
The relationship between hygienic behavior and suppression of mite reproduction as honey bee ( Apis...
10
31 Apidologie 37 (2006) 31–40 © INRA/DIB-AGIB/ EDP Sciences, 2005 DOI: 10.1051/apido:2005052 Original article The relationship between hygienic behavior and suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of resistance to Varroa destructor 1 Abdullah IBRAHIM, Marla SPIVAK* Department of Entomology, University of Minnesota 219 Hodson Hall, 1980 Folwell Av., St. Paul, MN 55108, USA Received 4 April 2005 – revised 3 June 2005 – accepted 22 June 2005 Abstract – We compared the mechanisms of resistance to Varroa destructor displayed by bees bred for Suppression of Mite Reproduction (SMR) and hygienic behavior (HYG). Mites from SMR and HYG source colonies were introduced into recently sealed SMR and HYG worker brood, and the infested pupae were placed either into recipient colonies or into an incubator. SMR colonies removed significantly more mite- infested pupae than the HYG colonies. The reproductive success (fertility and number of viable female offspring) of mites from both sources on pupae not removed by bees was significantly lower in SMR colonies. Within the incubator, the reproductive success of mites was also lower on SMR worker pupae, and lowest when mites from SMR colonies were introduced on SMR brood. Our findings indicated that bees bred for SMR express hygienic behavior; adult bees selectively remove pupae infested with mites. In addition, there is an effect of SMR pupae that reduces mite reproductive success that requires further investigation. Apis mellifera / suppression of mite reproduction / hygienic behavior / Varroa destructor 1. INTRODUCTION The parasitic mite, Varroa destructor Anderson and Truman is a serious pest of honey bee colonies derived from European subspe- cies of Apis mellifera L. The application of pesticide treatments in bee colonies has led to the development of resistance by the mites (reviewed in Milani, 1999), and the propaga- tion of susceptible colonies by beekeepers. A more sustainable solution to the mite problem is to select lines of honey bees that show natural mechanisms of resistance to the mite. Harbo and Hoopingarner (1997) bred a line of bees that maintained low mite levels because the mites appeared to have low reproductive success on worker brood. They found the trait to be a heritable trait of the bees, and called it Suppression of Mite Reproduction (SMR). In colonies bred for SMR, the mites entered worker brood cells to feed and reproduce; how- ever, the authors reported that the mites died in the cell without reproducing, produced no progeny, produced males only, or produced progeny too late to mature (Harbo and Harris, 1999). The mechanism for how the bees could suppress mite reproduction in these ways was not known (Harris and Harbo, 1999). Another factor that hinders mite reproduc- tion is hygienic behavior. Hygienic behavior is considered the primary mechanism of honey bee resistance to two diseases of brood, Amer- ican foulbrood (caused by the bacteria, Paeni- bacillus larvae subsp. larvae; Rothenbuhler, 1964; Spivak and Reuter, 2001a) and chalk- brood (caused by the fungus, Ascosphaera apis; * Corresponding author: [email protected] 1 Manuscript editor: Gudrun Koeniger Article published by EDP Sciences and available at http://www.edpsciences.org/apido or http://dx.doi.org/10.1051/apido:2005052
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The relationship between hygienic behavior and suppression of mite reproduction as honey bee ( Apis...

31Apidologie 37 (2006) 31–40© INRA/DIB-AGIB/ EDP Sciences, 2005DOI: 10.1051/apido:2005052
Original article
The relationship between hygienic behavior and suppression of mite reproduction as honey bee
(Apis mellifera) mechanisms of resistance to Varroa destructor1
Abdullah IBRAHIM, Marla SPIVAK*
Department of Entomology, University of Minnesota 219 Hodson Hall, 1980 Folwell Av., St. Paul, MN 55108, USA
Received 4 April 2005 – revised 3 June 2005 – accepted 22 June 2005
Abstract – We compared the mechanisms of resistance to Varroa destructor displayed by bees bred forSuppression of Mite Reproduction (SMR) and hygienic behavior (HYG). Mites from SMR and HYG sourcecolonies were introduced into recently sealed SMR and HYG worker brood, and the infested pupae wereplaced either into recipient colonies or into an incubator. SMR colonies removed significantly more mite-infested pupae than the HYG colonies. The reproductive success (fertility and number of viable femaleoffspring) of mites from both sources on pupae not removed by bees was significantly lower in SMRcolonies. Within the incubator, the reproductive success of mites was also lower on SMR worker pupae, andlowest when mites from SMR colonies were introduced on SMR brood. Our findings indicated that beesbred for SMR express hygienic behavior; adult bees selectively remove pupae infested with mites. Inaddition, there is an effect of SMR pupae that reduces mite reproductive success that requires furtherinvestigation.
Apis mellifera / suppression of mite reproduction / hygienic behavior / Varroa destructor
1. INTRODUCTION
The parasitic mite, Varroa destructorAnderson and Truman is a serious pest of honeybee colonies derived from European subspe-cies of Apis mellifera L. The application ofpesticide treatments in bee colonies has led tothe development of resistance by the mites(reviewed in Milani, 1999), and the propaga-tion of susceptible colonies by beekeepers. Amore sustainable solution to the mite problemis to select lines of honey bees that show naturalmechanisms of resistance to the mite.
Harbo and Hoopingarner (1997) bred a lineof bees that maintained low mite levels becausethe mites appeared to have low reproductivesuccess on worker brood. They found the traitto be a heritable trait of the bees, and called it
Suppression of Mite Reproduction (SMR). Incolonies bred for SMR, the mites enteredworker brood cells to feed and reproduce; how-ever, the authors reported that the mites died inthe cell without reproducing, produced noprogeny, produced males only, or producedprogeny too late to mature (Harbo and Harris,1999). The mechanism for how the bees couldsuppress mite reproduction in these ways wasnot known (Harris and Harbo, 1999).
