
PINUS SYLVESTRIS HEIGHT AND RADIAL GROWTH STRATEGIES IN THE NINE PHYTO-SOCIOLOGICAL ASSOCIATIONS OF...
10
B RAUN-BLANQUETIA, vol. 46, 2010 385 PINUS SYLVESTRIS HEIGHT AND RADIAL GROWTH STRATEGIES IN THE NINE PHYTO-SOCIOLOGICAL ASSOCIATIONS OF TRENTINO-ALTO ADIGE (NORTH-EASTERN ITALY) Christian R OLLAND * , Paolo MINGHETTI ** * 14 bois noir, 38250 - Lans en Vercors (France) E-mail: [email protected] ** Via S. Trinità 14, 38062 - Arco (Italy) E-mail: [email protected] ABSTRACT Scots pine height and radial growth are compared in nine Pinus sylvestris associations of north-eastern Italy (Tren- tino and Alto-Adige). Tree growth and growth dynamics are assessed using stem and tree-ring measurements in 31 Scots pine stands; on the whole 445 trees have been measured and 715 cores have been made and studied. It has been possible to evaluate the growth of the pine in the different plant associations and the site fertility accor- ding to parameters, such as the stem volume calculated at a cambial age of 100 years (V 100 ), which are independent from the tree age. It has been found that, among all associations, the lowest stem volume (mean V 100 = 82 dm 3 at 100 years) and consequently the lowest growth is rea- ched in oligotrophic Sphagnum peat bogs, where Vaccinio uliginosi-Pine- tum sylvestris develops. These sites are unproductive and their major interest is for vegetation preservation, due to the presence of rare plant species. Growth is slightly better (179 dm 3 ) for Molinio coeruleae- Pinetum sylve- stris in meso-eutrophic bogs on hydro- morphic flat soils, and for Chamaecyti- so purpurei-Pinetum sylvestris (143 dm 3 ) in warm sites (extreme southern location) with soil erosion. Higher va- lues are measured for Antherico liliagi- nis-Pinetum sylvestris (213 dm 3 ), esta- blished on dry eroded sites, and for Salici eleagni-Pinetum sylvestris (221 dm 3 ) on river shores with episodic allu- vial events. Volume growth is 245 dm 3 for Erico-Pinetum sylvestris , a climax on dry southern carbonate slopes. The endemic Astragalo vesicarii-Pinetum sylvestris association in dry and xeric inner valleys shows higher stem volu- mes (292 dm 3 ), probably due to longer sunny periods, and a similar tree growth (298 dm 3 ) is found in the Gramineae- rich Pinus association (Molinio litora- lis-Pinetum sylvestris) , on marly slo- pes. Optimal growth values are found in mesophilous Vaccinio vitis-ideae-Pine- tum sylvestris (470 dm 3 ), established on siliceous plateaux or gentle slopes fa- cing north. Only the sites of the two latter associations can be taken into account for timber production. As far as growth dynamics is con- cerned, four different patterns of yearly increase in basal area have been esta- blished: basal area growth curves show a plateau growth for both Chamaecyti- so purpurei- and Molinio coeruleae- Pinetum sylvestris, almost no ageing effect for Erico-, Molinio litoralis-, and Antherico liliaginis-Pinetum sylvestris, a quick start before decrease (pioneer behaviour) for Astragalo vesicarii-, Vaccinio vitis-ideae-, and Salici elea- gni-Pinetum sylvestris , and a linear sta- gnation for Vaccinio uliginosi-Pinetum sylvestris. Therefore, Pinus sylvestris ecolo- gical plasticity enables this species to withstand a huge number of growth conditions by using different radial growth strategies, and leads to large variability in wood production (over a 1 to 5.75 scale according to phytosocio- logy, and a 1-22 scale for individual stands). Furthermore these results sug- gest that growth tables for this conifer could be improved by including infor- mation of site vegetation. KEYWORDS: Pinus sylvestris, dendroeco- logy, forest typology, wood produc- tion, tree growth. RIASSUNTO È stato confrontato l'accrescimen- to radiale e verticale del pino silvestre in nove associazioni vegetali del Tren- tino-Alto Adige. Dallo studio, basato su 193 rilievi fitosociologici (in MINGHET - TI , 2003), 31 rilievi dendrometrici, 445 piante misurate e 715 carote analizzate, è emerso che il pino silvestre assume modelli di crescita e di dinamica della crescita ben definiti e differenziati a seconda delle diverse condizioni ecolo- giche dove si trova a crescere. Sulla base di indicatori di crescita indipendenti dall'età degli alberi, come il volume del tronco calcolato all'età cambiale di 100 anni (V 100 ), risulta che l'associazione tipica delle torbiere a sfa- gni oligotrofiche ( Vaccinio uliginosi- Pinetum sylvestris), mostra i minori valori in assoluto (media V 100 = 82 dm 3 all'età di 100 anni). La crescita degli alberi è migliore (179 dm 3 ) nel Molinio coeruleae-Pinetum sylvestris , associa- zione caratteristica nelle torbiere basse meso-eutrofiche, su suoli gleyformi, e nel Chamaecytiso purpurei-Pinetum sylvestris (143 dm 3 ), associazione pre- alpina di stazioni rupestri carbonatiche con suoli rendziniformi di erosione. Valori di crescita relativamente mag- giori sono espressi dall’Antherico lilia- ginis-Pinetum sylvestris (213 dm 3 ), un'associazione delle ripide e soleggia- te scarpate silicatiche su suoli aridi e soggetti a erosione, e dal Salici eleagni- Pinetum sylvestris (221 dm 3 ), una pine- ta dei greti carbonatici dei torrenti en- doalpini soggetti a periodiche alluvio- ni. Un volume di 179 dm 3 si riscontra nell'Erico-Pinetum sylvestris, la classi- ca pineta a erica endoalpina dei versanti calcarei e dolomitici esposti a sud, su suoli rendziniformi soggetti a rapido prosciugamento. L’Astragalo vesicarii- Pinetum sylvestris, associazione ende- mica delle aride valli endoalpine come la Val Venosta, mostra un volume del tronco leggermente superiore (292 dm 3 ), valore probabilmente da attribuire alla maggiore insolazione delle sue stazio- ni. Simili valori di crescita (298 dm 3 ) si riscontrano nel Molinio litoralis-Pine- tum sylvestris, pineta ricca in gramina- cee su substrati argillosi, mentre i valori ottimali (470 dm 3 ) sono espressi dal Vaccinio vitis-ideae-Pinetum sylvestris, che si insedia in stazioni mesofile su substrati silicatici. Per quanto riguarda la dinamica della crescita invece sono stati definiti quattro differenti modelli di accresci- mento dello spessore degli anelli in funzione dell’età: le curve di incremen- to dell'area basale mostrano un plateau di crescita sia nel Chamecytiso purpu- rei-P.s. sia nel Molinio litoralis-P.s.; non si notano invece variazione nell'in- cremento dell'area basale in funzione dell'età nell'Erico-, Molinio litoralis- e Antherico liliaginis-P.s., mentre l'Astra- galo vesicarii-, il Vaccinio vitis-idae- ae- e il Salici eleagni-P.s . evidenziano
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of PINUS SYLVESTRIS HEIGHT AND RADIAL GROWTH STRATEGIES IN THE NINE PHYTO-SOCIOLOGICAL ASSOCIATIONS OF...

