
Division of labor by division of risk according to worker life expectancy in the honey bee (Apis...
15
Original article Division of labor by division of risk according to worker life expectancy in the honey bee (Apis mellifera L.) Micha&jadnr; Woyciechowski’ Jan Koz&jadnr;owski b a Bee Research Department, Agricultural University, Al. 29 Listopada 52, 31-425 Krakow, Poland b Institute of Environmental Biology, Jagiellonian University, Oleandry 2a, 30-060 Krakow, Poland (Received 15 July 1997; accepted 24 November 1997) Abstract - The hypothesis that the division of labor in foraging honey bee workers is a conse- quence of division of risk among foragers with differing life expectancies was tested using a dynamic programming model and field experiment. Honey bee workers collecting water were cap- tured in the middle of one cold day and one warm day in April, May and July. We investigated whether workers infected with Nosema apis or workers with worn wings undertake foraging in poorer weather conditions more often than do healthy workers or workers with unworn wings. On each pair of days, the ratio of diseased workers and workers with worn wings was usually sig- nificantly higher on the cold day than on the warm day. The result that risky tasks are more often undertaken by workers with shorter life expectancy is in favor of the predictions of the model. © Inra/DIB/AGIB/Elsevier, Paris division of labor / division of risk / life expectancy / Apis mellifera / Nosema apis 1. INTRODUCTION Worker fitness depends on the repro- ductive success of the whole colony of related individuals, not only in the honey bee but also in other eusocial haplodiploids (Hamilton, 1964). Colony reproduction depends, however, on the total of nonre- productive labor carried out by workers. This demands that colony members under- take many tasks not associated with their direct reproduction, despite conflicting * Correspondence and reprints Tel.: (48) 12 411 9144 ext. 342; fax: (48) 12 633 6245; e-mail: [email protected]
-
Upload
jagiellonian -
Category
Documents
-
view
4 -
download
0
Transcript of Division of labor by division of risk according to worker life expectancy in the honey bee (Apis...

Original article
Division of labor by division of riskaccording to worker life expectancyin the honey bee (Apis mellifera L.)
Micha&jadnr; Woyciechowski’ Jan Koz&jadnr;owskib
aBee Research Department, Agricultural University,Al. 29 Listopada 52, 31-425 Krakow, PolandbInstitute of Environmental Biology, Jagiellonian University,
Oleandry 2a, 30-060 Krakow, Poland
(Received 15 July 1997; accepted 24 November 1997)
Abstract - The hypothesis that the division of labor in foraging honey bee workers is a conse-quence of division of risk among foragers with differing life expectancies was tested using adynamic programming model and field experiment. Honey bee workers collecting water were cap-tured in the middle of one cold day and one warm day in April, May and July. We investigatedwhether workers infected with Nosema apis or workers with worn wings undertake foraging inpoorer weather conditions more often than do healthy workers or workers with unworn wings. Oneach pair of days, the ratio of diseased workers and workers with worn wings was usually sig-nificantly higher on the cold day than on the warm day. The result that risky tasks are moreoften undertaken by workers with shorter life expectancy is in favor of the predictions of themodel. © Inra/DIB/AGIB/Elsevier, Paris
division of labor / division of risk / life expectancy / Apis mellifera / Nosema apis
1. INTRODUCTION
Worker fitness depends on the repro-ductive success of the whole colony ofrelated individuals, not only in the honeybee but also in other eusocial haplodiploids
(Hamilton, 1964). Colony reproductiondepends, however, on the total of nonre-productive labor carried out by workers.This demands that colony members under-take many tasks not associated with theirdirect reproduction, despite conflicting
* Correspondence and reprintsTel.: (48) 12 411 9144 ext. 342; fax: (48) 12 633 6245; e-mail: [email protected]

evolutionary interests between differentcastes and even particular nest members,manifested, for example, by worker-queenconflict over the sex ratio (Trivers andHare, 1976; Sunström et al., 1996) orworker policing of the production of maleeggs (Woyciechowski and Lomnicki,1987; Ratnieks and Visscher, 1989).
There are suggestions that the divisionof nonreproductive labor influences colonysurvival and reproduction (Oster and Wil-son, 1978; Robinson, 1992; Gordon,1996). However, the decision on whichindividual in a colony undertakes whichtask necessitates a compromise amongworkers, as different tasks entail differentmortality risks. Even the risk of the sametask can change depending on weatherconditions. External factors can lead tomuch higher mortality rates for tasks awayfrom the nest than for tasks in the nest
(Sakagami and Fukuda, 1968; Schmid-Hempel and Schmid-Hempel, 1984;Dukas and Visscher, 1994; but see alsoO’Donnell, 1996) and for tasks undertakenby foragers on a cold day as compared toa warm one. How workers reach this com-
promise in the division of nonreproduc-tive labor is one of the most intriguingproblems concerning insect societies.
Internal factors such as age, or more
specifically life expectancy, can explainthe compromise of workers in taking dif-ferent risks. The fact that life expectancydifferentiates workers results directly fromthe fact that in all eusocial colonies olderindividuals rear younger siblings.Although many authors have pointed outthat the organization of work in the colonyis influenced by internal and external fac-tors [for a review see Robinson (1992);Gordon (1996)], only Jeanne (1986) andO’Donnell and Jeanne (1995a) showedthat different levels of task risk and pro-grammed senescence of workers createsufficient conditions for the evolution of adivision of labor based on temporalpolyetism. This is because task sequencing
to avoid risky tasks at the beginning oflife and to undertake risky tasks in old ageprolongs the mean longevity of workers.On the other hand, it is well known thatprolonging worker life increases the repro-ductive success of the colony (Schmid-Hempel, 1987; Schmid-Hempel and Wolf,1988).
