The heterogeneity of bovine IgG2—III. The ion-exchange heterogeneity of IgG2a is the result of...
10
Molecular Immunology, Vol. 24, No. 12, pp. 1317-1326, 1987 Printed in Great Britain. 0161-5890187 $3.00 + 0.00 Pergamon Journals Ltd THE HETEROGENEITY OF BOVINE IgG2-III. THE ION-EXCHANGE HETEROGENEITY OF IgG2a IS THE RESULT OF ®ION VARIATION* J. E. BUTLER,~ M. V. BORCA, H. HEYERMANN, M. DILLENDER and M. BIELECKA The University of Iowa Medical School, Department of Microbiology, Iowa City, IA 52242, U.S.A (First received 20 February 1987; accepted in revised form 2 July 1987) Abstract-Bovine IgG2a from an animal homozygous at the A-locus could be fractionated on DEAE columns into subpopulations; studies using analytical size columns showed that the number of sub- populations was dependent on the ionic strength of equilibration buffer. Subpopulations of IgG2a could be purified by this method which did not differ in their carbohydrate content but had mean Ips consistent with their elution behavior. The spectrotype of their H-chains but not their L-chains, paralleled the spectrotype of the corresponding intact IgG2a subpopulations. When papain digests of each sub- population were fractionated by chromatofocusing, their Fc fragments eluted homogeneously with a pl ca 6.0 while their Fabs eluted heterogeneously with pls ranging from 8 to 5. The distribution of carbohydrate among such fractions corresponded to the distribution of the Fcs. The behavior of the IgG2a Fabs on chromatofocusing paralleled the elution behavior of the parent IgG2a subpopulations from DEAE-Sephadex and the isoelectric behavior of their H-chains. The antibody activity of Fab fragments, separated by chromatofocusing, are consistent with the concept of idiotypic variation. The differential antibody activity of each intact IgG2a subpopulation for seven different antigens suggests that the ion-exchange behavior of IgG2a. in which the influence of allotypes and sub-isotypes has been excluded and for which charge-related L-chain heterogeneity is minimal, must reside in the V,-regions of the different IgG2a subpopulations. Investigators are cautioned against assigning ruminant subclass desig- nations on the basis of subpopulations isolated by ion-exchange chromatography. INTRODUCTION subclass designations to other subpopulation ob- Anion-exchange chromatography has been tradi- tained using anion-exchange chromatography (Kick- tionally employed for the fractionation of normal hiifen et al., 1968; Mossmann et al., 1981). Despite serum immunoglobulins (Rowe and Fahey, 1965) the work of Duncan et al. (1972), the concept that b ovine including those of cattle and other ruminants (Mur- IgG subclasses can be separated by anion- phy et al., 1965; Milstein and Feinstein, 1968; Kick- exchange chromatography has persisted in the litera- hijfen et al., 1968; Aalund, 1968; Mach and Pahud, ture and subisotype-specific biological activities of 1971; Butler and Maxwell, 1972; Duncan et al., 1972). the major subclasses continue to be assigned accord- The differential ion-exchange behavior of ruminant ing to this criterion (Hajdu et al., 1974; Lisowski et IgGs permitted the identification and character- al., 1975; Schmitz, 1979; Mossmann et al., 1981; G ization of the two major recognized subclasses IgGl orczyca et al., 1986). Serum IgGl preparations and IgG2 (Milstein and Feinstein, 1968; see Butler, invariably contain IgG2 which possesses identical 1969). Some investigators have assigned additional ion-exchange behavior although the opposite can normally be avoided (see Whitney, 1976; Butler et al., 1980; Butler, 1981; Butler et al., 1987~). *Research supported by USDA Grant CSRS-2-2172 Immunoglobulin heterogeneity has been classified awarded to J. E. Butler, a fellowship awarded by CONICET to Manuel Borca and a fellowship awarded as isotypic, allotypic and idiotypic variation. The by the Food and Agriculture Organization of the United isotypic variants IgGl and IgG2, have long been Nations to M. Bielecka. M. Dillender is supported by recognized (Milstein and Feinstein, 1968) and we the Iowa Biotechnology Training Program (IBTP) have more recently described IgG2a and IgG2b (But- funded by the USDA. tAuthor to whom correspondence should be addressed. ler et al., 1987b). Allotypic variants of IgG2 (Blakes- Abbreviations: Al, A2, allotypic markers of bovine IgG2a; lee et al., 1971a, b) which we now regard as variants a-ELI%, amplified ELISA; AES, activated enzyme of IgG2a (Butler and Heyermann, 1986; Butler et al., solution; BASA, Bruceila abortus soluble antigen; CF, 1987~) are also recognized. It is possible to employ chromatofocusing; DHA, dehydroascorbate; DNP, chromatographic conditions to fractionate IgG2a X4-dinitrophenyl; ELISA, enzyme-linked immuno- sorbent assay; FLU, fluorescein; HPGPC, high pressure from an animal homozygous for the A-allotype such gel permeation chromatography; IEF, isoelectric focus- that contamination by IgG2b and IgGl can be ing; PC, phosphorylcholine; SDS-PAGE, poly- excluded; hence constant region allotypic hetero- acrylamide gel electrophoresis in sodium dodecyl sul- geneity can be excluded. Nevertheless homogeneous fate; SRD, single radial diffusion. anion-exchange elution of IgG2a is not observed 1317
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The heterogeneity of bovine IgG2—III. The ion-exchange heterogeneity of IgG2a is the result of...

Molecular Immunology, Vol. 24, No. 12, pp. 1317-1326, 1987 Printed in Great Britain.
0161-5890187 $3.00 + 0.00 Pergamon Journals Ltd
THE HETEROGENEITY OF BOVINE IgG2-III.
THE ION-EXCHANGE HETEROGENEITY OF IgG2a IS THE RESULT OF ®ION VARIATION*
J. E. BUTLER,~ M. V. BORCA, H. HEYERMANN, M. DILLENDER and M. BIELECKA
The University of Iowa Medical School, Department of Microbiology, Iowa City, IA 52242, U.S.A
(First received 20 February 1987; accepted in revised form 2 July 1987)
Abstract-Bovine IgG2a from an animal homozygous at the A-locus could be fractionated on DEAE columns into subpopulations; studies using analytical size columns showed that the number of sub- populations was dependent on the ionic strength of equilibration buffer. Subpopulations of IgG2a could be purified by this method which did not differ in their carbohydrate content but had mean Ips consistent with their elution behavior. The spectrotype of their H-chains but not their L-chains, paralleled the spectrotype of the corresponding intact IgG2a subpopulations. When papain digests of each sub- population were fractionated by chromatofocusing, their Fc fragments eluted homogeneously with a pl ca 6.0 while their Fabs eluted heterogeneously with pls ranging from 8 to 5. The distribution of carbohydrate among such fractions corresponded to the distribution of the Fcs. The behavior of the IgG2a Fabs on chromatofocusing paralleled the elution behavior of the parent IgG2a subpopulations from DEAE-Sephadex and the isoelectric behavior of their H-chains. The antibody activity of Fab fragments, separated by chromatofocusing, are consistent with the concept of idiotypic variation. The differential antibody activity of each intact IgG2a subpopulation for seven different antigens suggests that the ion-exchange behavior of IgG2a. in which the influence of allotypes and sub-isotypes has been excluded and for which charge-related L-chain heterogeneity is minimal, must reside in the V,-regions of the different IgG2a subpopulations. Investigators are cautioned against assigning ruminant subclass desig- nations on the basis of subpopulations isolated by ion-exchange chromatography.
