
The local, global, and neural aspects of visuospatial processing in autism spectrum disorders
9
The local, global, and neural aspects of visuospatial processing in autism spectrum disorders Rajesh K. Kana a,n , Yanni Liu b , Diane L. Williams c , Timothy A. Keller b , Sarah E. Schipul b , Nancy J. Minshew d , Marcel Adam Just b a Department of Psychology, University of Alabama at Birmingham, Civitan International Research Center CIRC 235G, 1719 6th Ave South, Birmingham, AL, United States b Department of Psychology, Carnegie Mellon University, Pittsburgh, PA, United States c Department of Speech Language Pathology, Duquesne University, Pittsburgh, PA, United States d Department of Psychiatry, University of Pittsburgh, Pittsburgh, PA, United States article info Article history: Received 10 May 2013 Received in revised form 21 October 2013 Accepted 23 October 2013 Available online 30 October 2013 Keywords: Autism fMRI Global–local processing Block design task abstract Behavioral studies have documented a relative advantage in some aspects of visuospatial cognition in autism although it is not consistently found in higher functioning individuals with autism. The purpose of this functional neuroimaging study was to examine the neural activity in high functioning individuals with autism while they performed a block design task that systematically varied with regard to whether a global pattern was present. Participants were 14 adults with high-functioning autism and 14 age and IQ matched typical controls. The task was to identify a missing block in target figures which had either an obvious global shape or was an arbitrary array of blocks. Behavioral results showed intact, but not superior, performance in our participants with autism. A key group difference was that the participants with autism showed reliably greater activation in occipital and parietal regions in both tasks suggesting an increased reliance of the autism group on posterior brain areas to mediate visuospatial tasks. Thus, increased reliance on relatively posterior brain regions in itself may not guarantee superior performance as seen in the present study. & 2013 Elsevier Ltd. All rights reserved. 1. Introduction Individuals with autism spectrum disorders (ASD) have been reported to have intact, sometimes superior (faster and more accu- rate), performance, relative to typically developing (TD) individuals, in tasks involving visuospatial processing. For instance, children with ASD tend to perform better than TD participants on visual search (O'Riordan, Plaisted, Driver, & Baron-Cohen, 2001; Plaisted, O'Riordan, & Baron-Cohen, 1998a; Plaisted, Saksida, Alcantara, & Weisblatt, 2003), feature discrimination (O'Riordan & Plaisted, 2001; Plaisted, O'Riordan, & Baron-Cohen, 1998b), and the embedded figures test (EFT; Jolliffe & Baron-Cohen, 1997; Ropar & Mitchell, 2001; Shah & Frith, 1983). In tasks that require the detection of visual elements embedded in larger fields, people with autism have been found to show increased reliance on local details (Frith, 1989; Shah & Frith, 1993; Jolliffe & Baron-Cohen, 1997; Mottron, Burack, Iarocci, Belleville, & Enns, 2003; Lahaie, Mottron, Arguin, Berthiaume, & Jemel, 2006) and hence show intact or superior performance relative to controls. Better performance has also been reported in individuals with autism on the block design task (BDT), first by Shah and Frith (1993) and later by others (Caron, Mottron, Berthiaume, & Dawson, 2006; Gilchrist et al., 2001; Goldstein, Beers, Siegel, & Minshew, 2001; Happé, 1994; Pellicano, Gibson, Mayberry, Durkin, & Badcock, 2005; Ropar & Mitchell, 2001). Such advantage in visuospatial processing in autism has been best explained by two theoretical accounts, the Weak Central Coherence (WCC) theory (Frith, 1989; Happé, 2013; Happé & Booth, 2008) and the Enhanced Perceptual Functioning (EPF) account (Mottron et al., 2003; Mottron, Dawson, Soulières, Hubert, & Burack, 2006). While WCC addresses visual advantage in autism as a byproduct of a deficit in seeing global form, the EPF attributes it to a default local-oriented processing with increased reliance on details. BDT, developed by Kohs (1923), requires an individual to replicate a target two-dimensional geometric design by assem- bling a set of component structures. The components are squares (blocks) that are entirely white, entirely black, or are half-white and half-black (The half and half squares are formed by a division by one of the two diagonals of the square or a bisecting horizontal or vertical boundary, and the colors and organization of the surface may vary in specific experiments). The target figure can always be constructed by some arrangement of the nine possible component blocks. Shah and Frith (1993) found that individuals with autism had no advantage in a control condition in which the Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/neuropsychologia Neuropsychologia 0028-3932/$ - see front matter & 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.neuropsychologia.2013.10.013 n Corresponding author. Tel.: þ1 205 934 3171; fax: þ1 205 975 6330. E-mail addresses: [email protected], [email protected] (R.K. Kana). Neuropsychologia 51 (2013) 2995–3003
Transcript of The local, global, and neural aspects of visuospatial processing in autism spectrum disorders

The local, global, and neural aspects of visuospatial processingin autism spectrum disorders
Rajesh K. Kana a,n, Yanni Liu b, Diane L. Williams c, Timothy A. Keller b, Sarah E. Schipul b,Nancy J. Minshewd, Marcel Adam Just b
a Department of Psychology, University of Alabama at Birmingham, Civitan International Research Center CIRC 235G,1719 6th Ave South, Birmingham, AL, United Statesb Department of Psychology, Carnegie Mellon University, Pittsburgh, PA, United Statesc Department of Speech Language Pathology, Duquesne University, Pittsburgh, PA, United Statesd Department of Psychiatry, University of Pittsburgh, Pittsburgh, PA, United States
a r t i c l e i n f o
Article history:Received 10 May 2013Received in revised form21 October 2013Accepted 23 October 2013Available online 30 October 2013
Keywords:AutismfMRIGlobal–local processingBlock design task
a b s t r a c t
Behavioral studies have documented a relative advantage in some aspects of visuospatial cognition inautism although it is not consistently found in higher functioning individuals with autism. The purposeof this functional neuroimaging study was to examine the neural activity in high functioning individualswith autism while they performed a block design task that systematically varied with regard to whethera global pattern was present. Participants were 14 adults with high-functioning autism and 14 age and IQmatched typical controls. The task was to identify a missing block in target figures which had either anobvious global shape or was an arbitrary array of blocks. Behavioral results showed intact, but notsuperior, performance in our participants with autism. A key group difference was that the participantswith autism showed reliably greater activation in occipital and parietal regions in both tasks suggestingan increased reliance of the autism group on posterior brain areas to mediate visuospatial tasks. Thus,increased reliance on relatively posterior brain regions in itself may not guarantee superior performanceas seen in the present study.
& 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Individuals with autism spectrum disorders (ASD) have beenreported to have intact, sometimes superior (faster and more accu-rate), performance, relative to typically developing (TD) individuals, intasks involving visuospatial processing. For instance, children withASD tend to perform better than TD participants on visual search(O'Riordan, Plaisted, Driver, & Baron-Cohen, 2001; Plaisted, O'Riordan,& Baron-Cohen,1998a; Plaisted, Saksida, Alcantara, & Weisblatt, 2003),feature discrimination (O'Riordan & Plaisted, 2001; Plaisted, O'Riordan,& Baron-Cohen, 1998b), and the embedded figures test (EFT; Jolliffe &Baron-Cohen, 1997; Ropar & Mitchell, 2001; Shah & Frith, 1983). Intasks that require the detection of visual elements embedded in largerfields, people with autism have been found to show increased relianceon local details (Frith, 1989; Shah & Frith, 1993; Jolliffe & Baron-Cohen,1997; Mottron, Burack, Iarocci, Belleville, & Enns, 2003; Lahaie,Mottron, Arguin, Berthiaume, & Jemel, 2006) and hence show intactor superior performance relative to controls. Better performance hasalso been reported in individuals with autism on the block design task
(BDT), first by Shah and Frith (1993) and later by others (Caron,Mottron, Berthiaume, & Dawson, 2006; Gilchrist et al., 2001;Goldstein, Beers, Siegel, & Minshew, 2001; Happé, 1994; Pellicano,Gibson, Mayberry, Durkin, & Badcock, 2005; Ropar & Mitchell, 2001).Such advantage in visuospatial processing in autism has been bestexplained by two theoretical accounts, the Weak Central Coherence(WCC) theory (Frith, 1989; Happé, 2013; Happé & Booth, 2008) andthe Enhanced Perceptual Functioning (EPF) account (Mottron et al.,2003; Mottron, Dawson, Soulières, Hubert, & Burack, 2006). WhileWCC addresses visual advantage in autism as a byproduct of a deficitin seeing global form, the EPF attributes it to a default local-orientedprocessing with increased reliance on details.
