
The expression of keratinocyte growth factor receptor (FGFR2-IIIb) correlates with the high...
10
The expression of keratinocyte growth factor receptor (FGFR2-IIIb) correlates with the high proliferative rate of HaCaT keratinocytes Introduction Keratinocyte growth factor receptor (FGFR2- IIIb ¼ KGFR) is a transmembrane tyrosine kin- ase receptor mediating the effect of several fibroblast growth factors (FGF) such as FGF-1 (a-FGF), FGF-7 (KGF-1), FGF-10 (KGF-2) and as indirect evidence suggests, FGF-22. KGF-1 and KGF-2 are distinguished from other members of the FGF family by their specific mitogenic effects on a wide variety of cells, like keratinocytes, hepatocytes, intestinal epithelial cells and osteoblasts (1). They play important roles in cellular stress responses protecting cells from various damaging effects like radiation or oxidative stress (1). All the above effects are mediated by FGFR2-IIIb, thus it plays Nagy N, Bata-Cso¨rg} o Zs, Kopasz N, Szeg Cs, Pivarcsi A, Koreck A, Dobozy A, Keme´ny L, Sze´ll M. The expression of keratinocyte growth factor receptor (FGFR2-IIIb) correlates with the high prolifer- ative rate of HaCaT keratinocytes. Exp Dermatol 2006: 15: 596–605. Ó 2006 Department of Dermatology and Allergology, University of Szeged. Journal compilation Ó 2006 Blackwell Munksgaard. Abstract: Keratinocyte growth factor receptor (KGFR ¼ FGFR2- IIIb) is a tyrosine kinase receptor expressed by keratinocytes, which mediates the effects of fibroblast growth factors (FGF). There are con- tradictory data in the literature regarding the role of FGFR2-IIIb dur- ing the proliferation/differentiation programme of keratinocytes. In this study, we aimed to investigate whether overexpression of FGFR2- IIIb may have a role in the regulation of keratinocyte proliferation. We analysed the expression of FGFR2-IIIb in an in vitro HaCaT model system representing different stages of proliferation and differ- entiation of keratinocytes. Real-time RT-PCR and Western blot ana- lyses demonstrated a correlation between FGFR2-IIIb mRNA and protein expression and the proportion of cells in S/G2/M phase in synchronized HaCaT keratinocytes and thus with proliferation activity (r ¼ 0.96). After treatment with the antipsoriatic drug, dithranol, FGFR2-IIIb is downregulated dose dependently both at mRNA and protein levels. Moreover, when the rate of proliferation is decreased by the lack of cell attachment to the culturing surface, FGFR2-IIIb mRNA (P ¼ 0.0315) and protein expressions were also reduced (P ¼ 0.0242), while a differentiation marker, keratin 10, mRNA (P ¼ 0.0003) and protein levels (P ¼ 0.001) were increased (r ¼ )0.92). Based on our results we conclude that FGFR2-IIIb expression in HaCaT keratinocytes corresponds with the proliferative activation of the cells and is not related to the differentiation programme. Nikoletta Nagy 1 , Zsuzsanna Bata-Cso¨rg} o 1,2 , Norbert Kopasz 1 , Csilla Szeg 1 , Andor Pivarcsi 2 , Andrea Koreck 1 , Attila Dobozy 1,2 , Lajos Keme´ny 1,2 and Ma´rta Sze´ ll 2 1 Department of Dermatology and Allergology, University of Szeged, Szeged, Hungary; 2 Dermatological Research Group of the Hungarian Academy of Sciences and the University of Szeged, Szeged, Hungary Key words: differentiation – dithranol – HaCaT keratinocytes – keratinocyte growth factor receptor – proliferation Ma ´rta Sze ´ll Dermatological Research Group of the Hungarian Academy of Sciences and the University of Szeged 6720 Szeged Kora ´nyi fasor 6 Hungary Tel.: 36 62 545 278 Fax: 36 62 545 954 e-mail: [email protected] Accepted for publication 9 May 2006 Abbreviations: AcC, acetone-treated control; C, untreated control; DMEM, Dulbecco’s modified Eagle’s medium; FBS, fetal bovine serum; FGF, fibroblast growth factor; FGFR2-IIIb (KGFR), ker- atinocyte growth factor receptor; K1, keratin 1; K10, keratin 10; KGF, keratinocyte growth factor; Q-RT-PCR, quantitative real- time polymerase chain reaction; SDS, sodium dodecyl sulphate. 596 Experimental Dermatology 2006: 15: 596–605 Blackwell Munksgaard . Printed in Singapore doi: 10.1111/j.0906-6705.2006.00450.x Ó 2006 Department of Dermatology and Allergology, University of Szeged. Journal compilation Ó 2006 Blackwell Munksgaard. Experimental Dermatology ISSN 0906-6705
Transcript of The expression of keratinocyte growth factor receptor (FGFR2-IIIb) correlates with the high...

The expression of keratinocyte growthfactor receptor (FGFR2-IIIb) correlateswith the high proliferative rate of HaCaTkeratinocytes
Introduction
Keratinocyte growth factor receptor (FGFR2-IIIb ¼ KGFR) is a transmembrane tyrosine kin-ase receptor mediating the effect of severalfibroblast growth factors (FGF) such as FGF-1
(a-FGF), FGF-7 (KGF-1), FGF-10 (KGF-2)and as indirect evidence suggests, FGF-22.KGF-1 and KGF-2 are distinguished from othermembers of the FGF family by their specificmitogenic effects on a wide variety of cells, likekeratinocytes, hepatocytes, intestinal epithelialcells and osteoblasts (1). They play importantroles in cellular stress responses protectingcells from various damaging effects like radiationor oxidative stress (1). All the above effectsare mediated by FGFR2-IIIb, thus it plays
Nagy N, Bata-Csorg}o Zs, Kopasz N, Szeg Cs, Pivarcsi A, Koreck A,Dobozy A, Kemeny L, Szell M. The expression of keratinocytegrowth factor receptor (FGFR2-IIIb) correlates with the high prolifer-ative rate of HaCaT keratinocytes.Exp Dermatol 2006: 15: 596–605. � 2006 Department of Dermatologyand Allergology, University of Szeged. Journal compilation � 2006Blackwell Munksgaard.
Abstract: Keratinocyte growth factor receptor (KGFR ¼ FGFR2-IIIb) is a tyrosine kinase receptor expressed by keratinocytes, whichmediates the effects of fibroblast growth factors (FGF). There are con-tradictory data in the literature regarding the role of FGFR2-IIIb dur-ing the proliferation/differentiation programme of keratinocytes. Inthis study, we aimed to investigate whether overexpression of FGFR2-IIIb may have a role in the regulation of keratinocyte proliferation.We analysed the expression of FGFR2-IIIb in an in vitro HaCaTmodel system representing different stages of proliferation and differ-entiation of keratinocytes. Real-time RT-PCR and Western blot ana-lyses demonstrated a correlation between FGFR2-IIIb mRNA andprotein expression and the proportion of cells in S/G2/M phase insynchronized HaCaT keratinocytes and thus with proliferation activity(r ¼ 0.96). After treatment with the antipsoriatic drug, dithranol,FGFR2-IIIb is downregulated dose dependently both at mRNA andprotein levels. Moreover, when the rate of proliferation is decreasedby the lack of cell attachment to the culturing surface, FGFR2-IIIbmRNA (P ¼ 0.0315) and protein expressions were also reduced (P ¼0.0242), while a differentiation marker, keratin 10, mRNA (P ¼0.0003) and protein levels (P ¼ 0.001) were increased (r ¼ )0.92).Based on our results we conclude that FGFR2-IIIb expression inHaCaT keratinocytes corresponds with the proliferative activation ofthe cells and is not related to the differentiation programme.
