
The ontogeny of immunity in the honey bee, Apis mellifera L. following an immune challenge
10
The ontogeny of immunity in the honey bee, Apis mellifera L. following an immune challenge Alice M. Laughton a,b, *, Michael Boots a , Michael T. Siva-Jothy a a Department of Animal and Plant Sciences, University of Sheffield, Sheffield S10 2TN, UK b Biology Department, Emory University, O. Wayne Rollins Research Center, 1510 Clifton Road NE, Atlanta, GA 30322, USA 1. Introduction The honey bee, Apis mellifera, is eusocial, with a caste system comprising a single queen, up to 80,000 sterile female (diploid) workers, and male (haploid) drones (Wilson, 1971). As a consequence of living in the confines of a hive with high levels of physical contact between closely related conspecifics, bees are likely to be subjected to increased levels of risk of parasitism (Schmid-Hempel, 1998). Despite evolving unique organisational, physiological and behavioural mechanisms to reduce the transfer and establishment of parasites within the colony (Cremer et al., 2007; Evans and Spivak, 2009), the potential risk of parasitic infection is also likely to select for the maintenance of a robust and comprehensive immune system. The innate immune system of insects can be split into constitutive and induced responses based on when they act (Schmid-Hempel, 2005). Constitutive responses comprise cellular responses including coagulation, phagocytosis, nodule formation and encapsulation, and the humoral prophenoloxidase activating system (ProPO-AS) (Gillespie et al., 1997). The ProPO-AS produces the enzyme phenoloxidase, and is (a) responsible for the biosynthesis of melanin, (b) involved in cuticle sclerotisation, (c) wound healing, (d) nodule formation, (e) encapsulation, and (f) phagocytosis stimulation (Mason, 1955; Ratcliffe et al., 1984; Cerenius et al., 2008). These immune responses are always present, although not maintained at maximum levels (Siva-Jothy et al., 2005; Cerenius et al., 2008). They are non-specific, and immediate in effect (Gillespie et al., 1997; Schmid-Hempel, 2005). By contrast, induced responses, which include antimicrobial peptides (AMPs) and lysozyme-like-activity, require specific pathogen recognition to stimulate their production. As such, they take longer to produce, but are more targeted and longer lasting in their effect (Boman and Hultmark, 1987). A combination of the level of parasite threat and the developmental stage may influence how much investment an individual bee will allocate to their immune system. The expression of immunity-related genes is known to differ both within and between honey bee developmental stages, but little is known about the impact of this variation on immune responses (Evans, 2004; Chan et al., 2006). Cellular and antimicrobial responses have been shown to vary with developmental stage Journal of Insect Physiology 57 (2011) 1023–1032 A R T I C L E I N F O Article history: Received 14 December 2010 Received in revised form 23 April 2011 Accepted 26 April 2011 Keywords: Honey bee Ontogeny Immunity Phenoloxidase Temporal polyethism Sex biased immunity A B S T R A C T The honey bee, Apis mellifera, is an ideal system for investigating ontogenetic changes in the immune system, because it combines holometabolous development within a eusocial caste system. As adults, male and female bees are subject to differing selective pressures: worker bees (females) exhibit temporal polyethism, while the male drones invest in mating. They are further influenced by changes in the threat of pathogen infection at different life stages. We investigated the immune response of workers and drones at all developmental phases, from larvae through to late stage adults, assaying both a constitutive (phenoloxidase, PO activity) and induced (antimicrobial peptide, AMP) immune response. We found that larval bees have low levels of PO activity. Adult workers produced stronger immune responses than drones, and a greater plasticity in immune investment. Immune challenge resulted in lower levels of PO activity in adult workers, which may be due to the rapid utilisation and a subsequent failure to replenish the constitutive phenoloxidase. Both adult workers and drones responded to an immune challenge by producing higher titres of AMPs, suggesting that the cost of this response prohibits its constant maintenance. Both castes showed signs of senescence in immune investment in the AMP response. Different sexes and life stages therefore alter their immune system management based on the combined factors of disease risk and life history. ß 2011 Elsevier Ltd. All rights reserved. * Corresponding author at: Biology Department, Emory University, O. Wayne Rollins Research Center, 1510 Clifton Road NE, Atlanta, GA 30322, USA. Tel.: +1 404 727 9344; fax: +1 404 727 2880. E-mail addresses: [email protected] (A.M. Laughton), m.boots@sheffield.ac.uk (M. Boots), m.siva-jothy@sheffield.ac.uk (M.T. Siva-Jothy). Contents lists available at ScienceDirect Journal of Insect Physiology jo u rn al h om ep ag e: ww w.els evier.c o m/lo c ate/jin sp h ys 0022-1910/$ – see front matter ß 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.jinsphys.2011.04.020
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The ontogeny of immunity in the honey bee, Apis mellifera L. following an immune challenge

Journal of Insect Physiology 57 (2011) 1023–1032
The ontogeny of immunity in the honey bee, Apis mellifera L. followingan immune challenge
Alice M. Laughton a,b,*, Michael Boots a, Michael T. Siva-Jothy a
a Department of Animal and Plant Sciences, University of Sheffield, Sheffield S10 2TN, UKb Biology Department, Emory University, O. Wayne Rollins Research Center, 1510 Clifton Road NE, Atlanta, GA 30322, USA
A R T I C L E I N F O
Article history:
Received 14 December 2010
Received in revised form 23 April 2011
Accepted 26 April 2011
Keywords:
Honey bee
Ontogeny
Immunity
Phenoloxidase
Temporal polyethism
Sex biased immunity
A B S T R A C T
The honey bee, Apis mellifera, is an ideal system for investigating ontogenetic changes in the immune
system, because it combines holometabolous development within a eusocial caste system. As adults,
male and female bees are subject to differing selective pressures: worker bees (females) exhibit temporal
polyethism, while the male drones invest in mating. They are further influenced by changes in the threat
of pathogen infection at different life stages. We investigated the immune response of workers and
drones at all developmental phases, from larvae through to late stage adults, assaying both a constitutive
(phenoloxidase, PO activity) and induced (antimicrobial peptide, AMP) immune response. We found that
larval bees have low levels of PO activity. Adult workers produced stronger immune responses than
drones, and a greater plasticity in immune investment. Immune challenge resulted in lower levels of PO
activity in adult workers, which may be due to the rapid utilisation and a subsequent failure to replenish
the constitutive phenoloxidase. Both adult workers and drones responded to an immune challenge by
producing higher titres of AMPs, suggesting that the cost of this response prohibits its constant
maintenance. Both castes showed signs of senescence in immune investment in the AMP response.
Different sexes and life stages therefore alter their immune system management based on the combined
factors of disease risk and life history.
� 2011 Elsevier Ltd. All rights reserved.
Contents lists available at ScienceDirect
Journal of Insect Physiology
jo u rn al h om ep ag e: ww w.els evier .c o m/lo c ate / j in sp h ys
1. Introduction
The honey bee, Apis mellifera, is eusocial, with a caste systemcomprising a single queen, up to 80,000 sterile female (diploid)workers, and male (haploid) drones (Wilson, 1971). As aconsequence of living in the confines of a hive with high levelsof physical contact between closely related conspecifics, bees arelikely to be subjected to increased levels of risk of parasitism(Schmid-Hempel, 1998). Despite evolving unique organisational,physiological and behavioural mechanisms to reduce the transferand establishment of parasites within the colony (Cremer et al.,2007; Evans and Spivak, 2009), the potential risk of parasiticinfection is also likely to select for the maintenance of a robust andcomprehensive immune system.
