
Defective Stimulus Set Attention ... Epilepsy
12
BRAIN AND COGNITION 3, 146151 (1984) Defective Stimulus Set Attention in Generalized Epilepsy JASON BRANDT The Johns Hopkins University This study examined auditory selective attention in primary generalized and complex-partial (temporal lobe) epileptics using binaural and dichotic versions of the Continuous Performance Test. The patient groups did not differ from normal controls when only one channel was presented or when divided attention (simultaneously monitoring two channels) was required. However, generalized epileptics had significantly depressed hit rates when directed attention (responding to critical stimuli in only one channel) was required. These results suggest a contribution of brain stem “arousal” mechanisms to stimulus set attention. In addition, right ear perceptual advantages were found on these dichotic monitoring tasks, lending support to the contention that short-term auditory memory and verbal report are not required for the dichotic ear effect. It is well known that patients with primary generalized epilepsy (par- ticularly of the absence type) exhibit deficits in arousal and attentiveness (Penfield & Jasper, 1954). Numerous studies using the Continuous Per- formance Test (CPT) and other sustained attention tasks have demonstrated defects in the interictal performances of generalized epileptics, but not temporal lobe epileptics (Landsell & Mirsky, 1964; Mirsky & Van Buren, 1965; Fedio & Mirsky, 1969). This has traditionally been attributed to dysfunction of “centrencephalic” arousal mechanisms, involving the mesencephalic reticular formation, in generalized seizure patients (Penfield & Jasper, 1954; Jasper, 1957; Gloor, 1968). More recently, however, it has been suggested that some forms of generalized epilepsy, particularly those associated with three per-second spike-wave paroxysms on EEG, are due not to pathology in a centrencephalic “pacemaker,” but to generalized hyperexcitability of cortical neurons (Gloor, 1979). Regardless This work was supported by NIMH Predoctoral Fellowship lF31MHO8148-02 to the author and by NIH Grant 2S07RR07041-16to the Johns Hopkins University. The author thanks Drs. Nelson Butters, Merle Orren, William Mackavey, and Allan Mirsky for their assistance in the conduct of this research. Portions of this paper were presented at the 1lth Annual Meeting of the International Neuropsychological Society, February 1983. Send correspondence to the author at: Department of Psychology, The Johns Hopkins University, Baltimore, MD 21218. 140 0278-2626/84/$3.00 Copyright Q 1984 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
johnshopkins -
Category
Documents
-
view
0 -
download
0
Transcript of Defective Stimulus Set Attention ... Epilepsy

BRAIN AND COGNITION 3, 146151 (1984)
Defective Stimulus Set Attention in Generalized Epilepsy
JASON BRANDT
The Johns Hopkins University
This study examined auditory selective attention in primary generalized and complex-partial (temporal lobe) epileptics using binaural and dichotic versions of the Continuous Performance Test. The patient groups did not differ from normal controls when only one channel was presented or when divided attention (simultaneously monitoring two channels) was required. However, generalized epileptics had significantly depressed hit rates when directed attention (responding to critical stimuli in only one channel) was required. These results suggest a contribution of brain stem “arousal” mechanisms to stimulus set attention. In addition, right ear perceptual advantages were found on these dichotic monitoring tasks, lending support to the contention that short-term auditory memory and verbal report are not required for the dichotic ear effect.
It is well known that patients with primary generalized epilepsy (par- ticularly of the absence type) exhibit deficits in arousal and attentiveness (Penfield & Jasper, 1954). Numerous studies using the Continuous Per- formance Test (CPT) and other sustained attention tasks have demonstrated defects in the interictal performances of generalized epileptics, but not temporal lobe epileptics (Landsell & Mirsky, 1964; Mirsky & Van Buren, 1965; Fedio & Mirsky, 1969). This has traditionally been attributed to dysfunction of “centrencephalic” arousal mechanisms, involving the mesencephalic reticular formation, in generalized seizure patients (Penfield & Jasper, 1954; Jasper, 1957; Gloor, 1968). More recently, however, it has been suggested that some forms of generalized epilepsy, particularly those associated with three per-second spike-wave paroxysms on EEG, are due not to pathology in a centrencephalic “pacemaker,” but to generalized hyperexcitability of cortical neurons (Gloor, 1979). Regardless
This work was supported by NIMH Predoctoral Fellowship lF31MHO8148-02 to the author and by NIH Grant 2S07RR07041-16 to the Johns Hopkins University. The author thanks Drs. Nelson Butters, Merle Orren, William Mackavey, and Allan Mirsky for their assistance in the conduct of this research. Portions of this paper were presented at the 1 lth Annual Meeting of the International Neuropsychological Society, February 1983. Send correspondence to the author at: Department of Psychology, The Johns Hopkins University, Baltimore, MD 21218.
