
THE DIETS OF MODERN AND HISTORIC BOTTLENOSE DOLPHIN POPULATIONS REFLECTED THROUGH STABLE ISOTOPES
16
MARINE MAMMAL SCIENCE, 15(2):335-350 (April 1999) 0 1999 by the Society for Marine Mammalogy THE DIETS OF MODERN AND HISTORIC BOTTLENOSE DOLPHIN POPULATIONS REFLECTED THROUGH STABLE ISOTOPES JANE L. WALKER Department of Environmental Sciences, University of Virginia, Charlottesville, Virginia 22903, U.S.A. CHARLES W. POTTER Marine Mammal Program, Smithsonian Institution, Washington, DC 20560, U.S.A. STEPHEN A. MACKO Department of Environmental Sciences, University of Virginia, Charlottesville, Virginia 22903, U.S.A. ABSTRACT The 6I3C and WN compositions of teeth used in combination with exist- ing data provide dietary information for different populations of western North Atlantic bottlenose dolphins (Ttlrsiops truncutus). The dental isotopic signatures of bottlenose dolphins collected during the 1980s significantly differ for coastal and offshore ecotypes and are consistent with reports that coastal forms feed primarily on fish whereas offshore individuals consume more squid. In a second study, the isotopic compositions of teeth from bot- tlenose dolphins that span a 100-yr period and data from published stomach content analyses as well as field observations made during the past 100 yr provide evidence that coastal bottlenose dolphins from the 1880s, 1920s, and 1980s had similar diets. Key words: bottlenose dolphin, Tursiops truncattls, carbon isotopes, nitrogen isotopes, diet. The carbon and nitrogen isotopic signatures of an organism provide valuable dietary information. One advantage to using the stable isotope technique is the documentation of nutrients assimilated and not simply those ingested. Because small amounts (5-10 mg) of body components, such as blood, hair, whiskers, or nails, contribute enough organic material for isotopic analysis, samples can be humanely obtained from living specimens. In addition, samples 335
Transcript of THE DIETS OF MODERN AND HISTORIC BOTTLENOSE DOLPHIN POPULATIONS REFLECTED THROUGH STABLE ISOTOPES

MARINE MAMMAL SCIENCE, 15(2):335-350 (April 1999) 0 1999 by the Society for Marine Mammalogy
THE DIETS OF MODERN AND HISTORIC BOTTLENOSE DOLPHIN POPULATIONS
REFLECTED THROUGH STABLE ISOTOPES JANE L. WALKER
Department of Environmental Sciences, University of Virginia,
Charlottesville, Virginia 22903, U.S.A.
CHARLES W. POTTER Marine Mammal Program,
Smithsonian Institution, Washington, DC 20560, U.S.A.
STEPHEN A. MACKO Department of Environmental Sciences,
University of Virginia, Charlottesville, Virginia 22903, U.S.A.
ABSTRACT
The 6I3C and W N compositions of teeth used in combination with exist- ing data provide dietary information for different populations of western North Atlantic bottlenose dolphins (Ttlrsiops truncutus). The dental isotopic signatures of bottlenose dolphins collected during the 1980s significantly differ for coastal and offshore ecotypes and are consistent with reports that coastal forms feed primarily on fish whereas offshore individuals consume more squid. In a second study, the isotopic compositions of teeth from bot- tlenose dolphins that span a 100-yr period and data from published stomach content analyses as well as field observations made during the past 100 yr provide evidence that coastal bottlenose dolphins from the 1880s, 1920s, and 1980s had similar diets.
Key words: bottlenose dolphin, Tursiops truncattls, carbon isotopes, nitrogen isotopes, diet.
The carbon and nitrogen isotopic signatures of an organism provide valuable dietary information. One advantage to using the stable isotope technique is the documentation of nutrients assimilated and not simply those ingested. Because small amounts (5-10 mg) of body components, such as blood, hair, whiskers, or nails, contribute enough organic material for isotopic analysis, samples can be humanely obtained from living specimens. In addition, samples
335

336 MARINE MAMMAL SCIENCE, VOL. 1 5 , NO. 2, 1999
for isotopic analysis can be acquired from the teeth and bones of museum specimens so that historic diets can be studied. The use of tooth samples has an additional benefit in that short-term and long-term diets can be examined from the same sample.
The premise of the isotopic technique lies in the metabolic discriminations between the heavy and light isotopes. Laboratory studies have shown that isotopic compositions of modern organisms reflect the nutritional sources of the animal (DeNiro and Epstein 1978, 1981; Macko et al. 1982, 1987; T' ieszen etal. 1983; Hobson et al. 1996). DeNiro and Epstein (1978, 1981) determined that animals were more enriched by about 1%0 for 13C and approximately 3%0 for lSN when compared to the isotopic compositions of the diet.l Hobson et al. (1996) utilized carbon and nitrogen isotopic signatures of various body components of captive seals to provide applicable information for use in future dietary reconstructions.
