
LifePort Kidney Transporter Operator's Manual 1.1 - Organ ...

The Na+ transporter AtHKT1;1 controls retrieval of Na+ fromthe xylem in Arabidopsis
ROMOLA JANE DAVENPORT1, ALICIA MUÑOZ-MAYOR2, DEEPA JHA3, PAULINE ADOBEA ESSAH1, ANA RUS4 &MARK TESTER3
1Department of Plant Sciences, Downing St, Cambridge CB2 3EA, UK, 2Centro de Edafología y Biología Aplicada delSegura-Consejo Superior de Investigaciones Científicas, Campus Universitario de Espinardo, 30100-Murcia, Spain,3Australian Centre for Plant Functional Genomics, Private Mail Bag 1,Glen Osmond, SA 5064, Australia and 4Centre forPlant Environmental Stress Physiology, Purdue University, West Lafayette, Indiana 47907-2010, USA
ABSTRACT
HKT-type transporters appear to play key roles in Na+
accumulation and salt sensitivity in plants. In ArabidopsisHKT1;1 has been proposed to influx Na+ into roots, recir-culate Na+ in the phloem and control root : shoot allocationof Na+. We tested these hypotheses using 22Na+ flux mea-surements and ion accumulation assays in an hkt1;1 mutantand demonstrated that AtHKT1;1 contributes to thecontrol of both root accumulation of Na+ and retrieval ofNa+ from the xylem, but is not involved in root influx orrecirculation in the phloem. Mathematical modelling indi-cated that the effects of the hkt1;1 mutation on root accu-mulation and xylem retrieval were independent. AlthoughAtHKT1;1 has been implicated in regulation of K+ trans-port and the hkt1;1 mutant showed altered net K+ accumu-lation, 86Rb+ uptake was unaffected by the hkt1;1 mutation.The hkt1;1 mutation has been shown previously to rescuegrowth of the sos1 mutant on low K+; however, HKT1;1knockout did not alter K+ or 86Rb+ accumulation in sos1.
Key-words: HKT1; salinity; salt tolerance; sodium transport.
INTRODUCTION
Salinization of soils represents a major threat to the main-tenance of crop yields, because most crop species are sen-sitive to the accumulation of sodium (Na+) in shoot tissues(Tester & Davenport 2003). One focus of salinity researchtherefore is the mechanisms by which plants accumulateNa+, a non-essential ion. Figure 1 shows the main pathwayspresumed to operate in the accumulation of Na+ in shoottissues. Na+ enters roots passively, via non-selective cationchannels and possibly other Na+ transporters such as HKTs(Amtmann & Sanders, 1999; Tester & Davenport 2003;Haro et al. 2005). It is likely that most of the Na+ that entersroot cells in the outer part of the root is pumped back outagain via plasma membrane Na+/H+ antiporters (Tester &Davenport 2003), and such a function has been
demonstrated for the Arabidopsis protein SOS1 at the rootapex (Qiu et al. 2003). Na+ remaining in the root can besequestered in vacuoles or transported to the shoot. Com-partmentation in vacuoles is achieved by tonoplast Na+/H+
antiporters such as those belonging to the NHX family inArabidopsis. Membranes exhibit bidirectional transport,and there is passive leakage of Na+ back to the cytosolfrom vacuoles (possibly via tonoplast non-selective cationchannels), requiring constant re-sequestration of Na+ intovacuoles. Overexpression of NHX1 or the tonoplast pyro-phosphatase AVP1 that contributes to the H+ gradient forNa+ storage in the vacuole increases both Na+ accumulationand Na+ tolerance in Arabidopsis, suggesting that moreefficient sequestration may improve osmotic toleranceand/or reduce cytosolic Na+ levels (Apse et al. 1999;Gaxiola et al. 2001).
Transport of Na+ to the shoot is poorly understood. Na+
moves in the symplast across the endodermis (where it isintact) and is released from mainly xylem parenchyma cellsinto the apoplastic xylem. The electrochemical gradient forNa+ release into the xylem is likely to vary and may evenfavour passive release under conditions where the xylemis relatively dilute (during high transpiration) and cellsloading the xylem are depolarized (as is possible in salineconditions). Passive release of Na+ to the xylem can involvenon-selective cation channels and/or other uniport mecha-nisms. Under conditions of low transpiration or in matureparts of the root where Na+ has accumulated along thexylem pathway, Na+ efflux to the xylem may require activetransport.The Na+/H+ antiporter SOS1 is expressed in stelarcells and has been implicated in active Na+ loading of thexylem. Re-uptake from the xylem back into root cells mayalso occur, and this would reduce Na+ delivery to the shoot.In this case, the thermodynamic conditions would bereversed, and Na+ could be retrieved passively in matureparts of the root where xylem Na+ is at high concentration.SOS1 has also been proposed to act in xylem retrieval (Shiet al. 2002) but requires some stoichiometric plasticity toovercome the H+ gradient favouring Na+ efflux (Tester &Davenport 2003).
The rate of delivery to the shoot depends on root Na+
transport processes and, to some extent, on the rate of
Correspondence: Romola J. Davenport. Oxford Institute of Ageing,Manor Rd Building, Manor Rd, Oxford, OX1 3UQ, UK. Fax: +441865 286191; e-mail: [email protected]
Plant, Cell and Environment (2007) 30, 497–507 doi: 10.1111/j.1365-3040.2007.01637.x
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd 497

transpiration (if xylem loading rates are influenced byxylem Na+ concentration, as previously suggested). Oncedelivered to the shoot in the xylem, Na+ is accumulated inshoot cells probably by processes similar to those describedin roots.A final factor affecting shoot concentrations of Na+,apart from the shoot growth rate, is the rate of recirculationof Na+ to the roots. Recirculation can occur via the phloem,but the contribution of recirculation to Na+ homeostasisremains in question. Little is known about the thermody-namics of Na+ loading and unloading of the phloem, butAtHKT1;1 has been proposed to function in both processes(Berthomieu et al. 2003).
Despite decades of intensive research, the kinetics of Na+
transport remains poorly understood, and the proteinscatalysing Na+ transport are largely unknown. However,research in a number of species is now converging onHKT-type proteins as major controllers of Na+ transportto the shoot. Here we address the role of AtHKT1;1 inArabidopsis.
The HKT family of proteins has been implicated in Na+
transport in a number of species, and members of the familyhave been demonstrated to function as Na+/K+ symportersand as Na+-selective transporters of both high and low affin-ity (Rubio, Gassmann & Schroeder 1995; Garciadeblás et al.2003; Haro et al. 2005). The wheat TaHKT2;1 functions asa Na+/K+ symporter when expressed in Xenopus oocytes(Rubio et al. 1995), and down-regulation of expression inplanta reduces root Na+ accumulation and improves growthin saline conditions (Laurie et al. 2002). The rice OsHKT1;5
functions as a Na+-selective transporter in Xenopus oocytesand is hypothesized to control shoot Na+ and influenceshoot K+ by withdrawing Na+ from the xylem stream intothe xylem parenchyma cells (Ren et al. 2005). The Arabi-dopsis genome contains a single HKT homolog, AtHKT1;1.AtHKT1;1 appears to function as a Na+-selective uniporterwhen expressed in Xenopus oocytes and yeast, but it alsocomplements an Escherichia coli K+ uptake-deficientmutant and increases its K+ accumulation, suggesting somerole in K+ transport (Uozumi et al. 2000). hkt1;1 mutants aresalt sensitive compared to wild type and hyperaccumulateNa+ in the shoot, but show hypoaccumulation of Na+ in theroot (Maser et al. 2002; Berthomieu et al. 2003; Rus et al.2004).
In addition to the evidence for a direct role of AtHKT1;1in Na+ transport in Arabidopsis, there is also evidence thatAtHKT1;1 plays a more complex role in controlling ionfluxes. Although hkt1;1 knockout plants are more salt sen-sitive than wild type, knockout of AtHKT1;1 partiallyrestores salt resistance in the salt overly sensitive (sos)mutants (sos1, sos2, sos3) (Rus et al. 2001, 2004). The SOS2and SOS3 proteins function in a signalling pathway, one ofthe outputs of which is activation of SOS1, a plasma mem-brane Na+/H+ antiporter. SOS1 is transcribed (at least innon-saline conditions) in the root tip and vasculature, and ishypothesized to efflux Na+ from roots to the external solu-tion and to pump Na+ into or retrieve Na+ from the xylem,depending on Na+ and H+ gradients in the stele (Shi et al.2002). sos1, 2 and 3 mutants are more salt sensitive thanwild type and show hypo- or hyperaccumulation of Na+
depending on growth conditions (Wu, Ding & Zhu 1996;Ding & Zhu 1997; Shi et al. 2002), and inhibition of rootgrowth in K+ starvation conditions (Wu et al. 1996; Rus et al.2004). Interestingly, knockout of AtHKT1;1 rescues boththe Na+ and K+ sensitivity phenotypes (Rus et al. 2001,2004).
