
The essential oil qualitative and quantitative composition in the needles of Pinus sylvestris L....
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of The essential oil qualitative and quantitative composition in the needles of Pinus sylvestris L....

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The essential oil qualitative and quantitative composition in the needlesof Pinus sylvestris L. growing along industrial transects
Eugenija Kupcinskiene a,*, Aida Stikliene b, Asta Judzentiene c
a Department of Biology, Vytautas Magnus University, Vileikos 8, LT-44404 Kaunas, Lithuaniab Department of Ecology, Lithuanian University of Agriculture, Studentu 11, LT-53361 Kaunas, Akademija, Lithuania
c Institute of Chemistry, A. Gostauto 9, LT-01108 Vilnius, Lithuania
Received 1 February 2008; accepted 5 February 2008
Quantities of components of essential oil in the needles of Scots pine growing around twofactories are described in relation to pollution.
Abstract
The aim of this study was to evaluate composition of the essential oils in the needles of Pinus sylvestris growing in the areas affected by acement factory (CF), and an oil refinery (OR). Volatile components of the needles were analyzed by GC and GC/MS. The most heavily pollutedCF stand had significantly higher concentration of g-Terpinene, Caryophyllene oxide in the current-year needles, while higher concentration ofd-3-Carene, a-Terpinene, g-Terpinene and Terpinolene was documented for 1-year-old needles. The most heavily polluted OR stand hada significantly higher concentration of Sabinene þ b-Pinene, 1-epi-Cubenol in the current-year needles and a significantly higher concentrationof Camphene, Sabinene þ b-Pinene, Myrcene, a-Cadinene, 1-epi-Cubenol in the 1-year-old needles than the least polluted site. Along transectsan increase in the amount of some diterpenes and a decrease in the components of the shorter chain essential oils was observed. These effectscould be at least partially attributed to SO2.� 2008 Elsevier Ltd. All rights reserved.
Keywords: Terpenes; Phytoindication; Conifers; Secondary metabolites; Air pollution; Cement dust pollution; Sulphur dioxide
1. Introduction
In many developed countries atmospheric pollution de-creased in the 1980s (Emberson, 2003; Wieser et al., 2006)and in the Central and Eastern European countries in the1990s (Mankovska, 1996; Staszewski et al., 1998; Sopaus-kiene et al., 2001). Despite this, during the past two decadesthere has been a growing interest in air pollutionevegetationeffects (Bytnerowicz et al., 2006). Up-to-date field studies,monitoring, and modelling work document air pollution andclimate change impacts on forests in Europe and NorthAmerica (Manninen and Huttunen, 1995; Krupa and Legge,2000; Bytnerowicz et al., 2002; Manning and Godzik, 2004;
Huttunen 2005; Huttunen and Manninen, 2005; Paoletti,2005, 2006; Gunthardt-Goerg and Vollenweider, 2006; Allenet al., 2007; Grulke et al. 2007). It is particularly challengingto identify specific or unique indicators for stresses.
Recently greater attention is paid to the secondary metabo-lites including essential oils of various plant species (Law-rence, 1991; Shu and Lawrence, 1997; Barnola and Cedeno,2000; Lawrence, 2001, 2006). Terpenes in conifers are signif-icant chemomarkers of environmental impact (Supuka andBerta, 1998). Diurnal (Barnola et al., 1997), seasonal andannual (Nault, 2003), climatic and edaphic (Kainulainenet al., 1992; Barnola et al., 1997), geographicalelatitudinalealtitudinal (Manninen et al., 1998, 2002; Nault, 2003) varia-tions of the concentration of terpenes in the needles of conifershave been documented.
Data concerning the effect of various anthropogenic factorson secondary compounds of the conifers is still controversial
* Corresponding author. Tel.: þ1 370 6122 3391.
E-mail address: [email protected] (E. Kupcinskiene).
0269-7491/$ - see front matter � 2008 Elsevier Ltd. All rights reserved.
doi:10.1016/j.envpol.2008.02.001
Available online at www.sciencedirect.com
Environmental Pollution 155 (2008) 481e491www.elsevier.com/locate/envpol

Author's personal copy
(Barnola and Cedeno, 2000; Zavala and Ravetta, 2002; Turtolaet al., 2006). Elevated CO2 concentration caused an increase inthe concentration of a-pinene (Heyworth et al., 1998; Sallaet al., 2001), while influence of elevated UV-B radiation onsecondary compounds of the conifers was not observed (Tur-tola et al., 2006). Increased limonene emission rates inozone-fumigated woody plants were described (Klusia et al.,2002). Wider surveys of the conifer essential oil compositionunder acidic and alkaline pollutant effects are still scarce(Heller et al., 1990; Fuksman, 2002; Kainulainen et al.,1992; Supuka and Berta, 1998). The potential for air pollutioneffects on biogenic VOC emissions should be considered whenassessing forest health (Erisman et al., 2005; Cape, 2007;Heath, 2007).
In Lithuania, since the 1990s the reduced level of emissionsin a cement factory (operating since 1952), and an oil refinery(since 1980) caused further adverse changes in the surround-ing forests (Kupcinskiene, 2001; Kupcinskiene and Huttunen,2005) and some disturbances were registered in the ongoingdecade (Ceburnis et al., 2002; Kupcinskiene, 2006; Judzen-tiene et al., 2007).
The present study was aimed to evaluate whether presentlevels of industrial pollution caused by the oil refinery andthe cement factory affects the qualitative and quantitativecharacteristics of the essential oils in the needles of Scotspine (Pinus sylvestris L.) growing under different levels ofpollution and at different distances from the pollution source.
2. Materials and methods
2.1. Study area
The Scots pine (Pinus sylvestris L.) growing along the transects from the
cement factory (CF) and the oil refinery (OR) in Lithuania were investigated.
In 2004e2005, total emissions from the CF comprised up to 3000 t year�1 and
emissions from the OR 24,000 t year�1 (data provided by the Ministry of En-
vironment of Lithuania). The total deposition of calcium and magnesium ions
along the CF transect ranged between 19.1 and 2.6 kg ha�1 month�1 (Armo-
laitis et al., 1999). Elevated concentrations of heavy metals in the mosses
were documented near CF and OR (Ceburnis et al., 2002). Sites near the
OR differed in the amounts of sulphur dioxide up to 6.7 times (with the highest
mean monthly concentration of 27 mg m�3; in 2005, being lower, 9.4 mg m�3),
and in the amounts of nitrogen dioxide up to 2 times (with the highest mean
monthly concentration of 6 mg m�3, registered in 2005). Sites near the CF dif-
fered in the amounts of sulphur dioxide up to 9 times (with the highest mean
monthly concentration of 8.3 mg m�3, registered in 2005; Kupcinskiene,
2006), and in the amounts of nitrogen dioxide up to 1.9 times (with the highest
mean monthly concentration of 3 mg m�3). In both transects, the most polluted
sites were the ones closest to the factories. The furthest stands were used as
control sites. Before sampling severe drought was registered (Table 1).
Middle-aged pines (51e62 years old) growing on histosols (near the CF),
and 46e75-year-old trees growing on luvisols (near the OR) were examined.
Selection of sites was based on availability of the stands and the prevailing
wind direction from each pollution source. Related to the CF, four stands be-
longing to Carico-sphagno-Pinetum (siccata) type in a 10-km transect (north-
east direction) were studied. Near the OR, four stands of Oxalido-Pinetum type
in a 5.4-km northeast transect and one stand (3.6 km away from the OR, east
direction) was examined.
2.2. Plant material
Branches facing the wind coming from the factory were cut from eight
pines at the height of 6e8 m above the ground (the third lowest part of the
crown). From each tree, four shoots with the current-year (c) and 1-year-old
(c þ 1) needles were used. July (10/07/2005) was chosen for sampling due
to the most vigorous growth of the new needles that expanded in June. For
analysis, four independent samples were prepared from each stand. A separate
sample was made by mixing equal amounts of the needles collected from eight
trees (Schulz et al., 1998) and drying the material at room temperature (20e
25 �C). Defoliation (expressed in %) of the sampled trees was estimated
according to EU standards (Anonymous, 1989).
2.3. Oil isolation and analysis
Oil yield was conducted by hydrodistillation of 50 g of dry needles. Pale
yellow oils were obtained in 0.2e0.6 and 0.2e0.3% (v/w) of the yield, respec-
tively, in the current-year and 1-year-old needles on a dry mass basis. GC anal-
ysis was done by an HP 5890(II) chromatograph equipped with FID and
capillary column HP-FFAP (30 m � 0.25 mm i.d., film thickness 0.25 mm).
