
The Discriminative Stimulus Effects of the Neurotensin NTS 1 Receptor Agonist PD149163 in Rats:...
26
Discriminative stimulus effects of PD149163 1 RUNNING HEAD: Discriminative stimulus effects of PD149163 The discriminative stimulus effects of the neurotensin NTS 1 receptor agonist PD149163 in rats: Stimulus generalization testing with dopamine D 1 and D 2 receptor ligands. Adam J. Prus*, Candice J. Schuck, Kristoffer R. Rusch, and Lawrence M. Carey Department of Psychology, Northern Michigan University, Marquette, MI *Corresponding author Department of Psychology Northern Michigan University 1401 Presque Isle Ave Marquette, MI 49855 Email: [email protected] Phone: 906-227-2941 Fax: 906-227-2954 Funding Experiment 1 of this study was supported by an NIH grant to AJP (1R15MH083241). Pages and Figures Abstract + Intro + Methods + Results + Discussion + References = 18 pages Figures = 7 figures
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The Discriminative Stimulus Effects of the Neurotensin NTS 1 Receptor Agonist PD149163 in Rats:...

Discriminative stimulus effects of PD149163 1
RUNNING HEAD: Discriminative stimulus effects of PD149163
The discriminative stimulus effects of the neurotensin NTS1 receptor agonist PD149163 in rats: Stimulus
generalization testing with dopamine D1 and D2 receptor ligands.
Adam J. Prus*, Candice J. Schuck, Kristoffer R. Rusch, and Lawrence M. Carey
Department of Psychology, Northern Michigan University, Marquette, MI
*Corresponding author
Department of Psychology Northern Michigan University 1401 Presque Isle Ave Marquette, MI 49855 Email: [email protected] Phone: 906-227-2941 Fax: 906-227-2954
Funding
Experiment 1 of this study was supported by an NIH grant to AJP (1R15MH083241).
Pages and Figures
Abstract + Intro + Methods + Results + Discussion + References = 18 pages
Figures = 7 figures
Copyright Wiley-Blackwell. DOI: 10.1002/ddr.21171

Discriminative stimulus effects of PD149163 2
ABSTRACT
Brain-penetrant neurotensin NTS1 receptor agonists produce antipsychotic drug-like effects in
animal models, including inhibition of conditioned avoidance responding and reversal of psychostimulant-
induced hyperactivity and stereotypy. Allosteric interactions between NTS1 receptors and dopamine D2
receptors may account for some of these antipsychotic effects. In order to determine the role that
dopamine receptors may play in the behavioral effects produced by activation of NTS1 receptors, a drug
discrimination approach was used in rats to evaluate the potential mediation of NTS1 receptor agonist
stimulus effects by dopamine D1 and D2 receptors. Rats were trained to discriminate either the NTS1
receptor agonist PD149163, the D1 receptor agonist SKF81297, or the D2 receptor agonist quinpirole from
vehicle in a two choice drug discrimination task. Full stimulus generalization occurred from PD149163 to
the typical antipsychotic drug and D2 receptor-preferring antagonist haloperidol. However, stimulus
generalization did not occur from SKF81297 or quinpirole to PD149163. The discriminative cue for
SKF91297 and quinpirole was fully blocked the D1 receptor antagonist SCH23390 and the D2/3 receptor
antagonist raclopride, respectively. Cross generalization did not occur between SKF91297 and quinpirole.
Based on these findings, the antipsychotic effects of PD149163 may be mediated, in part, through D2
receptor antagonism, but this may only be evident when PD149163 is used as the training drug.
KEYWORDS: Neurotensin, PD149163, quinpirole, SKF81297, raclopride, SCH23390, antipsychotic,
drug discrimination

Discriminative stimulus effects of PD149163 3
INTRODUCTION
Neurotensin is a neuropeptide neurotransmitter that has been evaluated as a pharmacological
target for treating schizophrenia [Tanganelli et al. 2012], anxiety disorders [Prus et al. 2014],
neurocognitive impairment [Azmi et al. 2006; Feifel et al. 1999], and drug addiction [Fredrickson et al.
2005]. The behavioral effects of neurotensin are primarily derived from activation of neurotensin NTS1
receptors, and the emergence of brain-penetrant NTS1 receptor agonists, particularly including NT69L
[Cusack et al. 2000] and PD149163 [Petrie et al. 2004], has stimulated dozens of studies on the putative
therapeutic efficacy of these drugs for mental disorders. NTS1 receptor agonists have consistently
engendered atypical antipsychotic-like effects in animal models, including an ability to attenuate
psychotomimetic-induced deficits in pre-pulse inhibition [Feifel et al. 1999; Shilling et al. 2003], inhibit
conditioned avoidance responding without eliciting catalepsy [Holly et al. 2011a], reverse
psychostimulant-induced hyperactivity and stereotypy [Cusack et al. 2000; Feifel et al. 2008], and
preferentially enhance dopamine concentrations in the prefrontal cortex compared to the nucleus
accumbens [Prus et al. 2007]. There is also emerging evidence that NTS1 receptor agonists have
anxiolytic effects based on the ability of systemically administered PD149163 to reduce fear-potentiated
startle responses [Shilling and Feifel 2008] and footshock-conditioned 22 kHz vocalizations [Prus et al.
2014] in rats, and that NTS1 receptor agonists may treat nicotine addiction, based upon the ability of
NT69L to reduce nicotine self-administration in rats [Boules et al. 2011].
While neurotensin NTS1 receptors have been found postsynaptically on GABA, serotonin, and
cholinergic neurons, these receptors are particularly located on dopamine neurons, including postsynaptic
localization on ventral tegmental area and substantia nigra dopamine neurons and presynaptic
localization on dopamine terminals in nucleus accumbens, prefrontal cortex, and striatum [Binder et al.
2001; St-Galais et al. 2006]. Neurotensin is also synthesized and released from mesocorticolimbic and
nigrostriatal dopamine neurons. When neurotensin binds to presynaptic NTS1 receptors, these receptors
inhibit D2 autoreceptor functioning through either an allosteric mechanism or by reducing the ability the
receptor to be bound by dopamine [Binder et al. 2001; St-Galais et al. 2006; Tanganelli et al. 2012].
Inhibition of D2 autoreceptors causes increased dopamine efflux and activation of postsynaptic receptors
of the D1 or D2 receptor family [Beaulieu and Gainetdinov 2011]. The findings that the NTS1 receptor is

Discriminative stimulus effects of PD149163 4
associated with three different G-proteins (Gq/11, Gi/o, and Gs) and that selective activation of any of these
G-proteins may occur depending on a ligand’s binding site on the receptor provides an additional facet of
complexity for understanding the mediation of behavioral effects by NTS1 receptor agonists [Tanganelli et
al. 2012].
Drug discrimination procedures have long been used to evaluate pharmacological actions that
may mediate a drug’s interoceptive stimulus effects [Porter and Prus 2009]. The procedure has a
remarkable degree of specificity for the stimulus effects elicited by a training drug (i.e., discriminative
stimulus effects, in this context) as indicated by stimulus generalization selectively occurring from the
training drug to compounds that elicit these same effects and by blockade of these discriminative stimulus
effects by compounds with opposing pharmacological actions (e.g., an antagonist). The results from
generalization testing highly correspond to the shared receptor binding affinities of the training and test
compounds.
The present study sought to determine the role that dopamine D1 and D2 receptors may play in
the stimulus effects of a NTS1 receptor agonist by using a two-choice drug discrimination procedure in
rats to assess stimulus generalization occurring between the NTS1 receptor agonist PD149163 and
compounds selective for dopamine D1 or D2 receptors. The first experiment employed PD149163 as the
training drug in one group of rats and the second experiment employed a D1 receptor agonist and a D2
receptor agonist as training drugs in separate groups of rats. The study’s findings provide some tentative
support for mediation of PD149163’s discriminative stimulus effects by D2 receptor antagonism.
METHODS AND MATERIALS
Experiment 1
Subjects
Ten male Sprague Dawley rats (Charles River, Portage, Michigan, USA) obtained at
approximately 90 days old were used and were housed in a temperature and humidity controlled vivarium
under a 12 hour light/dark schedule. Rats were individually housed and with free access to water, but with
restricted access to food in order to maintain 85% of free-feeding weight. All procedures were approved
by the Northern Michigan University Institutional Animal Care and Use Committee and followed the Guide
for the Care and Use of Laboratory Animals [2011].

