
The polysaccharide fraction of Propionibacterium acnes modulates the development of experimental...
11
Immunobiology 217 (2012) 831–841 Contents lists available at SciVerse ScienceDirect Immunobiology j ourna l homepage: www.elsevier.de/imbio The polysaccharide fraction of Propionibacterium acnes modulates the development of experimental focal segmental glomerulosclerosis Vanessa Oliveira Reis a , Janice Costa Silva a , Gabriela Trindade Souza c , Patricia Semedo a , Bruna Buscariollo a , Rafael Luiz Pereira a , Marcos Antonio Cenedeze a , Alvaro Pacheco-Silva a,b , Ieda M. Longo-Maugéri c , Niels Olsen Saraiva Câmara a,d , Alexandre Castro Keller a,c,∗ a Departamento de Medicina, Universidade Federal de São Paulo, São Paulo, Brazil b Unidade de Transplante Renal, Instituto Israelita de Ensino e Pesquisa Albert Einstein, São Paulo, Brazil c Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de São Paulo, São Paulo, Brazil d Departamento de Imunologia, Universidade de São Paulo, São Paulo, Brazil a r t i c l e i n f o Article history: Received 28 June 2011 Received in revised form 8 December 2011 Accepted 11 December 2011 Keywords: Adriamycin nephropathy FSGS Glomerulosclerosis Immunomodulation Propionibacterium acnes a b s t r a c t The pathogenesis of focal segmental glomerulosclerosis (FSGS) appears to be associated with type-2 cytokines and podocyte dysfunction. In this study, we tested the hypothesis that immunization with the polysaccharide fraction of Propionibacterium acnes (PS), a pro-Th1 agonist, may subvert the type-2 pro- file and protect podocytes from adriamycin-induced glomerulosclerosis. Adriamycin injection resulted in albuminuria and increased serum creatinine in association with loss of glomerular podocin and podoplanin expression, which is consistent with podocyte dysfunction. Renal tissue analysis revealed the expression of transcripts for GATA3 and fibrogenic-related proteins, such as TGF-, tissue inhibitor of metalloproteinase-1 (TIMP-1) and metalloproteinase 9 (MMP9). In association with the expression of fibrogenic transcripts, we observed peri-glomerular expression of -smooth muscle actin (-SMA), indicating epithelial-to-mesenchymal transition, and increased expression of proliferating cell nuclear antigen (PCNA) in tubular cells, suggesting intense proliferative activity. Previous immunization with PS inhibited albuminuria and serum creatinine in association with the preservation of podocyte proteins and inhibition of fibrogenic transcripts and the expression of -SMA and PCNA proteins. Tissue analysis also revealed that PS treatment induced expression of mRNA for GD3 synthase, which is a glycosiltrans- ferase related to the synthesis of GD3, a ganglioside associated with podocyte physiology. In addition, PS treatment inhibited the influx of inflammatory CD8 pos and CD11b pos cells to kidney tissue. Finally, PS treatment on day 4 post-ADM, a period when proteinuria was already established, was able to improve renal function. Thus, we demonstrate that the PS fraction of P. acnes can inhibit FSGS pathogenesis, suggesting that immunomodulation can represent an alternative approach for disease management. © 2011 Elsevier GmbH. All rights reserved. Introduction Renal end-stage failure, marked by glomerulosclerosis, tubu- lointerstitial inflammation and fibrosis, is a common result of the development of chronic kidney disease (CKD) due to focal segmen- tal glomerulosclerosis (FSGS). In accord with independent works, changes in podocyte physiology appear to be the first event dur- ing the development of FSGS (Barisoni et al. 2009). Although the Abbreviations: ADM, adriamycin; AN, adriamycin nephropathy; -SMA, - smooth muscle actin; CKD, chronic kidney disease; EMT, epithelial-to-mesenchymal transition; FSGS, focal segmental glomerulosclerosis; MMP9, metalloproteinase 9; PCNA, proliferating cell nuclear antigen; PS, polysaccharide fraction of P. acnes; TIMP-1, tissue inhibitor of metalloproteinase-1. ∗ Corresponding author at: Rua Botucatu, 862, 4 ◦ andar Disciplina de Imunologia, CEP 04023-900, São Paulo, SP, Brazil. Tel.: +55 11 5549 6073; fax: +55 11 5549 6073. E-mail addresses: [email protected], [email protected] (A.C. Keller). mechanisms involved in podocyte injury remain an object of inves- tigation, alterations in podocyte proteins, such as nephrin and podocin, have been extensively associated with the loss of glomeru- lar barrier filtration in both congenital nephrotic syndrome and acquired glomerular diseases (Kawachi et al. 2000, 2003). It is not clear what immune components are responsible for these alter- ations; however, there is accumulating evidence associating the development of a pro-Th2 profile with FSGS pathogenesis. For example, Yap et al. were the first to link relapses of idiopathic nephrotic syndrome (INS), a disease related to FSGS, with signif- icant increases in IL-13 mRNA expression in T lymphocytes (Yap et al. 1999). Similarly, different experimental models of CKD rein- force the correlation between Th2 immune responses and FSGS pathogenesis. Studying the spontaneous development of proteinuria in Buf- falo/Mna rats, Le Berre et al. observed an association between the loss of the glomerular barrier due to glomerular epithelial cells 0171-2985/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.imbio.2011.12.003
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The polysaccharide fraction of Propionibacterium acnes modulates the development of experimental...

Td
VBIa
b
c
d
a
ARRA
KAFGIP
I
ldtci
stPT
C
0d
Immunobiology 217 (2012) 831– 841
Contents lists available at SciVerse ScienceDirect
Immunobiology
j ourna l homepage: www.elsev ier .de / imbio
he polysaccharide fraction of Propionibacterium acnes modulates theevelopment of experimental focal segmental glomerulosclerosis
anessa Oliveira Reisa, Janice Costa Silvaa, Gabriela Trindade Souzac, Patricia Semedoa,runa Buscariolloa, Rafael Luiz Pereiraa, Marcos Antonio Cenedezea, Alvaro Pacheco-Silvaa,b,
eda M. Longo-Maugéri c, Niels Olsen Saraiva Câmaraa,d, Alexandre Castro Kellera,c,∗
Departamento de Medicina, Universidade Federal de São Paulo, São Paulo, BrazilUnidade de Transplante Renal, Instituto Israelita de Ensino e Pesquisa Albert Einstein, São Paulo, BrazilDepartamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de São Paulo, São Paulo, BrazilDepartamento de Imunologia, Universidade de São Paulo, São Paulo, Brazil
r t i c l e i n f o
rticle history:eceived 28 June 2011eceived in revised form 8 December 2011ccepted 11 December 2011
eywords:driamycin nephropathySGSlomerulosclerosis
mmunomodulationropionibacterium acnes
a b s t r a c t
The pathogenesis of focal segmental glomerulosclerosis (FSGS) appears to be associated with type-2cytokines and podocyte dysfunction. In this study, we tested the hypothesis that immunization with thepolysaccharide fraction of Propionibacterium acnes (PS), a pro-Th1 agonist, may subvert the type-2 pro-file and protect podocytes from adriamycin-induced glomerulosclerosis. Adriamycin injection resultedin albuminuria and increased serum creatinine in association with loss of glomerular podocin andpodoplanin expression, which is consistent with podocyte dysfunction. Renal tissue analysis revealedthe expression of transcripts for GATA3 and fibrogenic-related proteins, such as TGF-�, tissue inhibitorof metalloproteinase-1 (TIMP-1) and metalloproteinase 9 (MMP9). In association with the expressionof fibrogenic transcripts, we observed peri-glomerular expression of �-smooth muscle actin (�-SMA),indicating epithelial-to-mesenchymal transition, and increased expression of proliferating cell nuclearantigen (PCNA) in tubular cells, suggesting intense proliferative activity. Previous immunization with PSinhibited albuminuria and serum creatinine in association with the preservation of podocyte proteinsand inhibition of fibrogenic transcripts and the expression of �-SMA and PCNA proteins. Tissue analysis
also revealed that PS treatment induced expression of mRNA for GD3 synthase, which is a glycosiltrans-ferase related to the synthesis of GD3, a ganglioside associated with podocyte physiology. In addition,PS treatment inhibited the influx of inflammatory CD8pos and CD11bpos cells to kidney tissue. Finally, PStreatment on day 4 post-ADM, a period when proteinuria was already established, was able to improverenal function. Thus, we demonstrate that the PS fraction of P. acnes can inhibit FSGS pathogenesis,odu
suggesting that immunomntroduction
Renal end-stage failure, marked by glomerulosclerosis, tubu-ointerstitial inflammation and fibrosis, is a common result of theevelopment of chronic kidney disease (CKD) due to focal segmen-
al glomerulosclerosis (FSGS). In accord with independent works,hanges in podocyte physiology appear to be the first event dur-ng the development of FSGS (Barisoni et al. 2009). Although theAbbreviations: ADM, adriamycin; AN, adriamycin nephropathy; �-SMA, �-mooth muscle actin; CKD, chronic kidney disease; EMT, epithelial-to-mesenchymalransition; FSGS, focal segmental glomerulosclerosis; MMP9, metalloproteinase 9;CNA, proliferating cell nuclear antigen; PS, polysaccharide fraction of P. acnes;IMP-1, tissue inhibitor of metalloproteinase-1.∗ Corresponding author at: Rua Botucatu, 862, 4◦andar Disciplina de Imunologia,EP 04023-900, São Paulo, SP, Brazil. Tel.: +55 11 5549 6073; fax: +55 11 5549 6073.