Analyses by GC/MS were performed using an HP 5890 chromatograph inter-
faced to an HP 5971 mass spectrometer (ionization voltage 70 eV) and equip-
ped with a capillary column CP-Sil 8 CB (50 m � 0.32 mm i.d., film thickness
0.25 mm). Other details of oil analyses were described earlier (Mockute et al.,
2003; Mockute and Judzentiene, 2004). Qualitative analysis was based on
a comparison of retention times, indices with mass spectra libraries (Wiley
and NBS 54K), and other corresponding data (Adams, 2001). Quantity of
each component of essential oils was calculated as relative concentration
(peak area percentage) and amount (mg g�1 d.m., recalculated according to
the internal standart).
2.4. Statistical analysis
To compare the stands along the transects, dispersion and correlation anal-
yses were applied using EXCEL, SPSS, and SAS packages. Error bars in the
figures indicate an interval of 95% confidence. The normality of the data dis-
tribution was assessed and log transformations were performed. Data were
analysed with two-way analysis of variance. The significance of differences
between sites was assessed by Tukey’s multiple range test. Variances of con-
centration logarithms with their components (site, needle age and their inter-
action) were estimated by maximum likelihood (Schleppi et al., 2000).
3. Results
3.1. Needle essential oil general characteristics
There were no significant differences between the standsalong separate transects in the total yield of the essential oilsin the needles. Seventy identified components made up to89.1e95.1% of total oil content. The qualitative compositionof the main components appeared to be constant in the needlesof all pine stands investigated. However, there were consider-able differences in the amounts of separate components.The predominant fraction was found to be monoterpenes(19.0e40.0%), with the major constituents being a-pinene
Table 1
Precipitation (sum within decades, in mm) and temperature (mean values for
the decades) in JuneeJuly 2005, according to the data of the closest to the
transects Meteorological Station (Telsiai, Lithuania)
Month Decade Precipitation, mm Temperature�C
June I 3.1 11.9
II 0.6 15.8
III 2.0 16.0
July I 1.1 19.0
482 E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491
Author's personal copy
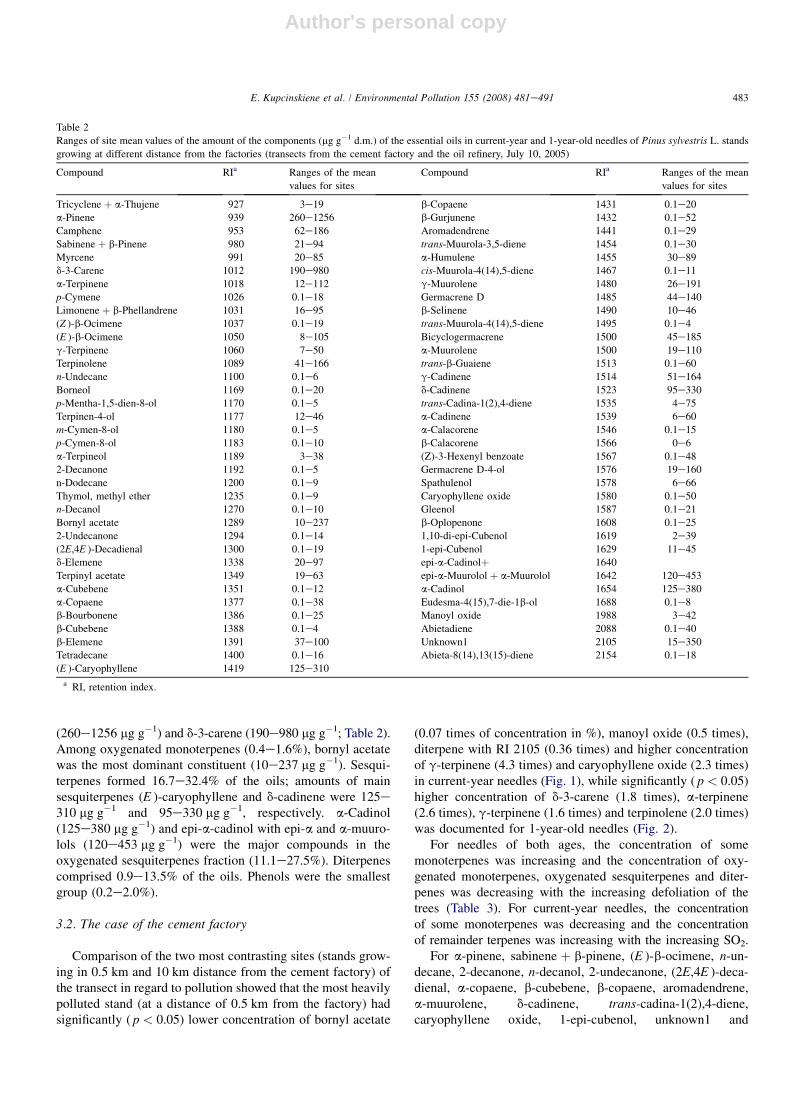
(260e1256 mg g�1) and d-3-carene (190e980 mg g�1; Table 2).Among oxygenated monoterpenes (0.4e1.6%), bornyl acetatewas the most dominant constituent (10e237 mg g�1). Sesqui-terpenes formed 16.7e32.4% of the oils; amounts of mainsesquiterpenes (E )-caryophyllene and d-cadinene were 125e310 mg g�1 and 95e330 mg g�1, respectively. a-Cadinol(125e380 mg g�1) and epi-a-cadinol with epi-a and a-muuro-lols (120e453 mg g�1) were the major compounds in theoxygenated sesquiterpenes fraction (11.1e27.5%). Diterpenescomprised 0.9e13.5% of the oils. Phenols were the smallestgroup (0.2e2.0%).
3.2. The case of the cement factory
Comparison of the two most contrasting sites (stands grow-ing in 0.5 km and 10 km distance from the cement factory) ofthe transect in regard to pollution showed that the most heavilypolluted stand (at a distance of 0.5 km from the factory) hadsignificantly ( p < 0.05) lower concentration of bornyl acetate
(0.07 times of concentration in %), manoyl oxide (0.5 times),diterpene with RI 2105 (0.36 times) and higher concentrationof g-terpinene (4.3 times) and caryophyllene oxide (2.3 times)in current-year needles (Fig. 1), while significantly ( p < 0.05)higher concentration of d-3-carene (1.8 times), a-terpinene(2.6 times), g-terpinene (1.6 times) and terpinolene (2.0 times)was documented for 1-year-old needles (Fig. 2).
For needles of both ages, the concentration of somemonoterpenes was increasing and the concentration of oxy-genated monoterpenes, oxygenated sesquiterpenes and diter-penes was decreasing with the increasing defoliation of thetrees (Table 3). For current-year needles, the concentrationof some monoterpenes was decreasing and the concentrationof remainder terpenes was increasing with the increasing SO2.