INTRODUCTION subclass designations to other subpopulation ob-
Anion-exchange chromatography has been tradi- tained using anion-exchange chromatography (Kick-
tionally employed for the fractionation of normal hiifen et al., 1968; Mossmann et al., 1981). Despite
serum immunoglobulins (Rowe and Fahey, 1965) the work of Duncan et al. (1972), the concept that b ovine
including those of cattle and other ruminants (Mur- IgG subclasses can be separated by anion-
phy et al., 1965; Milstein and Feinstein, 1968; Kick- exchange chromatography has persisted in the litera-
hijfen et al., 1968; Aalund, 1968; Mach and Pahud, ture and subisotype-specific biological activities of
1971; Butler and Maxwell, 1972; Duncan et al., 1972). the major subclasses continue to be assigned accord-
The differential ion-exchange behavior of ruminant ing to this criterion (Hajdu et al., 1974; Lisowski et
IgGs permitted the identification and character- al., 1975; Schmitz, 1979; Mossmann et al., 1981; G
ization of the two major recognized subclasses IgGl orczyca et al., 1986). Serum IgGl preparations
and IgG2 (Milstein and Feinstein, 1968; see Butler, invariably contain IgG2 which possesses identical
1969). Some investigators have assigned additional ion-exchange behavior although the opposite can normally be avoided (see Whitney, 1976; Butler et al.,
1980; Butler, 1981; Butler et al., 1987~).
*Research supported by USDA Grant CSRS-2-2172 Immunoglobulin heterogeneity has been classified
awarded to J. E. Butler, a fellowship awarded by CONICET to Manuel Borca and a fellowship awarded
as isotypic, allotypic and idiotypic variation. The
by the Food and Agriculture Organization of the United isotypic variants IgGl and IgG2, have long been
Nations to M. Bielecka. M. Dillender is supported by recognized (Milstein and Feinstein, 1968) and we
the Iowa Biotechnology Training Program (IBTP) have more recently described IgG2a and IgG2b (But- funded by the USDA.
tAuthor to whom correspondence should be addressed. ler et al., 1987b). Allotypic variants of IgG2 (Blakes-
Abbreviations: Al, A2, allotypic markers of bovine IgG2a; lee et al., 1971a, b) which we now regard as variants
a-ELI%, amplified ELISA; AES, activated enzyme of IgG2a (Butler and Heyermann, 1986; Butler et al.,
solution; BASA, Bruceila abortus soluble antigen; CF, 1987~) are also recognized. It is possible to employ
chromatofocusing; DHA, dehydroascorbate; DNP, chromatographic conditions to fractionate IgG2a X4-dinitrophenyl; ELISA, enzyme-linked immuno- sorbent assay; FLU, fluorescein; HPGPC, high pressure
from an animal homozygous for the A-allotype such
gel permeation chromatography; IEF, isoelectric focus- that contamination by IgG2b and IgGl can be
ing; PC, phosphorylcholine; SDS-PAGE, poly- excluded; hence constant region allotypic hetero-
acrylamide gel electrophoresis in sodium dodecyl sul- geneity can be excluded. Nevertheless homogeneous fate; SRD, single radial diffusion. anion-exchange elution of IgG2a is not observed
1317

1318 J. E. BUTLER et al.
(Butler, 1985; Butler et al., 1987a). Rather, multiple
“peaks”, i.e. ion-exchange subpopulations, can be obtained; the number of these subpopulations and the degree to which they may be separated depends on the type of anion-exchanger, ionic strength of the equilibrium buffer and column dimensions (Butler et al., 1987~).
The data presented in this report show that the charge-dependent subpopulations of IgG2a obtained from the serum of an Al/Al homozygous animal by DEAE anion-exchange chromatography, differ with respect to the pl of their respective Fabs. This charge-dependent heterogeneity is ascribable to the heavy chains of these IgG2a subpopulations. These
biochemical differences reside in their Vu-regions which could affect the antibody binding site and could explain the differential antibody activity we observed among fractions for seven different antigens.
DTE) was warmed to 27°C and 566~1 of activated enzyme solution (AES) was added. The reaction vessel was capped tightly to prevent evaporation of the thiol reagent, mixed and incubated for 2 hr at 37°C. The AES consisted of 530 pg of mercuripapain (Sigma Chemical, St Louis, MO) in 1.0 ml of di- gestion buffer. Ten mM dehydroascorbate (DHA) was prepared in a buffer containing 0.15 M NaCl, 0.05 M Tris and 2 mM EDTA. Six hundred ~1 of DHA was added. Digests were dialyzed against or diluted in the appropriate buffer for either high pressure gel permeation chromatography (HPGPC) or chromatofocusing.
MATERIALS AND METHODS
Source of bovine IgG
Bovine serum IgG was prepared from the serum of a Holstein-Friesian steer which was homozygous Al/Al (Blakeslee et al., 197la; Butler and Hey- ermann, 1986). The serum was precipitated with 40% saturated ammonium sulfate and washed once with the same concn of ammonium sulfate. Bovine IgGl was prepared from colostrum as reviewed elsewhere (Butler, 1983).
Biochemical methods
Chromatofocusing (CF). Chromatofocusing was conducted in 0.9 x 10 cm columns of DEAE-Biogel A as described by Voss (personal communications). Briefly, the Biogel A column was packed and equili- brated with 0.026 M Tris, pH 8.0. A flow rate of 0.5 ml/min was maintained and the effluent mon- itored with an ISCO UA-5 absorption meter (ISCO, Lincoln, NE) at 280 nm. One ml of papain digest (5 mg/ml) in equilibration buffer and 1 ml of final buffer was applied to the column and allowed to enter the gel. A buffer head of final buffer, sufficient to maintain a flow rate of 0.5 ml/min, was then applied and 1 ml fractions were collected. The final buffers were mixtures of Polybuffers 96 and 74 (Pharmacia Fine Chemicals, Piscataway, NJ) diluted 1: 16 and sufficient to provide either a pH of 5 or 4, depending on the papain digest being analyzed (see Fig. 6). The distribution of Fc and Fab was quantitated using an indirect competitive ELISA (see below).