140 0278-2626/84/$3.00 Copyright Q 1984 by Academic Press, Inc. All rights of reproduction in any form reserved.

ATTENTIONAL DEFECT IN EPILEPSY 141
of which of these models best describes the pathogenesis of primary generalized seizures, the fact remains that they are associated with both the absence of focal cortical pathology and a prominent disturbance of attentiveness.
The status of selective attention in its many varieties has rarely been studied in epileptic patients. While the precise neuropsychological mech- anisms underlying the ability to distribute mental effort unequally among competing tasks or stimuli remain poorly understood (Posner, 1975), several brain systems have been implicated. Luria maintained that the superior brain stem and ascending reticular activating formation are re- sponsible for only the most elementary condition of attention, the gen- eralized state of waking. He asserted that any more complex form of attention which requires selective recognition of a particular stimulus and inhibition of responses to irrelevant stimuli is accomplished by brain structures at higher levels, particularly in the limbic system and in the frontal cortex. (Luria, 1973, p. 271). The parietal cortex, especially of the right cerebral hemisphere, has also been implicated in selective attention (Weinstein & Friedland, 1977). Recently, Mesulem (1981) has attempted a synthesis of behavioral, electrophysiological, and anatomical data, and has described a cerebral network for divided attention which includes posterior parietal cortex, frontal cortex, and cingulate cortex, in addition to the reticular component.
The present investigation examined two varieties of auditory selective attention in generalized and focal (temporal lobe) epileptics using binaural and dichotic versions of CPT. One variety, “response set” attention, requires the selective recognition of particular target stimuli and usually a specific response to them (Broadbent, 1970). Thus, the standard CPT is best thought of as a response set task. “Stimulus set” attention requires the subject to distinguish between concurrent input channels and select one channel for immediate processing. The subjects in the present study performed three auditory attention tasks, all of which required response set attention, but only one of which required stimulus set attention. If the mechansims of stimulus set are cortical in origin (for example, a functional “priming” of one hemisphere, as suggested by Kinsbourne, 1973), impairments on the task requiring channel selection should be found in epileptic patients with temporal lobe foci. Either performance should be impaired bilaterally or subjects should be inattentive to stimuli contralateral to the seizure focus (but cf. Mazzucchi & Parma, 1978). On the other hand, if the selection of auditory channels is more highly dependent on the integrity of subcortical (e.g., brain stem) mechanisms, patients with generalized seizures should display performance deficits, and stimuli presented to the left and right ears should be missed equally often.

142 JASON BRANDT
METHOD
Subjects. Four groups of subjects were studied: primary generalized epileptics (GEs), complex-partial epileptics (CPEs) with left temporal lobe foci, complex-partial epileptics with right temporal lobe foci, and normal controls. Patients were selected using widely accepted EEG and clinical criteria (International League Against Epilepsy, 1981). The GE group was composed of 11 patients whose seizures were marked by impaired consciousness and which provided no clinical evidence of local onset. Their EEGs reflected bilaterally symmetrical paroxysmal activity which gave no indication of lateralized onset. This group consisted of two patients with pure absence seizures, one with absence seizures with automatisms, three with idiopathic tonic-clonic seizures, two with a combination of tonic-clonic and absence seizures, two with tonic-clonic and myoclonic seizures, and one patient with atonic-tonic seizures.
The two CPE groups were each composed of 11 patients with complex- partial seizures (marked by impaired consciousness with automatisms, and affective, cognitive, psychosensory, and/or psychomotor changes) and unequivocal spikes or slow waves emanating from temporal leads. One group was composed of patients whose EEGs consistently indicated left temporal lobe foci. Six patients had left temporal spike or sharp- wave foci, two had slow-wave foci, and three displayed both sharp waves and focal slowing. Six of these eleven patients had only complex-partial seizures, while five had experienced some generalized convulsions as well. These generalized seizures are thought to be secondarily generalized from temporal lobe foci.
The other complex-partial group was composed of 11 patients with right temporal lobe foci. Five had EEGs indicating sharp-wave or spike foci, four had slow-wave foci, and two had evidence of both. Six of the eleven right temporal patients had only complex-partial seizures, while five had secondarily generalized seizures as well.
CPE patients in whom primary foci could not be identified were excluded from the study, as were those with evidence of multifocal brain damage. Individuals with known or suspected hearing loss, audiogenic seizures, or frequent uncontrolled major motor seizures were likewise excluded. Subjects with multiple seizure types were classified according to the most frequent seizure type which was consistent with the EEG findings. All patients were maintained on anticonvulsant medications, and none had toxic serum levels of these medications.