Nikoletta Nagy1,Zsuzsanna Bata-Csorg}o1,2,Norbert Kopasz
1, Csilla Szeg
1,
Andor Pivarcsi2, Andrea
Koreck1, Attila Dobozy
1,2,
Lajos Kemeny1,2
and MartaSzell
2
1Department of Dermatology and Allergology,University of Szeged, Szeged, Hungary;2Dermatological Research Group of theHungarian Academy of Sciences and theUniversity of Szeged, Szeged, Hungary
Key words: differentiation – dithranol – HaCaTkeratinocytes – keratinocyte growth factorreceptor – proliferation
Marta SzellDermatological Research Group of theHungarian Academy of Sciences and theUniversity of Szeged6720 SzegedKoranyi fasor 6HungaryTel.: 36 62 545 278Fax: 36 62 545 954e-mail: [email protected]
Accepted for publication 9 May 2006
Abbreviations: AcC, acetone-treated control; C, untreated control;DMEM, Dulbecco’s modified Eagle’s medium; FBS, fetal bovineserum; FGF, fibroblast growth factor; FGFR2-IIIb (KGFR), ker-atinocyte growth factor receptor; K1, keratin 1; K10, keratin 10;KGF, keratinocyte growth factor; Q-RT-PCR, quantitative real-time polymerase chain reaction; SDS, sodium dodecyl sulphate.
596
Experimental Dermatology 2006: 15: 596–605Blackwell Munksgaard . Printed in Singaporedoi: 10.1111/j.0906-6705.2006.00450.x
� 2006 Department of Dermatology and Allergology, University of Szeged.Journal compilation � 2006 Blackwell Munksgaard.
Experimental Dermatology
ISSN 0906-6705

important roles in a wide variety of biologicalprocesses such as control of proliferation/differ-entiation, re-epithelization in wound healing (2)or osteoblast differentiation in osteogenesis.In healthy epidermis, FGFR2-IIIb is mostly
distributed in the suprabasal layers and isexpressed at a very low level in basal kera-tinocytes (3,4). In contrast, both KGF andFGFR2-IIIb are upregulated in the hyperpro-liferative skin disease, psoriasis: both in thebasal [keratin 1 (K1)/keratin 10 (K10))] andsuprabasal (K1/K10+) layers of involved pso-riatic epidermis, FGFR2-IIIb mRNA levelswere approximately 2.5-fold higher comparedwith non-involved epidermis. In the subepider-mal fibroblasts of involved skin, elevated lev-els of KGF mRNA have also been reportedcompared with non-involved skin (5). How-ever, in this study the expression of FGFR2-IIIb in psoriatic involved and non-involvedepidermis was not compared with healthy epi-dermis.Although a great body of evidence describes
the tissue localization of FGFR2-IIIb, there areless data on the expression profile of this cellsurface receptor at different proliferation anddifferentiation stages of keratinocytes. A previ-ous study examining FGFR2-IIIb expression inhuman epithelial cells has demonstrated thatFGFR2-IIIb expression was characteristic ofthose keratinocytes that have already startedtheir differentiation programme (6). FGFR2-IIIb expression is also regulated due to highdose of calcium, which promotes keratinocytesdifferentiation (7) and due to confluence-induced spontaneous differentiation (8). On theother hand, it has also been described that asensitizing agent, nickel-sulphate, is able toincrease FGFR2-IIIb expression without affect-ing keratinocyte differentiation (9) and thatantiproliferative pharmacological agents such asphorbol ester, interferon-c and ultraviolet Bsubstantially downregulated FGFR2-IIIbexpression (5,10).To clarify the above described contradictions,
we studied FGFR2-IIIb expression in differentproliferation and differentiation stages of kera-tinocytes. We used synchronized, immortalizedHaCaT cell cultures, because it is a suitablemodel system to study the in vitro events ofkeratinocyte proliferation and differentiation(11). We also investigated the effects of theantipsoriatic drug, dithranol (12,13), as well asthe effects of different cell culture surfaces onthe FGFR2-IIIb expression in HaCaT keratino-cytes.
Materials and methods
Cell culture, synchronization and treatments ofHaCaT keratinocytes
Human HaCaT keratinocytes, kindly provided by Dr N. E.Fusenig (Heidelberg, Germany), were cultured and syn-chronized as described in details in a previous publication(12). Briefly, HaCaT keratinocytes after reaching a 100%confluence were cultured for 5 days in Dulbecco’s modifiedEagle’s medium (DMEM) containing 10% fetal bovineserum (FBS), then the medium was replaced by a serum-freemedium. The cells were grown in a serum-free medium for1 week. The synchronized cells were trypsinized and wereseeded into 75-cm2 culture flasks at a density of 5 · 103
cells/cm2 in high-glucose DMEM containing 10% FBS.Samples for quantitative real-time polymerase chain reaction(Q-RT-PCR) and Western blot analysis were collected 12,24, 36, 48, 72, 96 and 168 h after the end of synchroniza-tion using TRIzol reagent and protein lysis buffer, respect-ively.
Highly proliferating subconfluent HaCaT keratinocytes(cultured in high-glucose DMEM containing 10% FBS) werewashed with PBS under sterile conditions and were incubatedfor 30 min at 37�C with increasing concentrations of dithra-nol (0.1, 0.25 and 0.5 lg/ml) in DMEM, containing 0.5%FBS. Dithranol was always freshly dissolved in acetone (Ac)and used immediately. Control cells were treated with the sol-vent (AcC) only, or left untreated (C). After dithranol treat-ment, cells were washed with PBS twice and 10% FBScontaining high-glucose DMEM was re-added to them.HaCaT keratinocytes were harvested 4 h after dithranol treat-ment, either by TRIzol or protein lysis buffer.
To see whether cell attachment was necessary for HaCaTkeratinocytes to express FGFR2-IIIb, subconfluent HaCaTkeratinocytes cultured in high-glucose and 10% FBS-contain-ing DMEM were passaged onto 9 cm Ø bacterial plates(Matraplast, Budapest, Hungary) and tissue culture plates(Sarstedt, Nunbrecht, Germany). HaCaT keratinocytes weregrown for 96 h under these different attachment conditionsthen harvested at 24, 48, 72 and 96 h after plating. For har-vest, either TRIzol reagent or protein lysis buffer was used.The cell cultures were maintained in a humidified atmospherecontaining 5% CO2.
Reverse transcription and Q-RT-PCR
Total RNA was isolated from the cell cultures by TRIzolreagent, following the instructions of the manual. cDNAwas generated with oligo(dT) and random hexamer primersfrom 1 lg of RNA using the iScriptTM cDNA Synthesis Kitof Bio-Rad Laboratories (Hercules, CA, USA) in a final vol-ume of 20 ll. After reverse transcription, Q-RT-PCR wasperformed to quantify the abundance of FGFR2-IIIbmRNA. Besides FGFR2-IIIb, the differentiation marker,K10 mRNA was also measured. RNA expression data werenormalized to the 18S ribosomal RNA expression data ofeach examined samples. The primer sets and TaqMan probesused for the reactions are listed in Fig. 1(a). TaqMan probeswere purchased from IDT Probes (Chicago, IL, USA). TheQ-RT-PCR reactions were performed using iQ Supermix(Bio-Rad Laboratories) in an iCycler (Bio-Rad Laborator-ies). All RT-PCRs were carried out using three independentcell culture experiments and are presented as mean ± SE inthe figures.
