity of the pRb protein, a regulator of the G1-phase, was investigated using a specific antibody against phosphorylated Ser780. Figure 4A shows that pRb phosphorylation was completely downregulated by DHS, but not by RSV, which was able to affect its phosphorylation status only at the higher concentrations of RSV tested (60 and 90 µM).
DHS inhibits cell adhesion, migration and invasion of MCF-7 cancer cellsThe collagen type I adhesion assay was performed to evaluate the inter-action between MCF-7 cells treated for 24 h with DHS or RSV and colla-gen type I. A significant inhibitory effect on adhesion was observed only in MCF-7 cells treated with higher DHS concentrations, as shown by
Fig. 2. Effect of DHS or RSV on cell transformation induced by chemical carcinogens in BALB/c 3T3 cells and evaluated with the CytoSelect 96-well cell transformation assay. (A) BALB/c 3T3 cells (5 × 105) were treated with MCA + TPA, DHS or RSV + (MCA + TPA) as described in the Materials and methods. After treatment, the cells were incubated for 8 days in semisolid agar media before being solubilized, lysed and detected by CyQuant Dye© in a fluorescence plate reader. Microphotographs, taken using phase contrast microscopy with a ×10 magnification objective, show representative images of the different conditions of soft agar colony formation. (B) Effect of 2.5 µM DHS or 10 µM RSV, i.e. their respective maximal non-toxic concentrations, on the transformation response of BALB/c 3T3 cells. The results shown are the mean ± SD from at least three independent experiments; values are expressed as percentage of transformation response and are relative to MCA + TPA transformed cells. (*P ≤ 0.05 and **P ≤ 0.01 compared with transformed cells; °°P ≤ 0.01 compared with RSV-treated cells).
DHS inhibits cell transformation, proliferation and invasion
2175
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

their limited growth in wells coated with collagen type I (Figure 5A and 5B). In particular, DHS at 60 and 90 µM significantly inhibited MCF-7 cell adhesion by 27 and 40%, respectively. As in the Boyden cham-ber assay, the treatments with 30, 60 and 90 µM of RSV significantly inhibited the migration of MCF-7 cells by approximately 38, 54 and 31%, respectively (Figure 5C). However, DHS showed a more power-ful dose-dependent effect, with reductions of approximately 73, 72, 89 and 91% at 10, 30, 60 and 90 µM, respectively. Figure 5D shows the similarity between the invasiveness of RSV-treated cells and their migra-tory ability. The invasiveness of MCF-7 cells was significantly reduced (by approximately 50%) at the higher concentrations, whereas stronger inhibition of 95% was observed in DHS-treated cells at all concentra-tions. It is important to note that DHS inhibition was, even at the lowest concentration, significantly higher than that observed with RSV. Given
the important role of E-cadherin in the mechanisms of intracellular adhe-sion and extracellular matrix invasion, its protein levels were measured by western blotting in DHS- or RSV-treated MCF-7 cells. As shown in Figure 4C, none of the concentrations of RSV had any significant effect on E-cadherin protein levels, whereas a dose-dependent increase was observed after DHS treatment. In particular, a higher E-cadherin pro-tein level (approximately sevenfold higher than that in control cells) was found even at the lowest concentration of DHS, and the E-cadherin pro-tein levels increased by up to ten times at 90 µM (Figure 4D).
