
Glosario de las partes o piezas de las armas de fuego portátiles.
Upload
independentCategory
view
0download
0
International Journal of OsteoarchaeologyInt. J. Osteoarchaeol. (in press)Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/oa.828
The Oldest Human Remainsfrom the Beagle ChannelRegion, Tierra del Fuego
R. MACCHIARELLI,a* L. BONDIOLI,b S. CAROPRESO,b A. MAZURIER,a
G. MERCERONa AND E. L. PIANAc
a Laboratoire de Geobiologie, Biochronologie et Paleontologie Humaine, CNRS UMR 6046,
Universite de Poitiers, Franceb Sezione di Antropologia, Museo Nazionale Preistorico Etnografico ‘‘Luigi Pigorini’’, Rome,
Italyc Centro Austral de Investigaciones Cientıficas (CADIC), Ushuaia, Tierra del Fuego, Argentina
ABSTRACT The biohistory of the human peopling of the Fuegian Archipelago and the processes ofbiocultural adaptation and microdifferentiation of the ethnohistorical ‘canoe’ (‘sea nomad’)and ‘foot’ natives are poorly known. Here we report the oldest human remains of a ‘sea nomad’native discovered so far in the Beagle Channel region. The specimen consists of a deciduousmolar from the site of Imiwaia I, dated to 5870� 145 years BP. The microwear patternindicates the consumption of hard food processed by long chewing cycles. The histomor-phometric analysis shows the presence of at least three episodes of stress between 8 and10.5 months after birth, and an unusually thin neonatal line, suggesting a mechanically non-stressing birth event. Copyright � 2006 John Wiley & Sons, Ltd.
Key words: Tierra del Fuego; Beagle Channel; earliest peopling; Imiwaia; human tooth
Introduction
The native ‘canoe Indians’—the so-called ‘seanomads’—peopling the eastern and southernregions of the Fuegian Archipelago, who calledthemselves Yamana, were first encountered byEuropeans at the beginning of the 17th century.Their land, encompassing the south and westernshores of the Tierra del Fuego Main Island andmore than 200 smaller islands down to CapeHorn, is characterised by forest-covered moun-tains and sea channels, with a profusion ofislands, fjords, bays and inlets with abundantmarine resources, sea mammals and sea birds,
and an hyperoceanic cold climate with veryfrequent rain/snow fall (Hyades & Deniker,1891; Lothrop, 1928; Gusinde, 1939).
The ‘sea nomads’ attracted the curiosity of theEuropean scholars because of their ‘primitive’culture and harsh lifestyle, and were labelled asa key example of an ‘early stage of humankind’, of‘low hierarchical placement’ and of ‘cultural fixity’(for a review, see Orquera & Piana, 1996). Untilrecently, most knowledge of these ‘canoe Indians’was restricted to ethnohistorical information.Accordingly, they were depicted as a marginal,poorly adapted group, with little antiquity in thearea (approximately 2000 years), and confined tothe most distant part of the globe by culturallymore advanced people (Orquera & Piana, 1999a).
We report the oldest human remains of a ‘seanomad’ native discovered so far in the FuegianArchipelago. The remains consist of a deciduous
Copyright # 2006 John Wiley & Sons, Ltd. Received 14 November 2004Revised 1 June 2005
Accepted 15 June 2005
* Correspondence to: Laboratoire de Geobiologie, Biochronologieet Paleontologie Humaine, UMR 6046, Universite de Poitiers, 40,av. du Recteur Pineau, 86022 Poitiers Cedex, France.e-mail: [email protected]

molar from the site of Imiwaia I, dated to5870� 145 years BP (Orquera & Piana, 2000a).
The archaeological evidence
Systematic archaeological research in the BeagleChannel region began only in mid-1970s. Withthe first discoveries, most historically rootedassumptions proved to be wrong. The peoplingof the region is currently seen as the result of afully successful long-term tradition, the so-calledAdaptive Tradition to the Magellan-Fuegianchannels and islands (Piana, 1984, 2002). Thistechno-cultural tradition is documented in thearchaeological record from about 6400 years BP(uncal. 14C) to the 19th century AD. The ances-tors of the ethnohistorical ‘canoe’ natives had ahunting–fishing–gathering economy mostlybased on sea-mammal fat consumption. Whilesea lions were the most important dietary source,the diet was complemented with marine birds,fish, guanaco, and daily shellfish gathering(Orquera & Piana, 1999a).
Although Tierra del Fuego was one of the lastland masses to be populated (Borrero, 1977),human presence on the Main Island is over10000 years old (Massone, 1987) and certainlyprecedes the opening of the Magellan Strait,which occurred some 9000 years BP (Rabassaet al., 2000). Those earliest inhabitants had nospecific adaptation to the littoral environment. Infact, the oldest archaeological record from theBeagle Channel region, including the evidencefrom the almost 7000-year-old lowest layersof the Tunel I site (Orquera & Piana, 1995),testifies to a purely inland hunting–gatheringlifestyle.