Western blot analysis
Total protein extracts were prepared from synchronizedHaCaT keratinocytes at 12, 24, 36, 48, 72, 96 and 168 h
The expression of keratinocyte growth factor receptor
597

after the end of the synchronization process, from dithra-nol-treated HaCaT keratinocytes 4 h after the treatment andfrom HaCaT keratinocytes cultured for 24, 48, 72 and 96 hon different culture surfaces. The composition of proteinlysis buffer used for all harvest was as follows: 1.5%sodium dodecyl sulphate (SDS), 62.5 mM Tris–HCl (pH6.8), 5 mM ethylenediaminetetraacetic acid, 5% 2-mercapto-ethanol, 1 lg/ml antipain, 1 lg/ml chymostatin and 1 lg/mlleupeptin (all chemicals were obtained from Sigma-Aldrich,St Louis, MO, USA). Lysates were precleared by centrifuga-tion and supernatants were stored at )20�C. In order toverify the equivalent loading of proteins in the wells, thefollowing procedure was carried out: based on the OD280
measured, density and protein concentration of each sam-ples were calculated, and the samples were then run on10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE).The gels were stained by Coomassie brilliant blue (CBB;Sigma-Aldrich), dried, scanned and all of the loaded laneswere analysed by densitometry. The amounts of the loadedprotein samples were further corrected and checked againon SDS-PAGE. For the Western blot analysis, equalamounts of proteins were run on SDS-PAGE, and thentransferred to nitrocellulose membrane (Bio-Rad Laborator-ies). Membranes were blocked by incubation in Tris-bufferedsaline (150 mM NaCl, 25 mM Tris, pH 7.4) containing0.05% Tween 20 (Sigma-Aldrich) and 3% non-fat dry milk(Fluka Chemie AG, Neu-Buchs, Switzerland) for 2 h atroom temperature and subsequently incubated overnight at4�C with rabbit antiserum to the carboxyl terminus ofFGFR2-IIIb receptor (BEK C-17: sc-122, Santa Cruz Bio-technology, Inc., Santa Cruz, CA, USA) in a dilution of1:100. Besides FGFR2-IIIb, a differentiation marker, K10protein was also detected by a mouse monoclonal antibody(Dako A/S, Glostrup, Denmark) applied in a dilution of1:500. Alkaline phosphatase-conjugated goat anti-rabbit IgGand anti-mouse IgG (Sigma-Aldrich) were used as secondaryantibodies at 1:4000 dilution in the blocking buffer for 2 hat room temperature. The blots were developed by using5-bromo-4-chloro-3-indolyl phosphate/nitroblue tetrazoliumas a substrate (BCIP/NBT; Sigma-Aldrich).
All of our Western blots were carried out using three inde-pendent sets of protein samples. In the figures we present oneof them, for densitometry and for the calculation of FGFR2-IIIb and K10 expression rate we used all three blots.
Statistical analysis
The statistical significances of differences among samples ofsynchronized HaCaT cells collected at different time pointsand dithranol-treated HaCaT cells were assessed by varianceanalysis: one-way ANOVA for independent or correlatedsamples. In case of HaCaT cells cultured on tissue cultureand bacterial plates, statistical significances of differences weredetermined by procedures applicable to ordinal data: Mann–Whitney U-test, Student’s t-test and two-way factorialANOVA for independent samples. Similarity between expres-sion profiles of different genes was calculated by basic linearcorrelation test. All statistical analyses were carried out usingVassarStats (http://faculty.vassar.edu/lowry/VassarStats.html).
Results
High-level FGFR2-IIIb expression correlateswith the high proliferative rate of synchronizedHaCaT keratinocytes
In order to receive detailed information aboutthe relationship between the proliferation/differ-entiation stage of HaCaT keratinocytes andtheir FGFR2-IIIb expression, cells were syn-chronized and FGFR2-IIIb mRNA and proteinlevel changes were studied for 1 week afterrelease from cell quiescence. We have previouslydemonstrated in this model (11) that HaCaTcells become highly proliferative after releasefrom cell quiescence (72–96 h), they express a5integrin and D1 cyclin, at the same time K1and K10 expression are suppressed. By the endof the 1-week culture period (168 h) keratinexpression re-occurs in the cells parallel withsuppression of a5 integrin and D1 cyclin expres-sion and with the substantial reduction of pro-liferation.
FGFR2-IIIb forward primer 5’ -ACTCGGGGATAAATAGTTCCAA-3’
FGFR2-IIIb reverse primer 5’ -CCTTACATATATATTCCCCAGCAT-3’
FGFR2-IIIb probe 5’ -56-FAM-ACATTGAACAGAGCCAGCACTT-BHQ-3’
K10 primers and probe Applied Biosystems, TaqMan® Gene Expression Assays, Hs00166289_ml
18S forward primer 5’ -CGGCTACCACATCCAAGGAA-3’
18S reverse primer 5’ -GCTGGAATTACCGCGGCT-3’
18S probe
(a)
(b)
FGFR-2 gene
FGFR2-IIIb
FGFR2-IIIb F
exon 8 exon 9 exon 10 exon 11
exon 8 exon 9
exon 8 exon 10
FGFR2-IIIc F
FGFR2-IIIc R
FGFR2-IIIb R
FGFR2-IIIc
5’ -TexasRed-TGCTGGCACCAGACTTGCCCTC-BHQ-3’
Figure 1. Sequences of primers and probes used for real-time RT-PCR experiments. (a) After reverse transcription, quantitativereal-time polymerase chain reactions (Q-RT-PCR) were performed to quantify the abundance of FGFR2-IIIb (KGFR) and K10mRNAs. mRNA expression data were normalized to the 18S ribosomal RNA expression data of each examined sample. (b)Primers were designed to detect exclusively the alternatively spliced FGFR2-IIIb variant of FGFR2 primary gene product. BesideFGFR2-IIIb, there is another alternatively spliced variant of FGFR2 gene, the bacterially expressed kinase (FGFR2-IIIc ¼BEK).
Nagy et al.
