
The release of hepatitis B core antigen from Escherichia coli by batch mode bead milling
7
Short communication The release of hepatitis B core antigen from Escherichia coli by batch mode bead milling Chin Woi Ho a , Wen Siang Tan b,c , Suryani Kamarudin a , Tau Chuan Ling d , Beng Ti Tey a,c, * a Department of Chemical & Environmental Engineering, Faculty of Engineering, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia b Department of Microbiology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia c Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia d Department of Process & Food Engineering, Faculty of Engineering, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia Received 6 July 2007; received in revised form 19 October 2007; accepted 7 November 2007 Abstract The performance of a batch model bead mill on the release of hepatitis B core antigen (HBcAg) from Escherichia coli was investigated in this study. The operating parameters examined were impeller tip speed (8–14 m/s), biomass concentration [5–20% (w/v)] and bead loading [65–80% (v/v)]. The highest yield (24.3 mg/g cell) and rate constant (0.471 l/min) of HBcAg release were achieved at impeller tip speed of 14 m/s. However, the high-shear stress under these operating conditions caused damage of the HBcAg. The highest yield (22.7 mg/g cell) and rate constant (0.344 l/ min) of HBcAg release were observed at biomass concentration of 20% (w/v). There was no significant effect of bead loading on the performance of bead milling being observed. In conclusion, the optimal operating condition for the release of HBcAg was at bead loading of 75% (v/v), biomass concentration of 20% (w/v) and impeller tip speed of 10 m/s. # 2007 Elsevier Ltd. All rights reserved. Keywords: Bead mill; Batch mode; Cell disruption; Hepatitis B core antigen; Downstream process; Escherichia coli 1. Introduction Liver cirrhosis and hepatocellular carcinoma (HCC) caused by hepatitis B virus (HBV) is one of the serious global health problems [1]. There is no effective treatment for acute hepatitis B and it can only be prevented by vaccination. The viral core protein also known as hepatitis B core antigen (HBcAg) is widely used as antigen for the detection of anti-HBcAg antibody [2–4] in serum sample. During viral hepatitis B infection, anti-HBcAg antibodies are produced at the early stage of infection and can persist years after the recovery from the illness [2,5]. Indeed, anti-HBcAg antibodies are the only detectable markers during the period when the hepatitis B surface antigen (HBsAg) has disappeared and anti-HBcAg antibodies indicate that HBV has not yet been eliminated. This characteristic has made anti-HBcAg antibodies a very useful serological marker in diagnosing HBV infections during the screening of populations at risk for HBV infection. Further- more, several observations have suggested that the screening for anti-HBcAg antibodies in blood donors could reduce the incidence of post-tranfusion hepatitis [6]. Therefore, HBcAg has great potential in the development of diagnostic tools for the detection of HBV. The gene encoding HBcAg was successfully expressed in Escherichia coli (E. coli), where it assembles into particles which are indistinguishable from the viral capsids isolated from virally infected liver [7]. We had successfully scaled up the production of this intracellular recombinant HBcAg in a stirred tank bioreactor [8]. There is an increased interest in the efficient and cost effective release of intracellular products like antibiotics, enzymes and therapeutic drugs from the impermeable cell wall and membrane of their host micro-organisms. Prior to the protein purification www.elsevier.com/locate/procbio Process Biochemistry 43 (2008) 206–212 * Corresponding author at: Department of Chemical & Environmental Engi- neering, Faculty of Engineering, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia. Tel.: +60 3 89466289; fax: +60 3 86567120. E-mail address: [email protected] (B.T. Tey). 1359-5113/$ – see front matter # 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.procbio.2007.11.004
-
Upload
umonash-my -
Category
Documents
-
view
1 -
download
0
Transcript of The release of hepatitis B core antigen from Escherichia coli by batch mode bead milling

www.elsevier.com/locate/procbio
Process Biochemistry 43 (2008) 206–212
Short communication
The release of hepatitis B core antigen from Escherichia coli
by batch mode bead milling
Chin Woi Ho a, Wen Siang Tan b,c, Suryani Kamarudin a,Tau Chuan Ling d, Beng Ti Tey a,c,*
a Department of Chemical & Environmental Engineering, Faculty of Engineering, Universiti Putra Malaysia,
43400 Serdang, Selangor Darul Ehsan, Malaysiab Department of Microbiology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia,
43400 Serdang, Selangor Darul Ehsan, Malaysiac Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia
d Department of Process & Food Engineering, Faculty of Engineering, Universiti Putra Malaysia,
43400 Serdang, Selangor Darul Ehsan, Malaysia
Received 6 July 2007; received in revised form 19 October 2007; accepted 7 November 2007
Abstract
The performance of a batch model bead mill on the release of hepatitis B core antigen (HBcAg) from Escherichia coli was investigated in this
study. The operating parameters examined were impeller tip speed (8–14 m/s), biomass concentration [5–20% (w/v)] and bead loading [65–80%
(v/v)]. The highest yield (24.3 mg/g cell) and rate constant (0.471 l/min) of HBcAg release were achieved at impeller tip speed of 14 m/s. However,
the high-shear stress under these operating conditions caused damage of the HBcAg. The highest yield (22.7 mg/g cell) and rate constant (0.344 l/
min) of HBcAg release were observed at biomass concentration of 20% (w/v). There was no significant effect of bead loading on the performance
of bead milling being observed. In conclusion, the optimal operating condition for the release of HBcAg was at bead loading of 75% (v/v), biomass
concentration of 20% (w/v) and impeller tip speed of 10 m/s.