Ion -exchange chromatography. Bovine serum IgG was fractionated on DEAE-Sephadex A-50. Anal- ytical columns of 0.9 x 10 cm were equilibrated with a 0.01 M Tris-HCI buffer, pH 8.6, at ionic strengths of 0.005, 0.01 and 0.05. The latter was controlled by the molarity of NaCl. Two hundred mg of IgG was applied to these analytical columns and the protein eluted isocratically. Columns of DEAE-Sephadex (2.5 x 18 cm and 2.5 x 50 cm) equilibrated with the Tris-HCI buffer containing 0.05 M NaCl were also used, the latter size to obtain preparative amounts of various IgG2a subpopulations. These columns were initially eluted isocratically and when protein was no
longer detectable in the effluent. a salt gradient employing a limit buffer containing 0.5 M NaCl was supplied to the column with an LKB ultragrad. Typical, complete elution patterns are seen in Figs 1 and 3.
Carbohydrate analysis. The concn of neutral hexose was determined using a micro version of the phenol-sulfuric acid method. One hundred to four hundred ~1 of each CF fraction were tested and the neutral hexose content obtained by reference to a standard curve prepared over a range of 2.5-25 pg of glucose.
Polyacrylamide gel electrophoresis (SDS-PAGE). SDS-PAGE was conducted in 7.5% acrylamide gels in the absence of 2-mercaptoethanol. All gels were stained with Coomassie blue. The general procedure is essentially that of Laemmli (1970).
Protein elution was monitored spectro- photometrically and the pH and ionic strength of every third fraction was determined. The concn of IgG2a and IgG 1 was determined by both single radial diffusion (SRD) and using an indirect competitive ELISA.
Analytical gel isoelectric focusing (IEF). IEF was conducted in precast Agarose gels (Resolve; Isolabs, Akron, OH) containing ampholytes spanning the range from pH 3 to 10. IEF was conducted in a thermoelectrically controlled chamber from Hoeffer Scientific (San Francisco, CA) which had been purged with ultrapure nitrogen for 2min. Electrode buffers consisted of 0.5 M NaOH containing 30 mM lysine and 0.5 M acetic acid. Samples contained 1 M glycine in H,O and were applied with 2 x 0.4cm Whatman No. 3 paper for 20min. Proteins were focused for 5 min at 5 W, 45 min at 10 W and finally for 5 min at 20 W. The IEF was conducted at 9°C.
Papain digestion. Thirty mg of each IgG2a fraction The focused gels were stained by a modification of (see Fig. 3) or IgGl, in 5 ml of digestion buffer the silver stain procedure as described for Agarose (0.15 M NaCl, 0.05 M Tris, 2mM EDTA and 1 mM gels by Willoughby and Lambert (1982). It was

Bovine IgG2 heterogeneity 1319
estimated that this method could detect as little as
20 ng of IgG.
High pressure gel permeation chromatography (HPGPC). HPGPC was performed on a 0.4 x 250mm GPC-100 column from Synchrom (Linden, IN). The mobile phase was 0.1 M Na,SO, in 0.01 M phosphate buffer at pH 6.6. This was deliv- ered with a dual piston LKB 2150 high pressure pump at 0.4 ml/min operated at 70 bar. Absorbancy was monitored using an ISCO V4 (Lincoln, NE).
Immunoassays
Source of reagents. Antisera specific for the con- stant region of bovine IgG2a, IgGI, IgGZb, Fab and the Al allotype were present in the laboratory and their preparation and specific absorption has been described elsewhere (Butler and Maxwell, 1972; But- ler, 1983; Butler and Heyermann, 1986; Butler et al., 19876; Heyermann and Butler, 1987). All other im- munochemical reagents used in the immunoassay have been described in the publications cited for the immunoassay.
Gel precipitin assays. The specificity of reagents employed in this work was evaluated by immunodiffusion using the modification described by Harrington et al. (1971). Single radial diffusion (SRD; Fahey and McKelvey, 1965) was used to measure quantitatively the IgGl and IgG2a eluted from certain columns.
Indirect competitive ELZSA. A competitive ELISA was used to determine quantitatively the Fab or Fc content of various column fractions. Briefly, micro- titer wells were coated with an excess of bovine IgG, Fab or an Fc-enriched fraction of the appropriate isotype. Then, a mixture of the test sample or a known amount of reference antigen, and a high dilution of the appropriate antibody-enzyme conju- gate was incubated for 2 hr in the microtiter wells. The wells were washed and the excess, uninhibited antibody-enzyme conjugate (which had become bound to the surfaced adsorbed antigen), was mea- sured. The quantity of antigen, i.e. Fc or Fab, in the test samples was determined from a standard curve established with known amounts of Fc or Fab used as inhibitors. All data were analyzed using EL- ISANALYSIS (Peterman and Butler, 1987; Butler et al., 19856). For indirect competitive assays measuring
IgGl-Fc or IgG2-Fc, an appropriate Fc-enriched fraction was used as a standard because use of intact IgGl or IgG2 failed to generate a titration plot with the same slope as that generated by digests. Data on the distribution of Fab among CF fractions is accu- rate in absolute units while the distribution of IgG2a-Fc and IgGl-Fc is estimated because neither Fc can be obtained in completely pure form for use as an absolute standard (see Fig. 5).
Determination of specific antibody activity. The IgG2a antibody activity of various subpopulations of IgG2a eluted from DEAE-Sephadex and the Fab subpopulations eluted during chromatofocusing were
tested against seven antigens using the a-ELISA (Butler et al., 1978): phosphorylcholine (PC), dinitro- phenyl (DNP), fluorescein (FLU), Brucella abortus soluble antigen (BASA), staphylococcal cc-toxin, bo- vine parvovirus and strain K-99 E. co/i. Escherichia coli was sonicated and adsorbed on microtiter wells
in the presence of methyl glyoxal (Czerkinsky et al., 1983) while the other antigens were adsorbed at pH9.6 in carbonate buffer (Butler et a/., 1978). All haptens were conjugated to a gelatin carrier. FLU-gelatin was prepared with ditriazinolamino fluorescein as described by Blakeslee and Baines (1976). PC-gelatin was prepared with diazonium phenylphosphorylcholine as previously described (Cheseboro and Metzger, 1972; Butler et al., 1985) and DNP-gelatin was prepared using dinitrobenzene sulfonate as described by Eisen (1964).