Several hypotheses have been advanced concerningAtHKT1;1 function in Arabidopsis. Because hkt1;1 muta-tions ameliorate the sos phenotypes and reduce whole seed-ling Na+ in sos3, Rus et al. (2001) proposed that AtHKT1;1was an influx pathway for Na+ uptake into the root.However, Berthomieu et al. (2003) and Essah, Davenport &Tester (2003) showed that hkt1;1 mutants did not havelower root Na+ influx (in excised roots), and Berthomieuet al. (2003) proposed instead that AtHKT1;1 functioned inNa+ recirculation from shoots to roots by loading Na+ fromthe shoot into phloem and then unloading it into the rootsfor efflux. This hypothesis accounted for the shoot Na+
hyperaccumulation and reduced root Na+ observed inhkt1;1 mutants, and was supported by evidence for reducedphloem Na+ content in the hkt1;1 (sas2) mutant andapparent phloem localization of AtHKT1;1 transcripts(Berthomieu et al. 2003). Subsequently, Rus et al. (2004)demonstrated that plants overexpressing AtHKT1;1 underthe control of the putative native AtHKT1;1 promoter werealso salt sensitive, and argued that AtHKT1;1 plays a role inregulation of K+ and Na+ homeostasis that is disrupted byknockout or misexpression. Most recently, Sunarpi et al.
Na+
H+
Shoot
Cortex
Na+
Xylem
Vacuole Cytosol
Stele
Na+
H+
Na+
H+ Na+Na+
Na+
1
2
3
4
5
6
7
8
9
Figure 1. A simple model of the unidirectional Na+ fluxescontributing to shoot net accumulation of Na+. In high NaClconditions, Na+ enters the root cytosol passively, probably mainlyvia non-selective cation channels (1). Na+ in the outer part of theroot may be pumped back to the soil via antiporters such asSOS1 (2) or compartmentalized in the vacuole by antiporterssuch as the tonoplast NHX transporters (3). Na+ also leakspassively from the vacuole back to the root cytosol (4). In thestele, Na+ is loaded into the xylem: it is likely that this occurspassively under at least some conditions (5) and also actively,possibly via SOS1 (6), in the mature parts of the root or underconditions of low transpiration. Retrieval from the xylem canalso occur via these processes (5 & 6). Na+ is probably unloadedpassively into leaf cells (7), and can be recirculated from leavesto roots via the phloem (8 & 9).
498 R. J. Davenport et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507

(2005) demonstrated that AtHKT1;1 was localized to theplasma membrane of xylem parenchyma cells in the shoot.They found both reduced phloem Na+ and elevated xylemNa+ in the shoot of hkt1;1 mutants and proposed thatAtHKT1;1 functioned primarily to retrieve Na+ from thexylem, at least in the shoot, and that retrieval of Na+ into thesymplast had a knock-on effect on phloem Na+ levels.
Hypotheses regarding the role of AtHKT1;1 in Na+ trans-port have relied mainly on measurements of tissue ion con-tents, which are the net result of a number of differenttransport processes (Fig. 1), or on disruptive measurementsof phloem and xylem contents. In addition, many of theprevious experiments have been conducted in plants grownon agar plates (where transpiration is extremely limited) orin soil (where the activity of nutrient and toxic ions is dif-ficult to control), and this makes it difficult to compareresults between experiments. For the present study, wedeveloped a hydroponic growth system and minimal mediathat would allow us to perform a range of physiologicalexperiments under standardized growth conditions, so thatwe could be confident that a biomass or ion accumulationphenotype observed in a growth experiment could bereproduced in plants grown for short-term flux experi-ments. We used radioactive tracers to dissect the individualtransport processes contributing to Na+ and K+ accumula-tion in intact, transpiring plants in order to test whetherAtHKT1;1 was involved in (1) root Na+ influx, (2) recircu-lation from shoot to root, (3) Na+ retrieval from the xylemand (4) K+ influx.
MATERIALS AND METHODS
Plant materials used were EMS-mutagenized sos1-1 (Shiet al. 2002:) kindly provided by J.-K. Zhu), T-DNA-mutagenized knockout mutant hkt1-4, sos1-1 hkt1-1 andgl1-pCAM (gl1) control plants (Rus et al. 2004). The gl1plants contained an empty pCAMBIA 2300 vector andwere used to replicate the genotypes used by Rus et al.(2004) in their systematic analysis of hkt1;1-sos1,2,3 inter-actions. For hydroponic experiments, seeds were sown onagar in 1.5 mL Eppendorf tubes (Eppendorf UK, Cam-bridge, UK), and then the bottom of each tube was cut offand the tubes suspended over aerated solution containing1.25 mM KNO3, 0.625 mM KH2PO4, 0.5 mM MgSO4,0.5 mM Ca(NO3)2, 0.045 mM FeNaEDTA and micronutri-ents (Arteca & Arteca 2000) (solution 1). The agar con-tained half-strength solution 1. The plants were maintainedin aerated solutions on an 8/16 h light/dark cycle, 21 °C, withan irradiance of 70 mmol m-2 s-1. For experiments thatincluded sos1, all genotypes were grown in solution 1 plus10 mM KCl before experimental treatments. For highNaCl experiments, all plants were grown for 5–7 d beforeexperiments in 50 mM NaCl + solution 1, and 22Na+ uptakewas measured in this solution except where Ca2+ activitywas modified by replacement of Ca(NO3)2 with NaNO3
and CaCl2. Rus et al. (2001, 2004) showed that rescue ofthe sos phenotypes in saline conditions by hkt1;1 muta-tions was more successful at higher Ca2+. Therefore, Ca2+
concentrations were manipulated to investigate whetherHKT1 function was Ca2+ dependent, using very low(0.03 mM) Ca2+ activity and a high Ca2+ activity (3 mM) atwhich effects of Ca2+ on Na+ influx were known to saturate(Essah et al. 2003).Where Ca2+ effects were not under inves-tigation, 0.5 mM Ca2+ concentration was used to duplicateprevious work. To test effects of high NH4
+, the plants weregrown in a modified version of the high NH4
+ solutionspreviously used to phenotype the sos mutants: 18.8 mMNH4NO3, 1.25 mM K2HPO4, 0.5 mM MgSO4, 0.5 mM CaCl2,0.045 mM FeNaEDTA and micronutrients (solution 2). ForK+ starvation experiments, K+ was excluded by replacingKNO3 and KH2PO4 with Na or NH4 salts, and 0.02 mMRbCl was included. The plants were maintained in solution1 or 2 for 5 d and then K+ starved for 12–24 h before experi-ments. To maximize K+ uptake, CaCl2 was reduced to0.05 mM and Ca(NO3)2 replaced (for solution 1) by NaNO3
or NH4NO3. For agar plate experiments, seeds were sownon vertical plates containing solution 1 or 2 with 1% bac-toagar, with modified K, Na and Ca as described earlier, and3% sucrose where indicated.
Unidirectional fluxes of Na+ and Rb+ were measuredusing radioisotopes 22Na+ and 86Rb+ (Amersham, Amer-sham, UK). The plants were pre-treated for 10 min in unla-belled solution of the same composition as the labelledexperimental solution. Whole seedlings were suspendedwith their roots in 80 mL labelled solution on a gently rotat-ing shaker. At the end of the experiment, the roots wererinsed for 2 ¥ 1.5 min in ice-cold rinse solution of the samecomposition as the uptake solution plus 10 mM CaCl2, andthen roots and shoots were separated, blotted and weighed.Samples were mixed with scintillation cocktail (OptiphaseHiSafe; Fisher Chemicals, Loughborough, UK) and countedwith a liquid scintillation counter (Beckman Instruments,Fullerton, CA, USA). For measurements of net ion uptake,the plants were grown hydroponically and separated intoroot and shoot at harvest, rinsed in deionized water,weighed and then dried at 60 °C for 2 d and dry weight(DW) determined. Tissue was digested for 5 h at 120 °C in1% nitric acid and Na and K concentrations measured usinga flame photometer (model 420; Sherwood Scientific, Cam-bridge, UK). Calcium activities were calculated using VisualMinteq version 2.30 (US Environmental ProtectionAgency; Environmental Research Laboratory, Athens,Georgia, USA), and mathematical modelling was per-formed with ModelMaker 4.0 (Cherwell Scientific,Oxford, UK).