The innate immune system of insects can be split intoconstitutive and induced responses based on when they act(Schmid-Hempel, 2005). Constitutive responses comprise cellular
* Corresponding author at: Biology Department, Emory University, O. Wayne
Rollins Research Center, 1510 Clifton Road NE, Atlanta, GA 30322, USA.
Tel.: +1 404 727 9344; fax: +1 404 727 2880.
E-mail addresses: [email protected] (A.M. Laughton),
[email protected] (M. Boots), [email protected] (M.T. Siva-Jothy).
0022-1910/$ – see front matter � 2011 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jinsphys.2011.04.020
responses including coagulation, phagocytosis, nodule formationand encapsulation, and the humoral prophenoloxidase activatingsystem (ProPO-AS) (Gillespie et al., 1997). The ProPO-AS producesthe enzyme phenoloxidase, and is (a) responsible for thebiosynthesis of melanin, (b) involved in cuticle sclerotisation, (c)wound healing, (d) nodule formation, (e) encapsulation, and (f)phagocytosis stimulation (Mason, 1955; Ratcliffe et al., 1984;Cerenius et al., 2008). These immune responses are always present,although not maintained at maximum levels (Siva-Jothy et al.,2005; Cerenius et al., 2008). They are non-specific, and immediatein effect (Gillespie et al., 1997; Schmid-Hempel, 2005). By contrast,induced responses, which include antimicrobial peptides (AMPs)and lysozyme-like-activity, require specific pathogen recognitionto stimulate their production. As such, they take longer to produce,but are more targeted and longer lasting in their effect (Boman andHultmark, 1987).
A combination of the level of parasite threat and thedevelopmental stage may influence how much investment anindividual bee will allocate to their immune system. Theexpression of immunity-related genes is known to differ bothwithin and between honey bee developmental stages, but little isknown about the impact of this variation on immune responses(Evans, 2004; Chan et al., 2006). Cellular and antimicrobialresponses have been shown to vary with developmental stage

A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–10321024
in several insect systems (Natori et al., 1999; Chung and Ourth,2000; Evans and Lopez, 2004; Meylaers et al., 2007; Freitak et al.,2007; Dubovskiy et al., 2008; Eleftherianos et al., 2008; Wilson-Rich et al., 2008), with small changes in age resulting in significantchanges in the immune response (e.g. Eleftherianos et al., 2008;Wilson-Rich et al., 2008). Larval honey bees, housed within broodcells, are susceptible to bacterial, fungal, viral and ectoparasiticinfections that can severely hamper their development andsurvival (Bailey and Ball, 1991; Allen and Ball, 1996; Hooper,1997). Due to their limited behavioural means of avoidingpathogens, developing honey bees may increase investment intheir immune response (Wilson-Rich et al., 2008); there is someevidence that larval and pupal honey bees have higher totalhaemocyte counts than adults (Wilson-Rich et al., 2008, althoughsee Schmid et al., 2008 for conflicting result). Conversely,phenoloxidase activity in honey bee pupae has been shown toincrease successively as pupae age and increase their cuticularmelanisation (Zufelato et al., 2004).
Adult honey bees are also subject to infection by a range ofpathogens (Bailey and Ball, 1991), although relatively few overlapwith those infecting larval and pupal stages (Bailey, 1968; Bailey andBall, 1991). Opportunities to contract infections are likely to beclosely linked to bee activity, which is, in turn, associated withdevelopmental age. Adult worker honey bees exhibit age polyeth-ism, a temporary, dynamic division of labour, with younger (nurse)workers employed within the hive, feeding and tending todeveloping larvae, and cleaning or grooming (Robinson, 1992). At2–3 weeks old, nurses switch to become foragers, leaving the colonyeach day to collect pollen and nectar (Hooper, 1997). While leavingthe hive exposes foragers to a higher risk of pathogen infection, andmay therefore favour an increased level of immune investment(Bocher et al., 2007), the process of age polyethism selects nutrientdepleted foragers to undertake the most hazardous tasks towardsthe end of their life, and therefore the ability to invest in an immuneresponse may be reduced. The transition to foragers in bumble beeshas been shown to correlate with a decrease in ‘‘immunocompe-tence’’ (Konig and Schmid-Hempel, 1995; Doums and Schmid-Hempel, 2000), but evidence for this in the honey bee is mixed(Amdam et al., 2005; Schmid et al., 2008; Wilson-Rich et al., 2008).The transition from nurse bee to forager is brought about byphysiological and behavioural changes (Robinson, 1992), including adecrease in the yolk precursor vitellogenin (Robinson, 1987; Omholtand Amdam, 2004), which causes pycnosis (cell death) of thehaemocytes (Falchuk, 1998; Amdam et al., 2004, 2005). A decrease infunctioning haemocyte number may reduce the bee’s capacity forcellular-based immune responses (Robinson, 1992; Bedick et al.,2001; Amdam et al., 2004, 2005; Omholt and Amdam, 2004), asshown by Bedick et al. (2001). Honey bee foragers also have a lowerfat body mass than nurse bees (Wilson-Rich et al., 2008). The fatbody synthesises haemolymph proteins including AMPs andlysozymes (Hultmark, 1996; Lehrer and Ganz, 1999; Tzou et al.,2000). It is therefore potentially possible that a reduction in fat bodymass could lead to a depleted ability to initiate the induced arm ofthe immune system. An increase in phenoloxidase activity inforagers compared to nurse bees (Wilson-Rich et al., 2008) maycompensate for the reduction of more ‘‘costly’’ induced immuneresponses with age.
While most work to date has focussed on worker bees (e.g.Navajas et al., 2008; Wilson-Rich et al., 2008; Chan et al., 2009),examining the drone (male) bees provides an opportunity toinvestigate the immune investment of sexuals in the colony(logistic constraints did not allow us to examine queen bees in thisstudy). Several insect systems show a trade-off between repro-ductive effort and immune investment (e.g. Adamo et al., 2001;Rolff, 2002; Simmons and Roberts, 2005). Although adult honeybee drones have different behavioural patterns to workers (drones
have no age polyethism, as their sole function is to mate with aqueen (Hooper, 1997)), both castes return to the hive at night,providing opportunities for the transfer of novel pathogens, whichmay select for the maintenance of a comparable immune system indrones (Hooper, 1997; Boomsma et al., 2005; Cremer et al., 2007).However, bumblebee drones were found to have a less intenseencapsulation response than workers (Gerloff et al., 2003; Baer andSchmid-Hempel, 2006), although there was no difference betweenmale and female susceptibility to transmissible parasites (Ruiz-Gonzalez and Brown, 2006). In studies on ants, males were foundto exhibit lower immune responses than either workers (Baer et al.,2005) or queens (Vainio et al., 2004).
