The protein kinase Cdk5 Structural aspects, roles in neurogenesis and involvement in Alzheimer's...
10
MINIREVIEW The protein kinase Cdk5 Structural aspects, roles in neurogenesis and involvement in Alzheimer’s pathology Ricardo B. Maccioni 1 , Carola Otth 2 , Ilona I. Concha 2 and Juan P. Mun ˜ oz 1 1 Laboratory of Cellular and Molecular Biology, Millennium Institute for Advanced Studies in Cell Biology and Biotechnology, Faculty of Sciences, University of Chile, N ˜ un ˜oa, Santiago, Chile; 2 Instituto de Bioquı ´mica, Universidad Austral, Valdivia, Chile A set of different protein kinases have been involved in tau phosphorylations, including glycogen synthase kinase 3b (GSK3b), MARK kinase, MAP kinase, the cyclin- dependent kinase 5 (Cdk5) system and others. The latter system include the catalytic component Cdk5 and the regulatory proteins p35, p25 and p39. Cdk5 and its neuron- specific activator p35 are essential molecules for neuronal migration and for the laminar configuration of the cerebral cortex. Recent evidence that the Cdk5/p35 complex con- centrates at the leading edge of axonal growth cones, together with the involvement of this system in the phos- phorylation of neuronal microtubule-asociated proteins (MAPs), provide further support to the role of this protein kinase in regulating axonal extension in developing brain neurons. Although the aminoacid sequence of p35 has little similarity with those of normal cyclins, studies have shown that its activation domain may adopt a conformation of the cyclin-folded structure. The computed structure for Cdk5 is compatible with experimental data obtained from studies on the Cdk5/p35 complex, and has allowed predictions on the protein interacting domains. This enzyme exhibits a wide cell distribution, even though a regulated Cdk5 activity has been shown only in neuronal cells. Cdk5 has been charac- terized as a proline-directed Ser/ Thr protein kinase, that contributes to phosphorylation of human tau on Ser202, Thr205, Ser235 and Ser404. Cdk5 is active in postmitiotic neurons, and it has been implicated in cytoskeleton assembly and its organization during axonal growth. In addition to tau and other MAPs, Cdk5 phosphorylates the high molecular weight neurofilament proteins at their C-terminal domain. Moreover, nestin, a protein that regulates cytoskeleton organization of neuronal and mus- cular cells during development of early embryos, and several other regulatory proteins appear to be substrates of Cdk5 and are phosphorylated by this kinase. Studies also suggest, that in addition to Cdk5 involvement in neuronal differentiation, its activity is induced during myogenesis, however, the mechanisms of how this activity is regulated during muscular differentiation has not yet been elucidated. Recent studies have shown that the b-amyloid peptide (Ab) induces a deregulation of Cdk5 in cultured brain cells, and raises the question on the possible roles of this tau- phosphorylating protein kinase in the sequence of mol- ecular events leading to neuronal death triggered by Ab. In this context, there are evidence that Cdk5 is involved in tau hyperphosphorylation promoted by Ab in its fibrillary form. Cdk5 inhibitors protect hippocampal neurons against both tau anomalous phosphorylations and neuronal death. The links between the studies on the Cdk5/p35 system in normal neurogenesis and its claimed participation in neurodegeneration, provide the framework to understand the regulatory relevance of this kinase system, and changes in its regulation that may be implicated in disturbances such as those occurring in Alzheimer disease. Keywords: cyclin-dependent kinase 5 (Cdk5); structural/ functional aspects; cytoskeleton; tau protein; protein phos- phorylation; neuronal development; neurodegenerative disorders; Alzheimer’s disease. A marked interest among cell biologists and neuroscientists has been generated by the findings that the cyclin- dependent protein kinase 5 (Cdk5) and the regulatory protein p35, also named neuronal Cdk5 activator (Nck5a), are involved in tau hyperphosphorylations and hence in the route by which b-amyloid protein (Ab) triggers neuronal death [1–3]. The neuronal microtubule-associated protein tau is a target of phosphorylations on a large number of sites, and a substrate for several kinases according with in vitro studies. Some of these are proline-directed kinases, e.g, MAP-kinase, GSK3b and Cdk5, while other enzymes including MARK kinase, protein kinase A and others are nonproline directed kinases [4,5]. Studies have claimed that some of tau phosphorylating kinases, such as GSK3b [5,6], Eur. J. Biochem. 268, 1518–1527 (2001) q FEBS 2001 Correspondence to R. B. Maccioni, Laboratory of Cellular and Molecular Biology, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, N ˜ un ˜oa, Santiago, Chile. Fax: 1 562 218 6245, Tel.: 1 562 260 0255, E-mail: [email protected] Abbreviations: GSK3b, glycogen synthase kinase 3b; cdk, cyclin-dependent kinase; MAP, microtubule-asociated protein; Ab, b-amyloid peptide; Nck5a, neuronal Cdk5 activator; PHF, paired helical filaments; GAK, cyclin G-associated kinase; frontotemporal, dementia and parkinsonism linked to chromosome-17. (Received 22 August 2000, revised 2 January 2001, accepted 15 January 2001)
Transcript of The protein kinase Cdk5 Structural aspects, roles in neurogenesis and involvement in Alzheimer's...

M I N I R E V I E W
The protein kinase Cdk5Structural aspects, roles in neurogenesis and involvement in Alzheimer's pathology
Ricardo B. Maccioni1, Carola Otth2, Ilona I. Concha2 and Juan P. MunÄ oz1
1Laboratory of Cellular and Molecular Biology, Millennium Institute for Advanced Studies in Cell Biology and Biotechnology,
Faculty of Sciences, University of Chile, NÄ unÄoa, Santiago, Chile; 2Instituto de BioquõÂmica, Universidad Austral, Valdivia, Chile
A set of different protein kinases have been involved in
tau phosphorylations, including glycogen synthase kinase
3b (GSK3b), MARK kinase, MAP kinase, the cyclin-
dependent kinase 5 (Cdk5) system and others. The latter
system include the catalytic component Cdk5 and the
regulatory proteins p35, p25 and p39. Cdk5 and its neuron-
specific activator p35 are essential molecules for neuronal
migration and for the laminar configuration of the cerebral
cortex. Recent evidence that the Cdk5/p35 complex con-
centrates at the leading edge of axonal growth cones,
together with the involvement of this system in the phos-
phorylation of neuronal microtubule-asociated proteins
(MAPs), provide further support to the role of this protein
kinase in regulating axonal extension in developing brain
neurons. Although the aminoacid sequence of p35 has little
similarity with those of normal cyclins, studies have shown
that its activation domain may adopt a conformation of the
cyclin-folded structure. The computed structure for Cdk5 is
compatible with experimental data obtained from studies on
the Cdk5/p35 complex, and has allowed predictions on the
protein interacting domains. This enzyme exhibits a wide
cell distribution, even though a regulated Cdk5 activity has
been shown only in neuronal cells. Cdk5 has been charac-
terized as a proline-directed Ser/Thr protein kinase, that
contributes to phosphorylation of human tau on Ser202,
Thr205, Ser235 and Ser404. Cdk5 is active in postmitiotic
neurons, and it has been implicated in cytoskeleton
assembly and its organization during axonal growth. In
addition to tau and other MAPs, Cdk5 phosphorylates
the high molecular weight neurofilament proteins at their
C-terminal domain. Moreover, nestin, a protein that
regulates cytoskeleton organization of neuronal and mus-
cular cells during development of early embryos, and
several other regulatory proteins appear to be substrates of
Cdk5 and are phosphorylated by this kinase. Studies also
suggest, that in addition to Cdk5 involvement in neuronal
differentiation, its activity is induced during myogenesis,
however, the mechanisms of how this activity is regulated
during muscular differentiation has not yet been elucidated.
Recent studies have shown that the b-amyloid peptide (Ab)
induces a deregulation of Cdk5 in cultured brain cells,
and raises the question on the possible roles of this tau-
phosphorylating protein kinase in the sequence of mol-
ecular events leading to neuronal death triggered by Ab. In
this context, there are evidence that Cdk5 is involved in tau
hyperphosphorylation promoted by Ab in its fibrillary
form. Cdk5 inhibitors protect hippocampal neurons against
both tau anomalous phosphorylations and neuronal death.
The links between the studies on the Cdk5/p35 system in
normal neurogenesis and its claimed participation in
neurodegeneration, provide the framework to understand
the regulatory relevance of this kinase system, and changes
in its regulation that may be implicated in disturbances such
as those occurring in Alzheimer disease.
Keywords: cyclin-dependent kinase 5 (Cdk5); structural/
functional aspects; cytoskeleton; tau protein; protein phos-
phorylation; neuronal development; neurodegenerative
disorders; Alzheimer's disease.
