
The Molecular Physiology of the Cardiac Transient Outward Potassium Current (I to) in Normal and...
22
J Mol Cell Cardiol 33, 851–872 (2001) doi:10.1006/jmcc.2001.1376, available online at http://www.idealibrary.com on Basic Cardiac Electrophysiology The Molecular Physiology of the Cardiac Transient Outward Potassium Current (I to ) in Normal and Diseased Myocardium Gavin Y. Oudit, Zamaneh Kassiri, Rajan Sah, Rafael J. Ramirez, Carsten Zobel and Peter H. Backx Department of Medicine and Physiology, Toronto General Hospital, University Health Network, University of Toronto, Toronto, Canada (Received 15 February 2001, accepted 2 March 2001) G. Y. O, Z. K, R. S, R. J. R, C. Z P. H. B. The Molecular Physiology of the Cardiac Transient Outward Potassium Current (I to ) in Normal and Diseased Myocardium. Journal of Molecular and Cellular Cardiology (2001) 33, 851–872. The Ca 2+ -independent transient outward potassium current (I to ) plays an important role in early repolarization of the cardiac action potential. I to has been clearly demonstrated in myocytes from different cardiac regions and species. Two kinetic variants of cardiac I to have been identified: fast I to , called I to,f , and slow I to , called I to,s . Recent findings suggest that I to,f is formed by assembly of K v4.2 and/or K v4.3 alpha pore- forming voltage-gated subunits while I to,s is comprised of K v1.4 and possibly K v1.7 subunits. In addition, several regulatory subunits and pathways modulating the level and biophysical properties of cardiac I to have been identified. Experimental findings and data from computer modeling of cardiac action potentials have conclusively established an important physiological role of I to in rodents, with its role in large mammals being less well defined due to complex interplay between a multitude of cardiac ionic currents. A central and consistent electrophysiological change in cardiac disease is the reduction in I to density with a loss of heterogeneity of I to expression and associated action potential prolongation. Alterations of I to in rodent cardiac disease have been linked to repolarization abnormalities and alterations in intracellular Ca 2+ homeostasis, while in larger mammals the link with functional changes is far less certain. We review the current literature on the molecular basis for cardiac I to and the functional consequences of changes in I to that occur in cardiovascular disease. 2001 Academic Press K W: Transient outward potassium current; Myocytes; Regulatory subunits; Cardiac action potential; Intracellular Ca 2+ ; Arrhythmia; Hypertrophy; Congestive heart failure. and developmental stages can be traced to a com- Introduction mon set of molecular correlates of K v channel pro- teins and their auxiliary subunits. 5,6 The The cardiac action potential (AP) profile is governed by the complex interaction of depolarizing and permutation of various combinations of the pore- forming K v channels, combined with the existence repolarizing currents. Variation in the expression and biophysical properties of voltage-gated K + of an ever-increasing number of accessory proteins and modulatory pathways, provides ongoing chal- channels (K v ) in cardiac myocytes from different species as well as intra-cardiac regional variation lenges for understanding the molecular basis of myocardial electrophysiology. largely accounts for the wide variety of myocardial AP profiles observed. 1–4 It is now becoming clear Generally, voltage-gated K + currents can be clas- sified into two groups based upon their time- and that electrophysiological diversity of K v repolarizing currents between different cardiac regions, species voltage-dependent properties. The first group is the Please address all correspondence to: Peter H. Backx, Toronto General Hospital, CCRW 3-802, 101 College St., Toronto, Ontario, M5G 2C4, Canada. E-mail: [email protected] 0022–2828/01/050851+22 $35.00/0 2001 Academic Press
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The Molecular Physiology of the Cardiac Transient Outward Potassium Current (I to) in Normal and...

J Mol Cell Cardiol 33, 851–872 (2001)
doi:10.1006/jmcc.2001.1376, available online at http://www.idealibrary.com on
Basic Cardiac Electrophysiology
The Molecular Physiology of the CardiacTransient Outward Potassium Current (Ito)in Normal and Diseased MyocardiumGavin Y. Oudit, Zamaneh Kassiri, Rajan Sah, Rafael J. Ramirez,Carsten Zobel and Peter H. BackxDepartment of Medicine and Physiology, Toronto General Hospital, University Health Network,University of Toronto, Toronto, Canada
(Received 15 February 2001, accepted 2 March 2001)
G. Y. O, Z. K, R. S, R. J. R, C. Z P. H. B. The Molecular Physiology of the CardiacTransient Outward Potassium Current (Ito) in Normal and Diseased Myocardium. Journal of Molecular and CellularCardiology (2001) 33, 851–872. The Ca2+-independent transient outward potassium current (Ito) plays animportant role in early repolarization of the cardiac action potential. Ito has been clearly demonstrated in myocytesfrom different cardiac regions and species. Two kinetic variants of cardiac Ito have been identified: fast Ito, calledIto,f, and slow Ito, called Ito,s. Recent findings suggest that Ito,f is formed by assembly of Kv4.2 and/or Kv4.3 alpha pore-forming voltage-gated subunits while Ito,s is comprised of Kv1.4 and possibly Kv1.7 subunits. In addition, severalregulatory subunits and pathways modulating the level and biophysical properties of cardiac Ito have beenidentified. Experimental findings and data from computer modeling of cardiac action potentials have conclusivelyestablished an important physiological role of Ito in rodents, with its role in large mammals being less well defineddue to complex interplay between a multitude of cardiac ionic currents. A central and consistent electrophysiologicalchange in cardiac disease is the reduction in Ito density with a loss of heterogeneity of Ito expression and associatedaction potential prolongation. Alterations of Ito in rodent cardiac disease have been linked to repolarizationabnormalities and alterations in intracellular Ca2+ homeostasis, while in larger mammals the link with functionalchanges is far less certain. We review the current literature on the molecular basis for cardiac Ito and thefunctional consequences of changes in Ito that occur in cardiovascular disease. 2001 Academic Press
K W: Transient outward potassium current; Myocytes; Regulatory subunits; Cardiac action potential;Intracellular Ca2+; Arrhythmia; Hypertrophy; Congestive heart failure.
and developmental stages can be traced to a com-Introductionmon set of molecular correlates of Kv channel pro-teins and their auxiliary subunits.5,6 TheThe cardiac action potential (AP) profile is governed
by the complex interaction of depolarizing and permutation of various combinations of the pore-forming Kv channels, combined with the existencerepolarizing currents. Variation in the expression
and biophysical properties of voltage-gated K+ of an ever-increasing number of accessory proteinsand modulatory pathways, provides ongoing chal-channels (Kv) in cardiac myocytes from different
species as well as intra-cardiac regional variation lenges for understanding the molecular basis ofmyocardial electrophysiology.largely accounts for the wide variety of myocardial
AP profiles observed.1–4 It is now becoming clear Generally, voltage-gated K+ currents can be clas-sified into two groups based upon their time- andthat electrophysiological diversity of Kv repolarizing
currents between different cardiac regions, species voltage-dependent properties. The first group is the
Please address all correspondence to: Peter H. Backx, Toronto General Hospital, CCRW 3-802, 101 College St., Toronto, Ontario,M5G 2C4, Canada. E-mail: [email protected]
0022–2828/01/050851+22 $35.00/0 2001 Academic Press

G. Y. Oudit et al.852
delayed rectifiers (IK), which activate at variable currents by themselves in heterologous systems andare thought to regulate the activity of the Kv2rates and either do not inactivate or inactivate very
slowly. While these currents can contribute to all subfamily.6
Assembly of four Kv�-subunits into a tetramericphases of repolarization, in most species they aremore important for the later phases of re- structure creates a functional Ito channel.16 The
preponderance of evidence suggests that only Kv�polarization. Delayed rectifiers can be classified intothe ultra-rapid, IKUR, rapid, IKR, and slow, IKS, delayed from the same family can assemble or co-assemble
to form a functional channel. This co-assemblyrectifiers based on their relative rate of activation.7
The other group of currents is the Ca2+-independent depends critically on highly conserved N-terminalregions called T- or Nab-domains17,18 although ex-transient outward potassium current (Ito) which
rapidly activates and inactivates following mem- ceptions occur.19 The pore-forming alpha subunit(Kv�) contains six transmembrane domains (S1–brane depolarization.2–4,8,9
As a result of its contribution to the early phase of S6), flanked by cytoplasmic amino and carboxyltermini. S4 is a highly positively charged segmentrepolarization, Ito strongly affects L-type Ca2+current
magnitude, thereby modulating excitation– that is involved in voltage sensing and channelactivation.16,20 The P-loop (between the S5 and S6contraction coupling processes and myocardial con-
tractility as well as repolarization. As such, Ito adds transmembrane domains) from each subunit projectcentrally to create the K+-selective pore structure20another level of diversity to its functional role via its
interaction with other ionic currents such as the L- with a structure likely to resemble that of the KcsAK+ channel.21type Ca2+and delayed rectifier currents.10–12 Because
of these complex interactions, changes in Ito can have With respect to Ito expression in the heart, onlyKv1.4, Kv1.7, Kv4.2, and Kv4.3 show substantial ex-unexpected consequences on the action potential
(AP) duration10,11 and susceptibility to lethal ar- pression in the mammalian myocardium (Table1),4,6,22 even though other Kv� genes can encoderhythmias.13–15 A more thorough understanding of
the electrical activity in the heart is clearly pivotal to for Ito-like currents (i.e. Kv3.4 and Kv4.1). Of thesecardiac Ito genes, only Kv4.3 is alternatively splicedenable further elucidation of the function of normal
and diseased myocardium as well as the development into two variants that are distinguished by theinsertion of 19 amino acids into the proximal C-of novel strategies for the treatment of cardiovascular
diseases. The purpose of this review article is to dis- terminus of the “long” variant.23–25 Human heartexpresses both alternatively spliced forms,23 whilecuss our current knowledge of the molecular basis
for Ito, including the role of auxillary subunits and the long Kv4.3 variant is predominantly expressedin rat.24 It appears that only the short variantregulatory pathways, and to approve the functional
importance of Ito in regulating the electrical and encodes for the functional Ito in dog heart.25
Following translation, Kv�-subunits are as-mechanical activities of the heart. The alterations inIto in pathological states such as myocardial hyper- sembled in the endoplasmic reticulum and then
undergo further post-translational modificationtrophy and heart failure and the mechanisms in-volved will be highlighted. and maturation in the Golgi apparatus prior to
transport to the sarcolemmae.26 The distributionof these Ito-encoding Kv channels in the sarcolem-mae displays a heterogeneous and tissue-specificpattern of expression. For example, im-Molecular Properties of Itomunofluorescent studies revealed that Kv4.2 wasintensely observed in the peripheral sarcolemmaePore composition and structure of Kv channelsand the intercalated disks in atrial myocytes,whereas in ventricular myocytes Kv4.2 was ob-Following the identification of four Drosophila sub-
families of voltage-gated K+ channels (shaker, shab, served predominantly in the transverse-axial tu-bular system.27 Although the basis for this patternshaw, shal), homologous families of pore-forming
alpha subunits were uncovered in mammals.3,4 is unknown, membrane targeting signals that areresponsible for their differential targeting to theThese K+ channels were classified using the Kvx.y
nomenclature, where Kv indicates a voltage-gated sarcolemmae are known to reside in the C-terminus of Kv channels.28 Such a C-terminal motifK+ channel, x represents the subfamily and y the
number of the gene within the subfamily.3,6 Pres- in Kv1.X channels (including Kv1.4) has recently beenshown to be necessary for plasma membraneently, nine families of pore-forming Kv�-subunits
(Kv1.x to Kv9.x) have been identified.4–6 Members of localization and glycosylation of the Kv subunitsas well as their interaction with beta-subunits.29the Kv5, Kv6, Kv8, and Kv9 subfamilies do not express