BRAUN-BLANQUETIA, vol. 46, 2010 385
PINUS SYLVESTRIS HEIGHT AND RADIAL GROWTH STRATEGIES IN THE NINE PHYTO-SOCIOLOGICALASSOCIATIONS OF TRENTINO-ALTO ADIGE (NORTH-EASTERN ITALY)
Christian ROLLAND*, Paolo MINGHETTI**
* 14 bois noir, 38250 - Lans en Vercors (France)E-mail: [email protected]** Via S. Trinità 14, 38062 - Arco (Italy)E-mail: [email protected]
ABSTRACT
Scots pineheight and radialgrowthare compared in nine Pinus sylvestrisassociationsofnorth-easternItaly (Tren-tino and Alto-Adige). Tree growth andgrowth dynamics are assessed usingstem and tree-ring measurements in 31Scots pine stands; on the whole 445trees have been measured and 715 coreshave been made and studied.
It has been possible to evaluate thegrowth of the pine in the different plantassociations and the site fertility accor-ding to parameters, such as the stemvolume calculated at a cambial age of100 years (V
100), which are independent
from the tree age.It has been found that, among all
associations, the lowest stem volume(mean V
100= 82 dm3 at 100 years) and
consequently the lowest growth is rea-ched in oligotrophic Sphagnum peatbogs, where Vaccinio uliginosi-Pine-tum sylvestrisdevelops. These sites areunproductive and their major interest isfor vegetation preservation, due to thepresence of rare plant species.
Growth is slightly better (179 dm3)for Molinio coeruleae- Pinetum sylve-stris in meso-eutrophic bogs on hydro-morphic flat soils, and for Chamaecyti-so purpurei-Pinetum sylvestris (143dm3) in warm sites (extreme southernlocation) with soil erosion. Higher va-lues are measured for Antherico liliagi-nis-Pinetum sylvestris (213 dm3), esta-blished on dry eroded sites, and forSalici eleagni-Pinetum sylvestris (221dm3) on river shores with episodic allu-vial events. Volume growth is 245 dm3
for Erico-Pinetum sylvestris , a climaxon dry southern carbonate slopes. Theendemic Astragalo vesicarii-Pinetumsylvestris association in dry and xericinner valleys shows higher stem volu-mes (292 dm3), probably due to longersunny periods, and a similar tree growth(298 dm3) is found in the Gramineae-rich Pinus association (Molinio litora-lis-Pinetum sylvestris), on marly slo-pes. Optimal growth values are found inmesophilousVaccinio vitis-ideae-Pine-tum sylvestris(470 dm 3), established onsiliceous plateaux or gentle slopes fa-
cing north. Only the sites of the twolatter associations can be taken intoaccount for timber production.
As far as growth dynamics is con-cerned, four different patterns of yearlyincrease in basal area have been esta-blished: basal area growth curves showa plateau growth for both Chamaecyti-so purpurei- and Molinio coeruleae-Pinetum sylvestris, almost no ageingeffect for Erico-, Molinio litoralis-, andAntherico liliaginis-Pinetum sylvestris,a quick start before decrease (pioneerbehaviour) for Astragalo vesicarii-,Vaccinio vitis-ideae-, and Salici elea-gni-Pinetum sylvestris, and a linear sta-gnation for Vaccinio uliginosi-Pinetumsylvestris.
Therefore, Pinus sylvestris ecolo-gical plasticity enables this species towithstand a huge number of growthconditions by using different radialgrowth strategies, and leads to largevariability in wood production (over a 1to 5.75 scale according to phytosocio-logy, and a 1-22 scale for individualstands). Furthermore these results sug-gest that growth tables for this conifercould be improved by including infor-mation of site vegetation.
KEYWORDS:Pinus sylvestris, dendroeco-logy, forest typology, wood produc-tion, tree growth.
RIASSUNTO
È stato confrontato l'accrescimen-to radiale e verticale del pino silvestrein nove associazioni vegetali del Tren-tino-Alto Adige. Dallo studio, basato su193 rilievi fitosociologici (in MINGHET-TI, 2003), 31 rilievi dendrometrici, 445piante misurate e 715 carote analizzate,è emerso che il pino silvestre assumemodelli di crescita e di dinamica dellacrescita ben definiti e differenziati aseconda delle diverse condizioni ecolo-giche dove si trova a crescere.
Sulla base di indicatori di crescitaindipendenti dall'età degli alberi, comeil volume del tronco calcolato all'etàcambiale di 100 anni (V100), risulta chel'associazione tipica delle torbiere a sfa-
gni oligotrofiche (Vaccinio uliginosi-Pinetum sylvestris), mostra i minorivalori in assoluto (media V
100= 82 dm3
all'età di 100 anni). La crescita deglialberi è migliore (179 dm3) nel Moliniocoeruleae-Pinetum sylvestris, associa-zione caratteristica nelle torbiere bassemeso-eutrofiche, su suoli gleyformi, enel Chamaecytiso purpurei-Pinetumsylvestris (143 dm3), associazione pre-alpina di stazioni rupestri carbonatichecon suoli rendziniformi di erosione.Valori di crescita relativamente mag-giori sono espressi dall’Antherico lilia-ginis-Pinetum sylvestris (213 dm3),un'associazione delle ripide e soleggia-te scarpate silicatiche su suoli aridi esoggetti a erosione, e dal Salici eleagni-Pinetum sylvestris (221 dm3), una pine-ta dei greti carbonatici dei torrenti en-doalpini soggetti a periodiche alluvio-ni. Un volume di 179 dm3 si riscontranell'Erico-Pinetum sylvestris, la classi-ca pineta a erica endoalpina dei versanticalcarei e dolomitici esposti a sud, susuoli rendziniformi soggetti a rapidoprosciugamento. L’Astragalovesicarii-Pinetum sylvestris, associazione ende-mica delle aride valli endoalpine comela Val Venosta, mostra un volume deltronco leggermentesuperiore (292dm3),valore probabilmente da attribuire allamaggiore insolazione delle sue stazio-ni. Simili valori di crescita (298 dm3) siriscontrano nel Molinio litoralis-Pine-tum sylvestris, pineta ricca in gramina-cee su substrati argillosi, mentre i valoriottimali (470 dm3) sono espressi dalVaccinio vitis-ideae-Pinetum sylvestris,che si insedia in stazioni mesofile susubstrati silicatici.
Per quanto riguarda la dinamicadella crescita invece sono stati definitiquattro differenti modelli di accresci-mento dello spessore degli anelli infunzione dell’età: le curve di incremen-to dell'area basale mostrano un plateaudi crescita sia nel Chamecytiso purpu-rei-P.s. sia nel Molinio litoralis-P.s.;non si notano invece variazione nell'in-cremento dell'area basale in funzionedell'età nell'Erico-, Molinio litoralis- eAntherico liliaginis-P.s.,mentre l'Astra-galo vesicarii-, il Vaccinio vitis-idae-ae- e il Salici eleagni-P.s. evidenziano

386 BENSETTITI A., BIORET F., BOULLET V., PEDROTTI F., Centenaire de la Phytosociologie
una partenza rapida prima di una decre-scita, andamento tipico delle piante pio-niere; la pineta delle torbiere alte oligo-trofiche, il Vaccinio uliginosi-P.s. , mo-stra invece una stagnazione lineare.
In definitiva, l'elevata plasticitàecologica del pino silvestre permette aquesta specie di far fronte a una grandevarietà di condizioni ecologiche adot-tando di volta in volta differenti strate-gie di crescita radiale e, di conseguenza,esprimendo una grande variabilità nellasua produzione in legno (una variabilitàfino a 5,7 volte tra le diverse associazio-ni e fino a 22 volte se si confrontanoinvece le singole aree di campionamen-to). Questi risultati inoltre suggerisco-no che le tabelle di crescita di questaconifera dovrebbero essere sempre cor-redate di informazioni ecologico-vege-tazionali.