The ‘Maximum Possible Antiquity’ hypoth-esis invokes the presence of extended woods inthe Fuegian islands as the essential requirementfor a fully sea littoral adaptation (Orquera &Piana, 1988). Accordingly, traces of this life-style older than 6700–6500 ca. years BP (uncal.14C) are not expected in the area because of thelate persistence of spread residual ice. Nearly athousand archaeological sites have been iden-tified in the Beagle Channel and surroundingarea, and some of them have been dated(Barcelo et al., 2002), but no evidence has
disproved the terminus ante quem predicted bythe palaeoecological model.
The first unambiguous evidence for a sea-nomadic adaptation comes from the sites ofAridos Guerrico (6495� 60 BP; Ocampo & Rivas,2000), Imiwaia I (6490� 120 BP on shells,5870� 145 BP on charcoal; Orquera & Piana,2000a), Tunel I Second Component (6400–5800BP; Orquera & Piana, 1999b), Grandi 1(6160� 100 BP, 6120� 80 BP; Legoupil, 1994,1995) and Lomada Alta Olivia (5600� 125 BP,unpublished)1 (Figure 1). The archaeologicalevidence displays a tool-kit fully developed forintensive use of the regional sea resources and theraw materials they provided.
Skeletal remains of sea-nomad ancestors fromdocumented contexts of the Beagle Channelregion and surrounding areas are very scarceand chronologically late compared with theavailable archaeological signals (Massone et al.,1985–86; Legoupil & Prieto, 1991; Perez-Perez,1996; Aspillaga et al., 1999).
The deciduous tooth from the Imiwaia I sitewas discovered in situ within a ring shell-middenadjacent to the base of a hut, dated 5870� 145years BP. So far, it represents the only humanremains from the middle Holocene sites listedabove (Macchiarelli & Piana, 2002).
Imiwaia I site
The Imiwaia I site (54�52.380S–67�17.750W), atbahıa Cambaceres Interior, is a field of intercon-nected annular and dome-like shell-middens builtup from very many short occupations. The baseof the excavated unit is 4.60m above currentsea-level, although when first settled the hut wasvery near the shore level. At that time, theenvironment was more open, with a much loweroccurrence of Nothofagus and a relative abundanceof gramineae and tubuliflora (Heusser, 1989).
Cultural items from the lower anthropogeniclayers (M and K) containing the human toothinclude cross-base toggling harpoon heads,multibarbed harpoon heads, wedges and chisels
1All uncalibrated 14C dates. Because of the so-called reservoir effect(Albero et al., 1988), dates on shells (e.g. Aridos Guerrico andImiwaia I) should be reduced by nearly 600 years.
R. Macchiarelli et al.
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

made from marine mammal bones, necklacebeads made with bird bones and shells, carvedbones, awls, and expeditive lithic technologyincluding side scrapers, used flakes, and stonefishing-line weights. These findings match thoseknown for the earliest phases of this adaptivetradition (Orquera & Piana, 1999b).
Faunal remains from the first excavationconsist of 2149 specimens (67% identified atgeneric level), representing pinnipeds (n¼ 633),various sea birds (n¼ 519), guanaco (n¼ 269),and other sea mammals (n¼ 17). The estimatedminimum number of individuals show seven furseals (Arctocephalus sp.), two guanacos (Lamaguanicoe), one large unidentified phocid, twocormorants (Phalacocrorax sp.), two albatross (Dio-medea sp.) and one penguin (Spheniscus magellanicus).Canine teeth suggest that all of the seals weremales, ranging from 4 to 12 years of age at death;three had probably been hunted in winter, two insummer, and one in summer–autumn (Orquera &Piana, 2000a). Most of the fish (103� 24 speci-mens per dm3) are hake and sardine, which todayenter the channels during summer, and occasion-ally strand.
The shell-midden also documents a huge con-sumption of shells (108� 25 individuals perdm3). Almost 91% of the mollusks are mussels,6% are limpets, and the remaining 3% representdifferent taxa of gastropods (Orquera & Piana,2000b, c).
The human tooth (IMI 384)
The human specimen from Imiwaia I (IMI 384) isan upper left deciduous second molar (m2).The crown is complete, but partially worn(Figure 2A). The lingual and mesiobuccal roots(height¼ 0.99 and 1.01mm, respectively) areremnants of the original trunks, while thedistobuccal root has been completely resorbed.The tooth was likely lost intra vitam whenthe individual was about 10 years old (usingUbelaker, 1978).