Structure and Function of Cardiac Ito 853
Table 1 Ito composition in different mammalian myocardial cell
Species Chamber Subtype Putative Clone Ito density# Referenceclone distribution
Mouse Atrium Ito,f Kv4.2 A<V 6, 22, 78, 80Kv4.3
Ventricle Ito,f Kv4.2 Epi>Endo Epi>EndoIto,s Kv1.4 Endo>Epi Apex>baseIto,s∗ Kv1.7
a Apex>SepRat Atrium Ito,f Kv4.2 A>V 50, 91, 96,
Kv4.3 A>V 106, 159, 237Ventricle Ito,f Kv4.2 RV>Sep Epi>Endo
Kv4.3 LV wall>SepIto,s Kv1.4
Ferret Atrium NR Kv4.2b RA, LA 82, 238
Kv4.3b RA>LA
Kv1.4b RA, LA
Ventricle Ito,f Kv4.2 Base, Epi, RV Epi>EndoKv4.3 Apical Epi
Ito,s Kv1.4 Apical EndoRabbit Atrium Ito,s Kv4.2/3 88, 89, 239,
Kv1.4 240, 241Figure 1
Ventricle Ito,s Kv4.2 Epi>Endo Epi>EndoKv1.4
SA node Ito,f NRAV node Ito,f
Ito,s
Dog Atrium Ito,f NR 92, 114, 189,242, 243
Ventricle Ito,f Kv4.3 Epi>Endo Epi>EndoIto,s Kv1.4 Endo>EpiNR Kv3.4
$ LVHuman Atrium Ito,f Kv4.3 6, 80, 89,
Kv4.2 103, 242, 244Ventricle Ito,f Kv4.3 Epi>Endo
Ito,s Kv1.4c
Kv4.1a
Ito,f and Ito,s refers to fast and slow inactivating Ito, respectively; A=atrium; V=ventricle; N=node; RA=right atrium; LA=left atrium;Sep=septum; Epi-epicardium; Endo=endocardium; #=overall Ito density; NR=not reported; ∗=studied in heterologous system(Xenopus oocytes); $=detected at mRNA level.242
a=detected in whole heart.b=detected in mRNA level, but no Ito density reported in ferret atrial myocytes.c=identified with RT-PCR244 but not with Western immunoblots.89
Regulatory subunits and cytoskeletal interaction reductase enzyme with its active site positioned topotentially interact with the K+ channel voltage
Assembly of the alpha-subunits into pore-forming sensor.5,34,36 �-subunits can also assemble with afour-fold symmetric structure and are able to co-tetramers is sufficient to generate functional K+
channels. However, the level of expression as well assemble with the T1 domain of the Kv�-subunit(T14�4)5,37 which occurs in the endoplasmic re-as the biophysical properties of voltage-gated potas-
sium channels are modulated by cytoplasmic pro- ticulum prior to plasmalemmal insertion.26
Kv�1.3 (human) and Kv�3 (human and ferret)teins, such as �-subunits, KChIP, frequenin, andKChAP. Three highly homologous �-subunit famil- have been cloned from the heart.30–32,38 The ex-
pression of �-subunits appears to be regulated in aies, Kv�1–Kv�3, have been cloned.30–34 The Kv�1(Kv�1.1, Kv�1.2, Kv�1.3), Kv�2 (Kv�2.1, Kv�2.2), and Kv�3 chamber-specific manner since the level of Kv�3
in human myocardium is two-fold higher in the(Kv�3.1, Kv�3.2) families are alternative spliced vari-ants of the same gene.34,35 Each beta-subunit con- ventricle than in the atrium.30 Kv�1, Kv�2, and
Kv�3 families can specifically associate with Kv1.1,tains a conserved core region with variable N-terminal peptides and functions as a complete oxido- Kv1.2, Kv1.4, Kv1.5 and Kv4.3�-subunits thereby mod-

G. Y. Oudit et al.854
ifying their functional properties, stability, and cell such as cell swelling and disruption of intracellularfilaments can alter ion conductivity indirectly bysurface expression.5,30,31,33,38–40 The N-terminal pep-
tide in Kv�1 and Kv�3 subunits confers a rapid altering the delivery, expression, clustering and/or phosphorylation of cardiac Na+ channel andinactivation to Kv1.1 and Kv1.5 subunits via a pro-
posed N-type inactivation5,39,41,42 while Kv�2.1 only inwardly rectifying K+ channels.52–54 Recent datahave provided supporting evidence for the role ofenhances C-type inactivation in Kv1.5 channels.43
Furthermore, human and ferret Kv�2 and Kv�3 the cytoskeleton in regulating the expression ofIto. For example, insulin-triggered synthesis of Itosubunits can specifically increase the rate of in-
activation of human and ferret Kv1.4 currents,32,38 channel proteins and augmentation of Ito in ratventricular myocytes was prevented by disruptionwhile Kv�2 also affects the level of its expression,
which appears to depend critically on the ox- of the actin microfilament network or microtubules,suggesting cytoskeletal involvement in traffickingidoreductase activity of the �-subunit.44 In contrast,
expression of Kv�1 or Kv�2 subunits does not affect of newly synthesized K+ channels to the plasmamembrane.55 Consistent with a role of the cyto-Kv4.3 channel gating, but does increase current
density and protein expression, which is dependent skeleton in affecting Ito, Kv4.2 subunit has beenshown to be directly linked to the scaffold protein,upon the presence of the C-terminus of Kv4.3.40
Kv channels can also be regulated by KChAP filamin, which mediates its interaction with theactin cytoskeleton resulting in upregulation of Kv4.2(Kv channel-associated protein), a member of the
transcription factor binding protein family, by in- currents56 while the Kv�1.1 subunit associates withcytoskeleton by its amino terminus.57,58creasing the functional cell-surface expression of
specific Kv channels without affecting other channelproperties.45–47 KChAP produces these effects byinteracting with the N-termini of Kv�2 and Kv�1 Mechanisms of Ito inactivationsubunits and the C-termini of Kv�-subunits.45 Co-expression with KChAP increases the amplitude of Inactivation is a hallmark feature of Ito currents in
the heart and refers to the process whereby channelKv1.3, Kv2.1, Kv2.2, and Kv4.3 currents without affectingthe channel kinetics and gating,45,46 suggesting that opening following depolarization leads to con-
formational changes resulting in a stable non-con-KChAP is a true chaperone. Complexes of KChAPand Kv4.3 have been co-immunoprecipitated from rat ducting (inactivated) state of the channel from
which the channel is incapable of reopening unlesshearts implying that KChAP may be an importantregulatory subunit for the native Ito in the heart.46 the membrane potential is repolarized. Recent stud-
ies of the K+ channel have elucidated three mech-Kv channel-interacting proteins (KChIPs) areCa2+-binding proteins that interact with the cyto- anistically different but related types of inactivation,
N-type, C-type and V-type.59,60 N- and C-type in-plasmic amino termini of Kv4 �-subunits and re-constitute several features of native Ito current by activation occurs in Kv3.4,61 Kv1.7
6,22 and Kv1.459,62,63
channels while V-type inactivation occurs in Kv4.2modulating the density, inactivation kinetics andrate of recovery from inactivation of Kv4 channels.48 and Kv4.3 channels.60 The inactivation of the native
cardiac Ito has been stratified into a slow, Ito,s, andAll three KChIPs (KChIP1, KChIP2 and KChIP3)co-localize and co-immunoprecipitate with the nat- a fast component, Ito,f, with distinct recovery from
inactivation in a wide variety of species and inive Kv4 channel and, as such, are considered integralcomponents of Kv channels.48 KChIP2 has tissue- different regions of the heart4 (see Table 1). For
example, in the mouse myocardium Ito,s and Ito,fspecific expression in the heart.48 Co-expression ofKv4.2 with KChIP1, 2 or 3 increased the density of appear to be encoded by Kv1.4 and Kv4.2/4.3 channels,
respectively4,64 (see Table 1).Kv4.2 currents, indicating that KChIPs may promoteand/or stabilize expression of Kv4.2 at the cell surface. N-type inactivation is the best-understood gating
transition in ion channels. It occurs by a “ball-and-In addition, co-expression changes the biophysicalproperties of heterologously expressed currents such chain” type mechanism wherein an N-terminal
domain (inactivation gate) binds to its receptor atthat they match almost precisely the Ito recordedin rat myocytes.48–50 Another KChIP-related Ca2+- the inner vestibule of the channel thereby physically
blocking the pore.63,65,66 Inactivation domains arebinding protein, called frequenin, is also expressedin heart and increases both Ito density and alters contributed by either the pore-forming alpha-sub-
units or certain auxiliary beta-subunits.34,35 Giventhe inactivation characteristics.51
The cytoskeleton is an intracellular filamentous the steric hindrance posed by the �-subunit/T1domain (T14�4) on the intracellular side of the pore,network of F-actin (microfilament), microtubules,
and intermediate filaments.52 Cytoskeletal changes inactivation peptides appear to gain access to the

Structure and Function of Cardiac Ito 855
pore via lateral openings in the T14/�4 macro- Functional Role of Itodomain.37
A second type of slower inactivation, termed C- Molecular basis for heterogeneity of Ito in the hearttype inactivation, has also been elucidated.59,67 Thistype of inactivation involves conformational Understanding the molecular basis for Ito expression,changes in the outer mouth of the pore.68–71 For function and regulation is central for the under-many channels, C- and N-type inactivation co-exist standing of normal and abnormal cardiac elec-and are, in fact, kinetically linked. Indeed, in the trophysiology. Ito currents can broadly be classifiedabsence of N-type inactivation, entry into the C- into two separate types although considerable di-inactivated state is extremely slow but appears to versity and discrepancies remain to be resolved.occur quickly in the presence of the N-terminal The “rapid” or “fast” Ito, referred to as Ito,f, and theinactivation ball for Kv1.4 channels69 and Shaker slower form of Ito, referred to as Ito,s, have verychannels, the fruit fly homologue of Kv1.4.65,66 Clear rapid activation (�a≈2–10 ms) but differing ratesevidence for the interplay between N- and C-type of inactivation, �i≈25–80 ms and 80–200 ms, andinactivation has been well documented in Kv1.4 recovery from steady-state inactivation,channels where the recovery from inactivation is �rec≈25–80 ms and 1–2 s, respectively.6,49,77–79,80
controlled by exit from the C-type inactivated state These distinct forms of Ito can be further distin-while the fast rate of inactivation is determined by guished on the basis of their differential sensitivityN-type inactivation.69 All members of the Kv4 family to blockade by toxins such as heteropodatox-show rapid inactivation and recovery from in- ins,80–82 hanatoxin83,84 and phrixotoxins85 with vary-activation.59 Simultaneous N-and C-terminal de- ing degrees of specificity. In addition, several phar-letions of the Kv4.1, but not individual deletions, macological agents like flecainide, quinidine andresulted in very modest slowing of inactivation,72 4-amino-pyridine (4-AP) can also block Ito.83,86
suggesting that structural interaction between the Marked differences in the densities of Ito,f andN- and C-termini is required for inactivation in Kv4.1 Ito,s exist between different species as well withinchannels. More recent work has definitively shown different regions in the same heart. Ito,f has beenthat inner vestibule residues not associated with identified in adult ventricular and atrial myocytesthe P-loops of Kv4.1 channel pore are critical for this from most species and has been reasonably wellunique inactivation, called V-type inactivation.60 studied in rat, mouse, human, dog and fer-
The interaction of Kv-subunits with a variety of ret.2,4,82,87 By contrast, rabbit atrial and ventricularancillary proteins can have profound effects on myocytes appear to produce primarily Ito,s cur-the inactivation kinetics (see above). In addition, rents.9,88,89 Ito,f density in rats and Ito,s density inphosphorylation of Kv subunits and/or subunit pep- rabbits are higher in atrium than in ven-tides provides another mechanism for regulating tricle,9,90,91 whereas Ito,f density in mouse showsthe inactivation of Ito. Ca2+/Calmodulin-dependent the opposite pattern.78 Regional variations in Ito,f
protein kinase II (CaMKII) are abundant in the density within the ventricle also appears in can-mammalian heart, with �-CaMKII being the pre- ine,92 cat,93 ferret,82 human,15,94 mouse77,78 anddominant isoform.73 CaMKII-mediated phos- rat95–97 hearts. Many species with significant Ito,f
phorylation of the NH2-terminus of Kv1.4 slows its express Ito,s either within the same cell or inrate of inactivation and accelerates the recovery different myocytes isolated from various regionskinetics.74 The inactivation of Ito,f is also slowed of the heart. Although regional differences in theby CaMKII in human atrial myocytes.75 In Kv3.4 expression of Ito,f and Ito,s exist, the general patternchannels, PKC-mediated phosphorylation of several observed is that Ito,s is preferentially expressed inN-terminal residues leads to reduced inactivation.61, regions of the heart with longer APs, such as76 Kv�1, �2, and �3-subunit protein sequences con- the septum, left ventricular endocardium andtain multiple putative PKA and PKC phos- apex of the heart, while Ito,f expression is favoredphorylation sites, which may modulate its role in in epicardial regions, the right ventricle and themediating inactivation of the Kv sub- base of the heart. Furthermore, the current densityunits.5,32,35 Although Ito has been traditionally con- of Ito,f when present appears to be far greatersidered to be a Ca2+-independent current, the as- than the corresponding levels of Ito,s. This pattern ofsociation of Kv subunits with calcium-binding Ito heterogeneity essentially contributes to regionalproteins such as KChIPs and frequenin as well as variations of AP profiles. This in turn leads to atheir regulation by Ca2+-dependent enzymes sug- more synchronous repolarization throughout thegests that functional Ito currents in heart might in ventricle1,98,99 and might also provide a mechanism
for regional modulation of contractility.100fact be regulated by Ca2+.48,74,75