# 2007 Elsevier Ltd. All rights reserved.
Keywords: Bead mill; Batch mode; Cell disruption; Hepatitis B core antigen; Downstream process; Escherichia coli
1. Introduction
Liver cirrhosis and hepatocellular carcinoma (HCC) caused
by hepatitis B virus (HBV) is one of the serious global health
problems [1]. There is no effective treatment for acute hepatitis
B and it can only be prevented by vaccination. The viral core
protein also known as hepatitis B core antigen (HBcAg) is
widely used as antigen for the detection of anti-HBcAg
antibody [2–4] in serum sample. During viral hepatitis B
infection, anti-HBcAg antibodies are produced at the early
stage of infection and can persist years after the recovery from
the illness [2,5]. Indeed, anti-HBcAg antibodies are the only
detectable markers during the period when the hepatitis B
* Corresponding author at: Department of Chemical & Environmental Engi-
neering, Faculty of Engineering, Universiti Putra Malaysia, 43400 Serdang,
Selangor Darul Ehsan, Malaysia. Tel.: +60 3 89466289; fax: +60 3 86567120.
E-mail address: [email protected] (B.T. Tey).
1359-5113/$ – see front matter # 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.procbio.2007.11.004
surface antigen (HBsAg) has disappeared and anti-HBcAg
antibodies indicate that HBV has not yet been eliminated. This
characteristic has made anti-HBcAg antibodies a very useful
serological marker in diagnosing HBV infections during the
screening of populations at risk for HBV infection. Further-
more, several observations have suggested that the screening
for anti-HBcAg antibodies in blood donors could reduce the
incidence of post-tranfusion hepatitis [6]. Therefore, HBcAg
has great potential in the development of diagnostic tools for
the detection of HBV. The gene encoding HBcAg was
successfully expressed in Escherichia coli (E. coli), where it
assembles into particles which are indistinguishable from the
viral capsids isolated from virally infected liver [7]. We had
successfully scaled up the production of this intracellular
recombinant HBcAg in a stirred tank bioreactor [8].
There is an increased interest in the efficient and cost effective
release of intracellular products like antibiotics, enzymes and
therapeutic drugs from the impermeable cell wall and membrane
of their host micro-organisms. Prior to the protein purification

C.W. Ho et al. / Process Biochemistry 43 (2008) 206–212 207
operations, the cell wall must be disrupted to allow the release of
these intracellular products into the surrounding medium.
Optimization of cell disruption is of economically important
because the nature of the disruption process may affect the extent
of product recovery, the ease of the subsequent purification steps,
the nature of the suspension processed and the form and quality
of the final products. In the production of recombinant proteins,
the ratio of variable recovery process costs to fermentation costs
vary from 1 to 3 for enzyme and antibiotic recovery, and the ratio
reached up to 10 for the recovery of intracellular recombinant
insulin [9,10]. Thus, continuous improvement has been made
both by adapting and refining existing cell disruption equipment
and by adding pre-treatment stages to the disruption processes
[11].
Bead milling has been an industrially applicable cell
disruption method due to its capability to process large volume
of biomass and relatively low-power consumption. It is
considered as the method of choice for the disruption of yeast
[12–17] and bacteria [18–21]. It has also been applied
successfully to plant and animal cell disruption [22]. Among
the different type of bead mills, Dyno mill is the most suitable
for scaling up [23]. It consists of several discs or impellers
mounted concentrically or eccentrically on a motor-driven shaft
inside a vertical or horizontal cylindrical chamber. The discs
rotating at high speed and thus generate high-shear force inside
the grinding chamber. The inclusion of small beads produces an
abrasive action to grind the cells. Under the shearing or
compaction of cells and energy transfer from beads to cells, the
microbial cells wall subsequently undergo disintegration, thus
allowing the release of intracellular proteins into the
surrounding medium [23–25]. The performance of bead
milling is influenced by a number of parameters such as
biomass concentration, agitator speed, bead loading, density
and size of bead, temperature, feed flow rate, residence time,
nature and state of the cell and size of organisms [26].
Therefore, the aim of this study was to investigate the effect of
impeller tip speed, biomass concentration and bead loading on
the performance of batch mode bead milling for the release of
HBcAg. The optimized conditions achieved can be used for the
development of an integration recovery process involving bead
milling and expanded bed adsorption chromatography.
2. Materials and methods
2.1. Cultivation of E. coli
The E. coli W3110IQ cells harbouring plasmid PR1-11E encoding trun-
cated HBcAg [7,27] was cultured in Luria Bertani (LB) broth supplemented
with 100 mg/ml ampicillin. The culture was incubated at 37 8C as previously
described by Tey et al. [28]. The cells were harvested by centrifugation at
3750 � g (Avanti JLA-16.250, USA) for 15 min at 4 8C.
2.2. Cell disruption by batch mode bead milling
The pelleted E. coli biomass was washed and resuspended in lysis buffer
[50 mM Tris–HCl (pH 8.0)] to the appropriate biomass concentration [5%,10%,
15%, and 20% (w/v)]. The biomass suspension was fed into the 600-ml glass
chamber of the Dynomill Type MultiLab (CH-4005, W.A. Bachofen, Switzer-
land) loaded with 0.3 mm Zirconia beads [bead loadings of 65%, 75% and 80%
(v/v)]. Cell disruption was carried out under several impeller tip speeds (8, 10
and 14 m/s) for 17 min. The disruption chamber was cooled by an outer jacket
circulated with ice-cooled water. The intracellular protein release generally
obeys the first order kinetics law, in which the rate of protein release is
proportional to the amount of unreleased proteins [25,26]. Samples were
collected at regular interval times and were clarified by centrifugation at
16,249 � g (Mikro 20, H. Zentrifugen, Germany) for 1 min to remove cell
debris.