By comparing the ontogeny of immunity in honey bee workerand drone castes throughout adulthood we can examine the agingof the immune system. Few studies have followed the changes thatoccur in invertebrate immune systems throughout development,metamorphosis and subsequent adult aging (Schmid et al., 2008;Wilson-Rich et al., 2008; Moret and Schmid-Hempel, 2009).Commonly, aging in both vertebrates and invertebrates isassociated with increased activation of the innate immune system(DeVeale et al., 2004; Zerofsky et al., 2005), but the maintenance ofan immune system throughout life is costly (e.g. Kaaya and Darji,1988; Zerofsky et al., 2005; Konig and Schmid-Hempel, 1995;Moret and Schmid-Hempel, 2000). Adult honey bee drones arerelatively short-lived compared to workers (22 days compared to36 days for workers during summer months (Hooper, 1997)) somay invest even less in an immune response with increasing age,instead channelling their resources into sperm production andmating flights.
Here we assay a component of the constitutive (phenoloxidase)and induced (antimicrobial peptides, AMP) immune responses(Barnes and Siva-Jothy, 2000; Thompson, 2002) in worker anddrone honey bees throughout larval development and adultontogeny. In addition, we stimulate an immune challenge in thehoney bees to determine whether assayed immune parameters arebeing utilised in immune defence or required in a different guise aspart of the development process. We predict that developing larvaland pupal honey bees will have a low immune response andreduced ability to respond to an immune challenge. Our argumentis that developing larvae and pupae reside in a relatively hygienicenvironment attended by nurse bees and are therefore selected toinvest resources into growth and development rather thanimmunity. Honey bee drones are predicted to have lower levelsof immune investment than workers throughout their life, and relyon the less costly phenoloxidase rather than induced AMPresponses as their primary defence.
2. Materials and methods
2.1. Bees
For most experiments, worker and drone bees were eachcollected from two different colonies of healthy British hybrid bees(A. mellifera L.) maintained at the University of Sheffield. Theexception to this is the adult drones collected for the ontogeny ofphenoloxidase and antimicrobial peptide experiments. In thesecases, poor weather resulted in minimal drone production in somecolonies, and consequently results are presented from dronescollected from a single colony. While the two different sexes werecollected from colonies matched in terms of size, health and broodproduction, these were not always the same colonies. Whereneeded, brood frames were incubated overnight (36 8C � 1 8C, 24 hdark photoperiod, 40% � 2% humidity) and newly emerged adultscollected the following morning. Adult bees were maintained in cages(36 8C � 1 8C, 12:12 h L:D photoperiod, 50% sucrose solution ad
libitum). All experimental bees were destructively sampled.

A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–1032 1025
2.2. Immune challenge
In all experiments we stimulated an immune challenge bythe injection of 1 ml lipopolysaccharide (LPS, 0.5 mg ml�1) ininsect Ringer (128 mM NaCl, 18 mM CaCl2, 1.3 mM KCl, 2.3 mMNaHCO3, 1 l dH2O, pH 7.2). LPS (derived from Escherichia coli) is acomponent of the outer membrane of Gram-negative bacteria. Itis an endotoxin and is widely used as an elicitor of the immuneresponse in insects (e.g. Moret and Schmid-Hempel, 2000). Inaddition, three control groups of (a) a full control (No Challenge),(b) a sterile needle stab (Needle), and (c) a vehicular controlinjection with 1 ml Ringer solution (Ringer) were used. Haemo-coelic injections were made using a glass microcapillary pulledto a fine needle (130 mm diameter). Survival analysis frompreliminary data showed a significant effect of treatment onadult worker bee survival (GLM, p < 0.001, d.f. = 3,281, gammaerrors), with the challenge of LPS leading to a reduction inlongevity (data not shown). However, the differential effects ofsurvival only began to manifest at 48 h post-challenge; as all ourexperimental samples were collected at 24 h post-challenge,reduced survival is not likely to impact on the data presentedhere.
2.3. Assaying phenoloxidase
Phenoloxidase is highly immunologically active, generating arange of cytotoxins when switched on, so is generally stored asits inactive precursor prophenoloxidase. Phenoloxidase wasmeasured as the total potential phenoloxidase response (POactivity), measured after the artificial activation of propheno-loxidase into phenoloxidase with a-chymotrypsin, a commonprophenoloxidase activator (Saul and Sugumaran, 1988; Kopa-cek et al., 1995). Levels of PO activity were assayed through itscatalysis of the conversion of L-dopa (3,4-dihydroxy-L-phenylal-anine, colourless) to dopachrome (red-brown colour), measuredphotometrically (Horowitz and Shen, 1952; Barnes and Siva-Jothy, 2000; Thompson, 2002). Following Laughton and Siva-Jothy (2010), 20 ml aliquots of haemolymph were mixed with135 ml dH2O and 20 ml PBS (phosphate buffered solution;150 mM NaCl, 10 mM Na2HPO4, pH 6.5) and 5 ml a-chymotryp-sin (0.5 mg ml�1) in a pre-chilled 96-well plate. The reactionmixture was incubated at room temperature (19 8C � 2 8C) for5 min to allow activation, before adding 20 ml L-dopa (saturated,between 2.88 mM and 5.75 mM; solution filtered before use) andrecording started at 30 8C in a spectrophotometer (VersaMaxtunable microplate reader, Molecular Devices). Readings weretaken at 490 nm every 15 s for 1 h and analysed using Softmax Prov5.0.1 software. Enzyme activity was measured as the maximumlinear rate of substrate conversion.
2.4. Developmental immunity
Developmental immunity was assessed as PO activity inworkers and drones at five different developmental stages: 4thinstar larvae (uncapped), 5th instar larvae (capped), white-eyedpupae (white-eyed), red-eye pupae (red-eyed) and newly eclosedadults (adult). Preliminary experiments found that phenolox-idase could not be measured reliably in earlier instar larvae(data not shown). Bees were gauged on their developmentalstage based on size, positioning within the cell and, in the caseof pupae, eye and thorax colouring (Hooper, 1997; Micheletteand Soares, 1993). They were removed from their brood cellsand maintained at a high humidity (45% humidity � 5%,36 8C � 1 8C, 24 h dark photoperiod) and, in the case of larvae,provided with brood food. Adults were incubated in bee boxesprovided with 50% sucrose solution ad libitum. Body mass (wet
weight) was recorded as a measure of body size. Each of the 5developmental stages was split into four groups to measure theeffect of an immune challenge on immune response (No Challenge,Needle, Ringer, LPS; worker n = 29–128, drone n = 14–67 pertreatment:age group). Individuals were assessed for survival24 h post-challenge, and haemolymph samples from survivingbees were collected via perfusion bleeds with 0.25 ml sodiumcacodylate buffer (NaCac: 0.001 M sodium cacodylate, 0.005 Mcalcium chloride, pH 6.5). Samples were immediately frozen(�80 8C) for a minimum 24 h to disrupt haemocytes. Prior touse, samples were defrosted and centrifuged (4 8C, 8000 � g,15 min) and assayed for PO activity as outlined above. Due to theconstraints of collecting and processing developing bees, sampleswere collected on several different dates over a period of 3 weeks.The date of collection was included in the analysis.