G. Y. Oudit et al.856
Regional heterogeneity of AP profiles and Ito have against Kv4.2 or Kv4.3,116 while only AODNs targetedagainst Kv4.2 attenuated Ito,f in rat atrial myocytes.50been well documented in human,94 ferret,82 rat,96
rabbit88 and mouse64,77,80 ventricles (Table 1). For Additional corroborative, but circumstantial, evi-dence for contribution of both Kv4.2 and Kv4.3 to Ito,fexample, in right ventricular or left ventricle apical
myocytes only Ito,f is present, while left ventricular in rat ventricle has been provided by expressionstudies at both the transcriptional and translationalseptal myocytes express Ito,s alone or a combination
of Ito,f and Ito,s.78 In addition, mouse Ito,f density is levels.49,96,113,118,120,121
Further evidence for the contribution of Kv4.Xsignificantly higher in apical than septum cells.78
Similarly, Ito,f and Ito,s in rat appear simultaneously currents to Ito,f has been provided in studies usingadenoviral transfection of dominant-negative Kv4.2in most myocytes although the relative level of Ito,f
compared to Ito,s is far greater in the epicardial mutants in adult myocytes.117 Similar methods wererecently used with neonatal myocytes to establishregions than septum and endocardium.96,97 For lar-
ger mammals like human, ferret and dog, Ito,s is that both Ito,f and Ito,s could be selectively eliminatedby adenoviral transfection with dominant-negativepreferentially expressed in left ventricular en-
docardial cells compared to the left ventricular mutant Kv4.2 (Kv4.2N) and mutant Kv1.4 (Kv1.4N) pro-teins, respectively.119 In transgenic mice expressingepicardial and right ventricular myocytes.82,94,101–103
Substantial changes in Ito density and its expression dominant-negative mutants of Kv4.2 (Kv4.2W362Fand Kv4.2N), Ito,f was largely abolished in atrium78pattern also occur during the postnatal de-
velopment in rats and dogs.104–109 For example, Ito and ventricle118,122 (see Table 2). Despite these find-ings, the issue of whether Kv4.2 or Kv4.3 or bothdensities are about two-fold lower in neonatal rat
ventricular mocytes compared with adult myo- genes are primarily responsible for Ito,f in mouseremains unresolved. Strong support for the con-cytes.106 Moreover, in neonatal myocytes Ito,s and
Ito,f contribute almost equally to the Ito density,105,106 nection between Ito,s and Kv1.4 in the mouse wasrecently generated in myocytes isolated from thewhich contrasts with adult rat myocytes where
Ito,s contributes less than 10% to total Ito.106 ventricular septum of Kv1.4 knockout (Kv1.4−/−)
transgenic mice115 and double transgenics createdAs already mentioned, a number of candidateKv� clones are known to generate Ito-like currents by crossing the Kv4.2(W362F) and Kv1.4
−/− mice.64
Finally, in human atrial myocytes Ito,f was at-when heterologously expressed. Specifically, Kv1.4,Kv1.7, Kv3.4 (low level), Kv4.2 and Kv4.3 clones are tenuated following exposure to Kv4.3 AODNs but
not Kv4.2 AODNs,89 which is consistent with theexpressed in the myocardium6,22 (see Table 1). Basedpurely on their biophysical properties Kv4.2 and/ observation that Kv4.3 is the predominant Ito-en-
coding subunit in humans.23–25 Less direct evidenceor Kv4.3 channels could theoretically contribute tocardiac Ito,f while Kv1.4 and Kv3.4
61 and Kv1.722 have supporting the hypothesis that Kv1.4 encodes for Ito,s
and Kv4.2/3 encode for Ito,f has been derived frommany properties in common with Ito,s. However, theability to relate specific Kv� genes to the distinct studies examining the relationship of the expression
patterns of Kv1.4, Kv4.2 and Kv4.3 with the magnitudecomponents has been hampered by inherent com-plexities associated with modulation of Ito channel of Ito,s and Ito,f. For example, regional variations in
ferret left ventricular epicardial and endocardialfunction by heterotetramer assembly, interactionswith accessory subunits, phosphorylation, electrical myocytes show clear relationships between regional
differences in the expression of Kv1.4 and Kv4.2/Kv4.3activity and other possible cellular factors. Never-theless, the general emerging consensus is that with Ito,s and Ito,f, respectively.82 Similarly, regional
differences in Ito,f and Ito,s in rat ventricles are closelyIto,s is encoded primarily by Kv1.4 channel genes,while Ito,f is generated principally by Kv4.2 and/or connected to the differential expression of Kv4.2/Kv4.3
and Kv1.4.96Kv4.3 genes. This assertion is consistent with earlywork suggesting that Kv1.4 encodes for cardiac The underlying basis for quantitative and qual-
itative differences in the endogenous expressionIto110,111 possibly as a heteromultimer with other
Kv1.X subunits112 as well as the later suggestion that pattern of the Kv1.4 and Kv4.2/3 is unknown, althoughcell-specific repressor elements for Kv� channelsa primary determinant of cardiac Ito is Kv4.2.113
Much more definitive evidence for the Kv� chan- have been described.123 A multitude of factors are,however, known to potentially contribute to thisnels underlying Ito,f and Ito,s has emerged recently
particularly in the mouse atrium114,115 and vent- Ito diversity, including hetero-tetrameric �-subunitformation within a gene family,5,112,124 alternativericle,77,78 rat ventricle,116–119 and the rabbit and
human atrium.89 Specifically, Ito,f is significantly splicing,23,24 post-translation modifications such asglycosylation and phosphorylation,6,125 interactionattenuated in rat ventricular myocytes exposed to
antisense oligodeoxynucleotides (AODNs) targeted with the cytoskeletal network and/or ancillary sub-

Structure and Function of Cardiac Ito 857
agonists and angiotensin II (Ang-II) have beenshown to modulate Ito. For example, acute ap-plication of �-adrenergic agonists causes rapid Ito
reduction in adult rat ventricular myocytes,130–132
rabbit atrial133–135 and ventricular136 myocytes viaPKC-dependent130,137 and independent133 pathways.However, recent evidence suggests that one of theagonists used in these studies, methoxamine, dir-ectly blocks Ito
138 thereby leading to an over-estimation of the ability of �-adrenergic stimulationto inhibit Ito.88,132–134 The acute effects of �-ad-renergic stimulation on Ito current density alsodisplay species specificity. For example, acute ap-
Figure 1 Kv1.4 and Kv4.2 protein levels in rabbit and rat plication of phenylephrine failed to inhibit canineright ventricular (RV) wall. Using specific anti-Kv1.4 andventricular epicardial Ito while purkinje myocyte Itoanti-Kv4.2 antibodies, representative gels from Westernwas inhibited.139 The acute reductions in Ito in rabbitblots are shown for Kv1.4 (A) and Kv4.2 (B) levels in samples
obtained from adult rat brain (positive control), adult atrial myocytes following �-adrenergic activationrabbit RV wall, and adult rat RV wall. Equivalent amount coincided with the observations that phorbol ester,of protein from each specimen was loaded; multiple bands an activator of PKC, produces biphasic effects onin a single column represent glycosylation products (see
currents in oocytes expressing Kv1.4 channels with96).an initial 14% increase followed by a 40% reductionin peak Ito.140 However, whether these changesdepend on in vivo phosphorylation of Kv1.4, as shownunits such as �-subunits, frequenin, KChAP and/in vitro, remains to be determined.140
or KChIP proteins (see above), local (autocrineAs for acute applications of �-adrenergic agonists,and/or paracrine) effects of thyroid hormone and
chronic exposure of cultured neonatal rat vent-angiotensin II,106,126–128 and changes in PKA, PKCricular myocytes to the �-adrenergic agonist,and CaMKII-mediated phosphorylation.55,74,75,129 Al-phenylephrine (PE) also produced reductions inthough it is generally accepted that Kv4.2/4.3 encodesIto
137 that were subsequently shown to be associatedfor Ito,f and Kv1.4 is related to Ito,s, there are ex-with reductions in Kv4.2 and increased in Kv1.4 pro-perimental findings which suggest that our under-tein.141 These changes are particularly relevantstanding is far from complete. For example, rabbitsince similar changes in K+ channel expressionIto shows characteristic features consistent withoccur in heart disease (see below) and increasedthis current being entirely generated by Kv1.4�-cardiac adrenergic stimulation is thought to con-subunits.9,49,79 However, treatment of cultured rab-tribute to disease progression. This observationbit atrial myocytes with ADONs targeted againstraises the intriguing question of whether reductionsKv1.4 resulted in only partial reduction of Ito
89 within Ito are mechanistically linked to the chroniceven large Ito reductions being produced by ADONsstimulatory effects from catecholamines. Althoughagainst Kv4.2 and Kv4.3. Our western blot analysis ofchronic application of endothelin-1 to cultured neo-rabbit myocardium (Fig. 1) suggests further thatnatal rat ventricular myoctes resulted in reducedlarge levels of Kv4.2 as well as Kv1.4 are expressed inKv4.2 protein levels,141 acute application of en-rabbit heart. It also seems likely that other Ito-dothelin-1 (End-1) resulted in no change in Itoforming channels might also contribute to cardiacdensity in adult142 and neonatal ventricular myo-Ito. Specifically, despite the fact that the Kv1.7 channelcytes (Kassiri and Backx, unpublished data).has been shown to be expressed in heart at a high
Acute PKC activation also leads to Ito reduction inlevel, no definitive role for this gene in cardiac Itorat ventricular myocytes and in oocytes expressinghas been established.22
Kv4.2 and Kv4.3 channels143 which does not requireprotein tyrosine kinase (PTK) phosphorylation.143
This PKC-mediated effect was not accompanied bychanges in the biophysical properties of Ito in theseRegulation of Itostudies.143 The modulation of Ito by PKC might bean important mechanism for the reduction of Ito inVarious hormonal and/or paracrine systems have
been shown to modulate the expression and mag- diseases since PKC inhibition was able to reversethe reductions in Ito observed in rat myocytes de-nitude of Ito. In particular, the G-protein-coupled
receptor agonists, endothelin-1, �1-adrenergic rived from hypothyroid126,144 and diabetic126,145

G. Y. Oudit et al.858
hearts.55 In contrast to the results in rat, PKC Changes in fatty acid metabolism have beenlinked to increased arrhythmogenesis and myo-activation has no effect on Ito in normal and failing
canine ventricular myocytes. As such, the shorter cardial dysfunction during myocardial ischemia andin diabetic hearts.151,152 Fatty acids (and their meta-spliced variant of Kv4.3 that presumably lacks a PKC
phosphorylation site encodes for functional Ito in bolites) and arachidonic acid inhibit Ito density withprolongation of the AP in rat ventricular myo-canine ventricle.25
Ang-II has also been shown to have direct effects cytes.142,153,154 The Kv4.3 current underwent a similarreduction in response to a n-3 polyunsaturatedon Ito in the myocardium. Incubation of canine epi-
cardial ventricular myocytes with Ang-II reduces Ito fatty acid.155 The Ito-suppressing activity ofarachidonic acid was PKC-dependent142 but in-density but more importantly transforms the epi-
cardial fast Ito, assumed to be Kv4.3-based,25,102 into dependent of the intracellular cyclooxygenase path-way.153 Although multiple other putative regulatorya current resembling slow Ito, typical of the canine
endocardium.128 Indeed, changes in biophysical factors of Ito have been described, such as glu-cocorticoid (dexamethasone) exposure,156 acido-properties of these currents resemble closely the dif-
ferences observed between Kv4.X-based currents and sis,157 and oxidative stress,87 their precise physio-logical roles remain to be defined.Kv1.4-based currents.49,80 Remarkably, the Ang-II-
mediated effect occurred without measurablechanges in Kv1.4 and Kv4.3 mRNA levels. Conversely,treatment of the endocardial myocytes with the AT1 Role of Ito in modulating action potential profile and
Ca2+ handlingblocker, losartan, converted slow endocardial Ito intoa much faster Ito resembling that observed in epi-
The influence of Ito on AP profile differs remarkablycardium.128 These observations have profound im-plications in the understanding of Ito by suggesting between different species. Differences in AP duration
resulting from variations in Ito depend critically onthat biophysical properties of Ito can be changed dra-matically without alterations in expression. the kinetics and magnitude of the other repolarizing
and depolarizing currents that are present.11,158Thyroid hormone, a member of the steroid hor-mone superfamily, has profound effects on Ito cur- Moreover, the complex interplay between the vari-
ous currents in myocytes can lead to changes inrents. In rat hearts, hypothyroidism prolongs APduration in conjunction with reduced Ito density, AP profile in response to variations in Ito that are
not readily apparent11 (see Fig. 2). Two currents ofdecreases Kv4.2 expression and increases Kv1.4
levels146 which is likely responsible for slowing Ito particular interest in all species when consideringthe net effects of Ito changes on AP profile are ICa,Lrate of recovery from inactivation.127 Conversely,
hyperthyroidism in rat ventricular myocardium in- and INa/Ca, because of their importance in regulatingmyocardial contractility and in generating arrhy-creases Ito current density with elevated Kv4.2 and
Kv4.3 expression and reduced Kv1.4 expression.106,127, thmias such as early and delayed after de-polarizations (EADs and DADs).141,146 The thyroid hormone T3 (tri-iodothyronine)
may be responsible for the postnatal changes in AP In mice and rats Ito dominates repolarization andis largely responsible for the brief triangular APduration, which have been linked to changes in
the transcriptional and protein patterns Kv1.4, Kv4.2 profile.4,78,106,159 Accordingly, numerous studieshave established that Ito reductions in rat97,116,120,and Kv4.3 genes.106,126,127,141,144,146,147 Indeed, in rat
myocardium, changes in pattern of expression of 160–165 (reviewed in 14) and mouse myocardium77,78,
118,122,166 cause prolongation of the AP during allIto-based genes following treatment with T3 mimicclosely the developmental changes.106,127,141 This phases which leads to enhanced myocardial con-
tractility in transgenic mice.118 Moreover, 4-amino-observation suggests that developmental changesin thyroid status play an important role in the age- pyridine (4-AP)-induced prolongation of the AP
leads to elevated contractility in the working mousedependent electrical remodeling that occurs,106,127
although the expression of many other proteins that heart model.167 In myocytes derived from normaland hypertrophied rat hearts, AP prolongation fol-affect electrical properties in heart are influenced by
thyroid hormone.148–150 This pattern of change in lowing Ito reduction has been shown to increaseL-type Ca2+ influx, enhance peak Ca2+ transientIto-encoding genes readily explains the changes in
Ito recovery kinetics which underlies the dis- amplitude and augment unloaded cell shorten-ing.12,13,120 This augmentation of Ca2+ transientappearance of rate-dependent changes in action
potential duration that occur during develop- amplitude occurs in response to a 75% reductionin Ito but not with a comparable reduction in thement,106 suggesting further that Kv1.4 is linked to
Ito,s while Kv4.2/3 encodes for Ito,f. inward rectifier current (IK1) as illustrated in Figure