RESULTS
Na+ and K+ accumulation in hydroponicallygrown plants
Control and mutant genotypes (hkt1-4, sos1-1, sos1-1-hkt1-1) were grown hydroponically and exposed to 50 mMNaCl or control solution for 10 d. Root growth in 50 mMNaCl was strongly reduced in sos1, and this inhibition waspartially relieved in the sos1-hkt1;1 double mutant (Fig. 2a).
AtHKT1;1 controls retrieval of Na+ from the xylem 499
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507

Although sos1 shoots showed visible symptoms of stress,shoot growth (DW) was relatively unaffected by 10 d NaCltreatment in all genotypes (Fig. 2b) and was unrelated toshoot Na+, as all mutant genotypes accumulated approxi-mately threefold more Na+ in the shoot than the parentalcontrol line (Fig. 2c). In the roots, hkt1;1 plants accumulatedless Na+ than wild type (Fig. 2c), as observed in previousstudies. sos1 plants accumulated a threefold higher root Na+
concentration than wild type, and this hyperaccumulationwas reduced to a twofold excess in sos1-hkt1;1 plants, sug-gesting that the salt sensitivity of sos1 mutants may be dueto excessive root Na+ accumulation that is partially reducedby knockout of AtHKT1;1. Salt sensitivity was not relatedto tissue K+ content. Shoot K+ accumulation (Fig. 2d) andtotal plant K+ accumulation was highest in sos1 and sos1-hkt1;1 plants, indicating that, at least in our conditions,excess Na+ uptake did not inhibit K+ acquisition in sos1, assuggested by Qi & Spalding (2004). hkt1;1 plants accumu-lated slightly more root K+ than controls; however, root K+
of both sos1 and sos1-hkt1;1 plants was 50% lower thancontrols, indicating that knockout of AtHKT1;1 did notrescue sos1 root growth via enhancement of K+ accumula-tion (Fig. 2d).
Transport of Na+ and K+ (Rb+) fits athree-compartment model in Arabidopsis
To characterize the transport processes that underlie rootand shoot accumulation of K+ and Na+, we first measured
time courses of Na+ and Rb+ uptake. Na+ uptake into Ara-bidopsis roots resembled typical measurements of cationuptake into excised roots and storage tissues. We thereforefitted a three-compartment model comprising the rootcytosol, root vacuole and the shoot (as previously describedin wheat, Davenport et al. 2005). The model was based onthe classical ‘four-compartment’ model used to describefluxes in algal cells and in roots with a xylem compartment,where uptake into an initial root compartment is followedby uptake in parallel into two compartments (in our modelthe vacuole and shoot) (Walker & Pitman 1976). The initiallinear phase of uptake was attributed to unidirectional Na+
influx across the plasma membrane of root cells, resulting inrapid labelling of the root cytosol.The second, slower linearphase of uptake was attributed to influx across the tono-plast to root vacuoles (which assumed a constant rate oncethe cytosol was approximately fully radioactively labelled).This model assumes that tonoplast fluxes of Na+ were muchslower than fluxes across the plasma membrane, as is com-monly assumed for plant cells. Alternatively the secondlinear phase could represent a second compartment withinthe root, such as the stele. We think that this is unlikely,given the relative sizes of the first and second compart-ments and the kinetics of uptake to the shoot. The first,rapidly saturating compartment was clearly small in volumecompared to the slower compartment, which continued tofill over many hours (Fig. 3b). It is most likely that the firstcompartment represents the cytosol, or some portion of thecytosol that includes a stelar component, because it appearsthat Na+ transported to the shoot derived from thiscompartment.
Transport of Na+ into the shoot occurred with a lag coin-ciding with the period required for labelling of the firstcompartment (Fig. 3a). Shoot uptake subsequently became
gl1 hkt1;1 sos1 sos1-hkt1;10.0
0.5
1.0
1.5
2.050 mM NaCl
Control
Tis
su
e D
W (
mg
)
gl1 hkt1;1 sos1 sos1-hkt1;10
2
4
6
8
gl1 hkt1;1 sos1 sos1-hkt1;10
50
100
150
200Root Shoot
Tis
su
e io
n (
mM
)
gl1 hkt1;1 sos1 sos1-hkt1;10
50
100
150
200
(a) Root DW (b) Shoot DW
(c) Root and shoot Na+ (d) Root and shoot K+
Figure 2. Growth and ion accumulation of 6-week-oldhydroponically grown plants maintained in control medium(solution 1) or exposed to 50 mM NaCl for 10 d beforemeasurements. (a & b) Tissue dry weights (DW) in control andNaCl-treated plants. (c & d) Tissue ion concentrations inNaCl-treated plants. Data represent mean � SEM, n = 5. SEM,standard error of the mean.
0 20 40 60 80 100
0
2
4
6
8
10Root
Shoot
t (min)
Na
+ u
pta
ke
(mm
ol g
–1 ro
ot
FW
)
0 5 10 15 20
0
5
10
15
Root
Shoot
t (h)
(a) (b)
Figure 3. Kinetics of Na+ uptake in control plants, measured asuptake of 22Na+. (a) Time course of Na+ uptake to root and shoot(50 mM NaCl, 0.03 mM Ca2+ activity). The second linear phase ofroot uptake was fitted to give y = 0.078 mmol g-1 root fresh weight(FW) min-1 + 1.95 mmol g-1. Shoot uptake occurred with a lag of10 min and was linear between 20 and 100 min, fitted with liney = 0.058 mmol g-1 root FW min-1 - 0.97 mmol g-1. (b) Na+ uptakeover 24 h was approximately linear in both roots and shoots (Na+
uptake measured in 50 mM NaCl, 0.2 mM Ca2+ activity, so uptakerates are not directly comparable between a & b). Data pointsrepresent mean � SEM, n = 4. SEM, standard error of the mean.
500 R. J. Davenport et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507
linear (indicating full labelling of the compartment fromwhich shoot loading occurred) and approximated the rateof unidirectional transfer from root to shoot. Root andshoot uptake continued for at least 24 h with no evidence ofsaturation (Fig. 3b). The lag and then linearity in shootuptake indicate that the first, rapidly exchanging compart-ment comprised the symplast from which xylem loadingoccurred, indicating that this compartment included a stelarcomponent. The data also suggest that the second phase ofroot uptake did not contribute to shoot uptake, consistentwith a vacuolar identity. A similar phenomenon wasobserved for Na+ uptake in Spergularia marina (Lazof &Cheeseman 1986) and wheat (Davenport et al. 2005). It is ofcourse possible that the apparently simple kinetics ofuptake represented the summed contribution of many het-erogeneous compartments; however, we think that therewas a clear functional difference between the first ‘compart-ment’ that was in contact with the external medium andcontributed to shoot uptake, and the second ‘compartment’that did not have these characteristics. A similar modelcould describe 86Rb+ uptake in K+-starved plants (Supple-mentary Fig. S1).