For a-pinene, sabinene þ b-pinene, (E )-b-ocimene, n-un-decane, 2-decanone, n-decanol, 2-undecanone, (2E,4E )-deca-dienal, a-copaene, b-cubebene, b-copaene, aromadendrene,a-muurolene, d-cadinene, trans-cadina-1(2),4-diene,caryophyllene oxide, 1-epi-cubenol, unknown1 and
Table 2
Ranges of site mean values of the amount of the components (mg g�1 d.m.) of the essential oils in current-year and 1-year-old needles of Pinus sylvestris L. stands
growing at different distance from the factories (transects from the cement factory and the oil refinery, July 10, 2005)
Compound RIa Ranges of the mean
values for sites
Compound RIa Ranges of the mean
values for sites
Tricyclene þ a-Thujene 927 3e19 b-Copaene 1431 0.1e20
a-Pinene 939 260e1256 b-Gurjunene 1432 0.1e52
Camphene 953 62e186 Aromadendrene 1441 0.1e29
Sabinene þ b-Pinene 980 21e94 trans-Muurola-3,5-diene 1454 0.1e30
Myrcene 991 20e85 a-Humulene 1455 30e89
d-3-Carene 1012 190e980 cis-Muurola-4(14),5-diene 1467 0.1e11
a-Terpinene 1018 12e112 g-Muurolene 1480 26e191
p-Cymene 1026 0.1e18 Germacrene D 1485 44e140
Limonene þ b-Phellandrene 1031 16e95 b-Selinene 1490 10e46
(Z )-b-Ocimene 1037 0.1e19 trans-Muurola-4(14),5-diene 1495 0.1e4
(E )-b-Ocimene 1050 8e105 Bicyclogermacrene 1500 45e185
g-Terpinene 1060 7e50 a-Muurolene 1500 19e110
Terpinolene 1089 41e166 trans-b-Guaiene 1513 0.1e60
n-Undecane 1100 0.1e6 g-Cadinene 1514 51e164
Borneol 1169 0.1e20 d-Cadinene 1523 95e330
p-Mentha-1,5-dien-8-ol 1170 0.1e5 trans-Cadina-1(2),4-diene 1535 4e75
Terpinen-4-ol 1177 12e46 a-Cadinene 1539 6e60
m-Cymen-8-ol 1180 0.1e5 a-Calacorene 1546 0.1e15
p-Cymen-8-ol 1183 0.1e10 b-Calacorene 1566 0e6
a-Terpineol 1189 3e38 (Z)-3-Hexenyl benzoate 1567 0.1e48
2-Decanone 1192 0.1e5 Germacrene D-4-ol 1576 19e160
n-Dodecane 1200 0.1e9 Spathulenol 1578 6e66
Thymol, methyl ether 1235 0.1e9 Caryophyllene oxide 1580 0.1e50
n-Decanol 1270 0.1e10 Gleenol 1587 0.1e21
Bornyl acetate 1289 10e237 b-Oplopenone 1608 0.1e25
2-Undecanone 1294 0.1e14 1,10-di-epi-Cubenol 1619 2e39
(2E,4E )-Decadienal 1300 0.1e19 1-epi-Cubenol 1629 11e45
d-Elemene 1338 20e97 epi-a-Cadinolþ 1640
Terpinyl acetate 1349 19e63 epi-a-Muurolol þ a-Muurolol 1642 120e453
a-Cubebene 1351 0.1e12 a-Cadinol 1654 125e380
a-Copaene 1377 0.1e38 Eudesma-4(15),7-die-1b-ol 1688 0.1e8
b-Bourbonene 1386 0.1e25 Manoyl oxide 1988 3e42
b-Cubebene 1388 0.1e4 Abietadiene 2088 0.1e40
b-Elemene 1391 37e100 Unknown1 2105 15e350
Tetradecane 1400 0.1e16 Abieta-8(14),13(15)-diene 2154 0.1e18
(E )-Caryophyllene 1419 125e310
a RI, retention index.
483E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
abieta-8(14),13(15)-diene, the main source of variation (Table4) was site and for the biggest part of the other compoundsneedle age differences were greater compared to sitedifferences.
3.3. The case of the oil refinery
Comparison of the two most contrasting sites of the transectin regards to pollution showed that the most heavily pollutedstand (at a distance of 2.0 km from the factory) had a signifi-cantly ( p < 0.05) higher concentration of sabinene þ
b-pinene (1.8 times), 1-epi-cubenol (2.2 times) and lower con-centration of manoyl oxide (0.1 times), diterpene with RI 2105(0.4 times) in the current-year-needles (Fig. 3) than the leastpolluted site (located at 5.4 km away from the oil refinery).Also, the 1-year-old needles of the most polluted site pre-sented a significantly ( p < 0.05) higher concentration of cam-phene (3.1 times), sabinene þ b-pinene (3.2 times), myrcene(2.4 times), a-cadinene (2.5 times), 1-epi-cubenol (2.4 times)and a lower percentage of manoyl oxide (0.1 times) and diter-pene with RI 2105 (0.2 times) than those sampled at the leastpolluted site (Fig. 4).
γ-Terpinene
020406080
0.5 1.5 3.5 10Distance, km
μg g
-1
μg g
-1
μg g
-1
μg g
-1
μg g
-1
Caryophyllene oxide
01020304050
0.5 1.5 3.5 10Distance, km
Manoyl oxide
0.1
10.1
20.1
30.1
0.5 1.5 3.5 10Distance, km
Unknown (RI 2105)
0100200300400
0.5 1.5 3.5 10Distance, km
Bornyl acetate
-50
0
50
100
150
0.5 1.5 3.5 10Distance, km
Fig. 1. Concentration (mg g�1 d.m.; mean values, n ¼ 4) of the components of essential oils in current-year needles of Scots pine (Pinus sylvestris L.) stands
growing at different distance from the cement factory (July 10, 2005). Error bars indicate an interval of 95% confidence.
δ-3-Carene
0
500
1000
1500
0.5 1.5 3.5 10Distance, km
μg g
-1
α-Terpinene
020406080
100
0.5 1.5 3.5 10Distance, km
γ-Terpinene
05
10152025
0.5 1.5 3.5 10Distance, km
μg g
-1
Terpinolene
0
50
100
150
200
0.5 1.5 3.5 10Distance, km
μg g
-1
μg g
-1
Fig. 2. Concentration (mg g�1 d.m.; mean values, n ¼ 4) of the components of essential oils in 1-year-old needles of Scots pine (Pinus sylvestris L.) stands growing
at different distance from the cement factory (July 10, 2005). Error bars indicate an interval of 95% confidence.
484 E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
In current-year needles and 1-year-old needles, the con-centration of the most monoterpenes, some sesquiterpenesand oxygenated sesquiterpenes was increasing with the in-creasing defoliation of the trees/increasing concentration ofSO2 (Table 5), while opposite relations were found forditerpenes.
Along the oil refinery transect, the most heavily pollutedstand (a distance of 2.0 km from the factory) had the lowest(by a large degree) concentration of diterpenes in both cur-rent-year and 1-year-old needles.
For tricyclene þ a-thujene, a-pinene, camphene, myrcene,d-3-carene, (Z )-b-ocimene, (E )-b-ocimene, g-terpinene, n-un-decane, borneol, p-mentha-1,5-dien-8-ol, m-cymen-8-ol,p-cymen-8-ol, n-dodecane, thymol methyl ether, n-decanol,bornyl acetate, terpinyl acetate, n-tetradecane, cis-muurola-4(14),5-diene, g-muurolene, germacrene D-4-ol, trans-muur-ola-4(14),5-diene, a-muurolene, a-cadinene, caryophyllene
Table 3
Correlation coefficients between the concentration of the components of the
essential oils in current-year (c) and 1-year-old needles (c þ 1) and defoliation
of the Scots pine (Pinus sylvestris L.) stand/sulphur dioxide concentration
(transect from the cement factory; July 10, 2005)
Compound Defoliation SO2
c needles c þ 1 needles c needles c þ 1 needles
Monoterpenes
Myrcene n.s. n.s. �0.664** n.s.
a-Terpinene n.s. 0.722** �0.891** n.s.
Limonene þb-Phellandrene
n.s. n.s. �0.511* n.s.
g-Terpinene 0.827** n.s. n.s. 0.610*
Oxygenated monoterpenesBornyl acetate �0.814** n.s. n.s. n.s.
Borneol �0.507* n.s. n.s. n.s.
Terpinen-4-ol
Sesquiterpenes
a-Cubebene n.s. n.s. n.s. 0.734**
b-Cubebene n.s. n.s. n.s. 0.641**
a-Copaene �0.507* �0.617* n.s. �0.702**
a-Humulene 0.498* n.s. n.s. n.s.
b-Selinene 0.504* �0.652** 0.647** �0.621**
Bicyclogermacrene n.s. n.s. 0.684** n.s.
g-Cadinene 0.499* n.s. n.s. n.s.
a-Calacorene n.s. n.s. n.s. �0.513*
Oxygenated sesquiterpenes
Germacrene D-4-ol n.s. n.s. 0.619* n.s.
Spathulenol n.s. n.s. 0.616* n.s.
Caryophyllene oxide �0.721** n.s. 0.499* n.s.
1,10-di-epi-Cubenol n.s. n.s. 0.537* n.s.
1-epi-Cubenol n.s. n.s. 0.570* n.s.
epi-a-Muurolol þa-Muurolol
n.s. �0.540* n.s. n.s.
a-Cadinol �0.558* 0.565*
DiterpenesManoyl oxide �0.752** n.s. n.s. n.s.
Unknown1 �0.851** n.s. n.s. n.s.
n.s., non-significant ( p>0.05) correlations; *p < 0.05; **p < 0.01.