Structure and Function of Cardiac Ito 859
Figure 2 Numerical simulations of a canine cardiac AP with varying amounts of Ito conductance. The top row showsmodel APs paced at 1 Hz under control conditions (left), and with Ito conductance reduced by 50% (middle) and 90%(right). APD90 (ms) is 173, 203, and 199, and APD50 (ms) is 52, 92, and 88, respectively from left to right.
3. The underlying basis for the potentiation of ICa,L Ito clearly influences the trajectory of early re-polarization of the AP in large mammals,102,103,169–172can be traced to the effects of Ito on the trajectory of
early repolarization of the AP and its close temporal but the net effect on late repolarization is complex.Although some earlier studies from human andassociation with the L-type Ca2+ channel (LCC)
gating.12 Indeed, slowing of early repolarization canine ventricular cells showed an association be-tween reductions in Ito and overall AP prolonga-increases net Ca2+ entry via ICa,L which, in turn,
enhances excitation–contraction coupling and sec- tion,102,170,173,174 more recent work in the rabbitMI model,171 canine rapid pacing model,169 and inondarily helps load the sarcoplasmic reticulum with
Ca2+.12,100,168 Collectively, these results also support computer modeling studies10,175,176 suggest that theconsequences of Ito reductions depend critically ona role for changes in Na+/Ca2+ exchanger activity
under conditions where Ito is reduced in rat myo- complex interactions of Ito with ICa,L, INa/Ca, anddelayed rectifier currents. For example, in the can-cytes (see 120, although this has not been previously
studied in a systematic manner). Model com- ine ventricular AP model, reductions in Ito levelsobserved in diseased myocytes had minimal effectsputations of a canine mid-myocardial cell have
nicely illustrated the complex interactions of Ito on AP duration after phase 1 repolarization innormal myocytes, but these Ito reductions producedwith ICa,L and INa/Ca as well as their effects on Ca2+
cycling.11,169 massive prolongation in diseased myocytes whenchanges in Na+/Ca2+ exchange activity as observedIn larger mammals, the role of Ito in setting the
profile of the action potential is more controversial. in disease were included.169,176 Similarly, in a canine

G. Y. Oudit et al.860
Figu
re3
[Ca2+
] itr
ansi
ents
inn
orm
alra
tve
ntr
icu
lar
myo
cyte
su
nde
req
ual
sarc
opla
smic
reti
culu
mC
a2+
load
sre
cord
edu
nde
rvo
ltag
ecl
amp
con
diti
ons.
Ch
ange
sin
acti
onpo
ten
tial
profi
les
wer
ege
ner
ated
thro
ugh
the
use
ofa
com
pute
rm
odel
ofa
rat
ven
tric
ula
rA
Pby
sele
ctiv
ely
chan
gin
gth
eam
oun
tsof
I toan
dI K
1(i
nw
ard
rect
ifier
curr
ent)
.

Structure and Function of Cardiac Ito 861
atrial AP model,177 reduction in Ito beyond 50% and molecular changes have been found to be eitherregionally dependent100,184 or regionally in-does not cause further AP prolongation (Fig. 2)
because of increased voltage-dependent recruitment dependent.181 By contrast, Ito,s as well as Kv1.4 mRNAand protein expression are increased in the ratof the delayed rectifier currents, IKR and IKS. More-
over, using the human atrial AP model,10 APD90 infarct model at 6–8 weeks but not at 3 weekspost-infarction120,121 and in renovascular hyper-is significantly reduced following 50% and 90%
reduction in Ito (APD90=299, 275 and 259 ms at tensive rats,178 suggesting that transcriptional con-trol of Kv1.4 is likely to be both time- and model-100%, 50% and 10% of control Ito, respectively).
On the other hand, simulations in both failing and dependent. A recent transgenic murine model ofheart failure has demonstrated a marked down-non-failing human ventricular myocyte models11
predict that a direct connection between reductions regulation of Ito associated with AP prolongationduring the hypertrophy stage.166in Ito and AP prolongation, as observed in rodents,
occurs when Ito densities are large, as might be the The electrophysiological changes in heart diseasehave also been studied in two canine models. Likecase for human epicardial myocytes.94,170
the rat, subendocardial canine myocytes survivinginfarction show AP prolongation as well as re-ductions in Ito density and changes in its kineticswithin 2–5 days of infarction.186–188 Unfortunately,Role of Ito in Disease Statesno molecular correlates for these electrical changeshave been investigated. In the pacing-induced can-Downregulation of Itoine cardiomyopathy model, AP prolongation hasbeen associated with reductions in fast Ito,f in bothA central and consistent electrophysiological
change in cardiac hypertrophy has been the reduc- the ventricular102 and atrial myocytes.189 The re-ductions in Ito,f in canine ventricle from this modeltion in Ito, namely Ito,f with concomitant changes in the
level of expression of the Kv�-subunits.25,102,103,120,178 are due entirely to reductions in the shorter splicedvariant of Kv4.3.25 Unfortunately, despite measurableCompared to normal patients, myocytes derived
from terminal heart failure patients have prolonged expression of Ito,s in the dog endocardium128 andsignificant increases in Kv1.4 levels in the myo-AP duration and reduced Ito densities in the epi-
cardial myocytes94,170,179 without significant dif- cardium of paced dogs,25 the changes of Ito in theendocardium of the pacing dog model remains toferences in endocardial myocytes.94 These electrical
differences occur without significant kinetic or be determined. In addition, pacing-induced failureis associated with decreased Ito and AP prolongationsteady-state gating changes in the properties of
Ito,180 which are fast in the epicardium (Ito,f) and in rabbits.172,190
As discussed above, the mechanism for Ito down-slow in the endocardium (Ito,s).94 The Ito,f reductionsin the epicardium are linked to decreased mRNA regulation and loss of transmural heterogeneity in
myocardial hypertrophy and heart failure is likelyexpression of Kv4.3, consistent with this protein en-coding for Ito,f in human heart.103 mediated by a wide array of changes in neuro-
humoral and paracrine factors, regulatory subunitsIn the rat ventricle, there are temporal changesin the biophysical properties and expression of Ito and/or cytoskeleton and cell metabolism. A clear
example of the importance of neurohumoral ac-following myocardial infarction, leading to AP pro-longation. Ito and Kv4.2 and/or Kv.3 protein levels are tivation in mediating dysregulation of Ito is the
ability of the angiotensin receptor (AT1) antagonist,decreased while APs are prolonged within 3 daysin the left non-infarcted myocardium163 (Kaprielian losartan, and ACE inhibitors to reverse the changes
in AP duration and Ito density in spontaneouslyand Backx, unpublished data). At 3–8 weeks post-infarction, Ito,f and Kv4.2/3 expression levels are fur- hypertensive rats.182,183 Moreover, the use of an
ACE inhibitor in renovascular hypertensive ratsther decreased in the non-infarcted left121 and rightventricle120 and these changes persist at 4–6 months completely restores Kv4.2 and Kv4.3 mRNA levels,
178 and short exposure to Ang-II modulates thepost-infarction.161,181 Similar reductions in Ito andAP prolongation have been observed in other rat biophysical characteristics of Ito in canine vent-
ricular myocytes.128 In the pacing-induced caninemodels of heart disease in spontaneously hyper-tensive rats,182,183 catecholamine toxicity,184 aortic- model of heart failure, a local renin–angiotensin
system (RAS) is upregulated with increased localbanded rats,97 and monocrotaline-induced rightventricular hypertrophy.185 The expression levels of production of Ang-II.191 The important patho-
physiological role for the intra-cardiac and systemicKv4.2 and Kv4.3 mRNA were reduced in renovascularhypertensive rats.178 Furthermore, these electrical RAS191–193 is highlighted by clinical data which

G. Y. Oudit et al.862
show that ACE inhibitors and AT1 receptor blockers Nevertheless, changes in early repolarization inresponse to decreases of Ito will influence the levelreduce morbidity and mortality, particularly from
sudden cardiac death, in patients with congestive of activity of the L-type Ca2+ channel (LCC), Na+/Ca2+ exchangers and other currents.11,12,100,171 APheart failure.194–197 It is intriguing to speculate
whether the protective effects of these agents are profile is an important regulator of intracellularCa2+ handling11,176,211–214 and primarily involvesrelated, in part, to effects on Ito.modulation of the LCC and electrogenic Na+/Ca2+
exchanger. In the rodent heart, prolonged de-polarization enhances Ca2+ entry via the LCC andLinking Altered Ito Physiology to Ca2+ Dysregulation and
Arrhythmogenesis retards Ca2+ extrusion via the Na+/Ca2+ exchanger,thereby loading the cell with Ca2+ and enhancing
About 50% of heart failure patients die from sudden SR Ca2+ release.12,13,215 Since increases in the amp-litude of the [Ca2+]i transient exert a positive ino-cardiac death,198 often as a result of ventricular
tachycardia or fibrillation.199,200 Heart failure and tropic effect, it seems plausible that the reductionin K+ channel expression and AP prolongationmyocardial hypertrophy are associated with elec-
trical remodeling and defects in repolarization in- represents an early compensatory response of theheart to increased demand. These membrane-de-volving a wide and complex array of changes in
ion channels, transporters and calcium-handling pendent changes in Ca2+ homeostasis can also beimportant determinants of gene expression in aproteins.201 In addition, changes in Ito may have a
pro-arrhythmogenic role in iron-overload (hemo- number of excitable cells and might be linked todisease pathogenesis.218–220 A cardiac-specific over-chromatosis) cardiomyopathy,202 atrial fib-
rillation203 and drug-induced arrhythmias.204,205 expression of the �-subunit of the LCC, which res-ulted in increased ICa,L current density, myocardialCurrently, the relationship between these ab-
normalities in repolarization and arrhythmias, hypertrophy and an age-dependent cardio-myopathy, has highlighted the pathophysiologicalcausing sudden death, are still poorly understood.
AP profiles in normal and disease myocytes depend importance of the LCC.216,217 Elevated [Ca2+]i tran-sients have also been recorded in rodent models ofon a delicate balance between depolarizing (L-type
Ca2+, ICaL, sodium, INa, and sodium-calcium ex- moderate cardiac hypertrophy120,162 and in 3–4-week-old transgenic mice over-expressing Kv4.2N tochanger, INa/Ca) and hyperpolarizing (Ito, IK, IK1, ICl
and Na+/K+-ATPase) currents. There is increasing reduce Ito,f.118 In addition, changes in gene ex-pression induced by neurohumoral factors (no-evidence that K+ current downregulation con-
tributes significantly to altered repolarization in repinephrine, End-1 and Ang-II) and the activationof their corresponding signaling cascades have allheart disease.99,206 As already discussed, Ito density
appears to be reduced in hypertrophied and failing been shown to be dependent on Ca2+.221–230
AP prolongation also predisposes the heart tohearts regardless of the species, precipitating factorsor the stage of the disease. characteristic EADs and/or DADs.231–233 EADs arise
from an imbalance between depolarizing currentsDecreased Ito would, therefore, appear to be aplausible contributor to AP prolongation in cardiac (primarily ICa,L with possible contributions from the
Na+/Ca2+ exchanger) and repolarizing K+ currentshypertrophy and failure, although the role of otherrepolarizing or depolarizing currents also needs to during late phase 2 of the AP. DADs on the other
hand are linked to elevated [Ca2+]i and spontaneousbe considered.14,169,207 Indeed, studies in the dogpacing model, in human patients and rat models Ca2+ release leading to membrane depolarization
via the Na+/Ca2+ exchanger.231,234–236 While theof disease have suggested that reductions in re-polarizing currents are critical factors in AP pro- precise contribution of reductions of Ito to AP pro-
longation and arrhythmias remains uncertain inlongation,102,120,160,208–210 despite the fact that Ito
primarily affects the early repolarization period. large mammals, its influence is almost certain tovary regionally and this might exaggerate the dis-While this result seems solid in rodents, as es-
tablished by the recent transgenic results in which persion of refractoriness which predispose to re-entry types of arrhythmias.76,233 By contrast, directIto,f and Ito,s have been eliminated,64,99,118,122 recent
results in the dog pacing model have suggested that links between reductions in Ito, AP prolongationand arrhythmias have recently been demonstratedchanges in other currents generated by inward
rectifier channels and Na+/Ca2+ exchangers are in binary transgenic mice lacking both Ito,s andIto,f.64 These binary mice show evidence for AV nodalessential for explaining AP prolongation.169,207 Spe-
cifically, a bimodal effect on AP duration is observed heart block, and multiple isorhythmic tachycardicventricular dissociation64 which are precursors forwhen Ito density changes (see above).
Structure and Function of Cardiac Ito 863
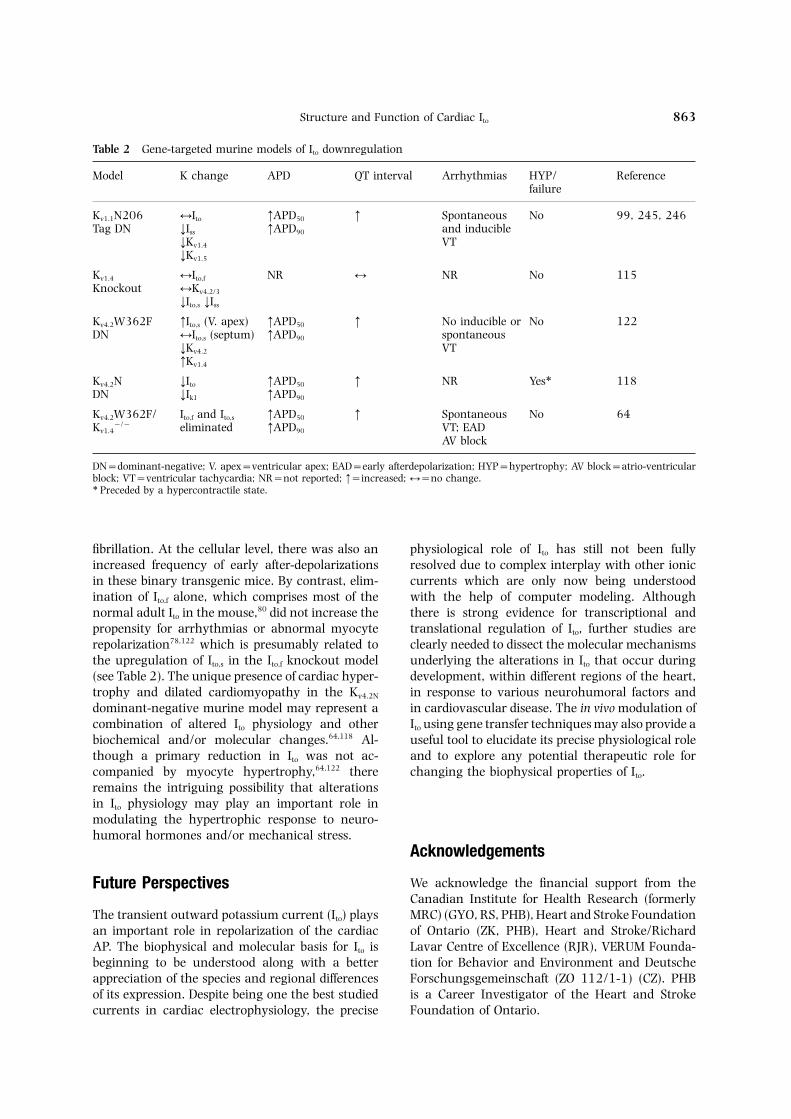
Table 2 Gene-targeted murine models of Ito downregulation
Model K change APD QT interval Arrhythmias HYP/ Referencefailure
Kv1.1N206 ↔Ito ↑APD50 ↑ Spontaneous No 99, 245, 246Tag DN ↓Iss ↑APD90 and inducible
↓Kv1.4 VT↓Kv1.5
Kv1.4 ↔Ito,f NR ↔ NR No 115Knockout ↔Kv4.2/3
↓Ito,s ↓Iss
Kv4.2W362F ↑Ito,s (V. apex) ↑APD50 ↑ No inducible or No 122DN ↔Ito,s (septum) ↑APD90 spontaneous
↓Kv4.2 VT↑Kv1.4
Kv4.2N ↓Ito ↑APD50 ↑ NR Yes∗ 118DN ↓Ik1 ↑APD90
Kv4.2W362F/ Ito,f and Ito,s ↑APD50 ↑ Spontaneous No 64Kv1.4
−/− eliminated ↑APD90 VT; EADAV block
DN=dominant-negative; V. apex=ventricular apex; EAD=early afterdepolarization; HYP=hypertrophy; AV block=atrio-ventricularblock; VT=ventricular tachycardia; NR=not reported; ↑=increased; ↔=no change.∗ Preceded by a hypercontractile state.
fibrillation. At the cellular level, there was also an physiological role of Ito has still not been fullyresolved due to complex interplay with other ionicincreased frequency of early after-depolarizations
in these binary transgenic mice. By contrast, elim- currents which are only now being understoodwith the help of computer modeling. Althoughination of Ito,f alone, which comprises most of the
normal adult Ito in the mouse,80 did not increase the there is strong evidence for transcriptional andtranslational regulation of Ito, further studies arepropensity for arrhythmias or abnormal myocyte
repolarization78,122 which is presumably related to clearly needed to dissect the molecular mechanismsunderlying the alterations in Ito that occur duringthe upregulation of Ito,s in the Ito,f knockout model
(see Table 2). The unique presence of cardiac hyper- development, within different regions of the heart,in response to various neurohumoral factors andtrophy and dilated cardiomyopathy in the Kv4.2N
dominant-negative murine model may represent a in cardiovascular disease. The in vivo modulation ofIto using gene transfer techniques may also provide acombination of altered Ito physiology and other
biochemical and/or molecular changes.64,118 Al- useful tool to elucidate its precise physiological roleand to explore any potential therapeutic role forthough a primary reduction in Ito was not ac-
companied by myocyte hypertrophy,64,122 there changing the biophysical properties of Ito.remains the intriguing possibility that alterationsin Ito physiology may play an important role inmodulating the hypertrophic response to neuro-humoral hormones and/or mechanical stress.
Acknowledgements
We acknowledge the financial support from theFuture PerspectivesCanadian Institute for Health Research (formerlyMRC) (GYO, RS, PHB), Heart and Stroke FoundationThe transient outward potassium current (Ito) plays
an important role in repolarization of the cardiac of Ontario (ZK, PHB), Heart and Stroke/RichardLavar Centre of Excellence (RJR), VERUM Founda-AP. The biophysical and molecular basis for Ito is
beginning to be understood along with a better tion for Behavior and Environment and DeutscheForschungsgemeinschaft (ZO 112/1-1) (CZ). PHBappreciation of the species and regional differences
of its expression. Despite being one the best studied is a Career Investigator of the Heart and StrokeFoundation of Ontario.currents in cardiac electrophysiology, the precise