E-mail addresses: [email protected], [email protected] (A.C. Keller).
171-2985/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.imbio.2011.12.003
lation can represent an alternative approach for disease management.© 2011 Elsevier GmbH. All rights reserved.
mechanisms involved in podocyte injury remain an object of inves-tigation, alterations in podocyte proteins, such as nephrin andpodocin, have been extensively associated with the loss of glomeru-lar barrier filtration in both congenital nephrotic syndrome andacquired glomerular diseases (Kawachi et al. 2000, 2003). It is notclear what immune components are responsible for these alter-ations; however, there is accumulating evidence associating thedevelopment of a pro-Th2 profile with FSGS pathogenesis. Forexample, Yap et al. were the first to link relapses of idiopathicnephrotic syndrome (INS), a disease related to FSGS, with signif-icant increases in IL-13 mRNA expression in T lymphocytes (Yapet al. 1999). Similarly, different experimental models of CKD rein-force the correlation between Th2 immune responses and FSGS
pathogenesis.Studying the spontaneous development of proteinuria in Buf-falo/Mna rats, Le Berre et al. observed an association between theloss of the glomerular barrier due to glomerular epithelial cells

8 obiolog
atsrotpuiopucSeatrpIiii
itpmm(
iafiswTo
M
E
stwn(
A
oSdwcl(iuptw
32 V.O. Reis et al. / Immun
lterations, e.g., foot process flattening, and early macrophage infil-ration and Th2 polarization (Le Berre et al. 2001, 2005). In theame vein, Lai et al. demonstrated that overexpression of IL-13esults in podocyte injury in association with impaired expressionf nephrin and podocin (Lai et al. 2007). Thus, we hypothesizedhat Th1 polarization of the immune response could alter the Th2rofile associated with the development of CKD and thereby mod-late disease pathology. Hence, to induce Th1 polarization of the
mmune responses we chose to use the polysaccharide fractionf Propionobacterium acnes, formerly known as Corynebacteriumarvum. The P. acnes is an anaerobic bacillus that is extensivelysed as an adjuvant in clinical trials and is therefore a suitableandidate for therapeutic approaches (Molife and Hancock 2002;hepherd 1997). The pro-Th1 properties of PS can be observed, forxample, in studies performed by Dr. Longo-Maugéri’s group. Theseuthors demonstrated that PS stimulation can induce the matura-ion and secretion of IL-12 by bone marrow cells and increase Th1esponses during vaccination with plasmid DNA containing a Try-anossoma cruzi gene (Mussalem et al. 2006; Squaiella et al. 2006).n the context of immunoregulation, they also demonstrated that PSmmunization was able to modulate the development of an OVA-nduced type I hypersensitivity late phase reaction, a typical Th2mmune response (Squaiella et al. 2008).
To understand the effects of PS on the progression of CKD renalnjury, we took advantage of adriamycin (ADM)-induced nephropa-hy (AN), an experimental model that involves consistent andredictable time-dependent injury. Because AN can mimic clinicalanifestation of human CKD due to primary FSGS, it is a valuableodel for studying interventions that can modify disease outcomes
Lee and Harris 2011).First, we demonstrate that PS immunization prior to ADM
njection inhibits renal failure due to podocyte injury, which is phenomenon associated with the inhibition of a pro-Th2 andbrogenic milieu in renal tissue and maintenance of glomerulartructure. In addition, we show that PS administration in periodshen proteinuria is already established recovers renal function.
herefore, our findings suggest that the immunomodulatory abilityf PS can be an alternative approach for CKD management.
aterials and methods
thical statements
The animals used in this work were housed in individualtandard cages and were kept on a 12 h light/dark cycle in aemperature-controlled room at 21–23 ◦C, with free access toater and food. All procedures were approved by the inter-al ethical committee of the Universidade Federal de São Paulono. 1136/09).
nimals and treatments
Isogenic male Balb/c mice aged 8–12 weeks (23–28 g) werebtained from the Animal Care Facility at the Federal University ofão Paulo (UNIFESP). All animals were housed in individual stan-ard cages and had free access to food and water. All proceduresere previously reviewed and approved by the internal ethical
ommittee of the institution. To induce focal segmental glomeru-osclerosis, experimental animals received 10 mg/kg adriamycindoxorubicin hydrochloride, Pfizer, NY, USA) via a single tail vein
njection at day 0, whereas control mice received an equal vol-me of saline (Wang et al. 2000; Zheng et al. 2005, 2006). Theolysaccharide fraction of P. acnes (25 �g/animal) was adminis-ered subcutaneously at days −7 and −1 or day 4 post-ADM. PSas obtained as previously described (Squaiella et al. 2006).y 217 (2012) 831– 841
Renal function
To determine renal function, urine samples were collected atdifferent time points for the quantification of proteinuria or thealbuminuria:creatininuria ratio. All samples were analyzed usingcommercially available colorimetric assays: Labtest (Minas Gerais,Brazil) for creatinine measurements and Sensiprot® (Minas Gerais,Brazil) for protein measurements. To estimate the urinary albuminconcentration, 10 �L of urine (1 mg/mL), corrected for urinary cre-atinine levels, was run on a 10% SDS-PAGE gel and stained withCoomassie blue. The densities of the bands present in the gel wereanalyzed using GeneSnap and Gene Tools software (Syngene, Cam-bridge, UK).