2.3. Total protein analysis and quantification
The total protein released was analysed with 12% SDS-PAGE and stained
with Coomassie Brilliant Blue R-250 as described by Laemmli [29]. The
amount of total protein was measured with the Bradford assay [30] using
bovine-serum albumin as standard. The absorbance of the sample was read with
a microplate reader (Model 550, Bio-Rad, Japan). All measurements were
carried out in triplicate.
2.4. Purity and quantitation of HBcAg
Purity and quantitation of HBcAg was based on the method previously
described by Ho et al. [31] and Ng et al. [32] with slight modifications. Briefly,
the purity of HBcAg (relative quantity of HBcAg band on the SDS-PAGE gels),
which represents the relative quantity of HBcAg in the sample, was determined
by using the Quantity One1 Quantitation software (Gel Doc, Bio-Rad, USA).
The amount of HBcAg was calculated from the relative quantity of the HBcAg
against the amount of total protein obtained from the Bradford assay. All
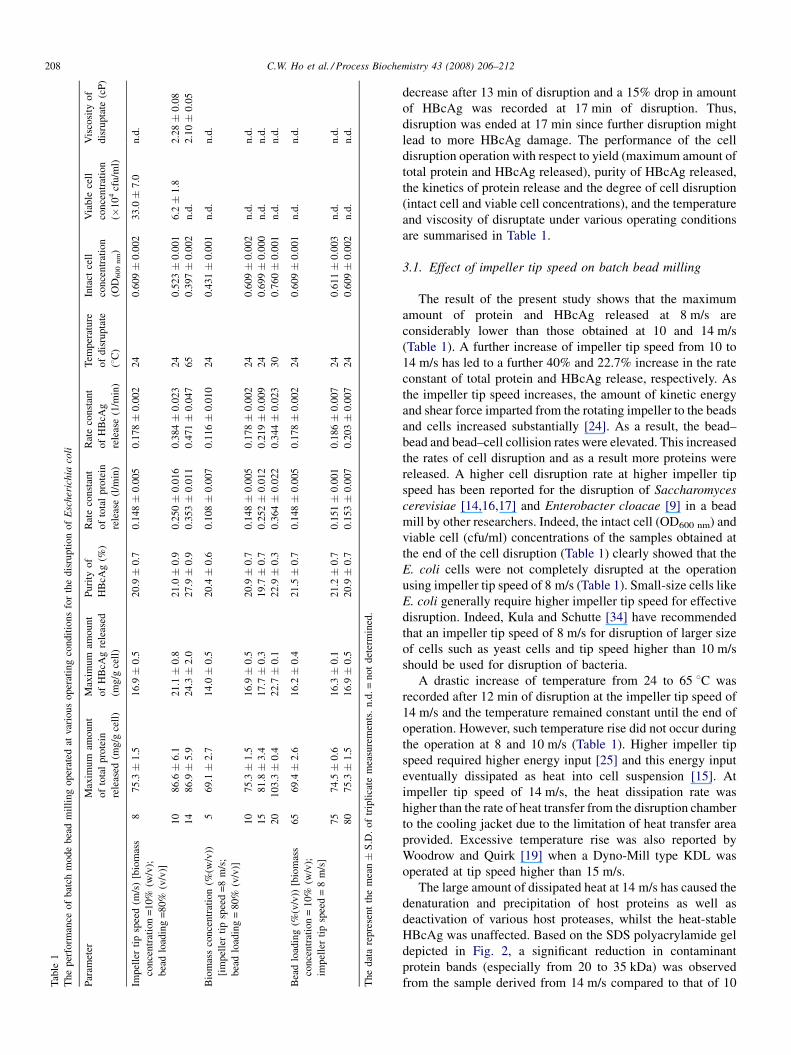
measurements were carried out in triplicate and the data presented in Table 1
were statistically significant.
2.5. Measurement of intact cell density and viable cell concentration
The intact cell density was measured spectrophotometrically under a
wavelength of 600 nm (OD600 nm) with a visible spectrophotometer (UVmine
1240, Shidmadzu, Japan). For the measurement of viable cell concentrations, a
10-fold dilution of disrupted sample was serially diluted under sterile condi-
tions. Diluted sample (50 ml) was spread on a LB agar surface (containing
100 mg/ml ampicillin) and colonies were counted as colony forming unit (cfu)
after incubating the plates at 37 8C for 16 h. All measurements were carried out
in triplicate.
2.6. Measurement of viscosity
Sample was loaded into a viscometer cone and its viscosity was determined
with Spindle SC 14-18 (rotating at a standard speed of 60 rpm) of a viscometer
(DV-II + Viscometer, Brookfield Eng. Lab., USA). All measurements were
carried out in triplicate.
2.7. Analysis of the intactness of HBcAg particles released
The intactness of the HBcAg particles released was analysed using sucrose
density gradient ultracentrifugation as previously described by Tan et al. [7] and
Ho et al. [31].