G. Y. Oudit et al.864
specific K+ channel functions. Annu Rev PhysiolReferences1992; 54: 537–555.
17. X J, Y W, J YN, J LY, L M. Assembly1. A C, S S, L SH, L A,of voltage-gated potassium channels. ConservedK SC, D D JM, G GA, L DW.hydrophilic motifs determine subfamily-specificHeterogeneity within the ventricular wall. Elec-interactions between the alpha-subunits. J Bioltrophysiology and pharmacology of epicardial, en-Chem 1995; 270: 24761–24768.docardial, and M cells. Circ Res 1991; 69:
18. L M, J YN, J LY. Specification of subunit1427–1449.assembly by the hydrophilic amino-terminal do-2. B DM, N JM. Myocardial potassiummain of the Shaker potassium channel. Sciencechannels: electrophysiological and molecular di-1992; 257: 1225–1230.versity. Annu Rev Physiol 1996; 58: 363–394.
19. K JW, P MA, B AM, K GE.3. S DJ. Structure and function of cardiac potas-Modulation of potassium channel gating by co-sium channels. Cardiovasc Res 1999; 42: 377–390.expression of Kv2.1 with regulatory Kv5.1 or Kv6.14. N JM. Molecular basis of functional voltage- alpha-subunits. Am J Physiol 1998; 274: C1501–gated K+ channel diversity in the mammalian myo- C1510.cardium. J Physiol 2000; 525 Pt 2: 285–298. 20. P O. Structural basis of voltage-gated K+ chan-
5. D KK, E SK, T MM, M KT, nel pharmacology. Trends Pharmacol Sci 1992; 13:F SA, P SS, H NN, S DJ, B 359–365.PB, L DM, F A. Molecular physiology 21. D DA, M C J, P RA, Kof cardiac potassium channels. Physiol Rev 1996; A, G JM, C SL, C BT, MK R.76: 49–67. The structure of the potassium channel: molecular
6. C WA, A Y, C J, C A, L D, basis of K+ conduction and selectivity. Science 1998;MC T, M H, N MS, O A, 280: 69–77.P D, S M, V-S M 22. K K, N A, T-C J, D ID,E, R B. Molecular diversity of K+ channels. Ann C G, H CM, C NG, J NA,N Y Acad Sci 1999; 868: 233–285. M H, B B, C M, G-
7. T-F M, C J, M JS, S- GA, C KG. Genomic organization, chro- MC. Molecular biology of K+ channels and mosomal localization, tissue distribution, andtheir role in cardiac arrhythmias. Am J Med 2001; biophysical characterization of a novel mammalian110: 50–59. Shaker-related voltage-gated potassium channel,
8. C DL, R RL, Q Y, S HC. The Kv1.7. J Biol Chem 1998; 273: 5851–5857.calcium-independent transient outward potassium 23. K W, P S, Y T, A MD, Scurrent in isolated ferret right ventricular myocytes. G, T GF. Isolation and characterization ofI. Basic characterization and kinetic analysis. J Gen the human gene encoding Ito: further diversity byPhysiol 1993; 101: 571–601. alternative mRNA splicing. Am J Physiol 1998;
9. G WR, I Y. Comparison of potassium 275: H1963–H1970.currents in rabbit atrial and ventricular cells. J 24. O S, T M, O T, A Y, W M,Physiol 1988; 405: 123–145. G WR, I Y. Molecular cloning and tissue
10. C M, R RJ, N S. Ionic distribution of an alternatively spliced variant ofmechanisms underlying human atrial action po- an A-type K+ channel alpha-subunit, Kv4.3 in thetential properties: insights from a mathematical rat. FEBS Lett 1997; 420: 47–53.model. Am J Physiol 1998; 275: H301–H321. 25. J G, K W, P S, Z MQ, W R, T-
11. G JL, W R, P S, T GF, W GF. Alteration of potassium channel ex-RL. Role of the calcium-independent transient out- pression in heart failure. Circulation 1999; 100ward current Ito1 in shaping action potential mor- (Suppl.): 2236A.phology and duration. Circ Res 2000; 87: 26. N N, P DM. Potassium channel1026–1033. alpha and beta subunits assemble in the endo-
12. S R, R RJ, K R, B PH. plasmic reticulum. J Biol Chem 1997; 272: 3022–Alterations in action potential profile enhance ex- 3027.citation-contraction coupling in rat cardiac myo- 27. T S, T Y, Y K, M Y,cytes. J Physiol 2001; In Press. T J, K I. Voltage-gated K+ channel,
13. B RA, C RB, G WR. Effects of Kv4.2, localizes predominantly to the transverse-axialaction potential duration on excitation-contraction tubular system of the rat myocyte. J Mol Cell Cardiolcoupling in rat ventricular myocytes. Action po- 2000; 32: 1361–1369.tential voltage-clamp measurements. Circ Res 1995; 28. P A, V-S M E, K C,76: 790–801. M H, T B, R B. K+ channel
14. W AD, K R, K Z, T subunit isoforms with divergent carboxy-terminalJN, T R, F GI, B PH. The role sequences carry distinct membrane targeting sig-of action potential prolongation and altered in- nals. J Membr Biol 1997; 159: 149–159.tracellular calcium handling in the pathogenesis of 29. L D, T K, L ES. Surface expressionheart failure. Cardiovasc Res 1998; 37: 312–323. of Kv1 channels is governed by a C-terminal motif.
15. N M. Electrical heterogeneity in the vent- J Biol Chem 2000; 275: 11597–11602.ricular wall and the M cell. Cardiovasc Res 1998; 30. E SK, U VN, S H, K J, B-40: 248–250. PB, T MM. Characterization of a voltage-
gated K+ channel beta subunit expressed in human16. J LY, J YN. Structural elements involved in