598
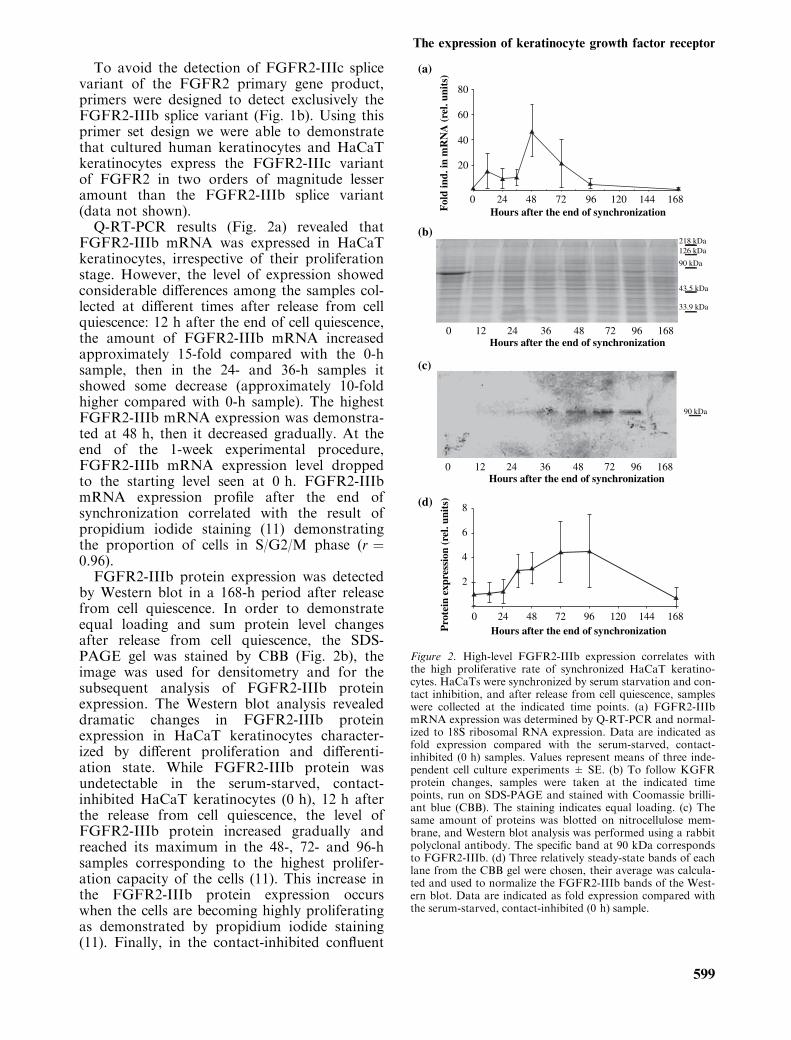
To avoid the detection of FGFR2-IIIc splicevariant of the FGFR2 primary gene product,primers were designed to detect exclusively theFGFR2-IIIb splice variant (Fig. 1b). Using thisprimer set design we were able to demonstratethat cultured human keratinocytes and HaCaTkeratinocytes express the FGFR2-IIIc variantof FGFR2 in two orders of magnitude lesseramount than the FGFR2-IIIb splice variant(data not shown).Q-RT-PCR results (Fig. 2a) revealed that
FGFR2-IIIb mRNA was expressed in HaCaTkeratinocytes, irrespective of their proliferationstage. However, the level of expression showedconsiderable differences among the samples col-lected at different times after release from cellquiescence: 12 h after the end of cell quiescence,the amount of FGFR2-IIIb mRNA increasedapproximately 15-fold compared with the 0-hsample, then in the 24- and 36-h samples itshowed some decrease (approximately 10-foldhigher compared with 0-h sample). The highestFGFR2-IIIb mRNA expression was demonstra-ted at 48 h, then it decreased gradually. At theend of the 1-week experimental procedure,FGFR2-IIIb mRNA expression level droppedto the starting level seen at 0 h. FGFR2-IIIbmRNA expression profile after the end ofsynchronization correlated with the result ofpropidium iodide staining (11) demonstratingthe proportion of cells in S/G2/M phase (r ¼0.96).FGFR2-IIIb protein expression was detected
by Western blot in a 168-h period after releasefrom cell quiescence. In order to demonstrateequal loading and sum protein level changesafter release from cell quiescence, the SDS-PAGE gel was stained by CBB (Fig. 2b), theimage was used for densitometry and for thesubsequent analysis of FGFR2-IIIb proteinexpression. The Western blot analysis revealeddramatic changes in FGFR2-IIIb proteinexpression in HaCaT keratinocytes character-ized by different proliferation and differenti-ation state. While FGFR2-IIIb protein wasundetectable in the serum-starved, contact-inhibited HaCaT keratinocytes (0 h), 12 h afterthe release from cell quiescence, the level ofFGFR2-IIIb protein increased gradually andreached its maximum in the 48-, 72- and 96-hsamples corresponding to the highest prolifer-ation capacity of the cells (11). This increase inthe FGFR2-IIIb protein expression occurswhen the cells are becoming highly proliferatingas demonstrated by propidium iodide staining(11). Finally, in the contact-inhibited confluent
0
20
40
60
80
24 48 72 96 120 144 168
0 2412 4836 72 96 168
90 kDa
218 kDa126 kDa
90 kDa
43.5 kDa
33.9 kDa
Hours after the end of synchronization
(a)
(b)
(c)
(d)
Hours after the end of synchronization
0 2412 4836 72 96 168Hours after the end of synchronization
0 24 48 72 96 168120 144Hours after the end of synchronization
Fol
d in
d. in
mR
NA
(re
l. un
its)
2
4
6
8
Pro
tein
exp
ress
ion
(rel
. uni
ts)
Figure 2. High-level FGFR2-IIIb expression correlates withthe high proliferative rate of synchronized HaCaT keratino-cytes. HaCaTs were synchronized by serum starvation and con-tact inhibition, and after release from cell quiescence, sampleswere collected at the indicated time points. (a) FGFR2-IIIbmRNA expression was determined by Q-RT-PCR and normal-ized to 18S ribosomal RNA expression. Data are indicated asfold expression compared with the serum-starved, contact-inhibited (0 h) samples. Values represent means of three inde-pendent cell culture experiments ± SE. (b) To follow KGFRprotein changes, samples were taken at the indicated timepoints, run on SDS-PAGE and stained with Coomassie brilli-ant blue (CBB). The staining indicates equal loading. (c) Thesame amount of proteins was blotted on nitrocellulose mem-brane, and Western blot analysis was performed using a rabbitpolyclonal antibody. The specific band at 90 kDa correspondsto FGFR2-IIIb. (d) Three relatively steady-state bands of eachlane from the CBB gel were chosen, their average was calcula-ted and used to normalize the FGFR2-IIIb bands of the West-ern blot. Data are indicated as fold expression compared withthe serum-starved, contact-inhibited (0 h) sample.
The expression of keratinocyte growth factor receptor
599
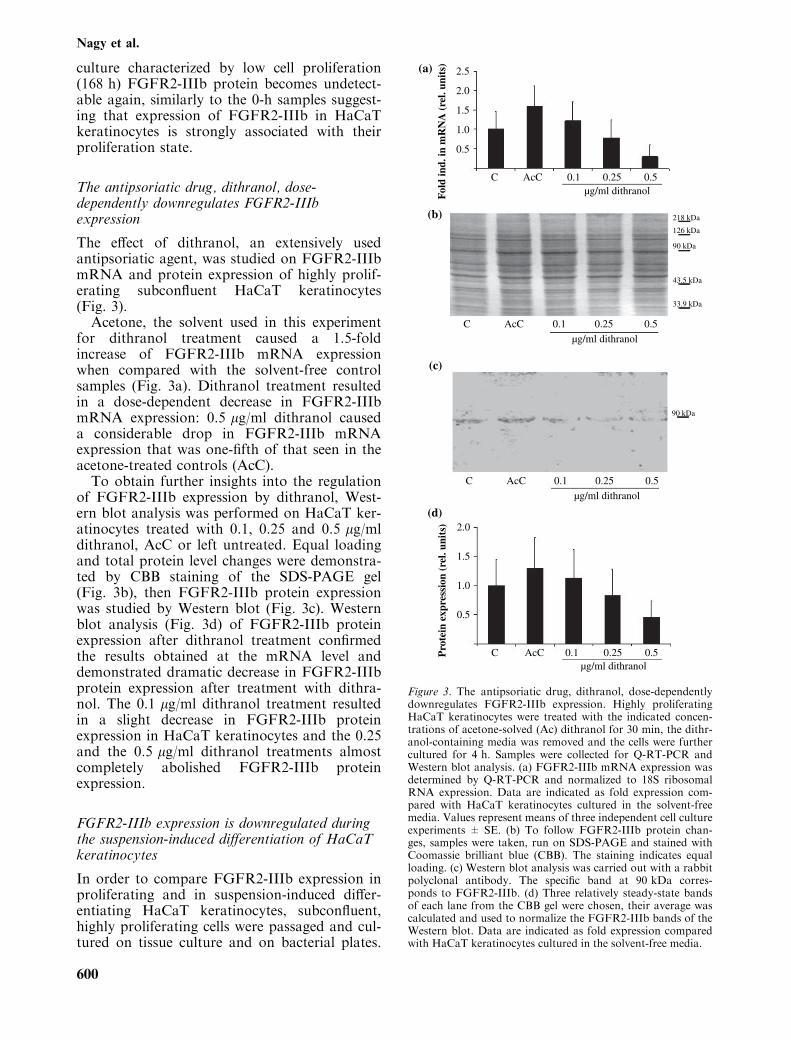
culture characterized by low cell proliferation(168 h) FGFR2-IIIb protein becomes undetect-able again, similarly to the 0-h samples suggest-ing that expression of FGFR2-IIIb in HaCaTkeratinocytes is strongly associated with theirproliferation state.