A marked interest among cell biologists and neuroscientistshas been generated by the findings that the cyclin-dependent protein kinase 5 (Cdk5) and the regulatoryprotein p35, also named neuronal Cdk5 activator (Nck5a),are involved in tau hyperphosphorylations and hence in theroute by which b-amyloid protein (Ab) triggers neuronaldeath [1±3]. The neuronal microtubule-associated proteintau is a target of phosphorylations on a large number ofsites, and a substrate for several kinases according within vitro studies. Some of these are proline-directed kinases,e.g, MAP-kinase, GSK3b and Cdk5, while other enzymesincluding MARK kinase, protein kinase A and others arenonproline directed kinases [4,5]. Studies have claimed thatsome of tau phosphorylating kinases, such as GSK3b [5,6],
Eur. J. Biochem. 268, 1518±1527 (2001) q FEBS 2001
Correspondence to R. B. Maccioni, Laboratory of Cellular and
Molecular Biology, Facultad de Ciencias, Universidad de Chile,
Las Palmeras 3425, NÄ unÄoa, Santiago, Chile. Fax: 1 562 218 6245,
Tel.: 1 562 260 0255, E-mail: [email protected]
Abbreviations: GSK3b, glycogen synthase kinase 3b; cdk,
cyclin-dependent kinase; MAP, microtubule-asociated protein;
Ab, b-amyloid peptide; Nck5a, neuronal Cdk5 activator; PHF, paired
helical filaments; GAK, cyclin G-associated kinase; frontotemporal,
dementia and parkinsonism linked to chromosome-17.
(Received 22 August 2000, revised 2 January 2001, accepted
15 January 2001)

MAP-kinase [4,7], microtubule-affinity regulating kinaseMARK [8] and Cdk5 [1,2] phospohorylate tau at sites thatare elevated in Alzheimer's paired helical filaments (PHFs).Hyperphosphorylations could lead to structural alteration intau, and decrease in its interaction with microtubules [9,10].It is possible that these kinases could be sensitive tochanges in their regulatory patterns or at the structural level,thus participating in the molecular pathway leading toneurodegeneration.
Cdk5 is a Ser-Thr protein kinase with postmitotic activitythat phosphorylates KSP protein motifs on MAP1b, KSPXKon tau [11] or KSPXX on neurofilament proteins, actin-binding protein caldesmon and proteins of the synapticvesicles [12±14]. This kinase was originally found inhuman cell lines as PSSALRE kinase [15], and almostsimultaneously identified and characterized as a brainproline-directed kinase [16]. Cdk5 was shown as an activeenzyme in postmitotic cells such as neurons [17], and as akinase that phosphorylates tau protein [18]. Soon after thesediscoveries, protein responsible for activation of brain Cdk5were found [19,20]. The Cdk5/p35 system participates innormal development of neuronal cells and the axonalgrowth [21,22]. Therefore, Cdk5 appears as one of theimportant kinases phosphorylating tau in neuronal cells.Cdk5 was identified on the basis of biochemical similaritiesto Cdc2 protein kinase, a regulator of mitosis. Unlike Cdc2,however, Cdk5 is not found in dividing cells, and exhibits apreferential distribution in brain differentiating neurons [11].The formation of a stable Cdk5/p35 complex followingphosphorylation of Cdk5 in hippocampal neurons [23], hasbeen postulated as a factor responsible for constitutiveactivation of this protein kinase with a consequent increasein phosphorylation at sites that have been found in taupathology. On the other hand, formation of a Cdk5 complexwith p25 [24], fragment derived from proteolytic cleavageof p35 by calpain or proteosomes, has been proposed as anstep toward Cdk5 activation. However, its involvement inthe pathway leading to neuronal degeneration in Alzheimer'sdisease is not clear at all. Besides the action of kinases inthe mechanisms of tau modifications as related withneurodegeneration, inhibition of phosphatases could bealso involved [25]. In this context, it is worth pointing outthat the detailed molecular aspects and the type of proteinkinases and phosphatases, and their implications inAlzheimer's physiopathology remain to be elucidated.The background on structural biological studies and theknowledge attained on the different enzymatic systems thatmodify tau protein, combined with future in vitro andin vivo studies are likely to provide clues to betterunderstand the changes in regulatory patterns and Cdk5implications in neuronal degeneration.
The Cdk5/p35 system: structural aspects
The processes that control the cell cycle are linked with afinely tuned set of molecular signals that operate in acoordinated fashion in animal development. Cyclin-dependent kinases are major regulators at the differentlevels of cell division. However, certain cyclin dependentkinases do not participate in controlling cell cycle in amature cellular system, but they are rather involved incontrolling cell differentiation in neuronal and muscle cells.They seem to have importance at the level of regulatory
processes in the cell nucleus and in the cytoskeletonorganization. Deregulation of these kinases appears to beinvolved in neuronal degeneration according with studies incellular models, an observation that may have implicationsfor human brain disorders such as Alzheimer and musculardiseases [1,23,24]. A set of seven proteins homologous withCdc2 derived from the gene Cdc2 have been found inmammalian cells, and are dependent on cyclins for theiractivity [26,27]. Cdc2 associates with type B cyclins andthe complex regulates the mitotic phase M of cell cycle[28]. Cdk2 associates with A and E cyclins, regulating thesynthesis phase S and the G1-S transition of the cell cycle[29]. Cdk4 and cdk6 associate with D cyclins and areimportant for progression of the G1 phase [29]. Cdk5 isactivated at the neuronal level by the cyclin-relatedproteins, p25, p35 and p39 [19,30], which bind to oneside of the catalytic cleft, interacting with both the N- andC-terminal lobes of Cdk5 to form a large, continuousprotein±protein interface [31].
Recent investigations have revealed that most of theCdk5 in cells forms large multimeric complexes of highmolecular weight, i.e. 60, 200, 450 and 670 kDa, inwhich Cdk5 is associated with p35, p25, synapsin, tau,CK1, b-catenins and N-cadherins [32±36]. These findingsindicate that Cdk5 is a multifunctional protein that associ-ates with other cell proteins to interact with the cyto-skeleton and form supramolecular complexes [37]. Cdk5may thus exhibit different domains for interactions with adiverse family of proteins, and can be activated by phos-phorylation at Ser159, similar to Thr160 phosphorylation inother kinases of the cell cycle. The activities of thesekinases increase several times in the phosphorylated state[38]. Therefore, Cdk5 could be regulated by phosphoryl-ation and alternative interaction with activators such as p35,p25 or p39 in a specific subset of neuronal cells. Furtherinsight into the regulatory roles of Cdk5 and p35 have beenobtained from studies on mutant mice with disruption ofgenes of these proteins [39]. However, the fact that theseproteins have not been crystallized yet has precluded adetailed knowledge of the structural interactions and theregulatory mechanisms of Cdk5. Several approaches havebeen used to explore the recognition of the enzyme forsubstrates and docking Cdk5/p25 [31]. The polypeptidesequence of Cdk5 has revealed a high degree of identitywith other cyclin-dependent kinases at the level of primaryand tridimensional structures [40]. The alignment of Cdk5with other cyclins as well as the putative sites for proteininteractions are shown in Fig. 1, where a high level ofidentity is observed. The alignment of p35 with p39 revealsidentity in potential regions for protein interaction. Despitethe sequence homology between p35, p25 or p39 activatorsand cyclins is low, the folding of these proteins is similar tothe different cyclins [40,41], a fact that may explain thefunctional relationships of these Cdk5 regulators with thefamily of cyclins involved in cell cycle control.
The finding of the potential phosphorylation of Ser159 inCdk5 indicates that the increased kinase activity may bedue to a conformational change, similarly to that occurringwith cdk2 [42], in which the cyclin binding triggers a 21 AÊ
change in the T-loop (146±166). These changes shouldpermit a better access of substrates and a favorablepositioning of ATP for the phosphate transfer [43,44].Moreover, Cdk5, by analogy with other cdk's, presents a
q FEBS 2001 Structure, function and pathology of protein kinase Cdk5 (Eur. J. Biochem. 268) 1519

PSSALRE helix domain in the N-terminal lobe, which isimportant for its interaction with its activator. A proteinmodel for the association Cdk5/p35 or Cdk5/p25 has beendesigned [31] on the basis of the crystallographic structureof the complex cdk2/cyclin A/ATP. This model agrees withthe different studies on the protein complex. This approachis very useful for the design of inhibitory substrates for thisenzyme, that appears to be implicated in neurodegenerativedisorders [31], and it may be of importance in drug designfor potential inhibitors with a neuroprotection action againstAlzheimer disease [1].