The activity of each fraction tested was compared to DEAE fraction A (Fig. 3) from DEAE-Sephadex A-50 which was assigned a value of 100 ELISA units/ml. Data acquisition and analysis were done as
described in ELISANALYSIS (Peterman and Butler, 1987; Butler et al., 1985b).
RESULTS
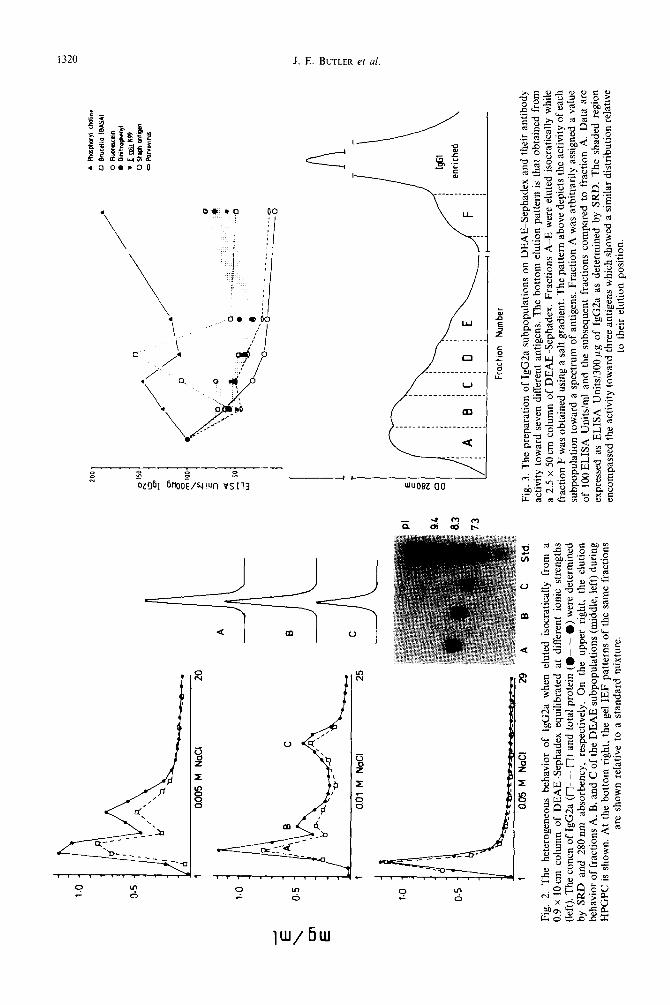
The data of Fig. 1 confirm the original obser- vations of Duncan et al. (1972) and have been repeatedly observed in our laboratory (Whitney, 1976; Butler et at., 1980); these data show that IgG2 elutes in virtually every fraction from a DEAE-anion exchanger. Figure 1 shows that: (1) the initial peak appearing with the equilibration buffer is pure IgG2a, (2) the initial peak tails off into smaller peaks, (3) the first peak which appears on addition of the gradient (the leading shoulder of the IgGl-enriched peak) is mostly IgG2a and (4) up to 20% of the IgGl-enriched peak is composed of IgG2a. This extensive ion-exchange heterogeneity was seen also in
I - 00 280 D 0 IgG, n
i FRACTION NUMBER PO
Fig. 1. The elution of bovine I&s from a 2.5 x 18 cm column of DEAE-Sephadex. The-initial peaks were eluted isocraticallv at DH 8.5. in 0.01 M Tris containing 0.05 M NaCl whilk the-last was eluted using an NaClgradient produced with a limit buffer containing 0.5 M NaCl. The distribution of total protein was determined by absorbancy at 280 nm and IgG2a and IgGl concns were determined by single radial diffusion. The salt gradient was initiated in the
trough separating the major two peaks.
0.00
5 M
Na
Cl
0.01
M
Na
Cl
Fig.
2.
T
he
hete
roge
neou
s be
havi
or
of
IgG
2a
whe
n el
uted
is
ocra
tical
ly
from
a
0.9
x IO
cm
colu
mn
of
DE
AE
-Sep
hade
x eq
uilib
rate
d at
di
ffer
ent
ioni
c st
reng
ths
(lef
t).
The
co
ncn
of
IgG
2a
(n--
-n)
and
tota
l pr
otei
n (@
---0
) w
ere
dete
rmin
ed
by
SRD
an
d 28
0nm
ab
sorb
ency
, re
spec
tivel
y.
On
the
uppe
r ri
ght,
the
elut
ion
beha
vior
of
fra
ctio
ns
A,
B,
and
C o
f th
e D
EA
E
subp
opul
atio
ns
(mid
dle,
le
ft)
duri
ng
HPG
PC
is s
how
n.
At
the
botto
m
righ
t, th
e ge
l IE
F
patte
rns
of
the
sam
e fr
actio
ns
Fig.
3.
The
pr
epar
atio
n of
Ig
G2a
su
bpop
ulat
ions
on
D
EA
E-S
epha
dex
and
thei
r an
tibod
y ac
tivity
to
war
d se
ven
diff
eren
t an
tigen
s.
The
bo
ttom
el
utio
n pa
ttern
is
tha
t ob
tain
ed
from
a
2.5
x 50
cm
co
lum
n of
D
EA
E-S
epha
dex.
Fr
actio
ns
A-E
w
ere
elut
ed
isoc
ratic
ally
w
hile
fr
actio
n F
was
ob
tain
ed
usin
g a
salt
grad
ient
. T
he
patte
rn
abov
e de
pict
s th
e ac
tivity
of
eac
h su
bpop
ulat
ion
tow
ard
a sp
ectr
um
of a
ntig
ens.
Fr
actio
n A
was
ar
bitr
arily
as
sign
ed
a va
lue
of
IOO
EL
ISA
U
nits
/ml
and
the
subs
eque
nt
frac
tions
co
mpa
red
to
frac
tion
A.
Dat
a ar
e ex
pres
sed
as
EL
ISA
U
nits
/300
pg
of
IgG
2a
as
dete
rmin
ed
by
SRD
. T
he
shad
ed
regi
on
enco
mpa
ssed
th
e ac
tivity
to
war
d th
ree
antig
ens
whi
ch
show
ed
a si
mila
r di
stri
butio
n re
lativ
e ar
e sh
own
rela
tive
to
a st
anda
rd
mix
ture
. to
th
eir
elut
ion
posi
tion.
Frac
tion
Num
ber

Bovine IgG2 heterogeneity 1321
the IgG2 which eluted isocratically and was studied in more detail using analytical DEAE-Sephadex col- umns equilibrated at different ionic strengths.