A control group of 11 nonepileptic, neurologically normal individuals was also studied. As seen in Table 1, the four subject groups were matched on age, education, age at onset of seizures, duration of illness, and seizure frequency. All subjects were right-handed as judged by self- report and preferred hand for writing.
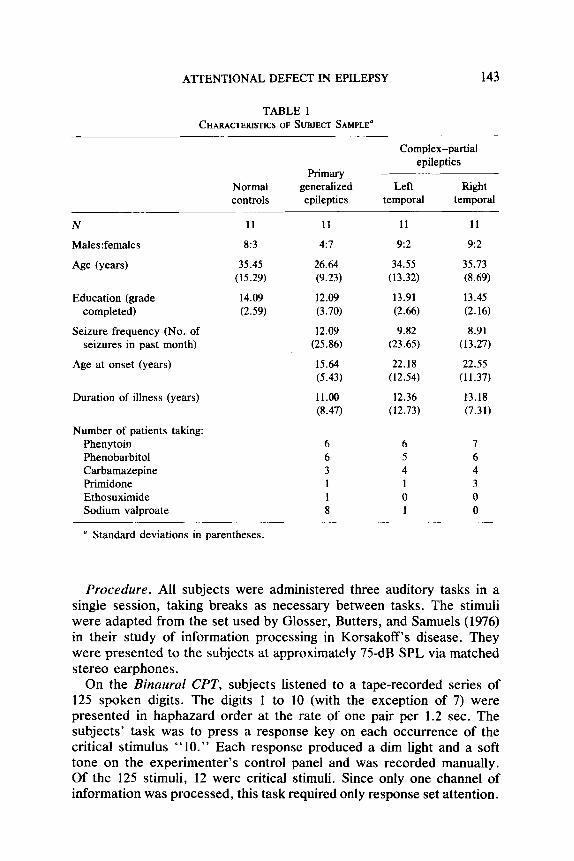
A’ITENTIONAL DEFECT IN EPILEPSY 143
TABLE 1 CHARACTERISTICS OF SUBJECT SAMPLE’
N
Males:females
Age (yem)
Education (grade completed)
Seizure frequency (No. of seizures in past month)
Age at onset (years)
Duration of illness (years)
Number of patients taking: Phenytoin Phenobarbital Carbamazepine Primidone Ethosuximide Sodium valproate
Normal controls
11
8:3
35.45 (15.29)
14.09 (2.59)
Primary generalized epileptics
11
4:7
26.64 (9.23)
12.09 (3.70)
12.09 (25.86)
15.64 (5.43)
11.00 (8.47)
Complex-partial epileptics
Left Bight temporal temporal
11 11
9:2 9~2
34.55 35.73 (13.32) (8.69)
13.91 13.45 (2.66) (2.16)
9.82 8.91 (23.65) (13.27)
22.18 22.55 (12.54) (11.37)
12.36 13.18 (12.73) (7.31)
6 6 7 6 5 6 3 4 4 1 1 3 1 0 0 8 1 0
” Standard deviations in parentheses.
Procedure. All subjects were administered three auditory tasks in a single session, taking breaks as necessary between tasks. The stimuli were adapted from the set used by Glosser, Butters, and Samuels (1976) in their study of information processing in Korsakoff’s disease. They were presented to the subjects at approximately 75dB SPL via matched stereo earphones.
On the Binaural CPT, subjects listened to a tape-recorded series of 125 spoken digits. The digits 1 to 10 (with the exception of 7) were presented in haphazard order at the rate of one pair per 1.2 sec. The subjects’ task was to press a response key on each occurrence of the critical stimulus “10.” Each response produced a dim light and a soft tone on the experimenter’s control panel and was recorded manually. Of the 125 stimuli, 12 were critical stimuli. Since only one channel of information was processed, this task required only response set attention.
144 JASON BRANDT
On the Dichotic CPT, Directed Attention Condition, subjects listened to 125 synchronized pairs of digits, presented at the rate of one pair every 1.2 sec. Each digit was paired with every other; none was paired with itself. Subjects heard this recording twice, with a 3-min rest break between the two blocks of trials. On the first block, subjects were told to attend only to the left (or right) channel and press the response key to “10”s on that side only. There were 25 such critical trials. On the second block of trials, the previously unattended channel was attended. Again, there were 25 critical trials. The order in which the two channels were attended was randomly selected for each subject. This task required both response set attention (listening for the target “10”) and stimulus set attention (listening at one ear only).