2.5. Adult ontogeny and phenoloxidase
To investigate the difference in immune responses of newlyemerged and old adult bees, workers and drones were collected attwo distinctly different life-stages. New adult worker bees werecollected as they emerged from a brood frame incubatedovernight (36 8C � 1 8C, 40% � 2% humidity). Old adult worker beeswere collected as they returned to the hive following a foraging trip.Although not directly aged, old worker bees had to exhibit foragingactivity (pollen collected on legs) in order to distinguish them fromyounger workers making test flights, and were likely to be between14 and 36 days old (Hooper, 1997). As drones do not forage, thismethod could not be employed. Instead, newly emerged adultdrones were collected and marked with a spot of non-toxic paint onthe thorax. Drones were released immediately into their natal hive,and marked bees were recaptured when 2 weeks old. This agecorresponds to the earliest foraging workers, although as drones areshorter lived they are approximately two thirds through adult life atthis time. To examine how the different ages responded to animmune challenge, each age group was split into four treatmentgroups (No Challenge, Needle, Ringer, LPS; worker n = 53–79, dronen = 11–26 for each age:treatment group). Treatment groups werehoused in separate bee boxes and haemolymph samples collected24 h post-challenge via perfusion bleed with 0.5 ml NaCac buffer.Samples were immediately frozen (�80 8C) for a minimum 24 h todisrupt haemocytes. Subsequently, samples were defrosted andcentrifuged (4 8C, 8000 � g, 15 min), before being assayed for POactivity (see above). Thorax width was recorded as a measure of bodysize.
2.6. Adult ontogeny and antimicrobial peptides
Adult workers and drones were sampled at two extremes of age(new and old) as described in the previous section. To investigatethe effect of an immune challenge, the bees in each age group werefurther split into four treatment groups (No Challenge, Needle,Ringer and LPS; worker n = 33–48, drone n = 5–31). Treatmentgroups were housed in separate bee boxes and neat haemolymphwas collected 24 h post-challenge via a leg wound, and frozenimmediately for 24 h (�80 8C). Samples were assayed for AMPactivity using a zone of inhibition assay (Haine et al., 2008), with1 ml neat haemolymph assayed for each bee. Briefly, a colony ofArthrobacter globiformis was grown overnight in 10 ml Luria broth(LB, 30 8C), and 100 ml of the resulting broth added to 50 ml 1% agarsolution and poured into Petri dishes. Wells were punched into theagar (2 mm2), and the haemolymph samples defrosted on ice,loaded into the wells and incubated (48 h, 30 8C). Clear zones ofinhibition were photographed and measured to calculate area(each zone measured three times, and mean area calculated usingQCapture Pro 51). Phenoloxidase in the samples lead to the

A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–10321026
production of melanin, seen as dark areas surrounding the wells.However, preliminary experiments using PO inhibitors such asphenylthiourea (PTU) found no effect of the presence of melanin onthe inhibition assay (Haine, unpubl. data). Thorax width wasrecorded as a measure of body size.
2.7. Analysis
In all experiments, the No Challenge and LPS treated groups werestatistically at the two extremes of the range of immune responsesrecorded. All analyses presented here were carried out on the fulldata sets incorporating all four treatment groups. However, forclarity, graphical data are presented for the No Challenge and LPS
treated groups only. Data were analysed using the statisticalpackage R (version 2.6.2) on the Mac OS X platform, and presented asthe minimal adequate model, with body size (wet body mass orthorax width) controlled for in all models. Where data from twocolonies were collected, colony effect was included as a fixed effectcontrol measure in all models. The date of sample collection was alsoincluded as a fixed effect in the developmental immunity models.Where appropriate, data were log 10 or square root transformedbefore analysis to conform to the assumptions of a normaldistribution. All figures present raw data for ease of interpretation.
Fig. 1. PO developmental immunity in worker bees at five developmental stages (1
5 = adults) for the No Challenge and LPS treatment groups.
Bee image portions reprinted by permission of the publisher from The Biology of the Ho
Copyright� 1987 by Mark L. Winston.
3. Results
3.1. Developmental immunity
3.1.1. Workers
There was a significant interaction between age and treatmenton PO activity (Fig. 1, ANOVA, F12,1140 = 3.56, p < 0.0001, datalog 10 transformed). In all treatments, including the Needle andRinger groups, PO activity was maintained at a relatively constantlevel throughout larval development, but increased following adulteclosion. PO activity was lower overall in LPS challenged bees(there was a significant main effect of treatment (ANOVA,F3,1140 = 20.2, p < 0.0001, data log 10 transformed)), while theinteraction effect arose from the higher levels of variation seenthroughout the No Challenge group during development, coupledwith the relatively larger increase in PO activity at eclosion. TheNeedle and Ringer treatment groups also showed a slight decreasein PO activity in the adult stage, but to a lesser extent than seen inthe LPS treatment group. There was a significant effect of body size(ANOVA, F1,1140 = 15.5, p < 0.0001, data log 10 transformed),colony (ANOVA, F1,1140 = 10.6, p < 0.01, data log 10 transformed)and collection date (ANOVA, F10,1140 = 92.8, p < 0.0001, data log 10transformed) on PO activity.
= uncapped larvae, 2 = capped larvae, 3 = white-eyed pupae, 4 = red-eyed pupae,
ney Bee by Mark L. Winston, pp. 50–51, Cambridge, MA: Harvard University Press,
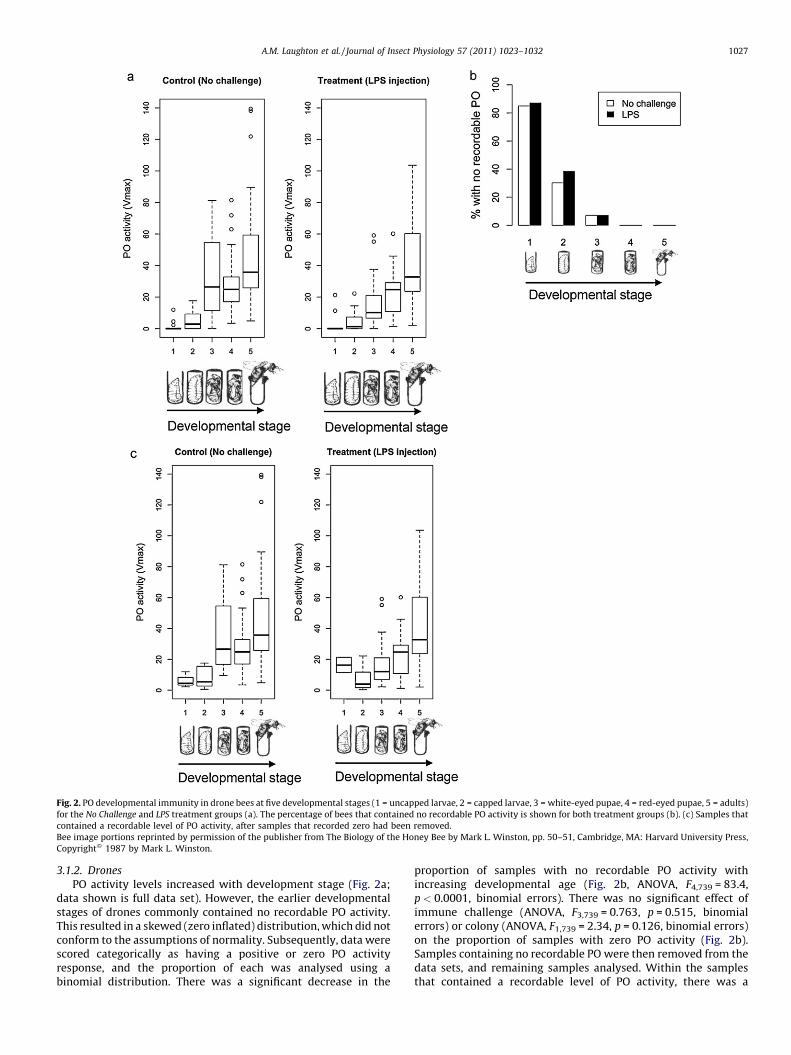
Fig. 2. PO developmental immunity in drone bees at five developmental stages (1 = uncapped larvae, 2 = capped larvae, 3 = white-eyed pupae, 4 = red-eyed pupae, 5 = adults)
for the No Challenge and LPS treatment groups (a). The percentage of bees that contained no recordable PO activity is shown for both treatment groups (b). (c) Samples that
contained a recordable level of PO activity, after samples that recorded zero had been removed.