Discriminative stimulus effects of PD149163 5
Apparatus
Six rat operant chambers equipped with two retractable levers, a food receptacle, and 45mg
pellet dispenser were used for all drug discrimination procedures (Med-Associates Inc., St. Albans,
Vermont, USA). A lever was located equidistantly on each side of the food pellet receptacle. The
chambers were housed in sound-attenuating cubicles with fans for ventilation and masking noise. The
chambers were controlled and data were recorded using Med-PC version 4 software (Med-Associates
Inc.).
Drugs
PD149163 (NIMH Chemical Synthesis and Drug Supply Program, National Institute of Mental
Health, Bethesda, Maryland, USA) and haloperidol HCl (Tocris, Ellisville, Missouri, USA) were
administered in 1.0 ml/kg volume. PD149163 and haloperidol were in salt form. PD149163 was
dissolved in sterile water and haloperidol HCl was dissolved in sterile water with a few drops of lactic acid.
The doses were determined based upon behaviorally effective doses used in previous studies [e.g., Feifel
et al. 1999; Holly et al. 2011b; Prus et al. 2004]. PD149163 was injected subcutaneously and haloperidol
was injected intraperitoneally 30 minutes prior to a session.
Drug Discrimination Training
After lever press training to fixed ratio (FR) 20 reinforcement schedule, 5 consecutive training
sessions were conducted with vehicle and with only the vehicle-appropriate lever present. Following this,
consecutive training sessions were conducted with PD149163 and with only the PD149163-lever present.
This initial phase of PD149163 lever training was planned for 5 training sessions, but response inhibition
by PD149163 led to lowering the training dose during this period (see results for experiment 1).
Following single-lever training sessions, two-lever drug discrimination training began. Vehicle (V)
and PD149163 (D) training days were assigned based on a single-double alternation pattern (i.e.,
VDVVDVDDV etc.). During two-lever training, every lever press reset the FR20 counter for the opposite
lever. Subjects were tested when the following criteria were met for 5 out of 6 consecutive training
sessions: 1) first FR20 occurring on the condition-appropriate lever, 2) 80% or greater condition-
appropriate responding during the first FR20, 3) 80% or greater condition-appropriate responding for the
total session, and 4) at least 0.1 responses per second for the session.

Discriminative stimulus effects of PD149163 6
Before every test session, subjects had to meet the above four criteria for both a vehicle and a
PD149163 training session. Test sessions ended after an FR20 occurred on either lever; otherwise, the
session ended after 20 minutes. No food pellet was delivered during or after a test session. As with
training sessions, every lever press reset the FR20 counter on the opposite lever.
Data Analysis
Percent PD149163-lever responding was expressed as a percentage based on the number of
PD149163-appropriate responses emitted divided by the total number of PD149163- and vehicle-
appropriate responses emitted. In order to be included in the percent PD149163-appropriate responding
calculation for a test session, rats were required to emit at least 10 responses. Percent PD149163-lever
responding and responses per second were reported as means +/- the standard error of the mean (SEM).
Full stimulus generalization from the PD149163 discriminative stimulus was defined as 80% or greater
drug-appropriate responding and partial stimulus generalization was defined as 60-80% drug-appropriate
responding. Statistically significant differences in response rates across doses and vehicle control were
determined using a repeated measures one way analysis of variance (ANOVA). Statistically significant
differences were followed by a Dunnett’s post hoc test for comparison to vehicle control. A nonlinear
regression analysis was used to calculate the ED50 dose (+/- 95% confidence intervals [CI]) for percent
PD149163-appropriate responding for drugs that produced full stimulus generalization. All analyses were
conducted using GraphPad Prism for Windows, version 5.02 (GraphPad Software, San Diego, California,
USA).
Experiment 2
Animals
A total of 20 male Sprague Dawley rats were used in experiment 2, with 10 animals assigned per
group. The housing procedures were identical to those described in experiment. All procedures in
experiment 2 were approved by the IACUC at Northern Michigan University and followed the Guide to the
Care and Use of Laboratory Animals [2011].
Drugs
The drugs assessed in experiment 1 were the D1 receptor agonist SKF81297 (0.15-1.2mg/kg;
generously provided by the NIMH Drug Repository), D2 receptor agonist quinpirole hydrochloride (0.0125-

Discriminative stimulus effects of PD149163 7
0.1mg/kg; NIMH Drug Repository), the D1 receptor antagonist SCH23390 (0.001875-0.015mg/kg; Sigma-
Aldrich), the D2 receptor antagonist raclopride (0.2-0.8mg/kg; Sigma-Aldrich), and PD149163 (0.0030125-
0.15mg/kg; NIMH Drug Repository). All drugs were in salt form and every drug was dissolved in saline,
which served as the vehicle in this experiment. Drug injections were given subcutaneously 30 min prior to
training or testing sessions. Doses were chosen based on previous studies [Gleason and Witkin 2006;
Reavill et al. 1993; Weathersby and Appel 1986].
Apparatus
The equipment and data collection procedures were identical to those described for experiment 1.
Drug Discrimination Training and Testing
Two groups of rats (n=10 each) were used for experiment 1, with one group of rats trained to
discriminate the D1 receptor agonist SKF81297 (0.3 mg/kg) from vehicle and the other group of rats
trained to discriminate the D2 receptor agonist quinpirole (0.025 mg/kg) from vehicle. Rats were trained
on a fixed ratio 30 reinforcement schedule, but otherwise, training and testing procedures were identical
to experiment 1.
Data Analysis
In order to be included in the percent drug-lever responding calculation for a test session, rats were
required to emit at least 15 responses. Otherwise these data analysis procedures were identical to
experiment 1.
RESULTS
Experiment 1
Training
The initial training dose of PD149163, 0.5mg/kg, completely abolished responding during single
lever PD149163 training sessions. A subsequent reduction to 0.25 mg/kg was less disruptive but still
precluded adequate responding for training. These first failed attempts to identify a non-disruptive
PD149163 training dose led to a short series of dose and route adjustments until a 0.0625 mg/kg doses,
administered, was utilized. Six of the ten rats met the training criteria for the 0.0625 mg/kg training dose.
In total, 121 mean training sessions were required to meet the training criteria. Poorly maintained
performance at the training criteria limited stimulus generalization testing to only PD149163 and