AtHKT1;1 is not a Na+ influx mechanism butaffects root vacuolar Na+ uptake
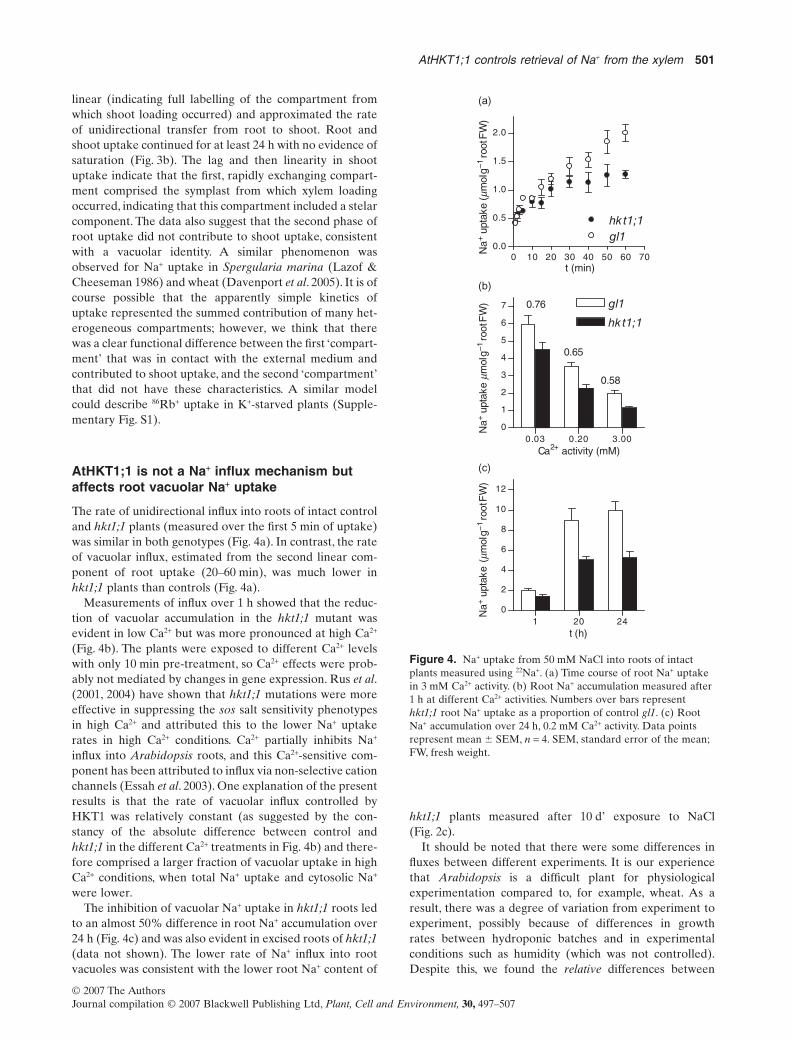
The rate of unidirectional influx into roots of intact controland hkt1;1 plants (measured over the first 5 min of uptake)was similar in both genotypes (Fig. 4a). In contrast, the rateof vacuolar influx, estimated from the second linear com-ponent of root uptake (20–60 min), was much lower inhkt1;1 plants than controls (Fig. 4a).
Measurements of influx over 1 h showed that the reduc-tion of vacuolar accumulation in the hkt1;1 mutant wasevident in low Ca2+ but was more pronounced at high Ca2+
(Fig. 4b). The plants were exposed to different Ca2+ levelswith only 10 min pre-treatment, so Ca2+ effects were prob-ably not mediated by changes in gene expression. Rus et al.(2001, 2004) have shown that hkt1;1 mutations were moreeffective in suppressing the sos salt sensitivity phenotypesin high Ca2+ and attributed this to the lower Na+ uptakerates in high Ca2+ conditions. Ca2+ partially inhibits Na+
influx into Arabidopsis roots, and this Ca2+-sensitive com-ponent has been attributed to influx via non-selective cationchannels (Essah et al. 2003). One explanation of the presentresults is that the rate of vacuolar influx controlled byHKT1 was relatively constant (as suggested by the con-stancy of the absolute difference between control andhkt1;1 in the different Ca2+ treatments in Fig. 4b) and there-fore comprised a larger fraction of vacuolar uptake in highCa2+ conditions, when total Na+ uptake and cytosolic Na+
were lower.The inhibition of vacuolar Na+ uptake in hkt1;1 roots led
to an almost 50% difference in root Na+ accumulation over24 h (Fig. 4c) and was also evident in excised roots of hkt1;1(data not shown). The lower rate of Na+ influx into rootvacuoles was consistent with the lower root Na+ content of
hkt1;1 plants measured after 10 d’ exposure to NaCl(Fig. 2c).
It should be noted that there were some differences influxes between different experiments. It is our experiencethat Arabidopsis is a difficult plant for physiologicalexperimentation compared to, for example, wheat. As aresult, there was a degree of variation from experiment toexperiment, possibly because of differences in growthrates between hydroponic batches and in experimentalconditions such as humidity (which was not controlled).Despite this, we found the relative differences between
0 10 20 30 40 50 60 700.0
0.5
1.0
1.5
2.0
hk t1;1gl1
t (min)
Na
+ u
pta
ke (mm
ol g
–1
root F
W)
0 .03 0.20 3.00
0
1
2
3
4
5
6
7 gl1
hk t1;1
0.76
0.58
0.65
Ca2+ activity (mM)
Na
+ u
pta
ke m
mol g
–1
root F
W)
1 20 24
0
2
4
6
8
10
12
t (h)
Na
+ u
pta
ke
(mm
ol g
–1
root F
W)
(a)
(b)
(c)
Figure 4. Na+ uptake from 50 mM NaCl into roots of intactplants measured using 22Na+. (a) Time course of root Na+ uptakein 3 mM Ca2+ activity. (b) Root Na+ accumulation measured after1 h at different Ca2+ activities. Numbers over bars representhkt1;1 root Na+ uptake as a proportion of control gl1. (c) RootNa+ accumulation over 24 h, 0.2 mM Ca2+ activity. Data pointsrepresent mean � SEM, n = 4. SEM, standard error of the mean;FW, fresh weight.
AtHKT1;1 controls retrieval of Na+ from the xylem 501
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507

hkt1;1 mutants and controls to be highly reproducible,although the absolute rates of transport varied betweenexperiments.
AtHKT1;1 affects xylem transport of Na+,not recirculation
The unidirectional flux of 22Na+ to the shoot was much morerapid in hkt1;1 plants than controls (Fig. 5a). Because thekinetics of initial root influx and saturation of the cytosolicphase of 22Na+ accumulation appeared similar in both hkt1;1and control (Fig. 4a), it was assumed that the specific activ-ity of Na+ loaded into the xylem (from the cytosolic phase)was similar in both genotypes, and therefore that the mea-sured fluxes represented actual rates of Na+ translocation.hkt1;1 plants accumulated approximately five times moreNa+ in the shoot within 1 h, and this difference was main-tained and did not increase significantly over 24 h (Fig. 5b),indicating that shoot Na+ hyperaccumulation in hkt1;1 wasthe result of processes occurring early in the pathway of Na+
uptake. The difference in the rate of unidirectional trans-port of Na+ to the shoot between genotypes was sufficientto explain the differences in shoot Na+ content (Fig. 2c),without recourse to explanations involving recirculation.
To test whether recirculation could contribute to controlof shoot Na+ concentration, the plants were loaded for 40 hwith 22Na+ and were then transferred to unlabelled solutionof the same composition and sampled over 52 h. Shoot22Na+ content declined by only 13% over 52 h in the controlplants (although this difference was statistically not signifi-cant), and showed little change in hkt1;1 plants (Fig. 5c).These results suggest that recirculation, if it occurs at all,does not significantly affect total shoot Na+ accumulation inArabidopsis. Certainly, the elimination of a possible recir-culation rate of 5–10% per day in hkt1;1 plants couldnot account for the fivefold difference observed in shootNa+ accumulation between hkt1;1 and control plants over40 h of labelling. Root 22Na+ content was low compared toshoot 22Na+ and did not change after 24 h of rinsing; there-fore, translocation of 22Na+ from the roots to the shootduring the unlabelled rinse period could not have maskedretranslocation.
AtHKT1;1 affects Na+ retrieval from the xylem
The rate of Na+ delivery to the shoot depends on the rate ofefflux of Na+ into the xylem from stelar cells, the rate ofwithdrawal of Na+ from the transpiration stream back intoroot stelar cells, and the transpiration rate. The hkt1;1mutant sas2 has been shown not to differ from controlplants in transpiration rate under a range of humidity (Ber-thomieu et al. 2003). When only the apical portion of theroot was exposed to 22Na+-labelled solution (Fig. 6a),control and hkt1;1 plants exported approximately the sameamount of Na+ from the apical root to the rest of the plant,suggesting that there was no difference in xylem loading, atleast in this part of the root (Fig. 6b, ‘translocated’).