Bee image portions reprinted by permission of the publisher from The Biology of the Honey Bee by Mark L. Winston, pp. 50–51, Cambridge, MA: Harvard University Press,
Copyright� 1987 by Mark L. Winston.
A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–1032 1027
3.1.2. Drones
PO activity levels increased with development stage (Fig. 2a;data shown is full data set). However, the earlier developmentalstages of drones commonly contained no recordable PO activity.This resulted in a skewed (zero inflated) distribution, which did notconform to the assumptions of normality. Subsequently, data werescored categorically as having a positive or zero PO activityresponse, and the proportion of each was analysed using abinomial distribution. There was a significant decrease in the
proportion of samples with no recordable PO activity withincreasing developmental age (Fig. 2b, ANOVA, F4,739 = 83.4,p < 0.0001, binomial errors). There was no significant effect ofimmune challenge (ANOVA, F3,739 = 0.763, p = 0.515, binomialerrors) or colony (ANOVA, F1,739 = 2.34, p = 0.126, binomial errors)on the proportion of samples with zero PO activity (Fig. 2b).Samples containing no recordable PO were then removed from thedata sets, and remaining samples analysed. Within the samplesthat contained a recordable level of PO activity, there was a

Fig. 3. Adult ontogeny and PO activity in adult worker (a) and drone (b) bees for the No Challenge and LPS treatment groups. There was a significant effect of age and treatment
on PO activity levels.
A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–10321028

A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–1032 1029
significant interaction of age with treatment (Fig. 2c, ANOVA,F11,639 = 1.99, p = 0.027). PO activity was lower overall in LPS
challenged bees (there was a significant main effect of treatment(ANOVA, F3,639 = 2.79, p = 0.040)), and increased sequentially withage (significant main effect of age (ANOVA, F4,639 = 32.6,p < 0.0001)). Adult bees in the Needle and Ringer treatment groupsalso showed a slight decrease in PO activity in the adult stage, butto a lesser extent than seen in the LPS treatment group. As with theworkers, the interaction effect arose from the high levels ofvariation within those samples that contained a recordable level ofPO activity. There was also a significant effect of body size (ANOVA,F1,639 = 270, p < 0.0001), colony (ANOVA, F1,639 = 17.9, p < 0.0001),and collection date (ANOVA, F4,639 = 9.29, p < 0.0001) on POactivity.
3.2. Adult ontogeny and phenoloxidase
3.2.1. Workers
All treatment groups recorded a higher level of PO activity in theold adult bees compared to the new adult bees (Fig. 3a). There wasa significant interaction with age and immune challenge on POactivity (ANOVA, F3,524 = 4.01, p < 0.01, data log 10 transformed);treatment with LPS resulted in lower levels of PO activity, but onlyin the new bees. This response was not seen in any of the othertreatment groups. There was a significant effect of colony on POactivity (ANOVA, F1,524 = 149.09, p < 0.0001, data log 10 trans-formed).
3.2.2. Drones
There was no significant effect of age on PO activity in adultdrones (Fig. 3b, ANOVA, F1,147 = 2.40, p = 0.123). There was asignificant effect of treatment on PO activity, with the LPS
challenged groups having lower levels of PO activity than theNo Challenge groups (Fig. 3b, ANOVA, F3,148 = 2.97, p = 0.0340, datalog 10 transformed). Old drones receiving in the Ringer treatmentgroup also showed a decrease in PO activity, although not to theextent of those in the LPS treatment group.
3.3. Adult ontogeny and antimicrobial peptides
3.3.1. Workers
There was a significant interaction between age and immunechallenge (Fig. 4a, ANOVA, F3,301 = 16.91, p < 0.0001, data squareroot transformed). AMP production increased following animmune challenge with Needle, Ringer and LPS in both age groups.In the No Challenge, Needle and Ringer groups, AMP production wasseen to increase in old adult bees. The LPS challenged groupshowed the opposite pattern, with a decrease in AMP productionwith increasing adult age. There was no significant effect of colonyon worker AMP production (ANOVA, F1,301 = 0.002, p = 0.963, datasquare root transformed).
3.3.2. Drones
A significant number of the No Challenge treatment samplesdid not produce a recordable AMP response (58% of young and71% of old drones), which produced a skewed (zero inflated)distribution (F3,114 = 18.8, p < 0.0001, binomial errors). Samplesthat did not contain a recordable level of AMPs were removedfrom the data set before analysis, so that data conformed tonormality. Following the removal of non-recording individuals,there was a significant interaction of age and treatment on AMPconcentration (Fig. 4b, ANOVA, F2,83 = 6.29, p < 0.01, data squareroot transformed). AMP activity increased following an immunechallenge: there was a slight increase in AMP production in theNeedle and Ringer groups, but drones showed a much greaterresponse to challenge with LPS. AMP production was lower in
old, LPS challenged drones, but this pattern was not seen in theother treatment groups.
4. Discussion
This work presents a unique insight into the ontogeny ofimmunity in the honey bee. Incorporating immune challenges andinvestigating multiple life stages of both workers and drones hasallowed us to examine bee immune responses in greater depth, andunderstand their relative importance in host immunity at differentlife stages.
We show that variation in immune activity is associated withdifferent phases of honey bee development within both workerand drone castes. Both workers and drones had lower levels of POactivity during larval and pupal stages than as adults, supportingprevious findings (Lourenco et al., 2005; Wilson-Rich et al., 2008;Chan et al., 2009). Phenoloxidase production may only increasewhen required for cuticle melanisation (Chmurzynska andWojtczak, 1963; Hopkins and Kramer, 1992), despite larval stagesbeing susceptible to many different pathogens (Bailey and Ball,1991). Only developing workers responded to an immunechallenge with LPS. The counter intuitive decrease in PO activityfollowing such a challenge may be due to the rapid utilisation ofphenoloxidase in the immune response, and a subsequent failureto replenish the supply. This trend has also been seen in the Africanarmyworm, Spodoptera exempta, and damselfly larvae, Ischnura
elegans (Povey, Slos, pers. comm.). Alternatively, PO activity may betraded off with other immune responses, such as AMP production,which was not assayed in developing bees. In larval drones, no suchresponse was seen. Although PO activity increased sequentiallyduring drone development, earlier developmental stages exhibitedlower levels than those seen in workers, suggesting that at thesestages drones have a lower level of immune investment thanworkers. Furthermore, the lack of a response 24 h after an immunechallenge suggests that in developing larval drones, phenoloxidasemay not be an important component of the immune response,although the ability of developing drone phenoloxidase to respondto alternative immune challenges was not tested. Alternativeimmune measures, such as increased haemocyte densities(Wilson-Rich et al., 2008), combined with the hygienic behavioursadministered by nurse bees (Evans and Spivak, 2009) may providea sufficient defence.