The occlusal cusp pattern is 4- (Dahlberg,1963), with slight hypocone reduction. Themesiobuccal cusp (paracone) bears a poorlydeveloped supernumerary cusplet. A distinctCarabelli’s cusp (degree 5; Turner et al., 1991),which is seen in varying frequencies in nativeAmericans (Scott & Turner, 1997), is present onthe lingual aspect. On its mesiobuccal aspect, amedium-sized parastyle (degree 3; Turner et al.,1991) is present.
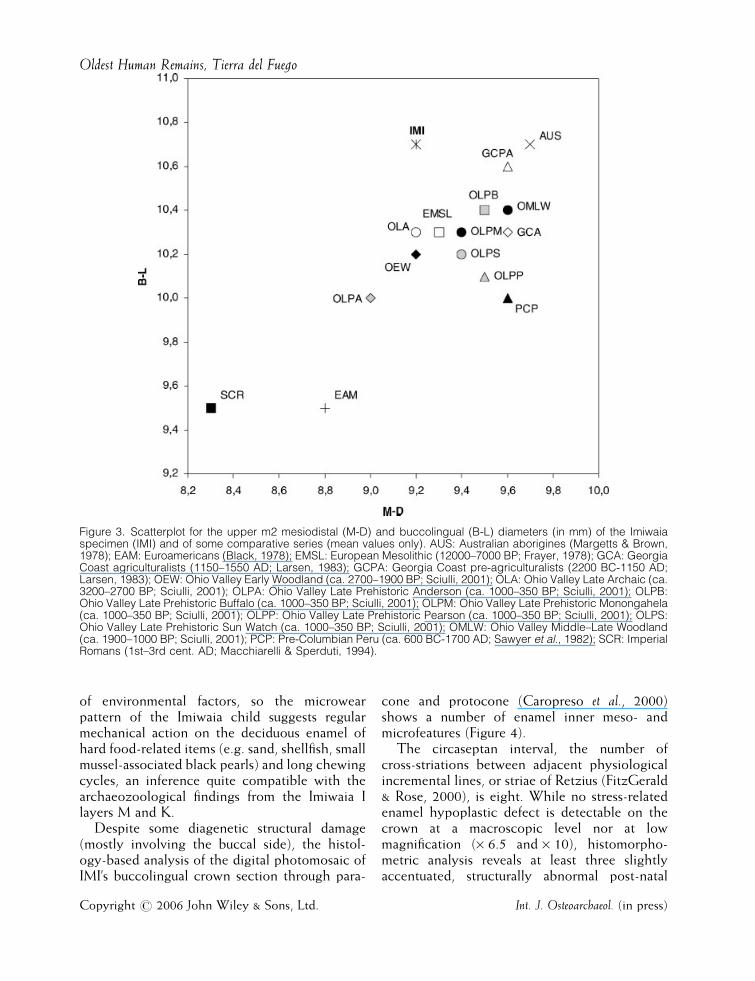
In proportion to its length (M-D, mesiodistaldiameter corrected for interproximal wear¼9.2mm), the Imiwaia tooth (IMI) shows a ratherbuccolingually expanded crown (B-L, buccolin-gual diameter¼ 10.7mm). This is apparent inFigure 3, a scatterplot showing decidious B-Land M-D tooth diameters from a variety ofarchaeological and recent samples (Black, 1978;Frayer, 1978; Margetts & Brown, 1978; Sawyer
Figure 1. The Fuegian Archipelago showing the archaeological sites documenting the earliest phases of humanadaptation to the littoral environment (sea-nomadic lifestyle according to the so-called Adaptive Tradition of theMagellan-Fuegian channels and islands). 1: Imiwaia I; 2: Tunel I; 3: Lomada Alta Olivia; 4: Aridos Guerrico; 5: Grandi I.See the text for chronology and references.
Oldest Human Remains, Tierra del Fuego
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

et al., 1982; Larsen, 1983; Macchiarelli & Sperduti,1994; Sciulli, 2001), including some nativeAmerican series (Sawyer et al., 1982; Larsen,1983; Sciulli, 2001).
No dental size values have been reported so farfor the deciduous dentition of the ethnohistoricalFuegians, nor for any southern Patagonian sample(Macchiarelli et al., 1988; Macchiarelli, 1996). Incross-sectional crown area, IMI (98.4mm2) over-laps or closely approximates the values of mostOhio Valley prehistoric samples (range:90.0–99.8mm2; calculated from Sciulli, 2001),those of the archaeological Georgia Coast agri-culturalists (99.1mm2; Larsen, 1983), and ofpre-Columbian Peruvians (96.0mm2; calculatedfrom Sawyer et al., 1982).