Real-time PCR analysis
For use in q-PCR, kidney samples were quickly frozen in liquidnitrogen. Total RNA was isolated using TRIzol Reagent (Invitrogen,Carlsbad, CA, USA) methodology. First-strand cDNAs were synthe-sized using MML-V reverse transcriptase (Promega, Madison, USA).Real-time PCR was performed via the TaqMan PCR assay to MMP9:Mm01240560 (Applied Biosystem, Foster City, CA, USA). For analy-ses of GATA3, T-bet, TGF-�, TGFBI, IFN-�, GM3 synthase (ST3Gal5),GD3 synthase (ST8Sia), podocin (NPHS2) and TIMP-1, real-timePCR was performed with SYBRGreen assays (Supplemental data1) (Applied Biosystems, Foster City, USA). The cycling conditionsfor the TaqMan and SYBRGreen assays were as follows: 10 min at95 ◦C, followed by 45 cycles of 30 s at 95 ◦C, 30 s at 60 ◦C and 30 s at72 ◦C. Relative quantification of mRNA levels was performed usingthe comparative threshold cycle method (Applied Biosystems, NJ,USA). Briefly, the amount of the target gene was normalized to anendogenous reference gene (HPRT, SYBR green), and it was thenrelated to a calibrator (control animals) using the formula 2−��Ct.Hence, all data were expressed as an N-fold difference related tothe expression of matched controls. Analyses were performed withSequence Detection Software 1.9 (Applied Biosystem, Foster City,USA).
Histological analysis
On the day of experiment, kidneys were fixed in 10% neu-tral formalin for 24 h and then embedded in paraffin. Sections(3 �m) were stained with hematoxylin/eosin. The sections wereanalyzed using a trinocular optical microscope (Olympus Corpora-tion, Toquio, Japan) to determine tubular dilation and/or atrophyand the presence of hyaline casts.
Immunohistochemical analysis
The glomerular expression of podocyte proteins, such aspodocin and podoplanin, was determined by immunohistochem-ical analysis of frozen kidney sections. Briefly, the kidneys wereembedded in OCT (Tissue Tek, USA) and preserved at −90 ◦C. Sec-tions of the kidneys with a thickness of 5 �m were incubatedfor 20 min with a solution containing 3% H2O2 to block endoge-nous peroxidase, washed with TBS and incubated for 20 min withan biotin/avidin blocking solution (Dako, Denmark), followed by2 h with a protein blocking solution (Dako, Denmark) plus 1%BSA. After washing, the primary antibodies rabbit anti-mousepodocin (Abcam, USA), biotin-conjugated Syrian hamster anti-mouse podoplanin (Biolegend, USA) or control antibodies were
added overnight. After washing in TBS, the slides were incu-bated for 30 min with EnVisionTM+ Dual Link Single Reagent orstreptoavidin-HRP (Dako, Denmark, followed by a 1 min incubationwith Liquid DAB+ (Dako, Denmark).
biolog
itseptwAEDD
C
uUbGtapMt
Aa
l1wtUTpS
Fiidr
V.O. Reis et al. / Immuno
The expression of �-SMA and PCNA was determined bymmunohistochemical analysis of kidney paraffin-embedded sec-ions. Briefly, 5 �m sections were deparaffinized, rehydrated, andubjected to antigen retrieval in Tris–EDTA, pH 9 at 95 ◦C. Thendogenous peroxidase activity was blocked with 3% hydrogeneroxide, and the sections were blocked with Protein Block Solu-ion (DAKO, Denmark). Then, the slides were incubated overnightith primary �-SMA, PCNA or control antibodies (Abcam, USA).fter washing in TBS, the slides were incubated for 30 min withnVisionTM+ Dual Link, Single Reagent or streptoavidin-HRP (Dako,enmark), respectively, followed by a 1 min incubation with LiquidAB+ (Dako, Denmark).
ytokine analysis
To quantify the levels of different kidney cytokines present, wesed a Bio-Plex mouse cytokine assay kit (Bio-Rad Laboratories,SA). Briefly, kidney tissue was homogenized in 500 �L of RIPAuffer plus a protease/phosphatase inhibitor cocktail (Roche,ermany), and the supernatant was collected and used to test
he samples for the presence of pro-inflammatory cytokines, suchs IL-13, KC and TNF-�. The assay was read in the Bio-Plex sus-ension array system, and the data were analyzed using Bio-Plexanager software version 4.0. Standard curves ranged from 1.95
o 32,000 pg/mL.
ctive TGFˇ-1 protein analysis by enzyme-linked immunosorbentssay (ELISA)
The active TGF�-1 protein was assessed from kidney tissuesysed with RIPA buffer (25 mM Tris·HCl pH 7.6, 150 mM NaCl,% NP-40, 1% sodium deoxycholate, 0.1% SDS), supplementedith a protease inhibitor (Roche, Germany). To this end, we used
he Human/Mouse TGF beta1 ELISA Ready-SET-Go (eBioscience,
SA), which is engineered to detect the mature/active form of theGF�-1 protein. The results are presented as TGF�-1 pg/mg of totalrotein measured using the Pierce 660 nM Protein Assay (Thermocientific, USA).ig. 1. PS treatment inhibits ADM-induced renal failure due to podocyte injury. BALB/mmunized with PS (days −7 and −1). (A) The time-dependent increase in the proteinurn comparison to control and PS + ADM animals, indicating a renoprotective effect of PS true to ADM cytotoxicity, which is inhibited by PS administration. (C) The high levels of senal failure, in contrast to the control and PS + ADM groups, confirming the renoprotecti
y 217 (2012) 831– 841 833
SMAD7 analysis
The levels of SMAD7 protein were determined by west-ern blot. Briefly, 60 �g of total protein from renal tissue wasdiluted in sample buffer (Bio-Rad, USA), containing 20 mg/mL of2-�-mercaptoethanol (Sigma–Aldrich, USA). The samples weredenatured for 5 min at 95 ◦C and then were separated on a 10% poly-acrylamide electrophoresis gel. Next, the proteins were transferredto a nitrocellulose membrane, blocked for 1 h with 5% albumindiluted in TBS-T solution, and then incubated with rabbit anti-SMAD7 (Santa Cruz, USA) or anti-HPRT (abcam, USA) primaryantibodies diluted in TBS. Finally, the membrane was washed withTBS for 1 h and incubated with the secondary goat anti-rabbit IgGHRP-conjugated antibody (abcam, USA). The proteins were deter-mined using an ECL plus detection kit (G&E, USA). The detectionassay was performed in the Alliance 4.7 equipment with data anal-ysis by UVIsoft software (Uvitec, UK).
FACS analysis
To perform the FACS analysis, kidneys were perfused via injec-tion of 10 mL PBS into the left ventricle, minced and incubatedfor 25 min at 37 ◦C in 1 mL PBS containing 1 mg/mL DNase and5 U/mL Dispase (Gibco, USA). Next, we gently passed the kidneyhomogenate through a 100 �M cell strainer (BD Biosciences, SanDiego, CA) and then separated mononuclear cells (MNCs) by gra-dient centrifugation in 35% Percoll (G&E, USA). We stained theMNCs (2 × 106 cells) with a mixture containing FITC-conjugatedanti-CD3, allophycocyanin (APC)-conjugated CD8 and Pacific Blue-conjugated anti-CD4 (ebioscience, USA) or with FITIC-conjugatedanti-CD11b (eBioscience, USA) (Supplemental data 2). The datawere analyzed on a FACS Canto II using DIVA software (Becton Dick-inson, USA). Further analyses of FACS data were performed usingFLOWJO 9.3 software (Tree Star, USA).
Statistical analysis
All data are presented as the mean ± SEM. Different resultsamong groups were compared using an ANOVA test. Significancewas established as p < 0.05. All statistical analyses were performed
c mice were injected at day 0 with 10 mg/kg of adriamycin (ADM) or previouslyia/creatininuria ratio observed in the ADM group reflects impaired renal functioneatment. (B) The albuminuria levels found in ADM animals reflect podocyte injuryerum creatinine found in ADM mice complete the manifestations associated with
ve effect of PS treatment. *p < 0.05 vs. control; #p < 0.05 vs. ADM.