Isotopic enrichment of an animal in relation to its food has been reported from field studies (McConnaughey and McRoy 1979, Gearing et al. 1984, Minagawa and Wada 1984, Fry 1988, Schell et al. 1989, Abend and Smith 1995, Smith et al. 1996). Schell et al. (1989) documented seasonal changes in the feeding behavior of bowhead whales (Balaena mysticetus) from the carbon isotopic signatures of baleen. Stable isotopic ratios of nitrogen in skin, muscle, blubber, and teeth were used to study the diets of different populations of long-finned pilot whales inhabiting the eastern and western North Atlantic (Abend and Smith 1995). Additionally, carbon and nitrogen isotopic infor- mation derived from hair samples of harbor seals was utilized to determine if freshwater or marine prey are more important to the diets of three distinct seal populations (Smith et al. 1996).
Isotopic analyses have also been successfully used with well-preserved animal fossils to show dietary patterns (Nelson et al. 1986; DeNiro and Weiner 1988; Tuross et al. 1988; Ostrom et al. 1989, 1993a; Hare et al. 1991; Hobson and Montevecchi 1991). Tuross et al. (1988) found the carbon and nitroge'n isotopic values of individual amino acids from fossilized whale bone to be similar to those of modern whale bone. If isotopic preservation can occur in fossils, it should be possible to also obtain historic dietary information from museum specimens collected within the past several centuries.
This study uses carbon and nitrogen isotopic signatures to examine the diets of modern and historic bottlenose dolphins (Tursiops trunsatus). It is unique because no isotopic research has utilized teeth from different populations of dolphins in order to assess their diets. In addition, this work is the first to use isotopic signatures from a single species collected at a particular location over an extended time period to determine possible dietary shifts.
Offshore and coastal populations-Several characteristics separate dolphins in- habiting cool, deep water (usually offshore) and those living in warm, shallow
' Isotopic compositions are expressed as: fiNE = [(Rsamplr/Rrrandard) - 11 X 1,000 where N is the heavy isotope of element E and R is the abundance ratio of the heavy to light isotope. Enriched values are more positive (heavier), and depleted signatures are more negative (lighter).

WALKER ETAL.: BOTTLENOSE DOLPHIN DIET 337
water. Northwest Atlantic bottlenose dolphins living in offshore waters appear genetically different from coastal Northwest Atlantic and Gulf of Mexico bot- tlenose dolphins in that the offshore dolphins have two types of hemoglobin whereas the coastal animals have only one (Hersh and Duffield 1990). A study of captive Atlantic and Pacific bottlenose dolphins suggest that offshore ani- mals and those inhabiting cool, deep water have increased concentrations of hemoglobin, packed cell volumes, and red blood cells-qualities necessary for the higher oxygen demand required of deeper dives (Duffield et al. 1983). The coastal animals are generally smaller and have proportionally larger flippers for more efficient movement and heat dissipation. Size differences for the skulls and rostrums of the two ecotypes may be correlated with the types of foods eaten (Hersh and Duffield 1990, Mead and Potter 1995). In this study stable isotopic ratios of teeth from coastal and offshore specimens of the western North Atlantic were compared.
18805, 1920s, and 1980s populations-Natural and anthropogenic changes have occurred within the coastal waters of the western North Atlantic during the past 100 yr that could have altered the diets of coastal bottlenose dolphins. Overfishing by humans, increased chemical pollution, and diseases are merely three examples of things that could have affected the prey of coastal bottlenose dolphins within the past century. A significant change in the populations of the food sources for dolphins would result in alterations of the diets for these coastal marine mammals and could be expressed in the isotopic signatures of their teeth. Therefore, the carbon and nitrogen isotopic compositions of teeth from coastal bottlenose dolphins captured in the western North Atlantic over a 100-yr period were compared.
METHODS
Tooth samples were taken from dolphin skulls collected by F. W. True in October 1884 and by A. J. Poole and A. R. Kellogg in May 1927. The specimens were collected at a shore-based drive fishery in Cape Hatteras, North Carolina. Because workers at this fishery frequently netted entire schools of coastal dolphins (Mead 1975), the animals captured at the same time for a given year may have been members of the same school. The gender, body length, and body weight of the specimens were not recorded. Stomach contents were not collected or analyzed. Also, the methods used to clean the skulls of these bottlenose dolphins were not documented.