0 10 20 30 40 50 60
0
10
20
30 hk t1;1Control
t (min)
Na
+ u
pta
ke
(mm
ol g
–1 ro
ot F
W)
1 20 24
0
50
100
150
200gl1hk t1;1
t (h)
Na
+ u
pta
ke (mm
ol g
–1 ro
ot F
W)
(a)
(b)
(c)
0 23 52
0.0
2.5
5.0
7.5
100
100
111
82 87
96
Time since transfer to rinse (h)
Shoot N
a+ (mm
ol/s
hoot)
Figure 5. Na+ uptake from 50 mM NaCl into shoots of intactplants measured using 22Na+. (a) Time course of shoot Na+ uptakein 3 mM Ca2+ activity. Uptake 20–60 min was linear in hkt1;1[y = 0.467 mmol g-1 root fresh weight (FW) min-1 - 6.500 mmol g-1]and in the control if the anomalous 50 min time point wasexcluded (y = 0.109 mmol g-1 root FW min-1 + 0.312 mmol g-1).hkt1;1 plants showed higher influx at all time points except50 min, and the difference between genotypes was significant(P = 0.018) when this time point was excluded. (b) Shoot Na+
accumulation over 24 h, 0.2 mM Ca2+ activity. (c) Na+
recirculation. NaCl-grown plants were loaded with 22Na+-labelled50 mM NaCl (3 mM Ca2+ activity) for 40 h then transferred tounlabelled rinse solution and sampled over 52 h for changes inshoot Na content (measured as 22Na+). Shoot Na+ content ratherthan concentration was measured because plant growth over thetime course of the experiment would have produced anartifactual reduction in shoot Na+ concentration. Numbers abovebars represent % change in shoot Na+ with respect to shoot Na+
content at the beginning of the rinse period. Data pointsrepresent mean � SEM, n = 4. Each panel represents data from aseparate experiment. SEM, standard error of the mean.
502 R. J. Davenport et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507
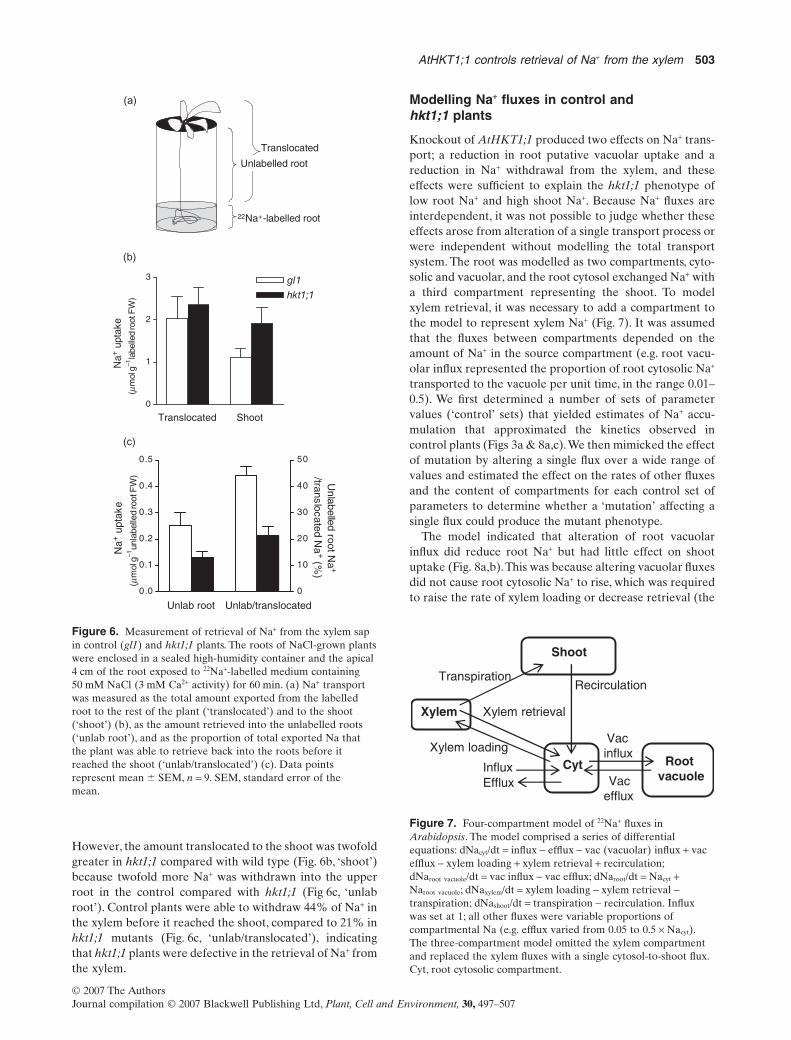
However, the amount translocated to the shoot was twofoldgreater in hkt1;1 compared with wild type (Fig. 6b, ‘shoot’)because twofold more Na+ was withdrawn into the upperroot in the control compared with hkt1;1 (Fig 6c, ‘unlabroot’). Control plants were able to withdraw 44% of Na+ inthe xylem before it reached the shoot, compared to 21% inhkt1;1 mutants (Fig. 6c, ‘unlab/translocated’), indicatingthat hkt1;1 plants were defective in the retrieval of Na+ fromthe xylem.
Modelling Na+ fluxes in control andhkt1;1 plants
Knockout of AtHKT1;1 produced two effects on Na+ trans-port; a reduction in root putative vacuolar uptake and areduction in Na+ withdrawal from the xylem, and theseeffects were sufficient to explain the hkt1;1 phenotype oflow root Na+ and high shoot Na+. Because Na+ fluxes areinterdependent, it was not possible to judge whether theseeffects arose from alteration of a single transport process orwere independent without modelling the total transportsystem. The root was modelled as two compartments, cyto-solic and vacuolar, and the root cytosol exchanged Na+ witha third compartment representing the shoot. To modelxylem retrieval, it was necessary to add a compartment tothe model to represent xylem Na+ (Fig. 7). It was assumedthat the fluxes between compartments depended on theamount of Na+ in the source compartment (e.g. root vacu-olar influx represented the proportion of root cytosolic Na+
transported to the vacuole per unit time, in the range 0.01–0.5). We first determined a number of sets of parametervalues (‘control’ sets) that yielded estimates of Na+ accu-mulation that approximated the kinetics observed incontrol plants (Figs 3a & 8a,c).We then mimicked the effectof mutation by altering a single flux over a wide range ofvalues and estimated the effect on the rates of other fluxesand the content of compartments for each control set ofparameters to determine whether a ‘mutation’ affecting asingle flux could produce the mutant phenotype.
The model indicated that alteration of root vacuolarinflux did reduce root Na+ but had little effect on shootuptake (Fig. 8a,b).This was because altering vacuolar fluxesdid not cause root cytosolic Na+ to rise, which was requiredto raise the rate of xylem loading or decrease retrieval (the
Translocated Shoot
0
1
2
3 gl1
hkt1;1
Na
+ u
pta
ke
(mm
ol g
–1 la
belle
d ro
ot F
W)
Unlab root Unlab/translocated
0.0
0.1
0.2
0.3
0.4
0.5
0
10
20
30
40
50
Na
+ u
pta
ke
(mm
ol g
–1 u
nla
belle
d ro
ot F
W)
Unla
belle
d ro
ot N
a+
/translo
cate
d N
a+ (%
)
Unlabelled root
22Na+-labelled root
Translocated
(a)
(b)
(c)
Figure 6. Measurement of retrieval of Na+ from the xylem sapin control (gl1) and hkt1;1 plants. The roots of NaCl-grown plantswere enclosed in a sealed high-humidity container and the apical4 cm of the root exposed to 22Na+-labelled medium containing50 mM NaCl (3 mM Ca2+ activity) for 60 min. (a) Na+ transportwas measured as the total amount exported from the labelledroot to the rest of the plant (‘translocated’) and to the shoot(‘shoot’) (b), as the amount retrieved into the unlabelled roots(‘unlab root’), and as the proportion of total exported Na thatthe plant was able to retrieve back into the roots before itreached the shoot (‘unlab/translocated’) (c). Data pointsrepresent mean � SEM, n = 9. SEM, standard error of themean.
Root vacuole
Shoot
Xylem
CytInflux
Efflux
Vac
influx
Vac
efflux
Xylem loading
Xylem retrieval
TranspirationRecirculation
Figure 7. Four-compartment model of 22Na+ fluxes inArabidopsis. The model comprised a series of differentialequations: dNacyt/dt = influx - efflux - vac (vacuolar) influx + vacefflux - xylem loading + xylem retrieval + recirculation;dNaroot vacuole/dt = vac influx - vac efflux; dNaroot/dt = Nacyt +Naroot vacuole; dNaxylem/dt = xylem loading - xylem retrieval -transpiration; dNashoot/dt = transpiration - recirculation. Influxwas set at 1; all other fluxes were variable proportions ofcompartmental Na (e.g. efflux varied from 0.05 to 0.5 ¥ Nacyt).The three-compartment model omitted the xylem compartmentand replaced the xylem fluxes with a single cytosol-to-shoot flux.Cyt, root cytosolic compartment.