2.8. Enzyme-linked immunosorbent assay (ELISA)
The antigenicity of the HBcAg was determined by ELISA using a micro-
plate reader (Model 550, Bio-Rad, Japan) as previously described by Ho et al.
[31,33]. All measurements were carried out in triplicate.
3. Results and discussion
The profile of total protein and HBcAg release from batch
mode bead milling is depicted in Fig. 1. Observations in Fig. 1
indicate that the amount of HBcAg in the disruptate started to
Tab
le1
Th
ep
erfo
rman
ceo
fb
atch
mo
de
bea
dm
illi
ng
op
erat
edat
var
iou
so
per
atin
gco
nd
itio
ns
for
the
dis
rup
tio
no
fE
sch
eric
hia
coli
Par
amet
erM
axim
um
amou
nt
of
tota
lp
rote
in
rele
ased
(mg/g
cell
)
Max
imu
mam
ou
nt
of
HB
cAg
rele
ased
(mg
/gce
ll)
Pu
rity
of
HB
cAg
(%)
Rat
eco
nst
ant
of
tota
lp
rote
in
rele
ase
(l/m
in)
Rat
eco
nst
ant
of
HB
cAg
rele
ase
(1/m
in)
Tem
per
atu
re
of
dis
rup
tate
(8C
)
Inta
ctce
ll
con
cen
trat
ion
(OD
600
nm
)
Via
ble
cell
con
cen
trat
ion
(�1
04
cfu
/ml)
Vis
cosi
tyo
f
dis
rup
tate
(cP
)
Imp
elle
rti
psp
eed
(m/s
)[b
iom
ass
con
cen
trat
ion
=1
0%
(w/v
);
bea
dlo
adin
g=
80
%(v
/v)]
87
5.3�
1.5
16
.9�
0.5
20
.9�
0.7
0.1
48�
0.0
05
0.1
78�
0.0
02
24
0.6
09�
0.0
02
33
.0�
7.0
n.d
.
10
86
.6�
6.1
21
.1�
0.8
21
.0�
0.9
0.2
50�
0.0
16
0.3
84�
0.0
23
24
0.5
23�
0.0
01
6.2�
1.8
2.2
8�
0.0
8
14
86
.9�
5.9
24
.3�
2.0
27
.9�
0.9
0.3
53�
0.0
11
0.4
71�
0.0
47
65
0.3
97�
0.0
02
n.d
.2
.10�
0.0
5
Bio
mas
sco
nce
ntr
atio
n(%
(w/v
))
[im
pel
ler
tip
spee
d=
8m
/s;
bea
dlo
adin
g=
80
%(v
/v)]
56
9.1�
2.7
14
.0�
0.5
20
.4�
0.6
0.1
08�
0.0
07
0.1
16�
0.0
10
24
0.4
31�
0.0
01
n.d
.n
.d.
10
75
.3�
1.5
16
.9�
0.5
20
.9�
0.7
0.1
48�
0.0
05
0.1
78�
0.0
02
24
0.6
09�
0.0
02
n.d
.n
.d.
15
81
.8�
3.4
17
.7�
0.3
19
.7�
0.7
0.2
52�
0.0
12
0.2
19�
0.0
09
24
0.6
99�
0.0
00
n.d
.n
.d.
20
10
3.3�
0.4
22
.7�
0.1
22
.9�
0.3
0.3
64�
0.0
22
0.3
44�
0.0
23
30
0.7
60�
0.0
01
n.d
.n
.d.
Bea
dlo
adin
g(%
(v/v
))[b
iom
ass
con
cen
trat
ion
=1
0%
(w/v
);
imp
elle
rti
psp
eed
=8
m/s
]
65
69
.4�
2.6
16
.2�
0.4
21
.5�
0.7
0.1
48�
0.0
05
0.1
78�
0.0
02
24
0.6
09�
0.0
01
n.d
.n
.d.
75
74
.5�
0.6
16
.3�
0.1
21
.2�
0.7
0.1
51�
0.0
01
0.1
86�
0.0
07
24
0.6
11�
0.0
03
n.d
.n
.d.
80
75
.3�
1.5
16
.9�
0.5
20
.9�
0.7
0.1
53�
0.0
07
0.2
03�
0.0
07
24
0.6
09�
0.0
02
n.d
.n
.d.
Th
ed
ata
repre
sen
tth
em
ean�
S.D
.o
ftr
ipli
cate
mea
sure
men
ts.
n.d
.=
no
td
eter
min
ed.
C.W. Ho et al. / Process Biochemistry 43 (2008) 206–212208
decrease after 13 min of disruption and a 15% drop in amount
of HBcAg was recorded at 17 min of disruption. Thus,
disruption was ended at 17 min since further disruption might
lead to more HBcAg damage. The performance of the cell
disruption operation with respect to yield (maximum amount of
total protein and HBcAg released), purity of HBcAg released,
the kinetics of protein release and the degree of cell disruption
(intact cell and viable cell concentrations), and the temperature
and viscosity of disruptate under various operating conditions
are summarised in Table 1.