Another factor that hinders mite reproduc-tion is hygienic behavior. Hygienic behavior isconsidered the primary mechanism of honeybee resistance to two diseases of brood, Amer-ican foulbrood (caused by the bacteria, Paeni-bacillus larvae subsp. larvae; Rothenbuhler,1964; Spivak and Reuter, 2001a) and chalk-brood (caused by the fungus, Ascosphaera apis;
* Corresponding author: [email protected] Manuscript editor: Gudrun Koeniger
Article published by EDP Sciences and available at http://www.edpsciences.org/apido or http://dx.doi.org/10.1051/apido:2005052

32 A. Ibrahim, M. Spivak
Gilliam et al., 1983, 1988), and is one of variousmechanisms of resistance against V. destructor(reviewed in Boecking and Spivak, 1999). Beesbred for hygienic behavior detect, uncap, andremove diseased brood from the colony beforethe disease reaches the infectious stage (Park,1937; Woodrow and Holst, 1942; Rothenbuhler,1964). Bees bred for hygienic behavior alsodetect and remove mite-infested pupae fromthe colony (Spivak, 1996). In contrast to theremoval of diseased infested brood which mustbe done before or during the 5th larval instar toavoid handling and transmitting disease spores,hygienic bees removed the majority of mite-infested brood at least 60 hours after the cell issealed, which is after the mite has initiated ovi-position (Donzé et al., 1996), when the bee isin the pupal stage (Spivak, 1996). Removingthe pupa at that time ensures destruction of anymite progeny. Extensive field studies carriedout by Spivak and Reuter (1998a, 2001b)showed that the colonies bred for hygienicbehavior produced as much honey as unse-lected control colonies and had lower levels ofmites than the control colonies, but the degreeof removal of infested pupae by hygienicbehavior was not sufficient to maintain the mitepopulation below an estimated economic thresh-old (Delaplane and Hood, 1999).
The aim of our study was to determine howbees from the SMR line suppressed mite repro-duction. During the course of other experimentsalso involving the SMR line, we conducted astandard freeze-killed brood assay for hygienicbehavior (Spivak and Reuter, 1998b) on somecolonies containing queens bred for SMRobtained from J. Harbo. We found that SMRcolonies removed 98.6% (± 0.9 s.d., n = 7 col-onies) of the freeze-killed brood within 48 hours,which indicated that they were expressinghygienic behavior. These results were surpris-ing because J. Harbo selectively bred only forreduced mite reproduction, not for hygienicbehavior (Harbo and Harris, 1999). Based onthese findings, and our previous data (Spivak,1996) which suggest that the bees detect themajority of mite-infested pupae only after themite has initiated oviposition, we hypothesizedthat the SMR bees could be detecting andremoving pupae on which the mites were repro-ducing, leaving mites that did not reproducesuccessfully. If so, the assay for SMR, whichinvolves inspecting mite reproductive success
on late stage pupae (Harbo and Harris, 1999)would reveal pupae with mites that did notreproduce successfully because the majority ofthe others would have been removed previ-ously by the bees. We used mite fertility and thenumber of viable female offspring, NVO, astwo measures of reproductive success. Fertilityis defined as the number of foundress mites thatproduced at least one male offspring (e.g.,Mondragon et al., 2005). NVO is defined morespecifically by the presence of at least one liveadult daughter mite and one live adult malewhen the bee pupa is in the gray wing-padstage, at least 230 h or approximately 10 dayspostcapping (Medina and Martin, 1999; Corrêa-Marques et al., 2003). At this stage of develop-ment of the bee pupa, there is a high probabilitythat the adult daughter would have been matedby the male (Donzé et al., 1996), and so can beconsidered a viable female offspring.
The removal of mite-infested pupae is aneffect of adult bee behavior. We also wanted toinvestigate the possibility that there could be abrood effect; i.e., there might be some physio-logical factor associated with SMR workerbrood that somehow reduces the reproductivesuccess of mites. Therefore, our experimentswere designed to answer three questions; thefirst two concerned the extent of the adult beeeffect on mite reproduction, and the third on thepossible brood effect. 1. Do bees bred for Suppression of Mite
Reproduction (SMR) detect and removemite-infested pupae?
2. If so, do SMR bees preferentially removepupae infested with mites with highreproductive success leaving pupae withmites with low reproductive success?
3. What is the reproductive success of mitesfrom SMR colonies when bees are notallowed to remove mite-infested brood?
2. MATERIALS AND METHODS
2.1. Lines of bees
The experiments were conducted on the St Paulcampus of the University of Minnesota in the sum-mer of 2004. The breeding program for hygienicbehavior was established in 1993 at the Universityof Minnesota by selecting colonies of Italian –derived A. mellifera bees using a freeze-killed broodassay (Spivak and Downey, 1998; Spivak and Reuter,

Mechanisms of resistance to Varroa 33
1998b). Colonies that uncapped and removed 95%freeze-killed brood within 48 h were consideredhygienic. To establish and maintain the hygieniclines, we raised daughter queens from colonies thatdisplayed the most rapid removal rates. For eachgeneration, the daughter hygienic queens wereinstrumentally inseminated with a mixture of semencollected from drones from different hygienic colo-nies (Spivak and Gilliam, 1998).
J. Harbo provided us with the instrumentallyinseminated queens from the SMR line. To breedthis line, Harbo and Hoopingarner (1997) beganselecting honey bees colonies for resistance toV. destructor by measuring several characteristicsthat impeded the growth of mite populations in beecolonies, such as hygienic behavior, groomingbehavior, short post capping period, and the fre-quency of non-reproducing mites. Of 43 colonies,only three had fewer mites than at the beginning ofthe test, and only “non-reproduction” (defined asmites that enter the cell to reproduce but produce noprogeny, produce males only, produce progeny toolate to mature, or die in the cell before they can pro-duce) was correlated with changes in mite popula-tion (Harbo and Harris, 1999). Daughter queenswere instrumentally inseminated with semen fromsingle drones to propagate a line of bees that sup-pressed mite reproduction.