Age polyethism resulted in a universal up-regulation in POactivity with increasing age in adult worker bees. Previous researchsuggests that the opposite should be true, as the increase inhaemocyte apoptosis with transition from hive bee to foragershould reduce the level of cellular immune responses available(Robinson, 1992; Bedick et al., 2001; Amdam et al., 2004, 2005;Omholt and Amdam, 2004). However, despite assaying differentmeasures of phenoloxidase activity, in addition to our work hereseveral recent studies have also found an increase in workerphenoloxidase activity with age, despite recording a juxtaposeddecrease in haemocyte counts (Schmid et al., 2008; Wilson-Richet al., 2008). Contrasting results have been found on the relativehaemocyte counts of nurse and foraging worker bees (Amdamet al., 2005; Schmid et al., 2008; Wilson-Rich et al., 2008). Thissuggests that variation in haemocyte numbers may depend onfactors other than developmental age, and that such counts maynot have a bearing on PO activity. Alternatively, as haemocytestypes have different functions in different insects, natural selectionmay act towards the survival of haemocytes with immune activitywhile causing pycnosis in those cell types with no link to immunity(Lavine and Strand, 2002).
Following an immune challenge with LPS, PO activity levelsdecreased in new adult bees while the response was reduced orlost altogether in the old adult bees. One explanation is that old

Fig. 4. Adult ontogeny and AMP activity in adult worker (a) and drone (b) bees for the No Challenge and LPS treatment groups. There was a significant interaction between age
and treatment on AMP activity for both castes.
A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–10321030

A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–1032 1031
bees may maintain a constant maximum level of phenoloxidaseproduction; if they face a continuous pathogen threat whileforaging, the mechanisms for manufacturing and replacingphenoloxidase as it is used up in the immune response mayalready be operational. Unlike workers, PO activity in drones wasmaintained at a relatively constant level throughout adult life thatwas lower than that found in corresponding worker bees. LPS
challenged drones had a lower PO activity response than thecontrols, which suggests that, as in workers, phenoloxidase doeshave a role in immunity in adult drone life, all be it at a reducedlevel to that of workers. Ringer challenged drones also showed aslight decrease in PO activity in old drones, suggesting that olddrones may be more sensitive to immune stimulation, althoughthis response was not seen in the Needle group, indicating that it isnot a function of cuticle wounding.
In line with previous studies, AMP production was found toincrease in both adult workers and drones following an immunechallenge with LPS (Boulanger et al., 2001, 2002; Evans, 2004). Inaddition, No Challenge, Needle and Ringer challenged workersshowed an increase in AMP activity with age. As predicted, theAMP response of drones was always lower than that of thecorresponding worker group, suggesting that following a challengewith LPS, drones have fewer resources to invest in this immuneresponse, while workers were shown to maintain AMP productioneven when an immune challenge was absent. Both castes exhibiteda decrease in AMP activity with age, but only in the LPS challengedbees, which may be a sign of immune senescence (Doums et al.,2002; Hillyer et al., 2005). Recent research found a decrease in thefat body mass in old bees compared to new bees (Wilson-Rich et al.,2008). Although the reduced fat body still produced AMPs in oldbees, age-imposed limits on functioning ability would support thedecrease in AMP production seen in responding to the challenge ofLPS in old bees. The increase in PO activity in foraging workers mayrepresent a compensatory effort by the immune system once theability to invest in innate responses falls into decline. In addition,sperm viability in drones reaches its peak in 2-week old adults,which may cause resources from the fat body to be channelled intosperm production in the older, sexually mature males (Page andPeng, 2001). It should however be noted that successfully mateddrones die post-copulation (Hooper, 1997). Therefore, by defini-tion, old drones recaptured for this work were, as yet, unsuccessfulat finding a mate. Failure to mate may be due to the drone being ofpoor quality overall, including their immune system. Alternatively,on entering the final week of their life with no mating success,drones may redirect resources away from immune responses andtowards mating flights or sperm production efforts, as any(negative) trade-off between energy expended for mating andlifespan is reduced (Rueppell et al., 2004).
Due to our desire to gather data from a high number ofindividuals, our work presented here focussed only on theresponses of two (and sometimes one) different colonies perexperiment. Colony had a significant effect in almost all ouranalyses but bees from different colonies exhibited qualitativelysimilar responses. While outside the scope of this work, colonyvariation may prove to be an important factor in both individualbee, and colony immunity and survival.
These results shed light on the ontogenetic differences existingbetween immune responses from the constitutive and inducedimmune systems in a eusocial insect, and point to an adaptiveontogenetic shift in the immune responses of the honey bee. Whilethe results for PO activity suggest that at certain life stages its usemay not directly relate to the constitutive immune response, butrather may be tied up in alternative physiological processesrelating to growth and development, the AMP results explicitlyshow such adaptations directly linked to caste and age. Tounderstand the consistently lower immune responses seen in
drones compared to workers, we must consider life history as wellas age. While there is much evidence that that males are the ‘‘sickersex’’ (Zuk, 2009), and it may be easy to add honey bee drones intothis category, the eusocial, haplo-diploid mating system of beesshould also be taken into consideration. The added complexities ofthis unusual organisation mean that bees differ both in terms ofsexual investment and behaviour within each caste and sex. Whilethe selection pressures that this exerts on workers and drones (andqueens) may ultimately result in the same immunologicalphenotypes seen in ‘‘traditional’’ mating systems, the underlyingcauses are likely to be multifaceted.
Our work adds to the growing body of research seeking tounderstand the complexities of the immune system of theeconomically important honey bee. While this subject has gainedmomentum in recent years, due in part to the phenomenon ofcolony collapse disease (Oldroyd, 2007), further research is neededto unravel the relative roles of constituents of the immune systemto natural pathogenic infections. In addition, while workersaccount for the majority of colony members, and correspondinglythe largest body of research to date, drones offer a secondaryplatform with which to study the impact of sexual life historystrategies on the immune response, which merits furtherinvestigation.
Acknowledgements
This research was funded by the Natural Environment ResearchCouncil. Thanks to Steve Martin for advice during this project, andto the Gerardo and de Roode labs for comments on the manuscript.
References
Adamo, S.A., Jensen, M., Younger, M., 2001. Changes in lifetime immunocompetencein male and female Gryllus texensis (formerly G. integer)*: trade-offs betweenimmunity and reproduction. Animal Behaviour 62, 417–425.
Allen, M., Ball, B., 1996. The incidence and world distribution of honey bee viruses.Bee World 77, 141–162.
Amdam, G.V., Aase, A.L.T.O., Seehuus, S., Fondrk, M.K., Norberg, K., Hartfelder, K.,2005. Social reversal of immunosenescence in honey bee workers. Experimen-tal Gerontology 40, 939–947.