INTRODUCTION AND AIMS
Scots pine (Pinus sylvestris L.) is arustic tree species that is able to growover a large range of situations (MIN-GHETTI, 2003), with a vast distributionarea thatextendsfromCatalogna(Spain)to Siberia (Russia). It can face harshgrowing conditions such as sandy xericareas (GUITTET & LABERCHE, 1972), cal-careous soils on steep slopes in dry
inner Alpine valleys (BARTOLI, 1966),sandy soils in Central Europe, marlyslopes in Jura mountains, as well as acidbogs with Sphagnum (ELLENBERG,1988).
This conifer produces wood ofgoodquality (NEPVEU &BLACHON, 1989)and can colonize very different soils,especially sites unfavourable for otherspecies, which explains its great valuefor foresters. Unfortunately, timber pro-duction is often reduced by the lowfertility of siteswhereScots pines usual-ly grow. Most of the time, the betterareas are colonized by species moreresistant to inter-specific competition,except when Scots pine is temporarilyestablished asapioneer specieson aban-doned lands.
Thus, large variations in treegrowth are expected due to the extremeecological plasticity of Scots pine.However, up to now little is knownabout how site conditions influenceScots pine growth, although some re-sults obtained in Finland have clearlyunderlined their importance (VANNINEN
et al., 1996).The purpose of this study, carried
out in Italy, is to investigate height andradial growth of Pinus sylvestris in dif-ferent phytosociological associations,and to determine the relationshipsbetween growth and site conditions.
Our aim is: 1) to quantify wood produc-tion and to discover which are the mostproductive associations; 2) to comparetree growth strategies in very differentsituations.
MATERIAL AND METHODS
SITES AND SAMPLING
The study area was located in theTrentino-Alto Adige region, in north-eastern Italy. Phytosociological studyof Scots pine forests has already beencarried out there (MINGHETTI, 2003)using Braun-Blanquet classical metho-ds, in forests where this species repre-sents more than 80% of the trees. Itcovered the entire region and all growthconditions where Pinus sylvestris canbe established. A vegetation matrix of193 relevés totalizing334differentplantspecies (and 5,569 abundance/domi-nancecoefficients)wasanalysed toesta-blishaforest typology.Ninemainphyto-sociological associations were distin-guished, four on limestone soils andfive on siliceous soils. Complete flori-stic lists were published by MINGHETTI
(2003), with details on classificationresults and soil analyses.
In each of these nine associations,from 2 to 5 stands were sampled (de-pending on the extension of each asso-ciation), with a total of 31 stands (Fig. 1,Tab. 1). The stand density, the develop-ment phase and the disturbance (naturalor anthropic) history were similar andtherefore comparisons were made pos-sible. In particular we chose stands thathave reached a stable mature phasewithout signs of recent human interven-tion.
In each site a complete soil analy-sis was carried out, with soil descrip-tion, granulometry analysis, determi-nation of soil pH at different depths, C,N, CO3--, Fe, Al, Si, and Mn measure-ments (in MINGHETTI, 2003). The domi-nant Scots pines of each site were mea-sured using a Blume-Leiss dendrome-ter for tree height, and with a tape forstem circumference, including bark(CORONA, 1969). By dominant trees wemean the 4 largest trees ha-1 in a multi-layered or unevenaged stand. Incrementcores were taken on those trees at 1.30m above ground using a Pressler borer,with two cores per stem taken in oppo-site directions perpendicularly to theslope line. A total of 445 pines (meanvalue of 15 trees per stand) and 715cores (mean value of 23.1 cores perstand) were studied. Ring widths weremeasured under a binocular microscope
Fig. 1 - Location map of the 31 dendro-sites in the Trentino and Alto-Adige region (northeastern Italy) within 9 phyto-sociological associations.

BRAUN-BLANQUETIA, vol. 46, 2010 387
(magnified by 12 to 50 times) and cross-dated with classical dendroecological te-chniques (SCHUELLER & ROLLAND, 1994).
MEASUREMENTS AND FORMULAE
For each tree the stem volume wascalculatedusingTABACCHI &TOSI(1992)equations, based onPinus sylvestris dia-meterand height values measured in 151forests growing in northern Italy (altitu-des from 380 m to 1,820 m), and inclu-ding the Trentino and Alto-Adige areas:
Stem volume (in dm3) = A + B x D 2
x H + C x D2 x H2
with A=2.8459, B=4.1776*10 -2,C=-2.4970*10-4, D=diameter at 1.30 m(in cm) and H=tree height (in m) (a, b,and c values published by TABACCHI &TOSI, 1992).
The mean dominant tree height,diameter and volume were calculatedfor each site. Similarly, mean valueswere obtained for each phyto-sociolo-gical group by averaging several den-dro-sites.
Site comparisons are only practi-cable when the same tree age is usedeverywhere, for instance at 100 years.We also computed the stem radius at100 years (R100) by adding up the 100
first ring-width values (from stem cen-tre). Tree height at 100 years (i.e. siteindex H100) was derived from a heightgrowth model based on a Chapman-Richards equation (similar to thoseobtained for Spruce trees by Brandini etal. 1994), in the form:
with B1=13.87983, B2=0.03474,and B3=1.536154. (B coefficients adju-sted with our data set). Tree volumes at100 years (V100) were finally obtainedusing TABACCHI & TOSI (1992) equationV100=A+BxD100
2 xH100 +CxD1002xH100
2.In each stand, all ring-widths for-
med on different trees at the same cam-bial age (but at different years) wereaveraged to compute the mean ring-width growth curve as a function of treeage. Then these values were added to-gether to give the radial growth curve ofthe whole stand (SCHUELLER & ROLLAND
1994). For a given forest, the meanring-width <Ck> as a function of cam-bial age (k) was given by:
, by averaging all Scots pine rings atevery given cambial age. Here, Cjk stan-
ds for the ring-width (expressed in 0.01mm) formed by the tree number j whenit was k years old. Thus, k ranges from1 (at pith) to Age j (under bark) for thistree j. Nk is the number of trees thatreach this age k.
Furthermore, themeanradius<Rk>as a function of cambial age was calcu-lated by:
, by cumulating the successive meanring-widths (from pith until a given agek). This value (hereafter called "radialgrowth") does not include bark. Becau-se of geometrical effects (circular ringsget larger), the mean ring-width <Ck >decreases after a few years. That is whya better estimation of the amount ofwood formed each year was given bythe area comprised between two suc-cessive rings, dSk = (Sk - Sk-1) with Sk =.Rk
2. This value dSk is hereafter called"yearly increase in basal area". Theyearly increase in tree height was notanalysed here. In a first step, these cal-culationswereachievedforeachof the31individual stands, and then by gatheringtrees of different locations to characterizetree growth of all stands that belong to thesame phytosociological group.
Tab. 1 - Topography (altitude, slope, aspect), location and bedrock of the 31 dendro-sites.