2.2. Mite source
All mites used in the experiments were collectedfrom one hygienic and one SMR colony to controlfor variability in reproductive success of the mitesdue to colony rearing environment. Previousresearch (Harbo and Harris, 1999) demonstrated thatthe reproductive success of mites within SMR colo-nies is not suppressed until after one reproductivecycle, a phenomenon they termed “Delayed Sup-pression of Mite Reproduction.” Therefore, themites we collected from the SMR mite-source col-ony had been present for at least one month beforethe experiments began. The use of mites from a hygi-enic colony served as a control because our prelim-inary research (Ibrahim and Spivak, 2004 andunpublished data) showed that the reproductive suc-cess of individual mites in worker brood within hygi-enic colonies is the same as the success of mites inunselected colonies over time.
2.3. Experimental methods
Question 1: We tested whether colonies from theHygienic line (HYG) and from the SMR line pref-erentially removed mites that have developed withintheir own line or the other line of bees. The methodsof Boecking (1992), and Spivak (1996) were used toobtain mite-infested brood. Female mites were dis-lodged from adult bees of the two mite-source col-
onies, one SMR and one HYG, by dusting about3000 bees in powdered sugar (Macedo and Ellis,2002). In the lab, mites from both mite source colo-nies were introduced into cells containing recentlysealed 5th instar larvae within the recipient threeSMR and three HYG colonies, so that each recipientcolony received 80 mites, 40 from each mite source.The larvae to be infested were obtained by markingunsealed 5th instars on a plastic transparency, andfour hours later, introducing mites into only thosethat had been sealed in the interim. The mites wereintroduced by making a small opening in the edge ofeach cell's wax capping using a sharp forceps, intro-ducing the mite through the opening with a finepaintbrush, and then carefully resealing the cell. Anadditional set of cells containing recently sealed 5thinstars were opened and resealed, without introduc-ing a mite, to serve as controls for the cell manipu-lation. Each location of mite-treated and control cellwas marked on a plastic transparency, and the framesof brood containing the treated cells were returnedto the original recipient colonies. Every two days for10 days, the number of mite-infested and controlcells removed by the bees were recorded.
Question 2: To test whether SMR bees preferen-tially remove pupae infested with mites with highreproductive success, leaving pupae with mites withlow reproductive success, we recorded fertility andthe number of viable female offspring of the miteson pupae that were not removed by the bees withinthe three SMR and three HYG colonies fromQuestion 1. On day 10 after the brood was sealed,remaining pupae in the gray wing-pad stage wereinspected for the presence of mites and their progenyunder a dissecting microscope. The pupae from eachcell were removed into a petri dish and each wasexamined for the presence of the mother mite, amale, and the different life stages of its offspring fol-lowing the ontogenetic developmental chart inMartin (1994). Mite fertility was calculated for eachcolony by dividing the number of reproductive foun-dresses (mites that produced at least one male off-spring) by the number of introduced mites thatremained on pupae. Number of viable offspring wasdetermined by dividing the total number of adultfemale daughters by the total number of remainingmites (including reproductive and non reproductivefoundress). Adult daughters can be recognized bythe presence of a live, adult male and the presenceof a female with a shed exuvia of the final molt, orif the exuvia could not be found, by light dark brown-ish color of the mite's body.
Question 3: In this experiment, we wanted toeliminate the effect of adult bee removal behavior onmite-reproductive success. Mites were collected andmanually introduced into brood cells, as describedabove in Question 1. The frames containing theinfested cells were placed in an incubator held at hive

34 A. Ibrahim, M. Spivak
temperature and humidity (34 °C and 70% RH). Thetwo measures of reproductive success of the miteson grey wing-pad pupae were recorded on day10,after the brood cells were sealed, following the meth-ods described above.
2.4. Statistical analysis
Question 1. The proportion of mites (from bothmite sources) removed by the SMR and HYG colo-nies of bees on days 2, 4, 6, 8 and 10 after mite intro-duction was analyzed by comparing Kaplan-Meiersurvivorship curves using the Survfit procedurefrom R statistical software.
Questions 2 and 3. Measures of mite fertility andthe number of female offspring produced were com-pared using a non-linear mixed-model (ProcNLMixed, SAS version 8.2; see also, McCullochand Searle, 2002), or log likelihood chi-square tests.
3. RESULTS
3.1. Question 1
By day 10 after mite introduction, the threeSMR colonies removed 84.5% ± 7.4 of the
pupae containing mites from the SMR mite-source colony, and 80.1% ± 14.9 of the pupaecontaining mites from the HYG mite-sourcecolony. In contrast, the three HYG coloniesremoved 65.7% ± 10.8 of the pupae containingmites from the SMR mite-source colony and61.7% ± 7.3 from the HYG mite-source colo-nies. Of the control cells (cells manipulatedwith no mite introduction) the SMR and HYGcolonies removed pupae from 10.8% ± 1.4 and12.4% ± 0.7 of the cells, respectively. Figure 1shows the data as a survival function; i.e., theproportion of treated and pooled control pupaethat survived (were not removed) at each timeinterval. A log rank test of the survival curvesrevealed a highly significant treatment effect(χ2 = 395; df = 4, P < 0.0001) between the sur-vival of pupae within mite-infested vs. controlcells. Subsequently, to determine if the sourceof mites affected the proportion of pupae thatsurvived (were not removed) in each line, thecontrol cells were excluded from the data set.There was no significant effect of mite sourceon the number of pupae removed by the bees(χ2 = 2.9; df = 1, P = 0.088). However, therewas a highly significant effect of bee type:(χ2 = 18.9; df = 1, P < 0.0001); the SMR col-onies removed significantly more infestedpupae than the HYG colonies, irrespective ofthe source of the mites.