Amdam, G.V., Hartfelder, K., Norberg, K., Hagen, A., Omholt, S.W., 2004. Alteredphysiology in worker honey bees (Hymenoptera: Apidae) infested with the miteVarroa destructor (Acari: Varroidae): a factor in colony loss duringoverwintering? Journal of Economic Entomology 97, 741–747.
Baer, B., Schmid-Hempel, P., 2006. Phenotypic variation in male and workerencapsulation response in the bumblebee Bombus terrestris. Ecological Ento-mology 31, 591–596.
Baer, B., Krug, A., Boomsma, J.J., Hughes, W.O.H., 2005. Examination of the immuneresponses of males and workers of the leaf-cutting ant Acromyrmex echinatiorand the effect of infection. Insectes Sociaux 52, 298–303.
Bailey, L., 1968. Honey bee pathology. Annual Review of Entomology 13, 191–212.Bailey, L., Ball, B.V., 1991. Honey Bee Pathology. Academic Press Limited, London.Barnes, A.I., Siva-Jothy, M.T., 2000. Density-dependent prophylaxis in the meal-
worm beetle Tenebrio molitor L. (Coleoptera: Tenebrionidae): cuticular melani-zation is an indicator of investment in immunity. Proceedings of the RoyalSociety of London, Series B 267, 177–182.
Bedick, J.C., Tunaz, H., Nor Aliza, A.R., Putnam, S.M., Ellis, M.D., Stanley, D.W., 2001.Eicosanoids act in nodulation reactions to bacterial infections in newly emergedadult honey bees Apis mellifera, but not in older foragers. Comparative Bio-chemistry and Physiology Part C 130, 107–117.
Bocher, A., Tirard, C., Doums, C., 2007. Phenotypic plasticity of immune defencelinked with foraging activity in the ant Cataglyphis velox. Journal of EvolutionaryBiology 20, 2228–2234.
Boman, H.G., Hultmark, D., 1987. Cell-free immunity in insects. Annual Review ofMicrobiology 41, 103–126.
Boomsma, J.J., Schmid-Hempel, P., Hughes, W.O.H., 2005. Life histories and parasitepressure across the major groups of social insects. In: Fellowes, M.D.E., Hol-loway, G.J., Rolff, J. (Eds.), Insect Evolutionary Ecology. CABI Publishing, Oxford,pp. 139–175.
Boulanger, N., Brun, R., Ehret-Sabatier, L., Kunz, C., Bulet, P., 2002. Immunopeptidesin the defense reactions of Glossina morsitans to bacterial and Trypanosomabrucei brucei infections. Insect Biochemistry and Molecular Biology 32, 369–375.
Boulanger, N., Ehret-Sabatier, L., Brun, R., Zachary, D., Bulet, P., Imler, J., 2001.Immune response of Drosophila melanogaster to infection with the flagellateparasite Crithidia spp. Insect Biochemistry and Molecular Biology 31, 129–137.

A.M. Laughton et al. / Journal of Insect Physiology 57 (2011) 1023–10321032
Cerenius, L., Lee, B.L., Soderhall, K., 2008. The proPO-system: pros and cons for itsrole in invertebrate immunity. Trends in Immunology 29, 263–271.
Chan, Q.W.T., Melathopoulos, A.P., Pernal, S.F., Foster, L.J., 2009. The innate immuneand systemic response in honey bees to a bacterial pathogen, Paenibacilluslarvae. BMC Genomics 10, 387.
Chan, Q.W.T., Howes, C.G., Foster, L.J., 2006. Quantitative comparison of castedifferences in honeybee hemolymph. Molecular and Cellular Proteomics 5,2252–2262.
Chmurzynska, W., Wojtczak, L., 1963. Effect of thiourea on moulting and pupationof the silkworm Bombyx mori L. Biological Bulletin 125, 61–68.
Chung, K.T., Ourth, D.D., 2000. Viresin. A novel antibacterial protein from immunehemolymph of Heliothis virescens pupae. European Journal of Biochemistry 267,677–683.
Cremer, S., Armitage, S.A., Schmid-Hempel, O.P., 2007. Social immunity. CurrentBiology 17, R693–R702.
DeVeale, B., Brummel, T., Seroude, L., 2004. Immunity and aging: the enemy within?Aging Cell 3, 195–208.
Doums, C., Moret, Y., Benelli, E., Schmid-Hempel, P., 2002. Senescence of immunedefence in Bombus workers. Ecological Entomology 27, 138–144.
Doums, C., Schmid-Hempel, P., 2000. Immunocompetence in workers of a socialinsect Bombus terrestris L., in relation to foraging activity and parasitic infection.Canadian Journal of Zoology 78, 1060–1066.
Dubovskiy, I.M., Krukova, N.A., Glupov, V.V., 2008. Phagocytic activity and encap-sulation rate of Galleria mellonella larval haemocytes during bacterial infectionby Bacillus thuringiensis. Journal of Invertebrate Pathology 98, 360–362.
Eleftherianos, I., Baldwin, H., ffrench-Constant, R.H., Reynolds, S.E., 2008. Develop-mental modulation of immunity: changes within the feeding period of the fifthlarval stage in the defence reactions of Manduca sexta to infection by Photo-rhabdus. Journal of Insect Physiology 54, 309–318.
Evans, J.D., 2004. Transcriptional immune responses by honey bee larvae duringinvasion by the bacterial pathogen, Paenibacillus larvae. Journal of InvertebratePathology 85, 105–111.
Evans, J.D., Lopez, D.L., 2004. Bacterial probiotics induce an immune response in thehoney bee (Hymenoptera: Apidae). Journal of Economical Entomology 97, 752–756.
Evans, J.D., Spivak, M., 2009. Socialized medicine: individual and communal diseasebarriers in honey bees. Journal of Invertebrate Pathology 103, S62–S72.
Falchuk, K.H., 1998. The molecular basis for the role of zinc in developmentalbiology. Molecular and Cellular Biochemistry 188, 41–48.
Freitak, D., Wheat, C., Heckel, C., Vogel, H., 2007. Immune system responses andfitness costs associated with consumption of bacteria in larvae of Trichoplusia ni.BMC Biology 5, 56.
Gerloff, C.U., Ottmer, B.K., Schmid-Hempel, P., 2003. Effects of inbreeding onimmune response and body size in a social insect, Bombus terrestris. FunctionalEcology 17, 582–589.
Gillespie, J.P., Kanost, M.R., Trenczek, T., 1997. Biological mediators of insectimmunity. Annual Review Entomology 42, 611–643.
Haine, E.R., Pollitt, L.C., Moret, Y., Siva-Jothy, M., Rolff, T.J., 2008. Temporal patternsin immune responses to a range of microbial insults (Tenebrio molitor). Journalof Insect Physiology 54, 1090–1097.
Hillyer, J.F., Schmidt, S.L., Fuchs, J.F., Boyle, J.P., Christensen, B.M., 2005. Age-associated mortality in immune challenged mosquitoes (Aedes aegypti) corre-lates with a decrease in haemocyte numbers. Cellular Microbiology 7, 39–51.