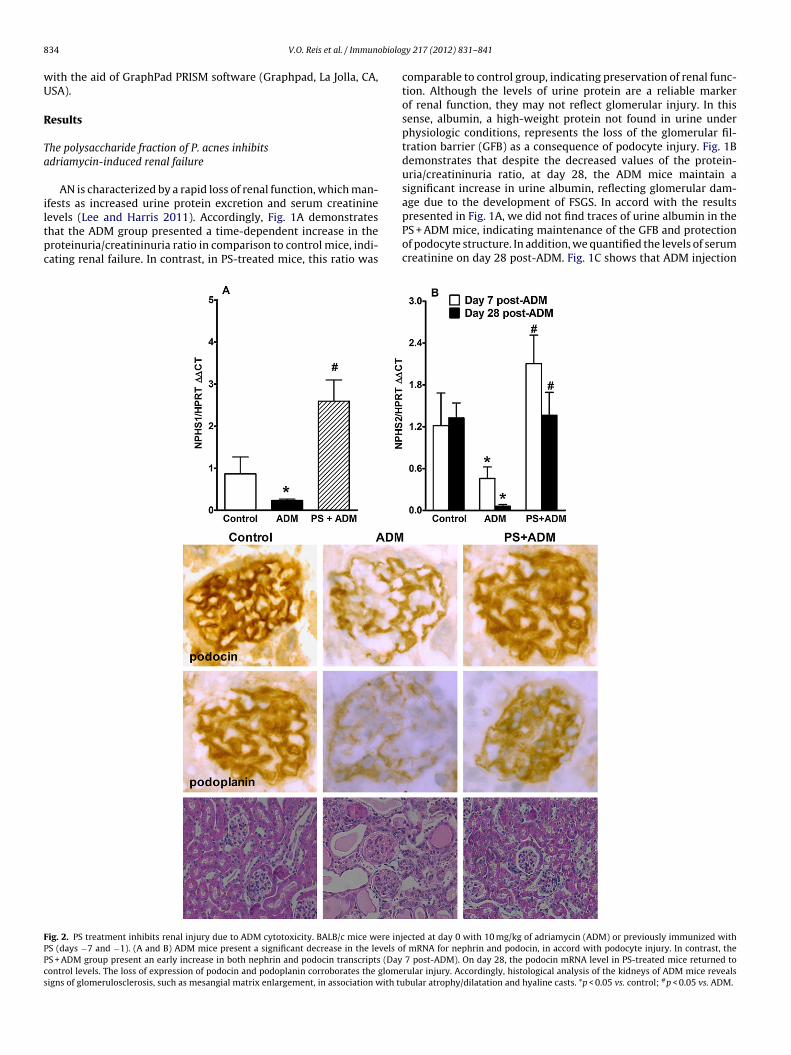
8 obiolog
wU
R
Ta
iltpc
FPPcs
34 V.O. Reis et al. / Immun
ith the aid of GraphPad PRISM software (Graphpad, La Jolla, CA,SA).
esults
he polysaccharide fraction of P. acnes inhibitsdriamycin-induced renal failure
AN is characterized by a rapid loss of renal function, which man-fests as increased urine protein excretion and serum creatinine
evels (Lee and Harris 2011). Accordingly, Fig. 1A demonstrateshat the ADM group presented a time-dependent increase in theroteinuria/creatininuria ratio in comparison to control mice, indi-ating renal failure. In contrast, in PS-treated mice, this ratio wasig. 2. PS treatment inhibits renal injury due to ADM cytotoxicity. BALB/c mice were injS (days −7 and −1). (A and B) ADM mice present a significant decrease in the levels oS + ADM group present an early increase in both nephrin and podocin transcripts (Dayontrol levels. The loss of expression of podocin and podoplanin corroborates the glomeigns of glomerulosclerosis, such as mesangial matrix enlargement, in association with tu
y 217 (2012) 831– 841
comparable to control group, indicating preservation of renal func-tion. Although the levels of urine protein are a reliable markerof renal function, they may not reflect glomerular injury. In thissense, albumin, a high-weight protein not found in urine underphysiologic conditions, represents the loss of the glomerular fil-tration barrier (GFB) as a consequence of podocyte injury. Fig. 1Bdemonstrates that despite the decreased values of the protein-uria/creatininuria ratio, at day 28, the ADM mice maintain asignificant increase in urine albumin, reflecting glomerular dam-age due to the development of FSGS. In accord with the results
presented in Fig. 1A, we did not find traces of urine albumin in thePS + ADM mice, indicating maintenance of the GFB and protectionof podocyte structure. In addition, we quantified the levels of serumcreatinine on day 28 post-ADM. Fig. 1C shows that ADM injectionected at day 0 with 10 mg/kg of adriamycin (ADM) or previously immunized withf mRNA for nephrin and podocin, in accord with podocyte injury. In contrast, the
7 post-ADM). On day 28, the podocin mRNA level in PS-treated mice returned torular injury. Accordingly, histological analysis of the kidneys of ADM mice revealsbular atrophy/dilatation and hyaline casts. *p < 0.05 vs. control; #p < 0.05 vs. ADM.

V.O. Reis et al. / Immunobiology 217 (2012) 831– 841 835
Fig. 3. PS treatment inhibits the ADM-induced Th2-like profile and induces a pro-Th1 environment. BALB/c mice were injected at day 0 with 10 mg/kg of adriamycin (ADM)or previously immunized with PS (days −7 and −1). (A) ADM administration resulted in increased levels of mRNA for GATA3, reflecting the induction of a Th2-like profile.I ith a( S is asv
idtc
Tg
wota(epatitaarap
Tmi
rfcwacrpp
n contrast, PS treatment decreased the levels of GATA3 transcripts in association wB and C, respectively). Thus, these data corroborate the idea that ADM-induced FSGs. control; #p < 0.05 vs. ADM.
ncreased serum creatinine, in line with the loss of renal functionemonstrated by the levels of proteinuria and albuminuria. In con-rast, in the PS + ADM group, serum creatinine was comparable toontrol group, reinforcing the protective effect of PS treatment.
he polysaccharide fraction of P. acnes inhibits ADM-inducedlomerular degeneration
Loss of the GFB due to podocyte injury is strongly associatedith decreased expression of podocyte-associated proteins, and
ur analysis of renal tissues by q-PCR revealed that ADM injec-ion induced significant decreases in the levels of nephrin (NPHS1)nd podocin (NPHS2) transcripts in comparison to control animalsFig. 2A and B, respectively). In contrast, PS treatment induced anarly increase in the levels of nephrin and podocin mRNA in com-arison to both ADM and control mice. The analysis of podocint day 28 post-ADM revealed that its transcripts were at con-rol levels, suggesting preservation of podocin expression. Indeed,mmunohistochemical evaluation of renal tissue showed that PSreatment conserved the glomerular expression of both the podocinnd podoplanin proteins compared to ADM animals. Histologicalnalysis revealed that ADM mice presented signs of glomeruloscle-osis, e.g., mesangial matrix swelling, in association with tubulartrophy and dilatation and hyaline casts. In contrast, PS treatmentreserved renal structure, confirming its renoprotective effect.
he polysaccharide fraction of P. acnes modulates the pro-Th2ilieu and the expression of fibrogenic-related transcripts
nduced by ADM
q-PCR analyses of renal tissue revealed that ADM injectionesulted in increased levels of mRNA for the pro-Th2 transcriptionactor GATA3 (Fig. 3A) (Lee et al. 2001; Zheng and Flavell 1997). Inontrast, PS treatment inhibited GATA3, a phenomenon associatedith increased levels of T-bet (T box expressed in T-cells) mRNA,
pro-Th1-related transcription factor, and INF-�, a classical Th1
ytokine (Fig. 3B and C, respectively) (Szabo et al. 2000). These dataeinforce the connection between Th2 immune responses and FSGSathogenesis and support the hypothesis that PS immunizationromotes pro-Th1 polarization and thereby kidney preservation.n increase in the levels of mRNA for T-bet and IFN-�, a characteristic Th1 cytokinesociated with a Th2-like profile that can be inhibited by PS immunization. *p < 0.05
Kidney fibrosis is strongly associated with CKD; therefore,we sought to determine the expression of mRNA for fibro-genic proteins, such as TGF-�, TIMP-1 (tissue inhibitor ofmetalloproteinase-1) and metalloproteinase 9 (MMP-9). ADMinjection induced a significant increase in TGF-� transcripts as wellas in the levels of TIMP-1 and MMP9 mRNA, supporting the rela-tionship between the development of fibrosis and AN (Fig. 4A–C,respectively). In contrast, in the PS + ADM group, these levels weresimilar to the levels in control animals, demonstrating that PS treat-ment inhibited the development of a fibrogenic milieu in the kidney.