3.2. Question 2
The reproductive success of the mites onpupae that were not removed by the bees isshown in Table I. Within the three SMR colo-nies combined, only 43 of the 241 experimen-tally infested pupae remained, and uponinspection, mites were missing in 14 (32.6%)of the pupal cells, and were found dead withinin 2 (4.6%) others. Of the 27 remaining pupaethat contained live mites in the SMR colonies,10 contained mites that did not oviposit, so theoverall fertility (proportion of mites that pro-duced at least one egg) was 63%. However,only 1 mite produced a viable female offspringthat would have mated with the male in the cell,resulting in an overall proportion of viablefemale offspring produced in SMR colonies ofonly 3.7%.
In contrast, 89 of 243 infested pupaeremained within the HYG colonies, and miteswere missing from 5 cells (5.6%) or were found
Figure 1. Survival curve showing the proportion ofmite-infested pupae surviving (i.e., not removed bybees) every two days from the day the mites wereintroduced (within 4 h of the cell being capped withwax by the bees) to day 10 post-capping within 3HYG and 3 SMR colonies. Control cells (pooledamong all colonies) refer to cells containing recentlysealed larvae that were sham-manipulated; the cellwas opened and resealed without introducing a mite.H–H = HYG bee colonies–HYG mite-sourcecolony; H–S = HYG bee colonies–SMR mite-source colony; S–H = SMR bee colonies–HYG mitesource colonies; S–S = SMR bee colonies–SMRmite-source colonies.

Mechanisms of resistance to Varroa 35
dead in 2 cells (2.2%). Of the 82 remainingpupae containing live mites, 70 (85.4%) of themites were fertile, and 22 (26.8%) produced atleast one viable adult female offspring.
Because of the low number of infested pupaeremaining within the SMR colonies, it was notpossible to use a full statistical model with col-onies as a random effect to examine measuresof mite fertility or number of viable female off-spring. Therefore, the data was pooled amongcolonies within each line and analyzed usinglog likelihood chi-square tests. Mites remain-ing within SMR colonies were significantlyless fertile than mites within HYG colonies(effect of bee type: χ2 = 6.00; df = 1, P = 0.014),irrespective of mite source (effect of mitesource: χ2 = 1.11; df = 1, P = 0.293). Likewise,mites remaining on pupae within SMR colo-nies produced significantly fewer viablefemale offspring compared to mites withinHYG colonies (χ2 = 4.69; df = 1, P = 0.030),irrespective of mite source (χ2 = 1.31; df = 1,P = 0.252).
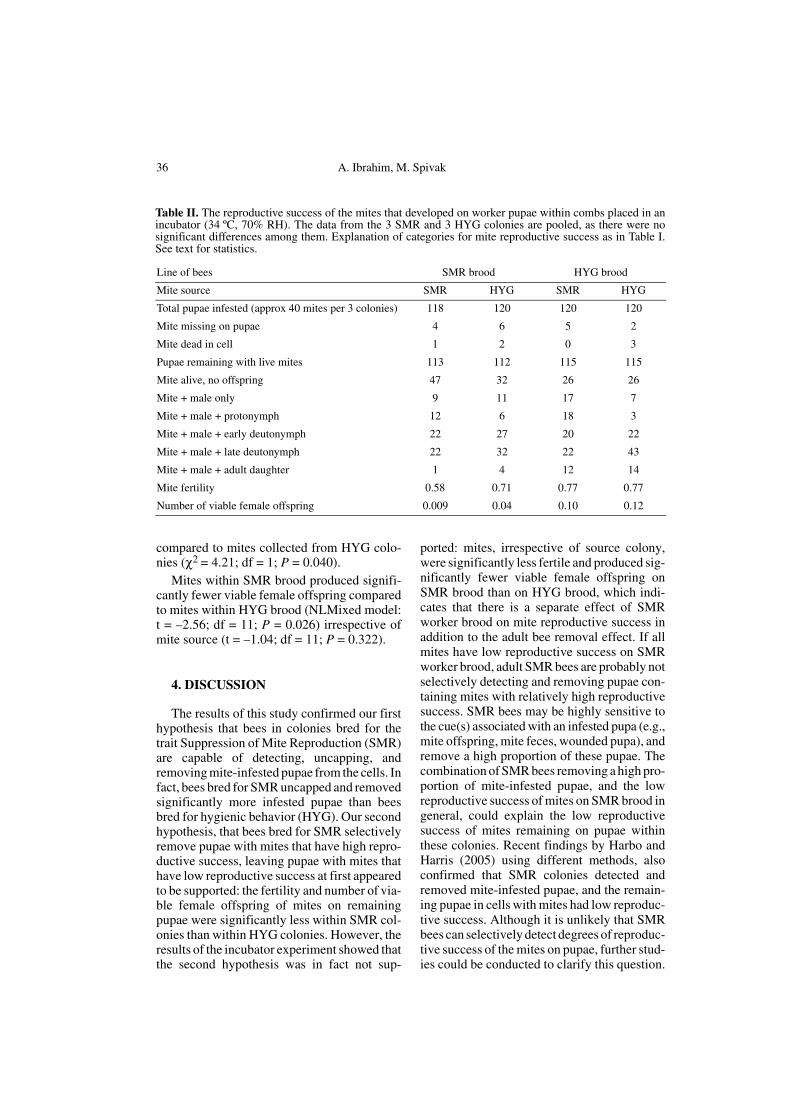
3.3. Question 3
Table II shows results from when the exper-imentally infested pupae developed in an incu-bator, so that the bees could not interrupt thereproductive cycle of the mites. The larger sam-ple sizes allowed us to use a non-linear mixedmodel (Proc NLMixed; SAS 8.2), to take intoconsideration the source of mites introducedonto the pupae within each line of bees as maineffects with individual colonies as a randomsource of variation. There was no interactionbetween brood type (SMR or HYG pupae) andmite source, so the model was run excluding theinteraction term. The mites were significantlyless fertile on SMR brood than on HYG brood(NLMixed model: t = –2.44; df = 11; P =0.033), irrespective of mite source (t = –0.27;df = 11; P = 0.795). A further comparison ofthe fertility of mites within SMR brood only,using a log likelihood chi-square test, indicatedthat mites collected from SMR colonies weresignificantly less fertile on their own pupae
Table I. Number of mite-infested worker pupae that were not removed by bees, and the reproductivesuccess of the surviving mites on the pupae. The data from the 3 SMR and 3 HYG colonies are pooled, asthere were no significant differences among them. See text for statistics.