Table 4
Variances of concentration (of the components of essential oils in the needles
of Pinus sylvestris L. growing at different distance from the cement factory)
logarithms with their components (site, needle age, site � needle age interac-
tion) estimated by maximum likelihood (July 10, 2005)
Components of essential oils Site Age Site � Age Residuals
Tricyclene þ a-Thujene 0.131 1.560 0.148 0.554
a-Pinene 0.478 0.002 0.113 0.080
Camphene 0.034 1.034 0.008 0.124
Sabinene þ b-Pinene 0.145 0.450 0.076 0.087
Myrcene 0.221 2.525 0.403 0.269
d-3-Carene 1.673 2.813 0.635 0.102
a-Terpinene 1.717 1.767 3.623 0.271
p-Cymene 0.524 3.811 1.566 0.757
Limonene þ b-Phellandrene 0.180 0.335 0.292 0.103
(Z )-b-Ocimene 0.986 2.781 0.443 0.955
(E )-b-Ocimene 4.175 2.486 0.480 0.118
g-Terpinene 2.037 0.020 0.857 0.119
Terpinolene 1.363 0.947 0.779 0.084
n-Undecane 0.307 0.005 0.307 0.206
Borneol 1.742 3.509 2.096 1.002
p-Mentha-1,5-dien-8-ol 0.827 0.199 0.920 1.127
Terpinen-4-ol 0.119 0.365 0.903 0.227
m-Cymen-8-ol 0 0 0 0
p-Cymen-8-ol 0.147 1.207 0.722 0.965
a-Terpineol 0.113 5.163 0.011 0.207
2-Decanone 2.547 0.045 0.045 0.286
n-Dodecane 0.272 0.659 0.851 0.282
Thymol, methyl ether 0.710 0.410 0.463 1.186
n-Decanol 1.043 0.080 0.045 0.255
Bornyl acetate 2.184 4.921 0.363 0.190
2-Undecanone 2.163 0.325 1.377 0.443
(2E,4E )-Decadienal 2.481 0.465 0.321 0.902
b-Elemene 0.077 1.822 0.127 0.421
Terpinyl acetate 0.162 1.390 0.384 0.242
a-Cubebene 1.284 3.602 1.385 0.203
a-Copaene 2.831 0.472 0.477 0.388
b-Bourbonene 1.752 0.060 2.053 0.745
b-Cubebene 2.490 0.166 0.690 0.458
b-Elemene 0.079 0.000 0.254 0.143
Tetradecane 0.007 0.007 0.305 0.271
(E )-Caryophyllene 0.038 1.529 0.225 0.098
b-Copaene 1.184 0.551 0.072 0.900
b-Gurjunene 0.558 2.064 0.248 1.028
Aromadendrene 1.013 0.017 0.502 0.601
trans-Muurola-3,5-diene 0.408 0.663 0.573 1.202
a-Humulene 0.084 2.187 0.097 0.109
cis-Muurola-4(14),5-diene 0.039 0.318 0.064 0.612
g-Muurolene 0.441 3.268 1.080 0.432
Germacrene D 0.151 0.802 0.490 0.324
b-Selinene 0.066 0.254 0.736 0.088
trans-Muurola-4(14),5-diene 0.297 0.081 0.081 1.337
Bicyclogermacrene 0.142 1.979 0.427 0.123
a-Muurolene 0.548 0.281 0.444 0.150
trans-b-Guaiene 0.166 0.166 0.166 0.166
g-Cadinene 0.249 0.510 0.200 0.087
d-Cadinene 0.179 0.040 0.245 0.113
trans-Cadina-1(2),4-diene 0.838 0.043 0.394 0.796
a-Cadinene 0.183 1.353 0.201 0.181
a-Calacorene 0.603 0.425 2.594 0.665
b-Calacorene 0.324 1.913 0.678 1.169
(Z)-3-Hexenyl benzoate 0.335 3.205 1.527 0.671
Germacrene D-4-ol 0.259 9.869 0.273 0.109
Spathulenol 1.5663 17.106 0.587 0.261
Caryophyllene oxide 1.594 0.067 0.185 0.120
Gleenol 0.726 2.481 1.202 0.830
(continued on next page)
485E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
oxide, 1-epi-cubenol, a-cadinol and unknown1, the mainsource of variation (Table 6) was the site.
4. Discussion
Varying amounts of terpene species in the needles of pineswere described by different research teams: 10 (Berta et al.,1997), 42 (Bojovic et al., 2005), 55 (Idzojtic et al., 2005),56 (De Simon et al., 2001); in our study 70 componentswere identified. Generally, our results concerning the compo-sition of the essential oils in the needles of P. sylvestris weresimilar to those reviewed by Lawrence (1991). Sixteen mono-terpenes are described among 10 pine species (Da Silva et al.,2001). In our Scots pine study 13 monoterpenes were identi-fied. Domination of monoterpenes and sesquiterpenes ob-served in our study are in agreement with other studies ofconifers (Dormont et al., 1998; De Simon et al., 2001; Manni-nen et al., 2002; Bojovic et al., 2005).
Decrease in the concentration of d-3-carene, terpinolene(near the cement factory), and an increase in the concentrationof camphene, sabinene þ b-pinene (near the oil refinery) wasdocumented in the needles of the pines growing closest to thefactories. Conifer changes in the needle a-pinene at thepolluted urban site (Berta et al., 1997; Supuka et al., 1997)
or under elevated CO2 concentration were reported (Heyworthet al., 1998; Salla et al., 2001).
Scots pine stands growing along transects from the factoriesin Lithuania differed significantly not only according to theconcentrations of some major (mentioned above) compounds,but also according to intermediate (camphene, myrcene, terpi-nolene, sabinene þ b-pinene, a-terpinene, bornyl acetate,d-elemene, b-elemene, g-muurolene, bicyclogermacrene, g-cadinene) and minor (g-terpinene, a-terpineol, a-cadinene,caryophyllene oxide, 1-epi-cubenol, manoyl oxide) compo-nents of essential oils in the needles. Continuous stress onthe pines due to the factories might induce, in part, the ob-served needle differences in quantities of separate componentsof the essential oils between stands growing along selectedtransects in Lithuania. This lends support to reported changesin the terpene production of several conifers under the effect ofcontaminated urban atmosphere (Juttner, 1988; Supuka andBerta, 1998).
Concentration correlations between air pollutants and theseparate components of essential oils in the needles corre-sponded to the data summed for monoterpenes, oxygenatedmonoterpenes, sesquiterpenes, oxygenated sesquiterpenesand diterpenes (Judzentiene et al., 2007). Along the CF tran-sect for current-year needles, the percentage of diterpeneswas decreasing with the increasing concentration of SO2
(r ¼ �0.573; p < 0.05). Along the OR transect, in both thecurrent-year and 1-year-old needles, the percentage of diter-penes was decreasing with the increasing SO2 (respectively,r ¼ �0.773, p < 0.01; r ¼ �0.486, p < 0.05); an opposite re-lation was true for sesquiterpenes (respectively, r ¼ �0.751,p < 0.01; r ¼ 0.785, p < 0.01). Similar to what was shownin many other countries, examinations of the industrial tran-sects in Lithuania showed that worsening of general tree con-dition could be related to the effects of the factories(Armolaitis et al., 1999; Kupcinskiene, 2001; Kupcinskieneand Huttunen, 2005). Defoliation in conifers has been associ-ated with changes in leaf primary and secondary metabolite
Table 4 (continued)
Components of essential oils Site Age Site � Age Residuals
b-Oplopenone 0.335 0.478 0.809 0.604
1,10-di-epi-Cubenol 0.161 0.115 0.543 0.384
1-epi-Cubenol 0.448 0.077 0.150 0.158
epi-a-Cadinolþepi-a-Muurolol þ a-Muurolol 0.606 0.950 0.358 0.166
a-Cadinol 0.341 1.119 0.209 0.114
Eudesma-4(15),7-1b-ol 0.3456 0.785 0.790 1.058
Manoyl oxide 0.486 2.620 1.075 0.098
Abietadiene 0.259 0.003 0.957 0.400
Unknown1 0.617 0.117 0.590 0.104
Abieta-8(14),13(15)-diene 2.110 0.019 1.733 0.731
Sabinene +β-Pinene
0
20
40
60
2.0 2.5 3.6 5.3 5.4
Distance, km
μg g
-1
μg g
-1
μg g
-1
μg g
-1
1-epi-Cubenol
-20
0
20
40
60
2.0 2.5 3.6 5.3 5.4
Distance, km
Manoyl oxide
0
10
20
30
40
2.0 2.5 3.6 5.3 5.4Distance, km
Unknown (RI 2105)
0100200300400500
2.0 2.5 3.6 5.3 5.4Distance, km
Fig. 3. Concentration (mg g�1 d.m.; mean values, n ¼ 4) of the components of essential oils in current-year needles of Scots pine (Pinus sylvestris L.) stands grow-
ing at different distance from the oil refinery (July 10, 2005). Error bars indicate an interval of 95% confidence.