The hypothesis that the division of laborin foraging workers is a consequence ofthe division of risk among foragers withdifferent life expectancies was tested usinga dynamic programming model and con-firmed experimentally. The model pre-dicts that honey bee workers vary theirforaging behavior with age or health status,foraging mainly under safe conditionswhen young or healthy and accepting riskyforaging when old or sick. In the fieldexperiment, workers collecting water onwarm and cold days were examined. Theirlife expectancies were scored on the basisof wing wear and the presence of theNosema apis Zander (Protista) parasite inthe intestine.
2. THE MODEL
Let us consider bee workers choosingeach day whether to stay in the nest (strat-egy j = 0) or to forage (strategy j = 1),depending on the weather. Weather is clas-sified according to the risk of foraging init, with the lowest risk for class i = 1 andthe highest risk for i = 160 (the numberof classes is arbitrary and adjusted to com-puter capacity). The probability of deathduring foraging one day under weather i ispi, and the probability of weather i is ci.Daily foraging yield is yij and alwaysequals zero for j = 0, that is, for the work-ers staying in the nest.
Maximum worker lifespan is T, and itdepends on health status. Physiologicalmortality (not dependent on foraging deci-sions) is μ(t), where t is age in days. Onlyworkers older than 19 days are consid-

ered, because younger bees work in thenest. This cutoff age is close to the real
value (Seeley, 1982).
The goal is to find the optimal strategy(stay in the nest or forage) for each age t,each weather class i and each health status
(healthy or sick) to maximize the expectedtotal yield toward the end of life T. This isa dynamic optimization problem, whichwill be solved here with dynamic pro-gramming (Bellman, 1957; Mangel andClark, 1988). The discrete time unit is 1
day. Bellman’s algorithm works back-ward. Let us consider first the last day T.It is always optimal to forage on this day,irrespective of the risk, because no morelife is left. Thus the expected yield on thisday is
for all kinds of weather i. The decisionwhether to forage or not on day T - 1
depends not only on the day’s weather butalso on the residual yield of the last daywhich will be lost if the worker dies.Because the next day’s weather is
unknown, the residual yield for the lastday under each weather multiplied by theprobability that this weather will occurmust be summed up across all weatherclasses k. Moreover, the expected resid-ual yield must be multiplied by the age-dependent probability of surviving to ageT - 1
for staying in the nest (j = 0), and for for-aging (j = 1) also multiplied by the prob-ability pi of dying in weather i. Thus
for j = 0, that is, for staying in the nest,and
for j = 1, that is, for foraging. Current yieldis added only for j = 1, because yi0 equalszero. Now the value of equation (3) mustbe compared against equation (4), and thestrategy giving higher F(i,T - 1) is cho-sen. The procedure is repeated for all i,and optimal strategies and correspondingtotal yields are saved for further use.
Exactly the same procedure is repeatedfor T - 2, yielding the expressions
For any t < T, this recursive procedureis represented by
where max means that the strategy j whichmaximizes the functional (5) should bechosen. Solving this problem backwardto age 20 days gives the best strategy forany foraging risk at each age. In otherwords, for each age we find the level ofrisk taking that maximizes the expectedyield of the whole life.
To illustrate the problem with a numer-ical example, we will assume that maxi-mum longevity T equals 100 days forhealthy workers and 80 for infected ones(referred to as sick hereafter). We alsoassume that physiological mortality is

Mortality approaches infinity when ttends toward T, and parameter q = 0.002defines how early in life aging has a pro-nounced effect on mortality. We alsoassume that the probability of survivingforaging is constant at 0.98 for the first 50weather classes, then decreases with i lin-early to 0.036 for i = 160. We assume thatall weather classes are equally likely, andthat yield does not depend on weather.These assumptions are not very realistic,but we shall show that they do not affect,or very weakly affect, the results we areinterested in: the proportions of old or sickworkers under good and poor weather.The last assumption (yield independent ofweather) was used because water collec-tion was studied here; for nectar or pollencollection, decreased yield on days withrisky foraging (wind, rain, cold) would bemore realistic.
The probability of surviving to a givenage (lt) under physiological mortality forhealthy and sick workers is representedby the thin lines in figure 1. Risk taking forworkers depends on health status as shownin figure 2. The lines on this graph repre-sent the maximum tolerable risk of forag-ing for healthy and sick workers at a givenage, which means that they forage at a risklower or equal to the value at the line andstay in the nest if the risk is higher. Allworkers older than 19 days (all foragersin the model) should leave the nest whenthe daily risk of foraging is equal to orlower than 0.05, although sick workersalso forage at a 0.06 probability of death.Higher risk is taken differentially, depend-ing on health status. For example, at a riskof 0.07, healthy workers younger than 43days should stay in the nest, and sickworkers should stay if younger than 26
days. At a risk of 0.13, healthy workersshould forage if older than 78 days, andsick ones if older than 60 days.
We are interested not only in age- andhealth-dependent risk taking, but also inthe proportions of healthy and sick work-ers foraging under different weather con-ditions. These proportions can be used totest the hypothesis that workers adjust theirdecisions optimally.