The third task was the Dichotic CPT, Divided Attention Condition. Using the same type of stimuli as in the Directed Attention Condition, subjects were here required to monitor both auditory channels simul- taneously and respond to all the “10”s. Of the 125 stimulus pairs, 24 contained critical stimuli; 12 on the left and 12 on the right. This task presented subjects with the same stimulus load and the same density of critical stimuli as in the Directed Attention Condition but, like the Binaural Condition, required only response set attention.
RESULTS
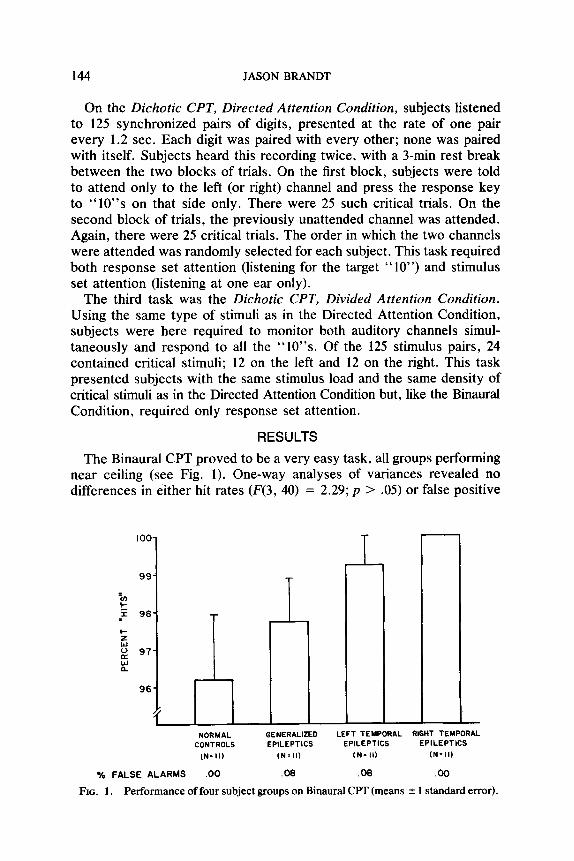
The Binaural CFT proved to be a very easy task, all groups performing near ceiling (see Fig. 1). One-way analyses of variances revealed no differences in either hit rates (F(3, 40) = 2.29; p > .05) or false positive
IOO-
I
- NORMAL GENERALIZED LEFT TEMPORAL RIGHT TEMPORAL
CONTROLS EPILEPTICS EPILEPTICS EPILEPTICS
IN= II) (N*ll) tN= II) (N= II)
% FALSE ALARMS .oo .08 .06 .oo
FIG. I. Performance of four subject groups on Binaural CPT (means f 1 standard error).
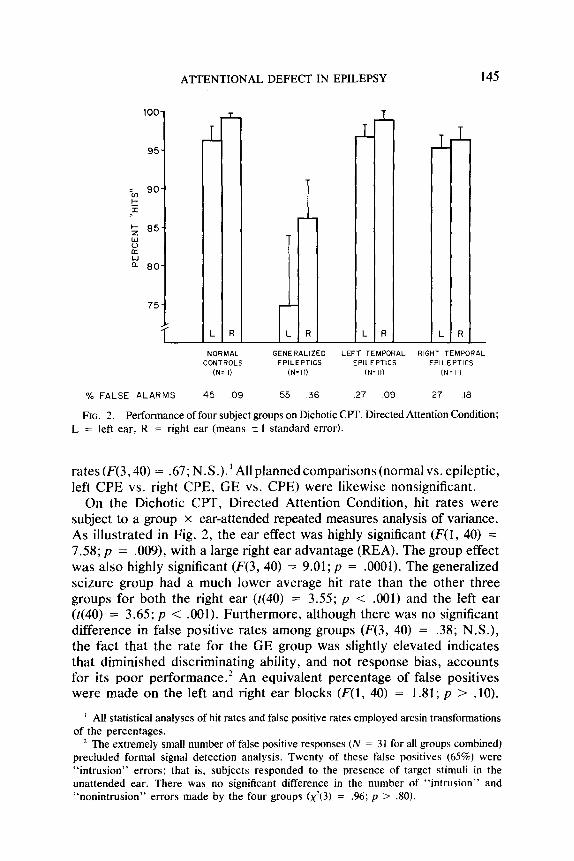
ATTENTIONAL DEFECT IN EPILEPSY 145
ii L R R - NORMAL GENERALIZED LEFT TEMPORAL RIGHT TEMPORAL
CONTROLS EPILEPTICS EPILEPTICS EPILEPTICS (N=II) (N=l I) IN- II) (N=II)
% FALSE ALARMS 45 09 55 36 27 09 .27 10
FIG. 2. Performance of four subject groups on Dichotic CPT, Directed Attention Condition; L = left ear, R = right ear (means 2 I standard error).
rates (F(3,40) = .67; N.S.). ’ All planned comparisons (normal vs. epileptic, left CPE vs. right CPE, GE vs. CPE) were likewise nonsignificant.