3.1. Effect of impeller tip speed on batch bead milling
The result of the present study shows that the maximum
amount of protein and HBcAg released at 8 m/s are
considerably lower than those obtained at 10 and 14 m/s
(Table 1). A further increase of impeller tip speed from 10 to
14 m/s has led to a further 40% and 22.7% increase in the rate
constant of total protein and HBcAg release, respectively. As
the impeller tip speed increases, the amount of kinetic energy
and shear force imparted from the rotating impeller to the beads
and cells increased substantially [24]. As a result, the bead–
bead and bead–cell collision rates were elevated. This increased
the rates of cell disruption and as a result more proteins were
released. A higher cell disruption rate at higher impeller tip
speed has been reported for the disruption of Saccharomyces
cerevisiae [14,16,17] and Enterobacter cloacae [9] in a bead
mill by other researchers. Indeed, the intact cell (OD600 nm) and
viable cell (cfu/ml) concentrations of the samples obtained at
the end of the cell disruption (Table 1) clearly showed that the
E. coli cells were not completely disrupted at the operation
using impeller tip speed of 8 m/s (Table 1). Small-size cells like
E. coli generally require higher impeller tip speed for effective
disruption. Indeed, Kula and Schutte [34] have recommended
that an impeller tip speed of 8 m/s for disruption of larger size
of cells such as yeast cells and tip speed higher than 10 m/s
should be used for disruption of bacteria.
A drastic increase of temperature from 24 to 65 8C was
recorded after 12 min of disruption at the impeller tip speed of
14 m/s and the temperature remained constant until the end of
operation. However, such temperature rise did not occur during
the operation at 8 and 10 m/s (Table 1). Higher impeller tip
speed required higher energy input [25] and this energy input
eventually dissipated as heat into cell suspension [15]. At
impeller tip speed of 14 m/s, the heat dissipation rate was
higher than the rate of heat transfer from the disruption chamber
to the cooling jacket due to the limitation of heat transfer area
provided. Excessive temperature rise was also reported by
Woodrow and Quirk [19] when a Dyno-Mill type KDL was
operated at tip speed higher than 15 m/s.
The large amount of dissipated heat at 14 m/s has caused the
denaturation and precipitation of host proteins as well as
deactivation of various host proteases, whilst the heat-stable
HBcAg was unaffected. Based on the SDS polyacrylamide gel
depicted in Fig. 2, a significant reduction in contaminant
protein bands (especially from 20 to 35 kDa) was observed
from the sample derived from 14 m/s compared to that of 10

Fig. 2. The SDS-PAGE of disruptates derived from various impeller tip speeds,
constant biomass concentration [(10% (w/v)] and constant bead loading [80%
(v/v)]. An equal volume of 0.5 ml of disruptate contained between 3.8 and
4.2 mg of total protein was loaded onto the polyacrylamide gel. Lane M:
molecular mass markers; lane 1: disruptate derived from 8 m/s; lane 2:
disruptate derived from 10 m/s; lane 3: disruptate derived from 14 m/s. Sig-
nificant reduction in the amount of contaminant bands of the SDS-PAGE
derived from impeller tip speed of 14 m/s indicates the denaturation and
precipitation of host proteins.
Fig. 1. A typical (a) total protein (~) and (b) HBcAg (*) profile of a batch
mode bead milling. The bead mill was operated at 10% (w/v) E. coli biomass at
impeller tip speed of 10 m/s and bead loading of 80% (v/v). The data represents
the mean � S.D. of triplicate measurements.
C.W. Ho et al. / Process Biochemistry 43 (2008) 206–212 209
and 8 m/s. Indeed, HBcAg was reported to be relatively stable
at temperature as high as 65 8C [32,35]. Ohori et al. [36] have
demonstrated that the antigenicity of HBcAg was still active
after 4 h incubation at 56 8C. Meanwhile, Naito et al. [37]
reported that the yield of HBcAg increased by approximately
50% after the E. coli disruptate was heated for 30 min at 65 8C.
The heat precipitation of host proteins may cause in slight
reduction in viscosity of disruptate (Table 1) as well as
circumvent the interaction of heat-stable HBcAg and E. coli
host proteins, which consequently increased the purity of
HBcAg.
The samples derived from 10 to 14 m/s were further
analysed with sucrose density gradient ultracentrifugation and
ELISA to examine the formation of HBcAg particles and its
antigenicity. The sucrose gradient profiles of the sample derived
from 14 m/s tip speed shows that the HBcAg was almost
uniformly distributed in fractions 5 to 24 (Fig. 3a). In a sucrose
gradient fractionation, heavy molecules are collected in earlier
fractions whilst the later fractions contained mainly lower
molecular weight particles. The uniform pattern indicates that
the HBcAg particles in the samples derived from 14 m/s tip
speed comprise aggregated (migrated to the bottom of the
gradient), ruptured, partially or fully formed core particles
(collected in the mid fractions) and denatured or unformed
HBcAg subunits (collected in the last fractions). In contrast, the
profile of sucrose gradient ultracentrifugation for 10 m/s tip
speed sample clearly shows the formation of intact core
particles in the sample (Fig. 3b). The HBcAg particles formed
were found in abundance (�90%) and migrated to form a bell-
shaped peak (fractions 9–14).
The result of ELISA test shows that the antigenicity of
HBcAg in the 14 m/s tip speed sample was affected (Fig. 4).
The A405 nm reading for 1000 ng of HBcAg for this sample after
1 h incubation with substrate was three-fold lower than that of
sample derived from 10 m/s after 20 min incubation with the
substrate. This is not surprising since a portion of the samples
derived from 14 m/s operation are comprised of ruptured,
partially formed core particles and denatured or unformed
HBcAg subunits as indicated by the sucrose gradient profile
(Fig. 3a). On the contrary, a strong reactivity signal was
detected in the sample derived from 10 m/s tip speed. The anti-
HBcAg monoclonal antibody could detect as low as 50 ng
(0.5 ng/mL) of HBcAg, confirming that HBcAg prepared by
10 m/s tip speed was functionally active and can be potentially
used as a diagnostic reagent for the detection of anti-HBcAg
antibodies.