In addition, further analysis of tissue cytokines revealed thatADM administration resulted in increased levels of IL-13, reinforc-ing the Th2/AN association, and inflammatory cytokines, such asKC and TNF-� (Fig. 4D–F, respectively). In contrast, PS treatmentinhibited the synthesis of IL-13, KC and TNF-�, associating the PSrenoprotective effect with the blockage of pro-Th2 and fibrogenictranscripts and inflammatory responses. Although we did not findany difference in the levels of active TGF-� in kidney tissue (datanot shown), western blot analysis revealed that AN developmentresulted in a decrease in SMAD7 protein, suggesting an associa-tion between disease pathology and impaired TGF-� regulation(Fig. 4G).
PS treatment inhibits the epithelial-to-mesenchymal transitionand cell proliferation, whereas it increases the levels ofganglioside synthase transcripts
Despite the Th2-FSGS association, the mechanisms involved inpodocyte degeneration are still not fully understood. In this regard,the epithelial-to-mesenchymal transition (EMT) due to TGF-� wasrecently described as a potential pathway associated with podocytedysfunction (Li et al. 2008). Because PS treatment decreased thelevels of TGF-� transcripts, we sought to determine the levels ofexpression of �-SMA, a protein extensively associated with EMT, inrenal tissue. Fig. 5A shows that ADM administration leads to the denovo expression of �-SMA in peri-glomerular epithelial cells, which
is in accord with the association between renal function degener-ation and EMT. In addition, we observed increased expression ofPCNA in the renal tubules of ADM mice, suggesting an intense pro-cess of cellular proliferation in these mice in comparison to the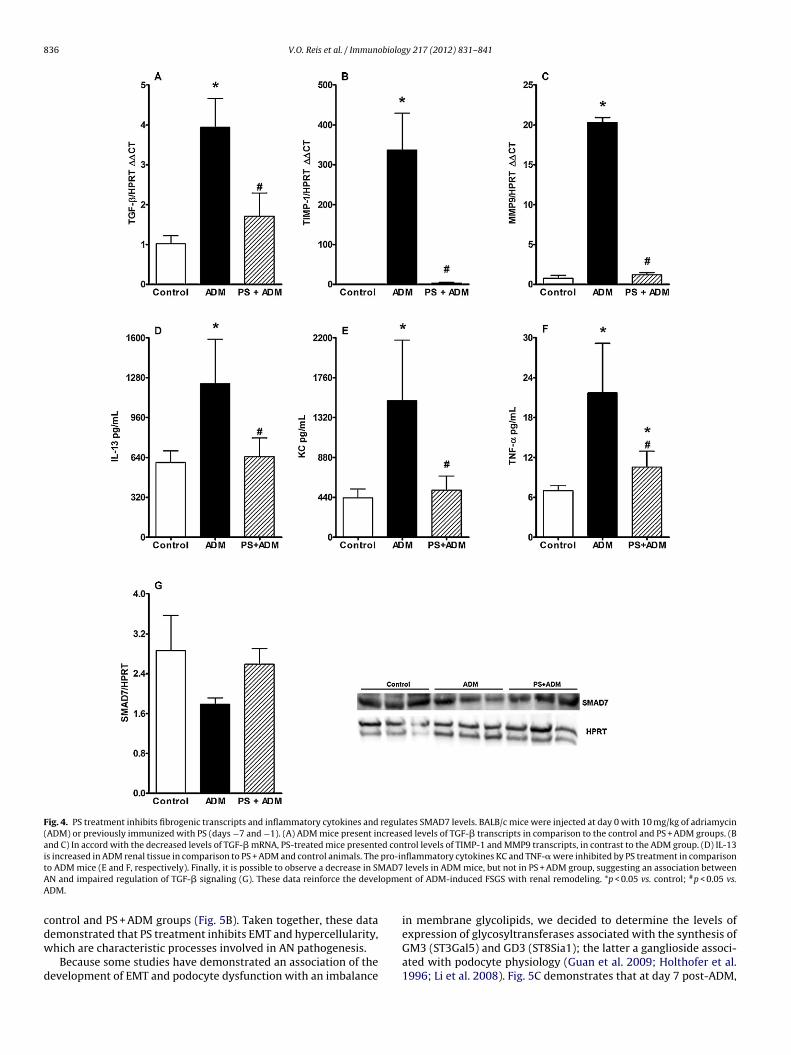
836 V.O. Reis et al. / Immunobiology 217 (2012) 831– 841
Fig. 4. PS treatment inhibits fibrogenic transcripts and inflammatory cytokines and regulates SMAD7 levels. BALB/c mice were injected at day 0 with 10 mg/kg of adriamycin(ADM) or previously immunized with PS (days −7 and −1). (A) ADM mice present increased levels of TGF-� transcripts in comparison to the control and PS + ADM groups. (Band C) In accord with the decreased levels of TGF-� mRNA, PS-treated mice presented control levels of TIMP-1 and MMP9 transcripts, in contrast to the ADM group. (D) IL-13is increased in ADM renal tissue in comparison to PS + ADM and control animals. The pro-inflammatory cytokines KC and TNF-� were inhibited by PS treatment in comparisont AD7A lopmeA
cdw
d
o ADM mice (E and F, respectively). Finally, it is possible to observe a decrease in SMN and impaired regulation of TGF-� signaling (G). These data reinforce the deveDM.
ontrol and PS + ADM groups (Fig. 5B). Taken together, these data
emonstrated that PS treatment inhibits EMT and hypercellularity,hich are characteristic processes involved in AN pathogenesis.Because some studies have demonstrated an association of theevelopment of EMT and podocyte dysfunction with an imbalance
levels in ADM mice, but not in PS + ADM group, suggesting an association betweennt of ADM-induced FSGS with renal remodeling. *p < 0.05 vs. control; #p < 0.05 vs.
in membrane glycolipids, we decided to determine the levels of
expression of glycosyltransferases associated with the synthesis ofGM3 (ST3Gal5) and GD3 (ST8Sia1); the latter a ganglioside associ-ated with podocyte physiology (Guan et al. 2009; Holthofer et al.1996; Li et al. 2008). Fig. 5C demonstrates that at day 7 post-ADM,
V.O. Reis et al. / Immunobiology 217 (2012) 831– 841 837
Fig. 5. PS treatment inhibited ADM-induced EMT and hypercellularity in association with the induction of ganglioside synthase transcripts. BALB/c mice were injected atday 0 with 10 mg/kg of adriamycin (ADM) or previously immunized with PS (days −7 and −1). (A) Immunohistochemical analysis of renal tissue demonstrates that ADMadministration induced de novo expression of �-SMA in peri-glomerular epithelial cells, in accord with the induction of EMT. (B) The presence of PCNA-stained nuclei (blackarrows) in the ADM group suggests an intense proliferative activity in ADM mice in comparison to PS + ADM and control animals. (C and D) In association with EMT andproliferative activity inhibition, ADM mice presented an early decrease in GD3 synthase (ST8Sia) expression, which was maintained at control levels by PS treatment. PSt s to co
tcd(ewpPgmr
reatment induced an early enhancement of GM3 synthase (ST3Gal5), which return
here is significant inhibition of the GD3 synthase transcripts inomparison to PS + ADM and control mice, although we did notetect significant alteration of the levels of GM3 synthase mRNAFig. 5C and D, respectively). However, PS treatment induced anarly increase in GM3 synthase mRNA, which can be associatedith the maintenance of GD3 synthase transcripts at levels com-arable to the control group. At day 28 post-ADM, the ADM and
S + ADM groups presented increased levels of transcripts for bothlycosyltransferases, suggesting that early alteration of gangliosideetabolism can be associated with ADM-induced pathology and aenoprotective effect of PS.
ntrol levels at day 28. *p < 0.05 vs. Control; #p < 0.05 vs. ADM.