AtHKT1;1 controls retrieval of Na+ from the xylem 503
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507
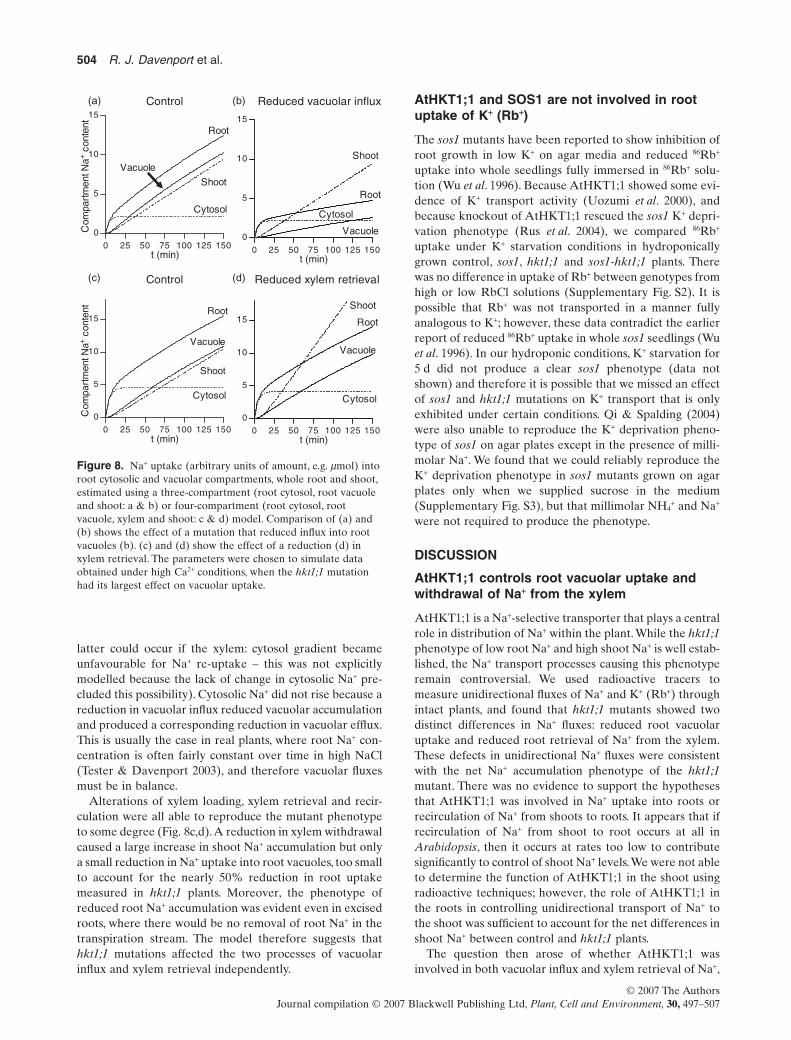
latter could occur if the xylem: cytosol gradient becameunfavourable for Na+ re-uptake – this was not explicitlymodelled because the lack of change in cytosolic Na+ pre-cluded this possibility). Cytosolic Na+ did not rise because areduction in vacuolar influx reduced vacuolar accumulationand produced a corresponding reduction in vacuolar efflux.This is usually the case in real plants, where root Na+ con-centration is often fairly constant over time in high NaCl(Tester & Davenport 2003), and therefore vacuolar fluxesmust be in balance.
Alterations of xylem loading, xylem retrieval and recir-culation were all able to reproduce the mutant phenotypeto some degree (Fig. 8c,d).A reduction in xylem withdrawalcaused a large increase in shoot Na+ accumulation but onlya small reduction in Na+ uptake into root vacuoles, too smallto account for the nearly 50% reduction in root uptakemeasured in hkt1;1 plants. Moreover, the phenotype ofreduced root Na+ accumulation was evident even in excisedroots, where there would be no removal of root Na+ in thetranspiration stream. The model therefore suggests thathkt1;1 mutations affected the two processes of vacuolarinflux and xylem retrieval independently.
AtHKT1;1 and SOS1 are not involved in rootuptake of K+ (Rb+)
The sos1 mutants have been reported to show inhibition ofroot growth in low K+ on agar media and reduced 86Rb+
uptake into whole seedlings fully immersed in 86Rb+ solu-tion (Wu et al. 1996). Because AtHKT1;1 showed some evi-dence of K+ transport activity (Uozumi et al. 2000), andbecause knockout of AtHKT1;1 rescued the sos1 K+ depri-vation phenotype (Rus et al. 2004), we compared 86Rb+
uptake under K+ starvation conditions in hydroponicallygrown control, sos1, hkt1;1 and sos1-hkt1;1 plants. Therewas no difference in uptake of Rb+ between genotypes fromhigh or low RbCl solutions (Supplementary Fig. S2). It ispossible that Rb+ was not transported in a manner fullyanalogous to K+; however, these data contradict the earlierreport of reduced 86Rb+ uptake in whole sos1 seedlings (Wuet al. 1996). In our hydroponic conditions, K+ starvation for5 d did not produce a clear sos1 phenotype (data notshown) and therefore it is possible that we missed an effectof sos1 and hkt1;1 mutations on K+ transport that is onlyexhibited under certain conditions. Qi & Spalding (2004)were also unable to reproduce the K+ deprivation pheno-type of sos1 on agar plates except in the presence of milli-molar Na+. We found that we could reliably reproduce theK+ deprivation phenotype in sos1 mutants grown on agarplates only when we supplied sucrose in the medium(Supplementary Fig. S3), but that millimolar NH4
+ and Na+
were not required to produce the phenotype.
DISCUSSION
AtHKT1;1 controls root vacuolar uptake andwithdrawal of Na+ from the xylem
AtHKT1;1 is a Na+-selective transporter that plays a centralrole in distribution of Na+ within the plant. While the hkt1;1phenotype of low root Na+ and high shoot Na+ is well estab-lished, the Na+ transport processes causing this phenotyperemain controversial. We used radioactive tracers tomeasure unidirectional fluxes of Na+ and K+ (Rb+) throughintact plants, and found that hkt1;1 mutants showed twodistinct differences in Na+ fluxes: reduced root vacuolaruptake and reduced root retrieval of Na+ from the xylem.These defects in unidirectional Na+ fluxes were consistentwith the net Na+ accumulation phenotype of the hkt1;1mutant. There was no evidence to support the hypothesesthat AtHKT1;1 was involved in Na+ uptake into roots orrecirculation of Na+ from shoots to roots. It appears that ifrecirculation of Na+ from shoot to root occurs at all inArabidopsis, then it occurs at rates too low to contributesignificantly to control of shoot Na+ levels.We were not ableto determine the function of AtHKT1;1 in the shoot usingradioactive techniques; however, the role of AtHKT1;1 inthe roots in controlling unidirectional transport of Na+ tothe shoot was sufficient to account for the net differences inshoot Na+ between control and hkt1;1 plants.
The question then arose of whether AtHKT1;1 wasinvolved in both vacuolar influx and xylem retrieval of Na+,
0 25 50 75 100 125 150
0
5
10
15
Cytosol
Shoot
Vacuole
Root
t (min)
Com
part
ment N
a+ c
onte
nt
0 25 50 75 100 125 150
0
5
10
15
Cytosol
Shoot
Vacuole
Root
t (min)
0 25 50 75 100 125 150
0
5
10
15
Cytosol
Shoot
Vacuole
Root
t (min)
Com
part
ment N
a+ c
onte
nt
0 25 50 75 100 125 150
0
5
10
15
Cytosol
Shoot
Vacuole
Root
t (min)
(a) (b)
(c) (d)
Control Reduced vacuolar influx
Control Reduced xylem retrieval
Figure 8. Na+ uptake (arbitrary units of amount, e.g. mmol) intoroot cytosolic and vacuolar compartments, whole root and shoot,estimated using a three-compartment (root cytosol, root vacuoleand shoot: a & b) or four-compartment (root cytosol, rootvacuole, xylem and shoot: c & d) model. Comparison of (a) and(b) shows the effect of a mutation that reduced influx into rootvacuoles (b). (c) and (d) show the effect of a reduction (d) inxylem retrieval. The parameters were chosen to simulate dataobtained under high Ca2+ conditions, when the hkt1;1 mutationhad its largest effect on vacuolar uptake.