Functional organization of Cdk5 in cells
While Cdk5 is widely distributed in mammalian tissues andin cultured cell lines, Cdk5 associated kinase activity hasbeen demonstrated only in mammalian brains where p35 isa main regulatory factor. Cdk5 and its neuron specificregulator p35 are essential for neuronal migration in thecerebral cortex [45,46]. In addition Cdk5/p35 concentrates
at the leading edge of axonal growth cones thus regulatingneurite outgrowth in cortical neurons [22]. In addition to theneuronal localization of p35, Zheng et al. [47] have found asignificant level of expression of mRNA for p35 inmyoblasts, suggesting that p35 may also activate Cdk5during myogenesis. Cultures of cerebellar macroneuronswere used to study expression, subcellular localization andfunctions of neuronal Cdk5 activator during laminin-enhanced axonal growth. Laminin, an extracellular matrixcomponent that promotes axonal extension stimulates p35expression, thus increasing its interaction with the sub-cortical cytoskeleton and its redistribution to axonal growthcones [21,48]. It has been shown that this effect is mediatedby selective phosphorylation of MAP1b at proline directedprotein kinase epitope. In differentiating neurons, regula-tion of cell architecture is especially relevant, and tauprotein also plays a major role in the outgrowth of neuritesand axonal development [49]. There is strong evidence thattau activity in determining neuronal polarity is modulatedby protein Cdk5. In neuroblastoma cells it has been shownthat neurite extension induced by dibutyryl cAMP isassociated with an increase in Cdk5 expression [50]. Thisinduction of Cdk5 was concomitant with changes in taudistribution, and an increase in the microtubule assemblingactivity of neuronal extracts of cells undergoing differenti-ation. The functional significance of Cdk5/p35 is furtherevidenced by studies that indicate that suppression of p35by antisense oligonucleotides, reduced Cdk5 activity, MAP1bphosphorylation and laminin-induced axonal growth [21].Therefore, the Cdk5 system appears to be an importantregulatory link between extracellular signals such aslaminin and the intracellular organization of MAPs andother cytoskeletal proteins involved in axonal elongation.
On the other hand, studies suggest that besides Cdk5involvement in neuronal differentiation, the protein kinaseactivity of this enzyme is induced during myogenesis[51,52]. However, the mechanisms of how this activity ismodulated during muscle differentiation has not yet beenelucidated. Lazaro et al. [51] have examined the expression,enzyme activity and localization of Cdk5 during myo-genesis. In murine C2 cells, the levels of Cdk5 protein andits kinase activity showed a marked increase during theonset of myogenesis with a maximum at 36±48 h ofdifferentiation. It was also reported that Cdk5 exhibited acytoplasmic distribution in myoblasts under proliferation[52], with a rapid increment in the detection of Cdk5 in thecell nucleus during the first 12 h of differentiation, and amarked immunodetection of Cdk5 in the myotubes nuclei.An overexpression of Cdk5 has been detected in somemyopathies, with accumulation in vesicles of the musclefibers. It has also been reported that this abnormal increasein Cdk5 levels leads to aberrant phosphorylations of cyto-skeletal proteins, results that suggest that Cdk5 over-expression produces damage in muscle fibers and arrest themyogenetic process. In addition, Zhang and coworkers havedescribed elevated Cdk5 expression during mouse develop-ment in cells undergoing apoptosis in such diverse tissuesas intestinal epithelium, testis and peripheral nerve gangliasuggesting that Cdk5 activation may be a common featureof apoptosis in a variety of settings [53]. An activator hasbeen shown in Xenopus named to as Xp35.2, which isexpressed in differentiating neurons and at the initial stagesof the amphibian early nervous system development [54].
Fig. 1. Sequence alignment of the protein components of Cdk5
system. (A) Sequence alignment of p39 with respect to p35 and cyclin
A. The a-helix forming segments in cyclin A are denoted in red. The
box represents sequence predictions for a-helix in p35 [30]. The points
denote the aminoacids identity within the primary structure. (B).
Sequence alignment of Cdk5 with respect to cdk2. The segment
PSTAIRE involved in protein-to±protein interactions is denoted in red.
The box in the Cdk2 structure (T-loop) indicates the region involved in
cyclin interactions. The same region in Cdk5 participates in the
interaction with p35. Serine 159 in Cdk5 and Thr160 in cdk2 are
phosphorylation sites on this kinase. The points represent aminoacid
identity. P-sites are in red.
1520 R. B. Maccioni et al. (Eur. J. Biochem. 268) q FEBS 2001

In a developmental context, recent evidence indicates thatthe Cdk5/p35 system plays a major role in neuronalmigration [39]. Studies suggest the involvement of thiskinase system in regulating the N-cadherin-mediated celladhesion during cortical development [35].
Cdk-5 interacting proteins
A variety of proteins exhibit the capacity to interact withCdk5 and as a result of these interactions to promoteregulatory effects on Cdk5 enzymatic activity. Amongthese, a set of inhibitors of the Cdk5±p35 complex such asthe ribosomal L34 protein, and several other componentsconstitute major Cdk5-interacting proteins. These findingshave led to postulating Cdk5 as a key molecule in theregulation of the translational machinery [53]. A DNAbinding protein named to as dbpA exhibits the ability tointeract with the Cdk5±p35 complex, and inhibits thecomplex activity [55,56], thus demonstrating a potentialrole of Cdk5 at the nuclear level. This finding is inagreement with the localization of Cdk5 in the nuclei ofneuronal cells. In this context, it is of interest that tau hasbeen found in the nucleus of neuroblastoma cells [57] andalso extracted from nuclear preparations of other cell types[58]. The possible links between Cdk5 and the nuclear tauare not known. In this way Cdk5 appears to be involved inregulatory processes different from the regulatory role ofthis kinase at the cytoskeletal level [59]. In addition, it hasbeen shown that D type cyclins bind Cdk5 in fibroblastcells, but without displaying its protein kinase activity [60].
There is agreement on the involvement of three majoractivators for Cdk5 at the neuronal level: p39 [61] and p35[40]. In addition the Munc-18/p67 protein appears to haveregulatory roles on Cdk5 activity [62], The best character-ized Cdk5 activator is p35, a protein factor regulated by aset of molecular signals that affect its stability. Itsinvolvement is a subject of further analysis in the contextof Alzheimer pathology, and also in relation with its normalrole in brain development in another review article of thisissue [63].
The participation of p39 in Cdk5 regulation is crucialconsidering its major localization with cell structures suchas lamellipodia, filopodia in different types of cells and atthe level of growth cones in neuronal cells. This activatorcoimmunoprecipitates with brain actin revealing a specificlocalization of the kinase activity of Cdk5 complex [30].Another interesting feature of this activator is its localiz-ation with neuronal and glial cells, in contrast with p35found only in neurons [64]. In hippocampal cells, it wasshown that p39 is a protein necessary and sufficient forneuronal development of these cells [65], but the preciseregulatory mechanisms on how p39 modulates neurito-genesis have not been elucidated. Also, no informationexists yet on how p67 regulates neuronal development.These studies point to the complexity of the Cdk5 regu-latory mechanisms, and show that there is an open and widehorizon to explore in order to understand Cdk5 regulationin normal brain development and the alterations in theregulatory patterns in neurodegenerative disorders.
Proteins such as tau, synapsin, CK1, b and d-cateninsand N-cadherins [32±36] specifically interact with Cdk5 inconnection with different signal transduction pathways. Inthis context, it is of utmost importance to point out the
association of Cdk5 with the Rac GTPase, protein impli-cated in neuronal migration and growth. The Cdk5±p35complex interact directly with Rac via p35, in a GTPdependent fashion [66]. In addition, there is another proteinPak1 that also associates with this complex, thus leading toa relationship between the dynamics of actin cytoskeletonreorganization in neurons with the activity of Cdk5±p35complex. The mechanisms imply a Rac-dependent modu-lated phosphorylation of Pak1, with a decrease in Pak1kinase activity, and therefore an impact on the actin cyto-skeleton dynamics [66]. In addition to this set of proteins,double hybrid experiments have revealed the existence ofthree new p35-interacting components, which are widelyexpressed in human tissues [67]. A new protein designatedas GAK (cyclin G-associated kinase) coimmunoprecipitateswith Cdk5 and the cyclin G [68].
Regulation of the Cdk5±p35 complex
The regulation process of Cdk5±p35 has been well studiedand revealed several essential features helpful towardsunderstanding neuronal development. It has been shownthat p35 is regulated at the transcriptional level [69], butrecent studies have shown that p35 has a half-life (t1/2) of20±30 min. This would induce a decrease in t1/2 of theCdk5±p35 complex once this is formed, considering thatthe catalytic subunit of Cdk5 has a t1/2 of 90 min. Thestability of p35 increases 2±3 times in the presence ofagents such as roscovitine, a Cdk5 inhibitor and lacta-cystine a proteosome inhibitor [70]. This increase instability has been associated with apoptosis, alterations inthe cytoskeleton and neurodegeneration [2,70].