Three modern specimens (571385, 571426, and 571439) were collected offshore from incidental catches in the western North Atlantic during 1989 and 1990. The remaining modern dolphin specimens came from stranded animals. These dolphins were classified as coastal or offshore based on mor- phometry, food habits, and internal parasite loads as described by Mead and Potter (1995). The teeth determined to be from coastal dolphins were stranded from Maryland to North Carolina. One individual was beached in 1979 and another in 1981. The other four were stranded on different days in 1987. The

338 MARINE MAMMAL SCIENCE, VOL. 15, NO. 2 , 1999
stranded offshore animals were collected between 1971 and 1992 from Mas- sachusetts to South Carolina.
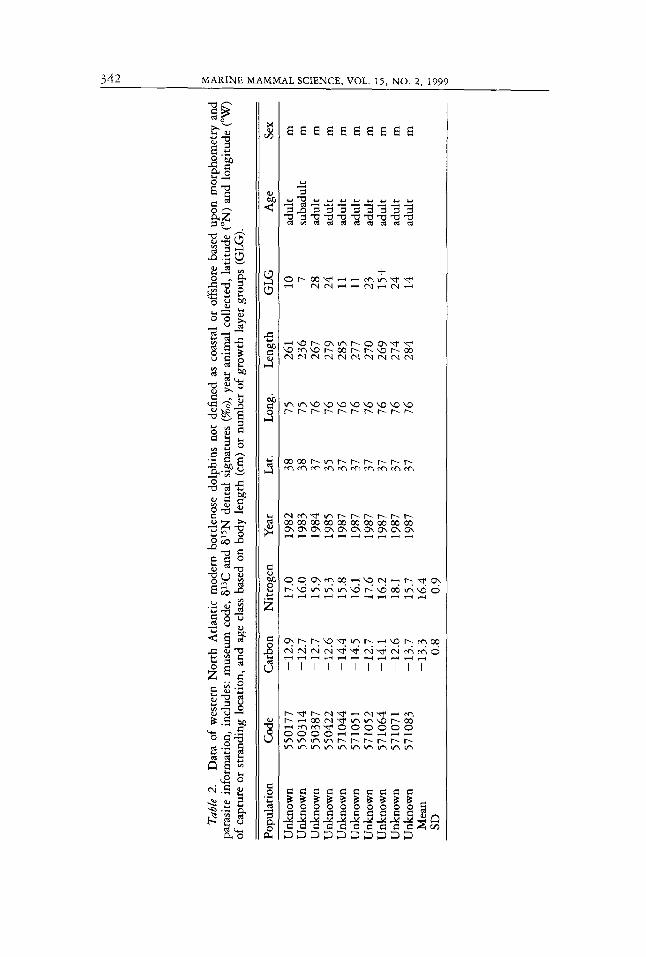
Not all stranded bottlenose dolphins from the western North Atlantic were able to be categorized into coastal or offshore populations based upon mor- phometry, food habits, and parasite information. The carbon and nitrogen isotopic signatures of teeth from ten stranded specimens whose population origin could not be determined with certainty were analyzed. They are noted as unknowns for the appropriate Table and Figure.
The modern specimens were defleshed by hand, allowed to dry for one or two days, and then stored with dermestid beetles (Dmestes spp.) for several weeks to months. Following this procedure, the skulls were washed in am- monium solution to clean the surface and rinsed. Excess organic material, such as cartilage, was removed from modern specimens by a solution of trypsin kept at 38°C for one or two days. The skulls were copiously rinsed in running water for several hours and dried at room temperature.
Containing both carbon and nitrogen, trypsin has the potential to contam- inate the isotopic signatures of the treated samples. Trypsin also has the ability to enzymatically digest non-collagenous and collagenous proteins found in bone and so could cause isotopic fractionation within the teeth. To test for the effects of trypsin, an isotopic comparison was made between one dolphin tooth that was cleaned using dermestid beetles and an adjacent tooth that was treated with beetles and trypsin. A sample of the trypsin was isotopically analyzed (6I3C = -20.1%0, 615N = 5.9%0) and found to be considerably depleted relative to the teeth. The two tooth samples gave similar results (Dermestid Treatment: S13C = -14.2%0, S1>N = 14.9%0; Dermestid and Trypsin Treatment: 613C = -14.0%0, 615N = 14.7%0), suggesting that the trypsin did not affect the isotopic values of the dental tissues.