321 exp1)( BAgeBBageH
k
Nj
jkj
kN
CC
k
1
ki
i
ik CR1

388 BENSETTITI A., BIORET F., BOULLET V., PEDROTTI F., Centenaire de la Phytosociologie
floristic associationsweredistinguished(MINGHETTI, 2003):
- On carbonate bedrocks:1a)The Erico-Pinetumsylvestris Braun-Blanquet in Braun-Blanquet et al. 1939is the inner alpine Spring Heath-Pineforest and represents the most characte-risticassociationoncarbonatebedrocks.Its floristic structure shows a clear basi-philous character: besides Erica herba-cea , the most frequent species are Co-ronila vaginalis , Polygala chamaebu-xus, Gymnadenia odoratissima andCarex alba . This association colonizessouthern steep slopes (30°), especiallyon dolomite bedrock and hard limesto-ne, from 1,000 to 1,600~1,700 m. Thetypical soil is a colluvial rendzina (Ren-dosol and Dolomitosol after AFES,1995, or Calcaric Phaeozems accor-ding to FAO classification, 1990): it issuperficial, with basic pH, rich in car-bonate and shows rapid drainage;1b) The Carex humilis Pinus forest for-ms a sub-association (Erico-Pinetumsylvestris caricicetosum humilis Min-ghetti 2003) during pioneer phases;2) The Chamaecytiso purpurei-Pine-tum sylvestris (Aichinger) Minghetti2003 is the south-alpine Spring Heath-Pine forest that grows in the southernpart of Trentino, in a less continentalclimate than Erico-P.s. It contains somebroadleaf species, like Fraxinus ornus,Ostrya carpinifolia, and other south-alpine or Illyrian species, like Chamae-cytisus purpureus, Asperula purpureaand Cotinus coggygria, not to be foundin the inner Alps. This association de-velops on superficial calcareous soilswith basic pH (Rendosols and Dolomi-tosols after AFES, 1995, or CalcaricPheozems according to the FAO classi-fication of 1990) and in general withcharacteristics very similar to those ofErico-P.s.;3) The Molinio litoralis-Pinetum sylve-stris Schmid ex Etter 1947 is located onmarly slopes with many Graminaceaesuch as Molinia arundinacea, Calama-grostis varia, Brachypodium rupestre,Brachypodium pinnatum and Cypera-cees like Carex flacca. The soils, belon-ging to the family of calcareous brownsoils, are clayey, basic and rich in car-bonate (Calcosols, according to AFES1995, or Calcaric Pheozems accordingto FAO classification, 1990). They arerepeatedly soaked with water althoughin the dry summer the topsoil may dry;4) On fluvial and river shores, withepisodic alluviation processes, the wa-ter balance is strongly contrasted withalternating drought and flooded perio-ds on the topsoil, whereas the water
1955) which explains the considerablevariations in the floristic relevés obser-ved here (MINGHETTI, 2003), as well asalready reported in France (BARTOLI,1966) or in Spain (PAUSAS, 1994). In theTrentino-Alto Adige region, nine main
3. RESULTS
3.1. PHYTO-SOCIOLOGICAL ASSOCIATIONS
Pinus sylvestris L. grows over alarge range of conditions (AGOSTINI,
Fig. 2 - Mean dominant tree height (in m.) for 31 Pinus sylvestris stands divided into 9phytosociological groups, and average values (horizontal lines) for each group.
Fig. 3 - Mean dominant tree diameter (in cm; with bark and at a height of 1,30 m) for 31Pinus sylvestris stands divided into 9 phytosociological groups, and average values(horizontal lines) for each group.
Fig. 4 - Mean dominant tree volume (whole stem, in dm3) calculated for 31 Pinus sylvestrisstands according to 9 phytosociological groups using H and D measurements. Averagevalues (horizontal lines) for each group.
BRAUN-BLANQUETIA, vol. 46, 2010 389
vestris Braun-Blanquet 1961 can befound in the dry areas of the Venostavalley, where average rainfalls do notreach 500 mm in a year, and it is cha-racterized by more or less endemic spe-cies of inner alpine dry valleys such asAstragalus vesicarius ssp. pastellianusand Oxytropis halleri ssp. velutina. Thisassociation develops between 1,000 to1,600 m a.s.l., on 20-40° slopes onmica-schist bedrocks. The soils (satura-ted Colluviosols after AFES 1995, orEutric Regosols after FAO, 1990) areweakly acid superficially and neutral-basic in depth;8) The Vaccinio uliginosi-Pinetum syl-vestris Kleist 1929 can be found inoligotrophic peat bogs with Sphagnumand Vaccinium uliginosum, V. oxycoc-cos, Eriophorum vaginatum, Carexpauciflora, Polytrichum strictum, Dro-sera rotundifolia and Andromeda poli-folia. It is established in only a few sitesfrom 1,300 to 1,600 m a.s.l., on Histo-sols sphagnofibriques (AFES, 1995) or
Fibric Histosols (FAO, 1990) with acidpeat;9) The Molinio coeruleae-Pinetum syl-vestris (Hofman) Passarge 1978 - Min-ghetti etPedrotti1999develops inmeso-eutrophic peat bogs and other wet siteson hydromorphic flat soils. Reductisolsstagniques and histosols mesiques eu-trophes (AFES, 1995) are typical soils,(or Eutric Gleysols and Eutric Histosolsaccording to FAO classification, 1990)and show a slightly acid pH. The under-storey is characterized by Molinia coe-rulea, Carex fusca, Carex canescens,Cirsium palustre, and Succisa praten-sis.
DENDROMETRICAL RESULTS
For all pine stands (31 sites), theaverage dominant tree height is 13.3 ±4.7 m, with extreme values rangingfrom 5.9 m to 27.6 meters, (Fig. 2). Theaverage value for the dominant tree
reserve remains quite constant at lowerdepths.Deeply rootedhygrophilousspe-cies (such as various willow species andAlnus incana) co-occurred with super-ficially rooted drought-tolerant species.Scots pine is there a temporary pioneerspecies and forms the Salici eleagni-PinetumsylvestrisOberdorfer1957sub-association ericetosum herbaceae. Itranges from 1,300 to 1,500 m a.s.l. andis characterized by Salix eleagnos, S.purpurea, S.glabra, Calamagrostis epi-gejos, C. pseudophragmites, Hieraciumstaticifolium, H. florentinum, and bySalix nigricans and Pinus mugo. Thesoils of this association are very youngand undeveloped, gravely, very poor infine particles and carbonate-rich. Theybelong mainly to Fluvisols (Fluviosols,afterAFES 1995, or Calcari-leptic Flu-viosols, after FAO classification, 1990).