A recent study suggests that the regulation process of p35degradation is originated by an intrinsic phosphorylation ofp35 by Cdk5, which would activate ubiquitination and thesubsequent proteosomal degradation [70]. An importantfeature of p35 is that it is anchored in the cell membraneand is sensitive to proteolytic cleavage originated by twopolypeptides p10 and p25, where p25 appears to be themajor activator protein for Cdk5 kinase [20]. This truncatedprotein would have a greater stability than p35 [2,70]. Theproteolytic processing of this activator is originated bycalpain, which produces an increase in p25 during oxidativestress, or an increase in calcium concentration and thereforean increment in Cdk5 activity, a phenomenon that has beenproposed to be linked with neurodegeneration [3,24].
The Cdk5 in Alzheimer pathology
Neurodegenerative diseases constitute a heterogeneous groupof brain disorders leading to dementia such as in Alzheimerdisease. They share in common the neurofibrillary degene-ration affecting neurons of the central nervous system,process that include decrease in cholinergic transmission,higher sensitivity to oxidative stress, alterations in thecytoskeleton and neuronal death [71±73]. Alzheimer diseaseis one of the most common types of dementia disordersaffecting the elderly, characterized by the formation of twomain protein aggregates: senile plaques and neurofibrillarytangles [72]. Senile plaques are mainly composed of fibrilsof the b-amyloid (Ab), a peptide derived from the proteo-lytic processing of the amyloid precursor protein (APP),through the action of a, b and g-secretases [74]. The
q FEBS 2001 Structure, function and pathology of protein kinase Cdk5 (Eur. J. Biochem. 268) 1521

a-secretase has a neuroprotective action as it cleaves APPin positions that prevent Ab formation. However cleavageby b and g-secretases produces Ab (1±40), which accountsfor around 90% of secreted Ab peptide. The remainingfraction corresponds to Ab(1±42) with a significantlyincreased capacity to generate fibrils, and therefore toincorporate into senile plaques [75]. APP mutations linkedto familial Alzheimer's disease (FAD) modify the activityof secretases and leads to an increased production of Ab.Other factors involved in FAD are presenilins (PS1 andPS2), proteins related to normal development of thenervous system, and that as a result of different types ofmutations, affect the normal APP processing [76]. Pre-senilins interact with b and d-catenins [77] involved in celladhesion functions in the pathway of Wnt signaling,essential for embryonic development, and are controlledin neurons by the Ser/Thr protein kinase GSK3b amongother regulators [78]. GSK3b is one the main kinasesinvolved in aberrant phosphorylation on tau and appears tobe implicated in tau pathology. This kinase interacts withpresenilin PS1 suggesting its involvement in the complexregulatory pathway of tau phosphorylation [79]. Besides theeffects of mutated form of PS1 on the activity of secretases,this protein seems to be involved in neurodegeneration as itinduces apoptosis and neuronal death, as a result of the lossof PS1 interaction with the antiapoptotic protein such asBcl-Xl [80].
Tau protein is the major component of PHFs, filamentsthat form a compact filamentous network called neuro-fibrillary tangles. Tau belongs to the family of microtubule-associated proteins (MAPs) (reviewed in [49]), it is theproduct of alternative splicing of a single gene leading tothe expression of six isotypes in neuronal axons of humanbrains [81±83]. A variable number of tau isoforms in othermammalian species and cultured cell lines have been found.The different tau isoforms contain either three or fourmicrotubule-binding repeats, and their presence in fetalbrain (isoforms with 3-repeats) or in adult brain (isoformswith 4-repeats) support the functional significance of tauisospecies as related with brain development [49,81]. In thecontext of Alzheimer's disease, no mutations have beenreported for tau molecule. However, different mutated tauforms have been identified in other neurodegenerativedisorders such as frontotemporal dementia and parkinson-ism linked to chromosome-17 (FTDP-17) [84,85], wheretau mutations at sites distributed in different proteindomains result in alterations of its conformation andcapacity to bind microtubules. Tau expression is develop-mentally regulated, and tau phosphorylation at sites that donot occur in the adult nervous system has been demon-strated in fetal neuronal tissue, with some similarities in themodification patterns to those exhibited for PHFs tau[4,86]. In normal brain, the equilibrium between tau phos-phorylation and dephosphorylation modulates the stabilityof cytoskeleton and thereby the axonal morphology. There-fore, the breakdown of this equilibrium under pathologicalconditions such as those of neurodegenerative disorders,results in deleterious consequences for the mature neuronalcells [6,49]. The earliest modification found in Alzheimerbrains is tau phosphorylation. The Alzheimer's phospho-epitopes corresponds to some of the tau sites for Cdk5 andGSK3b, which normally are not modified or modified at alow level, studies that point to the importance of the in vitro
and in vivo research on these protein kinases. Anomalousphosphorylations may be accompanied with other modifi-cation reactions linked with tau pathology, such as ubiquiti-nation, binding of proteoglycans and glycations at specificlysine residues on this protein [10]. These modificationsparticipate at later stages in the abnormal pathway leadingto PHFs assembly. In particular, tau glycation results indramatic changes in tau association with the cytoskeleton[10].
The hyperphosphorylation on tau by the actions ofdifferent protein kinase and phosphatase systems appear tolead to structural and conformational changes on tau, thusaffecting its binding with tubulin and capacity to promotemicrotubule assembly [6,87]. Even though there is cumu-lative information on the involvement of activated tauphosphorylating kinases in neurodegeneration, the preciserole and the nature of kinases in Alzheimer's disease hasnot been yet elucidated. The relevant protein kinasesinvolved in tau modifications as related with neurofibrillarydegeneration are GSK3b [5,88], PKA, MAP-kinase, MARKkinase and the Cdk5/p35 system [1,2,8,89]. Proline-directedkinase Cdk5 phosphorylates tau at residues Ser202, Thr205,Ser235 and Ser404 [89], while the sites sensitive to GSK3bappears to be Ser199, Ser396 and Ser413 [88]. Phos-phorylation has been detected at Ser396 and Ser404 whichare involved in the PHF-1 epitope [90]. Therefore, differentphosphorylated sites have been detected in PHFs purifiedfrom Alzheimer brains [6,89]. Evidence based on studies inhippocampal cultures support the idea that in neurodegene-ration, the initial anomalous tau phosphorylation by Cdk5stimulate subsequent modifications by GSK3b, thus pre-venting tau from incorporating into microtubules [1,91,92].