Discriminative stimulus effects of PD149163 8
haloperidol. Three of the four remaining rats were subsequently trained using a 0.03125 mg/kg dose and
met training criteria after 212 mean training sessions. However, these three rats failed to maintain the
training criteria, and therefore stimulus generalization data are unavailable for these subjects.
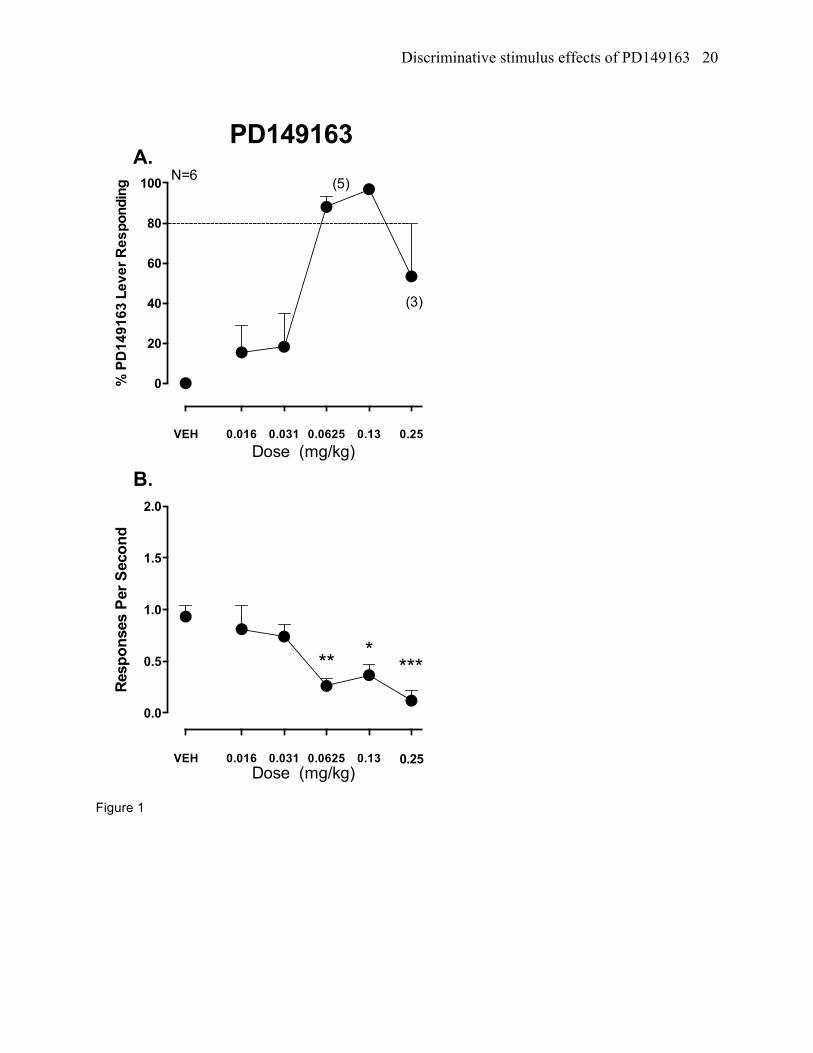
PD149163 testing in PD149163 trained rats
PD149163 produced full stimulus generalization to itself at the 0.0625 mg/kg training dose as well
as the 0.125 mg/kg dose (ED50= 0.04 mg/kg, 95% confidence-interval (CI)= 0.03- 0.05 mg/kg; figure 1a).
Full stimulus generalization did not occur to the 0.25 mg/kg dose, but this was largely influenced by one
rat (out of only three responding at this dose) that selected the vehicle lever. A statistically significant
reduction in response rates was shown (F[5,25]= 7.63, p<0.001; figure 1b). Multiple comparisons testing
revealed that these reductions occurred at the 0.0625, 0.125, and 0.25 mg/kg doses compared to vehicle.
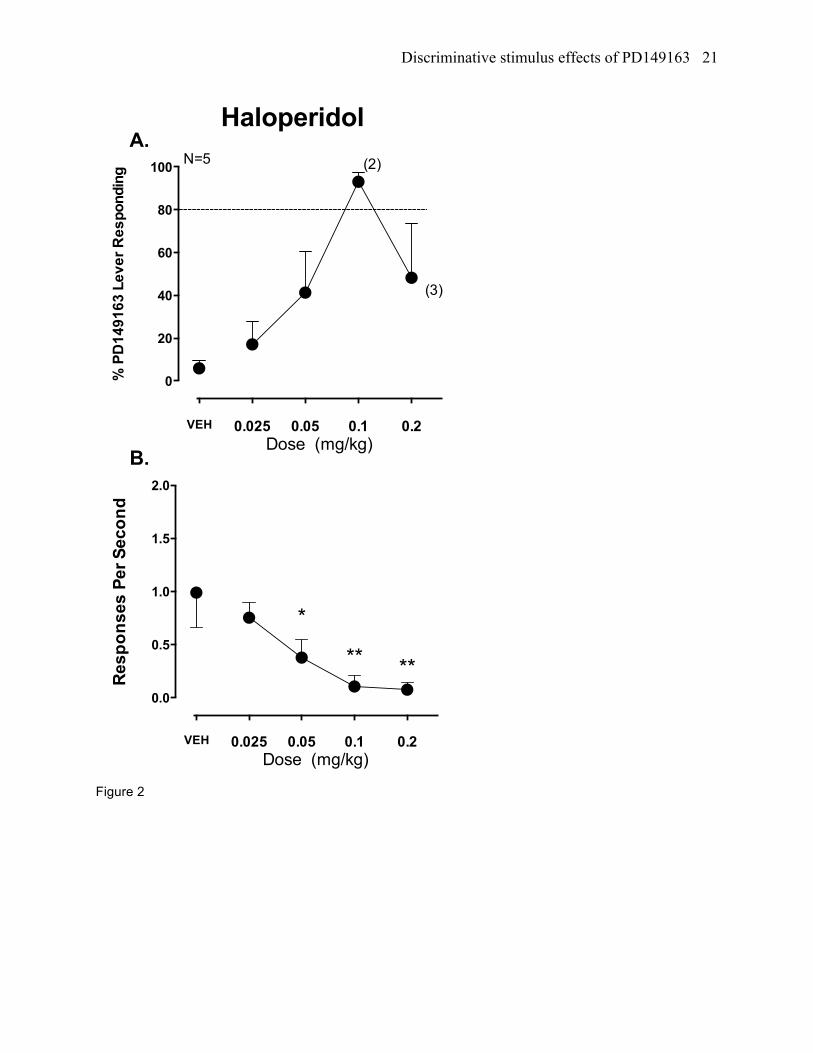
Haloperidol testing in PD149163 trained rats
Full stimulus generalization occurred to the 0.1 mg/kg dose of haloperidol (ED50 = 0.05 mg/kg,
95% CI= 0.03-0.08 mg/kg; figure 2a). Haloperidol significantly reduced response rates (F[4,16]= 6.41,
p<0.01; figure 2b) at the 0.05, 0.1, and 0.2 mg/kg doses compared to vehicle. Due to rate suppression,
two of five rats emitted enough responses to be included in the percent PD149163-appropriate
responding calculation for the 0.1 mg/kg dose and three of five rats were included in the calculation for
the 0.2 mg/kg dose.
Experiment 2
Training
The mean number of training sessions needed to meeting the training criteria was 42.6 sessions
in the group of rats trained to discriminate SKF81297 from vehicle and 26.0 sessions in the group of rats
trained to discriminate quinpirole from vehicle. Four rats from each group failed to meet the training
criteria within 200 training sessions and were removed from the study, leaving six rats each group.
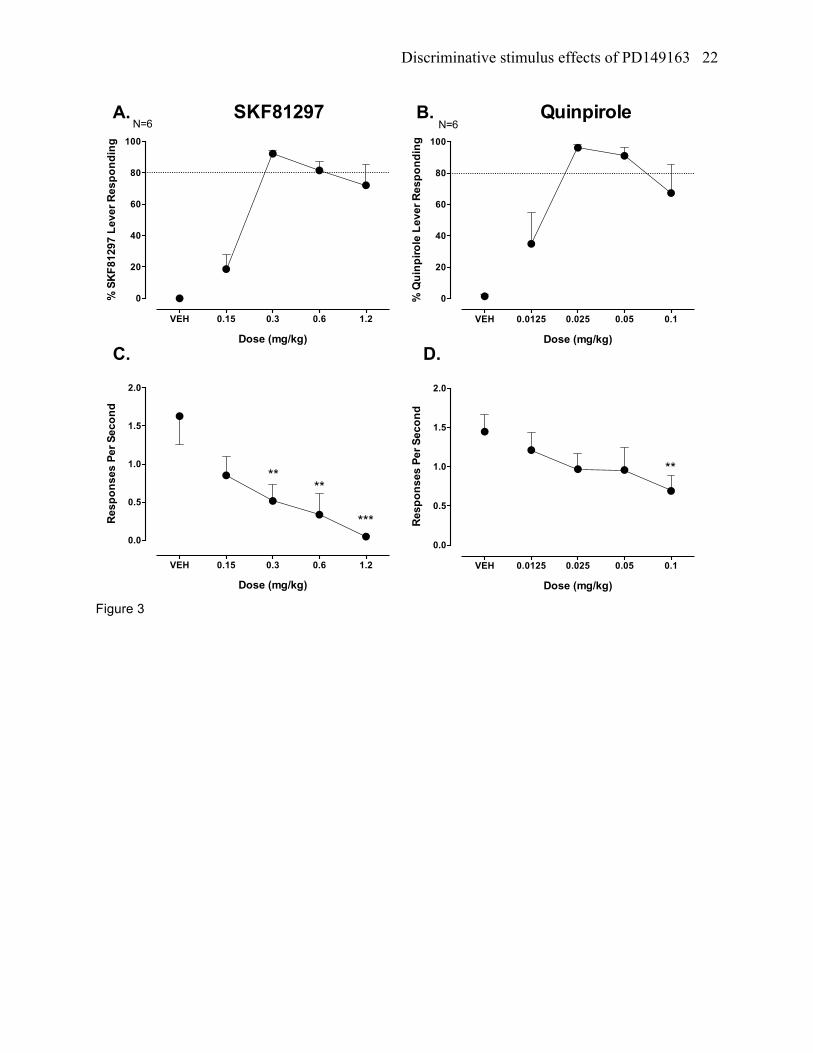
SKF81297 testing in SKF81297 trained rats
SKF81297 (0.15 to 1.2 mg/kg) led to full stimulus generalization to a 0.3 and 0.6 mg/kg dose in
rats trained to discriminate a 0.3 mg/kg dose of SKF81297 from vehicle, while partial stimulus
generalization occurred to a 1.2 mg/kg dose (ED50 = 0.2 mg/kg, 95% confidence interval [CI] = 0.12 to
0.34 mg/kg; figure 3a). SKF81297 also produced a statistically significantly reduction in response rates