504 R. J. Davenport et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507

or whether perturbation of one of these processes was suf-ficient to cause the hkt1;1 phenotype. Net accumulation ofions depends on the sum of ion fluxes, but the fluxes them-selves are interdependent, so alteration of one flux haseffects both on net accumulation and on other fluxes.There-fore, it was necessary to model the movements of Na+ withinthe plant to estimate how alterations of single fluxes couldimpact on other transport processes. Our very simple modelwas able to describe the observed fluxes and, in conjunctionwith the experimental evidence, indicated that the effects ofthe hkt1;1 mutation on vacuolar uptake and xylem retrievalof Na+ were independent.
The dual effects of AtHKT1;1 on root Na+ transportsupport the hypothesis of Rus et al. (2004) that AtHKT1;1functions to control ion homeostasis. How can these resultsbe reconciled with those of Berthomieu et al. (2003), whofound a fivefold elevation of Na+ in the phloem of hkt1;1(sas2) mutants compared to wild type and evidence forAtHKT1;1 localization to the phloem? Sunarpi et al. (2005)also found lower Na+ in the phloem of hkt1;1 mutants, butfound that the reduction in phloem Na+ between hkt1;1 andcontrols ranged between approximately sixfold and twofolddepending on the hkt1;1 allelic mutant. A key question iswhether the rate of Na+ export from the shoot in thephloem is high enough to have a significant effect on shootNa+ concentration. Berthomieu et al. (2003) compared onlyrelative amounts of phloem Na+ using the Na+ : glutamineratio; however, Sunarpi et al. (2005) reported the amount ofNa+ extruded from the phloem. They measured approxi-mately 30 mmol Na+ g-1 leaf DW, collected over a 4 h period,from plants exposed to 75 mM NaCl for 2 d. Xylem Na+
content in the controls was approximately 6 mmol Na+ mL-1
xylem sap. The transpiration rate was not reported so it isdifficult to compare these two measures. However, if weassume a transpiration rate of 0.3 mL g-1 leaf fresh weight(FW) h-1 (Hosy et al. 2003), then the delivery of Na+ to theleaf would be 2 mmol Na+/g-1 FW h-1. The retranslocation ofNa+ via the phloem would be 0.75 mmol g-1 FW h-1 (assum-ing a 10:1 leaf FW : DW ratio), suggesting that the phloemcould remove as much as 40% of Na+ delivered in thexylem. This is much higher than indicated in the recircula-tion experiment presented in Fig. 5. If the average transpi-ration rate were 1 mL g-1 leaf FW h-1,then the phloemwould remove only 12.5% of xylem-delivered Na+, closer toour estimates for control plants (Fig. 5c). However, even ifthe phloem removed as much as half the Na+ deliveredin the xylem, a mutation causing a fivefold reduction inphloem export would result in only a 1.8-fold increase inshoot Na+ accumulation, which is too low to account for thethree- to ninefold excess accumulation usually found inhkt1;1 mutants (Fig. 2c; Maser et al. 2002; Berthomieu et al.2003; Gong et al. 2004). For a fivefold reduction in phloemexport to cause a fivefold increase in shoot Na+ accumula-tion, phloem export would have to constitute around 85%of xylem import. We could find no evidence of such signifi-cant recirculation. By contrast, genotypic differences affect-ing xylem delivery would have a directly proportional effecton shoot Na+. Although Berthomieu et al. (2003) measured
only a statistically insignificant doubling of xylem Na+ inthe hkt1;1 (sas2) mutant, Sunarpi et al. (2005) found largeand significant increases in xylem Na+ in hkt1;1 mutantscompared with wild-type controls, and concluded thatwithdrawal of Na+ from the xylem constituted the mostsignificant function of AtHKT1;1 with respect to Na+ trans-port in the shoot. While it is very likely that hkt1;1 mutantshave lower rates of phloem transport of Na+, the morecritical effect on shoot Na+ is the early effect of the hkt1;1mutation in roots on unidirectional transfer of Na+ to theshoot in the xylem. There is little evidence for a significantcontribution of recirculation to control of shoot Na+ contentin any species (Tester & Davenport 2003).
The localization of AtHKT1;1 remains problematic. Ber-thomieu et al. (2003) presented evidence from RNA in situhybridization and promoter fusion for phloem localizationof AtHKT1;1 transcripts. While the images presented didnot localize AtHKT1;1 unequivocally to the phloem,AtHKT1;1 transcripts appeared to be concentrated in cellsaround the phloem rather than the xylem (Berthomieuet al. 2003). However, using an antibody raised toAtHKT1;1, Sunarpi et al. (2005) demonstrated localizationof the protein to the plasma membrane of xylem paren-chyma cells in the shoot, commensurate with a direct rolefor AtHKT1;1 in withdrawal of Na+ from the xylem in theshoot. It remains unclear, therefore, whether AtHKT1;1participates directly in retrieval of xylem Na+ in roots orcontrols the process indirectly. Plasma membrane localiza-tion of AtHKT1;1 is not compatible with a direct involve-ment in vacuolar Na+ fluxes, and the effect of the hkt1;1mutation in reducing vacuolar Na+ influx is likely to beindirect (unless the AtHKT1;1 protein has several intracel-lular locations). Overexpression of AtHKT1 under controlof its likely native promoter increased salt sensitivity (Ruset al. 2004), indicating that the role of AtHKT1;1 in Na+
transport involves more than simply a reduction of Na+
transport to the shoot.
How does hkt1;1 rescue sos1?
In hydroponic growth conditions, sos1 accumulated Na+ tothe same concentration as hkt1;1 and sos1-hkt1;1 mutants inthe shoot, and showed little reduction in shoot growth insaline conditions (although the shoot displayed visiblesymptoms of stress). Therefore, high shoot Na+ was not thecause of sos1 salt hypersensitivity, in accord with previousevidence (Ding & Zhu 1997; Shi et al. 2002). However, sos1plants showed a strong reduction in root growth and hyper-accumulated Na+ in roots. Root Na+ accumulation was par-tially reduced in the sos1-hkt1;1 double mutant relative tosos1, in conjunction with partial relief of growth inhibition.Because knockout of AtHKT1;1 caused a reduction in rootNa+ accumulation (via inhibition of vacuolar influx), theseresults suggest that hkt1;1 mutations relieve sos1 salt sensi-tivity by reducing root Na+ accumulation.
The mechanism by which the hkt1;1 mutation rescuessos1 growth in limiting K+ conditions remains unclear. Wefound no evidence for a role of AtHKT1;1 in K+ transport.
AtHKT1;1 controls retrieval of Na+ from the xylem 505
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507

We also found no evidence that the sos1 mutant was defec-tive in K+ acquisition (K+ influx or net accumulation). Thismay have been because the K+-specific defect in sos1 is onlyevident in certain growth conditions. Qi & Spalding (2004)could not reproduce the sos1 K+ deprivation phenotypeexcept in the presence of millimolar Na+ and speculatedthat excess root cytosolic Na+ accumulation in sos1 plantsinhibited K+ uptake via K+ channels. This would be compat-ible with our results showing that the hkt1;1 mutationreduced root Na+ in sos1 and restored root growth.However, sos1 did not demonstrate reduced K+ accumula-tion under saline conditions (although K+ distribution wasperturbed, with sos1 plants accumulating higher shoot K+
and lower root K+ than controls) (Fig. 2d). Tissue K+ levelswere not critical in determining salt sensitivity because thesos1-hkt1;1 mutant resembled sos1 in K+ accumulation butwas less salt sensitive. When we attempted to replicate pre-vious studies using agar plates, we found no genotype-specific inhibitory effect of low millimolar Na+ or NH4
+
(which blocks high-affinity K+ uptake) on sos1 growth onlow K+ agar plates, and could only reliably reproduce thesos1 K+ phenotype in the presence of sucrose (Supplemen-tary Fig. S3). Qi & Spalding (2004) did not include sucrosein their media, whereas sucrose was always present inexperiments demonstrating a clear sos1 K+ phenotype (Wuet al. 1996; Ding & Zhu 1997; Rus et al. 2001, 2004; Shi et al.2002). K+ is known to be important in sucrose transport(Deeken et al. 2002), and it is possible that indirect effects ofmutations of SOS1 and AtHKT1;1 on intra-root K+ trans-port perturb phloem transport processes.
In conclusion, we have demonstrated that AtHKT1;1 isnot involved in Na+ recirculation via the phloem and is notresponsible for Na+ influx into roots. Rather, AtHKT1;1appears to control both retrieval of Na+ from the xylem androot vacuolar loading.This paper demonstrates the utility ofmeasurements of unidirectional fluxes with radioactivetracers and the use of simple mathematical models to testintuitive descriptions of plant transport processes.