In Alzheimer disease, amyloid deposition is one of theprocesses that trigger the cascade of molecular eventsleading to neurofibrillary degeneration, however, senileplaques themselves is not the only factor triggeringneuronal death. In the context of the preceding information,it seems reasonable to think that the reactions leading toneuronal death in this disease are part of an integratedprocess involving extracellular and intracellular alterationsin the normal neuronal machinery. Ab deposition, oxidativestress, changes in the ECM appears to be among triggeringelements. Studies that link neurotoxic action of Ab withintracellular events leading to tau self-aggregation intoPHFs and apoptotic phenomena, are of interest towardelucidation of the molecular puzzle of Alzheimer disease.Busciglio and coworkers succeeded in demonstrating thatincubation of primary cultured hipoccampal cells with Abactivated tau phosphorylation [93]. Studies on Ab linkswith Cdk5 led to demonstration of an unusual Cdk5 acti-vation [1], concomitant with changes in the intraneuronaltau distribution [23] and phosphorylation at Cdk5 epitopeson tau. Deregulation of Cdk5 seems to involve formation ofa stable complex with p35, a process depending on Cdk5phosphorylation [23]. Interestingly, deregulation of Cdk5 isconcomitant with neuronal death of rat hippocampal neurons,but pretreatment of cells with Cdk5 specific inhibitors suchas butyrolactone I or by using antisense probes for Cdk5,protected cells against neuronal death [1]. Another com-pound, the bis-indole indirubin inhibits GSK3b and alsoCdk5/p25 complex with a concomitant inhibition in tauphsophorylation at those epitopes found in Alzheimerdisease [94]. Neuroprotection of cells to the neurotoxic
1522 R. B. Maccioni et al. (Eur. J. Biochem. 268) q FEBS 2001
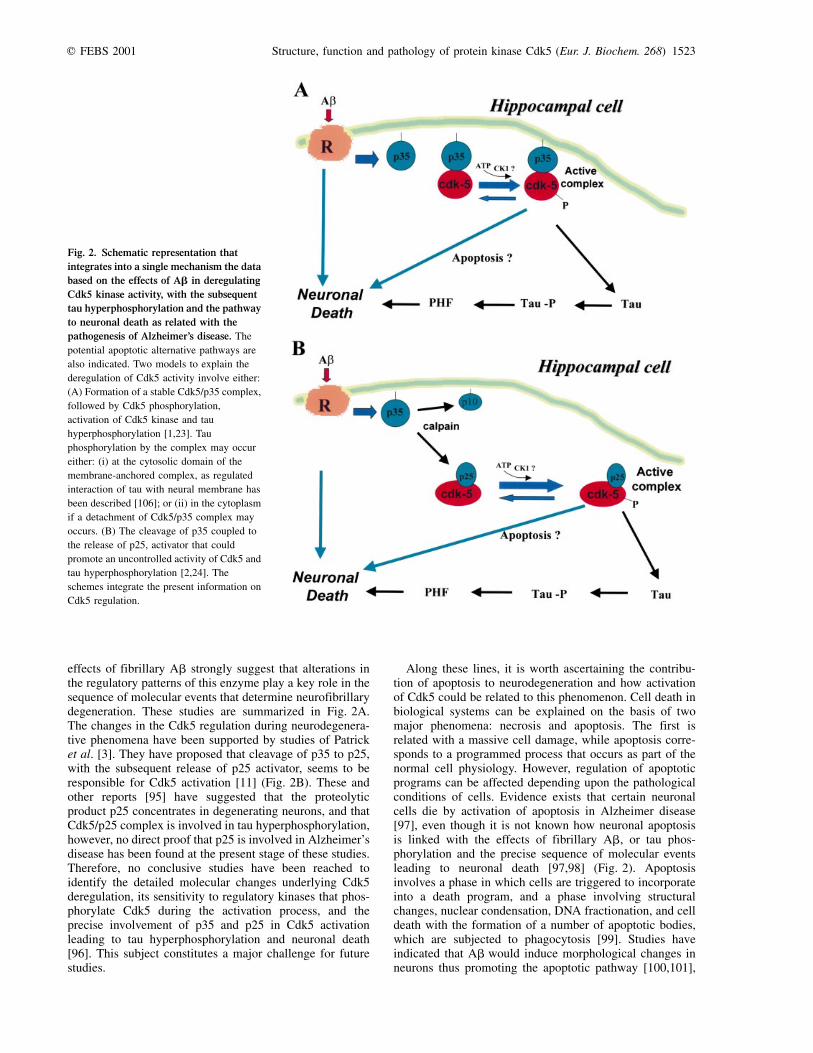
effects of fibrillary Ab strongly suggest that alterations inthe regulatory patterns of this enzyme play a key role in thesequence of molecular events that determine neurofibrillarydegeneration. These studies are summarized in Fig. 2A.The changes in the Cdk5 regulation during neurodegenera-tive phenomena have been supported by studies of Patricket al. [3]. They have proposed that cleavage of p35 to p25,with the subsequent release of p25 activator, seems to beresponsible for Cdk5 activation [11] (Fig. 2B). These andother reports [95] have suggested that the proteolyticproduct p25 concentrates in degenerating neurons, and thatCdk5/p25 complex is involved in tau hyperphosphorylation,however, no direct proof that p25 is involved in Alzheimer'sdisease has been found at the present stage of these studies.Therefore, no conclusive studies have been reached toidentify the detailed molecular changes underlying Cdk5deregulation, its sensitivity to regulatory kinases that phos-phorylate Cdk5 during the activation process, and theprecise involvement of p35 and p25 in Cdk5 activationleading to tau hyperphosphorylation and neuronal death[96]. This subject constitutes a major challenge for futurestudies.
Along these lines, it is worth ascertaining the contribu-tion of apoptosis to neurodegeneration and how activationof Cdk5 could be related to this phenomenon. Cell death inbiological systems can be explained on the basis of twomajor phenomena: necrosis and apoptosis. The first isrelated with a massive cell damage, while apoptosis corre-sponds to a programmed process that occurs as part of thenormal cell physiology. However, regulation of apoptoticprograms can be affected depending upon the pathologicalconditions of cells. Evidence exists that certain neuronalcells die by activation of apoptosis in Alzheimer disease[97], even though it is not known how neuronal apoptosisis linked with the effects of fibrillary Ab, or tau phos-phorylation and the precise sequence of molecular eventsleading to neuronal death [97,98] (Fig. 2). Apoptosisinvolves a phase in which cells are triggered to incorporateinto a death program, and a phase involving structuralchanges, nuclear condensation, DNA fractionation, and celldeath with the formation of a number of apoptotic bodies,which are subjected to phagocytosis [99]. Studies haveindicated that Ab would induce morphological changes inneurons thus promoting the apoptotic pathway [100,101],
Fig. 2. Schematic representation that
integrates into a single mechanism the data
based on the effects of Ab in deregulating
Cdk5 kinase activity, with the subsequent
tau hyperphosphorylation and the pathway
to neuronal death as related with the
pathogenesis of Alzheimer's disease. The
potential apoptotic alternative pathways are
also indicated. Two models to explain the
deregulation of Cdk5 activity involve either:
(A) Formation of a stable Cdk5/p35 complex,
followed by Cdk5 phosphorylation,
activation of Cdk5 kinase and tau
hyperphosphorylation [1,23]. Tau
phosphorylation by the complex may occur
either: (i) at the cytosolic domain of the
membrane-anchored complex, as regulated
interaction of tau with neural membrane has
been described [106]; or (ii) in the cytoplasm
if a detachment of Cdk5/p35 complex may
occurs. (B) The cleavage of p35 coupled to
the release of p25, activator that could
promote an uncontrolled activity of Cdk5 and
tau hyperphosphorylation [2,24]. The
schemes integrate the present information on
Cdk5 regulation.
q FEBS 2001 Structure, function and pathology of protein kinase Cdk5 (Eur. J. Biochem. 268) 1523

and that the cysteine aspartate proteases (caspases) play acritical role in Ab-apoptosis. Caspase inhibitors appear toblock the Ab-induced apoptotic process. Caspase 3 hasbeen directly involved in Ab-induced apoptosis of corticalneurons [102], and most recently, caspase 2 has beeninvolved in the mechanism of neuronal death caused byincubation with fibrillary Ab [103]. In addition, neuronalcells from knock-out mice for caspase 2 was resistant to theneurotoxic action of Ab, suggesting the involvement of thisenzyme in Ab-induced neuronal death [103]. Neuronaldeath can be a result of an activation of apoptotic programsinvolving the biochemical mechanisms underlying thisprocess. On the other hand, there is an integrated view thatAb and other extracellular signals may induce intracellularchanges involving anomalous tau phosphorylation, PHFsand neuronal death independently of the activity of caspases.Other lines of evidence link apoptosis with an overexpressionof Cdk5 but no other cdk's [104], especially in the contextof brain development [53]. Even though cells of thesubstantia nigra rather than hippocampal cells were thebiological system, these studies provide useful informationon the relationships between neuronal apoptosis [105]. Thepossible relationships between brain protein kinases, Cdk5deregulation, tau phosphorylation and apoptosis in the con-text of Alzheimer's disease are not understood. Elucidationof this and many other molecular and celullar problems, aswell as the projection of the in vitro and in vivo studiesdeveloped in several biological systems and animal modelsto the human brain constitute a major challenge for futureAlzheimer's research.
A C K N O W L E D G E M E N T S
This review has been prepared within the framework of the program of
the Millennium Institute for Advanced Studies in Cell Biology and
Biotechnology (CBB), and supported by project of the Millennium
Science Initiative P-99±031F MIDEPLAN, Chile. Support from a
Presidential Chair in Science and a Fondecyt grant 199±0002 (to
R. B. M.) are acknowledged.
R E F E R E N C E S
1. Alvarez, A., Toro, R., CaÂceres, A. & Maccioni, R.B. (1999)
Inhibition of tau phosphorylating protein kinase Cdk5 prevents
beta-amyloid induced neuronal death. FEBS Lett. 459, 421±426.
2. Patrick, G.N., Zuckerberg, L., Nikolic, M., de la Monte, S.,
Dikkes, P. & Tsai, L.H. (1999) Conversion of p35 to p25
deregulates Cdk5 activity and promotes neurodegeneration.
Nature 402, 605±622.
3. Kusakawa, G.I., Saito, T., Onuki, R., Ishiguro, K., Kishimoto, T.
& Hisanaga, S. (2000) Calpain-dependent proteolytic cleavage of
the p35 cyclin dependent kinase 5 activator to p25. J. Biol. Chem.
275, 17166±17172.
4. Schneider, A., Biernat, J., von Bergen, M., Mandelkow, E. &
Mandelkow, E.M. (2000) Phosphorylation that detaches tau
protein from microtubules (Ser262,Ser214) also protects it
against aggregation into Alzheimer paired helical filaments.
Biochemistry 38, 3549±3558.