Because we know maximal risk taking(figure 2), we can calculate the averageprobability of surviving to a given age (lt)for workers, taking into account both phys-iological mortality and mortality from for-aging (thick lines in figure 1; we mustremember, however, that short-termweather anomalies may change lt). After19 days, when workers start to forage, thethick lines go down much faster than thethin lines do, and converge at the end oflife.
The probability of surviving to a givenage for healthy workers is repeated in fig-ure 3A. This probability also means ’theexpected number of workers at a givenage’ assuming a stable age structure and astable total number of workers. The valueson this graph are standardized so that thenumber of hatched individuals equals one(t = 1). To obtain the number of workers20 or more days old we have to integratelt from 19 (because age 19 denotes the endof the 19th day) to infinity, and the integralis represented by the sum of the twohatched areas under this graph. The secondhatched area, starting at age 42, representsthe number of workers 43 days old andolder. We can calculate such areas for anyage. Their values are represented in fig-ure 3B. Using this graph together with theinformation about optimal risk taking rep-resented in figure 2, we can predict thenumber of healthy and sick bees foragingat a given risk. These numbers are markedwith squares for good weather when allworkers older than 19 days forage (riskr ≤ 0.05), triangles for intermediateweather (r = 0.07), and asterisks for harshweather (r = 0.13). Assuming a stable agedistribution, a stable total worker number,

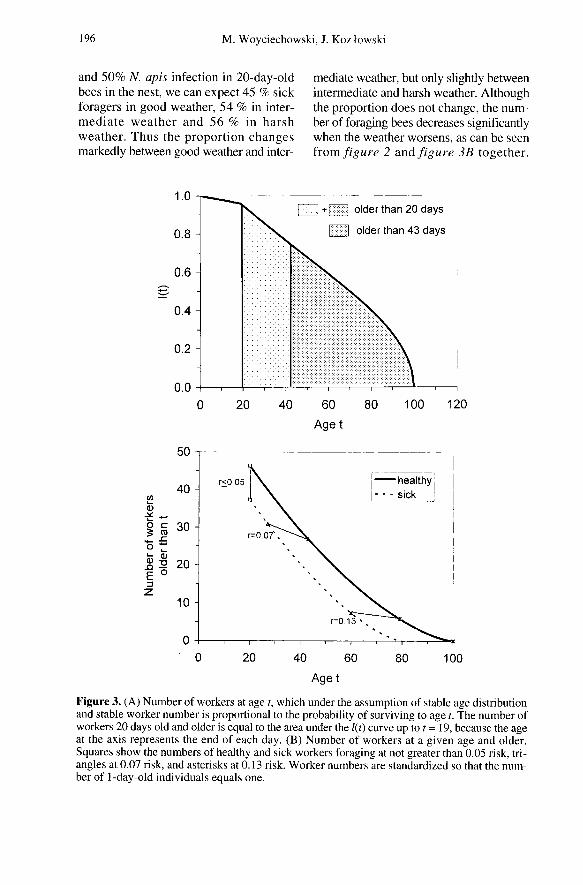
and 50% N. apis infection in 20-day-oldbees in the nest, we can expect 45 % sickforagers in good weather, 54 % in inter-mediate weather and 56 % in harshweather. Thus the proportion changesmarkedly between good weather and inter-
mediate weather, but only slightly betweenintermediate and harsh weather. Althoughthe proportion does not change, the num-ber of foraging bees decreases significantlywhen the weather worsens, as can be seen
from figure 2 and figure 3B together.

Although failure to satisfy the assump-tions of a stable age distribution and an
equal number of sick and healthy beesearly in life may change the proportions,we can still expect a greater percentageof sick workers under poor than under
good weather conditions.To test the robustness of the model, we
changed different parameters. After dou-bling or halving parameter q in the equa-tion for physiological age-dependent mor-tality [equation (6)], the fraction of sickforagers in good weather was still44-45 %, and 54 and 59 % under inter-mediate and harsh weather. Similarly,increasing the mortality risk in goodweather to 0.05 per day had only a smalleffect on the proportions (46 % for goodweather, 59 and 60 % for intermediate andharsh weather). Other changes in the dis-tribution of risk also had very weak effects.One parameter, however, was very impor-tant to the expected proportion of sick for-agers in good weather: the differencebetween the maximum lifespans of sickand healthy workers. Decreasing thisparameter for sick workers from 80 to 70days changed the proportion from 45 to41 %, and a further decrease to 60 daysreduced it to 36 %; increasing maximumlifespan to 90 days yielded 48 % sick for-agers in good weather. Interestingly, theproportion of sick foragers in bad weatheris still in a narrow range: 54-58 %. It
seems, therefore, that the proportion ofsick foragers is much more sensitive toshortening lifespan in good weather thanit is in bad weather. Thus, the qualitativeresult that the proportion of sick foragersshould become much higher in poor thanin good weather seems robust.
3. MATERIAL AND METHODSFOR FIELD EXPERIMENT
Honey bee workers collecting water werecaptured from a watering trough (not more than50 m from the farthest hive) in an apiary of
about 60 hives near Krakow (southern Poland)in spring and summer 1995, on three pairs ofcold-and-cloudy and warm-and-sunny days,between I 1 a.m. and 2 p.m. During the cap-ture of bees on cold days the maximum meantemperature was 15.7 °C and there was a strongwind. On the warm days the minimum meantemperature was 20.4 °C and there was nowind. The time gap between the days in eachpair was not longer than 3 days. The foragingactivity of workers on the pairs of compareddays differed significantly, being significantlylower on the cold days. This could be easilyobserved because of the small number of work-ers that left the hives and also because the num-ber of bees gathering water from the wateringtrough was several times lower on the colddays than on the warm days.