On the Dichotic CPT, Directed Attention Condition, hit rates were subject to a group x ear-attended repeated measures analysis of variance. As illustrated in Fig. 2, the ear effect was highly significant (F(1, 40) = 7.58; p = .009), with a large right ear advantage (REA). The group effect was also highly significant (F(3, 40) = 9.01; p = .OOOl). The generalized seizure group had a much lower average hit rate than the other three groups for both the right ear (t(40) = 3.55; p < .OOl) and the left ear (t(40) = 3.65; p < .OOl). Furthermore, although there was no significant difference in false positive rates among groups (F(3, 40) = .38; N.S.), the fact that the rate for the GE group was slightly elevated indicates that diminished discriminating ability, and not response bias, accounts for its poor performance.* An equivalent percentage of false positives were made on the left and right ear blocks (F(l) 40) = 1.81; p > .lO).
’ All statistical analyses of hit rates and false positive rates employed arcsin transformations of the percentages.
’ The extremely small number of false positive responses (N = 3 1 for all groups combined) precluded formal signal detection analysis. Twenty of these false positives (65%) were “intrusion” errors; that is, subjects responded to the presence of target stimuli in the unattended ear. There was no significant difference in the number of “intrusion” and “nonintrusion” errors made by the four groups (x2(3) = .96; p > JO).
146 JASON BRANDT
There were no group x ear interactions for either hit rate (F(3, 40) = .74; N.S.) or false positive rate (F(3, 40) = .14; N.S.).
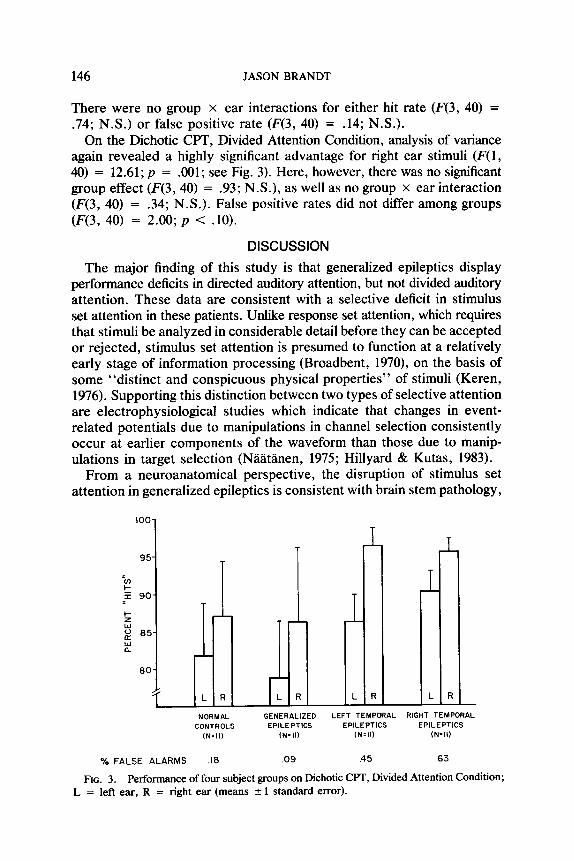
On the Dichotic CPT, Divided Attention Condition, analysis of variance again revealed a highly significant advantage for right ear stimuli (F(1, 40) = 12.61; p = .OOl; see Fig. 3). Here, however, there was no significant group effect (F(3,40) = .93; N.S.), as well as no group x ear interaction (F(3, 40) = .34; N.S.). False positive rates did not differ among groups (F(3, 40) = 2.00; p < .lO).
DISCUSSION
The major finding of this study is that generalized epileptics display performance deficits in directed auditory attention, but not divided auditory attention. These data are consistent with a selective deficit in stimulus set attention in these patients. Unlike response set attention, which requires that stimuli be analyzed in considerable detail before they can be accepted or rejected, stimulus set attention is presumed to function at a relatively early stage of information processing (Broadbent, 1970), on the basis of some “distinct and conspicuous physical properties” of stimuli (Keren, 1976). Supporting this distinction between two types of selective attention are electrophysiological studies which indicate that changes in event- related potentials due to manipulations in channel selection consistently occur at earlier components of the waveform than those due to manip- ulations in target selection (NZitanen, 1975; Hillyard & Kutas, 1983).