486 E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
chemistry (Wagner and Evans, 1985; Schonwitz et al., 1991);also relations between defoliation of conifers affected by in-sects and the concentration of terpenes in the needles weredocumented (Baranchikov et al., 1991), while some other ob-servations did not show a correlation between the percentageof needle loss and percentile terpene amounts in the needles(Schonwitz et al., 1991). Plant secondary metabolism relationswith defoliation caused by air pollution are poorly described.In our study, we observed a decrease in the concentration ofditerpenes in the needles for the stands (the nearest to the ce-ment factory and the oil refinery) that had the higher defolia-tion and shorter needle retention (Kupcinskiene, 2001;Kupcinskiene and Huttunen, 2005; Stikliene et al., 2006; Jud-zentiene et al., 2007). Among the stands of the selected indus-trial transects the extent of the differences for concentration ofseparate compounds of essential oils in the needles varied upto 10 times (transect from the cement factory) and also upto 10 times (transect from the oil refinery), while tree defolia-tion of the transects differed respectively 1.7, 1.8 and 1.3 timesand aerial pollutant concentrations differed along transects re-spectively 5.5 (SO2) and 1.8 (SO2) times. Variability of the
results (differences concerning distance, defoliation and SO2
effects) might be caused by several reasons. Due to differentresidence times for pollutants emitted from the factory ineach site, a special set (in terms of concentration) of toxiccompounds is formed. In addition to the SO2 effect, defolia-tion is a consequence of a variety of other stressors of anthro-pogenic and natural origin. Selection of sites was limited byavailability of pine stands, which is why only the furthest sitesof the transects corresponded exactly to the direction of theprevailing wind. Strong chemical reactivity of essential oilsalso might be an important source of variation.
Along two industrial transects the increase in the amount ofsome diterpenes and decrease in the components of the shorterchain essential oils was observed. These effects could be atleast partly attributed to sulphur dioxide that is omitted byboth factories. Changes in the concentration of some compo-nents of the essential oils in the needles of Scots pine werehigher to some extent compared to the differences in visible(defoliation of the trees) damage to the trees, and might bea useful tool for the detection of latent changes caused byindustrial, especially sulphur dioxide, pollution.
α-Cadinene
0
10
20
30
2.0 2.5 3.6 5.3 5.4Distance, km
Camphene
050
100150200250
2.0 2.5 3.6 5.3 5.4Distance, km
μg g
-1
μg g
-1
μg g
-1
μg g
-1
μg g
-1
μg g
-1
μg g
-1
Sabinene + β-Pinene
0
50
100
150
2.0 2.5 3.6 5.3 5.4Distance, km
Myrcene
020406080
100
2.0 2.5 3.6 5.3 5.4Distance, km
1-epi-Cubenol
0
50
100
150
2.0 2.5 3.6 5.3 5.4Distance, km
Manoyl oxide
0
20
40
60
80
2.0 2.5 3.6 5.3 5.4Distance, km
Unknown (RI 2105)
0
200
400
600
800
2.0 2.5 3.6 5.3 5.4Distance, km
Fig. 4. Concentration (mg g�1 d.m.; mean values, n ¼ 4) of the components of essential oils in 1-year-old needles of Scots pine (Pinus sylvestris L.) stands growing
at different distance from the oil refinery (July 10, 2005). Error bars indicate an interval of 95% confidence.
487E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
In the Picea abies study diminished content of the needlemonoterpenes was registered under 14 months of treatmentby ozone and acid mist (Heller et al., 1990), although diter-penes were not estimated. In another study involving ozoneand acid mist, different terpene patterns existed but couldnot be connected with any of the experimental conditions(Schonwitz and Merk, 1986). Further evidence, especially in
controlled-condition experiments, is needed to test ourfindings.
Relations between nutrient availability and the secondaryplant compounds in Scots pine have been described in otherstudies (Barnola and Cedeno, 2000; Lavola et al., 2003).Our results obtained in Scots pine nutritional studies alongtransects from the cement factory and the oil refinery (Kup-cinskiene, 2006) are consistent with those that have reportedchanges in volatile terpenoid concentrations in the leaves ofseveral plant species growing in conditions with low nitro-gen and phosphorus availability (Barnola and Cedeno,2000).
Qualitative and quantitative terpene variations between co-nifer needles of different age have been described (Cariseyand Bauce, 1997; Helanova et al., 2006; Lagalante et al.,2006; Campbell and Taylor, 2007). Seasonal and annual(Nault, 2003) geographicalelatitudinalealtitudinal (Manninenet al., 1998, 2002; Nault, 2003) variations in the concentrationof terpenes in the needles of conifers might be attributed atleast partially to the needle age-related effects, although infor-mation concerning pollutant effects on terpenes in needles ofdifferent age is scarce. It is documented that altered nitrogensupply affects allocation to secondary metabolites differently,depending on the age of the needles (Kainulainen et al.,1996). According to the present data needle age was themain source of variation for the biggest part of the compo-nents of essential oils. In our study comparison of the needlesof two age classes revealed diverse reactions concerning thecomponents of essential oils affected by pollution accordingto the presence/absence of the reaction, direction (increase/decrease) of the effect, and the extent (changes in concentra-tion). Correlations between concentration of the componentsof the essential oils and tree defoliation depended on theage of the needles. Cluster analysis of the concentration ofessential oils in the needles applied for industrial transectsshowed larger effects of pollution for current-year needlescompared to 1-year-old needles in the case of pine standsnear the oil refinery (Judzentiene et al., 2007). Despite presentlow-level dust emissions from the cement factory, both needleage classes showed significant factory distance-relatedchanges, confirming the opinion about the absence of thresh-old for dust effects (Farmer, 1993; Stikliene et al., 2006). For-mer studies of needle surface erosion along the sameindustrial pine stand transects revealed a higher indicativevalue of the older-age needles (Kupcinskiene and Huttunen,2005), while some nutritional transformations were observedfor current-year needles (Kupcinskiene, 2006). The presentstudy provides evidence that in a polluted environment impor-tant biochemical changes occur within the first month afterneedle expansion. It is well known that younger needles aremore resistant than older ones to various stressors. Biggerchanges in essential oils for freshly expanded (less than1 month old) current-year needles might reflect their protec-tive reaction to acute effects of pollutants including SO2.Older needles might be more adapted for chronic exposureto pollutants and their defence aids might also be alreadyexhausted.
Table 5
Correlation coefficients between the concentration of the components of essen-
tial oils in current-year (c) and 1-year-old needles (c þ 1) and defoliation of
the Scots pine (Pinus sylvestris L.) stand/sulphur dioxide concentration (tran-
sect from the oil refinery, July 10, 2005)
Compound Defoliation SO2
c needles c þ 1 needles c needles c þ 1 needles
Monoterpenes
a-Pinene n.s. n.s. 0.447* n.s.
Camphene n.s. 0.737** n.s. n.s.
Sabinene þ b-Pinene 0.633** 0.730** 0.596** n.s.
Myrcene 0.555* 0.577** 0.445* n.s.
d-3-Carene n.s. �0.540* n.s. �0.598**
(Z )-b-Ocimene 0.478* n.s. n.s. n.s.
Limonene þ b-
Phellandrene
n.s. 0.520* 0.448* n.s.
(E )-b-Ocimene 0.591** 0.488* 0.585** n.s.
c-Terpinene n.s. 0.541* n.s. 0.466*
Terpinolene �0.571** 0.483* �0.589** n.s.
Oxygenated monoterpenes
Borneol 0.448* 0.444* n.s. n.s.
p-Mentha-1,5-dien-8-ol �0.513* n.s. n.s. n.s.
Terpinen-4-ol 0.608** n.s. 0.556* n.s.
m-Cymen-8-ol 0.594** 0.471* n.s. n.s.
p-Cymen-8-ol n.s. n.s. n.s. 0.596*
a-Terpineol 0.443* n.s. 0.499* n.s.
Bornyl acetate n.s. n.s. n.s. �0.554*
Terpinyl acetate 0.497* 0.751** n.s. 0.581**
Sesquiterpenes
b-Bourbonene 0.449 * n.s. n.s. n.s.
a-Cubebene n.s. �0.471* n.s. �0.471*
a-Copaene n.s. 0.522* n.s. 0.562*
b-Cubebene �0.471* �0.594** �0.471** �0.594**
b-Elemene n.s. n.s. 0.585** 0.682**
(E )-Caryophyllene n.s. n.s. n.s. 0.463*
Aromadendrene n.s. 0.654** n.s. 0.783**
a-Humulene n.s. n.s. n.s. 0.780**
g-Muurolene n.s. n.s. n.s. 0.674**
Germacrene D n.s. n.s. n.s. 0.490*
Bicyclogermacrene 0.627** 0.449* 0.751** 0.793**
a-Muurolene 0.683** 0.605** 0.610** 0.675**
c-Cadinene 0.629** n.s. 0.513* 0.460*
d-Cadinene 0.529* 0.636** 0.461* 0.746**
a-Cadinene n.s. 0.822* n.s. 0.797**
Oxygenated sesquiterpenesGermacrene D-4-ol 0.492* n.s. n.s. n.s.