Hooper, T., 1997. Guide to Bees and Honey. Marston House, Yeovil.Hopkins, T.L., Kramer, K.J., 1992. Insect cuticle sclerotization. Annual Reviews in
Entomology 37, 273–302.Horowitz, N.H., Shen, S., 1952. Neurospora tyrosinase. Journal of Biological Chem-
istry 197, 513–520.Hultmark, D., 1996. Insect lysozymes. In: Jolles, P. (Ed.), Lysozymes: Model Enzymes
in Biochemistry and Biology. Birkhauser Verlag, Basel, Switzerland, pp. 87–102.Kaaya, G.P., Darji, N., 1988. The humoral defense system in tsetse: differences in
response due to age, sex and antigen types. Developmental & ComparativeImmunology 12, 255–268.
Konig, C., Schmid-Hempel, P., 1995. Foraging activity and immunocompetence inworkers of the bumble bee, Bombus terrestris L. Proceedings of the Royal Societyof London, Series B 260, 225–227.
Kopacek, P., Weise, C., Gotz, P., 1995. The prophenoloxidase from the wax mothGalleria mellonella: purification and characterization of the proenzyme. InsectBiochemistry and Molecular Biology 25, 1081–1091.
Laughton, A.M., Siva-Jothy, M.T., 2010. A standardised protocol for measuringphenoloxidase and prophenoloxidase in the honey bee Apis mellifera. Apidolo-gie, doi:10.1051/apido/2010046.
Lavine, M.D., Strand, M.R., 2002. Insect hemocytes and their role in immunity. InsectBiochemistry and Molecular Biology 32, 1295–1309.
Lehrer, R.I., Ganz, T., 1999. Antimicrobial peptides in mammalian and insect hostdefence. Current Opinion in Immunology 11, 23–27.
Lourenco, A.P., Zufelato, M.S., Bitondi, M.M.G., Simoes, Z.L.P., 2005. Molecularcharacterization of a cDNA encoding prophenoloxidase and its expression inApis mellifera. Insect Biochemistry and Molecular Biology 35, 541–552.
Mason, H.S., 1955. Comparative biochemistry of the phenolase complex. Advancesin Enzymology 16, 105–184.
Meylaers, K., Freitak, D., Schoofs, L., 2007. Immunocompetence of Galleria. mello-nella: sex- and stage-specific differences and the physiological cost of mountingan immune response during metamorphosis. Journal of Insect Physiology 53,146–156.
Michelette, E.R., Soares, A.E.E., 1993. Characterization of preimaginal developmentstages in Africanized honeybee workers (Apis mellifera L.). Apidologie 24, 431–440.
Moret, Y., Schmid-Hempel, P., 2009. Immune responses of bumblebee workers as afunction of individual and colony age: senescence versus plastic adjustment ofthe immune function. Oikos 118, 371–378.
Moret, Y., Schmid-Hempel, P., 2000. Survival for immunity: the price of immunesystem activation for bumblebee workers. Science 290, 1166–1168.
Natori, S., Shiraishi, H., Hori, S., Kobayashi, A., 1999. The roles of Sarcophaga defensemolecules in immunity and metamorphosis. Developmental and ComparativeImmunology 23, 317–328.
Navajas, M., Migeon, A., Alaux, C., Martin-Magniette, M.L., Robinson, G.E., Evans, J.D.,Cros-Arteil, S., Crauser, D., Le Conte, Y., 2008. Differential gene expression of thehoney bee Apis mellifera associated with Varroa destructor infection. BMCGenomics 9, 301.
Oldroyd, B.P., 2007. What’s killing American honey bees? PLoS Biology 5, e168.Omholt, S.W., Amdam, G.V., 2004. Epigenetic regulation of aging in honeybee
workers. Science of Aging Knowledge Environment 2004, 28.Page, R.E., Peng, C.Y.S., 2001. Aging and development in social insects with emphasis
on the honey bee, Apis mellifera L. Experimental Gerontology 36, 695–711.Ratcliffe, N.A., Leonard, C., Rowley, A.F., 1984. Prophenoloxidase activation: nonself
recognition and cell cooperation in insect immunity. Science 226, 55–559.Robinson, G.E., 1992. Regulation of division of labor in insect societies. Annual
Review Entomology 37, 637–665.Robinson, G.E., 1987. Regulation of honey bee age polyethism by juvenile hormone.
Behavioural Ecology and Sociobiology 20, 329–338.Rolff, J., 2002. Bateman’s principle and immunity. Proceedings of the Royal Society
of London, Series B 269, 867–872.Rueppell, O., Fondrk, M.K., Page Jr., R.E., 2004. Biodemographic analysis of male
honey bee mortality. Aging Cell 4, 13–19.Ruiz-Gonzalez, M.X., Brown, M.J.F., 2006. Males vs workers: testing the assumptions
of the haploid susceptibility hypothesis in bumblebees. Behavioural Ecologyand Sociobiology 60, 501–509.
Saul, S.J., Sugumaran, M., 1988. Prophenoloxidase activation in the hemolymph ofSarcophaga bullata larvae. Archives of Insect Biochemistry and Physiology 7, 91–103.
Schmid, M.R., Brockmann, A., Pirk, C.W.W., Stanley, D.W., Tautz, J., 2008. Adulthoneybees (Apis mellifera L.) abandon hemocytic but not phenoloxidase-basedimmunity. Journal of Insect Physiology 54, 439–444.
Schmid-Hempel, P., 2005. Evolutionary ecology of insect immune defenses. AnnualReview of Entomology 50, 529–551.
Schmid-Hempel, P., 1998. Parasites in Social Insects. Princeton University Press,Chichester, West Sussex.
Simmons, L.W., Roberts, B., 2005. Bacterial immunity traded for sperm viability inmale crickets. Science 309, 2031.
Siva-Jothy, M.T., Moret, Y., Rolff, J., 2005. Insect immunity: an evolutionary ecologyperspective. Advances in Insect Physiology 32, 1–48.
Thompson, J.J.W., 2002. Aspects of Melanin Production and Immunity in theMealworm Beetle Tenebrio molitor. MPhil., Department of Animal and PlantSciences, University of Sheffield.
Tzou, P., Ohresser, S., Ferrandon, D., Capovilla, M., Reichhart, J., Lemaitre, B., Hoff-man, J., Imler, J., 2000. Tissue-specific inducible expression of antimicrobialpeptide genes in Drosophila surface epithelia. Immunity 13, 737–748.
Vainio, L., Hakkarainen, H., Rantala, M.J., Sorvari, J., 2004. Individual variation inimmune function in the ant Formica exsecta; effects of the nest, body size andsex. Evolutionary Ecology 18, 75–84.
Wilson, E.O., 1971. The Insect Societies. Oxford University Press, London.Wilson-Rich, N., Dres, S.T., Starks, P.T., 2008. The ontogeny of immunity: develop-
ment of innate immune strength in the honey bee (Apis mellifera). Journal ofInsect Physiology 54, 1392–1399.
Zerofsky, M., Harel, E., Silverman, N., Tatar, M., 2005. Aging of the innate immuneresponse in Drosophila melanogaster. Aging Cell 4, 103–108.
Zufelato, M.S., Lourenco, A.P., Simoes, Z.L.P., Jorge, J.A., Bitondi, M.M.G., 2004.Phenoloxidase activity in Apis mellifera honey bee pupae, and ecdysteroid-dependent expression of the prophenoloxidase mRNA. Insect Biochemistry andMolecular Biology 34, 1257–1268.
Zuk, M., 2009. The sicker sex. PLoS Pathogens 5, e1000267.




