From a neuroanatomical perspective, the disruption of stimulus set attention in generalized epileptics is consistent with brain stem pathology,
IOO-
i L R
NORMAL GENERALIZED LEFT TEMPORAL RIGHT TEMPORAL CONTROLS EPILEPTICS EPILEPTICS EPILEPTICS
(N=ll) (N=II) (N=II) (N=II)
% FALSE ALARMS .I8 .09 .45 .63
FIG. 3. Performance of four subject groups on Dichotic CPT, Divided Attention Condition; L = left ear, R = right ear (means 2 1 standard error).

ATTENTIONAL DEFECT IN EPILEPSY 147
since afferents from the two ears are first combined at a brain stem nucleus, the superior olivary complex (Harrison & Howe, 1975). The recent report that an efferent pathway from the superior olivary complex to the cochlear nucleus attenuates irrelevant auditory information during selective attention (Lukas, 1981) lends strong support to the suggestion that significant filtering of sensory information is accomplished at the level of the brain stem. It has also recently been demonstrated that the midbrain reticular formation (together with a mediothalamic-frontocortical attentional network) has a major regulatory influence on the gating or modulation of sensory channels by the thalamic reticular nucleus (Skinner & Yingling, 1977).
The present report appears to be the first observation that patients with generalized seizures and bilaterally symmetrical EEG disturbances are deficient in their ability to selectively attend to stimuli on one side of space. Such a behavioral deficit is typically associated with unilateral lesions of the cerebral cortex, particularly of the right hemisphere (Brain, 1941; Weinstein & Friedland, 1977). However, similar deficits have also been found with brain stem lesions in animals. For example, Watson, Heilman, Miller, and King (1974) reported that unilateral lesions of the mesencephalic reticular formation in monkeys produce profound neglect of contralateral stimuli in all modalities, often accompanied by the EEG changes typical of absence epilepsy (three-per-second spike-wave dis- charges). To the extent that patients with generalized seizures are believed to have bilateral midbrain pathology, the experimental data reported here are consistent with Watson’s observation. However, why the behavioral defect emerges only when the subject must ignore one entire channel of irrelevant information (the Directed Attention Condition) remains un- answered. Clearly, it is an attention switching mechanism, rather than an “attentiveness” mechanism, which is impaired in these patients.
This study also finds that the dichotic REA can be obtained even on monitoring tasks which place minimal demands on memory and response mechanisms. This is noteworthy in that the work of several investigators (Satz, Achenbach, Pettishall, & Fennel, 1965; Shankweiler & Studdert- Kennedy, 1967; Goodglass & Peck, 1972) suggests that the demands of auditory short-term memory and verbal responding are responsible for the REA. Kimura’s (1967) observation that the REA is obtained even with single channel report (thereby minimizing memory demands) and Springer’s (1971) observation of a REA on dichotic monitoring of con- sonant-vowel syllables support the conclusion that the dichotic ear ad- vantage is perceptual, rather than mnemonic, in nature.
The data reported here provide no support for the hypothesis that the right hemisphere is superior to the left at detecting infrequent sensory events (Dimond, 1976, 1979a, 1979b) or, in fact, “dominates” behavior in sustained attention tasks (Warm, Schumsky, & Hawley, 1976; Warm,

148 JASON BRANDT
Richter, Sprague, Porter, & Schumsky, 1980). On both dichotic tasks, for all groups (including that with left hemisphere pathology), more right ear (left hemisphere) stimuli were detected. However, a variety of factors, including the verbal nature of our stimuli (Springer, 1971), their dichotic presentation (Kimura, 1967), the high signal density (Warm et al., 1976), and the short length of the vigil (Warm et al., 1980), may combine to explain the apparent discrepancy between the current findings and those cited above.
The present findings demonstrate that the REA is a relatively robust phenomenon, being clearly unaltered by the presence of epileptic foci in the left or right temporal lobe. This confirms several earlier reports for nonsurgical temporal lobe epileptics (Kimura, 1%1; Oxbury & Oxbury, 1969; Wale & Geffen, 1981; Brandt, 1982). It would appear, then, that the REA for verbal dichotic stimuli reflects left hemisphere language representation and is relatively insensitive to all but the grossest lateralized neuropathology (Schulhoff & Goodglass, 1969; Damasio & Damasio, 1979).