- On siliceous bedrocks, five other asso-ciations were distinguished:5) The most extended association onthis type of substrate is Antherico lilia-ginis-Pinetum sylvestris Mayer ex Min-ghetti 2003, which is characterised bythe presence of Carex humilis in theherb layer, together with Silene rupe-stris, Veronica officinalis, Polypodiumvulgare, Campanula rotundifolia, An-thericum liliago, mosses such as Rhyti-dium rugosum and Hypnum cupressi-forme, and several Sedum and Semper-vivum species. Antherico liliaginis-Pi-netum sylvestris colonises steep slopesfacing south where strong soil erosionand important colluvial processes arepresent. The soils, which may belong tocolluvial soils or to acid brown soils(ColluviosolsandAlocrisolsafterAFES1995; Eutric or Dystric Regosols, andDystric Cambiosols according to FAOclassification, 1990), are sandy and con-tinuously remixed by colluvial proces-ses and therefore are young and unde-veloped. They are oligosaturated andpH varies from weak acid to acid;6) In the underwood of Vaccinio vitis-ideae-Pinetum sylvestris Mayer et Ho-fmann 1969 Vaccinium myrtillus, V.vitis-idaeae, Pyrola media, Avenellaflexuosa and Rhododendron ferrugi-neum dominate. This association deve-lops at an altitude comprised between800 and 1,600 m a.s.l., on plateau andnorth facing slopes made of granite,quartzite, and rhyolithe. The soil is of apodzolic type (Podzosols ocriques andAlocrisols ocreux, after AFES 1995;Haplic or Cambic Podzols and Spodo-Dystric Cambisols after FAO, 1990),highly developed, from acid to veryacid and desaturated;7) TheAstragalo vesicarii-Pinetum syl-
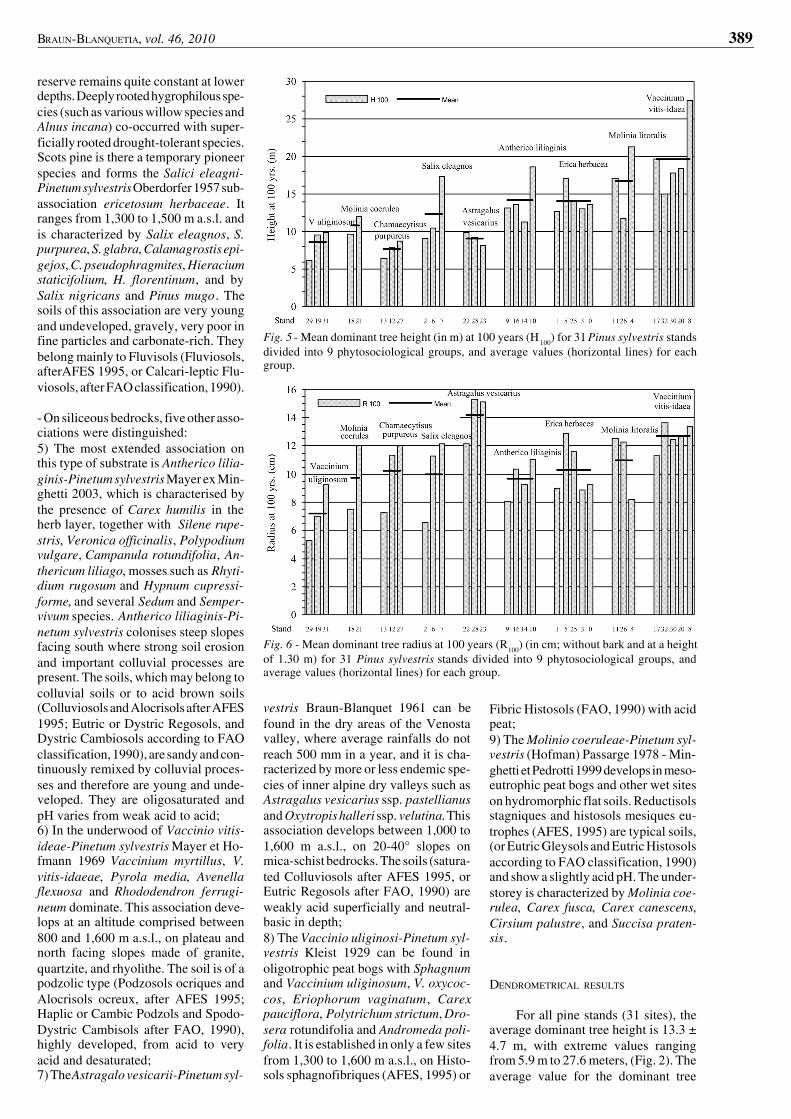
Fig. 5 - Mean dominant tree height (in m) at 100 years (H 100) for 31 Pinus sylvestris standsdivided into 9 phytosociological groups, and average values (horizontal lines) for eachgroup.
Fig. 6 - Mean dominant tree radius at 100 years (R100) (in cm; without bark and at a heightof 1.30 m) for 31 Pinus sylvestris stands divided into 9 phytosociological groups, andaverage values (horizontal lines) for each group.

390 BENSETTITI A., BIORET F., BOULLET V., PEDROTTI F., Centenaire de la Phytosociologie
used and is adopted in this chapter.More reliablecomparisons arepre-
sented in the following chapters on thebasis of tree volume and height calcula-ted at the 100-years threshold.
As expected, the lowest dominanttree height values are observed in thetwo kinds of bogs; in the Vaccinio uligi-nosi-Pinetum sylvestris (8.5 m) and inthe Molinia coerulea-P.s. stands (9.9m) (Fig.2).Similarly, treeheightgrowthappears strongly reduced in the Cha-maecytiso purpurei-Pinetum sylvestris ,with a mean tree height of 7.6 m only, aswell as in the Astragalo vesicarii-P.s.(9.4 m in average). Comparatively, hei-
ght growth appears to be better in theSalici eleagni-Pinetum sylvestris (12.3m), along riverbeds deposit on baresoils. The larger variability is probablyassociated with soil depth and watersupply differences within sites.
The last four phyto-sociologicalassociations appear more valuable fortimber production, since both the An-therigo liliaginis-P.s. and the Erico-Pinetum sylvestris are characterized bymoderately high growth values (14.7 mand 14.9 m respectively). The highesttrees are found in the Molinio litoralis-Pinetum sylvestris (17.2 m), establishedon marly slopes and more especially inthe Vaccinio vitis-ideae-Pinetum sylve-stris that appears to be the optimum forScots pine height growth in that region(19.8 m in average, with values up to27.6 m).
In addition to height growth, radialgrowth was also examined, despite thefact that stem diameter is more influen-ced by tree age compared to dominanttree height values (Fig. 3). The moststriking result is the opposition betweenheight and radial growth for the Astra-galovesicarii-Pinetum sylvestris. In thisparticular association, pine verticalgrowth is poor, but it is largely compen-sated by a sharply enhanced radialgrowth. The largest stems are foundhere, with a mean diameter of 42.4 cmthat even exceeds those measured in theoptimal growth conditions (42.1 cm inthe Molinio litoralis-P.s. and 38.3 cm inthe Vaccinio vitis-ideae-P.s.).
Tree volume is calculated usingTABACCHI & TOSI (1992) equations ba-sed on diameter and height measure-ments. This parameter is a direct esti-mation of wood production, and there-fore provides valuable information forsite comparisons (Fig. 4). Five growthlevels are distinguished:1) Extremely low wood production hasbeen recorded in both Vaccinio uligi-nos-Pinetum sylvestris and Molinio co-eruleae-P.s. associations, which corre-spond with unproductive sites (respec-tively 229 and 262 dm3). Hence themajor interest of such forest types, un-suitable for timber production, is forvegetation preservation due to the pre-sence of rare plant species.2) Very low wood production is obser-ved in the Chamaecytiso purpurei-P.s.and the Salici eleagni-P.s. (respecti-vely 280 and 370 dm 3), both with poorinterest for wood production. In thelater one, Scots pine plays an importantpart for soil formation and impedeserosion processes.3) Intermediate growth values weremeasured in the Astragalo vesicarii-
diameter is 33.8 ± 7.75 cm, with extre-me values ranging from 14.9 cm to 46.2cm (Fig. 3). The average stem volume is659.8 ± 360.0 dm3, with extreme valuesranging from 58.1 dm3 to 1,428.9 dm3
(Fig. 4).
HEIGHT, DIAMETER, AND VOLUME GROWTH
Height and volume of the domi-nant trees are measurements commonlyused by foresters as indicators of treegrowth. This method is not always re-liable, because height and volume arealso affected by tree age, but is largely
Fig. 7 - Mean tree volume (in dm3) at 100 years (V100) calculated for 31 Pinus sylvestrisstands according to 9 phytosociological groups using H100 and R100 values. Average values(horizontal lines) for each group.
Fig. 8 - Mean tree-ring width (expressed in 0.01 mm) as a function of cambial age for allScots pines of the same phytosociological group.

BRAUN-BLANQUETIA, vol. 46, 2010 391
juvenile phase of ring formation, evenwhen they grow in the same stand. Forthat reason we have also investigatedhere the dynamism of ring formationwith cambial age (i.e. the age of the treewhen the ring was formed).