Structure and Function of Cardiac Ito 865
heart. Proc Natl Acad Sci USA 1995; 92: 6309– chaperone for specific K+ channels key to re-polarization of the cardiac action potential. Focus6313.
31. E SK, U VN, K J, B PB, on KChAP as a chaperone for specific K+ channels.Am J Physiol Cell Physiol 2000; 278: C863–C864.T MM. A novel K+ channel beta-subunit
(hKv beta 1.3) is produced via alternative mRNA 48. A WF, B MR, B M, C J, L HP,M G, H JW, M KI, Ssplicing. J Biol Chem 1995; 270: 28531–28534.
32. M MJ, C RC, C AL, R BW, T JS, R KJ. Modulation of A-typepotassium channels by a family of calcium sensors.RL, S HC. A novel beta subunit increases rate
of inactivation of specific voltage-gated potassium Nature 2000; 403: 553–556.49. W AD, T RG, L VA, K-channel alpha subunits. J Biol Chem 1995; 270:
6272–6277. R, B PH. Effect of Cd2+ on Kv4.2 andKv1.4 expressed in Xenopus oocytes and on the33. S G, N K, H S, R KJ,
S LE, T JS. Beta subunits promote transient outward currents in rat and rabbit vent-ricular myocytes. Cell Physiol Biochem 1999; 9:K+ channel surface expression through effects early
in biosynthesis. Neuron 1996; 16: 843–852. 11–28.50. B-A E, N JM. Molecular correlates34. P O, L T, B M, R J, B
R, W D, G KP, S AJ, S JF. Functional of the calcium-independent, depolarization-activ-ated K+ currents in rat atrial myocytes. J Physioland molecular aspects of voltage-gated K+ channel
beta subunits. Ann N Y Acad Sci 1999; 868: 344– (Lond) 1999; 517: 407–420.51. N TY, N MS, R B, A M,355.
35. M JR, K Y, T MM. Modulation of C WA. Frequenin, a Ca2+-binding protein, isexpressed in heart and is a novel regulator of Kv4Kv channel alpha/beta subunit interactions. Trends
Cardiovasc Med 1999; 9: 253–258. currents. Circulation 2000; 102 (Suppl.): 432.52. J PA. The cytoskeleton and cell signaling:36. G JM, M S, MK R. Structure of a
voltage-dependent K+ channel beta subunit. Cell component localization and mechanical coupling.Physiol Rev 1998; 78: 763–781.1999; 97: 943–952.
37. G JM, Z M, M S, MK R. Struc- 53. M M, A R, F A, DFD. Cytoskeletal control of rectification and ex-ture of the cytoplasmic beta subunit-T1 assembly
of voltage-dependent K+ channels. Science 2000; pression of four substates in cardiac inward rectifierK+ channels. Faseb J 1996; 10: 357–361.289: 123–127.
38. M K, D B M, W Z, W BA. 54. F SC. Regulation of ion channel dis-tribution at synapses. Annu Rev Neurosci 1993; 16:Molecular cloning and functional expression of a
novel potassium channel beta-subunit from human 347–368.55. S Y, E HS, S D. Insulin stimu-atrium. FEBS Lett 1995; 361: 13–16.
39. R J, H SH, W F, L C, lation of rat ventricular K+ currents depends onthe integrity of the cytoskeleton. J Physiol (Lond)P DN, D JO, P O. Inactivation prop-
erties of voltage-gated K+ channels altered by pres- 1999; 514: 735–745.56. P K, M DM, S A. Localizationence of beta-subunit. Nature 1994; 369: 289–294.
40. Y EK, A MR, L ES, T K. and enhanced current density of the Kv4.2 potassiumchannel by interaction with the actin-binding pro-Kvbeta subunits increase expression of Kv4.3 chan-
nels by interacting with their C-termini. J Biol Chem tein filamin. J Neurosci 2000; 20: 8736–8744.57. J J, P T, S-L D, C D,2001; 276(7): 4839–4844.
41. H SH, R J, W F, P O. T WB, L I. Inactivation of a voltage-dependent K+ channel by beta subunit. ModulationMolecular and functional characterization of a rat
brain Kv beta 3 potassium channel subunit. FEBS by a phosphorylation-dependent interaction be-tween the distal C terminus of alpha subunit andLett 1995; 377: 383–389.
42. L T, B R, I D, P O. Co- cytoskeleton. J Biol Chem 1997; 272: 14021–14024.expression of the KCNA3B gene product with Kv1.5
leads to a novel A-type potassium channel. J Biol 58. N K, M MF, T JS. Differentialinteraction of voltage-gated K+ channel beta-sub-Chem 1998; 273: 35095–35101.
43. U VN, E SK, C A, T units with cytoskeleton is mediated by uniqueamino terminal domains. J Mol Neurosci 1998; 11:MM, S DJ. Functional differences in Kv1.5
currents expressed in mammalian cell lines are due 199–208.59. R RL, M MJ, W S, L S,to the presence of endogenous Kv beta 2.1 subunits.
J Biol Chem 1996; 271: 2406–2412. C DL, B MV, S HC. In-activation of voltage-gated cardiac K+ channels.44. P R, W BA, B AM. Mutations in the
Kvbeta 2 binding site for NADPH and their effects Circ Res 1998; 82: 739–750.60. J HH, S M, C M. In-on Kv1.4. J Biol Chem 2001; 276: 738–741.
45. W BA, Y Q, K YA, A EA, activation gating of Kv4 potassium channels: mo-lecular interactions involving the inner vestibule ofB AM. Cloning and expression of a novel K+
channel regulatory protein, KChAP. J Biol Chem the pore. J Gen Physiol 1999; 113: 641–660.61. B EJ, S RG, S SJ, C M.1998; 273: 11745–11751.
46. K YA, G TI, B AM, W BA. Interactions between multiple phosphorylation sitesin the inactivation particle of a K+ channel. InsightsKChAP as a chaperone for specific K+ channels.
Am J Physiol Cell Physiol 2000; 278: C931–C941. into the molecular mechanism of protein kinase Caction. J Gen Physiol 1998; 112: 71–84.47. A H, M H, K RS. KChAP: a novel

G. Y. Oudit et al.866
62. T-C J, Y JA, B MF, T GN. basis of transient outward K+ current diversity inmouse ventricular myocytes. J Physiol 1999; 521Functional role of the NH2-terminal cytoplasmic
domain of a mammalian A-type K channel. J Gen Pt 3: 587–599.78. X H, L H, N JM. Elimination of the tran-Physiol 1993; 102: 1057–1083.
63. C MB, C DL, R RL, L sient outward current and action potential pro-longation in mouse atrial myocytes expressing aDR, M MJ, Z Y, S HC. Cloning
and characterization of an Ito-like potassium chan- dominant negative Kv4 alpha subunit. J Physiol1999; 519 Pt 1: 11–21.nel from ferret ventricle. Am J Physiol 1994; 267:
H1383–H1395. 79. P KR, N JM. Expression en-vironment determines K+ current properties: Kv164. G W, L H, L B, N JM. Functional
consequences of elimination of Ito,f and Ito,s: early and Kv4 alpha-subunit-induced K+ currents inmammalian cell lines and cardiac myocytes. Pflu-afterdepolarizations, atrioventricular block, and
ventricular arrhythmias in mice lacking Kv1.4 and gers Arch 1999; 437: 381–392.80. X H, G W, N JM. Four kinetically distinctexpressing a dominant-negative Kv4 alpha subunit.
Circ Res 2000; 87: 73–79. depolarization-activated K+ currents in adult mouseventricular myocytes. J Gen Physiol 1999; 113:65. H T, Z WN, A RW. Biophysical
and molecular mechanisms of Shaker potassium 661–678.81. S MC, J JH, H LG,channel inactivation. Science 1990; 250: 533–538.
66. Z WN, A RW. Voltage-dependent gat- K PR, V RA, S NA,M AL. Heteropodatoxins: peptides isolateding of Shaker A-type potassium channels in Dro-
sophila muscle. J Gen Physiol 1990; 95: 29–60. from spider venom that block Kv4.2 potassiumchannels. Mol Pharmacol 1997; 51: 491–498.67. L-B J, H T, H SH, A
RW. Effects of external cations and mutations in 82. B MV, C DL, R RL,M MJ, T JS, N JM, Sthe pore region on C-type inactivation of Shaker
potassium channels. Receptors Channels 1993; 1: HC. Distinct transient outward potassium current(Ito) phenotypes and distribution of fast-inactivating61–71.
68. O EM, Z WN, H T, H potassium channel alpha subunits in ferret leftventricular myocytes. J Gen Physiol 1999; 113:SH, H J, A RW. Cooperative subunit inter-
actions in C-type inactivation of K channels. Biophys 581–600.83. H HM, W E, L Q, R U. FourJ 1995; 69: 2449–2457.
69. R RL, M MJ, C RC, Z different components contribute to outward currentin rat ventricular myocytes. Am J Physiol 1999;Y, C DL, S HC. C-type inactivation
controls recovery in a fast inactivating cardiac K+ 277: H107–H118.84. S KJ, MK R. Hanatoxin modifies thechannel (Kv1.4) expressed in Xenopus oocytes. J
Physiol 1995; 489: 709–721. gating of a voltage-dependent K+ channel throughmultiple binding sites. Neuron 1997; 18: 665–673.70. L Y, J ME, Y G. Dynamic rearrange-
ment of the outer mouth of a K+ channel during 85. D S, D MD, M D, F M, LM. Effects of phrixotoxins on the Kv4 family ofgating. Neuron 1996; 16: 859–867.
71. C NA, F SR, L DE, W GK. potassium channels and implications for the roleof Ito1 in cardiac electrogenesis. Br J Pharmacol4-Aminopyridine binding and slow inactivation are
mutually exclusive in rat Kv1.1 and Shaker potas- 1999; 126: 251–263.86. Y SW, S DJ. Electrophysiological andsium channels. Mol Pharmacol 1994; 46: 1175–
1181. pharmacological correspondence between Kv4.2 cur-rent and rat cardiac transient outward current.72. J HH, C M. K+ channel inactivation
mediated by the concerted action of the cytoplasmic Cardiovasc Res 1997; 33: 540–547.87. W CA, G WR. Ionic mechanism of the effectsN- and C-terminal domains. Biophys J 1997; 72:
163–174. of hydrogen peroxide in rat ventricular myocytes.J Physiol (Lond) 1997; 500: 631–642.73. H B, H H, S W, H D, M-
I, K EG, K P. Differentiation- 88. F D, G WR. Regional variations in actionpotentials and transient outward current in myo-dependent expression of cardiac delta-CaMKII iso-
forms. J Cell Biochem 1998; 68: 259–268. cytes isolated from rabbit left ventricle. J Physiol1991; 442: 191–209.74. R J, L C, P O. Frequency-dependent
inactivation of mammalian A-type K+ channel Kv1.4 89. W Z, F J, S H, P A, N JM,N S. Potential molecular basis of differentregulated by Ca2+/calmodulin-dependent protein
kinase. J Neurosci 1997; 17: 3379–3391. physiological properties of the transient outwardK+ current in rabbit and human atrial myocytes.75. T S, K P, K EG, P Y,
A C, L-L M, M JJ, H Circ Res 1999; 84: 551–561.90. B WA, N JM. A novel type of de-SN. Regulation of the transient outward K+ current
by Ca2+/calmodulin-dependent protein kinases II polarization-activated K+ current in isolated adultrat atrial myocytes. Am J Physiol 1991; 260:in human atrial myocytes. Circ Res 1999; 85:
810–819. H1236–H1247.91. B WA, N JM. Two functionally distinct76. A C. Ion channels and ventricular
arrhythmias: cellular and ionic mechanisms under- 4-aminopyridine-sensitive outward K+ currents inrat atrial myocytes. J Gen Physiol 1992; 100: 1041–lying the Brugada syndrome. Curr Opin Cardiol
1999; 14: 274–279. 1067.92. L SH, A C. Transient outward77. G W, X H, L B, N JM. Molecular

Structure and Function of Cardiac Ito 867
current prominent in canine ventricular epi- expression in rat ventricle. J Physiol (Lond) 1997;cardium but not endocardium. Circ Res 1988; 62: 504: 271–286.116–126. 107. X H, D JE, B DM, T JS, M
93. F T, K S, F N, B AL, JP, MK D, N JM. Developmental ana-M RJ. Potassium rectifier currents differ in lysis reveals mismatches in the expression of K+
myocytes of endocardial and epicardial origin. Circ channel alpha subunits and voltage-gated K+ chan-Res 1992; 70: 91–103. nel currents in rat ventricular myocytes. J Gen
94. N M, B DJ, U P, S- Physiol 1996; 108: 405–419. G. Regional differences in current density and 108. P LM, G RF. Developmentalrate-dependent properties of the transient outward changes of action potential configuration and Itocurrent in subepicardial and subendocardial myo- in canine epicardium. Am J Physiol 1995; 268:cytes of human left ventricle. Circulation 1996; 93: H2513–H2521.168–177. 109. J CD, B PA. Age-related appearance of
95. C RB, B RA, S-S E, outward currents may contribute to developmentalS-C J, G WR. Heterogeneity of differences in ventricular repolarization. Circ Resaction potential waveforms and potassium currents 1992; 71: 1390–1403.in rat ventricle. Cardiovasc Res 1993; 27: 1795– 110. T-C JC, T GN, S A, T1799. MA. Molecular cloning and functional expression
96. W AD, J TJ, K R, B PH. of a potassium channel cDNA isolated from a ratRegional contributions of Kv1.4, Kv4.2, and Kv4.3 to cardiac library. FEBS Lett 1990; 268: 63–68.transient outward K+ current in rat ventricle. Am 111. T MM, K KM, W JA, KJ Physiol 1999; 276: H1599–H1607. H, R DM, G DM. Molecular cloning and
97. V T, N TH, S JH, F J, H characterization of two voltage-gated K+ channelH. Regional alterations of repolarizing K+ currents cDNAs from human ventricle. Faseb J 1991; 5:among the left ventricular free wall of rats with 331–337.ascending aortic stenosis. J Physiol 2001; 530: 112. P S, R S, S DJ, T MM, B443–455. PB. Heteromultimeric assembly of human potas-
98. B MJ. Relation of ventricular repolarization to sium channels. Molecular basis of a transient out-electrocardiographic T wave-form and arrhythmia ward current? Circ Res 1993; 72: 1326–1336.vulnerability. Am J Physiol 1979; 236: H391– 113. D JE, MK D. Quantitative analysis ofH402. potassium channel mRNA expression in atrial and
99. B LC, L B, C BR, K G, S ventricular muscle of rats. Circ Res 1994; 75: 252–G. Enhanced dispersion of repolarization and re- 260.fractoriness in transgenic mouse hearts promotes 114. Y H, MK D, D JE, G J, W R,reentrant ventricular tachycardia. Circ Res 2000; C IS, D P, S A, A86: 396–407.
EP, S EA, H M, R MR. Transient100. V T, N TH, S JH, E H. Re-outward current, Ito1, is altered in cardiac memory.lationship between transient outward K+ currentCirculation 1999; 99: 1898–1905.and Ca2+ influx in rat cardiac myocytes of endo-
115. L B, W DW, H JA, B PB. Theand epicardial origin. J Physiol 1999; 519 Pt 3:transient outward current in mice lacking the841–850.potassium channel gene Kv1.4. J Physiol (Lond) 1998;101. L DW, G GA, A C. Ionic bases509: 171–182.for electrophysiological distinctions among epi-
116. F C, C RB, S Y, G WR. Shal-cardial, midmyocardial, and endocardial myocytestype channels contribute to the Ca2+-independentfrom the free wall of the canine left ventricle. Circtransient outward K+ current in rat ventricle. JRes 1993; 72: 671–687.Physiol 1997; 500: 51–64.102. K S, N HB, C N, O’R B,
117. J DC, N HB, M E. Suppression ofP PH, K DA, M E, T GF. Ionicneuronal and cardiac transient outward currentsmechanism of action potential prolongation in vent-by viral gene transfer of dominant-negative Kv4.2ricular myocytes from dogs with pacing-inducedconstructs. J Biol Chem 1997; 272: 31598–31603.heart failure. Circ Res 1996; 78: 262–273.
118. W AD, L P, S R, H Q, F103. K S, D J, D J, A D, N M,GI, B PH. Targeted expression of a dominant-B DJ, S G, MK D, T-negative Kv4.2 K+ channel subunit in the mouse GF. Molecular basis of transient outwardheart. Circ Res 1999; 85: 1067–1076.potassium current downregulation in human heart
119. K Z, Z C, H RJ, M JD, Jfailure: a decrease in Kv4.3 mRNA correlates with aDC, B PH. Down-regulation of Ito can inducereduction in current density. Circulation 1998; 98:myocyte hypertrophy in neonatal rat ventricular1383–1393.myocytes: manipulation of Kv1.4 and Kv4.2 genes104. MK D. Molecular identity of Ito: Kv1.4 redux.using adenoviral gene transfer. Circulation 2000;Circ Res 1999; 84: 620–622.102 (Suppl.): 91A.105. K MJ, F D. A study of the developmental
120. K R, W AD, K Z, Pchanges in outward currents of rat ventricularTG, L PP, B PH. Relationship between K+myocytes. J Physiol 1990; 430: 37–60.channel down-regulation and [Ca2+]i in rat vent-106. W AD, K R, P TG, Jricular myocytes following myocardial infarction. JOT, B PH. Effects of development and thyroid
hormone on K+ currents and K+ channel gene Physiol (Lond) 1999; 517: 229–245.