PS immunization inhibits the influx of inflammatory cells tokidneys
The balance between CD4pos and CD8pos lymphocytes and theearly influx of macrophages to renal tissue plays a pivotal role inAN pathogenesis (Wang et al. 2001a,b, 2005a). Thus, we sought todetermine the effect of PS treatment on the infiltration of inflam-
matory cells to the kidney after ADM administration. On day sevenpost-ADM, we observed a significant influx of T CD3pos lymphocytesfrom both the T CD4pos and CD8pos subsets and of CD11bpos cells tokidneys (Fig. 6A–D, respectively). In the PS + ADM group, although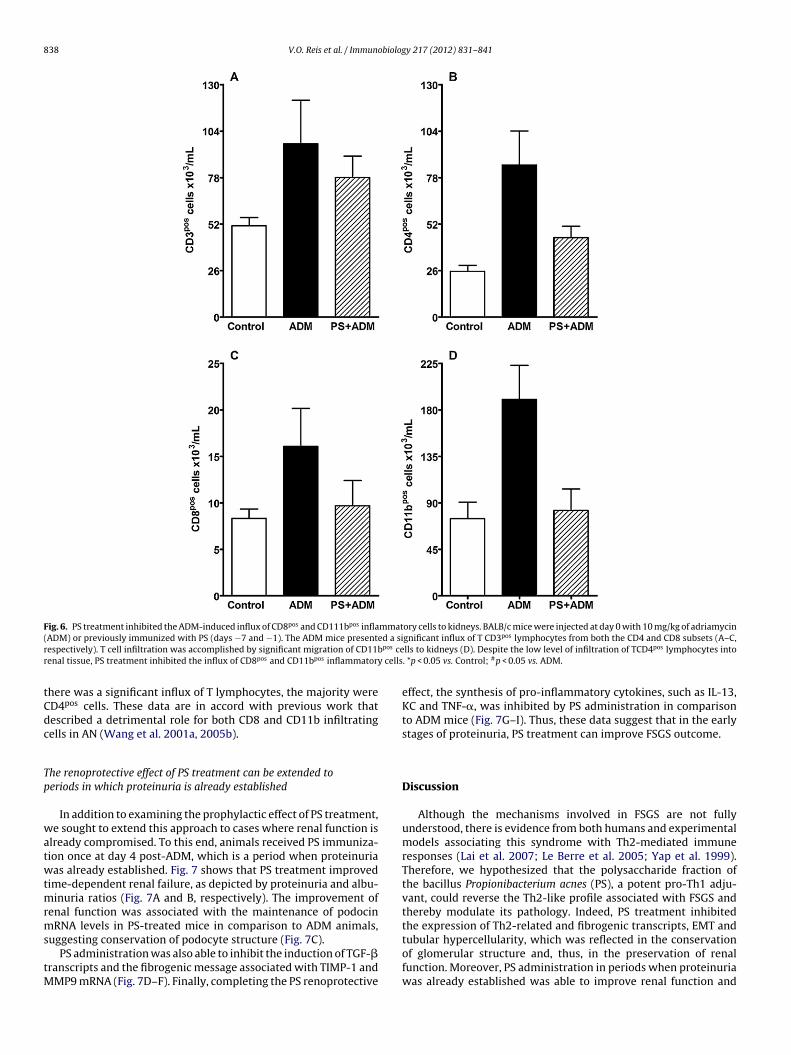
838 V.O. Reis et al. / Immunobiology 217 (2012) 831– 841
Fig. 6. PS treatment inhibited the ADM-induced influx of CD8pos and CD111bpos inflammatory cells to kidneys. BALB/c mice were injected at day 0 with 10 mg/kg of adriamycin( ed a sir pos cer cells
tCdc
Tp
watwtmrms
tM
ADM) or previously immunized with PS (days −7 and −1). The ADM mice presentespectively). T cell infiltration was accomplished by significant migration of CD11benal tissue, PS treatment inhibited the influx of CD8pos and CD11bpos inflammatory
here was a significant influx of T lymphocytes, the majority wereD4pos cells. These data are in accord with previous work thatescribed a detrimental role for both CD8 and CD11b infiltratingells in AN (Wang et al. 2001a, 2005b).
he renoprotective effect of PS treatment can be extended toeriods in which proteinuria is already established
In addition to examining the prophylactic effect of PS treatment,e sought to extend this approach to cases where renal function is
lready compromised. To this end, animals received PS immuniza-ion once at day 4 post-ADM, which is a period when proteinuriaas already established. Fig. 7 shows that PS treatment improved
ime-dependent renal failure, as depicted by proteinuria and albu-inuria ratios (Fig. 7A and B, respectively). The improvement of
enal function was associated with the maintenance of podocinRNA levels in PS-treated mice in comparison to ADM animals,
uggesting conservation of podocyte structure (Fig. 7C).PS administration was also able to inhibit the induction of TGF-�
ranscripts and the fibrogenic message associated with TIMP-1 andMP9 mRNA (Fig. 7D–F). Finally, completing the PS renoprotective
gnificant influx of T CD3pos lymphocytes from both the CD4 and CD8 subsets (A–C,lls to kidneys (D). Despite the low level of infiltration of TCD4pos lymphocytes into. *p < 0.05 vs. Control; #p < 0.05 vs. ADM.
effect, the synthesis of pro-inflammatory cytokines, such as IL-13,KC and TNF-�, was inhibited by PS administration in comparisonto ADM mice (Fig. 7G–I). Thus, these data suggest that in the earlystages of proteinuria, PS treatment can improve FSGS outcome.
Discussion
Although the mechanisms involved in FSGS are not fullyunderstood, there is evidence from both humans and experimentalmodels associating this syndrome with Th2-mediated immuneresponses (Lai et al. 2007; Le Berre et al. 2005; Yap et al. 1999).Therefore, we hypothesized that the polysaccharide fraction ofthe bacillus Propionibacterium acnes (PS), a potent pro-Th1 adju-vant, could reverse the Th2-like profile associated with FSGS andthereby modulate its pathology. Indeed, PS treatment inhibitedthe expression of Th2-related and fibrogenic transcripts, EMT and
tubular hypercellularity, which was reflected in the conservationof glomerular structure and, thus, in the preservation of renalfunction. Moreover, PS administration in periods when proteinuriawas already established was able to improve renal function and
V.O. Reis et al. / Immunobiology 217 (2012) 831– 841 839
Fig. 7. PS treatment improves time-dependent ADM-induced renal failure. BALB/c mice were injected at day 0 with 10 mg/kg of adriamycin (ADM) or received PS treatmentfour days post-ADM (ADM+PS). (A) PS treatment improved renal function in comparison to ADM mice. (B) The albuminuria levels found at day 28 post-ADM confirm ther in thP anscrt < 0.05
mf
TegBBHrteawi
ecovery of renal function by PS treatment. (C) PS treatment inhibited the decreasesS administration at day four post-ADM inhibited the induction of pro-fibrogenic trhe induction of IL-13 and other inflammatory cytokines, such as KC and TNF-�. *p
aintain kidney architecture, suggesting an alternative approachor FSGS management.