MEAN RING-WIDTH AS A FUNCTION OF CAM-BIAL AGE
Ring-widths variation with cam-bial age usually show an optimum whentrees are quite young, followed by agradual decrease for most pioneer spe-cies such as pines (BRÄKER, 1981;SCHUELLER & ROLLAND, 1994; PETITCO-LAS et al., 1997). In all stands the tree'slargest ring was formed at an age ran-ging from 3 to 10 years (Fig. 8). For allpines, regardless of site, this maximumring-width reaches 2.21 mm at 4 yearsold. Striking differences appear amongthe nine phyto-sociological groups. Thestands characterized by Astragalus ve-sicarius show the highest peak (3.15mm), above stands withErica herbacea(2.94 mm). On the contrary, in Vacci-nium uliginosum stands the largest rin-gs only reach 1.26 mm.
Another sharp contrast is visibleamong the different kinds of forest:most types show, after the initial phase,an exponential decline of ring widthwith increasing age, whereas theVacci-
nio vitis-idaeae-P.s. differs by a quitelineardecline,which is thereforeslower.
The mean ring-width at 100 yearsis 0.66 mm for all trees and ranges from0.37 mm in Salix eleagnos stands to0.75 mm in Vaccinium vitis-idaea stan-ds.
DYNAMICS OF RADIAL GROWTH
Such variations in ring formationpatterns according to forest typologylead to important differences in radialgrowth curves (Fig. 9). The nine mainradial growth curves are given here, toprovide a guideline to foresters. Notethat radial growth patterns in Vacci-niumuliginosum standswerequiteweakand linear, whereas the highest non-linearity was observed in Astragalusvesicarius stands.
YEARLY INCREASE IN BASAL AREA
The curves of increase in basalarea appear to be a highly efficientindicator for the understanding of thedifferences in stand growth dynamism.Four main patterns were distinguishedin these growth patterns (Fig. 10).
Type A, that includes the Chama-ecytiso purpurei-Pinetum sylvestrisandthe Molinio coeruleae-P.s. is characte-
P.s. and in the Antherigo liliaginis-P.s.(respectively 685 and 661 dm3). Theformer is characterized by short butlarge stems, whereas the second onepresents tree shapes more suitable fortimber production.4) Moderately productive sites are loca-ted in the Erico-Pinetum sylvestris (811dm3).5) Highly productive sites are located inboth the Molinio litoralis-P.s.and in theVaccinio vitis-ideae-P.s., with tree vo-lumes of 1,046 and 1,078 dm3 respecti-vely. In the later case, Pinus sylvestrisoften competes with Picea abies, whe-reas Spruce is virtually absent in theMolinio litoralis-P.s.
COMPARISONS AT 100 YEARS
Since the Scots pines do not haveexactly the same ages in the differentstands, tree growth was also analysed ata given cambial age (100 years). Thistechnique allows one to eliminate theinfluence of age on tree growth analy-sis. The results obtained for tree heightat 100 years (H100) (Fig. 5) appear to behighly consistent with those previouslyobtained with dominant tree height Hdom(Fig. 2), despite the effect of age hetero-geneity observed for Hdom. These twovalues show a strong correlation (H
domand H100: r = 0.991 n = 32), confirmingthat dominant tree height is an efficientpredictor of site fertility (PARDÉ & BOU-CHON, 1988). It does not depend on treeage, but levels off when trees are old.Moreover, the height growth thresholdonly depends on site fertility.
Contrarily, tree radius at 100 years(R100) obtained with ring-widths (Fig.6) differs from results obtained withstem diameter measurements (Fig. 3)due tobothage influenceandbarkwidth.As already observed with the heightvalues, the use of the tree volume at 100years (V100) (Fig. 7) was found moreefficient to compare inter-site produc-tion than the use of the tree volume (V)of dominant pines (Fig. 4). Considera-tions about comparisons at 100 yearsare made in the discussions and in theconclusions of this paper.
GROWTH DYNAMICS
Some previous studies about coni-fer growth (PETITCOLAS et al., 1997) hadrevealed that roughly similar values intree volumes could in fact hinder im-portant differences in tree growth pat-terns. For instance, Pinus cembra andPinus uncinata strongly differ in their
Fig. 9 - Mean tree radius (in cm) as a function of cambial age for all Scots pines of the samephytosociological group (derived from ring-width measurements, without bark). Thesquares indicate the value at 100 years.

392 BENSETTITI A., BIORET F., BOULLET V., PEDROTTI F., Centenaire de la Phytosociologie
TI, 1988). Pinus sylvestris is abundant inthe Centro-oriental area of the Alps (inTrentino and Alto-Adige, and in someparts of the Lombardian Alps) and, to alesser extent, in the western part (Pie-monte) (TABACCHI & TOSI, 1992). Ac-cording to the national forest survey(Inventario Forestale Nazionale), pinespecies cover 228,600 ha in Italy (TA-BACCHI & TOSI, 1992). As in neighbou-ring countries (DÉCOURT, 1965; OTTORI-NI & NYS, 1981a-b; VANNIÈRE, 1984;OFFICE NATIONAL FORÊTS, 1994; CLUZE-AU et al ., 1998), several growth tables(CASTELLANI, 1970) were established inItaly for Scots pine wood production:forVallediSusa (QUAGLINO etal., 1987),for Trentino-Alto Adige (CORONA &FERRARA, 1987), as well as for whole ofNorthern Italy (TABACCHI & TOSI, 1992).
Nevertheless,atourknowledge thisstudy is the first one that takes intoaccount the part played by site variabi-lity (described by forest phytosociolo-gy) on pine growth in Italy, and thatpoints out different ring-width growthpatterns.
Our sampling strategy covered theentire ecological range of Pinus sylve-stris in the Trentino-Alto Adige region;thus, allowing us to quantify the plasti-city of that species. From stand to stand,thevariations of the dendrometricalme-asurements are particularly high, sincea 3.1 times variability in dominant tree
diameter is reported here (Fig. 3), whe-reas the values of dominant tree heightvary up to 4.7 times (Fig. 2); this givesrise to considerable variation in stemvolume, over a 1 to 24.6 scale (Fig. 4).Regardless of the influence of ageing,results at a cambial age of 100 years(more valuable for site comparisons)lead to similar conclusions. Thus, stemdiameterD
100(withoutbark) fallsbetwe-
en 10.6 and 30.6 cm (variations over a2.9 times interval), tree height H
100va-
ries from 6.2 to 27.4 meters (showing a4.4 times change), and volume V
100falls
between 30.7 and 688.6 dm3 (i.e. itsvariations cover a 22.4 times interval).Such extreme high variability in treegrowth clearly underlines the extremerange of growth conditions that Pinussylvestris can tolerate.
Very smallheightgrowth (H100lessthan 10 m) is reported in the two kind ofbog communities (Vaccinio uliginosi-Pinetum sylvestris and Molino coerule-ae-P.s.), due to the particularly unfa-vourable growth conditions, characte-rized by insufficient drainage that cau-ses root asphyxia (KAUNISTO& TUKEVA,1986). Meso-eutrophic conditions inbogs on hydromorphic flat soils (withMolinia coerulea) lead to a slightlyenhanced treegrowth compared to moreoligotrophic conditions encompassedby pines in the acid peat bogs withSphagnum (+ 20% in height growthH
100, and + 54% for V
100). These findin-
gs are consistent with KAUNISTO (1987)results in peat sites, where P and Kfertilisation enhanced Pinus sylvestrisheight growth.