Each tooth was cleaned of outer material using abrasion and broken into large pieces. The dried pulp was removed and discarded. The remaining min- eralized components of the tooth were soaked in chilled 30% hydrochloric acid (HCl), rinsed with deionized water, dried, and crushed to a fine powder. The sample, which consisted of 5-10 mg of the powdered material, was com- busted in an evacuated quartz tube in the presence of copper and copper oxide at 850°C for one hour and allowed to cool slowly. The resulting N, was cryogenically isolated using a molecular sieve and liquid nitrogen, and the CO, was separated from the other combustion products (e.g., H,O and SO,) with a dry ice/ethanol slurry. The S1>N and S13C compositions of the N, and CO,, respectively, were assessed on a dual-inlet, triple collector VG PRISM stable-isotope ratio mass spectrometer. The sample gases were compared with laboratory tank standards that were calibrated against the international stan- dards of Peedee Belemnite (PDB) for carbon and atmospheric N Z for nitrogen. The typical overall errors for preparation and measurement of the 613C and 615N analyses were +0.1%0-+0.2%0.
The Statistical Package for the Social Sciences (SPSS) was utilized in the analysis of the data. Multivariate analysis of variance (MANOVA) tested for significant population differences for the two variables, 613C and sL5N. The

WALKER E T A L . : BOTTLENOSE DOLPHIN DIET 339
statistically analyzed data were reported in terms of F,,n (the transformed value of the test statistic for an approximate F distribution with m degrees of free- dom for the hypothesis and n degrees of freedom for the error) and P, which indicates the observed significance level. The power to detect differences be- tween populations was also determined (assuming a significance level of 0.05).
RESULTS
Isotopic signatures of offsshore and coastal populations-The six adult males de- termined to be of the modern coastal population had teeth with a mean 813C value of -13.0 5 0.7%0 and an average 615N signature of 16.8 2 0.9%0 (Table 1, Fig. 1). The bottlenose dolphins of the offshore ecotype had teeth with a mean P C value of -13.9 2 0.4%0 and an average 615N of 14.8 +0.8%0 (n = 9 , Table 1 , Fig. 1) . A comparison of the average isotopic com- positions for these two populations revealed that they differ significantly (FZ,12 = 13.50, P = 0.001, power = 99%). The teeth from the offshore animals were more depleted in I 3 C and lSN than were those of the coastal specimens.
Of the ten specimens that could not be categorized as either coastal ot offshore, three (550177, 571052, 571071) isotopically resembled the coastal bottlenose dolphins in '5N (Table 1 , 2 ; Fig. la ) . The results of the seven remaining specimens were istopically intermediate in 15N to most of the OR- shore and coastal animals (Table 2, Fig. la).
lsotopic signatures of bottlenose dolphins spanning 1 00 years-A comparison of the data from the three groups of coastal bottlenose dolphins spanning a 100- yr time period showed no statistically significant separations (F4,34 = 1.1 1 , P = 0.369, power = 31%). The mean isotopic compositions are progressively enriched from the animals captured in the 1880s (mean S13C = -13.4 t 0.3%0, mean 615N = 15.9 2 0.8%0, n = 6 ) through the 1920s (mean 6I3C = -13.1 2 0.8%0, mean V5N = 16.3 +-0.5%0, n = 8) to the 1980s (mean S13C = -13.0 ? 0.7%0, mean S15N = 16.8 5 0.9%0, n = 6 , Table 3, Fig. 2). This trend in the averages and the low power of the test call for isotopic analysis of additional samples. The capability for further isotopic study, how- ever, may not be possible unless other samples from these 100-yr old popu- lations exist.
DISCUSSION
The isotopic signatures in this study represented the long-term diets of the specimens because each sample was taken from the powder of an entire tooth. Dentin is continually deposited within a tooth until the pulp cavity is filled, which takes many years, and is therefore the major constituent of the tooth and the isotopic sample. By using a sample that incorporated all of the dentinal layers, variations in diet-such as those that occur with the age of the indi- vidual and seasonal prey fluctuations-were minimized, yielding an average dental isotopic value for the animal.
The 613C and 615N signatures reflect ingested nutrition, with the 6l5N

Tabl
e I.
D
ata
of m
oder
n bo
ttlen
ose
dolp
hins
rep
rese
ntin
g di
ffer
ent
popu
latio
ns f
rom
wes
tern
Nor
th A
tlant
ic,
incl
udes
: m
useu
m c
ode,
S1
3C an
dS'5
N d
enta
l si
gnat
ures
(%
& ye
ar a
nim
al c
olle
cted
, lat
itude
("N
) and
lon
gitu
de (
OW
) of
capt
ure
or s
trand
ing
loca
tion,
and
age
cla
ss
base
d on
bod
y le
ngth
(cm
) or
num
ber
of g
row
th la
yer g
roup
s (G
LG).