Discriminative stimulus effects of PD149163 9
(F[4,20]=7.02, p<0.01), which was found for a 0.3, 0.6, and 1.2 mg/kg dose compared to vehicle (figure
3c).
Quinpirole testing in quinpirole trained rats
Quinpirole (0.0125 – 0.1 mg/kg) produced full stimulus generalization to a 0.025 and 0.05 mg/kg
dose (ED50=0.007mg/kg, 95% CI = 0.002 to 0.018 mg/kg) and produced partial stimulus generalization to
a 0.1 mg/kg dose, although 4 out of 6 of the rats produced greater than 90% responding on the quinpirole
lever (figure 3b). Quinpirole also produced a significant decrease in response rates (F[4,20]=3.72,
p<0.05), which occurred at the 0.1 mg/kg dose compared to vehicle (figure 3d).
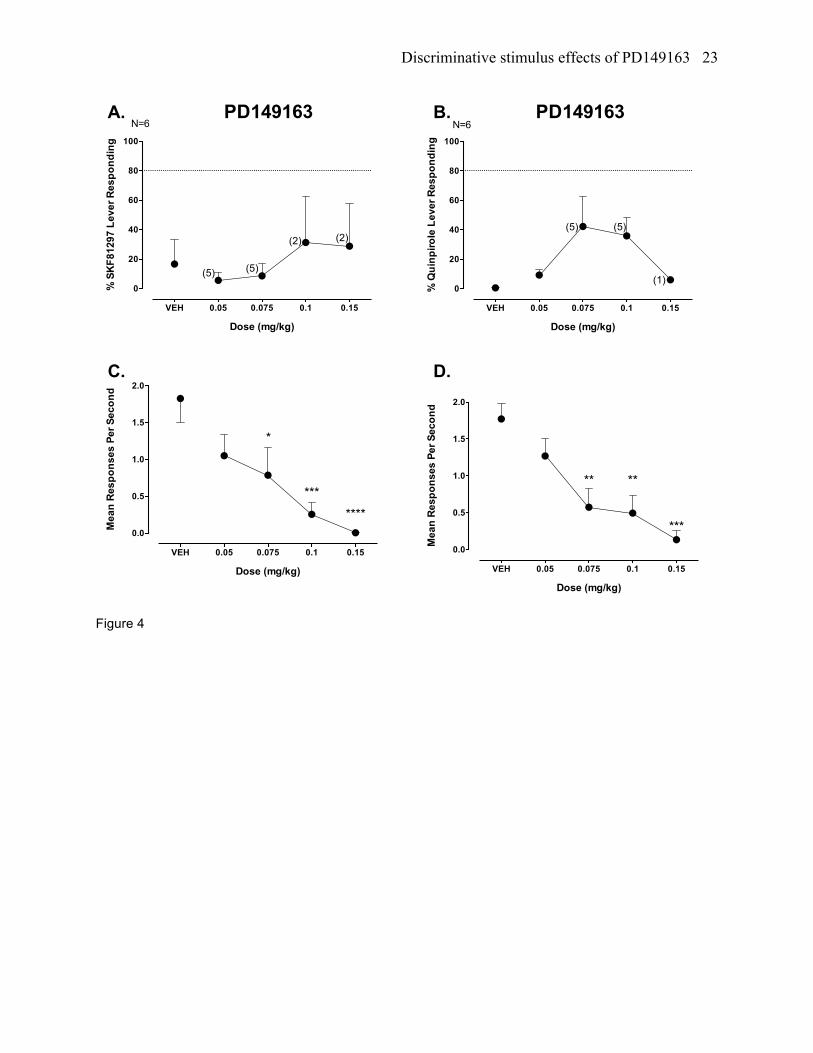
PD149163 testing in SKF81297 trained rats
PD149163 did not produce stimulus generalization from SKF81297, with the highest level of
generalized responding (32.15%) occurring to a 0.1 mg/kg dose (figure 4a). No animals produced full
generalized responding on the SKF81297 lever for any of the doses tested. PD149163 significantly
reduced response rates (F[4,20]=9.37, p<0.0001), which occurred at a 0.075, 0.1 and 0.15 mg/kg dose
compared to vehicle (figure 4c).
PD149163 testing in quinpirole trained rats
PD149163 neither engendered partial nor full generalized responding from quinpirole with the
highest level of generalized responding (42.24%) occurring to a 0.075 mg/kg dose (figure 4b). Two of the
five rats that responded at the 0.075 mg/kg dose emitted full generalized responding on the quinpirole
lever. PD149163 significantly reduced response rates (F[4,20]=8.82, p<0.0001), which occurred at a
0.075, 0.1 and 0.15 mg/kg dose compared to vehicle (figure 4d).
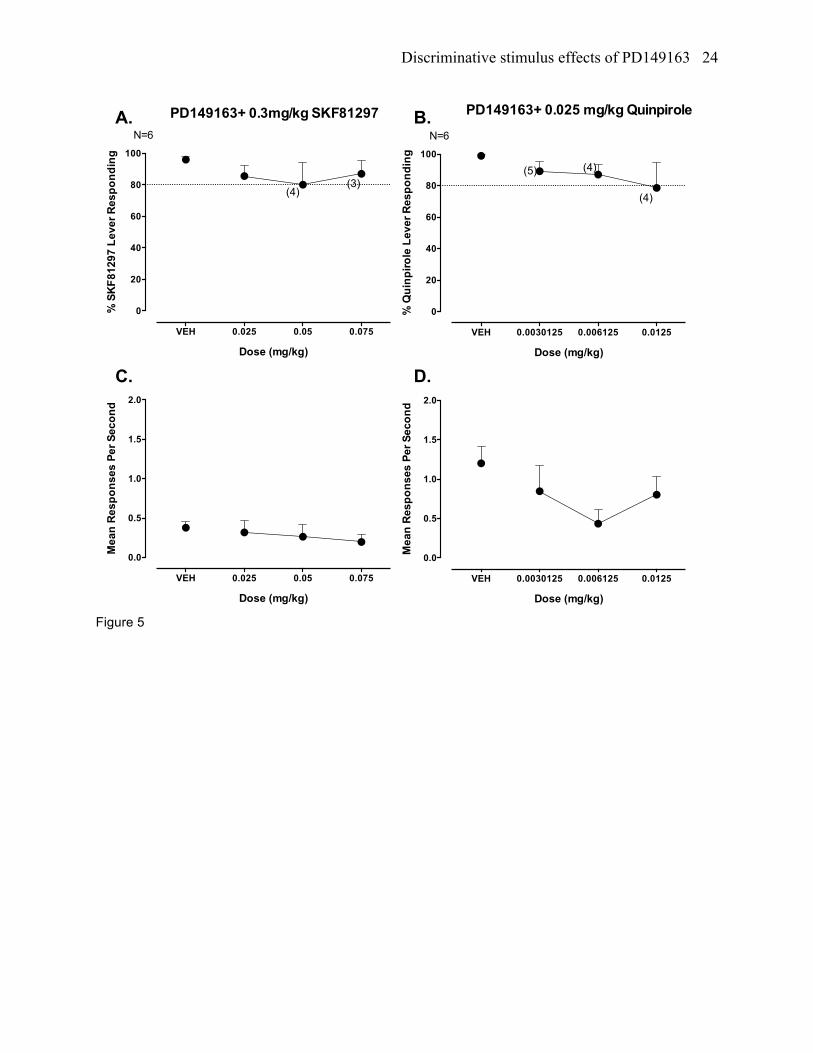
PD149163 + SKF81297 testing in SKF81297 trained rats
PD149163 did not produce partial or full blockade of the SKF81297 discriminative stimulus (figure
5a). Response rates did differ across the dose combinations of PD149163 and SKF81297 (figure 5c).
PD149163 + quinpirole testing in quinpirole trained rats
PD149163 did not fully block the quinpirole discriminative stimulus, although a 0.0125 mg/kg
dose of PD149163 did reduce quinpirole lever responding (78.6%) to slightly below the 80% criterion for
full stimulus generalization (figure 5b). Combination testing with these compounds did not lead to
statistically significant changes in response rates (figure 5d).