5. Godeman, R., Biernat, J., Mandelkow, E. & Mandelkow, E.M.
(1999) Phosphorylation of tau protein by recombinant GSK3b:
pronounced phosphorylation at select Ser/ Thr-Pro motifs but no
phosphorylation at Ser262 in the repeat domain. FEBS Lett. 454,
157±164.
6. Mandelkow, E.M., Biernat, J., Drewes, G., Gustke, N., Trinczek,
B. & Mandelkow, E. (1995) Tau domains, phosphorylation, and
interactions with microtubules. Neurobiol. Aging 16, 355±363.
7. Ferreira, A., Lu, Q., Orecchio, L. & Kosik, K. (1997) Selective
phosphorylation of adult tau isoforms in mature hippocampal
neurons exposed to fibrillar Ab. Mol. Cell Neurosci. 9, 220±234.
8. Chin, J.Y., Knowles, R.B., Schneider, A., Drewes, G., Man-
delkow, E.M. & Hyman, B.T. (2000) Microtubule-affinity
regulating kinase (MARK) is tightly associated with neuro-
fibrillary tangles in Alzheimer's brains: a fluorescence resonance
energy transfer study. J. Neuropathol. Exp. Neurol. 59, 966±971.
9. Alonso, A., Grundke-Iqbal, I. & Iqbal, K. (1997) Abnormal
phosphorylation of tau, and the mechanisms of Alzheimer
neurofibrillary degeneration: sequestration of MAP1 and MAP2
and the disassembly of microtubules by abnormal tau. Proc. Natl
Acad. Sci. USA 94, 298±303.
10. GonzaÂlez, C., FarõÂas, G. & Maccioni, R.B. (1998) Modification
of tau to an Alzheimer's protein interferes with interaction with
microtubules. Cell. Mol. Biol. 44, 1117±1127.
11. Kobayashi, S., Ishiguro, K., Omori, A., Takamatsu, M., Arioka,
M., Imahori, K. & Uchida, T. (1993) A Cdc2 related kinase
PSSALRE/Cdk5 is homologous with the 30 kDa subunit of tau
protein kinase II, a proline directed kinase associated with
microtubules. FEBS Lett. 335, 171±175.
12. Sun, D., Leung, C.L. & Liem, R.K. (1996) Phosphorylation of the
high molecular weight neurofilament protein (NF-H) by Cdk5
and p35. J. Biol. Chem. 271, 14245±14251.
13. Shuang, R., Zhang, L., Fletcher, A., Groblewski, G., Pevsner, J. &
Stuenkel, E.L. (1998) Regulation of Munc-18/Synthaxin 1A
interaction by cyclin-dependent kinase 5 in nerve endings. J. Biol.
Chem. 273, 4957±4966.
14. Shetty, K.T., Link, W.T. & Pant, H.C. (1993) Cdc2-like kinase
from rat spinal cord specifically phosphorylates KSPXK motifs
in neurofilament proteins: isolation and characterization. Proc.
Natl Acad. Sci. USA 90, 6844±6848.
15. Meyerson, M., Enders, G.H., Wu, C.L., Gorka, C., Nelson, C.,
Harlow, E. & Tsai, L.H. (1992) A family of human Cdc2-related
protein kinases. EMBO J. 11, 2909±2917.
16. Lew, J., Beaudette, K., Litwin, C.M. & Wang, J.H. (1992)
Purification and characterization of a novel proline directed
kinase from bovine brain. J. Biol. Chem. 267, 13383±13390.
17. Tsai, L.H., Takahashi, T., Caviness, V. & Harlow, E. (1993)
Activity and expression patterns of cyclin-dependent kinase 5
in the embryonic mouse nervous system. Development 119,
1029±1040.
18. Paudel, H.K., Lew, J., Ali, Z. & Wang, J. (1993) Brain proline-
directed protein kinase phosphorylates tau on sites that are
abnormally phosphorylated in tau associated with Alzheimer's
paired helical filaments. J. Biol. Chem. 268, 23512±23518.
19. Tsai, L.H., Delalle, I., Caviness, V.S., Chae, T. & Harlow, E.
(1994) p35 is a neural-specific regulatory subunit of cyclin-
dependent kinase 5. Nature 371, 419±423.
20. Lew, J., Huang, Q., Qi, Z., Winkfein, R.J., Aebersold, R., Hunt, T.
& Wang, J.H. (1994) A brain-specific activator of cyclin-
dependent kinase 5. Nature 371, 423±426.
21. Paglini, G., Pigino, G., Kunda, P., Morfini, G., Maccioni, R.B.,
Quiroga, S., Ferreira, A. & CaÂceres, A. (1998) Evidence for the
participation of the neuron-specific Cdk5 activator p35 during
laminin-enhanced axonal growth. J. Neurosci. 18, 9858±9869.
22. Pigino, G., Paglini, G., Ulloa, L., Avila, J. & CaÂceres, A. (1997)
Analysis of the expression, distribution and function of cyclin
dependent kinase 5 (Cdk5) in developing cerebellar macro-
neurons. J. Cell Sci. 110, 257±270.
23. Alvarez, A., Toro, R., MunÄoz, J.P. & Maccioni, R.B. (1999)
Inhibition of tau phosphorylating protein kinase Cdk5 prevents
Ab-induced neuronal death in Alzheimer's neuronal models. Mol
Biol. Cell 10, 61a.
1524 R. B. Maccioni et al. (Eur. J. Biochem. 268) q FEBS 2001

24. Lee, M.S., Kwon, Y., Li, M., Peng, J., Friedlander, R.M. & Tsai,
L.H. (2000) Neurotoxicity induces cleavage of p35 to p25 by
calpain. Nature 405, 360±364.
25. Gong, C., Lidsky, T., Wegiel, J., Zuck, L., Grundke-Iqbal, I. &
Iqbal, K. (2000) Phosphorylation of tau is regulated by protein
phosphatase 2A in mammalian brain. Implications for neuro-
fibrillary degeneration in Alzheimer's disease. J. Biol. Chem.
275, 5535±5544.
26. Van den Heuvel, S. & Harlow, E. (1993) Distinct roles for cyclin-
dependent kinases in cell cycle control. Science 262, 2050±2054.
27. Fisher, R.P. & Morgan, D.O. (1994) A novel cyclin associates
with MO15/cdk7 to form the cdk-activating kinase. Cell 78,
713±724.
28. King, R., Jackson, P. & Kirschner, M.W. (1994) Mitosis in
Transition. Cell 79, 563±571.
29. Sherr, C.J. (1994) G1 phase progression: cycling the cue. Cell 79,
551±555.
30. Humbert, S., Dhavan, R. & Tsai, L. (2000) p39 activates Cdk5 in
neurons, and is associated with the actin cytoskeleton. J. Cell Sci.
113, 975±983.
31. Chou, K.C., Watenpaugh, K.D. & Heinrikson, R.L. (1999) A
model of the complex between cyclin-dependent kinase 5 and the
activation domain of neuronal Cdk5 activator. Biochem. Biophys.
Res. Commun. 259, 420±428.
32. Rosales, J.L., Nodwell, M.J., Johnston, R.N. & Lee, K.Y. (2000)
Cdk5/p25 (nck5a) interaction with synaptic proteins in bovine
brain. J.Cell Biochem. 78, 151±159.
33. Sobue, K., Agarwal-Mawal, A., Li, W., Sun, W., Miura, Y. &
Paudel, H.K. (2000) Interaction of neuronal Cdc2-like protein
kinase with microtubule-associated protein tau. J. Biol. Chem.
275, 16673±16680.
34. Lee, K.Y., Rosales, J., Tang, D. & Wang, J.H. (1996) Interaction
of cyclin-dependent kinase 5 (Cdk5) and neuronal Cdk5 activator
in bovine brain. J. Biol. Chem. 271, 1538±1543.
35. Kwon, Y.T., Gupta, A., Zhou, Y., Nikolic, M. & Tsai, L.H. (2000)
Regulation of N-cadherin-mediated adhesion by the p35-Cdk5
kinase. Curr. Biol. 10, 363±372.
36. Takahashi, M., Amin, N., Grant, P. & Pant, H.C. (1995) P13suc1
associates with a Cdc2-like kinase in a multimeric cytoskeletal
complex in squid axoplasm Journal of. Neuroscience 15,
6222±6229.
37. Morgan, D.O. (1997) Cyclin-dependent kinases: engines, clocks,
and microprocessors. Annu. Rev. Cell Dev. Biol. 13, 261±291.
38. Sharma, P., Sharma, M., Amin, N.D., Albers, R.W. & Pant, H.C.
(1999) Regulation of cyclin-dependent kinase 5 catalytic activity
by phosphorylation. Proc. Natl Acad. Sci. USA 96, 11156±11160.