Surprisingly, the lowest heightgrowth is not recorded in bog sites, butin the Chamaecytiso purpurei-Pinetumsylvestris (H
100= 7.7 m). However, lar-
ger diameters partly compensate thisphenomenon. Volume growth there, isintermediatebetween the two bog types.Moreover, this association is the onlyone where some stands are infected bymistletoe (Viscum album). This is espe-cially the case of stands at lower altitu-des (mean altitude 852 m for the 6infected sites, versus 948 m for the 27uninfected stands of the same associa-tion where floristic relevés were done),facing S and SW, on lower slopes (24.2°versus 33.1°), and characterized by ahigher shrub cover (41% versus 29%).Such mistletoe infection may be consi-dered as an indication of tree waterstress. This is corroborated by the extre-me location of this association, develo-ped in the southern part of Trentinowith many thermophilous deciduousspecies such as Fraxinus ornus, Ostryacarpinifolia, and Quercus pubescens
Fig. 10 - Mean increase in basal area (cm2 / year) as a function of cambial age for all Scotspines of the same phytosociological group (derived from ring-width measurements,without bark).
rized by a rapid increase extending du-ring the 25 first years of tree growth,followed by a roughly constant plateau(about 3.5 cm2/year and 3.0 cm2/yearrespectively).
Type B, with 3 types of forests(Erico-Pinetum sylvestris, Molinio lito-ralis-P.s., and Antherico liliaginis-P.s.)does not show such a leveling off, buton the contrary presents a continuousyearly growth increase in basal areawith tree age. In such a case, tree sene-scence seems to be delayed, and Scotspine remains able to form more andmore wood as they get older.
Type C, with Astragalo vesicarii-P.s., Vaccinio vitis-idaeae-P.s. and Sa-lici eleagni-P.s. types, shows a maxi-mum in basal area yearly incrementfollowed by a decline. Thus, the maxi-mums are reached at 38 years for theSalix type (4.59 cm2 /year), at 44 yearsfor the Astragalus type (8.3 cm2/year),and at 68 years for the Vaccinium type(7.61 cm2/year).
Type D, for Vaccinio uliginosi-P.s. is characterized by a slow and line-ar trend with a continuous increase.
DISCUSSION
In Italy, pure Scots pine forestsgrow in the whole of the alpine area,especially in the inner valleys (PIGNAT-

BRAUN-BLANQUETIA, vol. 46, 2010 393
quick start during the juvenile phase.Canopy closure may be responsible forbasal area growth decline after 68 yearsin the Vaccinio vitis-idaeae-P.s., due tothe competition of Picea abies and La-rix decidua.
CONCLUSIONS
As hypothesized, Scots pine woodproduction shows considerable varia-tions depending on the phyto-sociolo-gical groups. By increasing order, thestem volumeat 100 years (without bark)(V
100) reaches only 82 dm3 in the Vacci-
nio uliginosi-Pinetum sylvestris, 143dm3 in the Chamaecytiso purpurei-P.s.,and 179 dm3 in the Molinia coerulea-P.s. Such extremely low pine growthseems to be caused by insufficient drai-nage in both Vaccinio uliginos-P.s. andMolinio coeruleae-P.s., leading to rootasphyxia and aggravated by soil oligo-trophy. Contrarily, in the Chamaecyti-so purpurei-P.s., soil erosion processesseem responsible for such poor treevigour, by reducing soil water storagecapacity (pines are confined in extremesites). Intermediate values are found forthe Antherico liliaginis-P.s. (213 dm3),for the Salici eleagni-P.s. (221 dm3),and for the Erico-Pinetum sylvestris(245 dm3). The large variability in theSalici eleagni-P.s. is probably associa-ted with differences in soil water tabledepths according to the sites. Highervalues were measured in both the Astra-galo vesicarii-P.s. (292 dm3) and theMolinio litoralis-P.s. (298 dm3). In theformer group, a high sunshine durationin the Val Venosta inner valley may beresponsible for better photosynthesis,in spite of high thermic contrast thatleads to water stress. Contrarily, thelater group seems favoured by marlysoils that provide sufficiently high wa-ter reserve. Timber production reachesits top in the Vaccinio vitis-idaeae-P.s.(470 dm3), where shade species such asNorway Spruce are able to establish,and probably eliminate pioneer Scotspines.
These values cover a 5.75 timevariation among the phyto-sociologicalgroup, whereas values for individualstands (that range from 31 to 689 dm3)cover a 22-time interval.
Our results clearly point out andquantify the part played by site condi-tions for Scots pine growth, a specieswith an extreme ecological plasticity.Thus,growth tables for thisconifercouldbe improved by including informationof site vegetation.
Moreover, important differences
according to the phytosociology are de-monstrated here in the radial growthstrategies, since four ring-width patter-ns are distinguished:- Both Chamaecytiso purpurei-P.s. andMolinio coeruleae-P.s. show "plateaugrowth";- The Erico-Pinetum sylvestris, Moli-nio litoralis-P.s. and Antherico liliagi-nis-P.s. show "no ageing effect";- The Astragalo vesicarii-P.s., Vacci-nio vitis-idaeae-P.s. and Salici eleagni-P.s. are characterized by "pioneer beha-viour" (i.e. a quick start followed by adecrease);- In acid bogs, (Vaccinio uliginosi-P.s.)a "linear stagnation" is observed.
Such unexpected results indicatethat, even though Pinus sylvestris cansurvive in a huge number of variousgrowth conditions, this species behavesspecifically in each of its phyto-socio-logical associations.
ACKNOWLEDGEMENTS
This research was supported bythe Centro di Ecologia Alpina (Viottedel Monte Bondone, I-38040 Trento).We also thank prof. R. Michalet (Bor-deaux, France) and prof. F. Pedrotti(Camerino, Italy) for their support.
REFERENCES
A.F.E.S., 1995 - Référentiel pédologi-que. Ed. INRA, Paris.
ANTONOROLLI R., BAGANARESI U., FER -RARI C., SPERANZA M., 1985 - Inda-gini su alcuni caraterri morfologi-ci del Pino silvestre (Pinus sylve-stris L.) in popolamenti spontaneinell’Appenino Emiliano. Ann.Accad. Ita. Sci For., 34: 151-160.
AGOSTINI R., 1955 - Le pinete di pinosilvestre in Italia. Atti CongressoNazionale Selvicoltura, vol 1, Ac-cad. It. Sc. For., Firenze.
BARTOLI C., 1966 - Études écologiquessur les associations forestières dela Haute-Maurienne. Ann. Sci.For., 23(3): 321 p.
BRÄKER O.U., 1981 - Der Alterstrendbei Jahrringdichten und Jahrring-breiten von Nadelhölzern und seinAusgleich. Mitt. Forst. Bund., 142:75-102.
BRANDINI P., CANTIANI M.G., FLORIS A.,TABACCHI G., 1994 - Curve di svi-luppo dell'altezza dominante perle formazioni di quota dell'abeterosso della valle di Fiemme. [Do-minant height growth curves forSpruce stands in Valle di Fiemme
(in the extreme sites).Poor growth is found in the Astra-
galo vesicarii-P.s., due to the extremeclimatic factors of the dry inner valleys.In fact, the climate of Val Venosta, thatrepresents the distribution centre of thisassociation, is characterizedby lowrain-fall amounts and high temperature con-trasts.For instance,yearly rainfallequals473 mm/year in Silandro (at 706 m),whereas the evapotranspiration is 664mm/year (GAFTA & PEDROTTI, 1996).However, this endemic association re-veals an unusualbehaviour patterncom-pared to all other Scots pine forests.Although height growth is particularlylow (H100 = 9.9 m), stem diameters arethe largest (D
100= 28.4 cm) compared to
results reported in all other stands, evenin the most productive ones. Conse-quently, tree volume (V100 = 292 dm3) ishigh, despite a tree shape characterizedby an extremely low H/D ratio, unfa-vourable for timber production. Suchtree form may be due to the presence ofa genetically different ecotype ofPinussylvestris there, as already reported byANTONOROLLI et al. (1985) in the Emi-lia-Romagna Appenines, and generallyaccepted for this species (ZOLLER, 1981).