3.2. Effect of initial cell biomass concentration on batch
bead milling
The maximum amount of both total protein and HBcAg
released was increased as the biomass concentration increased
from 5% to 20% (w/v) (Table 1). The bead milling operation
with 20% (w/v) biomass concentration has released a total of
103.3 mg protein/g cell, which was 26%, 37% and 49% higher
than that of 15%, 10% and 5% (w/v), respectively. A total of

Fig. 3. The sucrose gradient profile of HBcAg released from E. coli in batch mode bead milling. SDS-PAGE shows the presence of HBcAg, and graphical illustration
indicates the concentration of HBcAg in the sucrose gradient fractions. (a) Samples from batch mode bead milling at 14 m/s. Lane M: molecular mass markers; lanes
1–24: sucrose gradient fractions. The uniform pattern indicates that the HBcAg particles in the 14 m/s tip speed sample comprises evenly of aggregated, ruptured,
partially or fully formed core particles and denatured or unformed monomers. (b) Samples from batch mode bead milling at 10 m/s. Lane M: molecular mass markers;
lanes 1–24: sucrose gradient fractions. The majority of HBcAg was found in the fractions of 9–14 indicating that most of the HBcAg in the disruptate has formed
intact HBcAg particles.
Fig. 4. The antigenicity of the released HBcAg prepared from different tip
speed was tested using ELISA with an anti-HBcAg monoclonal antibody.
Microtiter plate wells were coated with different concentration of HBcAg.
The bound HBcAg was detected by anti-HBcAg monoclonal antibody (1:1000
dilution) and an anti-mouse antibody conjugated to alkaline phosphatase. The
antigenicity of HBcAg was detected by measuring the optical density at 405 nm
after different incubation time with addition of substrate for the enzyme.
Samples tested were HBcAg prepared from 14 m/s tip speed and incubated
for 20 min (*), and 1 h (&); and HBcAg prepared from 10 m/s tip speed and
incubated for 20 min (^). The anti-HBcAg monoclonal antibody was replaced
by milk diluent in the negative control (~). The data represents the
mean � S.D. of triplicate measurements.
C.W. Ho et al. / Process Biochemistry 43 (2008) 206–212210
22.7 mg of HBcAg/g cell was also obtained at the operation at
this biomass concentration, which was 29%, 34% and 62%
higher than that of 15%, 10% and 5% (w/v), respectively.
Besides, the rate constants of HBcAg release recorded at this
operation were 1.6-, 1.9- and three-fold higher than that
recorded at 15%, 10% and 5% (w/v), respectively. This means
that the operation of cell disruption can be operated at a shorter
period of time with higher biomass concentration loading. The
presence of higher biomass in the disruption chamber will
reduce the distances between centres of neighbouring cells and
consequently increases the intensity of mutual interactions of
the micro-organisms [38]. As a result, the cell disruption
efficiency and the rate of protein release are improved. Indeed,
other researchers have observed a similar correlation between
protein release and initial biomass concentration [16,21,39].
Apart from the higher protein release rate, the energy utilization
was also reported to be lower at a higher biomass concentration.
Bunge et al. [40] reported that a significantly lower mechanical
energy input was needed for the disruption of Arthrobacter sp at
biomass concentration of 55% (w/v) compared to that at 10%
(w/v). However, a further increase of biomass concentration
from 20% (w/v) is unfavorable in this study. At higher biomass
concentration, the viscosity of the disruptate was higher, which
poses problems to the subsequent steps of downstream
processing, for instance, chromatographic processes of size
exclusion and expanded bed adsorption. Besides, a viscous
sample not only requires more intense centrifugation force for
solid–liquid separation, but also impedes the separation
performance of centrifuges [41], pumps [41], ultrafiltration
[42], and cross-flow filtration systems [43,44]. High-viscosity
sample also has high-power dissipation under mixing or
agitating mode and thus promote high-heat generation [45].
This explained the moderate temperature increase (to 30 8C)
during the bead milling operation at 20% (w/v) biomass
concentration.

C.W. Ho et al. / Process Biochemistry 43 (2008) 206–212 211
3.3. Effect of bead loading on batch bead milling
There was no obvious improvement in terms of maximum
total protein and HBcAg released as the bead loading was
increased from 65% to 80% (v/v) in the present study. This is
not surprising since the degree of cell disruption is very similar
at various bead loadings as shown by the measurement of intact
cell particles (OD600 nm; Table 1). This result was somehow not
in accord with that reported by other researchers: higher protein
yield at higher bead loading has been reported by Schutte and
Kula [46] on the disruption of E. coli and baker’s yeast and
Tamer and Moo-Young [20] on the disruption of Alcaligenes
latus. However, a vibrated mixing chamber was used by Schutte
and Kula [46], while a horizontal agitation chamber was used in
the present study. A bead loading in the range of 80–85% (v/v)
is generally accepted as the optimum condition and has been
used in many studies [15,19,21,23]. Bead loading higher than
this range is not preferred for cell disruption due to the high-
heat generation and power consumption, which often out-
weighs the minimal increase in disruption efficiency [47].
Indeed, in the present study, a rise in chamber pressure occurred
when a bead loading of 85% (v/v) was used, which resulted in a
portion of feedstock seeping out from the bead chamber
throughout the process of batch bead milling.