Discriminative stimulus effects of PD149163 10
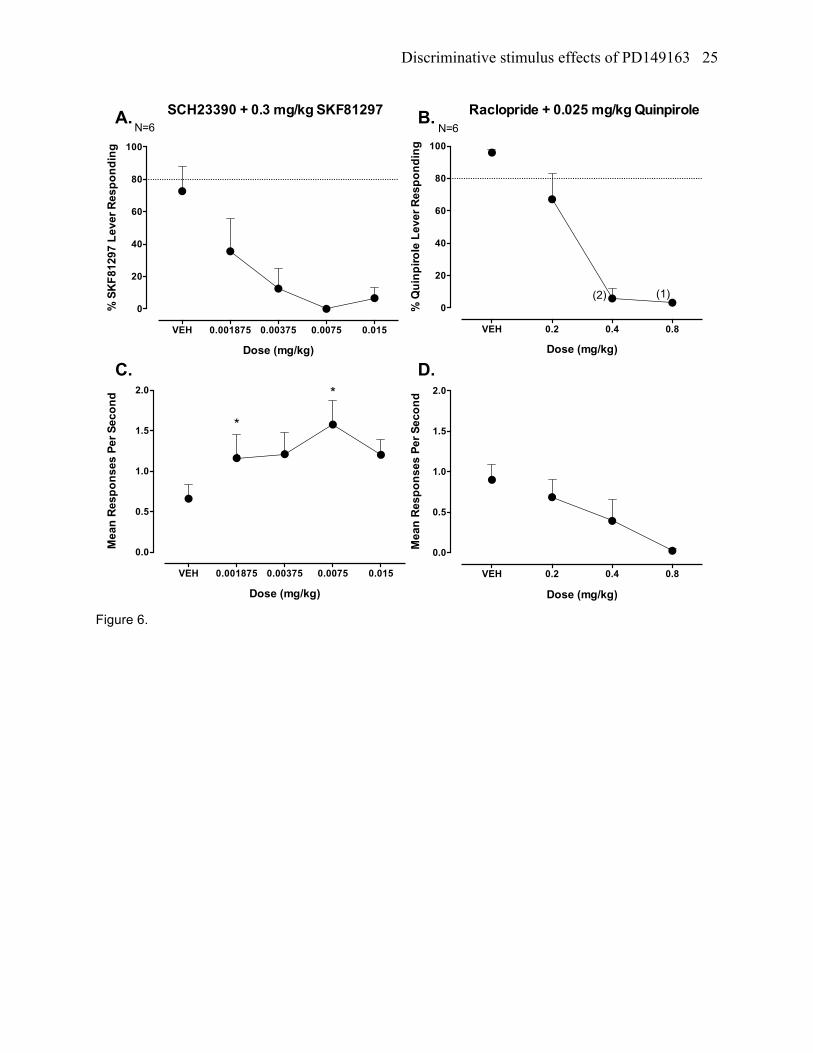
SCH22390+SKF81297 testing in SKF81297 trained rats
The D1 receptor antagonist SCH23390 (0.001875 – 0.015 mg/kg) produced full blockade of the
SKF81297 discriminative cue, which occurred after administration of a 0.00375, 0.075, and 0.015 mg/kg
dose (figure 6a). A statistically significant increase in response rates occurred (F[4,20], =5.90, p<0.05),
which was found at a 0.001875 mg/kg and 0.015 mg/kg dose compared to vehicle (figure 6c).
Raclopride + quinpirole testing in quinpirole trained rats
The D2 receptor antagonist raclopride (0.2 – 0.8 mg/kg) produced full blockade of the quinpirole
discriminative cue, which occurred after administration of a 0.4 and 0.8 mg/kg dose of raclopride (figure
6b). A statistically significant decrease in response rates occurred (F[3,15] =4.34, p<0.05), which was
found at a 0.8 mg/kg dose compared to vehicle (figure 6d).
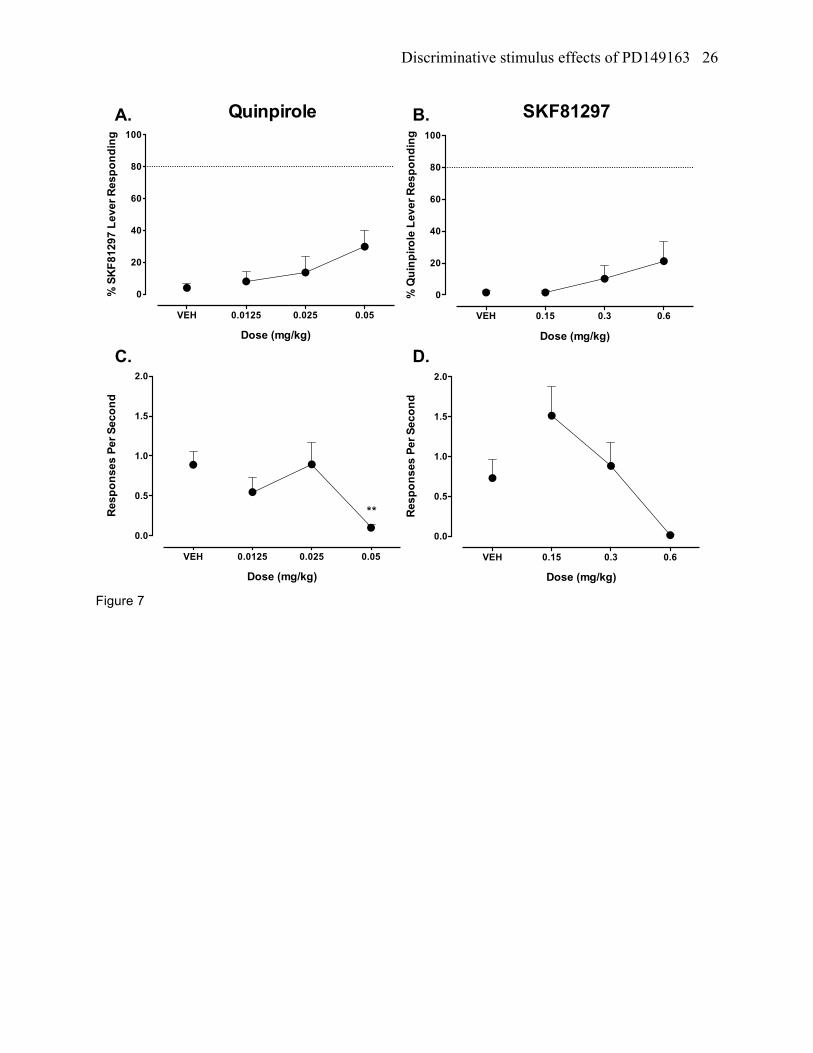
Quinpirole testing in SKF81297 trained rats
Quinpirole (0.0125 – 0.05 mg/kg) did not produce stimulus generalization from the SKF81297
discriminative cue (figure 7a). A significant reduction in response rates was found during quinpirole
testing (F[3,9]=6.75, p<0.05), which occurred at the 0.05 mg/kg dose compared to vehicle (figure 7c).
SKF81297 testing in quinpirole trained rats
SKF81297 (0.15-0.6 mg/kg) did not produce stimulus generalization from the quinpirole
discriminative cue (figure 7b). A significant reduction in rates occurred during SKF81297 testing
(F[3,9]=4.77, p<0.05), although a post hoc test failed to reveal a dose that produced statistically
significant differences in response rates compared to vehicle (figure 7d). The inability to find statistical
differences using a post hoc test is most likely due to the low sample size (N=4) that was available at this
later part of the study.
DISCUSSION
This is the first known report on the discriminative stimulus effects of a NTS1 receptor ligand.
Experiment 1 demonstrated that the NTS1 receptor agonist PD149163 could be established as a
discriminative stimulus in a drug discrimination procedure. Full stimulus generalization occurred from the
training dose of PD149163 to PD149163 and to the D2 receptor-preferring antagonist and typical
antipsychotic drug haloperidol. In experiment 2, PD149163 was neither found to produce stimulus