The presence of the parasite N. apis sporewas checked with a hematocytometer in1/40 mm3 samples of individual workerabdomens macerated in I mL distilled water. Ifthere were spores in a sample the individualwas classified as infected; if no spores werefound it was classified as uninfected.
The wear of each of the front wings of thecaptured workers was evaluated using a scalefrom 0 to 3. Zero meant that the outer wingmargin was not damaged; 1 meant that theouter margin was slightly damaged; 2 meantthat the wing was damaged one third of thedistance to the ends of the medial veins (M1,M2); 3 indicated damage two thirds of the dis-tance to the ends of the medial veins, or far-ther. The sum of two values, for the two frontwings, characterized each worker. If this sumwas ≤ 3 it was assumed that the individual’s
wings were in good condition; a sum > 3 wastaken to mean worn wings.
The proportions of uninfected/infected indi-viduals and also of individuals with wings ingood/worn condition were compared for eachpair of cold and warm days. For all individuals,the proportions of workers with good/wornwings in the infected and uninfected groupswere compared. For this analysis the G-testwith df = 1 and William’s correction wereused (Sokal and Rohlf, 1981).
4. EXPERIMENTAL RESULTS
Individuals infected with N. apis sporeswere significantly over-represented on

cold days in April (G = 8.903, P = 0.0028)and June (G = 10.156, P = 0.0014), whenthe total proportions of infected workerswere high on cold as well as on warm days(figure 4). The overall infection level waslow in July, as is usual in summer (Fur-gala and Mussen, 1978; Pickard and El-Shemy, 1989), which made the samplesize of sick workers too small (fiveinfected workers on cold and three onwarm days) to obtain a significant over-
representation of infected bees on the coldday (G = 0.193, P = 0.6608).
Comparing the wings of the sameworkers on each pair of days (figure 5),we found that there were significantlymore workers with worn wings collectingwater on the cold day in April (G = 4.187,P = 0.0407) and July (G = 5.014, P =0.0251). Only in June, when the proportionof workers with worn wings was very low,

over-representation of workers with badlyworn wings was not found (G = 0.887,P = 0.3462; two and four workers withworn wings on cold and warm days).
Analyzing the health status and wingcondition of the workers, we observed thatinfected individuals more often had worn
wings in each of the three pairs of days.Totaling the data for all workers capturedduring the experiment, worn wings werefound almost twice as often amonginfected (29.3 %) than among uninfected(15.8 %) workers. This difference was sig-nificant (G = 4.844, N = 189, P = 0.0277).
5. DISCUSSION
Most of the model results seem to con-form to intuition, our field experiment andmany well-known results of early obser-vations. It is obvious that the probability ofworker survival must drop abruptly whenforaging starts (figure 1). High mortality inworkers undertaking tasks outside the nestwas observed by Sakagami and Fukuda(1968) and recently by Visscher andDukas (1997). It is also patently true thatthe risk connected with foraging affectsboth healthy and sick foragers. The modelalso predicts that, independent of age andhealth status, all foragers (in the model,workers older than 19 days) are ready toundertake tasks if their risks are relativelylow (lower than 0.05 for numerical exam-ple; figure 2). Depending on the expectedlife span corresponding to health statusand age, foragers can maximize theexpected total life foraging yield by under-taking different maximal risks of forag-ing (figure 2). Thus, sick workers under-take higher risks earlier in life than healthyworkers do. This result is consistent withO’Donnell and Jeanne’s (1995a) predic-tions. Not so obvious is the result that evenat the beginning of the workers’ foraginglife the difference in the ages of sick and
healthy foragers undertaking the same risk
is only slightly less than the differencebetween the maximal longevities ofhealthy and sick workers (figure 2).
Not only physiological mortality, dif-ferent for healthy and sick workers, butalso the level of risk taking, dependent onlife expectancy, affects the age structureand health statistics among foragers in thenest. All foragers forage when the risk ofit is very low (r ≤ 0.05; figure 3B). Themodel predicts, however, that some of theforagers will remain in the nest while oth-ers leave it when the risk of foragingincreases. Such risky foraging (r = 0.07or r = 0.13) is not undertaken by theyoungest and the healthy workers (figure3B). This is why sick and old workers areover-represented among foragers com-pared to the ratio under safe conditions.
Our field experiment confirms themodel predictions of the age structure andhealth of foragers. Among water collectingworkers, the proportions of both diseasedindividuals (figure 4) and workers withsignificantly damaged wings (figure 5)show an over-representation of sick andold workers (presumably with shorter lifeexpectancy) during risky weather condi-tions compared to their proportions undersafe conditions. This does not mean thatthese individuals prefer to work undermore risky conditions, but only that theydecide to undertake more risky tasks thanworkers with longer life expectancies do.
The confirmation of the model predic-tion by field observations can be acceptedif certain assumptions are satisfied. Neitherof the two parameters, that is, the pres-ence of the parasite N. apis and wing wear,were expected to determine the exact ageof the tested honey bee workers. Each ofthese parameters was used only to dividethe workers into two groups differing inlife expectancy, following earlier obser-vations that both sickness and wing con-dition are good predictors of life
expectancy. Lifetime is significantlyreduced in honey bees infected with N.