DHS reduces MMP-2 and -9 activitiesTo determine whether MMPs were involved in the inhibition of MCF-7 cell migration and invasiveness induced by DHS, changes in
Fig. 3. Effect of DHS or RSV on cell proliferation of MCF-7 cells after treatment with increasing concentrations of both compounds. (A) Cell cycle phase distribution was evaluated by flow cytometry after staining with PI. Ten thousand cells were analyzed from each sample, and statistical analysis of the mean fluorescence intensity was performed with XL2 software. (B,C) DNA synthesis was analyzed by a biparametric flow cytometry analysis of BrdU incorporation versus DNA content. Representative profiles of cell cycle distribution and the relative quantification of the percentage of BrdU incorporation were calculated in the middle of the S phase. (D) Anchorage-dependent growth was evaluated as cell cloning efficiency. MCF-7 cells were treated with DHS or RSV (1, 5, 15, 30 or 90 µM) for 24 h, and after 15 days, the colonies formed were fixed, stained with crystal violet and counted. (E,F) Anchorage-independent growth was determined in a soft agar assay. Treatments were made directly by adding DHS or RSV to the cell suspension together with the semisolid agar. Representative images of DHS- or RSV-treated MCF-7 colonies in soft agar were taken after 21 days of growth by phase contrast microscopy using a ×20 magnification objective. The number of colonies in soft agar was counted, and relative quantification was performed. The results shown are the means ± SD of average cell counts from three Petri dishes and are representative of three independent experiments. (*P ≤ 0.05 and **P ≤ 0.01 compared with control cells; °P ≤ 0.05 and °°P ≤ 0.01 compared with RSV-treated cells).
C.Maccario et al.
2176
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

their activities were investigated by gelatin zymography. As shown in Figure 6A, RSV did not alter TPA-induced MMP-2 and -9 secre-tion at any of the tested concentrations, whereas treatments with DHS led to a significant inhibition of both MMPs in the 10–90 µM concentration range for MMP-2 and in the range of 60–90 µM for MMP-9. In particular, MMP-2 activity decreased by approximately 20% at 10 µM DHS in comparison to TPA-treated MCF-7 cells, with 49% inhibition being observed at 90 µM of DHS. The activity of MMP-9 decreased only at 60 and 90 µM, with 32 and 50% inhibition, respectively.
DHS induces changes in cell motilityThe effect of DHS or RSV on cell motility was investigated using the wound-healing assay. Figure 6B and 6C show the processed images and their relative quantification. The effect of RSV on the reconstitution of the cell monolayer started to be evident after 48 h and was greater at 96 h, when the gap size was increased in a concentration-dependent manner, with a reduction of wound closure of 90% at 90 µM, relative to the untreated MCF-7 cells. Additionally, cell motility and average cell velocities were dose-dependently reduced with increasing RSV concentrations, as shown in Figure 6B. DHS exhibited a significantly greater effect than RSV in inhibiting MCF-7 cell motility and the average cell velocities, with this inhibition being evident even at the lowest concentration of DHS. More specifically, when the wound was completely closed in untreated cells at 96 h, it remained approximately 80% open for all the DHS concentrations tested.
Discussion
Disruption in normal cell cycle regulation, cell division, cell migration and invasion are important events in cancer development. Targeting these critical events represents the goal of many cancer prevention and therapy regimens through dietary agents, mainly identified in fruits and vegetables (32,33). In this study, we demonstrated that DHS inhibits the transformation of BALB/c 3T3 cells in a multiphase carcinogenesis system. Although DHS was designed and synthesized to enhance the antiproliferative activity of RSV by introducing two hydroxyl groups at the 4 and 4′ positions in the stilbenic backbone, an efficient and interesting inhibition of cell transformation by DHS was observed. This inhibition was effective at a concentration that was low enough to be non-toxic, thus suggesting that DHS could be a good candidate as a potential anticancer agent. No effect was observed in our system after RSV treatment, which was in agreement with the study by Lee and coworkers (18), but in contrast with a previous study demonstrating that RSV suppressed epidermal growth factor- or TPA-induced anchorage-independent mouse cell transformation (17). This discrepancy is probably due to the different carcinogenesis models used (cells, chemicals and procedures). Cell cycle distribution experiments in DHS-treated MCF-7 cells demonstrated a reduction in the number of cells in the S-phase, with a concomitant increase in the number of cells in the G1-phase, whereas RSV-treated samples showed a delay at the beginning of the S-phase. Both compounds were also able to inhibit DNA synthesis, in agreement with the in vitro results reported by Locatelli and coworkers (29). A similar arrest of MCF-7 cells in the S-phase of the cell cycle was previously reported after RSV treatment,
Fig. 4. Effects of 24-h incubation with different concentrations of DHS or RSV on the levels of cell regulatory proteins and the adhesion molecule E-cadherin in MCF-7 cells. (A, C) MCF-7 cells were treated with 10, 30, 60 or 90 µM of DHS or RSV, and these cell lysates were subjected to SDS polyacrylamide gel electrophoresis followed by western blotting. Representative blots of p53, p21, pRb (both total and phosphorylated at Ser780) and E-cadherin are shown. Actin protein was used as loading control. (B,D) Quantification of the p53, p21, pRb (total and phosphorylated at Ser780) and E-cadherin protein was performed by densitometric analysis of the western blot and normalized to the internal loading control. Data are the mean ± SD from at least three independent experiments; values are expressed as percentages and are relative to untreated control cells. (*P ≤ 0.05 and **P ≤ 0.01 compared with control cells; °P ≤ 0.05 and °°P ≤ 0.01 compared with RSV-treated cells).