The antipsoriatic drug, dithranol, dose-dependently downregulates FGFR2-IIIbexpression
The effect of dithranol, an extensively usedantipsoriatic agent, was studied on FGFR2-IIIbmRNA and protein expression of highly prolif-erating subconfluent HaCaT keratinocytes(Fig. 3).Acetone, the solvent used in this experiment
for dithranol treatment caused a 1.5-foldincrease of FGFR2-IIIb mRNA expressionwhen compared with the solvent-free controlsamples (Fig. 3a). Dithranol treatment resultedin a dose-dependent decrease in FGFR2-IIIbmRNA expression: 0.5 lg/ml dithranol causeda considerable drop in FGFR2-IIIb mRNAexpression that was one-fifth of that seen in theacetone-treated controls (AcC).To obtain further insights into the regulation
of FGFR2-IIIb expression by dithranol, West-ern blot analysis was performed on HaCaT ker-atinocytes treated with 0.1, 0.25 and 0.5 lg/mldithranol, AcC or left untreated. Equal loadingand total protein level changes were demonstra-ted by CBB staining of the SDS-PAGE gel(Fig. 3b), then FGFR2-IIIb protein expressionwas studied by Western blot (Fig. 3c). Westernblot analysis (Fig. 3d) of FGFR2-IIIb proteinexpression after dithranol treatment confirmedthe results obtained at the mRNA level anddemonstrated dramatic decrease in FGFR2-IIIbprotein expression after treatment with dithra-nol. The 0.1 lg/ml dithranol treatment resultedin a slight decrease in FGFR2-IIIb proteinexpression in HaCaT keratinocytes and the 0.25and the 0.5 lg/ml dithranol treatments almostcompletely abolished FGFR2-IIIb proteinexpression.
FGFR2-IIIb expression is downregulated duringthe suspension-induced differentiation of HaCaTkeratinocytes
In order to compare FGFR2-IIIb expression inproliferating and in suspension-induced differ-entiating HaCaT keratinocytes, subconfluent,highly proliferating cells were passaged and cul-tured on tissue culture and on bacterial plates.
C
2.0
1.5
1.0
Pro
tein
exp
ress
ion
(rel
. uni
ts)
0.5
2.0
2.5(a)
(b)
(c)
(d)
1.5
1.0
Fol
d in
d. in
mR
NA
(re
l. un
its)
0.5
AcC 0.1µg/ml dithranol
0.25 0.5
C AcC 0.1
µg/ml dithranol
0.25 0.5
C AcC 0.1
µg/ml dithranol
0.25 0.5
C AcC 0.1µg/ml dithranol
0.25 0.5
90 kDa
218 kDa
126 kDa
90 kDa
43.5 kDa
33.9 kDa
Figure 3. The antipsoriatic drug, dithranol, dose-dependentlydownregulates FGFR2-IIIb expression. Highly proliferatingHaCaT keratinocytes were treated with the indicated concen-trations of acetone-solved (Ac) dithranol for 30 min, the dithr-anol-containing media was removed and the cells were furthercultured for 4 h. Samples were collected for Q-RT-PCR andWestern blot analysis. (a) FGFR2-IIIb mRNA expression wasdetermined by Q-RT-PCR and normalized to 18S ribosomalRNA expression. Data are indicated as fold expression com-pared with HaCaT keratinocytes cultured in the solvent-freemedia. Values represent means of three independent cell cultureexperiments ± SE. (b) To follow FGFR2-IIIb protein chan-ges, samples were taken, run on SDS-PAGE and stained withCoomassie brilliant blue (CBB). The staining indicates equalloading. (c) Western blot analysis was carried out with a rabbitpolyclonal antibody. The specific band at 90 kDa corres-ponds to FGFR2-IIIb. (d) Three relatively steady-state bandsof each lane from the CBB gel were chosen, their average wascalculated and used to normalize the FGFR2-IIIb bands of theWestern blot. Data are indicated as fold expression comparedwith HaCaT keratinocytes cultured in the solvent-free media.
Nagy et al.
600
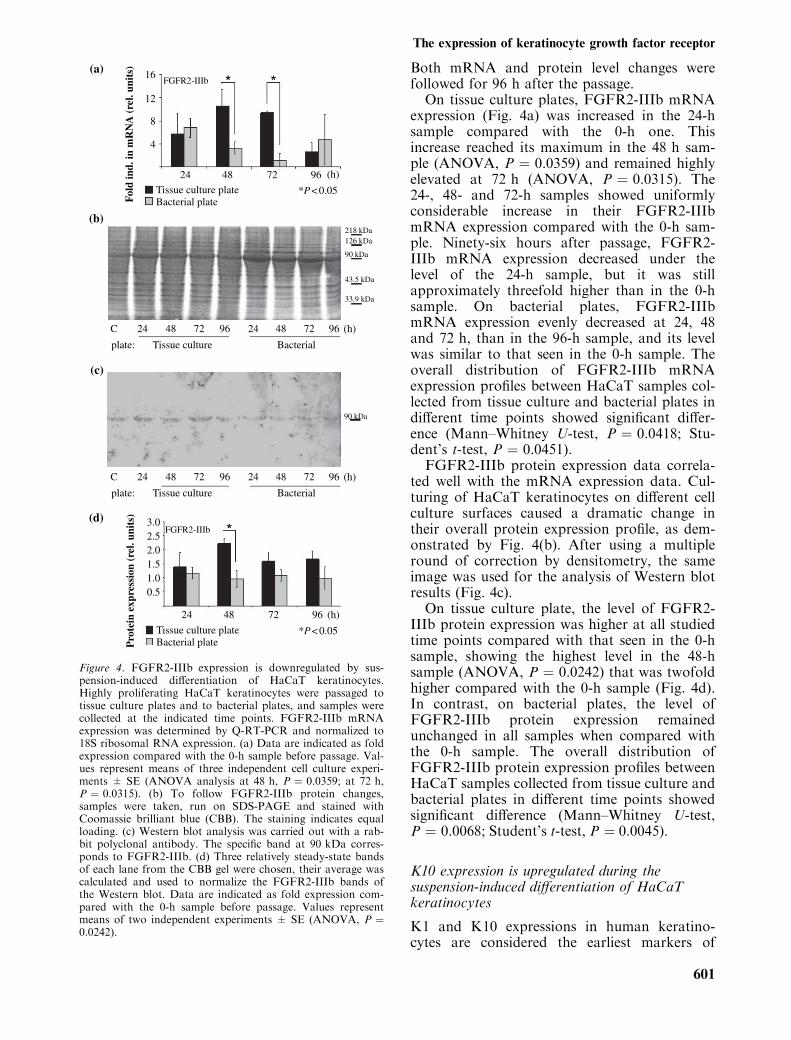
Both mRNA and protein level changes werefollowed for 96 h after the passage.