The mesial portion of IMI’s occlusal surface(Figure 2A), where an extended semilunar-shapeddentine patch occurs (length¼ 3.6mm; max.width¼ 1.0mm), is the most worn. The majorwear component is linguobuccally oriented. Twolarge interproximal wear facets are present(mesial: width¼ 4.4mm; height¼ 2.0mm; distal:4.7 and 2.6mm, respectively). Topographicalanalysis of the occlusal surface using scanning
electron microscopy and stereomicroscopyrevealed many large pits (>15mm) on thedistobuccal slope of the protocone (grindingfacet 9; Kay & Hiiemae, 1974) and on thedistolingual slope of the paracone (shearingfacet 3). On facet 9, 34 scratches, 51 pits (anysize), and 18 large pits exist on an investigatedsurface measuring 0.09mm2. Large scratches arealso found on the distolingual slope of the pro-tocone (shearing facet 6).
In their microwear analysis of the buccalaspect of permanent molars from a number ofprehistoric and recent population samples,Lalueza et al. (1996) included a group of ethno-historical Fuegians. Adults showed a prevalenceof long vertical scratches, a typical meat-eatingindicator. When measured similarly, IMI(Figure 2B) shows a nearly equal number ofvertical and horizontal scratches (n¼ 18 vs. 19,respectively), the latter being considerablylonger on average than the scratches found onthe adult counterparts (IMI¼ 240.5 mm; adultFuegian sample: 166.1� 60.2mm).
A high contribution of vegetable food inTierra del Fuego natives’ diet is unlikely because
Figure 2. IMI 384. A: Occlusal surface (scale bar is 2mm). B: Microfeatures (mostly scratches) on the buccal aspect(scale bar is 200mm).
R. Macchiarelli et al.
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)
of environmental factors, so the microwearpattern of the Imiwaia child suggests regularmechanical action on the deciduous enamel ofhard food-related items (e.g. sand, shellfish, smallmussel-associated black pearls) and long chewingcycles, an inference quite compatible with thearchaeozoological findings from the Imiwaia Ilayers M and K.
Despite some diagenetic structural damage(mostly involving the buccal side), the histol-ogy-based analysis of the digital photomosaic ofIMI’s buccolingual crown section through para-
cone and protocone (Caropreso et al., 2000)shows a number of enamel inner meso- andmicrofeatures (Figure 4).
The circaseptan interval, the number ofcross-striations between adjacent physiologicalincremental lines, or striae of Retzius (FitzGerald& Rose, 2000), is eight. While no stress-relatedenamel hypoplastic defect is detectable on thecrown at a macroscopic level nor at lowmagnification (� 6.5 and� 10), histomorpho-metric analysis reveals at least three slightlyaccentuated, structurally abnormal post-natal
Figure 3. Scatterplot for the upper m2 mesiodistal (M-D) and buccolingual (B-L) diameters (in mm) of the Imiwaiaspecimen (IMI) and of some comparative series (mean values only). AUS: Australian aborigines (Margetts & Brown,1978); EAM: Euroamericans (Black, 1978); EMSL: European Mesolithic (12000–7000 BP; Frayer, 1978); GCA: GeorgiaCoast agriculturalists (1150–1550 AD; Larsen, 1983); GCPA: Georgia Coast pre-agriculturalists (2200 BC-1150 AD;Larsen, 1983); OEW: Ohio Valley Early Woodland (ca. 2700–1900 BP; Sciulli, 2001); OLA: Ohio Valley Late Archaic (ca.3200–2700 BP; Sciulli, 2001); OLPA: Ohio Valley Late Prehistoric Anderson (ca. 1000–350 BP; Sciulli, 2001); OLPB:Ohio Valley Late Prehistoric Buffalo (ca. 1000–350 BP; Sciulli, 2001); OLPM: Ohio Valley Late Prehistoric Monongahela(ca. 1000–350 BP; Sciulli, 2001); OLPP: Ohio Valley Late Prehistoric Pearson (ca. 1000–350 BP; Sciulli, 2001); OLPS:Ohio Valley Late Prehistoric Sun Watch (ca. 1000–350 BP; Sciulli, 2001); OMLW: Ohio Valley Middle–Late Woodland(ca. 1900–1000 BP; Sciulli, 2001); PCP: Pre-Columbian Peru (ca. 600 BC-1700 AD; Sawyer et al., 1982); SCR: ImperialRomans (1st–3rd cent. AD; Macchiarelli & Sperduti, 1994).
Oldest Human Remains, Tierra del Fuego
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

striae of Retzius (Wilson & Shroff, 1970; Roseet al., 1985) (Figure 4A). The assessment of theoccurrence time of these three episodes of stress(FitzGerald et al., 1999) is about 8, 9.5 and 10.5months after birth, respectively.