The conclusions reached in this study are, of course, subject to con- firmation by replication. In addition, several limitations of this research must be acknowledged. Due to the small sample size, for example, patients with several types of generalized seizures (e.g., absence, tonic-clonic, myoclonic) and a variety of EEG abnormalities were included in a single group. It is believed that a variety of neuropathologic conditions underlie these various seizure disorders (Lockard & Ward, 1980). Additionally, there may be important differences within the complex-partial groups depending on the nature of the discharges (e.g., spike, sharp wave, slow wave) and their precise origins in the temporal region. However, the relative heterogeneity of our patient groups makes our ability to find intergroup differences even more noteworthy. Nonetheless, further studies with more precisely defined patient samples are clearly indicated.
It is also possible that some patients (particularly in the generalized group) were having subclinical seizures during testing. Concurrent EEG recording may be helpful in this regard. It could not, however, rule out paroxysmal discharging from deep structures undetected by scalp electrodes.
Mention should also be made of the possible role of different anti- convulsant drug regimens. The only anticonvulsant which was taken by a different number of patients in the three groups was sodium valproate (see Table 1). One of the few neuropsychological studies of this drug found few significant correlations between serum level and neuropsy- chological test performance and suggested that sodium valproate “may be the least behaviorally toxic of major anticonvulsants studied” (Hartlage & Mains, 1981). Moreover, while some anticonvulsants may have a generalized depressing effect on sustained attention, there is no reason to believe that they would selectively disrupt stimulus set attention.

ATTENTIONAL DEFECT IN EPILEPSY 149
Finally, this study examined only one type of stimulus set. Channels of information were here defined by the ear which received the information. The present set of data do not allow us to assume that channels defined in ways other than spatial separation would yield similar results. It would be most instructive to determine, for example, whether generalized seizure patients are also unable to accurately detect target stimuli spoken by a woman in a series of mixed male-spoken and female-spoken digits. (Thus, channels would be defined by the frequency range of the voices.) Such a finding would allow us to assert more strongly that the processing deficit in generalized epileptics is one of stimulus set attention per se, and not peculiar to the specific stimuli and procedures of the present study. An experiment of this type is currently being planned.
REFERENCES Brain. W. R. 1941. Visual disorientation with special reference to lesions of the right
cerebral hemisphere. Brain. 64, 244-272. Brandt, J. 1982. Disorders of uuditory attention in epileptic patients: Studies using the
dichotic listening technique. Unpublished doctoral dissertation, Boston University. Broadbent, D. E. 1970. Stimulus set and response set: Two kinds of selective attention.
In D. 1. Mostofsky (Ed.), Attention: Contemporary theory and analysis. New York: Appleton-Century-Crofts.
Damasio, H., & Damasio, A. 1979. “Paradoxic” ear extinction in dichotic listening: Possible anatomic significance. Neurology, 29, 644-653.
Dimond, S. J. 1976. Depletion of attentional capacity after total commissurotomy in man. Brain, 99, 347-356.
Dimond, S. J. 1979. Performance by split-brain humans on lateralized vigilance tasks. Cortex, 15, 43-50. (a)
Dimond, S. J. 1979. Tactual and auditory vigilance in split-brain man. Journal of Neurology, Neurosurgery, and Psychiatry, 42, 70-74. (b)
Fedio, P., & Mirsky. A. F. 1969. Selective intellectual deficits in children with temporal lobe or centrencephalic epilepsy. Neuropsychologia, 7, 287-300.
Gloor, P. 1968. Generalized cortico-reticular epilepsies. Some considerations on the patho- physiology of generalized bilaterally synchronous spike and wave discharge. Epilepsia, 9, 249-263.
Gloor, P. 1979. Generalized epilepsy and spike-wave discharge: A reinterpretation of its electrographic and clincial manifestations. Epilepsia, 20, 571-588.
Glosser, G., Butters, N., & Samuels, I. 1976. Failures in information processing in patients with Korsakoff’s Syndrome. Neuropsychologia, 14, 327-334.
Goodglass, H., & Peck, E. A. 1972. Dichotic ear order effects in Korsakoff and normal subjects. Neuropsychologia, 10, 211-217.
Harrison, J. M., & Howe, M. E. 1975. Anatomy of the afferent auditory nervous system of mammals. In W. D. Keidel & W. D. Neff (Eds.), Handbook of sensory physiology: Vol. 5. Auditory system. New York: Springer.
Hartlage, L. C., & Mains, M. R. 1981. Anticonvulsant medication as a detriment of neuropsychological test projiles. Poster presented at the Annual Meeting of the In- ternational Neuropsychological Society, February.
Hillyard, S. A. & Kutas, M. 1983. Electrophysiology of cognitive processing. Annual Review of Psychology, 34, 33-61.
International League Against Epilepsy. 1981. Proposal for revised clinical and electroen- cephalographic classification of epileptic seizures. Epilepsia, 22, 489-501.