The Molinio litoralis-Pinetum syl-vestris and the Vaccinio vitis-idaeae-Pinetum sylvestris appear to be the bestphytosociological types for Scots pinegrowth. This species is therefore able todevelop with high wood production onmarly calcareous soils as well as on acidsoils (schistes and rhyolithes), regard-less soil chemistry.
Such large ability of Scots pine tocolonize different kinds of substratesappears to be associated with differen-ces in ring formation patterns. Unfortu-nately, until now, such phenomenonhave only received little attention. Themaximum ring-width is found at veryyoung cambial tree age, that is in agre-ement with the pioneer behaviour ofPinus sylvestris. This species is therefo-re characterized by rapid early growth.Moreover, four main patterns of radialgrowth curves are distinguished in ourstudy. The slow linear increase in basalarea with cambial age (in the Vacciniouliginosi-P.s.) may be interpreted as asurvival strategy of trees facing soilflooding, and therefore requiring manyyears for root establishment. Such afeature is also visible for the pines esta-blished in the Molinio litoralis-P.s., alsogrowing on flooded soil, and for theAntherico liliaginis-P.s. sites with con-tinuous soil erosion. On the other hand,pioneer behaviour of pines may be in-ferred from the growth curve reportedin the Astragalo vesicarii-P.s., with a

394 BENSETTITI A., BIORET F., BOULLET V., PEDROTTI F., Centenaire de la Phytosociologie
(Trentino, Italy)]. ComunicazioniRicerca Istituto Sperimentale As-sestamento Forestale Alpicoltura(ISAFA): 94(2): 3-33.
CASTELLANI C., 1970 -Tavole stereome-tricheedalsometrichecostruiteperi boschi italiani. Annali IstitutoSperimentale Assestamento Fore-stale Alpicoltura, Trento, 1, n.s.:431 p.
CLUZEAU C., ULRICH E., LANIER F., GAR-NIER F., 1998 - Interprétation desmesures dendrométriques de 1991à 1995 des 102 peuplements duréseau.Ed.OfficeNational Forêts,Département Recherches Techni-ques, RENECOFOR, pp. 309.
CORONA E., 1969 - Ricerche sullo spes-sore corticale del pino silvestre.L'ItaliaForestaleeMontana,24(6):293-298.
CORONA P., FERRARA A., 1987 - Ricer-che dendrometriche sul pino silve-stredelTrentino-AltoAdige.Montie Boschi 38(6): 51-54.
DÉCOURTN., 1965 - Le pin sylvestre et lepin laricio de Corse en Sologne.Ann. Sci. For. 22(2): 259-317.
ELLENBERG H., 1988 -Vegetation ecolo-gy of Central Europe. CambridgeUniversity Press, Cambridge.
F.A.O.-UNESCO, 1990 - Soil map ofthe world. Revised legend. WorldSoil Resource Report, 60.
GAFTAD., PEDROTTIF., 1996 - Fitoclimadel Trentino-Alto Adige. StudiTrentini di Scienze Naturali - ActaBiologica, 73: 55-100.
GUITTET J., LABERCHE J.C., 1972 - L’im-plantation naturelle du pin sylve-stre sur pelouse xérophile en forêtde Fontainebleau. Bul. Soc. fr.Ecol. 4: 435-447.
KAUNISTO S., TUKEVA J., 1986 - Kasva-tustiheyden vaikutus männyn istu-tustaimikoiden kehitykseen turve-mailla. [Effect of tree spacing onthe development of pine planta-tions on peat]. Folia Forestalia,646, Institutum Forestale Fenniae:1-36.
KAUNISTO S., 1987 - Lannoituksen jamuokkauksen vaikutus männyn jaraudukoivun istutustaimien kasvu-un suonpohjilla. [Effect of fertili-sation and soil preparation on thedevelopment of Scots pine and sil-ver Birch plantations on peat cu-tover areas]. Folia Forestalia, 681,Institutum Forestale Fenniae: 1-23.
MINGHETTI P., 2003 - Le pinete a Pinussylvestris del Trentino-Alto Adige(Alpi italiane). Tipologia, ecolo-gia e corologia. Braun-Blanque-tia, 33: 1-95.
NEPVEU G., BLACHON J.L., 1989 - Lar-geur de cerne et aptitude à l'usageen structuredequelquesconifères:douglas, pin sylvestre, pin mariti-me, épicéa de Sitka, épicéa com-mun, sapin pectiné . Rev. For. Fr.6, XLI: 497-506.
OFFICE NATIONAL FORÊTS, 1994 - Guidede sylviculture du pin sylvestre enforêt de Hagenau. ONF DirectionRégionale Alsace, pp. 42.
OTTORINI J.M., NYS C., 1981 - Applica-tion des données de l'inventaireforestier national à l'étude de laproductiondupinsylvestreenMar-geride. A - Étude de la croissanceen hauteur. Ann. Sci. For. 38(2):223-236.
OTTORINI J.M., 1981 - Application desdonnées de l'inventaire forestiernational à l'étude de la productiondu pin sylvestre en Margueride. B- Étude de la production totale envolume. Ann. Sci. For. 38(4): 487-502.
PARDÉ J., BOUCHON J., 1988 - Den-drométrie. Ed. ÉcoleNationaledesEaux et Forêts, Nancy, 2de ed., pp.468.
PAUSAS J.G., 1994 - Species richnesspatterns in the understory of Pyre-neanPinus sylvestris forest. J.Veg.Sci., 5: 517-524.
PETITCOLAS V., ROLLAND C., MICHALET
R., 1997 - Croissance de l’épicéa,du mélèze, du pin cembro et du pinà crochets en limite supérieure dela forêt dans quatre régions desAlpes françaises. Ann. Sci. For.,54(8): 731-745.
PIGNATTI S., 1988 - I boschi d'Italia,sinecologia e biodiversità . UTET,Torino.
QUAGLINO A., NOSENZO A., MARCHISIO
C., DURANTE S., 1987 - Tavola dicubatura a doppia entrata del pinosilvestre cresciuto in fustaia coe-tanea prevalentemente pura del-l'Alta valle di Susa. Monti e Bo-schi, 38(4): 39-42.
SCHUELLER J., ROLLAND C., 1994 - In-fluence stationnelle sur la crois-sance du pin à crochets (Pinusuncinata Ramond). Schweizeri-sche Zeitschrift für Forstwesen,145(9): 739-755.
TABACCHI G., TOSI V., 1992 - Modelli diprevisione delle masse legnose delPino silvestrenell'Italia settentrio-nale. [Volume equations of Scotspine in Northern Italy] . Comuni-cazioni Ricerca Istituto Sperimen-tale Assestamento Forestale Al-picoltura (ISAFA), 92(1): 1-30.
VANNIÈRE B., 1984 - Tables de produc-tion pour les forêts françaises. Ed:
ENGREF, Nancy CNRF, INRA,2de ed., pp. 159.
VANNINEN P., YLITALO H., SIEVÄNEN R.,MÄKELÄ A., 1996 - Effects of ageand site quality on the distributionof biomass in Scots pine (Pinussylvestris L.). Trees, 10: 231-238.
ZOLLER H., 1981 - Genus Pinus. In:Hegi G. (ed.), Illustrierte Flora vonMitteleuropa, Bd. 1, Teil 2, Parey,Berlin.




