On tissue culture plates, FGFR2-IIIb mRNAexpression (Fig. 4a) was increased in the 24-hsample compared with the 0-h one. Thisincrease reached its maximum in the 48 h sam-ple (ANOVA, P ¼ 0.0359) and remained highlyelevated at 72 h (ANOVA, P ¼ 0.0315). The24-, 48- and 72-h samples showed uniformlyconsiderable increase in their FGFR2-IIIbmRNA expression compared with the 0-h sam-ple. Ninety-six hours after passage, FGFR2-IIIb mRNA expression decreased under thelevel of the 24-h sample, but it was stillapproximately threefold higher than in the 0-hsample. On bacterial plates, FGFR2-IIIbmRNA expression evenly decreased at 24, 48and 72 h, than in the 96-h sample, and its levelwas similar to that seen in the 0-h sample. Theoverall distribution of FGFR2-IIIb mRNAexpression profiles between HaCaT samples col-lected from tissue culture and bacterial plates indifferent time points showed significant differ-ence (Mann–Whitney U-test, P ¼ 0.0418; Stu-dent’s t-test, P ¼ 0.0451).
FGFR2-IIIb protein expression data correla-ted well with the mRNA expression data. Cul-turing of HaCaT keratinocytes on different cellculture surfaces caused a dramatic change intheir overall protein expression profile, as dem-onstrated by Fig. 4(b). After using a multipleround of correction by densitometry, the sameimage was used for the analysis of Western blotresults (Fig. 4c).
On tissue culture plate, the level of FGFR2-IIIb protein expression was higher at all studiedtime points compared with that seen in the 0-hsample, showing the highest level in the 48-hsample (ANOVA, P ¼ 0.0242) that was twofoldhigher compared with the 0-h sample (Fig. 4d).In contrast, on bacterial plates, the level ofFGFR2-IIIb protein expression remainedunchanged in all samples when compared withthe 0-h sample. The overall distribution ofFGFR2-IIIb protein expression profiles betweenHaCaT samples collected from tissue culture andbacterial plates in different time points showedsignificant difference (Mann–Whitney U-test,P ¼ 0.0068; Student’s t-test, P ¼ 0.0045).
K10 expression is upregulated during thesuspension-induced differentiation of HaCaTkeratinocytes
K1 and K10 expressions in human keratino-cytes are considered the earliest markers of
24
FGFR2-IIIb
FGFR2-IIIb
*
* *
Pro
tein
exp
ress
ion
(rel
. uni
ts)
Tissue culture plate
0.51.01.52.02.53.0
*P < 0.05Bacterial plate
48 72 96 (h)
24
Tissue culture plate
Fol
d in
d. in
mR
NA
(re
l. un
its)
4
8
12
16
*P < 0.05Bacterial plate
48 72 96 (h)
24C
plate: Tissue culture Bacterial
48 72 96 24 48 72 96 (h)
24C
plate: Tissue culture Bacterial
48 72 96 24 48 72 96 (h)
90 kDa
218 kDa126 kDa
90 kDa
43.5 kDa
33.9 kDa
(a)
(b)
(c)
(d)
Figure 4. FGFR2-IIIb expression is downregulated by sus-pension-induced differentiation of HaCaT keratinocytes.Highly proliferating HaCaT keratinocytes were passaged totissue culture plates and to bacterial plates, and samples werecollected at the indicated time points. FGFR2-IIIb mRNAexpression was determined by Q-RT-PCR and normalized to18S ribosomal RNA expression. (a) Data are indicated as foldexpression compared with the 0-h sample before passage. Val-ues represent means of three independent cell culture experi-ments ± SE (ANOVA analysis at 48 h, P ¼ 0.0359; at 72 h,P ¼ 0.0315). (b) To follow FGFR2-IIIb protein changes,samples were taken, run on SDS-PAGE and stained withCoomassie brilliant blue (CBB). The staining indicates equalloading. (c) Western blot analysis was carried out with a rab-bit polyclonal antibody. The specific band at 90 kDa corres-ponds to FGFR2-IIIb. (d) Three relatively steady-state bandsof each lane from the CBB gel were chosen, their average wascalculated and used to normalize the FGFR2-IIIb bands ofthe Western blot. Data are indicated as fold expression com-pared with the 0-h sample before passage. Values representmeans of two independent experiments ± SE (ANOVA, P ¼0.0242).
The expression of keratinocyte growth factor receptor
601

terminal differentiation (14). In order to inter-pret our results on FGFR2-IIIb mRNA andprotein expression during the suspension-induced differentiation of HaCaT keratinocytes,K10 gene and protein expression was also fol-lowed during this process.On tissue culture plates, K10 mRNA expres-
sion (Fig. 5a) was undetectable in the 24-, 48-,72- and 96-h samples after passage. In contrastwith this, on bacterial plate, the level of K10mRNA expression showed a significant, rapidupregulation (ANOVA, P ¼ 0.0003) thatreached its maximum in the 96-h sample. Theoverall distribution of K10 mRNA expressionprofiles between HaCaT samples collected fromtissue culture and bacterial plates in differenttime points showed a significant difference(Mann–Whitney U-test, P ¼ 0.0019; Student’st-test, P ¼ 0.0036).K10 protein changes correlated well with the
K10 mRNA changes: on tissue culture plate,K10 protein expression dropped approximatelyhalf of that seen in the 0-h sample than remainedat this level. The presence of this remaining pro-tein is explained by the high stability and lowturnover of K10 observed previously (11). Incontrast, HaCaT keratinocytes seeded onto bac-terial plates expressed gradually, significantlyincreasing amounts of K10 proteins at 48, 72and 96 h (ANOVA, P ¼ 0.0010), indicating theonward process of differentiation induced byculturing the HaCaT keratinocytes in suspension(Fig. 5b,c). The overall distribution of K10 pro-tein expression profiles between HaCaT samplescollected from tissue culture and bacterial platesin different time points showed significant differ-ence (Mann–Whitney U-test, P ¼ 0.0019; Stu-dent’s t-test, P ¼ 0.0007).
Discussion
Keratinocyte growth factor has been shown tobe a potent mitogen for a wide range of epithe-lial cells including human keratinocytes (15). Itacts through its tyrosine kinase receptor,FGFR2-IIIb. Some previous studies examiningFGFR2-IIIb expression in human epithelial cellshave concluded that FGFR2-IIIb expression isupregulated with differentiation, which is indica-ted by K1 expression in FGFR2-IIIb expressingcells (6–8). At the same time, there are evidencessuggesting that a high-level FGFR2-IIIb expres-sion is rather the characteristic of actively prolif-erating cells (7,9,10). In the present study, weaimed to clarify this contradiction by using theimmortalized keratinocyte cell line, HaCaT.
24
K10 ** **
Pro
tein
exp
ress
ion
(rel
. uni
ts)
Tissue culture plate
0.5
1.0
1.5
2.0
2.5
**P < 0.01Bacterial plate
48 72 96 (h)
24C
plate: Tissue culture Bacterial
48 72 96 24 48 72 96 (h)
24C
plate: Tissue culture Bacterial
48 72 96 24 48 72 96 (h)
126 kDa
218 kDa126 kDa
90 kDa
43.5 kDa
33.9 kDa
24Tissue culture plate ***P < 0.001Bacterial plate
48 72 96 (h)
Fol
d in
d. in
mR
NA
(re
l. un
its)
4
8
12
16K10 *** *** ***(a)
(b)
(c)
(d)
Figure 5. K10 expression is upregulated by suspension-induced differentiation of HaCaT keratinocytes. Highly prolif-erating HaCaT keratinocytes were passaged to tissue cultureplates and to bacterial plates, and samples were collected atthe indicated time points. (a) K10 mRNA expression wasdetermined by Q-RT-PCR and normalized to 18S ribosomalRNA expression. Data are indicated as fold expression com-pared with the 0-h sample before passage. Values representmeans of three independent cell culture experiments ± SE(ANOVA, P ¼ 0.0003). (b) To follow K10 protein changes,samples were taken, run on SDS-PAGE and stained withCoomassie brilliant blue (CBB). The staining indicates equalloading. (c) Western blot analysis was carried out with amouse monoclonal antibody. The specific band at 126 kDacorresponds to K10. (d) Three relatively steady-state bands ofeach lane from the CBB gel were chosen, their average wascalculated and used to normalize the K10 bands of the West-ern blot. Data are indicated as fold expression compared withthe 0-h sample before passage. Values represent means of twoindependent experiments ± SE (ANOVA, P ¼ 0.001).