Podocyte injury plays a pivotal role in the pathogenesis of FSGS.herefore, we took advantage of the AN model to test the hypoth-sis that the pro-Th1 adjuvant properties of PS could modulatelomerulosclerosis, based on the fact that the susceptibility ofALB/c mouse podocytes to ADM appears to be somehow linked toALB/c characteristic polarization through Th2 responses (Lee andarris 2011). Accordingly, we found an association between the
enoprotective effect of PS and the induction of transcripts relatedo Th1 responses to the detriment of those related to Th2 (Szabo
t al. 2000; Zheng and Flavell 1997). Moreover, the PS + ADM grouplso presented decreased levels of pro-inflammatory cytokines,hich is in agreement with the inhibition of the early influx ofnflammatory cells to kidneys.
e renal expression of transcripts for podocin, indicating podocyte protection. (D–F)ipts, such as TGF-�, TIMP-1 and MMP9. (G–I) PS administration was able to inhibit
vs. control; #p < 0.05 vs. ADM.
Although the association between the Th1/Th2 balance andresistance to ADM-induced FSGS is evident, it is not sufficientto explain the renoprotective effect of PS. In this regard, TGF-�is a cytokine that is extensively associated with CKD pathogen-esis. TGF-� overexpression in kidneys, as occurs in pathologicalinflammation, can be associated with podocyte apoptosis and pro-liferation (Wu et al. 2005; Yoshida et al. 2008). It can also lead toepithelial cell dedifferentiation into fibroblasts/myofibroblasts, aprocess known as EMT, resulting in matrix protein (MP) oversecre-tion and, thus, fibrosis (Liu, 2010). We found that in ADM mice,loss of the GFB due to podocyte injury, depicted in the level of
urine albumin, was associated with high levels of mRNA for TGF-� and peri-glomerular expression of �-SMA, which indicates EMT.In addition to TGF-�, the metalloproteinases modulate matrix pro-tein (MP) turnover and fibrosis due to their proteolytic activity. The
8 obiolog
ibitsbwPtoasBgirifpcctr
pdSiaad(Gpc(sPGc(bdietriftt
tup
ogoa
A
t
40 V.O. Reis et al. / Immun
ncreased levels of mRNA for MMP9 suggest a tentative of counter-alancing the excess of MP production, which is hampered by the
nduction of inhibitors of MMP proteolytic activity, as indicated byhe high levels of TIMP-1 transcripts. Taken together, these datauggest the induction of a vicious cycle of MP changes characterizedy protein deposition induced by TGF-� and impaired degradation,hich leads to the classical fibrosis associated with CKD. However,
S treatment inhibited the transcription of TGF-� and thereby con-rolled pathological MP turnover, which explains the maintenancef TIMP-1 and MMP9 mRNA at levels comparable to those in controlnimals. In order to clarify the role of TGF-� in AN pathogene-is, we first quantified the levels of active TGF-� in kidney tissue.ecause there was no significant difference in its levels among theroups, we decided to study SMAD7, an inhibitory protein involvedn downstream regulation of TGF-� signaling that is protective inenal fibrosis (Chen et al. 2011; Wang et al. 2005a, 2007). As shownn Fig. 4G, the SMAD7 levels in the ADM mice were under from thoseound in control animals, suggesting the association between ANathogenesis and impaired regulation of TGF-� signaling. In con-ordance, the PS treatment maintained SMAD7 protein at levelsomparable to control animals. Taken together, these data suggesthat PS protective effect can be, in part at least, associated withegulation of TGF-� cascade by sustaining SMAD7 expression.
An additional player in the intricate process that leads toodocyte injury is the alterations in podocyte surface proteinsue to variations in membrane lipid raft components. The work ofimons et al. demonstrated the involvement of lipid rafts contain-ng GD3 gangliosides in podocyte foot process effacement caused bylterations in nephrin (Simons et al. 2001). Additionally, Holthöfernd cols associated podocyte injury with decreases in glomerularisialogangliosides (GD3) in a model of puromycin induced-FSGSHolthofer et al. 1996). In addition to the direct involvement ofD3 in the downregulation of surface podocyte proteins and footrocess effacement, other classes of gangliosides have been impli-ated in EMT development as a consequence of TGF-� signalingGuan et al. 2009). Thus, it is possible to conjecture that ganglio-ides other than GD3 may be involved in the protective effect ofS. In this regard, the work of Noguchi et al. demonstrated thatM3 expression was able to inhibit doxorubicin cytotoxicity, whichan explain the podocyte protection observed in PS-treated miceNoguchi et al. 2006). Moreover, this ganglioside was found toe associated with inhibition of IL-4 priming and, thus, with theevelopment of Th2-associated responses as well as with the TGF�-
nduced proliferation of mesangial cells (Chen et al. 2000; Noguchit al. 2006). Consistent with these findings, we observed inhibi-ion of the message to transcription factors associated with Th2esponses (GATA3) and of tubular PCNA staining in PS-treated micen association with early increase in G3 synthase transcripts. There-ore, PS treatment can promote both podocyte stabilization throughhe synthesis of GD3 and podocyte protection against adriamycinoxicity via the induction of GM3.
Finally, PS administration in early proteinuria stages was ableo improve time-dependent renal failure in association with mod-lation of all of the deleterious parameters associated with FSGSathogenesis.
In conclusion, we demonstrated that the polysaccharide fractionf P. acnes can modulate the intricate network of Th1/Th2 cytokines,lycolipid metabolism and TGF-� signaling involved in AN devel-pment. Thus, this work suggests that PS immunization can be anlternative approach for FSGS management.
cknowledgments
The authors would like to thank Bernardo P. Albe for expertechnical assistance in the histological preparations. This work
y 217 (2012) 831– 841
was supported by grants from FAPESP (2007/07120-0) and CNPq(501848/2009-6 and 484445/2010-3).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.imbio.2011.12.003.
References
Barisoni, L., Schnaper, H.W., Kopp, J.B., 2009. Advances in the biology and genetics ofthe podocytopathies: implications for diagnosis and therapy. Arch. Pathol. Lab.Med. 133, 201.
Chen, H.Y., Huang, X.R., Wang, W., Li, J.H., Heuchel, R.L., Chung, A.C., Lan, H.Y., 2011.The protective role of Smad7 in diabetic kidney disease: mechanism and thera-peutic potential. Diabetes 60, 590.
Chen, X.P., Ding, X., Daynes, R.A., 2000. Ganglioside control over IL-4 priming andcytokine production in activated T cells. Cytokine 12, 972.
Guan, F., Handa, K., Hakomori, S.I., 2009. Specific glycosphingolipids mediateepithelial-to-mesenchymal transition of human and mouse epithelial cell lines.Proc. Natl. Acad. Sci. U.S.A. 106, 7461.
Holthofer, H., Reivinen, J., Solin, M.L., Haltia, A., Miettinen, A., 1996. Decrease ofglomerular disialogangliosides in puromycin nephrosis of the rat. Am. J. Pathol.149, 1009.
Kawachi, H., Koike, H., Kurihara, H., Sakai, T., Shimizu, F., 2003. Cloning of rat homo-logue of podocin: expression in proteinuric states and in developing glomeruli.J. Am. Soc. Nephrol. 14, 46.
Kawachi, H., Koike, H., Kurihara, H., Yaoita, E., Orikasa, M., Shia, M.A., Sakai, T.,Yamamoto, T., Salant, D.J., Shimizu, F., 2000. Cloning of rat nephrin: expressionin developing glomeruli and in proteinuric states. Kidney Int. 57, 1949.