Popu
latio
n C
ode
Car
bon
Nitr
ogen
Y
ear
Lat.
Long
. Le
ngth
G
LG
Age
Se
x ~
~~
~~~~
~
Coa
stal
50
4961
-1
2.9
15.3
19
79
36
76
272
32
adul
t m
C
oast
al
Coa
stal
co
asta
l C
oast
al
Coa
stal
M
ean
SD
Off
shor
e O
ffsh
ore
Off
shor
e O
ffsh
ore
Off
shor
e O
ffsh
ore
Off
shor
e O
ffsh
ore
Off
shor
e M
ean
SD
5500
95
5710
17
5710
24
5710
26
5710
68
5047
66
5048
79
5049
06
5503
75
5504
46
5713
85
5714
26
5714
39
57 1
496
-13.
0 -1
4.1
-12.
3 -1
2.3
- 13
.3
-13.
0 0.7
- 14
.4
-13.
7 -1
3.1
-13.
5 -1
3.7
- 14
.2
- 14
.0
- 14
.2
- 14
.2
-13.
9 0.4
16.7
16
.5
17.8
17
.1
17.3
16
.8
0.9
14.4
14
.8
13.6
15
.6
15.5
15
.5
13.7
14
.4
15.6
14
.8
0.8
1981
19
87
1987
19
87
1987
1971
19
79
1979
19
84
1984
19
89
1990
19
90
1992
38
36
37
37
37
36
36
37
41
42
40
41
37
39
75
75
76
76
76
76
75
79
72
70
67
66
75
74
277
264
281
270
267
309
284
288
310
279
159
293
174
202
22
adul
t ad
ult
22
adul
t 13
ad
ult
8 ad
ult
adul
t ad
ult
28
adul
t ad
ult
24
adul
t 0.
3 Y
oung
ad
ult
youn
g 0.
8 su
badu
lt
m
m
m
m
m
m
m
m
m
m
m
m
m
m

WALKER ETAL.: BOTTLENOSE DOLPHIN DIET 34 1
19 . . . . I . . . ~ I ~ ~ . . I ~ . ~ ~ I ~ . . . i ' . . .
18 - -
T - 1
+*----I Coastal 17 - - 16 - 6
z 2 u3
T 1
+*+ Offshore 15 -
14 -
13 . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
value providing more useful trophic level information (DeNiro and Epstein 1978, 1981; Minagawa and Wada 1984; Schoeninger and DeNiro 1984; Fry 1988). Deriving dietary information for high-trophic-level consumers from isotopic ratios is complex, however. It is possible for two populations to con- sume different prey yet have similar mean isotopic compositions. This occur- rence is less likely when two or more elements are used as isotopic tracers. Alternatively, populations from different geographic locations with dissimilar isotopic signatures may have similar diets, depending on the isotopic com- positions of their prey. Squid ( I l lex illecebroszls > 24.5 cm in length) from the coastal waters of Newfoundland were found to differ in their WSN composi- tions (1 5.1%0) compared to smaller 1. illecebroszls (9.3%0) collected offshore in the Grand Banks region (Ostrom et al. 19936).
Tabl
e 2.
D
ata
of w
este
rn N
orth
Atla
ntic
mod
ern
bottl
enos
e do
lphi
ns n
ot d
efin
ed a
s co
asta
l or
off
shor
e ba
sed
upon
mor
phom
etry
and
pa
rasi
te i
nfor
mat
ion,
inc
lude
s: m
useu
m c
ode,
6l3
C a
nd S
1’N
dent
al s
igna
ture
s (%
o),
year
ani
mal
col
lect
ed, l
atitu
de (O
N) a
nd l
ongi
tude
(”w
) of
cap
ture
or
stra
ndin
g lo
catio
n, a
nd a
ge c
lass
bas
ed o
n bo
dy l
engt
h (c
m) o
r nu
mbe
r of
gro
wth
laye
r gro
ups
(GL
G).
Popu
latio
n C
ode
Car
bon
Unk
now
n U
nkno
wn
Unk
now
n U
nkno
wn
Unk
now
n U
nkno
wn
Unk
now
n U
nkno
wn
Unk
now
n U
nkno
wn
Mea
n SD
5501
77
5503
14
5503
87
5504
22
5710
44
5710
51
5710
52
5710
64
5710
71
5710
83
-12.
9 - 12
.7
- 12
.7
-12.
6 - 1
4.4
- 1
4.5
-12.
7 -1
4.1
-12.
6 -1
3.7
-13.
3 0.8
Nitr
ogen
17.0
16
.0
15.9
15
.3
15.8
16
.1
17.6
16
.2
18.1
15
.7
16.4
0.