Figure 2 (left) illustrates that when bovine serum immunoglobulins from an Al/Al steer are iso- cratically eluted from DEAE ion-exchange columns, multiple subpopulations can be obtained. When tested by SRD and competitive ELISA, only lgG2a was present among the isocratically eluted fractions and the IgG2a concn approximately equalled the total protein concn estimated spec- trophotometrically.
When the molecular size of the different iso- cratically eluted IgG2a subpopulations was examined by HPGPC, all eluted as intact IgG2a (Fig. 2; upper right). When the subpopulations were tested by gel IEF, their mean Ips differed and decreased with increasing retention time during isocratic elution; this would be predicted if their elution behavior were charge-dependent (Fig. 2; lower right).
The basis of this charge-related heterogeneity of IgG2a was further investigated in four experiments. Firstly, we examined the spectrotype of the H- and L-chains from IgG2a populations which could be purified free of IgGl (Fig. 3, fractions A-E). Sec- ondly, we measured the carbohydrate content of each
DEAE subpopulation. Thirdly, we determined whether the charge-dependent heterogeneity, mea- sured by CF, was confined only to the Fab portions of each subpopulation and whether carbohydrate content was differently distributed among CF frac- tions. Finally, we examined the antibody activity of each DEAE subpopulation to seven different anti- gens.
The material necessary for such studies was pre- pared from 4g of serum globulins from an Al/Al animal. These were fractionated preparatively on a 2.5 x 50 cm column of DEAE-Sephadex equilibrated at 0.05 M NaCl in a 0.01 M Tris-HCl buffer. The IgG2a was eluted isocratically and subpopulations were assigned at regions in the elution patterns where a tendency for separate peaks to appear was evident (Fig. 3; bottom). It has been our experience that resolution among isocratically generated sub- population is lower when larger and longer columns are used; this phenomenon was also evident when analytical columns of different lengths were used (Butler et al., 1987~). The fractions which eluted isocratically were designated A-E and the protein, carbohydrate and IgG2a recovered in each was deter- mined (Table 1). While the actual concn values varied depending on the method of analysis, relative differences between fractions were the same and all protein could be accounted for as IgG2a. When no further protein eluted from the columns, a NaCl gradient was applied in which the final buffer was 0.01 M Tris-HCl, pH 8.3 and 0.5 M NaCl. This procedure generated an IgG2a-enriched fraction which preceded the elution of the major IgGl peak (see Fig. 1). The IgG2a in this fraction F was also

1322
Fig. 4. The spectrotypic analyses of DEAE fractions A-D. Patterns for the intact IgG2a subpopulation, H- and L-chains are. shown. D/E refers to the fact that in certain DEAE fractionations like that shown
in Fig. 3, fractions D and E were pooled. Legend on figure.
studied in experiments involving CF but not in those in which purified H- and L-chains were examined; because fraction F also contained IgG2b and IgGl (Butler, 1985) examination of H- and L-chains pre- pared from this fraction could only lead to ambigu- ous results.
Figure 4 shows the spectrotypic characteristics of the H- and L-chains obtained from four of the different IgG2a subpopulations indicated in Fig. 3; patterns for the H-chains, L-chains and intact IgG2a of these same subpopulations are shown. These data show that the spectrotype of the isolated H-chains from different subpopulations was heterogeneous but most similar to the spectrotype of the intact IgG2a subpopulations from which they were obtained. Light chains from all four subpopulations tested were also heterogeneous, showed no differences between sub- populations, and appeared to focus into groups in the regions of pH 8.7, 7.5 and 6.3. These results extend those of Fig. 2 which indicated that the ion-exchange behavior of various intact IgG2a subpopulations was paralleled by their IEF patterns; here we show that these differences in mean Ip are primarily a con- sequence of heavy chain spectrotypic variation. The mean pls of the different preparations are sum- marized in Table 1.
The data of Fig. 4 suggest that the heterogeneous behavior of Al/Al homozygous IgG2a during elut- ion from DEAE-Sephadex primarily resides in the H-chains of the IgG2a. Because no constant region isotypic or allotypic variation should be present among the Al/Al IgG2a subpopulations, such vari- ation must reside in the Vn-region and it is possible that the activity of the various IgG2a subpopulations for various antigens might differ. This was tested using seven antigens to which most cattle have antibodies. Methodologically, the concn of IgG2a in fractions A-F of the eluted protein from DEAE-Sephadex was carefully measured by SRD. Such analysis was especially important in the case of fraction F because this fraction also contained IgGl and some IgG2b. The antibody activity was measured using the a-ELISA and fraction A was
arbitrarily assigned a value of 100 E.U./ml. The results are plotted above the DEAE elution profile shown in Fig. 3 (top). Each point is the result of 2-3 determinations; each determination was obtained from ELISANALYSIS using a 7-well doubling dilu- tion sequence. The results show that at least four different patterns could be seen. PC activity increased with the decreased mean pI of the IgG2a while the response to both staphylococcal u-toxin and bovine parvovirus behaved in the opposite manner. The response to BASA was unique while the response to FLU, DNP, and K99 E. cofi all showed a similar pattern (these are highlighted by shading).
The third approach to testing the hypothesis that Vn-region heterogeneity could explain the isocratic elution behavior of IgG2a involved digestion of each fraction with mercuripapain. Digests of mouse mono- clonal antibodies contain only Fc and Fab fragments (Ed Voss, Jr, personal communication) and HPGPC confirmed that such digests of IgG2a contained no intact IgG2a after 2 hr (data not shown). All digests tested by HPGPC showed a major protein peak of ca 40 K mol. wt and a leading shoulder which could account for approx. 30% of the total protein and which had a molecular size of ca 56 K. These Fc/Fab mixtures for each fraction were then fractionated by chromatofocusing (Fig. 5). The quantity of Fab and IgG2a-Fc present in each chromatofocusing fraction was quantitated using an indirect competitive ELISA (Fig. 5). For fraction F, the quantity and distribution of IgGl-Fc was also determined. Because of the presence of both IgG2a and IgGl in fraction F, the distribution of Fab observed cannot be totally as- cribed to IgG2a. Finally, we measured the antibody activity of the Fab fragments separately by chro- matofocusing the digest of DEAE fraction E (Fig. 6).