Spathulenol 0.619** n.s. n.s. n.s.
1-epi-Cubenol n.s. 0.735** 0.446* n.s.
Diterpenes
Unknown1 �0.553* �0.914** �0.737** �0.521*
n.s., non-significant correlations; *p < 0.05; **p < 0.01.
488 E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
Because pollen and seeds of the pine are dispersed by wind,the role of the essential oils in the needles cannot be related tothe attraction of insects and might be quite unambiguouslyconnected with protection against pathogens and also para-sites. The essential oils or their components have been shownto exhibit antiviral, antibacterial, antimycotic, antitoxigenic,antiparasitic, and insecticidal properties, and activity againstmite pests and nematodes (Supuka and Berta, 1998; Barnolaand Cedeno, 2000). Monoterpenes in particular might be a fac-tor determining host plant resistance (Barnola et al., 1997;Tiberi et al., 1999; Chen et al., 2002; Petrakis et al., 2005).In our study, changes in concentration of some major, interme-diate or minor components of the essential oils of the needles,mentioned above, may modify tree relations with the sur-rounding biota including susceptibility of the polluted (by in-dustrial emissions) pine stands to the pathogens and parasites.
Higher production of the shorter-chain terpenes (observedalong the cement factory and oil refinery transects) might bea consequence of tree growth in the polluted environment re-quiring energy consumption for the reparation processes,while at the same time investing smaller resources for synthe-sis of protective compounds. A two-fold increase in monoter-pene and sesquiterpene accumulation in the needles has beenreported under methyl jasmonate treatment (Martin et al.,2003). This suggests the way in which phytohormones mightbe involved in the plant biochemical response to a pollutedenvironment.
Monoterpenes, coming from the conifer forests, form a sig-nificant part in BVOC emissions (Geron et al., 2000; Cape,2007). Alterations in concentration of major components ofthe essential oils like d-3-carene and terpinolene a-pinene inthe needles of the pines growing closest to the factories maycontribute to the changes in emissions of terpenes from thepolluted forests.
5. Conclusions
Under the effect of present-level industrial emissions,changes occur in the concentration of components of the es-sential oils in the needles of Scots pine. Significantly affectedby industrial emissions, the major components of essential oils
Table 6
Variances of concentration (of the components of essential oils in the needles
of Pinus sylvestris L. growing at different distance from the oil refinery) log-
arithms with their components (site, needle age, site � needle age interaction)
estimated by maximum likelihood (July 10, 2005)
Components of essential oils Source of variance
Site Age Site � Age Residuals
Tricyclene þ a-Thujene 1.115 0.368 0.550 0.587
a-Pinene 0.362 0.042 0.152 0.036
Camphene 0.380 0.006 1.072 0.095
Sabinene þ b-Pinene 1.058 1.928 0.390 0.115
Myrcene 0.956 0.452 0.199 0.100
d-3-Carene 0.364 0.114 0.081 0.027
a-Terpinene 0.199 0.196 0.276 0.434
p-Cymene 0.180 0.321 0.247 0.880
Limonene þ b-Phellandrene 0.281 0 0.326 0.123
(Z )-b-Ocimene 1.945 0.213 0.814 0.507
(E )-b-Ocimene 0.926 0.150 0.415 0.099
g-Terpinene 0.364 0.163 0.314 0.194
Terpinolene 0.453 0.989 0.515 0.145
n-Undecane 0.189 0.126 0.102 0.239
Borneol 2.376 0.417 0.417 0.806
p-Mentha-1,5-dien-8-ol 2.096 2.617 1.239 0.477
Terpinen-4-ol 0.218 6.072 0.483 0.193
m-Cymen-8-ol 3.314 0.133 0.133 0.309
p-Cymen-8-ol 0.919 0.007 1.542 0.973
a-Terpineol 1.197 4.560 0.817 0.467
2-Decanone 1.414 6.725 1.699 0.438
n-Dodecane 0.309 0.062 0.035 0.387
Thymol, methyl ether 2.660 0.023 0.048 0.892
n-Decanol 1.629 0.101 0.513 0.484
Bornyl acetate 1.230 0.527 0.495 0.132
2-Undecanone 0.444 0.632 0.334 0.360
(2E,4E )-Decadienal 0.004 0.236 0.223 0.261
d-Elemene 0.399 4.357 0.248 0.078
Terpinyl acetate 0.807 0.001 0.143 0.073
a-Cubebene 0.530 0.133 0.795 0.309
a-Copaene 0.721 0.578 2.382 0.269
b-Bourbonene 0.921 0.373 1.648 0.390
b-Cubebene 2.927 0.224 0.224 0.268
b-Elemene 0.672 1.412 0.102 0.044
Tetradecane 1.144 0.009 0.049 0.210
(E )-Caryophyllene 0.286 1.565 0.114 0.024
b-Copaene 0 0 0 0
b-Gurjunene 0.016 0.089 0.411 0.364
Aromadendrene 0.822 1.831 0.545 0.328
trans-Muurola-3,5-diene 0.244 2.121 1.814 0.719
a-Humulene 0.293 0.929 0.143 0.039
cis-Muurola-4(14),5-diene 1.502 0.843 0.881 0.794
g-Muurolene 1.447 0.006 0.383 0.083
Germacrene D 0.314 0.017 0.201 0.103
b-Selinene 0.099 1.370 1.352 0.149
trans-Muurola-4(14),5-diene 1.148 0.159 0.468 0.555
Bicyclogermacrene 0.487 3.417 0.101 0.029
a-Muurolene 1.386 0.011 0.200 0.090
trans-b-Guaiene 0 0 0 0
g-Cadinene 0.557 2.696 0.286 0.230
d-Cadinene 0.750 6.367 0.313 0.526
trans-Cadina-1(2),4-diene 0.170 0.769 0.462 0.586
a-Cadinene 0.527 0.124 0.212 0.177
a-Calacorene 0.890 1.634 0.382 0.962
b-Calacorene 0.343 0.065 0.396 0.595
(Z)-3-Hexenyl benzoate 0.489 7.893 0.255 0.671
Germacrene D-4-ol 0.218 15.334 0.131 0.153
Spathulenol 1.776 7.869 0.821 0.594
Caryophyllene oxide 1.041 0.097 1.121 0.542
Table 6 (continued )
Components of essential oils Source of variance
Site Age Site � Age Residuals
Gleenol 0.065 3.084 0.298 0.844
b-Oplopenone 0.459 0.932 1.654 0.511
1,10-di-epi-Cubenol 0.332 0.755 0.340 0.326
1-epi-Cubenol 0.518 0.455 0.238 0.120
epi-a-Cadinolþepi-a-Muurolol þ
a-Muurolol
0.468 0.757 0.044 0.015
a-Cadinol 0.415 0.095 0.067 0.071
Eudesma-4(15).7-die-1b-ol 0 0 0 0
Manoyl oxide 1.956 2.196 0.362 0.319
Abietadiene 0.491 0.224 0.985 0.453
Unknown1 3.414 0.490 0.382 0.056
Abieta-8(14),13(15)-diene 1.092 1.193 0.736 0.512
489E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
in the needles were d-3-carene, terpinolene (decrease near thecement factory), camphene, sabinene þ b-pinene (increasenear the oil refinery), and diterpenes (decrease near the cementfactory and the oil refinery). Correlation analyses showed re-lations between the concentrations of air pollutants and the to-tal concentration of the main classes of the essential oils ofseparate components of these classes. Current-year needleswere more informative indicators of the effects of pollutionthan 1-year-old needles in the case of the OR transect, andneedles of both ages were equally valuable in the case ofthe CF transect. In general, the pollution from the oil refineryor the cement factory had caused higher proportions ofshorter-chain terpenes and lower proportions of longer-chainterpenes. Along selected industrial transects the concentrationof separate compounds of essential oils in the needles of pinesvaried up to 10 times, while transect tree defoliation differedto lower extent (up to 1.8 times).
Acknowledgements
This work was supported by the Study and Science Founda-tion of Lithuania, Project ‘‘FIBISTRESS’’, grant No. C-05033/05. The authors thank Rima Potockiene for assistance in statis-tical analyses and Dr Almantas Kliucius for visual assessmentof tree conditions.
References
Adams, R.P., 2001. Essential Oil Components by Quadrupole GC/MS. Al-
lured, Carol Stream, IL.
Allen, E.B., Temple, P.J., Bytnerowicz, A., Arbaugh, M.J., Sirulnik, A.G.,
Rao, L.E., 2007. Patterns of understory diversity in mixed coniferous for-
ests of southern California impacted by air pollution. The Scientific World
JOURNAL 7 (S1), 247e263, doi:10.1100/tsw.2007.72.