39. Homayouni, R. & Curran, T. (2000) Cortical development: Cdk5
gets into sticky situations. Current Biol. 10, R331±R334.
40. Tang, D., Chun, A.C., Zhang, M. & Wang, J.H. (1997) Cyclin-
dependent kinase 5 (Cdk5) activation domain of neuronal Cdk5
activator. Evidence of the existence of cyclin fold in neuronal
Cdk5a activator. J. Biol. Chem 272, 12318±12327.
41. Poon, R.Y., Lew, J. & Hunter, T. (1997) Identification of
functional domains in the neuronal Cdk5 activator protein. J. Biol.
Chem. 272, 5703±5708.
42. Sharma, P., Steinbach, P.J., Sharma, M., Amin, N.D., Barchi, J.J.
Jr & Pant, H.C. (1999) Identification of substrate binding site of
cyclin-dependent kinase 5. J. Biol. Chem. 274, 9600±9606.
43. Russo, A.A., Jeffrey, P.D. & Pavletich, N.P. (1996) Structural
basis of cyclin-dependent kinase activation by phosphorylation.
Nat. Struct. Biol. 3, 696±700.
44. Jeffrey, P.D., Russo, A.A., Polyak, K., Gibbs, E., Hurwitz, J.,
MassagueÂ, J. & Pavletich, N.P. (1995) Mechanism of cdk
activation revealed by the structure of a cyclin-cdk2 complex.
Nature 376, 313±320.
45. Oshima, T., Ward, J.M., Huh, C.G., Longenecker, G., Veeranna,
C., Pant, H.C., Brady, R.O., Martin, L.J. & Kulkarni, A.B. (1996)
Targeted disruption of the cyclin-dependent kinase 5 gene results
in abnormal corticogenesis, neuronal pathology and perinathan
death. Proc. Natl Acad. Sci. USA 93, 11173±11178.
46. Chae, T., Kwon, Y.T., Bronson, R., Dikkes, P., Li, E. & Tsai, L.H.
(1997) Mice lacking p35, a neuronal specific activator of Cdk5
display cortical lamination defects, seizures, and adult lethality.
Neuron 18, 29±42.
47. Zheng, M., Leung, C. & Liem, R. (1998) Region-specific
expression of cyclin dependent kinase 5 (Cdk5) and its activators,
p35 and p39, in developing and adult rat central nervous system.
J. Neurobiol. 35, 141±159.
48. Ramirez, G., Alvarez, A., GarcõÂa, J., Gomes, F., Moura-Neto, V.
& Maccioni, R.B. (1999) Regulatory roles of microtubule-
associated proteins in neuronal morphogenesis. Involvement of
extracellular matrix. Braz. J. Med. Biol. Res. 32, 611±618.
49. Maccioni, R.B. & Cambiazo, V. (1995) Role of microtubule-
associated proteins in the control of microtubule assembly.
Pysiol. Revs. 23, 1±27.
50. MunÄoz, J.P., Alvarez, A. & Maccioni, R.B. (2000) Increase in the
expression of the neuronal cyclin-dependent protein kinase Cdk5
during differentiation of N2A neuroblastoma cells. Neuroreport
11, 12±21.
51. Lazaro, J.B., Kitzman, M., Poul, M.A., Vandromme, M., Lamb,
N.Y. & Fernandez, A. (1997) Cyclin-dependent kinase 5, Cdk5, is
a positive regulator of myogenesis in mouse C2 cells. J. Cell Sci.
110, 1251±1260.
52. Philpott, A., Porro, E., Kirschner, M.W. & Tsai, L. (1997) The
role of cyclin-dependent kinase 5 and a novel regulatory subunit
in regulating muscle differentiation and pattering. Genes Dev. 11,
1409±1421.
53. Zhang, Q., Ahuja, H.S., Zakeri, Z.F. & Wolgemuth, D.J. (1997)
Cyclin-dependent kinase 5 is associated with apoptotic cell death
during development and tissue remodeling. Dev Biol. 183,
222±233.
54. Philpott, A., Tsai, L. & Kirschner, M.W. (1999) Neuronal
differentiation and patterning in Xenopus: the role of Cdk5 and a
novel activator xp35.2. Dev. Biol. 207, 119±132.
55. Moorthamer, M., Zumstein-Mecker, S. & Chaudhuri, B. (1999)
DNA binding protein dbpA binds Cdk5 and inhibits its activity.
FEBS Lett 446, 343±350.
56. Moorthamer, M. & Chaudhuri, B. (1999) Identification of
ribosomal protein L34 as a novel Cdk5 inhibitor. Biochem.
Biophys. Res. Commun 255, 631±638.
57. Thurston, V., Pena, P., Pestell, R. & Binder, L.I. (1997) Nucleolar
localization of the microtubule-associated protein tau in neuro-
blastoma using sense and anti-sense transfection strategies. Cell
Motil. Cytoskel. 38, 100±110.
58. Cross, D., MunÄoz, J.P., HernaÂndez, P. & Maccioni, R.B. (2000)
Nuclear and cytoplasmic tau proteins from human nonneuronal
cells share common structural and functional features. J. Cell
Biochem. 78, 305±317.
59. Ino, H. & Chiba, T. (1996) Intracellular localization of cyclin-
dependent kinase 5 in mouse neuron: CDK5 is located in both
nucleus and cytoplasm. Brain Res. 732, 179±185.
60. Xiong, Y., Zhang, H. & Beach, D. (1992) D type cyclins associate
with multiple protein kinases and the DNA replication and repair
factor PCNA. Cell 71, 505±514.
61. Cai, X.H., Tomizawa, K., Tang, D., Lu, Y.F., Moriwaki, A.,
Tokuda, M., Nagahata, S., Hatase, O. & Matsui, H. (1997)
Changes in the expression of novel Cdk5 activator messenger
RNA (p39nck5ai mRNA) during rat brain development. Neurosci
Res. 28, 355±360.
62. Veeranna, A., Grant, P. & Pant, H.C. (1997) Expression of p67
(Munc-18), Cdk5, P-NFH and syntaxin during development of
the rat cerebellum. Dev Neurosci. 19, 172±183.
63. Paglini, G. & CaÂceres, A. (2001) The role of the Cdk5±p35
kinase in neuronal development. Eur. J. Biochem. 268, 1528±1533.
q FEBS 2001 Structure, function and pathology of protein kinase Cdk5 (Eur. J. Biochem. 268) 1525

64. Honjyo, Y., Kawamoto, Y., Nakamura, S., Nakano, S. &
Akigushi, I. (1999) Immunohistochemical localization of Cdk5
activator p39 in the rat brain. Neuroreport 10, 3375±3379.
65. Xiong, W., Pestell, R. & Rosner, M.R. (1997) Role of cyclins in
neuronal differentiation of immortalized hippocampal cells. Mol
Cell Biol. 17, 6585±6597.
66. Nikolic, M., Chou, M.M., Lu, W., Mayer, B.J. & Tsai, L.H.
(1998) The p35/Cdk5 kinase is a neuron-specific Rac effector
that inhibits Pak1 activity. Nature 395, 194±198.
67. Ching, Y.P., Qi, Z. & Wang, J.H. (2000) Cloning of three novel
neuronal Cdk5 activator binding proteins. Gene 242, 285±294.
68. Kanaoka, Y., Kimura, S.H., Okazaki, I., Ikeda, M. & Nojima, H.
(1997) GAK: a cyclin G associated kinase contains a tensin/
auxilin-like domain. FEBS Lett. 402, 73±80.
69. Delalle, I., Bhide, P.G., Caviness, V.S. & Tsai, L.H. (1997)
Temporal and spatial patterns of expression of p35, a regulatory
subunit of cyclin-dependent kinase 5, in the nervous system of
the mouse. J. Neurocytol. 26, 283±296.
70. Patrick, G.N., Zhou, P., Kwon, Y.T., Howley, P.M. & Tsai, L.H.
(1998) p35, the neuronal-specific activator of cyclin-dependent
kinase 5 (Cdk5) is degraded by the ubiquitin-proteasome
pathway. J. Biol. Chem. 273, 24057±24064.
71. Morrison, J.H. & Hof, P. (1997) Life and death of neurons in the
aging brain. Science 278, 412±419.
72. Maccioni, R.B. & Rodrigues, P. (1995) Alzheimer disease: an
attack of protein aggregates to human brain. Ciencia Hoje 20,
34±40.
73. Rossner, S., Ueberham, U., Schliebs, R., Perez-Polo, J. & Bigl, V.
(1998) The regulation of amyloid precursor protein metabolism
by cholinergic mechanisms and neurotrophin receptor signaling.
Prog. Neurobiol. 56, 541±569.
74. Pike, C., Overman, M.J. & Cotman, C.W. (1995) Amino-terminal
deletions enhance aggregation of b-amyloid peptide in vitro. J.