The data of Fig. 5 demonstrate numerous phenom- ena. Firstly, Fab fragments elute heterogeneously and in both CF peaks while the IgG2a-Fc elutes homoge- neously with a pI of ca 6.0. Some minor tailing of Fc, e.g. fraction A, was occasionally observed but ac- counted for < 10% of the total Fc. Secondly, the IgG2a-Fc eluted at the same pl regardless of which

Bovine IgG2 heterogeneity 1323
Fig. 5. The chromatofocusing behavior of selected digests of IgG2a subpopulations purified by DEAE-Sephadex. Patterns for fractions B and D were omitted for space consideration but produced a pattern intermediate between the alphabetically designed fractions either before or after. The thin, solid line indicates the quantitative distribution of protein, (---) = Fab, (-.-) = IgG2a-Fc and the distribu- tion of IgGI-Fc is indicated by shading. The units of the y-axis are exact for measurements of total protein (as determined by absorbance at 280 nm) and Fab but only estimates are shown for Fc because of the lack of a completely pure Fc (see text). The pH gradient is shown as a heavy dark line. IgG2a fractions which did not contain IgGl (fractions A-E) could be analyzed on pH 8-5 CF columns while those
containing IgGI (and IgG2) required 84 gradients. i.e. fraction F and purified IgGl.
DEAE fraction was tested while the distribution of
Fab in CF continuously shifted to lower pl as DEAE fractions A-F were sequentially fractionated (Fig. 5). Hence, there was a gradual decrease in the relative size of first CF fractions obtained as digests of DEAE fractions A-F were sequentially analyzed. Thirdly, the CF fractionation was reproducible when IgG2a sub-
populations obtained from different, but parallel, DEAE runs were tested. This is illustrated for the
fractions which contained IgGI. i.e. fraction F, had to be chromatofocused over a range from pH 8 to pH 4 rather than from pH 8 to 5 as was done for those fractions containing only IgG2a. When an 8-4 gradient was used for fractions containing only IgG2a, similar profiles as shown for fractions A-E (Fig. 5) were obtained (data not shown). Fifthly, the IgGZFc in fraction F (which contains also IgG2b and IgGI) eluted at the same pH as the IgG2a-Fc in
1.0.
0.6s
I _ SO.,-
,"
5 20.4.
I c
2 l.O-
pH 5.9 f
n
5 0.8-
a h r
I _
I % :: ,”
E
i
-25
-20
15
-10
3 6 9 12 15 16 3 6 9 12 15 16
C F Fraction CF Fraction
Fig. 6. The distribution of carbohydrate among CF fractions obtained with papain digests of DEAE fractions A and C. (o-0) Protein distribution measured by absorbancy at 280nm. (m--m) Neutral hexose content measured by the micro phenol-sulfuric acid test. The vertical arrows indicate the
pH and position at which the IgG2a-Fcs elute.

1324 J. E. BUTLER er al
70 - DNP Q
65 b-dFLU e--S A.
60 c? 0 PC.
/ 5 6 1 8 9 10 11 12 13 14 15
CF Fraction Number
Fig. 7. The antibody activity of the Fab fragments separated by CF from DEAE subpopulation E. All activity is nor- malized in terms of ELISA Units/2OOHg of Fab and CF fraction A is assigned a value of 100 ELkSA Units/200 pg. DNP = dinitroohenvl-gelatin: FLU = fiuorescvi eelatin: S.A. =purified’ an&n from S. aweus; and F.C. =
phosphoryl choline conjugated to gelatin.
digests of DEAE subpopuIations A-E. The results of the CF studies are also summarized in Table 1.
The data of Fig. 5 provide two additional pieces of information not directly related to the hypothesis under investigation, but which are nevertheless note- worthy. Firstly, the IgGl-Fc in fraction F appeared to behave more heterogeneously than does the con- trol IgGl preparation which had been digested. The control IgGl was purified from colostral whey and its Fc showed the same marked difference in charge from the Fc of IgG2a which we showed previously by elcctrophoresis (Butler, 1983). Secondly, unless only the IgG2a eluted isocratically from DEAE-Sephadex as fraction A was used as starting material, it was impossible to completely purify the Fc portion of polyclonal bovine IgG2a or IgGl from their Fabs using only charge-dependent methodology.
The possibility that differences in the carbohydrate concn of the various IgG2a subpopulations might explain their ion-exchange behavior or influence the behavior of papain digests in CF. was also examined. Table 1 shows that the carbohydrate content of the various DEAE-derived subpopulations did not differ significantly and any trend was opposite to that expected, if carbohydrate significantly influenced to- tal charge. Figure 6 presents CF data on papain digest of two representative subpopulations, “A” and “C”. In both digests, carbohydrate emerges as a single major peak, the pi of which is the same as that of the IgG2a-Fc for that subpopulation (Table 1); the elution of carbohydrate parallels the elution of Fc (Figs 5 and 6).
The heterogeneity of the IgG2a eluted from DEAE-Sephadex was further demonstrated by com- paratively testing the antibody activity of the Fab fractions obtained by CF from a representative IgG2a subpopulation. Figure 7 presents such data for DEAE fraction E. In each case, the antibody activity was normalized to total Fab content. Data showed that activities vary greatly among fractions; the same pattern seen when anti-FLU activity was measured, was also seen for anti-DNP. Activity to staphy- lococcat u-toxin was highest in CF fraction 5 and failed to show the high activity in CF fractions 8 and 15 which were characteristic of anti-FLU or anti- DNP.
DISCUSSION
The heterogeneous behavior of normal serum im- munoglobulins, especially IgG, during ion-exchange chromatography is commonly observed in all major species and can also be observed using charge- dependent methods like immunoelectrophoresis; het- erogeneity observed in the latter contrasts sharply with the restricted immun~lectrophoretic behavior of myeloma protein in monoclonal gammapathies (Clausen, 1971). The latter observation leads to the prediction that charge-related heterogeneity of nor- mal serum immunoglobulins is the result of their polyclonality, i.e. the result of V, and V, hetero- geneity. Among ruminants, a similar charge- dependent heterogeneity is seen (Murphy et al., 1965) but for this group of mammals, this observation encompasses another issue because the two major ruminant subclasses, IgGl and IgG2, are still defined by many on the basis of their charge-dependent properties (reviewed by Aalund, 1968; Butler. 1969, 1985). Such a definition has been extended to “sub- subclass” designations (Kickhofen et ul., 1968); such subsubclasses have been associated with different biological activities (Mossmann et al., 1981). The anion-exchange behavior of bovine serum IgGs is particularly complicated by the heterogeneity of IgG2 which elutes throughout the entire chromatogram (Fig. I). Furthermore, IgG2 in cattle can be differentiated into IgG2a which bears the Al or A2 allotypic markers, and IgG2b which does not (Butler et al., 1987b).