150 JASON BRANDT
Jasper, H. H. (Ed.). 1957. Reticular formation of the bruin. London: Churchill. Keren, G. 1976. Some considerations of two alleged kinds of selective attention. Journal
of Experimental Psychology: General, 105, 349-374. Kimura, D. 1961. Some effects of temporal-lobe damage on auditory perception. Canadian
Journal of Psychology, 15, 156-165. Kimura, D. 1967. Functional asymmetry of the brain in dichotic listening. Cortex, 3, 163-
178. Kinsboume, M. 1973. The control of attention by the interaction between the cerebral
hemispheres. In S. Komblum (Ed.), Attention andperformance. New York: Academic Press. Vol. 4.
Lansdell, H., & Mirsky, A. F. 1964. Attention in focal and centrencephalic epilepsy. Experimental Neurology, 9, 463-469.
Lockard, J. S., & Ward, A. A. (Eds.) 1980. Epilepsy: A window to brain mechanisms. New York: Raven Press.
Lukas, J. H. 1981. The role of efferent inhibition in human auditory attention: An examination of the auditory brain potentials. International Journal of Neuroscience, 12, 137-145.
Luria, A. R. 1973. The working bruin: An introduction to neuropsychology. New York: Basic Books.
Mazzucchi, A., & Parma, M. 1978. Responses to dichotic listening tasks in temporal epileptics with and without clinically evident lesions. Cortex, 14, 381-390.
Mesulem, M.-M. 1981. A cortical network for directed attention and unilateral neglect. Annals of Neurology, 10, 309-325.
Mirsky, A. F., & Van Buren, J. M. 1965. On the nature of the “absence” in centrencephalic epilepsy: A study of some behavioral, electroencephalographic, and autonomic factors. Electroencephalography and Clinical Neurophysiology, 18, 334-348.
NUtlnen, R. 1975. Selective attention and evoked potentials in humans: A critical review. Biological Psychology, 2, 237-307.
Oxbury, J. M., & Oxbury, S. M. 1969. Effects of temporal lobectomy on the report of dichotically presented digits. Cortex, 5, 3-14.
Penlield, W., & Jasper, H. 1954. Epilepsy and the functional anatomy of the human brain. Boston: Little Brown.
Posner, M. I. 1975. Psychobiology of attention. In M. S. Gazzaniga & C. Blakemore (Eds.) Handbook of psychobiology. New York: Academic Press.
Satz, P., Achenbach, K., Pattishall, E., & Fennel, E. 1965. Order of report, ear asymmetry, and handedness in dichotic listening. Cortex, 1, 377-396.
Schulhoff, C., & Goodglass, H. 1%9. Dichotic listening, side of brain injury, and cerebral dominance. Neuropsychologia, 7, 14-160.
Shankweiler, D., & Studdert-Kennedy, M. 1967. Identification of consonants and vowels presented to left and right ears. Quarterly Journal of Experimental Psychology, 19, 59-63.
Skinner, J. E., 8c Yingling, C. D. 1977. Central gating mechanisms that regulate event- related potentials and behavior. In J. E. Desmedt (Ed.), Program in clinical neuro- physiology: Vol. 1: Attention, voluntary contraction and event-related potentials. Basel: Karger.
Springer, S. P. 1971. Ear asymmetry in a dichotic detectiion task. Perception and Psy- chophysics, 10, 239-241.
Wale, J., t Geffen, G. 1981. Dichotic monitoring as a test of hemispheric dominance in cases of epilepsy. Cortex, 17, 135-140.
Warm, J. S., Schumsky, D. A., & Hawley, D. K. 1976. Ear asymmetry and the temporal uncertainty of signals in sustained attention. Bulletin of the Psychonomic Society, 7, 413-416.

ATTENTIONAL DEFECT IN EPILEPSY 151
Warm, .I. S., Richter, D. O., Sprague, R. L., Porter, P. K., & Schumsky, D. A. 1980. Listening with a dual brain: Hemispheric asymmetry in sustained attention. Bulletin of the Psychonomic Society, 15, 229-232.
Watson, R. T., Heihnan, K. M., Miller, B. D., & King, F. A. 1974. Neglect after mesencephalic reticular formation lesions. Neurology, 24, 294-298.
Watson, R. T., Miller, B. D., & Heilman, K. M., 1978. Nonsensory neglect. Annuls of Neurology, 3, 505-508.
Weinstein, E. A., & Friedland, R. P. (Eds.) 1977. Advances in Neurology: Vol. 18. Hemi- inattention and hemisphere specialization. New York: Raven Press.




