The neonatal line, separating the enamelformed in intrauterine life from that formed afterbirth (Skinner & Dupras, 1993; for evidence inarchaeological population samples, see Smith &Avishai, 2005), is very thin (Figure 4B). Digitalmeasurements (n¼ 15 at� 400) taken at variousdistances from the cemento-enamel junctiongave an average thickness of 6.57mm (range:5.11–7.87mm). Comparative estimates obtainedby the same method on 124 deciduous crowns (ofall morphological classes) from 102 modernEuropean children (‘Fatina’ collection; originalunpublished values) range between 6.5 and50.9mm (mean¼ 17.31 mm, SD¼ 7.97), and asample of 109 children (aged 6 months–9 years)from the Imperial Roman cemetery of IsolaSacra (Bondioli & Macchiarelli, 1999; Rossiet al., 1999) showed a mean of 16.7mm(SD¼ 4.4, n crowns¼ 209).
Following Eli et al. (1989), normal deliveriesresult in smaller neonatal line thickness(10–17mm, n¼ 147 modern children), whileoperative or stressful deliveries are associated
with wider neonatal lines (13–24mm). Althoughthe ethnohistorical record should not be adopteduncritically as a model for the past, it should benoted that according to 18th–19th centurychronicles (review in Orquera & Piana, 1999a),Yamana women gave birth on average to 4–6babies (some women produced up to 14 babies,most of whom died prematurely). The practice ofimmersing newborns in the cold seawater of theBeagle Channel just after the delivery, and even acouple of times per day regularly for a period of atleast one month, was supposed to be a cause ofinfant mortality, but was also said to invigoratenewborns and to make them healthier.
When a strict relationship between neonatalline thickness and delivery dynamics is assumed(Eli et al., 1989), the IMI tooth shows an unusuallythin neonatal line, suggesting an extremely gentleshift to and fast recovery from intra- to extra-uterine life through a mechanically non-stressingbirth event, actually compatible with the resultsreported following caesarean sections.
Discussion and concluding remarks
The human biohistory of the Fuegian Archipe-lago and the processes of adaptation and
Figure 4. IMI 384. A: Histology-based digital photomosaic of the buccolingual crown section through paracone (pa)and protocone (pr) (scale bar is 1mm), showing the three accentuated striae of Retzius (ASR) detected on the post-natal enamel. B: Detail of the neonatal line (scale bar is 400 mm).
R. Macchiarelli et al.
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

microdifferentiation between the ‘canoe’ Indiansdepending on marine resources and the inland‘foot’ Indians exploiting guanaco have beenconsidered from many perspectives, includingmorphology (Rothhammer & Silva, 1990; Lahr,1995; Hernandez et al., 1997; Gonzalez-Jose et al.,2001), population genetics (Moraga et al., 2000;Goicoechea et al., 2001) and aDNA (Lalueza,1996; Lalueza et al., 1997; Garcıa-Bour et al.,2004). However, despite the growing quantityand quality of the anthropological record, it isnot yet possible to answer convincingly thequestion of where the close ancestors of theImiwaia child did originate.
Rooted in the ‘Maximum Possible Antiquity’hypothesis (Orquera & Piana, 1988), the archae-ological perspective strongly suggests that theinfant that lost the IMI 384 tooth crown was partof a stock of ‘sea nomad’ pioneers who, some6700–6500 years BP, entered the area alreadyculturally and technologically equipped andpre-adapted to a littoral environment, followingthe post-Pleistocene climatic changes thatallowed the expansion of woods as raw materialindispensable to sustain a sea-dependent life-style, and probably coming from the southPacific islands of present-day Chile (Orquera &Piana, 1988, 1999b; Legoupil, 1995; Ocampoet al., 1999; for alternative scenarios, see Lanata,2002).
The deciduous tooth from Imiwaia I is theoldest human specimen discovered so far in theBeagle Channel region. Because of its unequi-vocal archaeological context, the tooth clearlybelonged to a ‘sea nomad’ native. It currentlyconstitutes a single, isolated find whose mor-phological, dimensional and (micro)structuralfeatures cannot be directly compared with anyother coeval nor penecontemporary Fuegianspecimen. Although the nature of the morpho-logical and dimensional differences affecting thepermanent dentition of the ethnohistorical Fue-gians is still unknown (Macchiarelli, 1996), wepredict that the discovery of additional dentalmaterial from early and middle Holocene sites inTierra del Fuego will introduce new key ele-ments to the discussion of the biocultural adap-tation (including social behaviour) and thedynamics of peopling of this remote part ofthe world.
Acknowledgements
This research was supported by CONICET(Argentina), the Italian Ministry for ForeignAffairs, the Fondazione C.M. Lerici-Politecnicodi Milano, the Italian National Research Council(‘Cultural Heritage’ Project). Special thanks areowed to O. Zanola, Director of the Museo del Findel Mundo of Ushuaia, Argentina, where the castof IMI 384 is stored. C.M. FitzGerald (London),D.W. Frayer (Lawrence), A. Nava (Rome),F. Ramirez-Rozzi (Paris), and D.S. Weaver(Sandia Park), and two anonymous reviewerskindly provided useful critical comments to theearly draft.