The work reported here examines the ion-exchange behavior of bovine IgG in a model system in which allotypic and isotypic differences play no role. Hence, we are able to focus on the ion-exchange behavior of a single isotype bearing a single allotype, IgG2a-Al. The data presented in Figs 1-3 indicate that within the subclass of IgG2a-A 1, multiple subpopulations can be obtained by Anton-exchange chromatography. Figure 2 shows that ionic strength influences the extent to which discrete subpopulation, i.e. “peaks”, are obtained. We show elsewhere (Butler et al.,
1987~) that bovine IgG behaves similarly on DEAE-Sephacel and DEAE-Sepharose and de- pending on column length and ionic strength, at least

Bovine IgG2 heterogeneity 1325
four lgG2a subpopulations can also be obtained. However, the higher DEAE substitution of DEAE-Sephadex means that higher ionic strength buffers are needed to yield the same result as obtained in DEAE-Sephacel at l/S the ionic strength. QAE-Sephadex behaves much like DEAE- Sephadex. Hence the generation of IgG2a sub- populations on anion-exchange columns is not a phenomenon unique to DEAE-Sephadex.
Figure 2 also shows that the IgG2a-Al sub- populations differ in their pl and are not a con- sequence of proteolytic degradation. When the heavy and light chain spectrotype of four of the DEAE subpopulations (Fig. 3) was examined, differences were almost exclusively associated with the heavy chains (Fig. 4). These subpopulations did not differ in their content of carbohydrate (Table 1). Further- more, gel IEF comparison can only be made on IgG2a subpopulations which are completely free of IgGl and IgG2b, and data obtained by IEF are difficult to quantitate accurately. Hence we choose to examine the heterogeneity of Fab and IgG2a-Fc by chromatofocusing so that Fc-specific reagents could be used to avoid ambiguity caused by contamination with IgGl and IgG2b. Figure 5 demonstrates that the charge-dependent heterogeneity of the IgG2A is asso- ciated with the Fab fragment, not the IgG2a-Fc. When the carbohydrate content of eluting CF frac- tions was measured: (1) carbohydrate concns paral- leled the elution of IgG2a-Fc and (2) no differences in carbohydrate with respect to DEAE fraction, were observed (Fig. 6; Table 1). Hence differences in heavy chain spectrotype (Fig. 4) most likely result from the Vn-region protein heterogeneity.
The data on differential antibody activity presented in Figs 3 and 7 indicate that the Vn-related differences behave like idiotypic variants which are charge- related and influence the antibody binding site. Evi- dence that the Vn-region is more important in the specificity of the antibody binding site than is the V,, is well documented (Haber and Richards, 1966; see Nisonoff et al., 1975).
The semi-discrete character or clustering of IgGZa-Al into subpopulations is in itself interesting. This clustering is seen in all three charge-dependent
methods used, i.e. anion-exchange chromatography, IEF and the CF distribution of Fab. Light chains which showed no intersubpopulation variation were nevertheless clustered into subpopulations, definable as having different mean pls (Fig. 4; Table 1). It is unlikely that such nonrandom distribution is circum- stantial; it may reflect constraints on the potential of V-D-J or V-J combinations as well as other factors beyond the scope of this report.
The data presented here indicate that the sub- classes or subsubclasses of bovine IgG should not be based on “peaks” obtained from anion exchange columns. Attempts to assign bovine IgG to the subclasses IgG2a, IgG2b or IgGl should be based on antigenic or molecular genetic criteria rather than
ion-exchange behavior. While observations on differences in biological activity are important in gaining an understanding of the ruminant immune system, the assignment of such activities to isotypic differences rather than to other forms of antibody variation which can influence their ion-exchange be- havior, should be done cautiously.
The mercuripapain digestion procedure used ini-
tially for mouse monoclonal antibodies (Voss. per- sonal communication) works well for both bovine IgG2a and IgGl. The behavior of the IgG2a digests seen by HPGPC indicates that no intact IgG remains and that the Fc portion is somewhat larger than the
Fab or that the gel permeation behavior of one or the
other is selectively influenced by either hydrophobic
or hydrophilic interactions with the Synchrom GPC column matrix.
In summary, IgG2a from a steer homozygous at the A-locus behaves heterogeneously on DEAE ion-
exchange columns. Such heterogeneity was shown biochemically to be associated with the Vn-region of the IgG2a H-chain and was associated with pro- nounced differences in activity toward various anti- gens. The invariant behavior of Fc fragments from different IgG2a subpopulations is consistent with isotypic homogeneity and differs by one full pH unit from the CF behavior of colostral IgGl-Fc. Hence, the heterogeneous behavior of bovine IgG2 on anion exchange columns appears to be a result of V, variations so that assignment of subsubclass status on the basis of charge-dependent behavior should not be done without supporting data on constant region variation.
REFERENCES
Aalund 0. (1968) Heterogeneity of Ruminant Immuno- globulins. Munksgaard, Copenhagen.
Blakeslee D. and Baines M. G. (1976) Immunofluorescense using DTAF I. Preparation and fractionation of labelled IgG. J. Immun. Merh. 13, 305-320.
Blakeslee D., Butler J. E. and Stone W. H. (1972a) Serum antigens of cattle II. Immunogenetics of two immu- noglobulin allotypes. J. Immun. 107, 227-235.
Blakeslee D., Rapacz J. and Butler J. E. (197lb) Bovine immunoglobulin allotypes. In Symposium: Bovine Im- mune Svstem (Edited bv Butler J. E.. Winter A. J. and Wagner-G. G.). J. Dair$ Sci. 54, 132LLl321.
Butler J. E. (1969) Bovine immunoglobulins: a review. J. Dairy Sci. 52, 1895-1909.
Butler J. E. (1981) A concept of humoral immunity among ruminants and an approach to its investigation. In The Ruminant Immune System (Edited by Butler J. E., Neilson K. and Duncan J. R.), pp. 3355. Plenum Press, New York.
Butler J. E. (1983) Bovine immunoglobulins: an augmented review. Pet. Immun. Immunopath. 4, 43-152.
Butler J. E. (1986) Biochemistry and biology of ruminant immunoglobulins. In Progress in Veterinary Microbiology and Immuno1og.v (Edited by Pandey R.), Vol 2, pp. l-53. S. Karger, Base].
Butler J.- E., Cambier J. C. and Klobasa F. (1985a) Identification and characterization of a hapten-modifiable TEPC-15 cross-reactive idiotype in swine. Mo/ec. Immun. 22, 1159--l 168.

I326 J. E. BUTLER er al.
Butler J. E. and Heyermann H. (1986) The heterogeneity of bovine IgG2-I. The Al allotypic determinant is the major aniigenic determinant recognized on bovine IgG2a bv oolvclonal rabbit anti-IgGZa. M&c. fmmun. 23, 291-1296.