G. Y. Oudit et al.868
121. G-J M, H B, J P, -S N. Dif- bit ventricular myocytes. J Physiol 1991; 441: 673–684.ferential expression of voltage-gated K+ channel
137. G JP, H CA, H SR. Elec-genes in left ventricular remodeled myocardiumtrophysiological properties of neonatal rat vent-after experimental myocardial infarction. Circ Resricular myocytes with alpha1-adrenergic-induced1996; 79: 669–675.hypertrophy. Am J Physiol 1998; 275: H577–H590.122. B DM, X H, S RB, N JM.
138. P C, L Q, F D. Non-specific action ofFunctional knockout of the transient outward cur-methoxamine on Ito, and the cloned channels hKv1.5rent, long-QT syndrome, and cardiac remodelingand Kv4.2. Br J Pharmacol 1999; 126: 595–606.in mice expressing a dominant-negative Kv4 alpha
139. R RB, L QY, R MR. Ionic basis forsubunit. Circ Res 1998; 83: 560–567.action potential prolongation by phenylephrine in123. V P, K G. Purification and preliminarycanine epicardial myocytes. J Cardiovasc Elec-characterization of a cardiac Kv1.5 repressor elementtrophysiol 2000; 11: 70–76.binding factor. Circ Res 1999; 84: 937–944.
140. M KT, F SA, D KK, P SS, H NN,124. F JF, C TP, R S, J JL, CS DJ, T MM, B PB. ModulationB, B A. Characterisation of Kv4.3 in HEK293of an inactivating human cardiac K+ channel bycells: comparison with the rat ventricular transientprotein kinase C. Circ Res 1994; 75: 999–1005.outward potassium current. Cardiovasc Res 1999;
141. G W, K K, H M, K I, T J.41: 188–199.Regulation of Kv4.2 and Kv1.4 K+ channel expression125. R DM, K S. From genes to channels:by myocardial hypertrophic factors in culturednormal mechanisms. Cardiovasc Res 1999; 42: 318–newborn rat ventricular cells. J Mol Cell Cardiol326.1998; 30: 1449–1455.126. S Y, S D, G W. Thyroid status and
142. D DS, V W DR, M CS, Bdiabetes modulate regional differences in potassiumM. Arachidonic acid and endothelin potentiate Ca2+
currents in rat ventricle. J Physiol 1995; 488:transients in rat cardiac myocytes via inhibition673–688.of distinct K+ channels. J Biol Chem 1993; 268:127. S Y, F C, C RB, D JE, MK27335–27344.D, G WR. Thyroid hormone regulates postnatal
143. N TY, C WA, V-S D Mexpression of transient K+ channel isoforms in ratE, A M, R B. Modulation of Kv4 channels,ventricle. J Physiol 1997; 500: 65–73.key components of rat ventricular transient out-128. Y H, G J, W H, W R, S S,ward K+ current, by PKC. Am J Physiol 1997;MK D, R MR, C IS. Effects of the273: H1775–H1786.renin-angiotensin system on the current Ito in epi-
144. S Y, S DL. Thyroid status and potas-cardial and endocardial ventricular myocytes fromsium currents in rat ventricular myocytes. Am Jthe canine heart. Circ Res 2000; 86: 1062–1068. Physiol 1995; 268: H576–H583.129. A AE, A JP, Q Y, C RG, P- 145. W DW, K T, S S, A M.
PJ, S JD. Kv4.2 phosphorylation by Abnormalities of K+ and Ca2+ currents in vent-cyclic AMP-dependent protein kinase. J Biol Chem ricular myocytes from rats with chronic diabetes.2000; 275: 5337–5346. Am J Physiol 1995; 269: H1288–H1296.
130. A M, N JM. Alpha 1-adrenergic agon- 146. N A, K F, K K, S H, Tists selectively suppress voltage-dependent K+ cur- J. Effects of thyroid status on expression of voltage-rent in rat ventricular myocytes. Proc Natl Acad Sci gated potassium channels in rat left ventricle. Car-USA 1988; 85: 8756–8760. diovasc Res 1998; 40: 343–351.
131. T N, N H, H Y, E M, K M. 147. N A, K F, K K, Y S,Inhibitory effect mediated by alpha 1-adrenoceptors M Y, S H, T J. Effects of thyroidon transient outward current in isolated rat vent- and glucocorticoid hormones on Kv1.5 potassiumricular cells. Pflugers Arch 1990; 415: 575–581. channel gene expression in the rat left ventricle.
132. F D, B RA. Mechanisms for the posi- Biochem Biophys Res Commun 1997; 237: 521–526.tive inotropic effect of alpha 1-adrenoceptor stimu- 148. P J, B LA, L PR. Thyroidlation in rat cardiac myocytes. Circ Res 1992; 71: hormone regulates hyperpolarization-activated673–688. cyclic nucleotide-gated channel (HCN2) mRNA in
133. B AP, F D, C RB, G WR. In- the rat heart. Circ Res 1999; 85: 498–503.tracellular mechanisms for alpha 1-adrenergic re- 149. H Y, I U, T Y, E H, M M,gulation of the transient outward current in rabbit O K, S T, S K. Thyroid hormoneatrial myocytes. J Physiol (Lond) 1990; 431: 689– stimulates Na+-Ca2+ exchanger expression in rat712. cardiac myocytes. J Cardiovasc Pharmacol 1997; 29:
134. F D, S Y, G WR. A novel effect of 75–80.norepinephrine on cardiac cells is mediated by 150. D WH. Biochemical basis of thyroid hor-alpha 1-adrenoceptors. Am J Physiol 1989; 256: mone action in the heart. Am J Med 1990; 88:H1500–H1504. 626–630.
135. F D, S Y, G WR. Alpha-adrenergic 151. MH J, Y KA, W J, Y GX, C PB.modulation of the transient outward current in Recent insights pertaining to sarcolemmal phos-rabbit atrial myocytes. J Physiol 1990; 423: 257– pholipid alterations underlying arrhythmogenesis277. in the ischemic heart. J Cardiovasc Electrophysiol
136. F D, B AP, G WR. Alpha 1-ad- 1993; 4: 288–310.152. MH J, C MH, H KK, J JH,renoceptors reduce background K+ current in rab-

Structure and Function of Cardiac Ito 869
MC R, K RH. Induction of Ca-in- activating sustained K+ current modulates re-polarization and excitation-contraction coupling independent PLA(2) and conservation of plasmalogen
polyunsaturated fatty acids in diabetic heart. Am J adult mouse ventricle. J Physiol 1997; 504: 557–563.Physiol Endocrinol Metab 2000; 279: E25–E32.
153. B KY, S HA, V TM, 168. C RB, B RA, G WR. Action po-tential duration modulates calcium influx, Na+-L EG. Modulation of the transient outward
current in adult rat ventricular myocytes by poly- Ca2+ exchange, and intracellular calcium releasein rat ventricular myocytes. Ann N Y Acad Sci 1996;unsaturated fatty acids. Am J Physiol 1998; 274:
H571–H579. 779: 417–429.169. O’R B, K DA, T GF, K S, T154. X Z, R GJ. K+ current inhibition by am-
phiphilic fatty acid metabolites in rat ventricular R, M E. Mechanisms of altered excitation-contraction coupling in canine tachycardia-in-myocytes. Am J Physiol 1998; 275: C1660–C1667.
155. S CB, V SM, W BD, T H, duced heart failure, I: experimental studies. CircRes 1999; 84: 562–570.W KR, B JA, Q MR, B SN, C
TJ. Blockade by N-3 polyunsaturated fatty acid of 170. B DJ, N M, E E. Alter-ations of K+ currents in isolated human ventricularthe Kv4.3 current stably expressed in Chinese ham-
ster ovary cells. Br J Pharmacol 1999; 127: 941– myocytes from patients with terminal heart failure.Circ Res 1993; 73: 379–385.948.
156. W L, F ZP, D HJ. Glucocorticoid re- 171. L SE, B JH. Enhanced Na+-Ca2+ ex-change in the infarcted heart. Implications for ex-gulation of cardiac K+ currents and L-type Ca2+
current in neonatal mice. Circ Res 1999; 85: 168– citation-contraction coupling. Circ Res 1997; 81:1083–1093.173.
157. C TW, B MR, S A, I 172. T Y, O T, K K, Y K, L W, LZ, K I. Pacing-induced heart failure causesK, O JM, O’B HA, L R, K K,
O CH. Inhibition of the K+ channel Kv1.4 by a reduction of delayed rectifier potassium currentsalong with decreases in calcium and transient out-acidosis: protonation of an extracellular histidine
slows the recovery from N-type inactivation. J ward currents in rabbit ventricle. Cardiovasc Res2000; 48: 300–309.Physiol 2000; 526 Pt 2: 253–264.
158. C M, R RJ, N S. Ionic 173. P D, G JP, F N. Depressed transientoutward current in single hypertrophied cardio-targets for drug therapy and atrial fibrillation-in-
duced electrical remodeling: insights from a math- myocytes isolated from the right ventricle of ferretheart. Cardiovasc Res 1995; 30: 440–448.ematical model. Cardiovasc Res 1999; 42: 477–489.
159. B DM, T JS, M JP, N JM. 174. W E, A GJ, P H, R U. Tran-sient outward current in human ventricular myo-Differential expression of voltage-gated K+ channel
subunits in adult rat heart. Relation to functional cytes of subepicardial and subendocardial origin.Circ Res 1994; 75: 473–482.K+ channels? Circ Res 1995; 77: 361–369.
160. Q D, Z ZH, C EB, B M, J P, 175. P L, B DJ. Simulation study ofcellular electric properties in heart failure. Circ Res-S N. Cellular and ionic basis of arrhythmias
in postinfarction remodeled ventricular myo- 1998; 82: 1206–1223.176. W RL, R J, J S, M E, O’Rcardium. Circ Res 1996; 79: 461–473.
161. R GJ, X Z, Z K, P KP. Altered K+ B. Mechanisms of altered excitation-contractioncoupling in canine tachycardia-induced heart fail-current of ventricular myocytes in rats with chronic
myocardial infarction. Am J Physiol 1998; 274: ure, II: model studies. Circ Res 1999; 84: 571–586.177. R RJ, N S, C M. Math-H259–H265.
162. B P, L AJ, J JV. Investigation of the ematical analysis of canine atrial action potentials:rate, regional factors, and electrical remodeling. Ammechanisms underlying the increased contraction
of hypertrophied ventricular myocytes isolated from J Physiol Heart Circ Physiol 2000; 279: H1767–H1785.the spontaneously hypertensive rat. Cardiovasc Res
1993; 27: 1268–1277. 178. T K, L D, H KM, L P, JEK, L ES. Decreased expression of Kv4.2 and163. Y JA, J M, F JS, Z YY, T GN.
Heterogeneous changes in K currents in rat vent- novel Kv4.3 K+ channel subunit mRNAs in ventriclesof renovascular hypertensive rats. Circ Res 1997;ricles three days after myocardial infarction. Car-
diovasc Res 1999; 44: 132–145. 81: 533–539.179. B P, B JP, M M, V164. W AD, K R, Y XM, B PH.
The thyroid hormone analog DITPA restores Ito in G, L P. Regional alteration of the transientoutward current in human left ventricular septumrats after myocardial infarction. Am J Physiol Heart
Circ Physiol 2000; 278: H1105–H1116. during compensated hypertrophy. Circulation 1997;96: 1266–1274.165. H UC, M E, J DC. Molecular dis-
section of cardiac repolarization by in vivo Kv4.3 180. N M, B DJ, E E. Char-acteristics of transient outward current in humangene transfer. J Clin Invest 2000; 105: 1077–1084.
166. K BC, K-R BE, W- ventricular myocytes from patients with terminalheart failure. Circ Res 1993; 73: 386–394. NJ, J LR, M M. Remodelling of ionic
currents in hypertrophied and failing hearts of 181. A F, A JL, R JM, L P,V G. Ionic basis of ventricular arrhythmiastransgenic mice overexpressing calsequestrin. J
Physiol 2000; 525 Pt 2: 483–498. in remodeled rat heart during long-term myocardialinfarction. Cardiovasc Res 1999; 42: 402–415.167. F C, C RB, L TS, G WR. A rapidly