References
Albero MC, Angiolin FE, Piana EL. 1988. Holocene14C reservoir effect at Beagle Channel (Tierra delFuego, Argentine Republic). Quaternary of SouthAmerica and Antartica Peninsula 5: 59–73.
Aspillaga E, Ocampo CE, Rivas PH. 1999. Restososeos humanos de contextos arqueologicos delarea de Isla de Navarino: indicadores de estilos devida en indıgenas canoeros. Anales del Instuto de laPatagonia 27: 123–135.
Barcelo JA, Piana EL, Martinioni DR. 2002. Archae-ological spatial modelling: a case study from BeagleChannel (Argentina). British Archaeological Reports1016: 351–360.
Black TK III. 1978. Sexual dimorphism in the tooth-crown diameters of the deciduous teeth. AmericanJournal of Physical Anthropology 48: 77–82.
Bondioli L, Macchiarelli R. 1999. Neonatal line thick-ness and delivery at Isola Sacra (2nd–3rd cent. AD,Rome, Italy). American Journal of Physical Anthropology(Suppl. 28): 94–95 (abstract).
Borrero LA. 1977. The origin of ethnographic sub-sistence patterns in Fuego-Patagonia. In Patagonia.Natural History, Prehistory and Ethnography at the Utter-most End of the Earth, MacEwan C, Borrero LA, PrietoA (eds). British Museum Press: London; 60–81.
Caropreso S, Bondioli L, Capannolo D, Cerroni L,Macchiarelli R, Condo SG. 2000. Thin sections forhard tissues histology: a new procedure. Journal ofMicroscopy 199: 244–247.
Dahlberg AA. 1963. Dental evolution and culture.Human Biology 35: 237–249.
Eli I, Sarnat H, Talmi E. 1989. Effect of the birthprocess on the neonatal line in primary toothenamel. Pediatric Dentistry 11: 220–223.
Oldest Human Remains, Tierra del Fuego
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

FitzGerald CM, Rose JC. 2000. Reading between thelines: dental development and subadult age assess-ment using the microstructural growth markers ofteeth. In Skeletal Biology of Past Peoples: Research Meth-ods, Katzenberg MA, Saunders SR (eds). JohnWiley& Sons: New York; 163–186.
FitzGerald CM, Saunders SR, Macchiarelli R, BondioliL. 1999. Large scale histological assessment ofdeciduous crown formation. In Dental Morphology1998, Mayhall JT, Heikkinen T (eds). OuluUniversity Press: Oulu; 92–101.
Frayer DW. 1978. Evolution of the Dentition in UpperPaleolithic and Mesolithic Europe. University of Kansas:Lawrence.
Garcıa-Bour J, Perez-Perez A, Alvarez S, Fernandez E,Lopez-Parra AM, Arroyo-Pardo E, Turbon D. 2004.Early population differentiation in extinct abori-gines from Tierra del Fuego-Patagonia: ancientmtDNA sequences and Y-chromosome STR char-acterization. American Journal of Physical Anthropology123: 361–370.
Goicoechea AS, Carnese FR, Dejean C, Avena SA,Weimer TA, Franco MHLP, Callegari-Jacques SM,Estalote AC, Simoes MLMS, Palatnik M, SalzanoFM. 2001. Genetic relationships between Amerin-dian populations of Argentina. American Journal ofPhysical Anthropology 115: 133–143.
Gonzalez-Jose R, Dahinten SL, Kuis MA, HernandezM, Pucciarelli HM. 2001. Craniometric variationand the settlement of the Americas: testing hypoth-eses by means of r-matrix and matrix correlationanalyses. American Journal of Physical Anthropology116: 154–165.
Gusinde M. 1939. Die Feuerland-Indianer. III/2. Anthro-pologie der Feuerland-Indianer. Modling: Vienna.
Hernandez M, Lalueza Fox C, Garcıa-Moro C. 1997.Fuegian cranial morphology: the adaptation to acold, harsh environment. American Journal of PhysicalAnthropology 103: 103–117.
Heusser CJ. 1989. Climate and chronology ofAntarctica and adjacent South America over thepast 30,000 yr. Palaeogeography PalaeoclimatologyPalaeoecology 76: 31–37.
Hyades P, Deniker J. 1891. Mission scientifique du CapHorn. 1882–1883. 7. Anthropologie, Ethnographie.Gauthier-Villars: Paris.
Kay RF, Hiiemae KM. 1974. Jaw movement and toothuse in recent and fossil primates. American Journal ofPhysical Anthropology 40: 227–256.
Lahr MM. 1995. Patterns of modern human diversifi-cation: implications for Amerindian origins. Year-book of Physical Anthropology 38: 163–198.