BDT, developed by Kohs (1923), requires an individual toreplicate a target two-dimensional geometric design by assem-bling a set of component structures. The components are squares(blocks) that are entirely white, entirely black, or are half-whiteand half-black (The half and half squares are formed by a divisionby one of the two diagonals of the square or a bisecting horizontalor vertical boundary, and the colors and organization of thesurface may vary in specific experiments). The target figure canalways be constructed by some arrangement of the nine possiblecomponent blocks. Shah and Frith (1993) found that individualswith autism had no advantage in a control condition in which the
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/neuropsychologia
Neuropsychologia
0028-3932/$ - see front matter & 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.neuropsychologia.2013.10.013
n Corresponding author. Tel.: þ1 205 934 3171; fax: þ1 205 975 6330.E-mail addresses: [email protected], [email protected] (R.K. Kana).
Neuropsychologia 51 (2013) 2995–3003

geometric design to be constructed was segmented such that eachcomponent square of the design was individually identifiable. Allthe participants had to do to construct the design in this conditionwas to iteratively select the component blocks and put them in thecorrect location. However, in the conventional presentation con-dition, the component blocks of the target design perceptuallymerge into each other forming global patterns that are larger thanindividual blocks. In this conventional presentation, the partici-pants with autism completed the block design task faster and withfewer errors than the typically developing individuals.
Of late, several research reports have questioned the idea ofAue, Lavelle, & Cacioppo, 2009; Edgin & Pennington, 2005). Forinstance, better performance on the BDT, relative to other tasks, isnot found in individuals with high-functioning autism and Asper-ger syndrome (Kaland, Mortensen, & Smith, 2007; Mayes &Calhoun, 2003; Ropar & Mitchell, 2001; Ehlers et al., 1997). In acomprehensive study on BDT, Caron et al. (2006) examinedindividuals with autism and controls with and without a visuos-patial peak. This study showed that while diminished detrimentalinfluence of perceptual coherence on BDT performance is bothsensitive and specific to autism, outstanding BDT performance wasseen only in a subgroup of individuals with autism. Thus, it ispossible that the autism spectrum has some individuals withsuperior visuospatial ability and some without. In studies thatfocused on broader phenotypes, superior performance on BDT wasnot found in parents of individuals with autism (de Jonge, Kemner,Naber, & van Engeland, 2009; Fombonne, Bolton, Prior, Jordan, &Rutter, 1997; Piven & Palmer, 1997; Szatmari et al., 1993). There arealso reports that visuospatial peaks within the autism spectrumare limited to individuals with a language delay (Mottron,Soulieres, Simard-Meileur, & Dawson, 2008). Furthermore, visuos-patial peak may be a developmental process as such peaks areseen more in children with autism aged 8 years 11 months than inchildren aged 5 years 5 months (Joseph, Tager-Flusberg, & Lord,2002). Overall, findings from these studies point to the fact thatBDT superiority may be characteristic of some but not all indivi-duals on the autism spectrum (Soulières et al., 2011; Stewart,Watson, Allcock, & Yaqoob, 2009; Caron et al., 2006).
Although visuospatial processing in autism has been exten-sively studied using behavioral measures, only a few studies havebeen conducted using neuroimaging measures. Some previousneuroimaging studies used different versions of the embeddedfigures task (Damarla et al., 2010; Manjaly et al., 2007; Ring et al.,1999), and two other studies (McGrath et al., 2012; Silk et al.,2006) used a mental rotation task. All of these studies foundincreased activation in participants with autism in the occipitaland superior parietal regions and reduced activation mainly in thefrontal regions, suggesting more low level, perceptually orientedprocessing in autism. A recent fMRI study (Spencer et al., 2012)also used EFT and examined the brain responses in autism andcontrol participants along with unaffected siblings of individualswith autism. This study found atypical activation in temporal andfrontal regions in autism and in unaffected siblings. Yet anotherrecent study (Liu, Cherkassky, Minshew, & Just, 2011) used aperceptual line-counting task and found decreased medial pre-frontal activation and decreased connectivity of this region withposterior regions in participants with autism. Overall, most ofthese studies point to an altered neural response in autism despiteintact or superior behavioral performance.
The primary goal of the present study was to use functionalmagnetic resonance imaging (fMRI) to examine the neural activityin high functioning individuals with autism while they performeda BDT that systematically varied with regard to whether a globalpattern was present. There was only one previous fMRI study ofBDT in autism (Bolte, Hubl, Dierks, Holtmann, & Poistka, 2008).This study found reduced activation response in autism in neurons
in the visual cortex that respond to shape representation, figure-ground, and gestalt organization. Our study is different from theBolte et al. study in terms of the variations in the stimuli and ourfocus on the whole brain. The present study also differs fromprevious neuroimaging studies of visuospatial processing in aut-ism in the following ways. The Silk et al. (2006) and Ring et al.(1999) studies should be considered as exploratory due to thelimited number of participants with autism, 6 and 7 respectively.In addition, our study used BDT unlike the EFT in Damarla et al.(2010), Spencer et al. (2012), and Ring et al. (1999), and unlike themental rotation task in Silk et al. (2006). If individuals with autismhave a performance advantage in BDTs, an fMRI study has thepotential of revealing the neural basis of that advantage.
According to previous reports of locally oriented processing inautism (Shah & Frith, 1993; Caron et al., 2006), the detrimentaleffect of a global pattern (which can obscure the attributes of aparticular location in the design) should have less impact on theindividuals with autism as compared to TD controls, due to thehypothesized decreased influence of global information for thosewith autism. Frith (2003) suggested that weak central coherence(WCC) in those with autism is the basis for the superior perfor-mance in BDTs, whereas the EPF model (Mottron et al., 2003,2006) attributes it to enhanced perceptual ability. However, theremay be other factors involved, such as an atypical bias toward localprocessing with a local to global interference (Booth and Happé,2010; Rinehart, Bradshaw, Moss, Brereton, & Tonge, 2000) and apreference toward detecting local targets in divided attentionconditions (Plaisted, Swettenham, & Rees, 1999). At the neurallevel, the EPF model (Mottron et al., 2003, 2006) would predict alocal overconnectivity, especially in the occipital areas in ASD. Just,Keller, Malave, Kana, and Varma (2012) also point to autonomy inthe posterior brain areas in ASD which mediates the recruitmentof the visuospatial route to accomplish tasks. Such a pattern ofincreased use of posterior brain areas has been found in manyneuroimaging studies of a variety of tasks in autism (see Samson,Mottron, Soulieres, & Zeffiro, 2012). The present study examinesthe neural bases of visuospatial processing in individuals withautism in the context of a block design task.
2. Method
2.1. Participants
The participants were 14 adults with high-functioning autism and 14 typicalcontrols. All participants were male and were matched for age (t(28)¼0.2, ns), FullScale IQ (t(28)¼0.23, ns), Verbal IQ (t(28)¼1.14, ns), and Performance IQ (t(28)¼0.89, ns) as determined by the Wechsler Adult Intelligence Scale-III (WAIS-III;Wechsler, 1997) (see Table 1 for demographic information).
The diagnosis of autism was established using two structured researchdiagnostic instruments, the Autism Diagnostic Interview-Revised (ADI-R; Lord,Rutter, & Le Couteur, 1994) and the Autism Diagnostic Observation Schedule(ADOS-WPS; Lord, Rutter, DiLavore, & Risi, 2001), supplemented with expertclinical opinion according to the accepted criteria for high-functioning autism(Minshew, 1996). Potential participants with autism were excluded if they had anidentifiable cause for their autism (such as fragile-X syndrome, tuberous sclerosis,or fetal cytomegalovirus infection) or if there was evidence of birth asphyxia, head
Table 1Demographic information of the participants.