Nagy et al.
602

Here, we provide data suggesting that inHaCaT keratinocytes FGFR2-IIIb expressioncorrelates with the proliferation state of thecells regardless of their differentiation state.When HaCaT keratinocytes are forced into cellquiescence and then released from that state,they almost synchronously enter a highly prolif-erative phase, while K10 and K1 mRNAbecome undetectable (11). Parallel with theiractivation, FGFR2-IIIb protein expression isconsiderably induced in the cells. FGFR2-IIIbprotein was undetectable in confluent cultureswhen cell proliferation was inhibited, both inK1+ (0 h) and K1) (168 h) cells.Quantitative RT-PCR results revealed that
FGFR2-IIIb mRNA was expressed in HaCaTkeratinocytes irrespective of their proliferation/differentiation stages. However, the level ofexpression showed considerable differences atdifferent time points after release from cellquiescence. We saw two peaks in FGFR2-IIIbmRNA at 12 and 48 h after the end of syn-chronization. Thus, we hypothesize that therecovery from a stressed situation such as con-tact inhibition and serum starvation in ourexperiment might be coupled with dramaticre-arrangements in the transcriptional machin-ery of the HaCaT keratinocytes leading todrastic transient changes in the expression ofcertain genes. Later, parallel with the cellsentering S/G2/M phase, demonstrated withpropidium iodide staining (12), the level ofFGFR2-IIIb mRNA expression increasesgradually and reaches its maximum at 48 h(r ¼ 0.96).Changes in the FGFR2-IIIb protein amounts
after release from cell quiescence show a some-what similar pattern to the FGFR2-IIIb mRNAbut some differences also occur. Parallel withthe mRNA level, the amount of FGFR2-IIIbprotein increases gradually after 0 h andreaches its maximum in the samples, when theculture of HaCaT keratinocytes is in a pre-con-fluent stage and the cells are proliferating veryactively. In contrast, in the serum-starved, con-tact-inhibited cells and in the 168-h, confluentHaCaT keratinocytes there is no detectableamount of FGFR2-IIIb protein. Comparison ofthe mRNA and protein expression patterns ofFGFR2-IIIb in this synchronized HaCaT modelsystem suggest that the expression of FGFR2-IIIb is regulated both at transcriptional andpost-transcriptional levels.Beside the well-characterized model system of
synchronized HaCaT keratinocytes, we studiedthe connections of FGFR2-IIIb expression and
cell proliferation/differentiation in in vitroexperiments where proliferation of HaCaT ker-atinocytes was inhibited by an antipsoriaticagent, dithranol and by culturing them in sus-pension.
Dithranol is an extensively used agent for thetreatment of psoriasis. It has been well estab-lished that it exerts its antipsoriatic action viainhibition of the hyperproliferation of psoriatickeratinocytes (16,17). Here we demonstrate thatdithranol downregulates the expression ofFGFR2-IIIb in HaCaT keratinocytes, suggest-ing that this inhibitory effect could be a compo-nent of the antipsoriatic property of this agent.Downregulation of FGFR2-IIIb by dithranolfits into the general view about the possibleeffect of this agent in psoriasis. Several reportssuggest that cell surface receptors that are posit-ive regulators of keratinocyte proliferation andinflammatory processes are downregulated bydithranol (13,18,19), while the cell surfacereceptor of the anti-inflammatory IL-10 whichis downregulated in psoriatic skin is upregulat-ed by dithranol (20). The fact that the dithra-nol-induced inhibition of proliferation iscoupled with the downregulation of FGFR2-IIIb expression further supports the hypothesisthat FGFR2-IIIb expression is rather the char-acteristic of proliferating than differentiatingkeratinocytes.
The effects of different cell culture surfaces,which either induced proliferation or differenti-ation of HaCaT keratinocytes also demonstra-ted that FGFR2-IIIb expression was related toproliferation and not to differentiation. Here wehave demonstrated that during the 96-h cultureperiod, FGFR2-IIIb expression was signifi-cantly upregulated only in HaCaT keratinocytescultured on tissue culture plate, where cells wereable to adhere to the surface and to proliferate.In contrast, on bacterial plates, cells wereunable to adhere to the surface, thus their dif-ferentiation was induced as in suspension cul-tures (21). It is also known from the literaturethat HaCaT cells cultured on bacterial platesundergo anoikis. The ratio of the survivingHaCaT cells depends on the ratio of the prolif-erating population before cells were plated onbacterial plates (22). Thus our results were cor-rected with the ratio of the surviving cells,which was 40–60% (23).
Under these culture conditions FGFR2-IIIband K10 expression profiles showed anti-parallel patterns: while FGFR2-IIIb expres-sion remained around the starting level, K10expression was upregulated (r ¼ )0.092). The
The expression of keratinocyte growth factor receptor
603

opposing tendency of the regulation of FGFR2-IIIb and K10 in suspension-cultured HaCaTkeratinocytes was pronounced when the expres-sion profiles were compared: the expression ofFGFR2-IIIb completely diminished by the endof the 24-h period of suspension culture andstayed undetectable for the entire 96-h experi-mental procedure, while the expression of K10showed an evenly increasing pattern during thisperiod. The seemingly contradictory results inFGFR2-IIIb mRNA and protein expressioncould be explained by the possible slow turn-over of FGFR2-IIIb protein (24).To date, there are only in situ hybridization
results on the comparison of FGFR2-IIIbmRNA expression in psoriatic involved andhealthy epidermis demonstrating an upregulatedFGFR2-IIIb mRNA expression in psoriasis (5).Recently, Kovacs et al. have demonstrated theoverexpression of FGF-7 and FGF-10 in psori-atic lesional skin by immunohistochemistry(25). The result of our Q-RT-PCR analysisrevealed increased FGFR2-IIIb mRNA expres-sion not only in psoriatic involved (2.65-foldinduction), but also in psoriatic uninvolved(3.08-fold induction) epidermis compared withhealthy epidermis (data not shown). The factthat we detected elevated levels of FGFR2-IIIbexpression both in involved and non-involvedepidermis of psoriatic patients supports our pre-vious findings that keratinocytes of the unin-volved epidermis are in an activated stage,compared with the keratinocytes of healthy epi-dermis. This activated state is associated withelevated levels of a5 integrin expression (26)with an accelerated spreading and focal adhe-sion kinase responsiveness to fibronectin (27)and with an enhanced ability to produceEDA+ fibronectin (28). The elevated FGFR2-IIIb mRNA expression may also contribute tothe pre-activated state of uninvolved psoriatickeratinocytes.Based on the results of our experiments we
conclude that FGFR2-IIIb expression inHaCaT keratinocytes corresponds with the pro-liferative activation of the cells and is not rela-ted to the differentiation programme (K10expression). The fact that in normal epidermisFGFR2-IIIb is expressed mainly on suprabasalK10+ keratinocytes does not exclude the possi-bility that the same applies for normal keratino-cytes, as it has been shown that the activelyproliferating keratinocyte population in normalepidermis is the K1/K10 expressing suprabasal-ly localized transiently amplifying keratinocytepopulation (29).