DHS inhibits cell transformation, proliferation and invasion
2177
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

and this effect appeared to be dose-dependent up to concentrations of 150 µM, at which no significant alteration in cell cycle distribution was observed despite an increased rate of apoptosis (34). Conversely, no significant induction of apoptosis was observed in our experimental conditions (data not shown) for either DHS or RSV, as confirmed by the lack of the sub-G1 peak and the absence of morphological apoptotic features. These data suggest that the antiproliferative effect and the impairment of cell cycle progression mechanisms involve an activation of cell cycle regulatory proteins. In fact, marked increases in p53 and p21 protein levels were detected after treatment with 10 µM DHS, whereas these increases were detected only with 90 µM of RSV. Similarly, cell cycle arrest was reported in normal human fibroblasts, but it was mediated only by DHS, and not by RSV, via the p53/p21 pathway (28). Hsieh and coworkers reported that another RSV derivative, trimethoxy-trans-stilbene, led to an increase in p53 levels, activation of p21 and induction of apoptosis in MCF-7 cells (35). The activation of the p53/p21 pathway by DHS may be linked to DNA damage response. In fact, it has been previously found that RSV and its synthetic analogs show in vitro pro-oxidant and DNA-damaging activities in the presence of Cu(II) ions. Among these analogs, DHS exhibited a higher activity than RSV in damaging the supercoiled DNA of the pBR322 plasmid (36). These findings might be consistent with the activation of p53/p21 by 10–90 µM DHS, but only by 90 µM RSV, in MCF-7 cells observed in this study. These data are also in accordance with the greater degree of oxidation of DNA bases induced by DHS than by RSV, as observed in the Comet test (data not shown). It is known that p21 inhibits the activity of the cyclin/ cyclin-dependent kinase (CDK) complexes, which are implicated in the sequential phosphorylation of pRb. In our experiments, a significant decrease in the hyperphosphorylated form of pRb was observed only at the highest RSV concentration, whereas DHS-treated cells showed low or barely detectable levels of the phosphorylated protein at all DHS concentrations. These results
indicated a progression through the restriction point in the late G1-phase in RSV-treated cells and arrest in the early–middle G1-phase in DHS-treated samples. Similar results were demonstrated by a previous study, in which normal human fibroblasts activated the S-phase checkpoint when treated with RSV, but not with DHS (28). A recent study conducted by Arima and coworkers revealed that pRb also controls cell–cell adhesion in epithelial cells through positive regulation of E-cadherin and that its inactivation plays a role in both the loss of cell proliferation control and tumor progression toward infiltrating malignancy (37). In fact, E-cadherin is frequently down-regulated in several cancers, particularly in breast cancer, and its down-regulation probably facilitates the detachment of cells from the primary tumor, thus promoting metastasis. In our experiments, although RSV was ineffective, a dose-dependent increase in E-cadherin levels was observed after DHS treatment. This result indicates an induced improvement of intercellular adhesion and, consequently, a decrease in cell–extracellular matrix interaction, as assessed by the inhibition of MCF-7 adhesiveness to collagen type I. On the basis of several reports, gelatinases A (MMP-2) and B (MMP-9) are two enzymes associated with an aggressive, invasive or metastatic tumor phenotype. MMP-2 is expressed in breast cancer cells, whereas MMP-9 needs to be induced by treatment with phorbol esters, such as TPA (38). In this case, DHS caused a down-regulation of the activities of both gelatinases, whereas RSV was ineffective. Our data are in agreement with those reported by Woo and coworkers that demonstrated no significant reduction in MMP-2 activity after RSV treatment in cervical cancer Caski cells; additionally, they showed a weak inhibition of MMP-9, albeit at a relatively high concentration (100 µM) (39). Moreover, a similar behavior was detected in the human colon 26 cancer cell line with a more potent inhibition of MMP-9 activity induced by the different dihydroxystilbenes, including DHS, rather than by RSV, suggesting a close structure/activity relationship (40). Finally, in vitro migration and