Autism Control
Age (years) Mean7SD 21.575.7 21.874.0VIQ Mean7SD 107.679.0 110.976.3PIQ Mean7SD 110.878.6 108.177.1FSIQ Mean7SD 110.378.0 110.976.5Handedness Right: left 13: 1 12: 2Gender Male: Female 14: 0 14: 0
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–30032996

injury, or a seizure disorder. Exclusionary criteria were based on neurologic historyand examination and chromosomal analysis or metabolic testing if indicated.
The control participants were community volunteers recruited to match theautism participants on age, IQ, gender, race, and socioeconomic status of family oforigin, as measured by the Hollingshead method (Hollingshead, 1957). Potentialcontrol participants were screened by questionnaire, telephone, face-to-face inter-view, and observation during screening psychometric tests. Exclusionary criteriawere evaluated through these procedures and included the following: current orpast psychiatric and neurologic disorders, birth injury, developmental delay, schoolproblems, acquired brain injury, learning disabilities, substance abuse, and medicaldisorders with implications for the central nervous system or those requiringregular medication. Potential control participants were also screened to excludethose with a family history (in parents, siblings, and offspring) of autism,developmental cognitive disorders, affective disorders, anxiety disorders, schizo-phrenia, obsessive compulsive disorder, substance abuse, or other neurologic orpsychiatric disorders thought to have a genetic component. Handedness wasdetermined with the Lateral Dominance Examination from the Halstead-ReitanNeuropsychological Test Battery (Reitan & Wolfson, 1985). One participant withautism and two control participants were left-handed. However, the brain activa-tion data from these left-handers were clearly similar to their respective groups;therefore, the data were not separated by handedness.
2.2. Materials and procedure
During fMRI scanning, the participants were presented with a target design onthe left side of the screen (See Fig. 1 for an example). The pattern in each figure wasmade of nine blocks that were all black, all white, or half black and half white(divided into two triangles or two rectangles of equal size). On the right side of thescreen, a blank matrix of the same size, with a question mark in the location of oneof the nine blocks, specifying the block to be identified within the pattern, wasshown. At the bottom of the screen were four answer choices. The participantexamined the pattern and the blank matrix and then chose which of the fouralternatives matched the cued location. In the global condition, the patterninvolved a simple and recognizable object such as a house or a sailboat, as shownin Fig. 1A. We chose to call this condition “global” as opposed to “gestalt” sincethere are clear differences between these two terms (see Brosnan, Scott, Fox, & Pye,2004), and the term “global” may fit better for our stimuli. In the random arraycondition (henceforth random), the design was a random configuration of blocks,with the restriction that the target square was clearly defined on all four sides sothat no obvious global pattern was present. In order for the target square to beclearly defined, the borders of the squares around it had contrasting color. Anexample of the random condition is shown in Fig. 1B.
At the start of each experimental trial, the pattern on the left side of the screenwas presented for 4000 ms (the onset of the first item in each block was timelocked to the acquisition of the superior most slice in the prescription). The blankmatrix on the right side of the screen appeared 750 ms after the start of the patternand remained on screen for 3250 ms, disappearing at the same time as the pattern.The answer choices at the bottom of the screen appeared at the same time as thepattern and remained on screen for 7000 ms, thus staying on the screen for3000 ms after the pattern and blank matrix disappeared. The participant was ableto answer any time during the 7000 ms when the answer choices were on thescreen. The reaction time data was recorded from the onset of the appearance ofthe figure on the left side of the screen. The next trial started immediately. Trials ofeach condition (global or random) were presented in blocks of six. A 12,000-msrest period followed each block during which an asterisk appeared on screen. Theblocks alternated between conditions, and counterbalanced across participants.There were three blocks of each condition, for a total of 18 trials in each condition.An asterisk was presented for four 24,000-ms fixation periods, evenly distributed
among the blocks, with one at the beginning and one after each random conditionblock. The participants were instructed that the asterisk was “a rest period for youto relax.” On the day of the scan, the participants completed a short practice set ofone block of four trials in each condition outside of the scanner to familiarize themwith the task.
The stimuli used in the present study is different from the BDT used in Shahand Frith (1993) in a few ways: (1) While their task had segmented andunsegmented designs, our task had global and random patterns without segmen-tation; (2) The Shah and Frith stimuli had 4 squares/blocks that made up a largershape, whereas our stimuli consisted of 9 blocks/squares perhaps making our taskmore complex; and (3) In their task, participants were shown a two-dimensionalpattern on a card and were asked to construct the pattern using 4 three-dimensional blocks; whereas our task was computerized and the participants' taskwas to identify a missing block in a larger array. We tried to preserve a criticalaspect of BDT, which is the ability to break down the visual gestalt into singleelements. Block design tasks serve as a measure of general intelligence (Royer et al.,1984), and as indicators of visualization ability (Snow, Kyllonen, & Marshalek,1984). In Koh's Block Design as well as in our version, the participant may rely onanalytic (the displayed design is mentally segmented into units) or holistic (thedesign is viewed as a whole and is not differentiated into units) strategies to solvethe problem at hand. Thus, the BDT used in our study, in addition to testing globaland analytical skills, tests problem-solving in general.
2.3. fMRI procedures
The data were collected using a Siemens Allegra 3.0T scanner (Siemens Inc.,Erlangen, Germany) at the Brain Imaging Research Center (BIRC) of Carnegie MellonUniversity and the University of Pittsburgh. The study was performed with agradient echo, EPI sequence with TR¼1000 ms, TE¼30 ms and a 601 flip angle.Seventeen oblique-axial slices were acquired; each slice was 5-mm thick with a gapof 1-mm between slices. The acquisition matrix was 64�64 with3.125 mm�3.125 mm�5 mm voxels.
2.4. fMRI analyses—Distribution of activation
To compare the participating groups in terms of the distribution of activation,the data were analyzed using SPM2 (Wellcome Department of Cognitive Neurology,London). Images were corrected for slice acquisition timing, motion-corrected,normalized to the Montreal Neurological Institute (MNI) template, resampled to2�2�2 mm voxels, and smoothed with an 8-mm Gaussian kernel to decreasespatial noise. Statistical analyses were performed on individual and group data byusing the general linear model and Gaussian random field theory as implementedin SPM2 (Friston et al., 1995). Group analyses were performed using a random-effects model. Statistical maps were superimposed on normalized T1-weightedimages. An uncorrected height threshold of t¼3.36 (P¼0.001) and an extentthreshold of twenty-four 8 mm3 voxels was used.
2.5. fMRI analyses—Functional connectivity
The functional connectivity was computed (separately for each participant) as acorrelation between the average time course of signal intensity of all the activatedvoxels in each member of a pair of ROIs. Fifteen functional ROIs were defined toencompass the main clusters of activation in the group activation map for eachgroup in both experimental conditions versus fixation (global and random). Labelsfor these 13 ROIs [bilateral middle frontal gyrus (MFG), bilateral superior frontalgyrus (SFG), bilateral insula (INS), the supplementary motor area (SMA), bilateralinferior parietal lobule (IPL), bilateral superior parietal lobule (SPL), and bilateral
Fig. 1. (A) and (B) Example stimuli from the Global (A) and the Random (B) condition. Figures with a target pattern that has an obvious global shape (A) and a random arraytarget pattern (B).
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–3003 2997
middle occipital gyrus (MOG)] were assigned with reference to the parcellation ofthe Montreal Neurological Institute (MNI) single subject T1 weighted datasetcarried out by Tzourio-Mazoyer and colleagues (Tzourio-Mazoyer et al., 2002).A sphere was defined for each cluster (with a radius ranging from 5 to 12 mm) thatbest captured the cluster of activation in the map for each group. The ROIs used inthe analysis were the union of the two spheres defined for the two groups. Theactivation time course extracted for each participant over the activated voxelswithin the ROI originated from the normalized and smoothed images, which werehigh-pass filtered and had the linear trend removed. Participants who did not showactivation in a given functional ROI were excluded from further analyses involvingthat ROI. The functional connectivity correlation was computed on the imagesbelonging only to the experimental conditions, so it reflects the synchronizationbetween the activation in two areas while the participant is performing the taskand not during the baseline condition. Fisher's r to z transformation was applied tothe correlation coefficients for each participant prior to averaging and statisticalcomparison of the two groups.