apis (Wang and Moller, 1969, 1970; Mof-fet and Lawson, 1975). The relationshipbetween age and infection level has beendemonstrated (El-Shemy and Pickard,1989); it seems to be a general rule inworkers infected with other parasites also(Schmid-Hempel and Schmid-Hempel,1993). Moreover, it has been proved thatage and duration of foraging affect workerwing wear not only in honey bees (Viss-cher and Dukas, 1997) but also in bum-ble bees (Alford, 1975) and solitary bees(Michener, 1969). In our experiment therelation between the presence of parasitesand wing wear confirms the correctnessof the choice of these parameters, which
probably both correlate with worker lifeexpectancy.
There are no exact data on how senes-cence influences mortality in workers withlimited maximum longevity (figure 1). Anexperiment with a small sample of work-ers has been interpreted to mean that long-living foragers undergo senescence (Dukasand Visscher, 1994) and that their proba-bility of death is not age-dependent (Viss-cher and Dukas, 1997). However, lifetables of honey bees (Sakagami andFukuda, 1968), and other work [for areview see O’Donnell and Jeanne (1995a)]support our assumption that senescenceinfluences mortality.
Survival curves represented in figure 1may look not very realistic: they are notsmooth at the beginning of foraging (20days), and they go abruptly down at theend of life. Real survival curves for bee
workers, although concave downward forsome time, have a concave up ’tail’ at theend (Sakagami and Fukuda, 1968; Viss-cher and Dukas, 1997). We must remem-ber, however, that our curves are not con-ventional curves representing probabilitiesof survival of an average individual. Theyrepresent survival curves for two ’clean’strategies, healthy and sick, and when for-aging starts exactly at the 20th day ifweather is not too harsh. If foraging of
different workers starts at a different time,for example between 20th and 40th day,the survival curve becomes smooth at the
beginning. Moreover, if health status istreated as a continuous variable, whichmeans in the model that lifespan can takeany value between, say, 60 and 100 days,the concave upward ’tail’ appears towardthe end of life (Woyciechowski andKoz&jadnr;owski, unpublished data). This isbecause workers in the poorest state dieearlier and those in a better state survive
longer. A similar phenomenon has beendescribed in a nematode Caenorhabdities
elegans for which, unlike the honey bee,genes for senescence are well known. Inthe mixture of genotypes, the survivalcurve is strongly elongated, whereas instrains developed from either short- orlong-lived individuals the curves end moreabruptly (Brooks et al., 1994). A closelook at the Sakagami and Fukuda (1968)paper supports our view: mortality ratesincrease rapidly toward the end of life upto some age, then remain constant or evendecrease slightly, and further increase at aslower rate. It is difficult to explain thisresult on a physiological ground, but suchan effect is straightforward if the data rep-resent a mixture of workers with differ-ent genotypes and different health status.Milne (1980) and Guzmán-Novoa andGary (1993) demonstrated probablegenetic variability for longevity in workerhoney bees.
We assumed that weather conditionsdetermine honey bee workers’ foragingdecisions and we ignored the potential roleof the distance to water sources. This dis-tance was relatively short in our experi-ment. Gathering water probably has beena greater risk on cold days throughouthoney bee evolution; workers returningto the hive with water are more likely todie of starvation than those engaged ingathering nectar (Visscher et al., 1996).By testing workers collecting only water atabout the same distance from their hives,

we eliminated other factors connected withdistance to resources in the worker life
strategy (Schmid-Hempel, 1987) or totheir genetic predisposition to undertakeone task (Robinson and Page, 1988).
Our field experiment results can beinterpreted in several different ways. It ispossible that on cold days less water isneeded and therefore new (young) beesare not recruited to collect it. It can also be
suggested that only more experiencedworkers undertake foraging during coldweather conditions. These alternative
explanations should be studied. In anycase both the model predictions and thefield results are in agreement with the con-tention that worker task sequencing basedon temporal polyethism is a result of thevarying risk levels of the tasks to be under-taken. It is generally known that safe workin the nest precedes dangerous foragingin all eusocial insect groups that do nothave polymorphic workers [ants: for areview see Hölldobler and Wilson (1990);bees: Seeley and Kolmes (1991); wasps:Jeanne (1991), but see O’Donnell (1996);termites: Gerber et al. (1988)]. This basicdivision of tasks is probably the main rea-son for (O’Donnell and Jeanne, 1995b)and not a result of (Fénéron et al., 1996)morphological and physiological changesduring the ontogenesis of adult individu-als. As a consequence of these changes,sequences of safe duties inside the hiveare most likely driven by physiology. Thisis why the division of these risk-indepen-dent tasks is so difficult to distinguish(Seeley, 1982) that its existence is evenquestioned (Kolmes, 1986).
The increase of outside duties observedin workers infected with N. apis (Has-sanein, 1953; Wang and Moeller, 1970)is not caused directly by the parasite. Nordoes it have to be an adaptation by whichdiseased workers avoid tasks whichincrease the chance of parasite spreadamong colony members as suggested byWang and Moeller (1970) and Schmid-
Hempel (1995). More likely, sick work-ers undertake risky duties at an early agebecause of their shortened life expectancy.Consequently, the same strategy shouldbe anticipated not only in sick individu-als but in all workers receiving cues ofshortened life expectancy.