ACKNOWLEDGMENTS
We thank J. Banfield, R. Hoskings and Dr S. Roy for assis-tance. This work was supported by the Royal Society (R.J.Davenport), the Spanish government (A. Muñoz-Mayor)and the Australian Research Council (M. Tester andD. Jha).
REFERENCES
Amtmann A. & Sanders D. (1999) Mechanisms of Na+ uptake byplant cells. Advances in Botanical Research 29, 75–112.
Apse M.P., Aharon G.S., Sneddon W.A. & Blumwald E. (1999) Salttolerance conferred by overexpression of a vacuolar Na+/H+
antiport in Arabidopsis. Science 285, 1256–1258.Arteca R.N. & Arteca J.M. (2000) A novel method for growing
Arabidopsis thaliana plants hydroponically. Physiologia Plan-tarum 108, 188–193.
Berthomieu P., Conéjéro G., Nublat A., et al. (2003) Functionalanalysis of AtHKT1 in Arabidopsis shows that Na+ recirculationby the phloem is crucial for salt tolerance. EMBO Journal 22,2004–2014.
Davenport R.J., James R.A., Zakrisson-Plogander A., Tester M. &Munns R. (2005) Control of sodium transport in durum wheat.Plant Physiology 137, 807–818.
Deeken R., Geiger D., Fromm J., Koroleva O., Ache P., Langenfeld-Heyser R., Sauer N., May S.T. & Hedrich R. (2002) Loss of theAKT2/3 potassium channel affects sugar loading into the phloemof Arabidopsis. Planta 216, 334–344.
Ding L. & Zhu J.-K. (1997) Reduced Na+ uptake in the NaCl-hypersensitive sos1 mutant of Arabidopsis thaliana. PlantPhysiology 113, 795–799.
Essah P.A., Davenport R. & Tester M. (2003) Sodium influx andaccumulation in Arabidopsis. Plant Physiology 133, 307–318.
Garciadeblás B., Senn M.E., Bañuelos M.A. & Rodríguez-NavarroA. (2003) Sodium transport and HKT transporters: the ricemodel. Plant Journal 34, 788–801.
Gaxiola R., Li J., Undurraga S., Dang L.M., Allen G.J., Alper S.L.& Fink G.R. (2001) Drought- and salt-tolerant plants resultsfrom overexpression of the AVP1 H+ pump. Proceedings ofthe National Academy of Sciences of the USA 98, 11444–11449.
Gong J.-M., Waner D.A., Horie T., Li S.L., Horie R., Abid K.B. &Schroeder J.I. (2004) Microarray-based rapid cloning of an ionaccumulation deletion mutant in Arabidopsis thaliana. Proceed-ings of the National Academy of Sciences of the USA 101, 15404–15409.
Haro R., Bañuelos M.A., Senn M.E., Barrero-Gil J. & Rodríguez-Navarro A. (2005) HKT1 mediates Na+ uniport in roots. Pitfallsin the expression of HKT1 in yeast. Plant Physiology 139, 1495–1506.
Hosy E., Vavasseur A., Mouline K., et al. (2003) The Arabidopsisoutward K+ channel GORK is involved in regulation of stomatalmovements and plant transpiration. Proceedings of the NationalAcademy of Sciences of the USA 100, 5549–5554.
Laurie S., Feeney K.A., Maathuis F.J.M., Heard P.J., Brown S.J. &Leigh R.A. (2002) A role for HKT1 in sodium uptake by wheatroots. Plant Journal 32, 139–149.
Lazof D. & Cheeseman J.M. (1986) Sodium transport and com-partmentation in Spergularia marina. Plant Physiology 81, 742–747.
Maser P., Eckelman B., Vaidyanathan R., et al. (2002) Alteredshoot/root Na+ distribution and bifurcating salt sensitivity inArabidopsis by genetic disruption of the Na+ transporterAtHKT1. FEBS Letters 531, 157–161.
Qi Z. & Spalding E.P. (2004) Protection of plasma membrane K+
transport by the salt overly sensitive Na+-H+ antiporter duringsalinity stress. Plant Physiology 136, 2548–2555.
Qiu Q.-S., Barkla B.J., Vera-Estrella R., Zhu J.-K. & SchumakerK.S. (2003) Na+/H+ exchange activity in the plasma membrane ofArabidopsis. Plant Physiology 132, 1041–1052.
Ren Z.-H., Gao J.-P., Li L.-G., Cai X.-L., Huang W., Chao D.-Y.,Zhu M.-Z., Wang Z.-Y., Luan S. & Lin H.-X. (2005) A ricequantitative trait locus for salt tolerance encodes a sodium trans-porter. Nature Genetics 37, 1141–1146.
Rubio F., Gassmann W. & Schroeder J.I. (1995) Sodium-drivenpotassium uptake by the plant potassium transporter HKT1 andmutations conferring salt tolerance. Science 270, 1660–1663.
Rus A., Yokoi S., Altanbadralt S., Reddy M., Lee B., MatsumotoT.K., Koiwa H., Zhu J.-K., Bressan R.A. & Hasegawa P.M. (2001)AtHKT1 is a salt tolerance determinant that controls Na+ entryinto plant roots. Proceedings of the National Acadeny of Sciencesof the USA 98, 14150–14155.
Rus A., Lee B., Muñoz-Mayor A., Sharkuu A., Miura K., Zhu J.-K.,
506 R. J. Davenport et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507

Bressan R.A. & Hasegawa P.M. (2004) AtHKT1 facilitates Na+
homeostasis and K+ nutrition in planta. Plant Physiology 136,2500–2511.
Shi H., Quintero F.J., Pardo J.M. & Zhu J.-K. (2002) The putativeplasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 14, 465–477.
Shin R. & Schachtman D.P. (2004) Hydrogen peroxide mediatesplant root cell response to nutrient deprivation. Proceedings ofthe National Acadeny of Sciences of the USA 101, 8827–8832.
Sunarpi H.T., Motoda J., Kubo M., et al. (2005) Enhanced salt tol-erance mediated by AtHKT1 transporter-induced unloadingfrom xylem vessels to xylem parenchyma cells. Plant Journal 44,928–938.
Tester M. & Davenport R. (2003) Na+ tolerance and Na+ transportin higher plants. Annals of Botany 91, 503–527.
Uozumi N., Kim E.J., Rubio F., Yamaguchi T., Muto S., Tsuboi A.,Bakker E.P., Nakamura T. & Schroeder J.I. (2000) The Arabi-dopsis HKT1 gene homolog mediates inward currents inXenopus laevis oocytes and Na+ uptake in Saccharomyces cerevi-siae. Plant Physiology 122, 1249–1259.
Walker N.A. & Pitman M.G. (1976) Measurements of fluxes acrossmembranes. In Encyclopedia of Plant Physiology (eds E. Luttge& M. Pitman) Vol. 2 (A), pp. 93–126. Springer-Verlag, Berlin.
Wu S.-J., Ding L. & Zhu J.-K. (1996) SOS1, a genetic locus essentialfor salt tolerance and potassium acquisition. Plant Cell 8, 617–627.
Received 12 September 2006; received in revised form 24 November2006; accepted for publication 12 December 2006
SUPPLEMENTARY MATERIAL
The following supplementary materials are available forthis article:
Figure S1. 86Rb+ uptake in Arabidopsis. Kinetics of Rb+
uptake in high NH4+-grown (solution 2, K+ replaced with
NH4 salts) K+-starved control plants.Figure S2. Comparison of 86Rb+ uptake in hkt1;1, sos1, sos1-htk1;1 and control (gl1) plants. Rb+ uptake into roots (a)and transport to the shoot (b) in intact K+-starved plantsgrown in high NH4
+ (solution 2), measured as 86Rb+ uptake.Figure S3. Effect of sucrose on sos1 K+ starvation pheno-type. Root growth assayed after 8 d growth on vertical agarplates in the presence or absence of 3% sucrose with low(0.02 mM) or high (20 mM) KCl.
This material is available as part of the online articlefrom: http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-3040.2007.01637x(This link will take you to the article abstract)
Please note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materialssupplied by the authors. Any queries (other than missingmaterial) should be directed to the corresponding authorfor the article.
AtHKT1;1 controls retrieval of Na+ from the xylem 507
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Plant, Cell and Environment, 30, 497–507
Copyright © 2022 FDOKUMEN




