2.6. Statistical analyses
For the behavioral data, reaction time and error rate were recorded for eachtrial of the experiment. We conducted a 2 (Group: autism vs. control)�2(Condition: global vs. random) mixed ANOVA on the reaction time data as wellas on the accuracy data separately. This analysis provided the main effect of group,main effect of condition, and group by condition interactions. For the reaction timedata, we used data from correct trials. For within-group measure of brainactivation, we conducted a one-sample t-test and for between-group activation, atwo-sample t-test. Moreover, multiple regression analyses were performed usingPIQ and VIQ as covariates to predict activation and functional connectivity, whichdid not reveal any statistically significant relationships.
3. Results
3.1. Group differences in brain activation
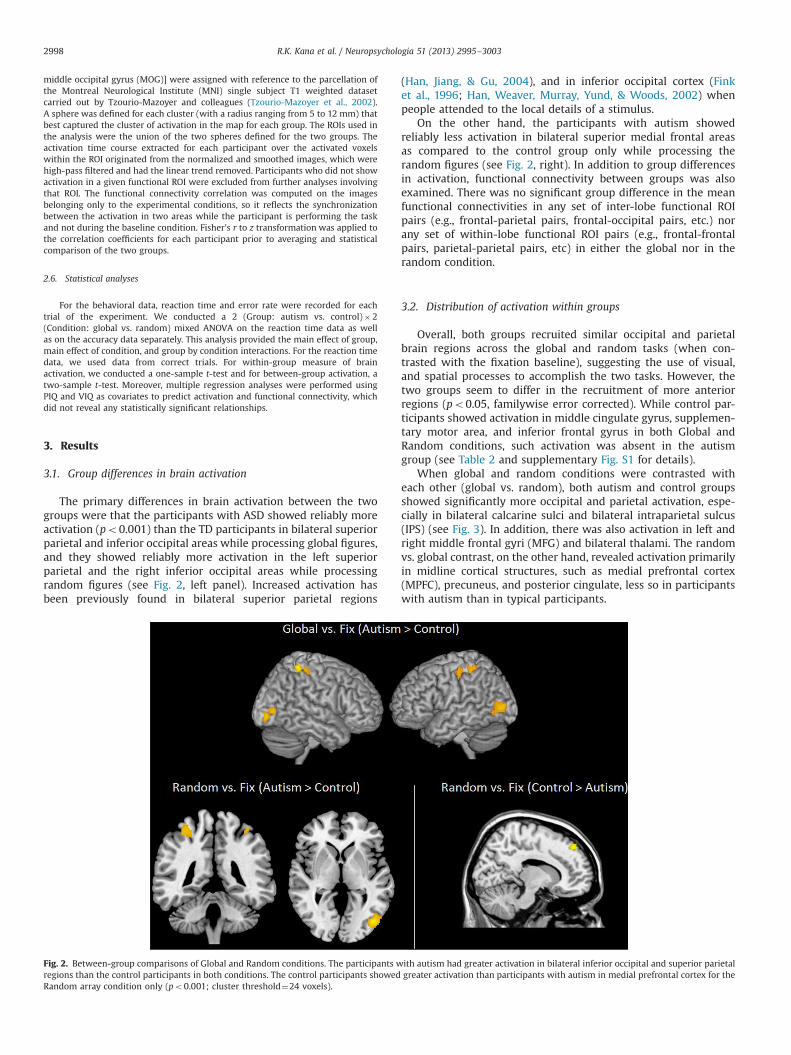
The primary differences in brain activation between the twogroups were that the participants with ASD showed reliably moreactivation (po0.001) than the TD participants in bilateral superiorparietal and inferior occipital areas while processing global figures,and they showed reliably more activation in the left superiorparietal and the right inferior occipital areas while processingrandom figures (see Fig. 2, left panel). Increased activation hasbeen previously found in bilateral superior parietal regions
(Han, Jiang, & Gu, 2004), and in inferior occipital cortex (Finket al., 1996; Han, Weaver, Murray, Yund, & Woods, 2002) whenpeople attended to the local details of a stimulus.
On the other hand, the participants with autism showedreliably less activation in bilateral superior medial frontal areasas compared to the control group only while processing therandom figures (see Fig. 2, right). In addition to group differencesin activation, functional connectivity between groups was alsoexamined. There was no significant group difference in the meanfunctional connectivities in any set of inter-lobe functional ROIpairs (e.g., frontal-parietal pairs, frontal-occipital pairs, etc.) norany set of within-lobe functional ROI pairs (e.g., frontal-frontalpairs, parietal-parietal pairs, etc) in either the global nor in therandom condition.
3.2. Distribution of activation within groups
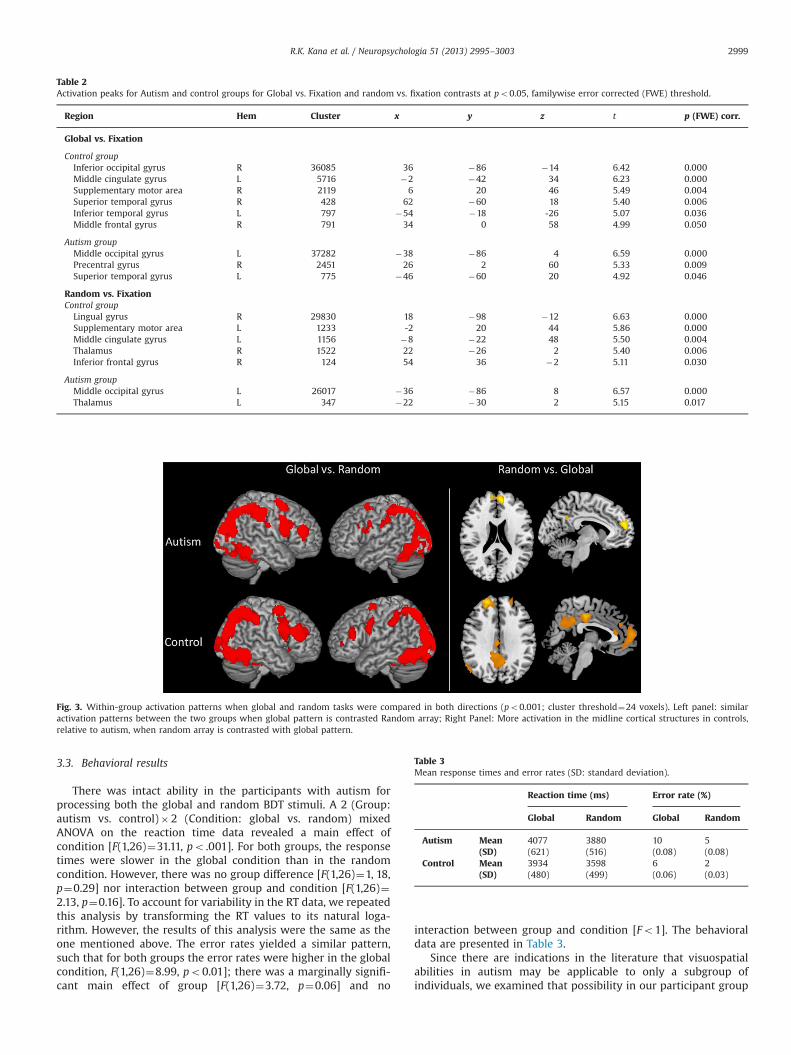
Overall, both groups recruited similar occipital and parietalbrain regions across the global and random tasks (when con-trasted with the fixation baseline), suggesting the use of visual,and spatial processes to accomplish the two tasks. However, thetwo groups seem to differ in the recruitment of more anteriorregions (po0.05, familywise error corrected). While control par-ticipants showed activation in middle cingulate gyrus, supplemen-tary motor area, and inferior frontal gyrus in both Global andRandom conditions, such activation was absent in the autismgroup (see Table 2 and supplementary Fig. S1 for details).
When global and random conditions were contrasted witheach other (global vs. random), both autism and control groupsshowed significantly more occipital and parietal activation, espe-cially in bilateral calcarine sulci and bilateral intraparietal sulcus(IPS) (see Fig. 3). In addition, there was also activation in left andright middle frontal gyri (MFG) and bilateral thalami. The randomvs. global contrast, on the other hand, revealed activation primarilyin midline cortical structures, such as medial prefrontal cortex(MPFC), precuneus, and posterior cingulate, less so in participantswith autism than in typical participants.