Fig. 5. Effects of increasing concentrations of DHS or RSV on cell adhesion, migration and invasion in MCF-7 cells after 24 h of incubation. (A) MCF-7 cells were treated for 24 h with 10, 30, 60 or 90 µM of DHS or RSV. The cells were then stained and quantified (B) as described in the Materials and methods. (C, D) Cell migration and invasion were evaluated through the use of the Boyden chamber and filters treated with collagen type I and Matrigel, respectively. Serum-free medium (C−) and medium containing 10% FBS (C) were applied to the bottom wells as negative control or chemoattractant, respectively. Data, obtained by counting the migrated cells under an inverted microscope, are the mean ± SD from at least three independent experiments; values are expressed as percentages and are relative to untreated control cells. (*P ≤ 0.05 and **P ≤ 0.01 compared with control cells; °P ≤ 0.05 and °°P ≤ 0.01 compared with RSV-treated cells).
C.Maccario et al.
2178
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

invasion assays demonstrated that both compounds can significantly reduce the motility of MCF-7 cells, with DHS being the most active molecule. It is important to note that the average velocity of MCF-7 cell motility was RSV dose-dependent and DHS dose-independent, demonstrating maximum inhibition at the lowest concentration of DHS tested. Similar results were obtained for MCF-7 cell migration in the Boyden chamber model. Our findings were also consistent with the data reported by Kimura and coworkers that demonstrated the inhibition of
vascular endothelial growth factor-induced Human Umbilical Vein Endothelial Cells (HUVEC) migration after treatment with different dihydroxystilbenes, including DHS (40). Through the use of the Boyden chamber, it was also possible to assess the ability of the stilbenic compounds to interfere with extracellular matrix overstepping, a process carried out actively by tumor cells that allows them to infiltrate tissues. Treatments with RSV induced a reduction in the tumor cell invasiveness, but only at the higher concentrations, whereas a stronger
Fig. 6. Effects of DHS or RSV on TPA-induced MMP activities and cell motility after the indicated incubation periods. (A) Conditioned media of DHS- or RSV-treated cells were subjected to 10% SDS polyacrylamide gel electrophoresis with 0.1% w/v gelatin. Gelatinolytic activities of MMP-2 and -9 were determined by gelatin zymography as described in the Materials and methods. Data are the mean ± SD from at least three independent experiments; values corresponding to the relative density of the bands are expressed as percentages and are relative to TPA treatment. *P ≤ 0.05 and **P ≤ 0.01 compared with TPA treatment only; °P ≤ 0.05 and °°P ≤ 0.01 compared with RSV-treated cells. (B) Representative processed images (×10 magnification objective) of MCF-7 cells treated with DHS or RSV for the indicated periods. The dashed white lines indicate the original gap of 500 µm, and average cell velocities are given under the photographs. (C) Migration rate quantification of MCF-7 cells; data are the mean ± SD from at least three independent experiments, normalized to 100% wound closure for untreated cells. °10–60 µM DHS at 48 and 72 h, °°10 and 30 µM at 96 h (*P ≤ 0.05 and **P ≤ 0.01 compared with control cells; °P ≤ 0.05 and °°P ≤ 0.01 compared with RSV-treated cells).