Although it is known that safe tasks inthe nest usually are undertaken prior torisky tasks outside the nest in many insectsocieties, there are only preliminary resultson the age dependence of foraging taskssequenced according to risk (Woy-ciechowski and Lomnicki 1995; Woy-ciechowski et al., 1996). So far as weknow, our findings are the first to be infavor of the hypothesis that in the honeybee and possibly also in other eusocialinsects the division of labor results fromthe division of risks among workers withdifferent life expectancies.
ACKNOWLEDGMENT
We thank Gra¿yna Grzanka for valuableassistance in the field and laboratory. Thanks toStefan Fuchs, Michael Jacobs, Eduard Scher-tiger, Ewa Woyciechowska and anonymousreviewers for their helpful comments on earlierversions of this paper.
Résumé - La division du travail par la
répartition des risques en fonction del’espérance de vie des ouvrières chezl’abeille (Apis mellifera L.). Chez l’abeillemellifère, ainsi que chez d’autres Hymé-noptères eusociaux, la reproduction de lacolonie dépend en totalité du travail nondestiné à la reproduction qu’accomplis-sent les ouvrières. Cela implique que lesmembres de la colonie entreprennent denombreuses tâches qui ne sont pas en rela-tion directe avec leur reproduction. Pour-tant décider quel individu dans une colo-nie va accomplir quelle tâche nécessite uncompromis entre les ouvrières, car lesdiverses tâches occasionnent des risques

de mortalité différents. C’est une règleque les facteurs externes conduisent à destaux de mortalité beaucoup plus élevéspour les tâches accomplies hors du nidque pour celles accomplies dans le nid.Le risque d’une tâche extérieure donnéepeut varier lui-même selon les conditionsmétéorologiques.
L’hypothèse, selon laquelle la divisiondu travail chez les ouvrières qui butinentest une conséquence de la répartition durisque parmi les butineuses ayant des espé-rances de vies différentes, a été testée àl’aide d’un programme de modélisation
dynamique et confirmée expérimentale-ment. Dans le modèle seules les ouvrièresde plus de 19 j ont été prises en compte,car nous estimons que les ouvrières plusjeunes travaillent dans le nid. Nous esti-mons aussi que le vieillissement influencela mortalité des ouvrières et délimite une
longévité maximale qui est de 100 j pourles ouvrières en bonne santé et de 80 jpour les ouvrières infectées (figure 1).
Le modèle prédit que, indépendammentde l’âge et de l’état de santé, toutes lesbutineuses sont prêtes à entreprendre destâches si les risques sont relativementfaibles (figure 2). En fonction de l’espé-rance de vie correspondant à l’état de santéet à l’âge, les butineuses peuvent maxi-miser la production du butinage surl’ensemble de leur espérance de vie enprenant des risques maximaux plus oumoins élevés au cours du butinage(figure 2). Les ouvrières modifient ainsileur comportement de butinage avec l’âgeet l’état de santé, butinant surtout dans desconditions sûres lorsqu’elles sont jeunes ouen bonne santé et acceptant un butinageplus risqué lorsqu’elles sont vieilles oumalades. C’est pourquoi les ouvrièresmalades et vieilles sont sur-représentéesparmi les butineuses lors de conditionsmétéorologiques risquées en comparaisonavec des conditions météorologiques sûres(figure 3B).
Dans l’expérience en champ, lesouvrières récoltant de l’eau ont été captu-rées à un abreuvoir dans un rucher d’envi-ron 60 ruches, au printemps et en été, aucours de trois jours froids et nuageux etde trois jours chauds et ensoleillés. Leurespérance de vie a été évaluée d’aprèsl’usure des ailes et la présence du parasiteNosema apis dans les intestins. Les résul-tats confirment les prédictions du modèlequant à la structure de l’âge et la santé desbutineuses. Parmi les récolteuses d’eau,la proportion à la fois d’individus maladeset d’ouvrières avec des ailes significati-vement endommagées (figure 5) est sur-représentée dans les conditions météoro-logiques à risques en comparaison avecdes conditions météorologiques sûres.
Il est bien connu que, dans de nom-breuses sociétés, les tâches sans dangerdans le nid sont généralement accompliesavant les tâches à risques hors du nid, maisnos résultats sont les premiers, à notreconnnaissance, à confirmer l’hypothèseselon laquelle, chez les abeilles et peut-être aussi chez les autres insectes sociaux,la division du travail pourrait découler dela répartition des risques parmi lesouvrières ayant différentes espérances devie. © Inra/DIB/AGIB/Elsevier, Paris
Apis mellifera / division du travail /répartition risques / espérance de vie /Nosema apis
Zusammenfassung - Arbeitsteilungdurch eine der Lebenserwartung ent-sprechende Risikoverteilung bei Honig-bienen (Apis mellifera L.). Bei Honig-bienen, aber auch bei anderen eusozialenHymenopteren hängt die Vermehrung vonKolonien von der insgesamt durch dieArbeiterinnen verrichteten nichtrepro-duktiven Arbeit ab. Dies bedingt, daß die
Koloniemitglieder viele Arbeiten ver-richten, die nicht mit ihrer direkten Repro-duktion im Zusammenhang stehen. Da

unterschiedliche Arbeiten mit unter-schiedlichem Überlebensrisiko behaftetsind, erfordert die Entscheidung, welchesEinzeltier in einem Volk welche Arbeit
ausführt, einen Kompromiß zwischen denArbeiterinnen. Es ist ein Regelfall, daßäußere Einwirkungen wesentlich mehrzur Sterblichkeit beitragen, sobald dieTätigkeiten abseits vom Nest ausgeführtwerden, als bei Arbeiten innerhalb desNestes. Bei Tätigkeiten ausserhalb desNestes kann das Risiko sich hierbei in
Abhängigkeit von Wetterbedingungenändern.