Discriminative stimulus effects of PD149163 11
generalization from the D1 receptor agonist SKF81297 nor the D2 receptor agonist quinpirole. The
selectivity of the discriminative stimulus effects of SKF81297 and quinpirole for D1 and D2 receptors,
respectively, was demonstrated by full stimulus blockade by antagonists for these receptors and by a lack
of cross generalization between these training drugs.
While PD149163 was established as a discriminative stimulus in experiment 1, these stimulus
effects were not robust. Only 6 of the 10 rats met the training criteria after a lengthy number of sessions,
and the discriminative stimulus effects of PD149163 could not be maintained after stimulus generalization
testing with PD149163 and haloperidol was completed. As reviewed in the introduction, NTS1 receptor
agonists likely functionally antagonize D2 receptors, and therefore the discriminative stimulus effects of a
NTS1 receptor agonist may in part be mediated by D2 receptor antagonism, which would account for
stimulus generalization to haloperidol. D2 receptor antagonism is a central pharmacological mechanism
for antipsychotic drugs, and therefore, this is a potential mechanism through which NTS1 receptor
agonists may engender antipsychotic-like effects in preclinical models. However, D2 receptor antagonism
may only mediate the stimulus properties of PD149163 when this compound is used as a training drug,
since D2 receptor antagonism was not apparent when PD149163 was tested in the quinpirole-trained rats.
However, response suppression caused by PD149163 precluded higher testing doses that were found to
produce full stimulus generalization from PD149163 in experiment 1. Thus, the lack of stimulus blockade
of quinpirole by PD149163 may be due to testing doses that were too low to adequately counteract these
stimulus effects.
Antipsychotic drugs have been thoroughly evaluated in drug discrimination procedures.
Generally, atypical antipsychotic drugs, which exhibit an extensive binding profile for monoaminergic and
cholinergic receptors, are readily established and robustly maintained as discriminative stimuli (i.e., as
training drugs in drug discrimination procedures). For example, the atypical antipsychotic drugs clozapine
[Porter et al. 2005; Prus et al. 2004; Prus et al. 2005], olanzapine [Porter and Strong 1996], and
quetiapine [Smith and Goudie 2002] have been established as discriminative stimuli in rats. On the other
hand, typical antipsychotic drugs, which engender a more selective receptor profile that primarily involves
D2 receptor antagonism, are less readily established as discriminative stimuli in rats.

Discriminative stimulus effects of PD149163 12
While the typical antipsychotic drug chlorpromazine, which exhibits a more general
monoaminergic receptor binding profile than most other typical antipsychotic drugs [Kroeze et al. 2003],
can be readily established as a discriminative stimulus in rats [Goas and Boston 1978; Porter et al. 1998],
haloperidol establishes a weaker discriminative stimulus. Colpaert [1976] was one of the first to establish
a haloperidol discriminative stimulus in rats but did so with difficulty, requiring a dose reduction from a
0.04 mg/kg to 0.02 mg/kg training dose. Subsequently, a haloperidol discriminative stimulus was
established after 80 mean training sessions. McElroy et al. [1989] later trained rats to discriminate
haloperidol 0.04 mg/kg from vehicle in a mean of 45 sessions and found that full stimulus generalization
occurred to chlorpromazine and that the haloperidol discriminative stimulus was blocked by cocaine and
d-amphetamine. Wiley and Porter [1993] also attempted to train rats to discriminate haloperidol from
clozapine but found that pharmacologically dissimilar drugs produced stimulus generalization from
haloperidol. These authors concluded that haloperidol was treated as the “other” condition (similar to a
vehicle condition in most two-lever drug discrimination studies) versus clozapine, rather than an
independent discriminative stimulus. If a PD149163 discriminative stimulus were mediated in a significant
way by D2 receptor antagonism, then this may account for the difficulty in establishing and maintaining
these stimulus effects.
The training data for SKF81297 and quinpirole confirm previous studies that these compounds
can be established as discriminative stimuli. Reavill et al. [1993] first established SKF81297, at a 0.1 and
0.2 mg/kg dose, as a discriminative stimulus in rats and reported full stimulus to SKF91297 up to a 0.4
mg/kg dose. In the present study, the 0.3 mg/kg training dose of SKF81297 produced full stimulus
generalization to a 0.3 and 0.6 mg/kg dose and partial stimulus generalization to a 1.2 mg/kg dose. The
lack of full stimulus generalization to a 1.2 mg/kg dose of SKF81297 was likely due to rate disruptive
effects or other nonspecific behavioral effects (e.g., impaired working memory [Zahrt et al. 1997]) of this
dose. The discriminative stimulus effects of quinpirole have been established in previous studies [Baladi
et al. 2010; Weathersby and Appel 1986] and appear to be primarily mediated by agonism of both D2 and
D3 receptors [Baladi et al. 2010]. In the present study, full stimulus generalization occurred from the 0.025
mg/kg training dose of quinpirole to a 0.025 and 0.05 mg/kg dose and partial generalization occurred to a
0.1 mg/kg dose. The reduction to partial generalization at a 0.2 mg/kg dose of quinpirole may be have

Discriminative stimulus effects of PD149163 13
been due to rate disruptive effects of this dose. The selectivity of the discriminative stimulus effects of
SKF81297 and quinpirole for D1 and D2 receptors, respectively, was supported by full stimulus blockade
by antagonists for these receptors and by a lack of cross generalization between SKF81297 and
quinpirole.
While generalization from a D2 receptor agonist to PD149163 was not considered likely based on
past molecular and behavioral studies, the disinhibition of dopamine release caused by D2 autoreceptor
inhibition was thought to possibly lead to postsynaptic D1 receptor binding [Binder et al. 2001]. Stimulus
generalization testing with PD149163 indicated a lack of D1 receptor or D2 receptor agonism for the
stimulus effects of PD149163. PD149163 significantly disrupted responding in both the SKF81297- and
quinpirole-trained rats, although fewer animals responded to higher doses of PD149163 in the
SKF81297-trained rats than in the quinpirole-trained rats. A 0.0125 mg/kg dose of PD149163 reduced
responding on the quinpirole lever to below the full stimulus generalization criterion. These effects might
be reflective of D2 receptor antagonism by PD149163, but if this is true, the effects are less pronounced
than were found in experiment 1, in which PD149163 was used as a discriminative stimulus.
In conclusion, the present study provided tentative evidence for the mediation of the
discriminative stimulus effects of PD149163 by dopamine D2 receptor antagonism, although these effects
were most pronounced when PD149163 was used as the training drug. These findings are consistent
with the putative antipsychotic effects found in preclinical models, although the atypical antipsychotic
features of NTS1 receptor agonists suggest that they may involve more than antagonism of D2 receptors.
Currently available NTS1 receptor antagonists (i.e., SR14297 and SR48692) have poor solubility and
therefore precluded their systemic use in this study to determine if NTS1 receptor antagonism may block
the discriminative stimulus effects of PD149163. Moreover, other NTS1 receptor agonists, which could be
used in stimulus generalization testing, are not commercially available. Further elucidation of the
discriminative stimulus effects of NTS1 receptor agonists will be improved upon the availability of other
brain penetrant NTS1 receptor ligands and with stimulus generalization testing conducted with ligands for
other receptors (e.g., receptors important for atypical antipsychotic drugs, such as 5-HT2A receptors) that
may be important for the behavioral effects of neurotensin.