DHS inhibits cell transformation, proliferation and invasion
2179
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from

inhibition was observed in DHS-treated cells, independent of the DHS concentration, as reported previously for the MMP activity assay. Furthermore, treatments with DHS were efficient in maintaining the rounded morphology of the migrated and invasive cells, thus reducing their migratory and invasive abilities. In sum, our results indicate that although the only chemical differences between RSV and its analog are the absence of one hydroxyl group and a different position of the second hydroxyl group, these simple structural modifications make DHS a more efficient molecule in inhibiting neoplastic transformation, cancer cell proliferation and invasion. Although part of its mechanism of action is yet to be clarified, DHS is clearly involved with the regulation of proteins implicated in G1 checkpoint activation and in the modulation of cellular adhesion molecules. E-cadherin is one of these cell adhesion molecules, which probably acts in concert with other integrins responsible for cell adhesion to collagen, cell motility and migration, all effects that are inhibited by DHS. Further in vivo studies to better characterize DHS absorption, metabolism, biosafety and the mechanisms of its anticancer activities are in progress.
Funding
MIUR (Ministero dell' Istruzione, dell'Università e della Ricerca, Italy) grant (PRIN 2008, PK2WCW).
Acknowledgements
The authors wish to thank Prof. M.Maggiolini for providing MCF-7 cells, Dr M.P.Vairetti for fluorometric analyses and J.Morini for general technical assistance.
Conflict of Interest Statement: None declared.
References
1. Cucciolla,V. et al. (2007) Resveratrol: from basic science to the clinic. Cell Cycle, 6, 2495–2510.
2. Kimura,Y. et al. (1985) Effects of stilbene derivates on arachidonate metab-olism in leukocytes. Biochim. Biophys. Acta, 837, 209–212.
3. Rotondo,S. et al. (1998) Effect of trans-resveratrol, a natural polyphe-nolic compound, on human polymorphonuclear leukocyte function. Br. J. Pharmacol., 123, 1691–1699.
4. Jang,D.S. et al. (1999) Inhibitory effects of resveratrol analogs on unopso-nized zymosan-induced oxygen radical production. Biochem. Pharmacol., 57, 705–712.
5. Pace-Asciak,C.R. et al. (1995) The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: implications for protection against coronary heart disease. Clin. Chim. Acta, 235, 207–219.
6. Pace-Asciak,C.R. et al. (1996) Wines and grape juices as modulators of plate-let aggregation in healthy human subjects. Clin. Chim. Acta, 246, 163–182.
7. Klinge,C.M. et al. (2008) Resveratrol stimulates nitric oxide production by increasing estrogen receptor alpha-Src-caveolin-1 interaction and phospho-rylation in human umbilical vein endothelial cells. FASEB J., 22, 2185–2197.
8. Jang,M. et al. (1997) Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science, 275, 218–220.
9. Mgbonyebi, O.P. et al. (1998) Antiproliferative effect of synthetic resvera-trol on human breast epithelial cells. Int. J. Oncol., 12, 865–869.
10. Hsieh,T.C. et al. (1999) Cell cycle effects and control of gene expression by resveratrol in human breast carcinoma cell lines with different metastatic potentials. Int. J. Oncol., 15, 245–252.
11. Frankel,E.N. et al. (1993) Inhibition of human LDL oxidation by resvera-trol. Lancet, 341, 1103–1104.
12. Kerry,N.L. et al. (1997) Red wine and fractionated phenolic compounds prepared from red wine inhibit low density lipoprotein oxidation in vitro. Atherosclerosis, 135, 93–102.