Anonymous, 1989. Manual on methodologies and criteria for harmonized
sampling, assessment, monitoring and analysis of the effects of air pollu-
tion on forests. UN/ECE, Hamburg/Geneva.
Armolaitis, K., Vaicys, M., Raguotis, A., Kubertaviciene, L., 1999. Affects of
pollutants from J/V ‘‘Akmen _es cementas’’ on forest ecosystems. In:
Ozolin�cius, R. (Ed.), Monitoring of Forest Ecosystems in Lithuania. Lu-
tute, Kaunas, pp. 65e76.
Baranchikov, Y.N., Ryzhkova, T.S., Safonova, L.V., Kudashova, F.N., 1991.
Energy and nitrogen-balance in gypsy-moth caterpillars when feeding on
the needles of larches damaged by miner insects. Soviet Journal of Ecol-
ogy 22, 388e393.
Barnola, L.F., Cedeno, A., 2000. Inter-population differences in the essential
oils of Pinus caribaea needles. Biochemical Systematics and Ecology
28, 923e931.
Barnola, L.F., Cedeno, A., Hasegawa, M., 1997. Intraindividual variations of
volatile terpene contents in Pinus caribaea needles and its possible rela-
tionship to Atta laevigata herbivory. Biochemical Systematics and Ecology
25, 707e716.
Berta, F., Supuka, J., Chladna, A., 1997. The composition of terpenes in nee-
dles of Pinus sylvestris in a relatively clear and in a city environment.
Biologia 52, 71e78.
Bojovic, S., Jurc, M., Drazic, D., Pavlovic, P., Mitrovic, M., Djurdjevic, L.,
Dodd, R.S., Afzal-Rafii, Z., Barbero, M., 2005. Origin identification of Pi-
nus nigra populations in southwestern Europe using terpene composition
variations. Trees: Structure and Function 19, 531e538.
Bytnerowicz, A., Tausz, M., Alonso, R., Jones, D., Johnson, R., Grulke, N.,
2002. Summer-time distribution of air pollutants in Sequoia National
Park, California. Environmental Pollution 118, 187e203.
Bytnerowicz, A., Omasa, K., Paoletti, E., 2006. Integrated effects of air pollu-
tion and climate change on forests: A northern hemisphere perspective.
Environmental Pollution 147 (3), 438e445.
Cape, J.N., 2007. Secondary air pollutants and forestsdnew perspectives. The
Scientific World JOURNAL 7 (S1), 9e14, doi:10.1100/tsw.2007.18.
Ceburnis, D., Sakalys, J., Armolaitis, K., Valiulis, D., Kvietkus, K., 2002. In-
stack emissions of heavy metals estimated by moss biomonitoring method
and snow-pack analysis. Atmospheric Environment 36, 1465e1474.
Emberson, L., 2003. Air pollution impacts on crops and forests: an introduc-
tion. In: Emberson, L., Ashmore, M., Murray, F. (Eds.), Air Pollution
Reviews. Air Pollution Impacts on Crops and Forests: A Global Assess-
ment, Vol. 4. Imperial College Press, London, pp. 3e34.
Farmer, M.A., 1993. The effects of dust on vegetationda review. Environmen-
tal Pollution 79, 63e75.
Campbell, E.S., Taylor, C.A., 2007. Monoterpene production in redberry
juniper foliage following fire. Rangeland Ecology and Management 60,
104e109.
Carisey, N., Bauce, E., 1997. Impact of balsam fir foliage age on sixth-instar
spruce budworm growth, development, and food utilization. Canadian
Journal of Forest Research 27, 257e264.
Chen, Z., Kolb, T.E., Clancy, K.M., 2002. The role of monoterpenes in resis-
tance of Douglas fir to western spruce budworm defoliation. Journal of
Chemical Ecology 28, 897e920.
Da Silva, M.D.R.G., Mateus, E.P., Munha, J., Prazyk, A., Farrall, M.H.,
Paiva, M.R., das Neves, H.J.C., Mosandl, A., 2001. Differentiation of
ten pine species from central Portugal by monoterpene enantiomer-selec-
tive composition analysis using multidimensional gas chromatography.
Chromatographia 53, S412eS416.
De Simon, B.F., Vallejo, M.C.G., Cadahia, E., Miguel, C.A., Martinez, M.C.,
2001. Analysis of lipophilic compounds in needles of Pinus pinea L.
Annals of Forest Science 58, 449e454.
Dormont, L., Roques, A., Malosse, C., 1998. Cone and foliage volatiles emit-
ted by Pinus cembra and some related conifer species. Phytochemistry 49,
1269e1277.
Erisman, J.W., Vermeulen, A., Hensen, A., Flechard, C., Dammgen, U.,
Fowler, D., Sutton, M., Grunhage, L., Tuovinen, J.-P., 2005. Monitoring
and modelling of biosphere/atmosphere exchange of gases and aerosols
in Europe. Environmental Pollution 133, 403e413.
Fuksman, I.L., 2002. Biochemical response of Pinus sylvestris L. to stress
factors. Rastitelnyje Resursy 38 (2), 126e133.
Geron, Ch., Rasmussen, R., Arnts, R.R., Guenther, A., 2000. A review and
synthesis of monoterpene speciation from forests in the United States.
Atmospheric Environment 34 1791e1781.
Grulke, N.E., Paoletti, E., Heath, R., 2007. Chronic vs. short-term acute O3
exposure effects on nocturnal transpiration in two Californian oaks.
The Scientific World JOURNAL 7 (S1), 34e40, doi:10.1100/tsw.
2007.33.
Gunthardt-Goerg, M.S., Vollenweider, P., 2006. Linking stress with macro-
scopic and microscopic leaf response in trees: New diagnostic perspec-
tives. Environmental Pollution 139, 1e22.
Heath, R.L., 2007. Alterations of the biochemical pathways of plants by the air
pollutant ozone: Which are the true gauges of injury? The Scientific World
JOURNAL 7 (S1), 110e118, doi:10.1100/tsw.2007.19.
Helanova, V., Chvilickova, I., Martinkova, M., Meloun, M., Kuban, V., 2006.
Application of multivariate statistical analysis to biological data. Varia-
tions of monoterpene content in fresh needles of Picea abies (L.) Karst.
Chemia Analityczna 51, 0551e0565.
Heller, W., Rosemann, D., Osswald, W.F., Benz, B., Schonwitz, R.,
Lohwasser, K., Kloos, M., Sandermann, H., 1990. Biochemical response
of Norway Spruce (Picea abies (L.) Karst.) towards 14-month exposure
to ozone and acid mist: part Ideffects on polyphenol and monoterpene
metabolism. Environmental Pollution 64, 353e366.
Heyworth, C.J., Iason, G.R., Temperon, V., Jarvis, P.G., Duncan, A.J., 1998.
The effect of elevated CO2 concentration and nutrient supply on carbon-
based plant secondary metabolites in Pinus sylvestris L. Oecologia 115,
344e350.
Huttunen, S., 2005. Light and UV acclimation responses in sub-arctic ever-
greens. Phyton 45 (3), 267e277.
490 E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491

Author's personal copy
Huttunen, S., Manninen, S., 2005. Scots pine and the changing environ-
mentdneedle responses. Polish Botanical Studies 19, 133e141.
Idzojtic, M., Kajba, D., Franjic, J., 2005. Differentiation of F-1 hybrids P. ni-
gra J.F. Arnold x P. sylvestris L., P. nigra J.F. Arnold x P. densiflora
Siebold et Zucc, P. nigra J.F. Arnold x P. thunbergiana Franco and their
parental species by needle volatile composition. Biochemical Systematics
and Ecology 33, 427e439.
Judzentiene, A., Stikline, A., Kupcinskiene, E., 2007. Changes in the essential
oil composition in the needles of Scots pine (Pinus sylvestris L.) under
anthropogenic stress. The Scientific World JOURNAL 7 (S1), 141e150,
doi:10.1100/tsw.2007.34.
Juttner, F., 1988. Changes of monoterpene concentrations in needles of pollu-
tion-injured Picea abies exhibiting montane yellowing. Physiologia Planta-
rum 72, 48e56.
Kainulainen, P., Oksanen, J., Palomaki, V., Holopainen, T., 1992. Effect of
drought and waterlogging stress on needle monoterpenes of Picea abies.
Canadian Journal of Botany 70, 1613e1616.
Kainulainen, P., Holopainen, J., Palomaki, V., Holopainen, T., 1996. Effects of
nitrogen fertilization on secondary chemistry and ectomycorrhizal state of
Scots pine seedlings and on growth of grey pine aphid. Journal of Chem-
ical Ecology 22, 617e636.