Line of bees SMR colonies HYG colonies
Mite source SMR HYG SMR HYG
Total pupae infested (approx 40 mites per 3 colonies) 122 119 123 120
Pupae not removed by bees 19 24 42 47
Mite missing on pupae1 7 7 3 2
Mite dead in cell 0 2 2 0
Pupae remaining with live mites 12 15 37 45
Mite alive, no offspring 6 4 6 6
Mite + male only 2 3 1 3
Mite + male + protonymph2 3 1 2 1
Mite + male + early deutonymph2 0 5 4 2
Mite + male + late deutonymph2 0 2 17 18
Mite + male + adult daughter2 1 0 7 15
Mite fertility (proportion of mites that oviposited) 0.50 0.73 0.84 0.87
Number of viable female offspring (proportion of adult offspring/cell)
0.08 0 0.19 0.33
1 The pupa was experimentally infested with a mite, but upon inspection on day 10, the mite was not present in the cell. 2 The presence of a protonymph, deutonymph, or adult daughter refers to the oldest female progeny in the cell when the pupa was in the grey wing-pad stage, although younger female progeny were also present in all cases.
36 A. Ibrahim, M. Spivak
compared to mites collected from HYG colo-nies (χ2 = 4.21; df = 1; P = 0.040).
Mites within SMR brood produced signifi-cantly fewer viable female offspring comparedto mites within HYG brood (NLMixed model:t = –2.56; df = 11; P = 0.026) irrespective ofmite source (t = –1.04; df = 11; P = 0.322).
4. DISCUSSION
The results of this study confirmed our firsthypothesis that bees in colonies bred for thetrait Suppression of Mite Reproduction (SMR)are capable of detecting, uncapping, andremoving mite-infested pupae from the cells. Infact, bees bred for SMR uncapped and removedsignificantly more infested pupae than beesbred for hygienic behavior (HYG). Our secondhypothesis, that bees bred for SMR selectivelyremove pupae with mites that have high repro-ductive success, leaving pupae with mites thathave low reproductive success at first appearedto be supported: the fertility and number of via-ble female offspring of mites on remainingpupae were significantly less within SMR col-onies than within HYG colonies. However, theresults of the incubator experiment showed thatthe second hypothesis was in fact not sup-
ported: mites, irrespective of source colony,were significantly less fertile and produced sig-nificantly fewer viable female offspring onSMR brood than on HYG brood, which indi-cates that there is a separate effect of SMRworker brood on mite reproductive success inaddition to the adult bee removal effect. If allmites have low reproductive success on SMRworker brood, adult SMR bees are probably notselectively detecting and removing pupae con-taining mites with relatively high reproductivesuccess. SMR bees may be highly sensitive tothe cue(s) associated with an infested pupa (e.g.,mite offspring, mite feces, wounded pupa), andremove a high proportion of these pupae. Thecombination of SMR bees removing a high pro-portion of mite-infested pupae, and the lowreproductive success of mites on SMR brood ingeneral, could explain the low reproductivesuccess of mites remaining on pupae withinthese colonies. Recent findings by Harbo andHarris (2005) using different methods, alsoconfirmed that SMR colonies detected andremoved mite-infested pupae, and the remain-ing pupae in cells with mites had low reproduc-tive success. Although it is unlikely that SMRbees can selectively detect degrees of reproduc-tive success of the mites on pupae, further stud-ies could be conducted to clarify this question.
Table II. The reproductive success of the mites that developed on worker pupae within combs placed in anincubator (34 ºC, 70% RH). The data from the 3 SMR and 3 HYG colonies are pooled, as there were nosignificant differences among them. Explanation of categories for mite reproductive success as in Table I.See text for statistics.
Line of bees SMR brood HYG brood
Mite source SMR HYG SMR HYG
Total pupae infested (approx 40 mites per 3 colonies) 118 120 120 120
Mite missing on pupae 4 6 5 2
Mite dead in cell 1 2 0 3
Pupae remaining with live mites 113 112 115 115
Mite alive, no offspring 47 32 26 26
Mite + male only 9 11 17 7
Mite + male + protonymph 12 6 18 3
Mite + male + early deutonymph 22 27 20 22
Mite + male + late deutonymph 22 32 22 43
Mite + male + adult daughter 1 4 12 14
Mite fertility 0.58 0.71 0.77 0.77
Number of viable female offspring 0.009 0.04 0.10 0.12

Mechanisms of resistance to Varroa 37
The source of the introduced mites, whethercollected from the SMR or HYG colony, didnot have an effect on either line of bees'removal response or on the reproductive suc-cess of the mites on pupae that were notremoved. However, within the incubator, mitesfrom both sources were significantly less fertileon SMR brood than on HYG brood, and thelowest measures of reproductive success wererecorded when mites from SMR colonies wereintroduced on SMR brood.
Although this study was based on a lownumber of colonies (three SMR and three HYG),and the mite source colonies were not repli-cated, our findings, together with those ofHarbo and Harris (2005) provide strong evi-dence that colonies selected for Suppression ofMite Reproduction express hygienic behaviorby detecting and removing mite-infested pupae.Our measures of mite fertility within hygieniccolonies in the field (84–87%) were compara-ble to those reported elsewhere for colonies thatdid not display hygienic behavior (e.g., 88% inMondragon et al., 2005), supporting our use ofhygienic colonies, and mites from hygienic col-onies as controls. For unknown reasons, the fer-tility of mites was 10% lower in brood placedin the incubator compared to in field colonies.It is possible that the humidity level in the incu-bator, which is known to affect mite reproduc-tion (Le Conte et al., 1990; Kraus and Velthuis,1997), may have contributed to this difference.