Lai, K.W., Wei, C.L., Tan, L.K., Tan, P.H., Chiang, G.S., Lee, C.G., Jordan, S.C., Yap,H.K., 2007. Overexpression of interleukin-13 induces minimal-change-likenephropathy in rats. J. Am. Soc. Nephrol. 18, 1476.
Le Berre, L., Godfrin, Y., Perretto, S., Smit, H., Buzelin, F., Kerjaschki, D., Usal, C., Cuturi,C., Soulillou, J.P., Dantal, J., 2001. The Buffalo/Mna rat, an animal model of FSGSrecurrence after renal transplantation. Transplant. Proc. 33, 3338.
Le Berre, L., Herve, C., Buzelin, F., Usal, C., Soulillou, J.P., Dantal, J., 2005. Renalmacrophage activation and Th2 polarization precedes the development ofnephrotic syndrome in Buffalo/Mna rats. Kidney Int. 68, 2079.
Lee, G.R., Fields, P.E., Flavell, R.A., 2001. Regulation of IL-4 gene expression by distalregulatory elements and GATA-3 at the chromatin level. Immunity 14, 447.
Lee, V.W., Harris, D.C., 2011. Adriamycin nephropathy: a model of focal segmentalglomerulosclerosis. Nephrology (Carlton) 16, 30.
Li, Y., Kang, Y.S., Dai, C., Kiss, L.P., Wen, X., Liu, Y., 2008. Epithelial-to-mesenchymaltransition is a potential pathway leading to podocyte dysfunction and protein-uria. Am. J. Pathol. 172, 299.
Liu, Y., 2010. New insights into epithelial-mesenchymal transition in kidney fibrosis.J. Am. Soc. Nephrol. 21, 212.
Molife, R., Hancock, B.W., 2002. Adjuvant therapy of malignant melanoma. Crit. Rev.Oncol. Hematol. 44, 81.
Mussalem, J.S., Vasconcelos, J.R., Squaiella, C.C., Ananias, R.Z., Braga, E.G., Rodrigues,M.M., Longo-Maugeri, I.M., 2006. Adjuvant effect of the Propionibacterium acnesand its purified soluble polysaccharide on the immunization with plasmidialDNA containing a Trypanosoma cruzi gene. Microbiol. Immunol. 50, 253.
Noguchi, M., Kabayama, K., Uemura, S., Kang, B.W., Saito, M., Igarashi, Y., Inokuchi, J.,2006. Endogenously produced ganglioside GM3 endows etoposide and doxoru-bicin resistance by up-regulating Bcl-2 expression in 3LL Lewis lung carcinomacells. Glycobiology 16, 641.
Shepherd, F.A., 1997. Alternatives to chemotherapy and radiotherapy as adjuvanttreatment for lung cancer. Lung Cancer 17 (Suppl. 1), S121.
Simons, M., Schwarz, K., Kriz, W., Miettinen, A., Reiser, J., Mundel, P., Holthofer, H.,2001. Involvement of lipid rafts in nephrin phosphorylation and organization ofthe glomerular slit diaphragm. Am. J. Pathol. 159, 1069.
Squaiella, C.C., Ananias, R.Z., Mussalem, J.S., Braga, E.G., Rodrigues, E.G., Travassos,L.R., Lopes, J.D., Longo-Maugeri, I.M., 2006. In vivo and in vitro effect of killedPropionibacterium acnes and its purified soluble polysaccharide on mouse bonemarrow stem cells and dendritic cell differentiation. Immunobiology 211, 105.
Squaiella, C.C., Longhini, A.L., Braga, E.G., Mussalem, J.S., Ananias, R.Z., Yendo, T.M.,Straus, A.H., Toledo, M.S., Takahashi, H.K., Hirata, I.Y., Longo-Maugeri, I.M., 2008.Modulation of the type I hypersensitivity late phase reaction to OVA by Propi-onibacterium acnes-soluble polysaccharide. Immunol. Lett. 121, 157.
Szabo, S.J., Kim, S.T., Costa, G.L., Zhang, X., Fathman, C.G., Glimcher, L.H., 2000. Anovel transcription factor T-bet, directs Th1 lineage commitment. Cell 100, 655.
Wang, W., Huang, X.R., Li, A.G., Liu, F., Li, J.H., Truong, L.D., Wang, X.J., Lan, H.Y., 2005a.Signaling mechanism of TGF-beta1 in prevention of renal inflammation: role ofSmad7. J. Am. Soc. Nephrol. 16, 1371.
Wang, W., Soltero, L., Zhang, P., Huang, X.R., Lan, H.Y., Adrogue, H.J., 2007. Renal
inflammation is modulated by potassium in chronic kidney disease: possiblerole of Smad7. Am. J. Physiol. Renal Physiol. 293, F1123.Wang, Y., Feng, X., Bao, S., Yi, S., Kairaitis, L., Tay, Y.C., Rangan, G.K., Harris, D.C.,2001a. Depletion of CD4(+) T cells aggravates glomerular and interstitial injuryin murine adriamycin nephropathy. Kidney Int. 59, 975.

biolog
W
W
W
W
Y
V.O. Reis et al. / Immuno
ang, Y., Mahajan, D., Tay, Y.C., Bao, S., Spicer, T., Kairaitis, L., Ran-gan, G.K., Harris, D.C., 2005b. Partial depletion of macrophages by ED7reduces renal injury in Adriamycin nephropathy. Nephrology (Carlton) 10,470.
ang, Y., Wang, Y.P., Tay, Y.C., Harris, D.C., 2000. Progressive adriamycin nephropa-thy in mice: sequence of histologic and immunohistochemical events. KidneyInt. 58, 1797.
ang, Y., Wang, Y.P., Tay, Y.C., Harris, D.C., 2001b. Role of CD8(+) cells in the pro-gression of murine adriamycin nephropathy. Kidney Int. 59, 941.
u, D.T., Bitzer, M., Ju, W., Mundel, P., Bottinger, E.P., 2005. TGF-beta concentra-tion specifies differential signaling profiles of growth arrest/differentiation andapoptosis in podocytes. J. Am. Soc. Nephrol. 16, 3211.
ap, H.K., Cheung, W., Murugasu, B., Sim, S.K., Seah, C.C., Jordan, S.C., 1999. Th1and Th2 cytokine mRNA profiles in childhood nephrotic syndrome: evidence
y 217 (2012) 831– 841 841
for increased IL-13 mRNA expression in relapse. J. Am. Soc. Nephrol. 10,529.
Yoshida, S., Nagase, M., Shibata, S., Fujita, T., 2008. Podocyte injury induced by albu-min overload in vivo and in vitro: involvement of TGF-beta and p38 MAPK.Nephron Exp. Nephrol. 108, e57.
Zheng, W., Flavell, R.A., 1997. The transcription factor GATA-3 is necessary andsufficient for Th2 cytokine gene expression in CD4 T cells. Cell 89, 587.
Zheng, Z., Pavlidis, P., Chua, S., D’Agati, V.D., Gharavi, A.G., 2006. An ancestral haplo-type defines susceptibility to doxorubicin nephropathy in the laboratory mouse.
J. Am. Soc. Nephrol. 17, 1796.Zheng, Z., Schmidt-Ott, K.M., Chua, S., Foster, K.A., Frankel, R.Z., Pavlidis, P., Barasch,J., D’Agati, V.D., Gharavi, A.G., 2005. A Mendelian locus on chromosome 16 deter-mines susceptibility to doxorubicin nephropathy in the mouse. Proc. Natl. Acad.Sci. U.S.A. 102, 2502.




