9
Yea
r La
c.
1982
19
83
1984
19
85
1987
19
87
1987
19
87
1987
19
87
38
38
37
35
37
37
37
37
37
37
Long
.
75
75
76
76
76
76
76
76
76
76
Leng
th
261
236
267
279
285
277
270
269
274
284
GLG
A
ge
Sex
10 7 28
24
11
11
23
15+
24
14
adul
t m
m
su
badu
lt ad
ult
m
adul
t m
ad
ult
m
adul
t m
ad
ult
m
adul
t m
ad
ult
m
adul
t m
w

Tabl
e 3.
Dat
a of
coa
stal
bot
tleno
se d
olph
ins
colle
cted
ove
r 10
0 yr
, inc
lude
s: m
useu
m c
ode,
813
C an
d 81
5N d
enta
l si
gnat
ures
(%&
date
of
capt
ure
or s
tran
ding
, col
lect
ion
latit
ude ("N
) and
lon
gitu
de (O
W), a
nd a
ge c
lass
bas
ed o
n bo
dy l
engt
h (c
m) o
r nu
mbe
r of g
row
th la
yer g
roup
s (G
LG).
Suba
dult
mea
ns n
ot o
f nu
rsin
g ag
e an
d no
t se
xual
ly m
atur
e.
Dat
e C
ode
Car
bon
Nitr
ogen
Po
p.
Lat.
Long
. Se
x Le
ngth
G
LG
A
ge
10/?
/188
4 10
/?/1
884
1 0/?
/188
4 10
/?/1
884
10/?
/188
4 10
/?/1
884
Mea
n SD
5/
?1 19
27
5/?/
1927
5/
?/19
27
5/?1
1927
5/
?/19
27
51?/
1927
5/
?/19
27
5/?/
1927
M
ean
SD
5122
11 97
9 91
1 011
98 1
5/1/
1987
7/
61 19
87
7112
11 98
7 7/
20/1
987
Mea
n SD
2207
9 22
080
2208
1 22
082
2208
3 22
084
2521
03
2521
04
2521
05
2521
07
2521
12
2521
14
2521
16
2521
23
5049
61
5500
95
5710
17
5710
24
5710
26
5710
68
-13.
5 - 1
3.4
-13.
5 -1
3.8
- 13
.4
-13.
0 - 13
.4
-11.
7 - 1
3.4
- 1
4.5
-13.
3 - 1
2.9
-12.
8 -1
3.1
- 1
3.3
-13.
1 0.8
- 12
.9
- 1
3.0
-14.
1 - 12
.3
- 1
2.3
-13.
3 -1
3.0 0.7 0.3
15.9
15
.6
16.5
15
.2
15.2
17
.1
15.9
0.
8 16
.6
16.8
15
.5
16.3
16
.5
16.8
16
.2
15.9
16
.3
0.5
15.3
16
.7
16.5
17
.8
17.1
17
.3
16.8
0.
9
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
Coa
stal
C
oast
al
35
35
35
35
35
35
35
35
35
35
35
35
35
35
36
38
36
37
37
37
76
76
76
76
76
76
76
76
76
76
76
76
76
76
76
75
75
76
76
76
3.5
14
29 7 23
30
21
24
m
272
32
m
277
22
m
264
m
281
22
m
270
13
m
267
8
suba
dul t
adul
t ad
ult
adul
t ad
ult
adul
t ad
ult
adul
t ad
ult
adul
t ad
ult
adul
t ad
ult

344 MARINE MAMMAL SCIENCE, VOL. 15, NO. 2, 1999
19
18
1 7
0 h
16; z n co 15
14
1 3
0 1880's
0 1920's
A 1980's
. . . . l . . . . l . . . . , . . . . l . . . . l . . . .
- -
[ 19801s - -
%!iGls lmtski -
- -
- -
. . . . I . . . . I . . . . l , . . . l , . . . l . . . .
Isotopic variations for consumers from different regions may result from contrasting 613C and 615N compositions at the base of the respective food webs. Environmental differences such as light intensity, nutrient concentra- tions, and species composition affect the 613C and 615N values of the primary producers in a region (Wong and Sackett 1978, Gearing e t al. 1984, Macko e t al, 1984, Fogel and Cifuentes 1993). Sackett e t al. (1965) and Rau e t a/. (1989) found that the 6I3C signal of phytoplankton tends to be more depleted in higher latitudes. Enriched 6 ' W values have been reported for samples of suspended particulate organic matter collected below the photic zone in off- shore waters and along salinity transects in estuaries (Fogel and Cifuentes

WALKER E T A L . : BOTTLENOSE DOLPHIN DIET 345
1993). Influences from terrestrial inputs-such as fertilizer, sewage, and ter- restrial plant debris-can alter the isotopic composition at the base of the of the food web for coastal waters (Kohl et al. 1971, Sweeney et al. 1980, Fogel and Cifuentes 1993, Macko and Ostrom 1994, Hansson et al. 1997).