The findings from the first question indi-cated that the main mechanism of the trait, Sup-pression of Mite Reproduction, is similar to, ifnot the same as, hygienic behavior. Adult beesfrom SMR colonies detect and remove mite-infested pupae. Bees bred for SMR appeared tobe highly sensitive to the cues that elicitremoval behavior and detected and removedmore mite-infested pupae than did the HYGcolonies. One possible explanation for the dif-ference in effectiveness of mite removal by thetwo lines of bees could be due to the differentassays used to select the lines for mite resist-ance. The line bred for hygienic behavior wasselected based on colonies' removal of freeze-killed brood; whereas, the line bred for SMRwas selected based on the low number of mitesremaining in cells just before worker pupaeemerged. The freeze-killed brood assay hasbeen used as an indirect correlate of actualhygienic behavior, which is the removal of dis-
eased or mite infested brood. The results of thefreeze-killed brood assay do not perfectly pre-dict whether a colony will be resistant to dis-ease (Gilliam et al., 1988; Spivak and Gilliam,1993; Spivak and Reuter, 2001a) and bees bredfor hygienic behavior based on this assay do notshow complete resistance to V. destructor(Spivak and Reuter, 1998a, 2001b). Selectionfor colonies that had negative mite populationgrowth after a certain period of time, as wasdone to select the SMR line, apparently putgreater selection pressure on adult bees to detectand remove infested pupae. Therefore, in con-trast to the indirect freeze-killed brood assay,the assay for SMR was direct, and the bees’response was quite specific; they were able todetect and remove more mite-infested pupae.
It is not clear whether HYG and SMR arecontrolled by the same genes. We speculate thatthe SMR bees have greater olfactory sensitivityto the cues associated with mite infested pupae,and respond to those cues with greater speedand acuity than do HYG bees. We base thisspeculation on our previous studies on theolfactory sensitivity and responsiveness ofhygienic vs. non-hygienic bees to the odors ofdiseased brood (Masterman et al., 2001; Spivaket al., 2003). It would be interesting to test theolfactory sensitivity of bees bred for SMRusing the same electroantennogram and pro-boscis-extension response conditioning assaysused in these previous studies. It would also beinteresting to test whether SMR colonies areresistant to the bacterial disease American foul-brood, and the fungal disease, chalkbrood andcompare their removal response with coloniesselected for HYG behavior based on the freeze-killed assay. If SMR colonies are detect andremove a greater proportion of diseased brood,it would indicate the basic genetic and neuro-logical mechanisms are the same between theHYG and SMR traits.
The results of our third question add newinformation to the recent study by Harbo andHarris (2005) and indicate that there may begenetic or environmentally induced differencesbetween worker pupae from HYG and SMRcolonies that influence mite reproductive suc-cess. Mites collected from HYG and SMRsource colonies were less fertile and producedfewer viable female offspring on SMR pupaethan on HYG pupae. This finding suggests thatthere may be a physiological effect of SMR

38 A. Ibrahim, M. Spivak
worker pupae that reduces mite reproductivesuccess. The mechanism for this effect of broodtype on mite reproduction is unknown andremains to be investigated. It is possible that thesame factor that hinders mite reproduction alsoaffects the development and viability of thepupa itself, as some colonies bred for SMRhave very low brood viability.
ACKNOWLEDGEMENTS
We thank Gary Reuter for his assistance withdata collection, colony maintenance, and commentson original versions of the manuscript. Dr. SanfordWeisberg, School of Statistics, University ofMinnesota and Dr. Matt Carroll provided statisticaladvice. The research was supported by North Cen-tral Sustainable Agriculture, Research and Educa-tion grant LNC02-202, and by beekeeping associa-tions in MN, ND, SD, IA, and WI. Special thanks tothe MN Honey Producers Association for their con-tinued encouragement and support.
Résumé – Relation entre le comportement hygié-nique et la suppression de la reproduction desacariens comme mécanisme de résistance desabeilles (Apis mellifera) à Varroa destructor. Nousavons étudié le mécanisme selon lequel des abeilles(Apis mellifera), sélectionnées pour le caractère« suppression de la reproduction des acariens »(abeilles SMR), pouvaient agir négativement sur lesuccès reproductif (S.R.) de l’acarien Varroa des-tructor, notre hypothèse étant que les abeilles SMRdétectent et éliminent les nymphes qui ont des aca-riens reproducteurs, donnant ainsi l’impression queles acariens restant sur les nymphes au stade pré-éclosion se reproduisent mal. Nous avons utilisé lafertilité des acariens et le nombre de descendantsfemelles viables comme mesure du S.R. Nous avonsaussi étudié s’il existe un facteur physiologiqueassocié au couvain d’ouvrières SMR qui réduit leS.R. des acariens. Nous avons comparé des coloniessélectionnées pour le caractère SMR, fournies parHarbo, avec des colonies sélectionnées pour le com-portement hygiénique (abeilles HYG) élevées àl’université du Minnesota. Pour tester si les abeillesSMR pouvaient détecter et éliminer les nymphesinfestées, nous avons introduit des acariens des colo-nies sources SMR et HYG dans du couvaind’ouvrières SMR et HYG récemment operculé. Au10e jour après l’introduction des acariens, les troiscolonies SMR avaient éliminé significativementplus de nymphes infestées que les trois coloniesHYG, quelle que fût la source des acariens (P <0,0001) (Fig. 1). Ensuite nous avons relevé le S.R.des acariens sur les nymphes restantes (Tab. I). Lesacariens dans les colonies SMR étaient significati-vement moins fertiles (moins d’acariens ont pondu
au moins un œuf) que ceux des colonies HYG (P =0,014). Ils ont aussi produit significativement moinsde descendantes viables (définies par la présenced’un acarien adulte fille et d’un acarien adulte mâlesur une nymphe au stade du bourrelet alaire gris) queceux des colonies HYG (P = 0,030). Pour étudierl’action du couvain SMR sur le S.R. des acariens,nous avons placé du couvain infesté dans une étuveréglée à la température et à l’hygrométrie de la ruche(Tab. II). Les acariens étaient significativementmoins fertiles (P = 0,033) et ont produit significati-vement moins de descendantes viables (P = 0,026)sur le couvain SMR que sur le couvain HYG. Lesrésultats confirment donc notre hypothèse : lesabeilles SMR détectent, désoperculent et éliminentsignificativement plus de nymphes infestées que lesabeilles HYG. Il ne semble pas que les abeilles SMRéliminent sélectivement les nymphes avec acariensà S.R. élevé et laissent les nymphes avec acariens àS.R. faible, parce que l’étude en étuve a montré quetous les acariens avaient un S.R. plus faible sur lecouvain SMR que sur le couvain HYG. En consé-quence la combinaison des abeilles SMR, quiéliminent une forte proportion d’abeilles infestées,et le faible succès reproductif des acariens sur le cou-vain SMR en général pourraient expliquer le faibleS.R. des acariens restants. D’autres expériences sontnécessaires pour déterminer l’action physiologiquedu couvain SMR sur le succès reproductif des aca-riens et savoir si les caractères HYG et SMR sontrégulés par les mêmes gènes.