Butler J. E., Heyermann H., Bielecka M. and Dillender M. (1987a) The heterogeneity of bovine IgG2. IV. IgG subpopulations obtained by anion-exchange chro- matography do not reflect isotypic variation. Am. J. ret. Res. (submitted).
Butler J. E.. Heyermann H., Borca M. and Bielecka M. (1987~) The isotypic, allotypic and idiotypic hetero- geneity of bovine IgG2. Proc. First Int. Congr. Vet. Immun. (Edited by Wilkie B. N.). Vet. Immun. Immuno- path. (in press).
Butler J. E.. Heyermann H., Frenyo V. L. and Borca M. (1987b) The heterogeneity of bovine IgG2. II. The identification of IgG2b. Immun. Lett. (pending).
Butler J. E. and Kennedy N. (1978) The differential enzyme susceptibility of bovine IgGl and IgG2 to pepsin and papain. Biochim. biophys. Acta 535, 125-137.
Butler J. E., McGivern P. L., Cantarero L. A. and Peterson L. (1980) Application of the amplified enzyme-linked immunosorbent assay: comparative quantitation of bo-
(1986) Immunoglobulins of colostrum-VI. Comparative studies of cytophilic properties of bovine serum and colostral IgG. Molec. Immun. 23, 961-964.
Haber E. and Richards F. F. (1966) The specificity of antigenic recognition of antibody heavy chain. Discussion of chemistry and biology of immunoglobulin. Proc. R. Sot. 166B, 176.
Hajdu S., Beseda I. and Polek B. (1974) Physical chemical and serological properties of Brucellu IgGl and IgG2 antibodies in cattle. Vet. Med. Prague 19, 141~ 150.
Harrington J. C., Fenton J. W., III and Pert J. H. (1971) Polymer-induced precipitation of antigen-antibody com- plexes: “Precipiplex” reactions. Immunochemistrv 8, 413421.
Heyermann H. and Butler J. E. (1987) The heterogeneity of bovine IgGZ-IV. Structural differences between IgG2a molecules of the Al and A2 allotypes. M&c. ~mmun. 24, 1317 1324.
Kickhofen B., Hammer D. K. and Scheel D. (1968) Iso- lation and characterization of gG type immunoglobuhns from bovine serum and colostrum. Hoppr-Sevler’.~ %. physiol. Chem. 349, 1755 -I 773.
Nature, Lund. 221, 68OG685. Lisowski J., Janusz M., Tyran B., Morawiecki A., Golab S.
Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4.
Butler J. E., McGivern P. L. and Swanson P. (1978)
vine serum IgGl, IgG2, IgA and IgM~antibodies. Am. J.
Amplification of the enzyme-linked immunosorbent assay
wt. Res. 41. 1479-1491.
(ELISA) in the detection of class-soecific antibodies. J. immun.‘Meth. 20, 365-383. .
Butler J. E. and Maxwell C. F. (1972) Preparation of bovine immunoglobulins and free secretory component and their specific antisera J. Dairy Sci. 55, 151 -164.
Butler J. E.. Peterman J. H. and Koertge T. E. (19856) The amplified enzyme-linked immunosorbent assay (a- ELISA). In Enzyme Mediated Immunoassay (Edited by Ngo ‘T. T. and Lenhoff H. M.), pp. 241-276. Plenum Press, New York.
and Bialkowska H. (1975) Immunoglobulins of colostrum IV. Comparative studies of bovine serum and colostral IgGl and IgG2. Immunochemistr,v 12, 167.
Mach J.-P. and Pahud J.-J. (1971) Secretory IgA, a major immunoglobulin in most bovine external secretions. J. Immun. 106, 552-563.
Milstein C. P. and Feinstein A. (1968) Comparative studies of two types of bovine immunoglobulin heavy chains. Biochem. J. 107, 559-564.
Mossmann H., Schmitz B., Possart P. and Hammer D. K. (1981) Antibody-dependent ceil-mediated cytotoxicity in cattle: transfer of IgG2 subclasses in relation to the population of the newborn calf. In The Ruminant Immune System (Edited by Butler J. E., Neilson K. and Duncan J. R.), pp. 279-292. Plenum Press, New York.
Murphy F. A., Osebold J. W. and Aalund 0. (1965) Physical heterogeneity of bovine TM and YG globulins. Archs Biochem. Biophys. 112, 126-136.
Nisonoff A., Hopper J. E. and Spring S. B. (1975) The Antibody Molecule, p, 254. Academic Press, New York.
Peterman J. H. and Butler J. E. (1987) Apphcation of the Mass hW to the development of a computerized system for the analysis of ELISA data. J. Immun. Mrfh. (submit- ted).
Cheseboro B. and Metzger H. (1972) Affinity labelling of a phosphorylcholine binding mouse myeloma protein. Bio- chemistry 11, 76677 1.
Clausen J. (1971) Immunoelectrophoresis-a survey of its application in clinical chemistry. In Handbook of Immunodiffision and Immunoelectrophoresis (Edited by Ouchterlony 0.) pp. 1899201. Ann Arbor Science, Ann Arbor.
Czerkinsky C., Rees A. S., Bergmeier L. A. and Challacombe S. J. (1983) The detection and specificity of class specific antibodies to whole bacterial cells using a solid phase radioimmunoassay. C[in. rxp. Immun. 53, 192-200.
Duncan J. R.. Wilkie B. N., Hiestand F. and Winter A. J. (1972) The serum and secretory immunoglobulins of cattle: characterization and quantitation. J. Immun. 108, 965 -976.
Eisen H. (1964) Preparation of purified anti-2,4- dinitrophenyl antibodies. In Mefh. Medical Research IO (Edited by Eisen H.), pp. 94102. Year Book Medical Publishers, Chicago.
Fahey J. L. and McKelvey E. M. (1965) Quantitative determination of serum immunoglobulins in antibody- agar plates. J. Immun. 94, 8490.
Gorczyca W., Ugorski M., Nowacki W. and Lisowski J.
Rowe D. and Fahey J. L. (1965) A new class of human immunoglobulins. II. Normal serum IgD. J. rxp. Med. 121? 1855199.
Schmttz B. (1979) Antikoerpervermittelte Zytotoxizitaet der peripheren Leukozyten des Rindes: Untersuchungen zur Altersabhaengigkeit und IgG Subklassenspezihtaet. Inaug. Diss. Justus-Liebig Univ., Giessen. 82 pp.
Whitney R. (1976) Nomenclature of the proteins of cow’s milk. Fourth version. J. Duiry Sci. 59, 7955815.
Willoughby E. W. and Lambert A. (1982) A sensitive silver stain for proteins in agarose gels. Anulyl. Biochem. 130, 353-358.