Fig. 2. Between-group comparisons of Global and Random conditions. The participants with autism had greater activation in bilateral inferior occipital and superior parietalregions than the control participants in both conditions. The control participants showed greater activation than participants with autism in medial prefrontal cortex for theRandom array condition only (po0.001; cluster threshold¼24 voxels).
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–30032998
3.3. Behavioral results
There was intact ability in the participants with autism forprocessing both the global and random BDT stimuli. A 2 (Group:autism vs. control)�2 (Condition: global vs. random) mixedANOVA on the reaction time data revealed a main effect ofcondition [F(1,26)¼31.11, po .001]. For both groups, the responsetimes were slower in the global condition than in the randomcondition. However, there was no group difference [F(1,26)¼1, 18,p¼0.29] nor interaction between group and condition [F(1,26)¼2.13, p¼0.16]. To account for variability in the RT data, we repeatedthis analysis by transforming the RT values to its natural loga-rithm. However, the results of this analysis were the same as theone mentioned above. The error rates yielded a similar pattern,such that for both groups the error rates were higher in the globalcondition, F(1,26)¼8.99, po0.01]; there was a marginally signifi-cant main effect of group [F(1,26)¼3.72, p¼0.06] and no
interaction between group and condition [Fo1]. The behavioraldata are presented in Table 3.
Since there are indications in the literature that visuospatialabilities in autism may be applicable to only a subgroup ofindividuals, we examined that possibility in our participant group
Table 2Activation peaks for Autism and control groups for Global vs. Fixation and random vs. fixation contrasts at po0.05, familywise error corrected (FWE) threshold.
Region Hem Cluster x y z t p (FWE) corr.
Global vs. Fixation
Control groupInferior occipital gyrus R 36085 36 �86 �14 6.42 0.000Middle cingulate gyrus L 5716 �2 �42 34 6.23 0.000Supplementary motor area R 2119 6 20 46 5.49 0.004Superior temporal gyrus R 428 62 �60 18 5.40 0.006Inferior temporal gyrus L 797 �54 �18 -26 5.07 0.036Middle frontal gyrus R 791 34 0 58 4.99 0.050
Autism groupMiddle occipital gyrus L 37282 �38 �86 4 6.59 0.000Precentral gyrus R 2451 26 2 60 5.33 0.009Superior temporal gyrus L 775 �46 �60 20 4.92 0.046
Random vs. FixationControl group
Lingual gyrus R 29830 18 �98 �12 6.63 0.000Supplementary motor area L 1233 -2 20 44 5.86 0.000Middle cingulate gyrus L 1156 �8 �22 48 5.50 0.004Thalamus R 1522 22 �26 2 5.40 0.006Inferior frontal gyrus R 124 54 36 �2 5.11 0.030
Autism groupMiddle occipital gyrus L 26017 �36 �86 8 6.57 0.000Thalamus L 347 �22 �30 2 5.15 0.017
Fig. 3. Within-group activation patterns when global and random tasks were compared in both directions (po0.001; cluster threshold¼24 voxels). Left panel: similaractivation patterns between the two groups when global pattern is contrasted Random array; Right Panel: More activation in the midline cortical structures in controls,relative to autism, when random array is contrasted with global pattern.
Table 3Mean response times and error rates (SD: standard deviation).
Reaction time (ms) Error rate (%)
Global Random Global Random
Autism Mean 4077 3880 10 5(SD) (621) (516) (0.08) (0.08)
Control Mean 3934 3598 6 2(SD) (480) (499) (0.06) (0.03)
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–3003 2999

by looking at the participants' scores on subtests of the WASI, thetest used to measure the intelligence level of the participants. Inparticular, we examined the raw scores as well as t-scores for theBlock Design subtest. The test scores did not reveal the existenceof any subgroup in ASD. Nor did it reveal any statistical differencebetween the ASD and TD groups (see Table 4).
4. Discussion
The main findings of this study are these: first, the participantswith autism had intact, but not enhanced, behavioral performanceon a variation of the BDT. This performance on the task created forthe fMRI study was consistent with the lack of a peak on the BDTsubtest of the WASI for the participants with autism. Second,similar behavioral performance across the participant groups wasaccompanied by differences in levels of brain activation. Specifi-cally, the autism group had relatively increased activation inposterior occipital and parietal regions as compared to the TDgroup, whereas the TD group was also using frontal resources.Third, there were no statistically significant group differences infunctional connectivity. Therefore, the difference for the autismgroup was in the relative use of neural resources in particularregions (less frontal/more posterior) not in the coordination ofthese regions.
Although it is difficult to delineate the specific relationshipbetween behavioral performance and levels of brain activation, anincrease in activation has frequently been interpreted as indicatingincreased use of the processing resources in the relevant brainregions (Aue et al., 2009). Therefore, the increase in activation inthe posterior occipital and parietal regions by the autism groupcould suggest that they were using the resources of these areas toa greater extent than the TD group, but for a similar behavioralresult. The increased reliance on posterior brain areas did nottranslate into superior performance in our participants withautism. This result is consistent with the results of a number ofother neuroimaging studies of visuospatial processing in indivi-duals with autism. For example, in a meta-analysis of 26 neuroi-maging studies of autism that utilized visual stimuli, Samson et al.(2012) found no significant group difference in performance in69% of the studies. They also found a generally higher task-related
activity in posterior brain areas and lower activity in frontal cortexjust as in the current study. Previous studies have found posteriorparietal cortex (e.g., Donner, Kettermann, Diesch, Villringer, &Brandt, 2003; Coull, Walsh, Frith, & Nobre, 2003) as well as anetwork of parietal and occipital areas (Nobre, Coull, Walsh, &Frith, 2003) to be extensively involved in visual search, a task thatcan be accomplished with low-level perceptual processing. Thegreater activation of posterior regions by the autism group may bereflective of an increased reliance by this group on low-levelperceptual processing centers to accomplish this task.
The additional recruitment of posterior processing resources bythe autism group occurs in the context of a lack of recruitment offrontal regions. The TD group relied on a frontal-parietal-occipitalnetwork to accomplish the task; the autism group had a similarlevel of coordination with the frontal regions (as indicated by thelack of a difference in functional connectivity) but did not have theexpected increased processing in frontal regions. The autism groupwas using more posterior and less frontal resources but wascoordinating the timing of the activation between these areas toa similar extent as indicated by the lack of a difference infunctional connectivity between the two groups. In the BDT task,the resources that were recruited were adequate and no differenceoccurred in behavioral performance. However, the marginallydifferent error rate suggests that the autism group may have beenworking less not more efficiently than the age and IQ-matched TDcontrols (thus the increase in activation in the posterior regions)for a similar behavioral result.
Direct comparison of processing global and random figuresgenerated relatively more activation for global figures in bothparticipant groups. This may be because of (a) the associatedmeanings and thought processes generated by the global figure ineach picture, e.g., boat, or bird, and/or (b) the difficulty in mentallysegmenting the global pattern in order to determine the locationof the interrogated block in the intact figure. On the other hand,when random stimuli were compared to global ones, the controlparticipants exhibited more activation in the bilateral superiormedial frontal areas (MPFC) than the participants with autism. TheMPFC has been found to be involved in stimulus-oriented and instimulus-independent attention, especially in tasks involvingnavigating around a visually presented shape versus imaginingthe same shape and navigating around it (Gilbert, Frith, & Burgess,2005; Gilbert et al., 2007; Burgess, Dumontheil, & Gilbert, 2007).Therefore the MPFC in control participants may be mediating thisconstant search for meaning and coherence. Activation in MPFChas also been found when participants switch from one way ofperforming a task to another (Rushworth et al., 2002). In thepresent study, the control participants may be shifting back andforth between global and local oriented perception of the visualstimuli to identify the missing block. The lack of activation inMPFC in participants with autism is consistent with the findings ofa recent fMRI study of a lower-level perceptual line-counting task(Liu et al., 2011) and also with a generally limited level ofactivation in frontal areas in autism (Samson et al., 2012).