Apis mellifera / Varroa destructor / comportementhygiénique / reproduction des acariens /suppression
Zusammenfassung – Wirkungsweisen vonHygiene Verhalten und Unterdrückung der Mil-benvermehrung beim Resistenzmechanismusder Honigbiene (Apis mellifera) gegen Varroa des-tructor. Wir untersuchten den Mechanismus durchden für den Faktor „Unterdrückung der MilbenReproduktion“ (SMR) gezüchtete Bienen den Ver-mehrungserfolg von Varroa destructor negativbeeinflussen. Wir untersuchten die Hypothese, dassSMR-Bienen mit vermehrungsfähigen Milbenbefallene Puppen erkennen und entfernen und damitden Eindruck erwecken, dass die Puppen einenreproduktionshemmenden Faktor besitzen, da dieübriggebliebenen Milben ohne Nachkommenwären. Als Maßsystem für reproduktiven Erfolguntersuchten wir die Milbenfruchtbarkeit undAnzahl von lebensfähigen weiblichen Nachkom-men. Wir untersuchten weiterhin, ob es einen phy-siologischen Faktor gibt, der bei SMR Arbeiterin-nenbrut den reproduktiven Erfolg beeinträchtigt.Wir verglichen SMR Völker von Harbo mit Völkernder Zuchtlinien „Hygiene Verhalten“ (HYG) vonder Universität von Minnesota. Zunächst untersuch-ten wir, ob SMR-Bienen Milben auf befallenen Pup-pen erkennen und entfernen können. Milben von

Mechanisms of resistance to Varroa 39
SMR und HYG Völkern wurden in gerade verdek-kelte Brutwaben von SMR and HYG Arbeiterinnen-zellen eingesetzt und in SMR bzw. HYG Völkergegeben. Zehn Tage nach dem Milbeneinsatz hattendie 3 SMR Völker signifikant mehr befallenen Pup-pen entfernt als die 3 HYG Völker, unabhängig vomUrsprung der Milben (P < 0,0001) (Abb. 1). Alszweites verglichen wir den Reproduktionserfolg derMilben auf den verbliebenen Puppen (Tab. I).Milben, die in den SMR Völkern verblieben,waren signifikant weniger fruchtbar (weniger Mil-ben legten mindestens ein Ei) als Milben in den HYGVölkern (P = 0,014), auch hatten sie signifikantweniger lebensfähige junge Weibchen (definiert alsPräsenz einer lebenden adulten Tochtermilbe undeinem lebenden adulten Männchen auf einem Pup-penstadium mit grauem Flügelansatz) als Milben inHYG Völkern (P = 0,030). Um die Wirkung vonSMR Brut auf die Milbenvermehrung zu prüfen,hielten wir befallene Brut im Wärmeschrank beiTemperatur und Luftfeuchtigkeit wie im Volk(Tab. II). Die Milben waren auf SMR Brut signifi-kant weniger fruchtbar (P = 0,033) und hatten signi-fikant weniger lebensfähige weibliche Nachkom-men (P = 0,026) als auf HYG Brut. Die Ergebnissebestätigen, dass Bienen der SMR Linien mit Milbenbefallenen Puppen signifikant häufiger erkennenund entfernen als Bienen der HYG Linien. SMR Bie-nen scheinen Puppen mit sich besonders erfolgreichvermehrenden Milben nicht selektiv zu entfernen,sodass nicht, wie anfangs angenommen, nur Puppenmit wenig Nachkommen übrig bleiben würden. DerVersuch im Wärmeschrank zeigte, dass sich alleMilben auf der SMR Brut weniger erfolgreich ver-mehrten als die auf HYG Brut. Demnach könnte beiSMR Linien die Kombination aus Entfernung voneinem hohen Prozentsatz von infizierten Puppenzusammen mit der niedrigen Reproduktionsrate derMilben auf SMR Brut die niedrige Reproduktions-rate der Milben dieser SMR Völker erklären. ZurBestimmung der physiologische Wirkungsweise derSMR Brut auf die Vermehrungsrate der Milben undob die Eigenschaften von HYG und SMR durch die-selben Gene reguliert werden, sind weitere Versucheerforderlich.
Apis mellifera / Unterdrückung derMilbenreproduktion / hygienisches Verhalten /Varroa destructor
REFERENCES
Boecking O. (1992) Removal behavior of Apis mellif-era colonies towards sealed brood cells infestedwith Varroa jacobsoni: techniques, extent andefficacy, Apidologie 23, 371–373.
Boecking O., Spivak M. (1999) Behavioral defenses ofhoney bees against Varroa jacobsoni Oud., Apid-ologie 30, 141–158.