Biol. Chem. 270, 23895±23898.
75. Citron, M., Diehl, T., Gordon, G., Biere, A., Seubert, P. & Selkoe,
D. (1996) Evidence that the 42 and 40 aminoacids forms of
amyloid b protein are generated from the b-amyloid precursor
protein by different protease activities. Proc. Natl Acad. Sci. USA
93, 13170±13175.
76. Selkoe, D. (1999) Translating cell biology into therapeutic
advances in Alzheimer's disease. Nature 399, A23±A31.
77. Noll, E., Medina, M., Hartley, D., Zhou, J., Perrimon, N. &
Kosik, K.S. (2000) Presenilin affects Arm/beta-catenin localiz-
ation and function in Drosophila. Dev. Biol. 227, 450±464.
78. Sherrington, R., Rogaev, E., Liang, Y., Rogaev, E.A., Levesque,
G., Ikeda, M., Chi, H., Lin, C., Li, G. & Holman, K. (1995)
Cloning of a gene bearing missense mutations in early-onset
familial Alzheimer's disease. Nature 375, 754±760.
79. Takashima, A., Murayama, M., Murayama, O., Kohno, T.,
Honda, T., Yasatuka, K., Nihonmatsu, N., Mercken, M.,
Yamagushi, H., Sugihara, S. & Wolozin, B. (1998) Presenilin 1
associates with glycogen synthase 3 beta and its substrate tau.
Proc. Natl Acad. Sci. USA 95, 9637±9641.
80. Passer, B.J., Pellegrini, L., Vito, P., Ganjei, J.K. & DaÂdamo, L.
(1999) Interaction of Alzheimer's presenilin-1 and presenilin-2
with Bcl-XL. J. Biol. Chem. 274, 24007±24013.
81. Goedert, M., Spillantini, M.G., Jakes, R., Rutherford, D. &
Crowther, R.A. (1989) Multiple isoforms of human microtubule-
associated protein tau: sequences and localization in neurofi-
brillary tangles of Alzheimer's disease. Neuron 3, 519±526.
82. Andreadis, A., Brown, M.W. & Kosik, K.S. (1992) Structure
and novel exons of the human tau gene. Biochemistry 31,
10626±10633.
83. Maccioni, R.B., Vera, J.C., Dominguez, J. & Avila, J. (1989) A
discrete sequence defines a tubulin binding domain on micro-
tubule-associated protein tau. Arch. Biochem. Biophys. 275,
568±579.
84. Spillantini, M.G. & Goedert, M. (1998) Tau protein pathology in
neurodegenerative diseases. Trends Neurosci. 21, 428±433.
85. Delacourte, A. (1999) Biochemical and molecular characteriz-
ation of neurofibrillary degeneration in frontotemporal dementias.
Dement. Geriatr. Cogn. Disord. 10 (Suppl. 1), 75±79.
86. Schweers, O., Mandelkow, E.M., Biernat, J. & Mandelkow, E.
(1995) Oxidation of cysteine-322 in the repeat domain of
microtubule-associated protein tau controls the in vitro assembly
of paired helical filaments. Proc. Natl Acad. Sci. USA 92,
8463±8467.
87. Von Bergen, M., : Friedhoff, P., Biernat, J., Heberle, J.,
Mandelkow, E.M. & Mandelkow, E. (2000) Assembly of tau
protein into Alzheimer paired helical filaments depends on a
local sequence motif (306VQIVYK311) forming beta structure.
Proc. Natl Acad. Sci. USA 97, 5129±5134.
88. Imahori, K. & Uchida, T. (1997) Physiology and pathology of tau
protein kinases in relation to Alzheimer's disease. J. Biochem.
121, 179±188.
89. Michel, G., Mercken, M., Murayama, M., Nogushi, K., Ishiguro,
K., Imahori, K. & Takashima, A. (1998) Characterization of tau
phosphorylation in GSK3b and cyclin dependent kinase-5
activator transfected cells. Biochim. Biophys. Acta 1380,
177±182.
90. Illenberger, S., Zhen-Frischhofer, Q., Preuss, U., Stamer, K.,
Bauman, K., Trincczek, B., Biernat, J., Godeman, R.,
Mandelkow, E.M. & Mandelkow, E. (1998) The endogenous
and cell cycle-dependent phosphorylation of tau protein in living
cells: implications for Alzheimer's disease. Mol. Biol. Cell 9,
1495±1512.
91. Sengupta, A., Wu, Q., Grundke-Iqbal, I., Iqbal, K. & Singh, T.
(1997) Alzheimer-like phosphorylation of human tau by Cdk5.
Mol. Cell. Biochem. 167, 99±105.
92. Alvarez, G., MunÄoz, J., Satrustegui, J., Avila, J., Bogonez, E. &
Diaz-Nido, J. (1999) Lithium protects cultured neurons
against beta-amyloid-induced neurodegeneration. FEBS Lett.
453, 260±264.
93. Busciglio, J., Lorenzo, A., Yeh, J. & Yankner, B.A. (1995)
Beta-amyloid fibrils induce tau phosphorylation and loss of
microtubule binding. Neuron 14, 879±888.
94. Leclerc, S., Garnier, M., Hoessel, R., Marko, D., Bibb, J.A.,
Snyder, G.L., Greengard, P., Biernat, J., Wu, Y., Mandelkow,
E.M. & Mandelkow, E. (2001) Indirubins inhibit glycogen
synthase kinase-3 beta and Cdk5p25, two protein kinases
involved in absnormal tau phosphorylation in Alzheimer's
disease- A property common to most cdk inhibitors? J. Biol.
Chem. 276, 251±260.
95. Anderson, B. (1999) Alzheimer's disease: clues from flies and
worms. Current Biol. 9, 106±109.
96. Ahlijanian, M., Barrezueta, N., Williams, R., Jakowski, A.,
Kowsz, K., McCarthy, S., Coskran, T., Carlo, A., Seymour, P.,
Burkhardt, J., Nelson, R. & McNeish, J. (2000) Hyper phos-
phorylated tau, neurofilaments and cytoskeletal disruption in
mice overexpressing human p25, an activator of Cdk5. Proc. Natl
Acad. Sci. USA 97, 2910±2915.
97. Anderson, J., Pike, C. & Cotman, C. (1995) Differential induction
of immediate early gene proteins in cultured neurons by
b-amyloid (Ab): association of c-jun with Ab-induced apoptosis.
J. Neurochem. 65, 1487±1498.
98. Earnshaw, W., Martins, L. & Kaufmann, S. (1999) Mammalian
caspases structure, activation, substrates and functions during
apoptosis. Ann. Rev. Biochem. 68, 383±424.
99. Mattson, M. & Duan, W. (1999) Apoptotic biochemical cascades
in synaptic compartments: roles in adaptative plasticity and
neurodegenerative disorders. J. Neurosci. Res. 58, 152±166.
100. Estus, S., Tucker, H., van Rooven, C., Wright, C., Brugham, S.,
Wogulis, M. & Rydel, R. (1997) Aggregated b-amyloid protein
1526 R. B. Maccioni et al. (Eur. J. Biochem. 268) q FEBS 2001

induces cortical neuronal apoptosis and concomitant apoptotic
patterns of gene induction. J. Neurosci. 17, 7736±7745.
101. Watt, J., Pike, C., Walencewicz, A. & Cotman, C. (1994)
Ultrastructural analysis of b-amyloid-induced apoptosis in
cultured hippocampal neurons. Brain Res. 661, 147±156.
102. Harada, J. & Sugimoto, M. (1999) Activation of caspase-3 in
b-amyloid-induced apoptosis of cultured rat cortical neurons.
Brain Res. 842, 311±323.
103. Troy, C., Rabacchi, S., Friedman, W., Frappier, T., Brown, K. &
Shelanski, M. (2000) Caspase-2 mediates neuronal death induced
by b-amyloid. J. Neurosci. 20, 1386±1392.
104. Ahuja, H., Zhu, Y. & Zakeri, A. (1997) Association of cyclin
dependent protein kinase 5 and its activator p35 with apoptotic
cell death. Dev. Genet. 21, 258±267.
105. Henchcliffe, C. & Burke, R. (1997) Increased expression of
cyclin-dependent kinase-5 in induced apoptotic neurons death in
rat substantia nigra. Neurosci. Lett. 230, 41±44.
106. Maas, T., Eidenmuller, J. & Brandt, R. (2000) Interaction of tau
with the neural membrane cortex is regulated by phopshorylation
at sites that are modified in paired helical filaments. J. Biol.
Chem. 275, 15733±15740.
q FEBS 2001 Structure, function and pathology of protein kinase Cdk5 (Eur. J. Biochem. 268) 1527