Despite several potential problems, useful dietary information can be ob- tained from isotopic compositions. Walker and Macko (1999) utilized the aL3C and 615N values of teeth from eight species of marine mammals from different geographic locations and trophic levels. The specimens were collected from the western North Atlantic and eastern North Pacific. The trophic position of the specimens ranged from the herbivorous manatee to the highly predaceous killer whale. The S'3C and SISN signatures of the marine mammal teeth were found to reflect the respective isotopic compositions of the diets.
Conclusions based on isotopic signatures for dietary differences and simi- larities of consumers from two potentially distinct environments need to be made carefully. When possible, it is best to have isotopic data from non- chemically preserved samples of the potential prey and primary producers collected from the inhabited geographic areas and time periods of the speci- mens. Unfortunately, samples for such supporting isotopic analyses are often not available, as was the case in this study. For this research, the isotopic data of bottlenose dolphin teeth were compared to published data of feeding ob- servations, stomach content analyses, and parasite examination studies for bot- tlenose dolphins taken from the same general areas and time periods as the isotopically analyzed specimens.
Isotopic signatzlres of offihore and coastal popdations-The offshore animals tended to be more depleted in 13C and '5N and were isotopically distinct from the coastal population (Table 1, Fig. 1). One interpretation of the results is that the diets differ for the offshore and coastal bottlenose dolphins. It is possible, however, that the depleted values correspond to an isotopically lighter food web and that the two ecotypes consume the same prey. Therefore, ad- ditional information about the diets o€ the populations or isotopic data for the base of the food webs for the respective areas are required.
The stomach content information for the specimens used in this study did not reveal much about the diets of the populations. Stomach contents were obtained for only two of the six coastal specimens. One stomach (504961) contained fish and a small squid, and the other specimen (571026) had fish bones. Intact squid and squid beaks were found in one of the offshore speci- mens (57 1426), and three other offshore individuals had squid beaks. Speci- men 571426 also had intact fish, fish bones, and otoliths. Three other offshore individuals had fish bones and/or otoliths. The two young offshore specimens (571385, 571439) had only milk in their stomachs.
Published stomach content analyses for coastal and offshore bottlenose dol- phins from the western North Atlantic indicate a more distinct difference in diets. A bottlenose dolphin determined to be of the offshore ecotype based on morphometry was found to have fed on squid (Illex sp. and Ornithotezlthis antillarzlm, Barros and Ode11 1990). Mercer (1973) analyzed the stomach con- tents of a female bottlenose dolphin captured in waters over 270 m deep off

346 MARINE MAMMAL SCIENCE, VOL. 15, NO. 2 , 1999
North Carolina and found the remains of one juvenile Anisakis sp., Illex ille- cebrosus (including over 70 upper and lower beaks), octopods (three sets of beaks and spermatophores), and an unidentified fish (an otolith). Stomach an- alyses for 18 offshore bottlenose dolphins from the western North Atlantic indicated they fed upon pelagic squid and fish, with the fish being primarily deep-water species of the family Myctophidae (Mead and Potter 1995). In contrast, the coastal form generally chose shoaling and bottom-dwelling fish. The stomachs of coastal animals from the eastern United States contained mainly sciaenid fish (drums or croakers): seatrout (Cynoscion spp.), spot (Leios- tomus xanthurus), and Atlantic croaker (Micropogon undulatus) (Barros and Ode11 1990; Mead and Potter 1990, 1995). Squid remains were found in only 20 of 117 stomachs collected from coastal bottlenose dolphins (Mead and Potter 1995).
The presence or absence of certain parasites can suggest a difference in diet because some parasites, such as species within Braunina, Phyl l~bo~hri~m, Mon- orhygma, and Crassicauda, are acquired through ingestion. Thus, if different dolphin populations have distinct diets, they may carry different parasites. Mead and Potter (1990, 1995) found coastal bottlenose dolphins to have Braunina but not Phyllobothrium, Monorhygma, and Crassicauda. Alternatively, offshore dolphins tended to have Phyllobothrium, Monorhygma, and Crassicauda but not Braunina. Thus, published comparisons of internal parasites also in- dicate a difference in diet for offshore and coastal bottlenose dolphins from the western North Atlantic (Mead and Potter 1990, 1995).