Corrêa-Marques M.H., Medina L.S., Martin S.J., DeJong D. (2003) Comparing data on the reproduc-tion of Varroa destructor, Genet. Mol. Res. 2, 1–6.
Delaplane K.S., Hood W.M. (1999) Economic thresh-old for Varroa jacobsoni Oud. in the southeasternUSA, Apidologie 30, 383–395.
Donzé G., Herrmann M., Bachofen B., Guerin P.M.(1996) Effect of mating frequency and brood cellinfestation rate on the reproductive success of thehoney bee parasite Varroa jacobsoni, Ecol. Ento-mol. 21, 17–26.
Gilliam M., Taber S., Richardson G. (1983) Hygienicbehavior of honey bee in relation to chalkbrooddisease, Apidologie 14, 29–39.
Gilliam M., Taber S., Lorenz B., Prest D. (1988) Fac-tors affecting development of chalkbrood diseasesin colonies of honey bees, Apis mellifera, fed pol-len contaminated with Ascosphaera apis, J. Inver-tebr. Pathol. 52, 314–325.
Harbo J.R., Harris J.W. (1999) Selecting honey beesfor resistance to Varroa jacobsoni, Apidologie 30,183–196.
Harbo J.R., Harris J.W. (2005) Suppressed mite repro-duction linked to the behavior of adult bees, J.Apic. Res. 44, 21–23.
Harbo J.R., Hoopingarner R. (1997) Honey bee(Hymenoptera: Apidae) in the United States thatexpress resistance to Varroa jacobsoni (Mesostig-mata: Varroidae), J. Econ. Entomol. 90, 893–898.
Harris J.W., Harbo J.R. (1999) Low sperm counts andreduced fecundity of mites in colonies of honeybees (Hymenoptera; Apidae) resistant to Varroajacobsoni (Mesostigmata: Varroidae), J. Econ.Entomol. 92, 83–88.
Ibrahim A., Spivak M. (2004) The relationshipbetween suppression of mite reproduction (SMR)and hygienic behavior, Am. Bee J. 144, 406.
Kraus B., Velthuis H. (1997) High humidity in thehoney bee (Apis mellifera L) brood nest limitsreproduction of the parasitic mite Varroa jacob-soni Oud., Naturwissenschaften 84, 217–218.
Le Conte Y., Arnold G., Desenfant P.H. (1990)Influence of brood temperature and hybrometryvariations on the development of the honey beeectoparasite Varroa jacobsoni (Mesostigmata:Varroidae), J. Econ. Entomol. 19, 1780–1785.
Macedo P., Ellis M. (2002) Using inert dusts to detectand access Varroa infestation in honey bee colo-nies, J. Apic. Res. 40, 3–7.
Martin S.J. (1994) Ontogenesis of the mite Varroajacobsoni Oud. in worker brood of honeybee Apismellifera L. under natural conditions, Exp. Appl.Acarol. 18, 78–100.
Masterman R., Ross R., Mesce K., Spivak M. (2001)Olfactory and behavioral response thresholds toodors of diseased brood differ between hygienicand non-hygienic honey bees (Apis mellifera L.),J. Comp. Physiol. A 187, 441–452.

40 A. Ibrahim, M. Spivak
McCulloch C., Searle S. (2002) Generalized, Linearand Mixed Models, Wiley, New York.
Medina L.M., Martin S.L. (1999) A comparative studyof Varroa jacobsoni reproduction in worker cellsof honey bees (Apis mellifera) in England andAfricanized bees in Yucatan, Mexico, Exp. Appl.Acarol. 23, 659–667.
Milani N. (1999) The resistance of Varroa jacobsoniOud. to acaridices, Apidologie 30, 229–234.
Mondragón L., Spivak M., Vandame R. (2005) A mul-tifactorial study of the resistance of Africanizedand hybrid honeybee Apis mellifera to the miteVarroa destructor over one year in Mexico, Api-dologie 36, 345–358.
Park O.W. (1937) Testing for resistance to Americanfoulbrood in honey bees, J. Econ. Entomol. 30,504–512.
Rothenbuhler W.C. (1964) Behavior genetics of nestcleaning in honey bees. IV. Responses of F1 andbackcross generations to disease-killed brood,Am. Zool. 4, 111–123.
Spivak M. (1996) Honey bee hygienic behavior anddefense against Varroa jacobsoni, Apidologie 27,245–260.
Spivak M., Downey D.L. (1998) Field assays for hygi-enic behavior in honey bees (Apidae: Hymenop-tera), J. Econ. Entomol. 91, 64–70.
Spivak M., Gilliam M. (1993) Facultative expressionof hygienic behavior of honey bees in relation todiseases resistance, J. Apic. Res. 32, 147–157.
Spivak M., Gilliam M. (1998) Hygienic behavior ofhoney bees and its application for control of brooddiseases and varroa, Bee World 79, 169–186.
Spivak M., Reuter G.S. (1998a) Performance of hygi-enic honey bee colonies in a commercial apiary,Apidologie 29, 291–302.
Spivak M., Reuter G.S. (1998b) Honey bee hygienicbehavior, Am. Bee J. 138, 283–286.
Spivak M., Reuter G.S. (2001a) Resistance to Ameri-can foulbrood diseases by honey bee colonies Apismellifera bred for hygienic behavior, Apidologie32, 555–565.
Spivak M., Reuter G.S. (2001b) Varroa destructorinfestation in untreated honey bee (Hymenoptera:Apidae) colonies selected for hygienic behavior,J. Econ. Entomol. 94, 326–331.
Spivak M., Masterman R., Ross R., Mesce K.A. (2003)Hygienic behavior in the honey bee (Apis mellif-era L.) and the modulatory role of octopamine, J.Neurobiol. 55, 341–354.
Woodrow A.W., Holst E.C. (1942) The mechanismsof colony resistance to American foulbrood, J.Econ. Entomol. 35, 327–330.
To access this journal online:www.edpsciences.org




