In sum, an atypical pattern of resource allocation (more poster-ior less frontal) during visuospatial processing may be reflective ofthe nature of cognitive and neural mechanisms in individuals withautism. The increased activity in posterior areas and intact con-nectivity we found may be a pattern that is indicative of thestronger role of perceptual processes in autism. Alternately, it maybe indicative of a processing network that is reflective of aberrantfrontal processing with a resultant reliance on posterior processingresources. Either explanation is consistent with the results of thecurrent study.
Altered brain response with intact behavioral performance, inautism in the present study is consistent with previous studies ofvisual processing in autism (McGrath et al., 2012; Damarla et al.,
Table 4Raw and T scores of the block design subtest of WASI for both participant groups.
Block design subtest
Raw score T-score
ASD TD ASD TD
52 56 57 5754 59 64 6162 52 62 5456 43 58 4957 40 58 4866 45 65 5025 53 39 5645 50 51 5366 56 64 5737 60 45 6148 46 52 5069 49 70 5646 44 52 5257 47 58 51
Mean 52.5 50.2 56.7 54.2S.D. 12.5 6.4 8.7 4.3S.E. 3.5 1.8 2.4 1.2
Raw score: t(26)¼0.78; p40.05; T score: t(26)¼1.14; p40.05.
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–30033000

2010; Lee et al., 2007; Manjaly et al., 2007; Silk et al., 2006) andgenerates interesting questions. The lack of superior performancein autism may point to the heterogeneity in autism and thepossible existence of individuals within the autism group withdiffering visuospatial abilities (Soulières, Zeffiro, Girard, &Mottron, 2011; Stewart et al., 2009; Caron et al., 2006). We testedthis in our sample with different indices but did not find anyvisuospatial peaks in autism. For instance, participants in bothgroups were matched on verbal, performance, and full scale IQs. Inaddition, our regression analysis using PIQ as a covariate onbehavioral and fMRI data did not result in statistically significantrelationship. We also examined the block design subtest of WASIand it did not show any significant variation either in ASD or in TDparticipant groups. It should be noted that such peaks were notseen in some previous studies conducted by our group (Williams,Goldstein, Kojkowski, & Minshew, 2008; Minshew, Turner, &Goldstein, 2005).
There have been several neuroimaging studies that haveprovided evidence for decreased frontal-posterior functional con-nectivity in more complex cognitive tasks in people with autism(e.g. Just, Cherkassky, Keller, & Minshew, 2004; Just, Cherkassky,Keller, Kana, & Minshew, 2007; Kana, Keller, Cherkassky, Minshew,& Just, 2006; Kana, Keller, Minshew, & Just, 2007; Kana, Keller,Cherkassky, Minshew, & Just, 2009; Koshino et al., 2008; Mason,Williams, Kana, Minshew, & Just, 2008). The findings pertaining tofunctional connectivity in autism from the current study aredifferent from previous studies in that there are no overall groupdifferences. It is possible that the visual search task in the presentstudy may be one where the proposed enhanced perceptualfunctioning in autism may manifest (Mottron et al., 2006) result-ing in intact connectivity. The BDT used in the present study maybe relatively less affected by other higher cognitive componentslike executive planning and motor control, at least for theparticipants with autism. It is possible that the default strategythe individuals with autism may use (one that is primarily basedon the recruitment of occipital and parietal areas) may work inthis task more so than a complex sentence comprehension task ora problem-solving task. Thus, the overall stronger engagement ofthe visual cortex and parietal areas by participants with autismentailed greater activation and intact coordination. It should benoted that a recent magnetoencephalography study (Khan et al.,2013) of face viewing showed reduced connectivity in local as wellas long-range connections in autism.
The EPF account (Mottron et al., 2006) and an updated WCCaccount (Happé, 2013; Happé & Booth, 2008; Happé & Frith, 2006)attribute visuospatial advantage in autism as due to superiority inlocal processing and to a bias towards local processing in peoplewith autism. At the brain level, such processing may underliegreater recruitment of relatively posterior areas. Mottron et al.(2006) suggest that perception in autism may be different fromthat in typical individuals in overall superior functioning, involve-ment, and autonomy of posterior regions like parietal and occipitalcortices. The results of the present study, increased parietal andoccipital area activation and intact functional connectivity, sup-ports that and can be the result of a more autonomous posteriorbrain functioning in our participants with autism (Just et al., 2012).The increased recruitment of posterior regions we found in autismcan also be the result of reduced top-down feedback as predictedby WCC and the cortical underconnectivity theory (Just et al.,2004; Kana et al., 2006). It has been found that brain activationand connectivity in lower brain regions can be altered by top-down feedback (Friston & Buechel, 2000) and a lack of which mayresult in abnormal connectivity (Frith, 2004). Hence EPF, WCC, andunderconnectivity theory suggest that a posterior-oriented brainfunctioning may underlie intact or superior visuospatial proces-sing in autism.
Although this study revealed important new information aboutvisuospatial processing in autism, there are a few limitations. First,the target figures in this study were presented at the left visualfield. While we do not think this would have had a significantimpact on the results, future studies may account for this; andsecond, although we matched the black and white blocks in globaland random stimuli, in some stimuli we could not achieve 100%matching due to the constraints of a given shape.
Future studies should examine what would happen if the taskdemand could be metrically increased. Would there be a point atwhich the increased activation or use of local processing resourceswould not be sufficient and would the autism group be able toincrease the coordination with frontal areas under this constraintor would we begin to see the decrease in functional connectivitythat has been found in more demanding tasks? It might also beinformative to seek out individuals with autism who display thereported BDT peak to compare their pattern of processing relativeto individuals with autism, such as those in the current study, whodid not display this behavioral pattern. In summary, this study hasnot only looked at processing during a visuospatial task but hasprovided information about what happens with functional con-nectivity during a task that the processing resources the groupwith autism have are adequate for the task and the task can beperformed without extensive recruitment of frontal resources.These are two elements that should be systematically varied infuture studies.
Acknowledgments
This research was supported by the Collaborative Program ofExcellence in Autism Grant HD35469 from the National Institute ofChild Health and Human Development, and a postdoctoral traininggrant from the Cure Autism Now Foundation.
Appendix A. Supporting information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.neuropsychologia.2013.10.013.
References
Aue, T., Lavelle, L. A., & Cacioppo, J. T. (2009). Great expectations: What can fMRIresearch tell us about psychological phenomena? International Journal ofPsychophysiology, 73(1), 10–16.
Bolte, S., Hubl, D., Dierks, T., Holtmann, M., & Poistka, F. (2008). An fMRI-study oflocally oriented perception in autism: Altered early visual processing of theblock design test. Journal of Neural Transmission, 115, 545–552.
Booth, R., & Happe, H. (2010). “Hunting with a knife and … fork”: Examining centralcoherence in autism, attention deficit/hyperactivity disorder, and typicaldevelopment with a linguistic task. Journal of Experimental Child Psychology,107(4-5), 377–393.
Brosnan, M. J., Scott, F. J., Fox, S., & Pye, J. (2004). Gestalt processing in autism:Failure to process perceptual relationships and the implications for contextualunderstanding. Journal of Child Psychology and Psychiatry, 45, 459–469.
Burgess, P. W., Dumontheil, I., & Gilbert, S. J. (2007). The gateway hypothesis ofrostral prefrontal cortex (area 10) function. Trends in Cognitive Science, 11,290–298.
Caron, M. J., Mottron, L., Berthiaume, C., & Dawson, M. (2006). Cognitive mechan-isms, specificity and neural underpinnings of visuospatial peaks in autism.Brain, 129, 1789–1802.
Coull, J. T., Walsh, V., Frith, C. D., & Nobre, A. C. (2003). Distinct neural substrates forvisual search amongst spatial versus temporal distractors. Cognitive BrainResearch, 17, 368–379.
Damarla, S. R., Keller, T., Kana, R. K., Cherkassky, V., Williams, D., Minshew, N., et al.(2010). Cortical underconnectivity coupled with preserved visuospatial cogni-tion in autism: Evidence from an fMRI study of an embedded figures task.Autism Research, 3, 1–7.
de Jonge, M., Kemner, C., Naber, F., & van Engeland, H. (2009). Block designreconstruction skills: Not a good candidate for an endophenotypic marker inautism research. European Child & Adolescent Psychiatry, 18(4), 197–205.