Lalueza C. 1996. Mitochondrial DNA haplogroups infour tribes from Tierra del Fuego-Patagonia: infer-
ences about the peopling of the Americas. HumanBiology 68: 855–871.
Lalueza C, Perez-Perez A, Turbon D. 1996. Dietaryinferences through buccal microwear analysis ofMiddle and Upper Pleistocene human fossils.American Journal of Physical Anthropology 100: 367–387.
Lalueza C, Perez-Perez A, Prats E, Cornudella L,Turbon D. 1997. Lack of founding Amerindianmitochondrial DNA lineages in extinct aboriginesfrom Tierra del Fuego-Patagonia. Human MolecularGenetics 6: 41–46.
Lanata JL. 2002. The world’s southernmost foragers:the native diversity of Tierra del Fuego. InArchaeological and Anthropological Perspectives on theNative Peoples of Pampa, Patagonia, and Tierra del Fuegoto the Nineteenth Century, Briones C, Lanata JL (eds).Bergin & Garvey: London; 57–73.
Larsen CS. 1983. Deciduous tooth size and subsis-tence change in prehistoric Georgian coast popula-tions. Current Anthropology 24: 225–226.
Legoupil D. 1994. El archipielago del Cabo deHornos y la costa sur de la isla Navarino: pobla-miento y modelos economicos. Anales del Instuto de laPatagonia 22: 101–121.
Legoupil D. 1995. Des indigenes au Cap Horn: con-quete d’un territoire et modele de peuplement auxconfins du continent sud-americain. Journal de laSociete des Americanistes 81: 9–45.
Legoupil D, Prieto A. 1991. Sepultura de ninos ca-noeros en un abrigo pintado en Ultima Esperanza,Chile. Anales del Instuto de la Patagonia 20: 133–138.
Lothrop SK. 1928. The Indians of Tierra del Fuego. HeyeFoundation: New York.
Macchiarelli R. 1996. The peopling of SouthernPatagonia and Tierra del Fuego: anthropologicalevidence. In 13th International Congress of Prehistoric andProtohistoric Sciences. The Sections 1. ABACO: Forli; 506(abstract).
Macchiarelli R, Corruccini RS, Bondioli L. 1988.Dental variation and oral health status in the Ona,Yahmana and Alakaluf (Tierra del Fuego). CollegiumAntropologicum 12: 217 (abstract).
Macchiarelli R, Piana EL. 2002. The earliestpeopling of the Beagle Channel region: humanpaleobiological evidence. Bullettins et Memoires de laSociete d’Anthropologie de Paris 14: 216–217(abstract).
Macchiarelli R, Sperduti A. 1994. Variazioni dimen-sionali delle corone dentarie umane dal Paleoliticosuperiore all’eta moderna in Italia. Bullettino diPaletnologia Italiana 85: 215–243.
Margetts B, Brown T. 1978. Crown diameters of thedeciduous teeth in Australian aborigines. AmericanJournal of Physical Anthropology 48: 493–502.
R. Macchiarelli et al.
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

Massone MM. 1987. Los cazadores paleoindios deTres Arroyos (Tierra del Fuego). Anales del Instuto dela Patagonia 17: 47–60.
Massone MM, Prieto AI, Cardenas PS. 1985–86.Contexto arqueologico de un enterratorioTehuelche excavado en la localidad de San Gre-gorio, Magallanes. Anales del Instuto de la Patagonia16: 95–101.
Moraga ML, Rocco P, Miquel JF, Nervi F, Llop E,Chakraborty R, Rothhammer F, Carvallo P. 2000.Mitichondrial DNA polymorphism in Chileanaboriginal populations: implications for the peo-pling of the southern cone of the continent.American Journal of Physical Anthropology 113:19–29.
Ocampo CE, Rivas HP. 2000. Nuevos fechados 14Cde la costa norte de la Isla Navarino, costa sur delCanal Beagle, Provincia Antartica Chilena, Regionde Magallanes. Anales del Instuto de la Patagonia 28:197–214.
Ocampo CE, Rivas HP, Aspillaga E. 1999. Pobla-miento temprano de los canales patagonicos: elnucleo ecotonal septentrional. Anales del Instuto de laPatagonia 27: 221–230.
Orquera LA, Piana EL. 1988. Human littoral adapta-tion in the Beagle Channel region: the maximumpossible age. Quaternary of South America and AntarcticPeninsula 5: 133–165.
Orquera LA, Piana EL. 1995. Tunel VII en la secuenciaarqueologica del canal Beagle: hipotesis y expecta-tivas de los investigadores argentinos. In Encuentrosen los Conchales Fueguinos, Estevez J, Vila Mitja A(eds). CSIC-UAB: Barcelona; 25–45.
Orquera LA, Piana EL. 1996. La imagen de loscanoeros magallanico-fueguinos: conceptos y ten-dencias. Runa 22: 187–245.