4. Conclusion
In this study, we have developed and optimized a scale-able
cell disruption method for the release of intracellular
recombinant HBcAg from E. coli cells. The highest HBcAg
release rate was achieved at 14 m/s impeller tip speed.
However, the high level of shear stress imparted to the cells
at high-tip speed operation ruptured HBcAg particles and
affected the antigenicity. The yield of protein was increased
with the increasing of biomass concentration due to the higher
mutual interactions, which occur in the grinding chamber. A
slightly increased of temperature to 30 8C was recorded at
biomass concentration of 20% (w/v); therefore a further
increase of biomass concentration is unfavorable. There were
no obvious effects of bead loading on the performance of bead
milling operation for HBcAg release observed. Hence, the
optimal operating conditions for the disruption of E. coli to
release HBcAg is at impeller tip speed of 10 m/s, biomass
concentration of 20% (w/v) and bead loading of 75% (v/v).
Acknowledgement
This study was supported by the IRPA Grant (09-02-04-
0766-EA001) and an E-Science Grant (02-01-04-SF0486) from
the Ministry of Science, Technology and Innovation of
Malaysia.
References
[1] World Health Organization. Introduction of hepatitis b vaccine into
childhood immunization services. Geneva: WHO; 2001. pp. 1–4.
[2] Hoofnagle JH, Gerety RJ, Ni LY, Barker LF. Antibody to hepatitis B core
antigen—a sensitive indicator of hepatitis B virus replication. N Engl J
Med 1974;290:1336–40.
[3] Korec E, Korcove J, Konig J, Hlozanek I. Detection of antibodies against
hepatitis B core antigen using the avidin–biotin system. J Virol Methods
1989;24:321–6.
[4] Tordjeman M, Rabillon V, Abouth D, Trepo C, Hoffenbach A, Somme G.
Specific detection of anti-HBc antibodies with an enzyme immunoassay
using recombinant HBcAg and monoclonal antibodies. J Virol Methods
1992;43:21–30.
[5] Milich DR, McLachlan A. The nucleocapsid of hepatitis B virus is
both a T-cell independent and T-cell dependent antigen. Science
1986;234:1398–401.
[6] Lai KN, Lai FM, Leung NWY, Lo ST, Tam JS. Hepatitis with isolated
serum antibody to hepatitis B core antigen. A variant of non-A, non-B
hepatitis? Am J Clin Pathol 1990;413:79–84.
[7] Tan WS, Dyson MR, Murray K, Hepatitis. B virus core antigen: enhance-
ment of its production in Escherichia coli, and interaction of the
core particles with the viral surface antigen. Biol Chem 2003;384:
363–71.
[8] Tey BT, Chua MI, Chua GS, Ng MYT, Awang Biak DR, Tan WS, Ling TC.
Influence of temperature and level of mixing on the growth and HBcAg
production of Escherichia coli in stirred tank bioreactor.. Biotechnol
Bioprocess Eng 2006;11:164–7.
[9] Fish NM, Lilly MD. The interactions between fermentation and protein
recovery. Biotechnology 1984;2:623–7.
[10] Datar R, Rosen CG. Studies on the removal of Escherichia coli cell debris
by aqueous two-phase polymer extraction. J Biotechnol 1986;3:207–19.
[11] Foster D. Optimizing recombinant product recovery through improve-
ments in cell-disruption technologies. Curr Opin Biotechnol 1995;6:
523–6.
[12] Currie JA, Dunnili P, Lilly MD. Release of protein from bakers’ yeast
(Saccharomyces cerevisiae) by disruption in an industrial agitator mill.
Biotechnol Bioeng 1972;14:725–36.
[13] Marffy F, Kula MR. Enzyme yields from cells of brewer’s yeast disrupted
by treatment in a horizontal disintegrator. Biotechnol Bioeng
1974;16:623–34.
[14] Limon-Lason J, Hoare M, Orsborn CB, Doyle DJ, Dunnill P. Reactor
properties of a high-speed bead mill for microbial rupture. Biotechnol
Bioeng 1979;21:745–74.
[15] Canales M, Buxado JA, Heynngnezz L, Eriquez A. Mechanical disruption
of Pichia pastoris yeast to recover the recombinant glycoprotein Bm86.
Enzyme Microb Technol 1998;23:58–63.
[16] Ricci-Silva ME, Vitolo M, Abhahao-Neto J. Protein and glucose 6-
phosphate dehydrogenase releasing from baker’s yeast cells disrupted
by a vertical bead mill. Process Biochem 2000;35:831–5.
[17] Chow YM, Tey BT, Ibrahim MN, Ariff A, Ling TC. The disruption of
Saccharomyces cerevisiae cells and release of glucose 6-phosphate dehy-
drogenase (G6PDH) in a horizontal Dyno Bead Mill operated in con-
tinuous recycling mode.. Biotechnol Bioprocess Eng 2005;10:184–288.
[18] Rehacek J. Continuous disintegration of micro-organisms in a new
laboratory apparatus. Experientia 1971;27:1103–4.
[19] Woodrow JR, Quirk AV. Evaluation of the potential of a bead mill for the
release of intracellular bacterial enzymes. Enzyme Microb Technol
1982;4:385–9.
[20] Tamer IM, Moo-Young M. Disruption of Alcaligenes latus for recovery of
poly (b-hydroxybutyric acid): comparison of high-pressure homogeniza-
tion, bead milling and chemically induced lysis. Ind Eng Chem Res
1998;37:1807–14.