Acknowledgements
This work was supported by grants OTKA TS044826, OTKANI62007, OTKA 061541, GVOP-3.1.1-2004-05-0149/3.0 andNKFP1A/0012/2002. Marta Szell was supported by the Bol-yai Foundation of the Hungarian Academy of Sciences. Zsu-zsanna Bata-Csorg}o is a recipient of the Szechenyi IstvanFellowship of the Hungarian Ministry of Education.
References
1. Braun S, auf dem K U, Steiling H, Werner S. Fibro-blast growth factors in epithelial repair and cytopro-tection. Philos Trans R Soc Lond B Biol Sci 2004:359: 753–757.
2. Werner S, Smola H, Liao X et al. The function ofKGF in morphogenesis of epithelium and reepitheliali-zation of wounds. Science 1994: 266: 819–822.
3. LaRochelle W J, Dirsch O R, Finch P W et al. Speci-fic receptor detection by a functional keratinocytegrowth factor-immunoglobulin chimera. J Cell Biol1995: 129: 357–366.
4. Rio-Tsonis K, Jung J C, Chiu I M, Tsonis P A. Con-servation of fibroblast growth factor function in lensregeneration. Proc Natl Acad Sci U S A 1997: 94:13701–13706.
5. Finch P W, Murphy F, Cardinale I, Krueger J G.Altered expression of keratinocyte growth factor andits receptor in psoriasis. Am J Pathol 1997: 151: 1619–1628.
6. Capone A, Visco V, Belleudi F et al. Up-modulationof the expression of functional keratinocyte growthfactor receptors induced by high cell density in thehuman keratinocyte HaCaT cell line. Cell Growth Dif-fer 2000: 11: 607–614.
7. Marchese C, Sorice M, De Stefano C, Frati L, TorrisiM R. Modulation of keratinocyte growth factor recep-tor expression in human cultured keratinocytes. CellGrowth Differ 1997: 8: 989–997.
8. Visco V, Belleudi F, Marchese C et al. Differentialresponse to keratinocyte growth factor receptor andepidermal growth factor receptor ligands of prolifer-ating and differentiating intestinal epithelial cells.J Cell Physiol 2004: 200: 31–44.
9. Marchese C, Visco V, Aimati L et al. Nickel-inducedkeratinocyte proliferation and up-modulation of thekeratinocyte growth factor receptor expression. ExpDermatol 2003: 12: 497–505.
10. Zhou Y, Lee H S, Kooshesh F et al. Effects of UVBirradiation on keratinocyte growth factor (KGF) andreceptor (KGFR) expression in cultured human kera-tinocytes. Exp Dermatol 1996: 5: 138–144.
11. Pivarcsi A, Szell M, Kemeny L, Dobozy A, Bata-Csorgo Z. Serum factors regulate the expression of theproliferation-related genes alpha5 integrin and keratin1, but not keratin 10, in HaCaT keratinocytes. ArchDermatol Res 2001: 293: 206–213.
12. Kemeny L, Gross E, Arenberger P, Ruzicka T. Dithr-anol-induced down-regulation of 12(S)-hydroxyeicosa-tetraenoic acid [12(S)-HETE] receptors in a humanepidermal cell line. Arch Dermatol Res 1991: 283:333–336.
13. Kemeny L, Michel G, Arenberger P, Ruzicka T et al.Down-regulation of epidermal growth factor receptorsby dithranol. Acta Derm Venereol 1993: 73: 37–40.
14. Woodcock-Mitchell J, Eichner R, Nelson W G, Sun TT. Immunolocalization of keratin polypeptides in
Nagy et al.
604

human epidermis using monoclonal antibodies. J CellBiol 1982: 95: 580–588.
15. Marchese C, Rubin J, Ron D et al. Human keratino-cyte growth factor activity on proliferation and differ-entiation of human keratinocytes: differentiationresponse distinguishes KGF from EGF family. J CellPhysiol 1990: 144: 326–332.
16. Walter J F, Stoughton R B, DeQuoy P R. Suppres-sion of epidermal proliferation by ultraviolet light,coal tar and anthralin. Br J Dermatol 1978: 99: 89–96.
17. Pol A, Bergers M, Schalkwijk J. Comparison of anti-proliferative effects of experimental and establishedantipsoriatic drugs on human keratinocytes, using asimple 96-well-plate assay. In Vitro Cell Dev BiolAnim 2003: 39: 36–42.
18. Kemeny L, Ruzicka T, Braun-Falco O. Dithranol: areview of the mechanism of action in the treatment ofpsoriasis vulgaris. Skin Pharmacol 1990: 3: 1–20.
19. Kemeny L, Kenderessy A S, Olasz E et al. The inter-leukin-8 receptor: a potential target for antipsoriatictherapy? Eur J Pharmacol 1994: 258: 269–272.
20. Farkas A, Kemeny L, Szony B J et al. Dithranolupregulates IL-10 receptors on the cultured humankeratinocyte cell line HaCaT. Inflamm Res 2001: 50:44–49.
21. Hotchin N A, Kovach N L, Watt F M. Functionaldown-regulation of alpha 5 beta 1 integrin in keratino-cytes is reversible but commitment to terminal differ-entiation is not. J Cell Sci 1993: 106: 1131–1138.
22. Bretland A J, Lawry J, Sharrard R M. A study ofdeath by anoikis in cultured epithelial cells. Cell Prolif2001: 34: 199–210.
23. Frisch S M, Francis H. Disruption of epithelial cell-matrix interactions induces apoptosis. J Cell Biol1994: 124: 619–626.
24. Pedchenko V K, Imagawa W T. Mammogenic hor-mones differentially modulate keratinocyte growth fac-tor (KGF)-induced proliferation and KGF receptorexpression in cultured mouse mammary gland epithe-lium. Endocrinology 1998: 139: 2519–2526.
25. Kovacs D, Falchi M, Cardinali G et al. Immunhisto-chemical analysis of keratinocyte growth factor andfibroblast growth factor 10 expression in psoriasis.Exp Dermatol 2005: 14: 130–137.
26. Bata-Csorgo Z, Hammerberg C, Voorhees J J, CooperK D. Flow cytometric identification of proliferativesubpopulations within normal human epidermis andthe localization of the primary hyperproliferativepopulation in psoriasis. J Exp Med 1993: 178: 1271–1281.
27. Chen G, McCormick T S, Hammerberg C, Ryder-Diggs S, Stevens S R, Cooper K D. Basal keratino-cytes from uninvolved psoriatic skin exhibit acceleratedspreading and focal adhesion kinase responsiveness tofibronectin. J Invest Dermatol 2001: 117: 1538–1545.
28. Szell M, Bata-Csorgo Z, Koreck A et al. Proliferatingkeratinocytes are putative sources of the psoriasis sus-ceptibility-related EDA (extra domain A of fibronec-tin) oncofetal fibronectin. J Invest Dermatol 2004:123: 537–546.
29. Bata-Csorgo Z, Hammerberg C, Voorhees J J, CooperK D. Kinetics and regulation of human keratinocytestem cell growth in short-term primary ex vivo culture.Cooperative growth factors from psoriatic lesional Tlymphocytes stimulate proliferation among psoriaticuninvolved, but not normal, stem keratinocytes. J ClinInvest 1995: 95: 317–327.
The expression of keratinocyte growth factor receptor
605
