G. Y. Oudit et al.870
182. C E, C A, S L, D P P, P from individual patients. ACE-Inhibitor MyocardialR, R ML, G G, M A. Long- Infarction Collaborative Group. Lancet 2000; 355:term treatment of spontaneously hypertensive rats 1575–1581.with losartan and electrophysiological remodeling 196. P B, P-W PA, S R, M FA,of cardiac myocytes. Cardiovasc Res 2000; 45: 388– D K, C AJ, K MA, R G,396. K GH, N J, S D, T
183. Y H, K T, T F, T N, N B. Effect of losartan compared with captopril onH, K M, K A. Restoration of action mortality in patients with symptomatic heart fail-potential duration and transient outward current ure: randomised trial—the Losartan Heart Failureby regression of left ventricular hypertrophy. J Mol Survival Study ELITE II. Lancet 2000; 355: 1582–Cell Cardiol 1997; 29: 1331–1339. 1587.
184. B SM, S SJ, H G. Normal regional 197. L MJ. Sudden death: ACE inhibitors anddistribution of membrane current density in rat beta-blockers. Basic Res Cardiol 2000; 95 (Suppl.left ventricle is altered in catecholamine-induced 1): I37–I40.hypertrophy. Cardiovasc Res 1999; 42: 391–401. 198. K WB, P JF, C LA. Cardiac failure
185. L JK, K I, H H, A T, K K, and sudden death in the Framingham Study. AmT J. Stage-dependent changes in membrane Heart J 1988; 115: 869–875.currents in rats with monocrotaline-induced right 199. B L A, C P, L JF. Am-ventricular hypertrophy. Am J Physiol 1997; 272: bulatory sudden cardiac death: mechanisms of pro-H2833–H2842. duction of fatal arrhythmia on the basis of data
186. L WM, B PA. Abnormal electrical properties from 157 cases. Am Heart J 1989; 117: 151–159.of myocytes from chronically infarcted canine 200. S WG, S LW, M HR,heart. Alterations in Vmax and the transient out- S LA. Sudden death prevention in patientsward current. Circulation 1992; 85: 1175–1188. with advanced ventricular dysfunction. Circulation
187. P JM, B PA. Electrical remodeling in 1993; 88: 2953–2961.ischemia and infarction. Cardiovasc Res 1999; 42: 201. T GF, M E. Electrophysiological re-284–297. modeling in hypertrophy and heart failure. Car-
188. W SS, B AL, C JS, E K, diovasc Res 1999; 42: 270–283.K P, M RJ. Dissimilarities in the 202. K YA, B GM, F H, Kelectrophysiological abnormalities of lateral border P, S CC, C SA, B AM. Decreasedand central infarct zone cells after healing of myo- sodium and increased transient outward potassiumcardial infarction in cats. Circ Res 1982; 51: 486– currents in iron-loaded cardiac myocytes. Im-493. plications for the arrhythmogenesis of human sider-
189. Y L, F J, G R, L GR, W Z, N otic heart disease. Circulation 1999; 100: 675–S. Ionic remodeling underlying action potential 683.changes in a canine model of atrial fibrillation. Circ203. N S, L D, Y L. Basic mechanisms of atrialRes 1997; 81: 512–525.
fibrillation—very new insights into very old ideas.190. R GJ, X Z, W RT, M H,Annu Rev Physiol 2000; 62: 51–77.Z IH. Electrophysiology of rabbit ventricular
204. W H, S H, Z L, P M, Y B,myocytes following sustained rapid ventricular pa-N S, W Z. Nicotine is a potent blocker ofcing. J Mol Cell Cardiol 1997; 29: 721–732.the cardiac A-type K+ channels. Effects on cloned191. B L, L A, D DE, F F,Kv4.3 channels and native transient outward cur-T H, H TH, K J, N-G B,rent. Circulation 2000; 102: 1165–1171.A P. Canine ventricular myocytes possess a
205. H W, B G, C AJ, J MJ,renin-angiotensin system that is upregulated withR MR, A C, E D, F M,heart failure. Circ Res 2001; 88: 298–304.M M, M A, S R. The potential for QT192. D DE, H RA, K CE, B GJ, Kprolongation and pro-arrhythmia by non-anti-V, MW CD, B KM. Molecular mech-arrhythmic drugs: clinical and regulatory im-anisms of angiotensin II in modulating cardiacplications. Report on a Policy Conference of thefunction: intracardiac effects and signal trans-European Society of Cardiology. Cardiovasc Resduction pathways. J Mol Cell Cardiol 1997; 29:2000; 47: 219–233.2893–2902.
206. H B, Q D, E-S N. Early down-re-193. S Y, M T, Y K, D R, Mgulation of K+ channel genes and currents in theT, K T, M T, T H, H M. Reninpostinfarction heart. J Cardiovasc Electrophysiolangiotensin system-dependent hypertrophy as a2000; 11: 1252–1261.contributor to heart failure in hypertensive rats:
207. N HB, K S, K DA, T GF, Mdifferent characteristics from renin angiotensin sys-E. Cellular basis of ventricular arrhythmias andtem-independent hypertrophy. J Am Coll Cardiolabnormal automaticity in heart failure. Am J Physiol2001; 37: 293–299.1999; 277: H80–H91.194. C JN. Prognosis in congestive heart failure. J
208. N M, K S. Potassium channel down-Card Fail 1996; 2: S225–S229.regulation in heart failure. Cardiovasc Res 1998;195. F MD, Y S, K L, P M, H37: 324–334.A, M G, T-P C, B S, P J,
209. T GF, B DJ, C HG, B-M L, B E. Long-term ACE-inhibitor RD, K PD, L JH, K D, Ftherapy in patients with heart failure or left-vent-
ricular dysfunction: a systematic overview of data AM, M E. Sudden cardiac death in heart

Structure and Function of Cardiac Ito 871
failure. The role of abnormal repolarization. Cir- 224. I H, H Y, H M, T M, A S,culation 1994; 90: 2534–2539. T T, N M, T K, M F.
210. P PH, N HB, T RS, K S, T GF, Endothelin-1 induces hypertrophy with enhancedM E, K DA. Repolarization abnormalities, expression of muscle-specific genes in cultured neo-arrhythmia and sudden death in canine tachy- natal rat cardiomyocytes. Circ Res 1991; 69: 209–cardia-induced cardiomyopathy. J Am Coll Cardiol 215.1997; 30: 576–584. 225. F S, G ME. Ca(2+)-dependent
211. W EH, H RL, W S. Inotropic routes to Ras: mechanisms for neuronal survival,effects of electric currents. I. Positive and negative differentiation, and plasticity? Neuron 1996; 16:effects of constant electric currents or current pulses 233–236.applied during cardiac action potentials. II. Hy- 226. M A, Z MJ, S WS, H-potheses: calcium movements, excitation-con- C. Modulation of SERCA2 expression bytraction coupling and inotropic effects. Circ Res thyroid hormone and norepinephrine in car-1969; 24: 409–445. diocytes: role of contractility. Am J Physiol 1997;
212. A DG. On the relationship between action 272: H1876–H1885.potential duration and tension in cat papillary 227. G C, N J, V H, N L. Effects ofmuscle. Cardiovasc Res 1977; 11: 210–218. nisoldipine on endothelin-1- and angiotensin II-
213. W B. Relationships between peak force, induced immediate/early gene expression and pro-action potential duration and stimulus interval in tein synthesis in adult rat ventricular cardio-rabbit myocardium. Acta Physiol Scand 1979; 106: myocytes. J Cardiovasc Pharmacol 1994; 24: 13–16.395–409. 228. B MA, A MB, G-B
214. S VJ. The negative correlation between J, C A, G PE, F SJ, S PH.action potential duration and force of contraction Adrenergic receptor stimulation of the mitogen-during restitution in rat myocardium. J Mol Cell activated protein kinase cascade and cardiac hyper-Cardiol 1986; 18: 1033–1045. trophy. Biochem J 1996; 314: 115–121.
215. B RA, C RB, G WR. Role of so- 229. R LB, G ME. Stimulation of growthdium-calcium exchange in activation of contraction factor receptor signal transduction by activation ofin rat ventricle. J Physiol 1993; 472: 391–413. voltage-sensitive calcium channels. Proc Natl Acad
216. M JN, Y H, M G, G IL, L Sci USA 1996; 93: 1113–1118.W, C H, S LS, L EG, V G, 230. H JJ, C KR. Signaling pathways for car-S A. Cardiac-specific overexpression of the diac hypertrophy and failure. N Engl J Med 1999;alpha1 subunit of the L-type voltage-dependent 341: 1276–1283.Ca2+ channel in transgenic mice. Loss of iso- 231. C PF, A RS. Cardiac Arrhythmias:proterenol-induced contraction. J Biol Chem 1999; The role of triggered activity and other mechanisms.274: 21503–21506. Mount Kisco, NY: Futura 1988.217. M JN, B I, L W, V G, S
232. A C, S S. Clinical relevance ofA. A Ca2+-dependent transgenic model of cardiaccardiac arrhythmias generated by after-hypertrophy: A role for protein kinase calpha. Cir-depolarizations. Role of M cells in the generationculation 2001; 103: 140–147.of U waves, triggered activity and torsade de pointes.218. S J. Depolarization-transcription coupling inJ Am Coll Cardiol 1994; 23: 259–277.excitable cells. Rev Physiol Biochem Pharmacol 1996;
233. A C. Basic mechanisms of reentrant127: 251–279.arrhythmias. Curr Opin Cardiol 2001; 16: 1–7.219. O J, C WA. Electrical activity, cAMP,
234. B DM. Calcium fluxes involved in control ofand cytosolic calcium regulate mRNA encodingcardiac myocyte contraction. Circ Res 2000; 87:sodium channel alpha subunits in rat muscle cells.275–281.Neuron 1989; 2: 1447–1452.
235. V AO, V MW, B LN, 220. A-S SR, C AJ, A RN, DG AC. Calcium-activated Cl− current con-DB. Cholinergic stimulation of skeletal muscle cellstributes to delayed afterdepolarizations in singleinduces rapid immediate early gene expression: rolePurkinje and ventricular myocytes. Circulationof intracellular calcium. Brain Res Mol Brain Res2000; 101: 2639–2644.1994; 26: 55–60.
236. F H. The heart and cardiovascular system:221. S J, I S. Signal transduction path-Scientific foundations. New York: Raven Press,ways of angiotensin II-induced c-fos gene ex-1992.pression in cardiac myocytes in vitro. Roles of
237. S P, V-S M E, R B. Cloningphospholipid-derived second messengers. Circ Resof a novel component of A-type K+ channels op-1993; 73: 424–438.erating at subthreshold potentials with unique ex-222. Y T, T K, H E, M K, K T,pression in heart and brain. J Neurophysiol 1996;K I, T H, K T, N R,75: 2174–2179.Y Y. Mechanical loading activates mitogen-
238. B MV, M MJ, L S, Ractivated protein kinase and S6 peptide kinase inRL, C DL, S HC. In situ hybridizationcultured rat cardiac myocytes. J Biol Chem 1993;reveals extensive diversity of K+ channel mRNA in268: 12069–12076.isolated ferret cardiac myocytes. Circ Res 1996; 78:223. B MA, G PE, S PH. En-1083–1089.dothelin-1, phorbol esters and phenylephrine
239. L M, H H, K I, B MR. Char-stimulate MAP kinase activities in ventricular car-diomyocytes. FEBS Lett 1993; 317: 271–275. acterisation of the transient outward K+ current

G. Y. Oudit et al.872
in rabbit sinoatrial node cells. Cardiovasc Res 2000; kinje cells and ventricular myocytes. Am J PhysiolHeart Circ Physiol 2000; 279: H466–H474.46: 433–441.
240. U K, H N, M T, K H. 244. G JB, B RF, K V, S L.Molecular remodeling of Kv4.3 potassium channels inProperties of the transient outward current in rabbit
sino-atrial node cells. J Mol Cell Cardiol 1999; 31: human atrial fibrillation. J Cardiovasc Electrophysiol2000; 11: 626–633.1975–1984.
241. M JS, H JC. Characteristics of a tran- 245. L B, J A, Z J, B P, H X,M GF, K G. Long QT and ventricularsient outward current (sensitive to 4-amino-pyr-
idine) in Ca2+-tolerant myocytes isolated from the arrhythmias in transgenic mice expressing the Nterminus and first transmembrane segment of arabbit atrioventricular node. Pflugers Arch 1999;
438: 68–78. voltage-gated potassium channel. Proc Natl AcadSci USA 1998; 95: 2926–2931.242. D JE, S W, W HS, MD C, Y H,
W RS, C IS, MK D. Role of the 246. J A, M GF, Z J, M M, LB, B P, W SD, K G. InducibleKv4.3 K+ channel in ventricular muscle. A molecular
correlate for the transient outward current. Circ polymorphic ventricular tachyarrhythmias in atransgenic mouse model with a long Q-T phenotype.Res 1996; 79: 659–668.
243. H W, W Z, N S. A comparison of Am J Physiol Heart Circ Physiol 2000; 278: H1891–H1898.transient outward currents in canine cardiac Pur-




