[21] Bury D, Jelen P, Kalab M. Disruption of Lactobacillus delbrueckii ssp.
Bulgaricus 11,842 cells for lactose hydrolysis ion dairy products: a
comparison of sonication, high-pressure homogenization and bead
milling.. Innov Food Sci Emerg Technol 2001;2:23–9.
[22] Hopkins T. Laboratory cell disruptors: a review of apparatus & techniques.
Int Lab News 2000;16–7. October.
[23] Garcia FAP. Cell wall disruption. In: Kennedy JF, editor. Cabral JMS
recovery process for biological materials. New York: John Wiley & Sons;
1993. p. 47–69.

C.W. Ho et al. / Process Biochemistry 43 (2008) 206–212212
[24] Melendres AV, Unno H, Shiragami N, Honda H. A concept of critical
velocity for cell disruption by bead mill. J Chem Eng Jpn 1992;25:354–6.
[25] Middelberg APG. Process cell disruption of micro-organisms. Biotechnol
Adv 1995;13:491–551.
[26] Harisson STL. Bacterial cell disruption: a key unit operation in the
recovery of intracellular products. Biotechnol Adv 1991;9:217–40.
[27] Tan WS, McNae IW, Ho KL, Walkinshaw MD, Crystallization. X-ray
analysis of the T=4 particle of hepatitis B capsid protein with an
N-terminal extension. Acta Cryst 2007;F63:642–7.
[28] Tey BT, Yong KH, Ong HP, Ling TC, Ong ST, Tan YP, Ariff A, Tan WS.
Optimal conditions for hepatitis B core antigen production in shaked flask
fermentation. Biotechnol Bioprocess Eng 2004;9:374–8.
[29] Laemmli UK. Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 1970;227:680–5.
[30] Bradford MM. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein–dye
binding. Anal Biochem 1976;72:248–54.
[31] Ho CW, Tan WS, Kamaruddin S, Ling TC, Tey BT. The direct recovery of
recombinant hepatitis B core antigen (HBcAg) from disruptate derived
from continuous flow bead milling. Biotechnol Appl Biochem 2007. doi:
10.1042/BA20070088.
[32] Ng MYT, Tan WS, Abdullah N, Ling TC, Tey BT. Heat treatment of
unclarified Escherichia coli homogenate improved the recovery effi-
ciency of recombinant hepatitis B core antigen. J Virol Methods
2006;137:134–9.
[33] Ho CW, Chew TK, Ling TC, Kamaruddin S, Tan WS, Tey BT. Efficient
mechanical cell disruption of Escherichia coli by an ultraconicator and
recovery of intracellular hepatitis B core antigen. Process Biochem
2006;41:1829–34.
[34] Kula MR, Schutte H. Purification of proteins and the disruption of
microbial cells. Biotechnol Prog 1987;3:31–42.
[35] Dyson MR, Murray K. Selection of peptide inhibitors of interactions
involved in complex protein assemblies: association of the core and
surface antigens of hepatitis B virus. Proc Natl Acad Sci USA
1995;92:2194–8.
[36] Ohori H, Shimizu N, Yamada E, Onodera S, Ishida N. Immunological and
morphological properties of HBeAg subtypes (HBeAg/1 and HBeAg/2) in
hepatitis B virus core particles. J Gen Virol 1984;65:405–14.
[37] Naito M, Ishii K, Nakamura Y, Kobayashi M, Takada S, Koike K. Simple
method for efficient production of hepatitis B virus core antigen in
Escherichia coli. Res Virol 1997;148:299–305.
[38] Heim A, Kamionowska U, Solecki M. The effect of micro-organism
concentration on yeast cell disruption in a bead mill. J Food Eng
2007;83:121–8.
[39] Morohashi S, Okada S, Hataya T, Sasaki T, Hoshino K, Sasakura T.
Release process of protein from Baker’s yeast by disruption in agitating
bead mill. J Chem Eng Jpn 1997;30:182–6.
[40] Bunge F, Pietzsch M, Mfiller R, Slydatk C. Mechanical disruption of
Arthrobacter sp. DSM 3747 in stirred ball mills for the release of
hydrantoin-cleaving enzymes. Chem Eng Sci 1992;47:225–32.
[41] Li WG. Effects of viscosity of fluids on centrifugal pump performance and
flow pattern in the impeller. Int J Heat Fluid Flow 2000;21:207–12.
[42] Wang SS. Effect of solution viscosity on ultrafiltration flux. J Membr Sci
1988;39:187–94.
[43] Brocklebank MP. Downstream processing plant and equipment. In:
Asenjo JA, editor. Separation processes in biotechnology. New York:
Marcel Dekker; 1990. p. 617–740.
[44] Fane AG, Radovich JM. Membrane systems. In: Asenjo JA, editor.
Separation processes in biotechnology. New York: Marcel Dekker; 1990.
p. 209–62.
[45] Khalaf WG, Sastry SK. Effect of fluid viscosity on the ohmic heating rate
of solid–liquid mixtures. J Food Eng 1996;27:145–58.
[46] Schutte H, Kula MR. Analytical disruption of micro-organisms in a mixer
mill. Enzyme Microb Technol 1988;10:552–8.
[47] Geciova J, Bury D, Jelen P. Methods for disruption of microbial cells for
potential use in the dairy industry—a review. Int Dairy J 2002;12:541–53.




