Klusia, J., Penuelas, J., Gimeno, B.S., 2002. Seasonal and species-specific re-
sponse of VOC emissions by Mediterranean woody plant to elevated ozone
concentrations. Atmospheric Environment 36, 3931e3938.
Krupa, S.V., Legge, A.H., 2000. Passive sampling of ambient, gaseous air
pollutants: an assessment from an ecological perspective. Environmental
Pollution 107, 31e45.
Kupcinskiene, E., 2001. Annual variations of needle surface characteristics of
Pinus sylvestris growing near the emission source. Water, Air and Soil
Pollution 130, 923e928.
Kupcinskiene, E., 2006. Latent Injuries of Scots Pine (Pinus sylvestris L.)
Under Influence of Local Pollution. Monograph. Kaunas, Lutute, ISBN
9955-692-40-5, 256 p. (in Lithuanian, in English).
Kupcinskiene, E., Huttunen, S., 2005. Long-term evaluation of the needle sur-
face wax condition of Pinus sylvestris around different industries in Lith-
uania. Environmental Pollution 137 (3), 610e618.
Lagalante, A.F., Lewis, N., Montgomery, M.E., Shields, K.S., 2006. Temporal
and spatial variation of terpenoids in eastern hemlock (Tsuga canadensis)in relation to feeding by Adelges tsugae. Journal of Chemical Ecology 32,
2389e2403.
Lavola, A., Aphalo, P.J., Lahti, M., Julkunen-Tiitto, R., 2003. Nutrient avail-
ability and the effects of increasing UV-B radiation on secondary plant
compounds in Scots pine. Environmental and Experimental Botany 49,
49e60.
Lawrence, B.M., 1991. Progress in essential oils. Perfum Flavor 16 (2), 59e67.
Lawrence, B.M., 2001. Progress in essential oils. Perfum Flavor 25, 33e44.
Lawrence, B.M., 2006. Analysis/compositiondpreface. Journal of Essential
Oil Research 18 1e1.
Mankovska, B., 1996. Variations in sulphur and nitrogen foliar concentration
of deciduous and conifers vegetation in Slovakia. Water, Air and Soil Pol-
lution 25, 329e345.
Manninen, S., Huttunen, S., 1995. Scots pine needles as bioindicators of sulphur
deposition. Canadian Journal of Forest Research 25, 1559e1569.
Manninen, A.M., Vuorinen, M., Holopainen, J.K., 1998. Variation in growth,
chemical defense, and herbivore resistance in Scots pine provenances.
Journal of Chemical Ecology 24, 1315e1331.
Manninen, A.M., Tarhanen, S., Vuorinen, M., Kainulainen, P., 2002. Compar-
ing the variation of needle and wood terpenoids in Scots pine provenances.
Journal of Chemical Ecology 28, 211e228.
Manning, W.J., Godzik, B., 2004. Bioindicator plants for ambient ozone in
Central and Eastern Europe. Environmental Pollution 130 (1), 33e39.
Martin, D.M., Gershenzon, J., Bohlmann, J., 2003. Induction of volatile ter-
pene biosynthesis and diurnal emission by methyl jasmonate in foliage
of Norway spruce. Plant Physiology 132, 1586e1599.
Mockute, D., Bernotiene, G., Judzentiene, A., 2003. The b-ocimene chemo-
type of essential oils of the inflorescences and the leaves with stems
from Origanum vulgare ssp. vulgare growing wild in Lithuania. Biochem-
ical Systematics and Ecology 31, 269e278.
Mockute, D., Judzentiene, A., 2004. Composition of the essential oils of
Tanacetum vulgare L. growing wild in Vilnius district (Lithuania). The
Journal of Essential Oils Research 16 (6), 550e553.
Nault, J.R., 2003. Site temperatures influence seasonal changes in terpene
composition in Douglas-fir vegetative buds and current-year foliage. Cana-
dian Journal of Forest Research 33, 2269e2273.
Paoletti, E., 2005. UV-B and Mediterranean forest species: Direct effects and
ecological consequences. Environmental Pollution 137, 372e379.
Paoletti, E., 2006. Impact of ozone on Mediterranean forests: A review. Envi-
ronmental Pollution 144, 463e474.
Petrakis, P.V., Roussis, V., Papadimitriou, D., Vagias, C., Tsitsimpikou, C.,
2005. The effect of terpenoid extracts from 15 pine species on the feeding
behavioural sequence of the late instars of the pine processionary caterpil-
lar Thaumetopoea pityocampa. Behavioural Processes 69, 303e322.
Salla, S.L., Kainulainen, P., Utriainen, J., Holopainen, T., Holopainen, J.K.,
2001. The influence of elevated O3 and CO2 concentrations on secondary
metabolites of Scots pine (Pinus sylvestris L.) seedlings. Global Change
Biology 7, 303e311.
Schleppi, P., Tobler, L., Bucher, J.B., Wyttenbach, A., 2000. Multivariate inter-
pretation of the foliar chemical composition of Norway spruce (Picea
abies). Plant and Soil 219, 251e262.
Schonwitz, R., Merk, L., 1986. Influence of mineral nutrition, ozone and acid
mist on the monoterpene pattern of Picea abies (L.) Karst. Forstw. Cbl 105,
258e261.
Schonwitz, R., Merk, L., Kloos, M., Ziegler, H., 1991. Influence of needle loss,
yellowing and mineral content on monoterpenes in needles of Picea abies
(L.) Karst. Trees 5, 208e214.
Schulz, H., Huhn, G., Hartling, S., 1998. Responses of sulphur- and nitrogen-
containing compounds in Scots pine needles. In: Huttl, R.F., Bellmann, K.
(Eds.), Changes of Atmospheric Chemistry and Effects on Forest Ecosys-
tems. Kluwer Academic Publishers, Great Britain, pp. 37e63.
Shu, C.K., Lawrence, B.M., 1997. Reasons for the variation in composition of
some commercial essential oils. American Chemical Society Symposium
Series 660, 138e159.
Sopauskiene, D., Jasineviciene, D., Stapcinskaite, S., 2001. The effect of
changes in European anthropogenic emissions on the concentrations of sul-
phur and nitrogen components in air and precipitation in Lithuania. Water,
Air and Soil Pollution 130 (1-4), 517e522.
Staszewski, T., Lukasik, W., Godzik, S., Szdzuj, J., Uzieblo, A.K., 1998. Cli-
matic and air pollution gradient studies on coniferous trees health status,
needles wettability and chemical characteristics. Chemosphere 36, 901e
905.
Stikliene, A., Deltuvas, R., Kupcinskiene, E., 2006. Needle asymmetry of
Scots pine (Pinus sylvestris L.) trees growing close to industrial areas.
Miskininkyste (Forestry) 59 (1), 15e21. in Lithuanian.
Supuka, J., Berta, F., 1998. The composition of terpenes in needles of white
pine (Pinus strobus L.) growing in urban environment. Ekologia (Brati-
slava) 17 (4), 419e433.
Supuka, J., Berta, F., Chladna, A., 1997. The Influence of the urban environ-
ment on the composition of terpenes in the needles of Black Pine (Pinus
nigra Arnold). Trees Structure and Function 11, 176e182.
Tiberi, R., Niccoli, A., Curini, M., Epifano, F., Marcotullio, M.C., Rosati, O.,
1999. The role of the monoterpene composition in Pinus spp. needles, in
host selection by the pine processionary caterpillar, Thaumetopoea pityo-
campa. Phytoparasitica 27, 263e272.
Turtola, S., Salla, S.L., Holopainen, J.K., Julkunen-Tiitto, R.,
Kainulainen, P., 2006. Long-term exposure to enhanced UV-B radiation
has no significant effects on growth or secondary compounds of outdoor-
grown Scots pine and Norway spruce seedlings. Environmental Pollution
144, 166e171.
Wagner, M.R., Evans, P.D., 1985. Defoliation increases nutritional quality and
allelochemics of pine seedlings. Oecologia 67, 235e237.
Wieser, G., Manning, W.J., Tausz, M., Bytnerowicz, A., 2006. Evidence for
potential impacts of ozone on Pinus cembra L. at mountain sites in Europe:
An overview. Environmental Pollution 139, 53e58.
Zavala, J.A., Ravetta, D.A., 2002. The effect of solar UV-B radiation on
terpenes and biomass production in Grindelia chiloensis (Asteraceae),
a woody perennial of Patagonia, Argentina. Plant Ecology 161, 185e191.
491E. Kupcinskiene et al. / Environmental Pollution 155 (2008) 481e491




