Discriminative stimulus effects of PD149163 14
ACKNOWLEDGEMENT
PD149163 was generously provided by the NIMH Chemical Synthesis and Drug Supply Program.
Experiment 1 of this study was supported by an NIH grant to AJP (1R15MH083241). The authors thank
Alexis Bench and Casey Cook for their technical assistance.
REFERENCES
Animals CftUotGftCaUoL. 2011. Guide for the care and use of laboratory animals. Washington DC:
National Academies Press.
Azmi N, Norman C, Spicer CH, Bennett GW. 2006. Effects of a neurotensin analogue (PD149163) and
antagonist (SR142948A) on the scopolamine-induced deficits in a novel object discrimination
task. Behavioural pharmacology 17(4):357-62.
Baladi MG, Newman AH, France CP. 2010. Dopamine D3 receptors mediate the discriminative stimulus
effects of quinpirole in free-feeding rats. The Journal of pharmacology and experimental
therapeutics 332(1):308-15.
Beaulieu JM, Gainetdinov RR. 2011. The physiology, signaling, and pharmacology of dopamine
receptors. Pharmacol Rev 63(1):182-217.
Binder EB, Kinkead B, Owens MJ, Nemeroff CB. 2001. Neurotensin and dopamine interactions.
Pharmacol Rev 53(4):453-86.
Boules M, Oliveros A, Liang Y, Williams K, Shaw A, Robinson J, Fredrickson P, Richelson E. 2011. A
neurotensin analog, NT69L, attenuates intravenous nicotine self-administration in rats.
Neuropeptides 45(1):9-16.
Colpaert FC, Niemegeers CJ, Janssen PA. 1976. Theoretical and methodological considerations on drug
discrimination learning. Psychopharmacologia 46(2):169-77.
Cusack B, Boules M, Tyler BM, Fauq A, McCormick DJ, Richelson E. 2000. Effects of a novel neurotensin
peptide analog given extracranially on CNS behaviors mediated by apomorphine and haloperidol.
Brain research 856(1-2):48-54.

Discriminative stimulus effects of PD149163 15
Feifel D, Melendez G, Murray RJ, Tina Tran DN, Rullan MA, Shilling PD. 2008. The reversal of
amphetamine-induced locomotor activation by a selective neurotensin-1 receptor agonist does
not exhibit tolerance. Psychopharmacology 200(2):197-203.
Feifel D, Reza TL, Wustrow DJ, Davis MD. 1999. Novel antipsychotic-like effects on prepulse inhibition of
startle produced by a neurotensin agonist. The Journal of pharmacology and experimental
therapeutics 288(2):710-3.
Fredrickson P, Boules M, Lin SC, Richelson E. 2005. Neurobiologic basis of nicotine addiction and
psychostimulant abuse: a role for neurotensin? The Psychiatric clinics of North America
28(3):737-51, 746.
Gleason SD, Witkin JM. 2006. Effects of dopamine D1 receptor agonists in rats trained to discriminate
dihydrexidine. Psychopharmacology 186(1):25-31.
Goas JA, Boston JE, Jr. 1978. Discriminative stimulus properties of clozapine and chlorpromazine.
Pharmacology Biochemistry and Behavior 8(3):235-41.
Holly EN, Ebrecht B, Prus AJ. 2011a. The neurotensin-1 receptor agonist PD149163 inhibits conditioned
avoidance responding without producing catalepsy in rats. Eur Neuropsychopharmacol
21(7):526-31.
Holly EN, Ebrecht B, Prus AJ. 2011b. The neurotensin-1 receptor agonist PD149163 inhibits conditioned
avoidance responding without producing catalepsy in rats. Eur Neuropsychopharmacol.
Kroeze WK, Hufeisen SJ, Popadak BA, Renock SM, Steinberg S, Ernsberger P, Jayathilake K, Meltzer
HY, Roth BL. 2003. H1-histamine receptor affinity predicts short-term weight gain for typical and
atypical antipsychotic drugs. Neuropsychopharmacology 28(3):519-26.
McElroy JF, Stimmel JJ, O'Donnell JM. 1989. Discriminative stimulus properties of haloperidol. Drug
Development Research 18:47-55.
Petrie KA, Bubser M, Casey CD, Davis MD, Roth BL, Deutch AY. 2004. The neurotensin agonist
PD149163 increases Fos expression in the prefrontal cortex of the rat.
Neuropsychopharmacology 29(10):1878-88.

Discriminative stimulus effects of PD149163 16
Porter JH, Covington HE, Varvel SA, Vann RE, Warren TA. 1998. Chlorpromazine as a discriminative
stimulus in rats: generalization to typical and atypical antipsychotics. Drug Development
Research 48:38-44.
Porter JH, Prus AJ. 2009. Drug discrimination: 30 years of progress. Psychopharmacology 203(2):189-
91.
Porter JH, Prus AJ, Vann RE, Varvel SA. 2005. Discriminative stimulus properties of the atypical
antipsychotic clozapine and the typical antipsychotic chlorpromazine in a three-choice drug
discrimination procedure in rats. Psychopharmacology 178(1):67-77.
Porter JH, Strong SE. 1996. Discriminative stimulus control with olanzapine: generalization to the atypical
antipsychotic clozapine. Psychopharmacology 128(2):216-9.
Prus AJ, Baker LE, Meltzer HY. 2004. Discriminative stimulus properties of 1.25 and 5.0 mg/kg doses of
clozapine in rats: examination of the role of dopamine, serotonin, and muscarinic receptor
mechanisms. Pharmacology, biochemistry, and behavior 77(2):199-208.
Prus AJ, Hillhouse TM, LaCrosse AL. 2014. Acute, but not repeated, administration of the neurotensin
NTS1 receptor agonist PD149163 decreases conditioned footshock-induced ultrasonic
vocalizations in rats. Progress in neuro-psychopharmacology & biological psychiatry 49:78-84.
Prus AJ, Huang M, Li Z, Dai J, Meltzer HY. 2007. The neurotensin analog NT69L enhances medial
prefrontal cortical dopamine and acetylcholine efflux: Potentiation of risperidone-, but not
haloperidol-, induced dopamine efflux. Brain research 1184:354-64.
Prus AJ, Philibin SD, Pehrson AL, Stephens CL, Cooper RN, Wise LE, Porter JH. 2005. Generalization
testing with atypical and typical antipsychotic drugs in rats trained to discriminate 5.0 mg/kg
clozapine from vehicle in a two-choice drug discrimination task. Drug Dev Res 64:55-65.
Reavill C, Bond B, Overend P, Hunter AJ. 1993. Pharmacological characterization of the discriminative
stimulus properties of the dopamine D1 agonist, SKF 81297. Behavioural pharmacology 4(2):135-
146.
Shilling PD, Feifel D. 2008. The neurotensin-1 receptor agonist PD149163 blocks fear-potentiated startle.
Pharmacology, biochemistry, and behavior 90(4):748-52.

Discriminative stimulus effects of PD149163 17
Shilling PD, Richelson E, Feifel D. 2003. The effects of systemic NT69L, a neurotensin agonist, on
baseline and drug-disrupted prepulse inhibition. Behavioural brain research 143(1):7-14.
Smith JA, Goudie AJ. 2002. Discriminative stimulus properties in rats of the novel antipsychotic
quetiapine. Exp Clin Psychopharmacol 10(4):376-84.
St-Galais F, Jomphe C, Trudeau L-E. 2006. The role of neurotensin in central nervous system
pathophysiology: What is the evidence? J Psychatry Neurosci 31:229-245.
Tanganelli S, Antonelli T, C. Tomasini M, Beggiato S, Fuxe K, Ferraro L. 2012. Relevance of Dopamine
D2/Neurotensin NTS1 and NMDA/Neurotensin NTS1 Receptor Interaction in Psychiatric and
Neurodegenerative Disorders. Current Medicinal Chemistry 19(3):304-316.
Weathersby RT, Appel JB. 1986. Dopamine D2 receptor mediation of the discriminative stimulus
properties of LY 171555 (quinpirole). European journal of pharmacology 132(1):87-91.
Wiley JL, Porter JH. 1993. Effects of serotonergic drugs in rats trained to discriminate clozapine from
haloperidol. Bull Psychonomic Soc 31:94-96.
Zahrt J, Taylor JR, Mathew RG, Arnsten AFT. 1997. Supranormal Stimulation of D1 Dopamine Receptors
in the Rodent Prefrontal Cortex Impairs Spatial Working Memory Performance. The Journal of
Neuroscience 17(21):8528-8535.