Orquera LA, Piana EL. 1999a. La Vida Social y Materialde los Yamana. Eudeba-IFIC: Buenos Aires.
Orquera LA, Piana EL. 1999b. Arqueologıa de la Region delCanal Beagle (Tierra del Fuego, Republica Argentina).Publ. Sociedad Argentina Antropologıa: BuenosAires.
Orquera LA, Piana EL. 2000a. Imiwaia I: un sitiode canoeros del sexto milenio A.P. en la costanorte del canal Beagle. In Desde el Paıs de losGigantes. Perspectivas Arqueologicas en Patagonia, BelardiJB, Carballo FC, Espinosa S (eds). UniversidadNacional de la Patagonia Austral: Rıo Gallegos;441–453.
Orquera LA, Piana EL. 2000b. Composicion de con-chales de la costa del Canal Beagle (Tierra delFuego, Republica Argentina). Primera parte.Relaciones de la Sociedad Argentina de Antropologıa 25:249–274.
Orquera LA, Piana EL. 2000c. Composicion de con-chales de la costa del Canal Beagle (Tierra delFuego, Republica Argentina). Segunda parte.Relaciones de la Sociedad Argentina de Antropologıa 26:345–368.
Perez-Perez A. 1996. Patologıa oral, indicadores deestres y dieta en una muestra esqueletica de abor-ıgenes de Tierra del Fuego. In Salud, Enfermedad yMuerte en el Pasado. Consecuencias Biologicas del Estres y laPatologıa, Perez-Perez A (ed.). Fundacion Uriach:Barcelona; 87–98.
Piana EL. 1984. Arrinconamiento o Adaptacion en Tierra delFuego. Editorial Universidad Belgrano: BuenosAires.
Piana EL. 2002. Magellan-Fueguian. In Encyclopedia ofPrehistory 7, Peregrine PN, Ember M (eds). YaleUniv. & Plenum Publ.: New York; 255–271.
Rabassa J, Coronato A, Bujalesky G, Salemme M, RoigC, Meglioli A, Heusser C, Gordillo S, Roig F,Borromei A, Quattrocchio M. 2000. Quaternaryof Tierra del Fuego, Southernmost South America:an updated review. Quaternary International 68–71:217–240.
Rose JC, Condon KW, Goodman AH. 1985. Diet anddentition: developmental disturbances. In TheAnalysis of Prehistoric Diets, Gilbert RI, Mielke J(eds). Academic Press: Orlando; 281–305.
Rossi PF, Bondioli L, Geusa G, Macchiarelli R. 1999.Osteodental Biology of the People of Portus Romae(Necropolis of Isola Sacra, 2nd–3rd Cent. AD). I.Enamel Microstructure and Developmental Defectsof the Primary Dentition. Digital Archives of HumanPaleobiology 1. Museo Nazionale ‘L. Pigorini’: Rome(CD-ROM).
Rothhammer F, Silva C. 1990. Craniometrical varia-tion among South American prehistoric popula-tions: climatic, altitudinal, chronological, andgeographic contributions. American Journal of PhysicalAnthropology 82: 9–17.
Sawyer DR, Allison MJ, Pezzia A, Mosadomi A. 1982.Crown dimensions of deciduous teeth frompre-Columbian Peru. American Journal of PhysicalAnthropology 59: 373–376.
Sciulli PW. 2001. Evolution of dentition in prehistoricOhio Valley native Americans III. Metrics of decid-uous dentition. American Journal of Physical Anthropol-ogy 116: 140–153.
Scott GR, Turner CG II. 1997. The Anthropology ofModern Human Teeth. Cambridge University Press:Cambridge.
Skinner MF, Dupras T. 1993. Variation in birthtiming and location of the neonatal line inhuman enamel. Journal of Forensic Sciences 38: 1383–1390.
Oldest Human Remains, Tierra del Fuego
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)

Smith P, Avishai G. 2005. The use of dental criteriafor estimating postnatal survival in skeletal remainsof infants. Journal of Archaeological Science 32: 83–89.
Turner CG II, Nichol CR, Scott GR. 1991. Scoringprocedures for key morphological traits of thepermanent dentition: the Arizona State UniversityDental Anthropology System. In Advances in Dental
Anthropology, Kelley MA, Larsen CS (eds). Wiley-Liss: New York; 13–31.
Ubelaker DH. 1978. Human Skeletal Remains. Excavation,Analysis, Interpretation. Aldine: Chicago.
Wilson PR, Shroff FR. 1970. The nature of the striaeof Retzius as seen with the optical microscope.Australian Dental Journal 15: 162–171.
R. Macchiarelli et al.
Copyright # 2006 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (in press)
Copyright © 2022 FDOKUMEN




