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–3003 3001

Donner, T. H., Kettermann, A., Diesch, E., Villringer, A., & Brandt, S. A. (2003).Parietal activation during visual search in the absence of multiple distractors.NeuroReport, 14, 2257–2261.
Edgin, J. O., & Pennington, B. F. (2005). Spatial cognition in autism spectrumdisorders: Superior, impaired, or just intact? Journal of Autism and Develop-mental Disorders, 35, 729–745.
Ehlers, S., Nyden, A., Gillberg, C., Sandberg, A. D., Dahlgren, S. O., Hjelmquist, E.,et al. (1997). Asperger syndrome, autism and attention disorders: A compara-tive study of the cognitive profiles of 120 children. Journal of Child Psychologyand Psychiatry, 38, 207–217.
Fink, G. R., Halligan, P. W., Marshall, J. C., Frith, C. D., Frackowiak, R. S. J., & Dolan, R. J.(1996). Where in the brain does visual attention select the forest and the trees?Nature, 382, 626–628.
Fombonne, E., Bolton, P., Prior, J., Jordan, H., & Rutter, M. (1997). A family study ofautism: Cognitive patterns and levels in parents and siblings. Journal of ChildPsychology and Psychiatry, 38, 667–683.
Friston, K. J., & Buechel, C. (2000). Attentional modulation of effective connectivityfrom V2 to V5/MT in humans. Proceedings of the National Academy of Sciences,97, 75912–75916.
Friston, K. A., Holmes, A. P., Worsley, K. J., Poline, J. B., Frith, C., & Frackowiak, R. S. J.(1995). Statistical parametric maps in functional imaging: A general linearapproach. Human Brain Mapping, 2, 189–210.
Frith, C. (2003). What do imaging studies tell us about the neural basis of autism.In: Bock, & Goode (Eds.), Autism: Neural basis and treatment possibilities (pp.149–176). Chichester, U.K.: John Wiley & Sons.
Frith, C. (2004). Is autism a disconnection disorder? Lancet Neurology, 3, 577.Frith, U. (1989). Autism: Explaining the enigma. Oxford: Blackwell.Gilbert, S. J., Frith, C. D., & Burgess, P. W. (2005). Involvement of rostral prefrontal
cortex in selection between stimulus-oriented and stimulus-independentthought. European Journal of Neuroscience, 21, 1423–1431.
Gilbert, S. J., Williamson, I. D. M., Dumontheil, I., Simons, J. S., Frith, C. D., & Burgess, P.W. (2007). Distinct regions of medial rostral prefrontal cortex supporting socialand non-social functions. Social Cognitive and Affective Neuroscience, 2, 217–226.
Gilchrist, A., Green, J., Cox, A., Burton, D., Rutter, M., & Le Couteur, A. (2001).Development and current functioning in adolescents with Asperger syndrome:A comparative study. Journal of Child Psychology and Psychiatry, 42, 227–240.
Goldstein, G., Beers, S. R., Siegel, D. J., & Minshew, N. J. (2001). A comparison ofWAIS-R profiles in adults with high-functioning autism or differing subtypes oflearning disability. Applied Neuropsychology, 8, 148–154.
Han, S., Jiang, Y., & Gu, H. (2004). Neural substrates differentiating global/localprocessing of bilateral visual inputs. Human Brain Mapping, 22, 321–328.
Han, S., Weaver, J., Murray., S., Yund, E. W., & Woods, D. L. (2002). Hemisphericasymmetry in global/local processing: Effects of stimulus position and spatialfrequency. NeuroImage, 17, 1290–1299.
Happé, F. G. E. (2013). Weak central coherence. In: F. E. Volkmar (Ed.), Theencyclopedia of autism spectrum disorders (pp. 3344–3346). NY: Springer.
Happé, F. G. E. (1994). An advanced test of theory of mind: Understanding of storycharacters' thoughts and feelings by able autistic, mentally handicapped andnormal children and adults. Journal of Autism and Developmental Disorders, 24,129–154.
Happé, F. G. E., & Booth, R. D. L. (2008). The power of the positive: Revisiting weakcoherence in autism spectrum disorders. The Quarterly Journal of ExperimentalPsychology, 61(1), 50–63.
Happé, F. G. E., & Frith, U. (2006). The weak coherence account: Detail-focusedcognitive style in autism spectrum disorders. Journal of Autism and Develop-mental Disorders, 36, 5–25.
Hollingshead, A. (1957). Four-factor index of social status. Unpublished manuscript.New Haven, CT: Yale University.
Jolliffe, T., & Baron-Cohen, S. (1997). Are people with autism and Aspergersyndrome faster than normal on the embedded figures test? Journal of ChildPsychology & Psychiatry, 38, 527–534.
Joseph, R. M., Tager-Flusberg, H., Lord, C. (2002). Cognitive profiles and socialcommunicative functioning in children with autism spectrum disorder. Journalof Child Psychology and Psychiatry, 43: 807–821.
Just, M. A., Cherkassky, V. L., Keller, T. A., & Minshew, N. J. (2004). Cortical activationand synchronization during sentence comprehension in high-functioningautism: Evidence of underconnectivity. Brain, 127, 1811–1821.
Just, M. A., Cherkassky, V. L., Keller, T. A., Kana, R. K., & Minshew, N. J. (2007).Functional and anatomical cortical underconnectivity in autism: Evidence froman fMRI study of an executive function task and corpus callosum morphometry.Cerebral Cortex, 17, 951–961.
Just, M. A., Keller, T. A., Malave, V. L., Kana, R. K., & Varma, S. (2012). Autism as aneural systems disorder: A theory of frontal-posterior underconnectivity.Neuroscience and Biobehavioral Reviews, 36(4), 1292–1313.
Kaland, N., Mortensen, E. L., & Smith, L. (2007). Disembedding performance inchildren and adolescents with Asperger syndrome or high-functioning autism.Autism, 11, 81–92.
Kana, R. K., Keller, T. A., Cherkassky, V. L., Minshew, N. J., & Just, M. A. (2006).Sentence comprehension in autism: Thinking in pictures with decreasedfunctional connectivity. Brain, 129, 2484–2493.
Kana, R. K., Keller, T. A., Minshew, N. J., & Just, M. A. (2007). Inhibitory control inhigh functioning autism: Decreased activation and underconnectivity in inhibi-tion networks. Biological Psychiatry, 62, 198–206.
Kana, R. K., Keller, T. A., Cherkassky, V. L., Minshew, N. J., & Just, M. A. (2009).Atypical frontal-posterior synchronization of Theory of Mind regions in autismduring mental state attribution. Social Neuroscience, 4, 135–152.
Khan, S., Gramfort, A., Shetty, N. R., Kitzbilchler, M. G., Ganesan, S., Kenet, T, et al.(2013). Local and long-range functional connectivity is reduced in concert inautism spectrum disorders. Proceedings of the National Academy of Sciences ofthe United States of America, 110(8), 3107–3112.
Koshino, H., Kana, R. K., Keller, T. A., Cherkassky, V. L., Minshew, N. J., & Just, M. A.(2008). fMRI investigation of working memory for faces in autism: Visualcoding and underconnectivity with frontal areas. Cerebral Cortex, 18(389-300).
Kohs, S. C. (1923). Intelligence measurement: A psychological and statistical studybased upon the block-design tests (pp. 39–63) New York: MacMillan.
Lahaie, A., Mottron, L., Arguin, M., Berthiaume, C., Jemel, B., et al. (2006). Faceperception in high-functioning autistic adults: Evidence for superior processingof face parts, not for a configural face-processing deficit. Neuropsychology, 20,30–41.
Lee, P. S., Foss-Feig, J., Henderson, J. G., Kenworthy, L. E., Gilotty, L., et al. (2007).Atypical neural substrates of embedded figures. Task performance in childrenwith autism spectrum disorder. NeuroImage, 38, 184–193.
Liu, Y., Cherkassky, V. L., Minshew, N. J., & Just, M. A. (2011). Autonomy of lower-level perception from global processing in autism: Evidence from brainactivation and functional connectivity. Neuropsychologia, 49, 2105–2111.
Lord, C., Rutter, M., DiLavore, P. C., & Risi, S. (2001). Autism diagnostic observationschedule. Los Angeles: Western Psychological Services.