Die Hypothese, daß die Verteilung derArbeit bei Sammlerinnen eine Folge derVerteilung des Risikos zwischen Arbei-terinnen mit unterschiedlicher Lebenser-
wartung ist, wurde unter Verwendungeines dynamischen Modellprogrammsuntersucht und durch Experimentegestützt. In diesem Modell wurden unterder Annahme, dass jüngere Arbeiterin-nen Arbeiten innerhalb des Nestes ver-
richten, nur Arbeiterinnen mit einem Altervon mehr als 19 Tagen berücksichtigt.Ebenso nahmen wir an, daß das Alterndie Sterblichkeit der Arbeiterinnen beein-
flußt, wobei von einer begrenzten maxi-malen Lebensdauer von 100 Tagen beigesunden und 80 Tagen bei infiziertenArbeiterinnen ausgegangen wurde
(Abb. 1).
Das Rechenmodell sagt voraus, daß bei
verhältnismäßig geringem Risiko alleArbeiterinnen unabhängig von ihrem Alteroder Gesundheitszustand zur Übernahmevon Arbeiten bereit sein sollten (Abb. 2).In Abhängigkeit von der entsprechenddem Gesundheitszustand und dem Alterzu erwartetenden Lebensdauer, könnendie Sammlerinnen ihren Lebens-Gesamt-
eintrag dadurch maximieren, daß sie unter-schiedlich hohe Maximalrisiken währenddes Sammelns eingehen (Abb. 2). Daherwürden Arbeiterinnen ihr Sammelverhal-ten nach ihrem Alter oder Gesundheits-zustand richten. Solange sie jung sind,
würden sie hauptsächlich unter sicherenBedingungen sammeln, wenn sie alt oderkrank sind aber auch riskantes Sammeln
akzeptieren. Als Folge wären kranke undalte Arbeiterinnen unter risikoreichen Wet-
terbedingungen im Vergleich zu sicherenBedingungen überrepräsentiert (Abb. 3B).
In einem Freilandversuch wurden andrei gepaarten Beobachtungen mit kalt-bewölkten oder warm-sonnigen Tagen imFrühjahr und Sommer wassersammelndeHonigbienen von einer Wasserstelle aneinem Bienenstand mit etwa 60 Völkern
abgefangen. Ihre zu erwartende Lebens-dauer wurde anhand der Flügelabnutzungund dem Befall mit dem Parasiten Nosema
apis im Mitteldarm bewertet. Das Ergeb-nis dieses Freilandversuchs bestätigt dieVorhersage aus dem Modell über dieAltersstruktur und den Gesundheitszu-stand der Sammlerinnen. Bei den wasser-sammelnden Arbeiterinnen zeigte derAnteil sowohl von befallenen als auch vonArbeiterinnen mit deutlich beschädigtenFlügeln eine Überrepräsentation alter undkranker Bienen bei risikoreichen Wetter-
bedingungen im Vergleich zu den sicherenBedingungen.
Obwohl allgemein bekannt ist, daß beivielen Insektensozietäten die sicheren Auf-
gaben innerhalb des Nests gewöhnlich injüngerem Alter ausgeführt werden als dieriskanten Arbeiten außerhalb des Nestes,sind diese Befunde unseres Wissens die
ersten, die die Hypothese unterstützen,daß bei Honigbienen und möglicherweiseauch bei anderen eusozialen Insekten die
Arbeitsteilung auf die Risikoverteilungzwischen Arbeiterinnen unterschiedlicher
Lebenserwartung zurückzuführen seinkönnte. © Inra/DIB/AGIB/Elsevier, Paris
Apis mellifera / Arbeitsteilung / Risiko-verteilung / Lebenserwartung / Nosemaapis

REFERENCES
Alford DV (1975) Bumblebees. Davis-Poynter, Lon-don
Bellman R (1957) Dynamic Programming. PrincetonUniv Press, Princeton, NJ
Brooks A, Lithgow GJ, Johnson TE (1994) Mortal-ity rates in a genetically heterogeneous populationof Caenorhabditis elegans. Science 263, 668-671
Dukas R, Visscher K (1994) Lifetime learning byforaging honey bees. Anim Behav 48, 1007-1012
El-Shemy AAM, Pickard RS (1989) Nosema apisZander infection levels in honeybees of knownage. J Apic Res 28, 101-106
Fénéron R, Durand JL, Jaisson P (1996) Relationbetween behaviour and physiological maturati.Behaviour 133, 791-806
Furgala B, Mussen EC (1978) Protozoa. In: HoneyBee Pests, Predators and Diseases (R Morse,ed), Cornell Univ Press, Ithaca, 63-77
Gerber C, Badertscher S, Leuthold RH (1988)Polyethism in Macrotermes bellicosus (Isoptera).Insectes Soc 35, 226-240
Gordon DM (1996) The organization of work insocial insect colonies. Nature 380, 121-124
Guzmán-Novoa E, Gary NE (1993) Genotypic vari-ability of components of foraging behavior inhoney bees (Hymenoptera: Apidae). J Econ Ento-mol 86, 715-721
Hamilton WD (1964) The genetical evolution ofsocial behaviour. I and II. J Theor Biol 7, 1-52
Hassanein MH (1953) The influence of infectionwith Nosema apis on the activities and longevityof worker honeybee. Ann Appl Biol 40, 418-423