13. MacCarrone,M. et al. (1999) Resveratrol prevents apoptosis in K562 cells by inhibiting lipoxygenase and cyclooxygenase activity. Eur. J. Biochem., 265, 27–34.
14. Szewczuk,L.M. et al. (2004) Resveratrol is a peroxidase-mediated inacti-vator of COX-1 but not COX-2: a mechanistic approach to the design of COX-1 selective agents. J. Biol. Chem., 279, 22727–22737.
15. Delmas,D. et al. (2006) Resveratrol as a chemopreventive agent: a promis-ing molecule for fighting cancer. Curr. Drug Targets, 7, 423–442.
16. Huang,C. et al. (1999) Resveratrol suppresses cell transformation and induces apoptosis through a p53-dependent pathway. Carcinogenesis, 20, 237–242.
17. She,Q.B. et al. (2001) Resveratrol-induced activation of p53 and apopto-sis is mediated by extracellular-signal-regulated protein kinases and p38 kinase. Cancer Res., 61, 1604–1610.
18. Lee,K.W. et al. (2008) The resveratrol analogue 3,5,3′,4′,5′-pentahydroxy- trans-stilbene inhibits cell transformation via MEK. Int. J. Cancer, 123, 2487–2496.
19. Sun,C. et al. (2006) Resveratrol downregulates the constitutional activation of nuclear factor-kappaB in multiple myeloma cells, leading to suppres-sion of proliferation and invasion, arrest of the cell cycle, and induction of apoptosis. Cancer Genet. Cytogenet., 165, 9–19.
20. Wu,H. et al. (2008) Resveratrol inhibits hypoxia-induced metastasis potential enhancement by restricting hypoxia-induced factor-1 alpha expression in colon carcinoma cells. Biomed. Pharmacother., 62, 611–621.
21. Weng,C.J. et al. (2012) Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: phenolic acids, monophenol, poly-phenol, and their derivatives. Cancer Treat. Rev., 38, 76–87.
22. Stivala,L.A. et al. (2001) Specific structural determinants are responsible for the antioxidant activity and the cell cycle effects of resveratrol. J. Biol. Chem., 276, 22586–22594.
23. Fang,J.G. et al. (2002) Antioxidant effects of resveratrol and its analogues against the free-radical-induced peroxidation of linoleic acid in micelles. Chemistry, 8, 4191–4198.
24. Cai,Y.J. et al. (2003) Inhibition of free radical-induced peroxidation of rat liver microsomes by resveratrol and its analogues. Biochim. Biophys. Acta, 1637, 31–38.
25. Cheng,J.C. et al. (2006) Structure-activity relationship studies of resvera-trol and its analogues by the reaction kinetics of low density lipoprotein peroxidation. Bioorg. Chem., 34, 142–157.
26. Fang,H. et al. (2001). Structure-activity relationships for a large diverse set of natural, synthetic, and environmental estrogens. Chem. Res. Toxicol., 14, 280–294.
27. Balan,K.V. et al. (2006) Proteasome-independent down-regulation of estrogen receptor-alpha (ERalpha) in breast cancer cells treated with 4,4'-dihydroxy-trans-stilbene. Biochem. Pharmacol., 72, 573–581.
28. Savio,M. et al. (2009) The resveratrol analogue 4,4′-dihydroxy-trans-stilbene inhibits cell proliferation with higher efficiency but different mechanism from resveratrol. Int. J. Biochem. Cell Biol., 41, 2493–2502.
29. Locatelli,G.A. et al. (2005) Inhibition of mammalian DNA polymerases by resveratrol: mechanism and structural determinants. Biochem. J., 389, 259–268.
30. Mosmann,T. (1983) Rapid colorimetric assay for cellular growth and sur-vival: application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65, 55–63.
31. Muscella,A. et al. (2010) Sublethal concentrations of the platinum(II) com-plex [Pt(O,O′-acac)(γ-acac)(DMS)] alter the motility and induce anoikis in MCF-7 cells. Br. J. Pharmacol., 160, 1362–1377.