Discriminative stimulus effects of PD149163 18
FIGURES
Figure 1. The neurotensin NT1 receptor agonist PD149163 was tested for stimulus generalization in rats
trained to discriminate PD149163, 0.0625 mg/kg, from vehicle (VEH) in a two-choice drug discrimination
task. Mean (+/- SEM) percent drug lever responding is shown in panel A and the mean (+/-) responses
per second is shown in panel B. The sample size is shown in parentheses; otherwise, N=6. The dashed
line indicates the full stimulus generalization criterion. *P<0.05, **P<0.01, ***P<0.001 versus VEH
control.
Figure 2. Stimulus generalization results for the typical antipsychotic drug and D2-receptor preferring
antagonist haloperidol (N=5). See figure 1 for further details.
Figure 3. Stimulus generalization testing with the D1 receptor agonist SKF81297 in rats trained
discriminate SKF81297 (0.3 mg/kg) from VEH (panels A and C) and with the D2 receptor agonist
quinpirole in rats trained to discriminate quinpirole (0.025 mg/kg) from VEH (panels B and D) in a two
choice drug discrimination task. Mean percent (+/- SEM) lever responding for SKF81297 and quinpirole
are shown in panels A and B, respectively, and the mean responses per second (+/- SEM) for SKF81297
and quinpirole are shown in panel C and D, respectively. **P<0.01 and ***P<0.001 versus VEH control.
Figure 4. Stimulus generalization testing with PD149163 in SKF81297- (panel A, percent SKF81297-
appropriate responding; panel C, responses per minute) and quinpirole-trained rats (panel B, quinpirole-
appropriate responding; panel D, responses per minute). *P<0.05, **P<0.01, ***P<0.001, and
****P<0.0001 versus VEH control. See figure 3 for further details.
Figure 5. Generalization testing with PD149163 in combination with SKF81297 (0.3 mg/kg) in
SKF81297-trained rats (A & C) and in combination with quinpirole (0.025 mg/kg) in quinpirole-trained rats
(B & D). See figure 3 for further details.

Discriminative stimulus effects of PD149163 19
Figure 6. Generalization testing with either the dopamine D1 receptor antagonist SCH23390 in
combination with SKF81297 (0.3 mg/kg) in SKF81297-trained rats (A & C) and the dopamine D2/D3
receptor antagonist raclopride in combination with quinpirole (0.025 mg/kg) in quinpirole-trained rats (B &
D). See figure 3 for further details.
Discriminative stimulus effects of PD149163 20
Figure 1
VEH 0.016 0.031 0.0625 0.13 0.25
0
20
40
60
80
100
Dose (mg/kg)
% P
D14
9163
Lev
er R
espo
ndin
g
(3)
N=6 (5)
VEH 0.016 0.031 0.0625 0.13 0.25
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Res
pons
es P
er S
econ
d
** ****
PD149163A.
B.
Discriminative stimulus effects of PD149163 21
Figure 2
VEH 0.025 0.05 0.1 0.2
0
20
40
60
80
100
Dose (mg/kg)
% P
D14
9163
Lev
er R
espo
ndin
g
(3)
N=5 (2)
VEH 0.025 0.05 0.1 0.2
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Res
pons
es P
er S
econ
d
*
** **
HaloperidolA.
B.
Discriminative stimulus effects of PD149163 22
Figure 3
VEH 0.15 0.3 0.6 1.2
0
20
40
60
80
100
Dose (mg/kg)
% S
KF8
1297
Lev
er R
espo
ndin
gN=6
VEH 0.0125 0.025 0.05 0.1
0
20
40
60
80
100
% Q
uinp
irole
Lev
er R
espo
ndin
g
Dose (mg/kg)
N=6
VEH 0.15 0.3 0.6 1.2
0.0
0.5
1.0
1.5
2.0
Res
pons
es P
er S
econ
d
Dose (mg/kg)
****
***
VEH 0.0125 0.025 0.05 0.1
0.0
0.5
1.0
1.5
2.0
Res
pons
es P
er S
econ
d
Dose (mg/kg)
**
SKF81297 QuinpiroleA. B.
C. D.
Discriminative stimulus effects of PD149163 23
Figure 4
Dose (mg/kg)
% S
KF8
1297
Lev
er R
espo
ndin
g
VEH 0.05 0.075 0.1 0.15
0
20
40
60
80
100
N=6
(5) (5)
(2) (2)
VEH 0.05 0.075 0.1 0.15
0
20
40
60
80
100
Dose (mg/kg)
% Q
uinp
irole
Lev
er R
espo
ndin
g
N=6
(5) (5)
(1)
VEH 0.05 0.075 0.1 0.15
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Mea
n R
espo
nses
Per
Sec
ond
**
*******
VEH 0.05 0.075 0.1 0.15
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Mea
n R
espo
nses
Per
Sec
ond
** **
***
PD149163 PD149163A. B.
C. D.
Discriminative stimulus effects of PD149163 24
Figure 5
VEH 0.025 0.05 0.075
0
20
40
60
80
100
Dose (mg/kg)
% S
KF8
1297
Lev
er R
espo
ndin
g
N=6
(4)(3)
VEH 0.0030125 0.006125 0.0125
0
20
40
60
80
100
Dose (mg/kg)
% Q
uinp
irole
Lev
er R
espo
ndin
g
N=6
(5) (4)
(4)
VEH 0.025 0.05 0.075
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Mea
n R
espo
nses
Per
Sec
ond
VEH 0.0030125 0.006125 0.0125
0.0
0.5
1.0
1.5
2.0M
ean
Res
pons
es P
er S
econ
d
Dose (mg/kg)
PD149163+ 0.3mg/kg SKF81297 PD149163+ 0.025 mg/kg QuinpiroleA. B.
C. D.
Discriminative stimulus effects of PD149163 25
Figure 6.
VEH 0.001875 0.00375 0.0075 0.015
0
20
40
60
80
100
Dose (mg/kg)
% S
KF8
1297
Lev
er R
espo
ndin
gN=6
VEH 0.2 0.4 0.8
0
20
40
60
80
100
% Q
uinp
irole
Lev
er R
espo
ndin
g
Dose (mg/kg)
(2) (1)
N=6
VEH 0.001875 0.00375 0.0075 0.015
0.0
0.5
1.0
1.5
2.0
Mea
n R
espo
nses
Per
Sec
ond
Dose (mg/kg)
*
*
*
VEH 0.2 0.4 0.8
0.0
0.5
1.0
1.5
2.0
Mea
n R
espo
nses
Per
Sec
ond
Dose (mg/kg)
SCH23390 + 0.3 mg/kg SKF81297 Raclopride + 0.025 mg/kg QuinpiroleA. B.
C. D.
Discriminative stimulus effects of PD149163 26
Figure 7
VEH 0.0125 0.025 0.05
0
20
40
60
80
100
Dose (mg/kg)
% S
KF8
1297
Lev
er R
espo
ndin
g
VEH 0.15 0.3 0.6
0
20
40
60
80
100
Dose (mg/kg)
% Q
uinp
irole
Lev
er R
espo
ndin
g
VEH 0.0125 0.025 0.05
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Res
pons
es P
er S
econ
d
**
VEH 0.15 0.3 0.6
0.0
0.5
1.0
1.5
2.0
Dose (mg/kg)
Res
pons
es P
er S
econ
d
Quinpirole SKF81297A. B.
C. D.




