Hölldobler B, Wilson EO (1990) The Ants. Belk-nap/Harvard Univ Press, Cambridge, MA
Jeanne RL (1986) The evolution of the organizationof work in social insects. Monitore Zool Ital (NS)20, 119-133
Jeanne RL (1991) Polyethism. In: The Social Biologyof Wasps (KG Ross, RW Matthews, eds),389-425
Kolmes SA (1986) Age polyethism in worker honeybees. Ethology 71, 252-255
Mangel M, Clark CW (1988) Dynamic Modeling inBehavioral Ecology. Princeton Univ Press,Princeton, NJ
Michener CD (1969) Comparative social behavior ofbees. Annu Rev Entomol 14, 299-342
Milne CP (1980) Laboratory measurement of honeyproduction in the honeybee. 2. Longevity orlength of life of caged workers. J Apic Res 19,172-175
Moffet JO, Lawson FA (1975) Effects of Nosemainfection on O2 consumption by honey bee. JEcon Entomol 68, 627-629
O’Donnell S (1996) Reproductive potential and divi-sion of labor in wasps: are queen and worker
behavior alternative strategies? Ethol Ecol Evol8, 305-308
O’Donnell S, Jeanne RL (1995a) Implications ofsenescence patterns for the evolution of agepolyethism in eusocial insects. Behav Ecol 6,269-273
O’Donnell S, Jeanne RL (1995b) Worker lipid storesdecrease with outside-nest task performance inwasps: implications for the evolution of agepolyethism. Experientia 51, 749-752
Oster GF, Wilson EO (1978) Caste and Ecology inthe Social Insects. Princeton Univ Press, Prince-ton, NJ
Pickard RS, El-Shemy AAM (1989) Seasonal vari-ation in the infection of honeybee colonies withNosema apis Zander. J Apic Res 28, 93-100
Ratnieks FL, Visscher K (1989) Worker policing inthe honeybee. Nature 342, 796-797
Robinson GE (1992) Regulation of division of laborin insect societies. Annu Rev Entomol 37,637-665
Robinson GE, Page RE Jr (1988) Genetic determi-nation of guarding and undertaking in honey beecolonies. Nature 333, 356-358
Sakagami SF, Fukuda H (1968) Life tables forworker honeybees. Res Pop Ecol 10, 127-139
Schmid-Hempel P (1987) Efficient nectar-collect-ing by honeybees. I. Economic models. J AnimEcol 56, 209-218
Schmid-Hempel P (1995) Parasites and social insects.Apidologie 26, 255-271
Schmid-Hempel P, Schmid-Hempel R (1984) Lifeduration and turnover of foragers in the antCataglyphis bicolor (Hymenoptera, Formicidae).Insectes Soc 31, 345-360
Schmid-Hempel P, Schmid-Hempel R (1993) Trans-mission of a pathogen in Bombus terrestris, witha note on division of labour in social insects.Behav Ecol Sociobiol 33, 319-327
Schmid-Hempel P, Wolf T (1988) Foraging effortand life span of workers in a social insect. JAnimEcol 57, 509-521
Seeley TD (1982) Adaptive significance of the agepolyethism schedule in honeybee colonies. BehavEcol Sociobiol 11, 287-293
Seeley TD, Kolmes SA (1991) Age polyethism forhive duties in honey bees - illusion or reality?Ethology 87, 284-297
Sokal RR, Rohlf FJ (1981) Biometry. WH Freemanand Company, New York
Sundström L, Chapuisat M, Keller L (1996) Condi-tional manipulation of sex ratios by ant workers:a test of kin selection theory. Science 274,993-995
Trivers RL, Hare H (1976) Haplodiploidy and theevolution of social insects. Science 191, 249-263
Visscher PK, Crailsheim K, Sherman G (1996) Howdo honey bee (Apis mellifera) fuel their waterforaging flights? J Insect Physiol 42, 1089-1094

Visscher PK, Dukas R (1997) Survivorship of for-aging honey bees. Insectes Soc 44, 1-5
Wang Der-I, Moeller FE (1969) Histological com-parisons of the development of hypopharyngealglands in healthy and nosema-infected workerhoney bee. J Invertebr Pathol 14, 135-142
Wang Der-I, Moeller FE (1970) The division of laborand queen attendance behavior of nosema-infected worker honey bees. J Econ Entomol 63,1539-1541
Woyciechowski M, Lomnicki A (1987) Multiplemating of queens and the sterility of workers
among eusocial Hymenoptera. J Theor Biol 128,317-327
Woyciechowski M, Lomnicki A (1995) Honey beeworkers parasitized by Nosema apis chooseriskier foraging strategies. Pszczel Zesz Nauk 39,209-21
Woyciechowski M, Koz&jadnr;owski J, Grzanka G (1996)Life expectancy and risk-taking in foraging honeybee workers. Proceedings XX InternationalCongress of Entomology, Firenze, Italy, August,394




