32. Baur,J.A. et al. (2006) Therapeutic potential of resveratrol: the in vivo evi-dence. Nat. Rev. Drug Discov., 5, 493–506.
33. Meeran,S.M. et al. (2008) Cell cycle control as basis for cancer chemopre-vention through dietary agents. Front. Biosci., 13, 2191–2202.
34. Pozo-Guisado,E. et al. (2002) The antiproliferative activity of resveratrol results in apoptosis in MCF-7 but not in MDA-MB-231 human breast can-cer cells: cell-specific alteration of the cell cycle. Biochem. Pharmacol., 64, 1375–1386.
35. Hsieh,T.C. et al. (2011) Regulation of p53 and cell proliferation by resvera-trol and its derivatives in breast cancer cells: an in silico and biochemical approach targeting integrin αVβ3. Int. J. Cancer, 129, 2732–2743.
36. Zheng,L.F. et al. (2006) DNA damage induced by resveratrol and its synthetic analogues in the presence of Cu (II) ions: mechanism and structure-activity relationship. Free Radic. Biol. Med., 41, 1807–1816.
37. Arima,Y. et al. (2008) Rb depletion results in deregulation of E-cadherin and induction of cellular phenotypic changes that are characteristic of the epithelial-to-mesenchymal transition. Cancer Res., 68, 5104–5112.
38. Liu,J.F. et al. (2002) FGF-2 and TPA induce matrix metalloproteinase-9 secretion in MCF-7 cells through PKC activation of the Ras/ERK pathway. Biochem. Biophys. Res. Commun., 293, 1174–1182.
39. Woo, J.H. et al. (2004) Resveratrol inhibits phorbol myristate acetate-induced matrix metalloproteinase-9 expression by inhibiting JNK and PKC delta signal transduction. Oncogene, 23, 1845–1853.
40. Kimura, Y. et al. (2008) Antitumor activities of synthetic and natural stil-benes through antiangiogenic action. Cancer Sci., 99, 2083–2096.
Received March 5, 2012; revised July 12, 2012; accepted July 13, 2012
C.Maccario et al.
2180
by guest on October 29, 2012
http://carcin.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN



![The photophysics of singlet, triplet, and degradation trap states in 4,4-N,N[sup ʹ]-dicarbazolyl-1,1[sup ʹ]-biphenyl](https://static.fdokumen.com/doc/165x107/634397aac405478ed30633d9/the-photophysics-of-singlet-triplet-and-degradation-trap-states-in-44-nnsup.jpg)



![Diethyl 4,4′-dihydroxy-3,3′-{[(3a RS ,7a RS )-2,3,3a,4,5,6,7,7a-octahydro-1 H -1,3-benzimidazole-1,3-diyl]bis(methylene)}dibenzoate](https://static.fdokumen.com/doc/165x107/63258a10584e51a9ab0ba0e1/diethyl-44-dihydroxy-33-3a-rs-7a-rs-233a45677a-octahydro-1.jpg)
![Mono and binuclear complexes involving [Pd(N,N-dimethylethylenediamine)(H2O)2], 4,4′-bipiperidine and DNA constituents](https://static.fdokumen.com/doc/165x107/631fd774d85b325bc2095ade/mono-and-binuclear-complexes-involving-pdnn-dimethylethylenediamineh2o2.jpg)




![Discovery of (2,4-Dihydroxy-5-isopropylphenyl)-[5-(4-methylpiperazin-1-ylmethyl)-1,3-dihydroisoindol-2-yl]methanone (AT13387), a Novel Inhibitor of the Molecular Chaperone Hsp90 by](https://static.fdokumen.com/doc/165x107/6337b1196f78ac31240eb3a7/discovery-of-24-dihydroxy-5-isopropylphenyl-5-4-methylpiperazin-1-ylmethyl-13-dihydroisoindol-2-ylmethanone.jpg)