Lord, C., Rutter, M., & Le Couteur, A. (1994). Autism diagnostic interview-revised: Arevised version of a diagnostic interview for caregivers of individuals withpossible pervasive developmental disorders. Journal of Autism and Develop-mental Disorders, 24, 659–685.
Manjaly, Z. M., Bruning, N., Neufang, S., Stephan, K. E., Brieber, S., Marshall, J. C.,et al. (2007). Neurophysiological correlates of relatively enhanced local visualsearch in autistic adolescents. NeuroImage, 35, 283–291.
Mason, R. A., Williams, D. L., Kana, R. K., Minshew, N., & Just, M. A. (2008). Theory ofMind disruption and recruitment of the right hemisphere during narrativecomprehension in autism. Neuropsychologia, 46, 269–280.
Mayes, S. D., & Calhoun, S. L. (2003). Analysis of WISC-II, Stanford-Binet: IV, andacademic achievement test scores in children with autism. Journal of Autismand Developmental Disorders, 33, 329–341.
McGrath, J., Johnson, K., Ecker, C., O'Hanlon, E., Gill, M., Gallagher, L., et al. (2012).Atypical visuospatial processing in autism: Insights from functional connectiv-ity analysis. Autism Research
Minshew, N. J. (1996). Autism. In: B. Berg (Ed.), Principles of child neurology (pp.1713–1729). New York: McGraw-Hill.
Minshew, N. J., Turner, C. A., & Goldstein, G. (2005). The application of short formsof the Wechsler Intelligence Scales in adults and children with high functioningautism. Journal of Autism and Developmental Disorders, 35, 45–52.
Mottron, L., Burack, J. A., Iarocci, G., Belleville, S., & Enns, J. T. (2003). Locallyoriented perception with intact global processing among adolescents withhigh-functioning autism: Evidence from multiple paradigms. Journal of ChildPsychology and Psychiatry, 44, 904–913.
Mottron, L., Dawson, M., Soulières, I., Hubert, B., & Burack, J. A. (2006). Enhancedperceptual functioning in autism: An update, and eight principle of autisticperception. Journal of Autism and Developmental Disorders, 36, 27–43.
Mottron, L., Soulieres, I., Simard-Meileur, A. A., & Dawson, M. (2008). Peaks of abilityas a subtyping tool for autism. In: International meeting for autism research,London, England.
Nobre, A. T., Coull, J. T., Walsh, V., & Frith, C. D. (2003). Brain activations duringvisual search: Contributions of search efficiency versus feature binding. Neuro-Image, 18, 91–103.
O'Riordan, M. A., Plaisted, K. C., Driver, J., & Baron-Cohen, S. (2001). Superior visualsearch in autism. Journal of Experimental Psychology: Human Perception andPerformance, 27, 719–730.
O'Riordan, M., & Plaisted, K. (2001). Enhanced discrimination in autism. TheQuarterly Journal of Experimental Psychology, 54, 961–979.
Pellicano, E., Gibson, L., Mayberry, M., Durkin, K., & Badcock, D. R. (2005). Abnormalglobal processing along the dorsal visual pathway in autism: A possiblemechanism for weak central coherence. Neuropsychologia, 43, 1044–1053.
Piven, J., & Palmer, P. (1997). Cognitive deficits in parents from multiple-incidenceautism families. Journal of Child Psychology and Psychiatry, 41, 491–502.
Plaisted, K., O'Riordan, M., & Baron-Cohen, S. (1998a). Enhanced discrimination ofnovel, highly similar stimuli by adults with autism during a perceptual learningtask. Journal of Child Psychology and Psychiatry, 39, 765–775.
Plaisted, K., O'Riordan, M., & Baron-Cohen, S. (1998b). Enhanced discrimination ofnovel, highly similar stimuli by adults with autism during a perceptual learningtask. Journal of Child Psychology and Psychiatry, 39, 765–775.
Plaisted, K., Saksida, L., Alcantara, J., & Weisblatt, E. (2003). Towards an under-standing of the mechanisms of weak central coherence effects: Experiments invisual configural learning and auditory perception. Philosophical Transactions ofthe Royal Society of London, Series B, 358, 375–386.
Plaisted, K., Swettenham, J., & Rees, L. (1999). Children with autism show localprecedence in a divided attention task and global precedence in a selectiveattention task. Journal of Child Psychology and Psychiatry, 40, 733–742.
Reitan, R. M., & Wolfson, D. (1985). The Halstead-Reitan neuropsychological testbattery: Theory and clinical interpretation. Tucson, AZ: Neuropsychology Press.
Rinehart, N. J., Bradshaw, J. L., Moss, S. A., Brereton, A. V., & Tonge, B. J. (2000).Atypical interference of local detail on global processing in high-functioningautism and Asperger's disorder. Journal of Child Psychology and Psychiatry, 41,769–778.
Ring, H., Baron-Cohen, S., Williams, S., Wheelwright, S., Bullmore, E., Brammer, M.,et al. (1999). Cerebral correlates of preserved cognitive skills in autism. A
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–30033002

functional MRI study of embedded figures task performance. Brain, 122,1305–1315.
Ropar, D., & Mitchell, P. (2001). Susceptibility to illusions and performance onvisuospatial tasks in individuals with autism. Journal of child Psychology andPsychiatry, 42, 539–549.
Royer, F. L., Gilmore, G. C., & Gruhn, J. J. (1984). Stimulus parameters that produceage differences in block design performance. Journal of Clinical Psychology, 40,1474–1485.
Samson, F., Mottron, L., Soulieres, I., & Zeffiro, T. A. (2012). Enhanced visualfunctioning in autism: An ALE meta-analysis. Human Brain Mapping, 33,1553–1581.
Shah, A., & Frith, U. (1983). An islet of ability in autistic children: A research note.Journal of Child Psychology and Psychiatry, 24, 613–620.
Shah, A., & Frith, U. (1993). Why do autistic individuals show superior performanceon the block design task? Journal of Child Psychology and Psychiatry, 34,1351–1364.
Snow, R. E., Killonen, P. C., & Marshalek, B. (1984). The topography of ability andlearning correlations. In R. J. Sternberg (Ed.), Advances in the psychology ofhuman intelligence (Vol. 2). Hillsdale, NJ: Erlbaum.
Silk, T. J., Rinehart, N., Bradshaw, J. L., Tonge, B., Egan, G., O'Boyle, M. W., et al.(2006). Visuospatial processing and the function of prefrontal-parietal
networks in autism spectrum disorders: A functional MRI study. AmericanJournal of Psychiatry, 163, 1440–1443.
Soulières, I., Zeffiro, T. A., Girard, M. L., & Mottron, L. (2011). Enhanced mental imagemapping in autism. Neuropsychologia, 49(5), 848–857.
Spencer, M. D., Holt, R. J., Chura, L. R., Calder, A. J., Suckling, J., Bullmore, E. T., et al.(2012). Atypical activation during the embedded figures task as a functionalmagnetic resonance imaging endophenotype of autism. Brain, 135, 3469–3480.
Stewart, M. E., Watson, J., Allcock, A., & Yaqoob, T. (2009). Autistic traits predictperformance on the block design. Autism, 13(2), 133–142.
Szatmari, P., Jones, M. B., Tuff, L., Bartolucci, G., Fisman, S., & Mahoney, W. (1993).Lack of cognitive impairment in first-degree relatives of childrenwith pervasivedevelopmental disorders. Journal of American Academy of Child and AdolescentPsychiatry, 32, 1264–1273.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix,N., et al. (2002). Automated anatomical labeling of activations in SPM using amacroscopic anatomical parcellation of the MNI MRI single-subject brain.NeuroImage, 15, 273–289.
Wechsler, D. (1997). Wechsler Adult Intelligence Scale—3rd Edition (WAIS-3®) SanAntonio, TX: Harcourt Assessment.
Williams, D. L., Goldstein, G., Kojkowski, N., & Minshew, N. J. (2008). Do Individualswith high-functioning Autism have the IQ profine associated with non-verballearning disability? Research on Autism Spectrum Disorders, 2(2), 353–361.
R.K. Kana et al. / Neuropsychologia 51 (2013) 2995–3003 3003




