Based on the distinct isotopic values, published stomach content informa- tion, and published parasite data, we believe the diets of offshore and coastal populations of bottlenose dolphins from the western North Atlantic differ. The a15N signatures of the teeth of offshore bottlenose dolphins (mean = 14.8%0) more closely resemble those of organisms that consume large amounts of squid (long-finned pilot whales = 14.0%0, Abend and Smith 1995; pygmy sperm whales = 12.4%0, Walker and Macko 1999) and published 615N values for squid (1 1 .O%O, Fry 1988; 9 . 3 % ~ Ostrom et al. 19933). Consequently, we believe that squid comprise more of the diet for the offshore bottlenose dol- phins than they do for the coastal dolphins. In addition, the sciaenid fishes taken from the stomachs of coastal bottlenose dolphins tend to be benthic consumers of crustaceans and small fish (Mead and Potter 1990), and benthic organisms usually have enriched 813C and 615N values relative to pelagic or- ganisms (McConnaughey and McRoy 1979, Fry and Sherr 1984, France 1995). Fry (1 988) found planktivorous, opportunistic generalist, and piscivorous fish from Georges Bank to have mean 61SN values of 1 0 . 6 % ~ 12.2%0, and 13.6%0, respectively. These generally enriched 615N values for fish relative to those reported for small squid are consistent with the belief that coastal bottlenose dolphins consume more piscivorous fish.
Tooth samples from ten bottlenose dolphins that could not be classified as coming from a coastal or offshore population based on the morphological and ecological differences examined at the museum were isotopically analyzed. Stomach contents of three of these specimens (550387, 571051, 571083) were

WALKER E T A L . : BOTTLENOSE DOLPHIN DIET 347
analyzed and found to contain evidence of fish but not squid. The 613C and 6ISN isotopic compositions of three of these specimens (550177, 571052, 571071) look like those of the coastal bottlenose dolphins and may indicate that these specimens belong to the coastal population (Table 1, 2; Fig. la). The remaining seven specimens had 615N signatures between most of the offshore and coastal specimens, which supports the belief proposed by Mead and Potter (1995) that there may be more than two populations of bottlenose dolphins in the western North Atlantic (Table 1, 2; Fig. la). Thus, isotopic data may be useful in determining if an animal belongs to a coastal, offshore, or other population. Additional evidence is required, however, before definite conclusions can be made.
Isotopic signatares of bottlenose dolphins spanning 1 00 yr-The isotopic results in combination with published feeding observations and stomach content analyses indicate that the habitual diets of coastal bottlenose dolphins in the western North Atlantic were similar during the 1880s, 1920s, and 1980s. Overall comparisons of 613C and 6ISN values from the coastal bottlenose dol- phins collected for the three time periods appear similar, although the mean isotopic compositions have a slight, progressive enrichment from the 1880s to the 1980s (Table 3, Fig. 2 ) . Published stomach content analyses and feeding observations also indicate similar diets for coastal bottlenose dolphins of the western North Atlantic from the 1880s to the 1980s. Common gunnard (Prionotus carolinus) was found in the stomachs of two adult female bottlenose dolphins captured in the 1880s (True 1885). Fishermen from the early 20th century reported that bottlenose dolphins ate mainly weakfish (Cynoscion spp.), a sciaenid (Townsend 1914). Mead and Potter (1990, 1995) found the stom- achs of coastal bottlenose dolphins off the eastern United States to contain mainly sciaenid fish (e.g., Cynoscion spp., Leiostornus xantburus, Microjogon un- dulatus, and Bairdiella cbrysura). All of these fish are benthic feeders, consum- ing mainly crustaceans and small fish.
In summary, this research illustrates an application of the isotopic technique for addressing questions about the natural histories of animals. The conclusions drawn from the isotopic data agree with earlier work concerning the feeding preferences of bottlenose dolphins and thus indicate that stable isotopes can be used with existing ecological information to assist in interpreting diets.
ACKNOWLEDGMENTS
We appreciate the Odum Foundation and the Department of Environmental Sciences at the University of Virginia for providing funds for this research. The Smithsonian Institution, under the supervision of James Mead, provided the teeth samples.
LITERATURE CITED
ABEND, A. G., AND T. D. SMITH. 1995. Differences in ratios of stable isotopes of nitrogen in long-finned pilot whales (Globicephala melas) in the western and east- ern North Atlantic. International Council for the Exploration of the Seas Journal of Marine Science 52:837-841.

348 MARINE MAMMAL SCIENCE, VOL. 15, NO. 2, 1999
BARROS, N. B., AND D. K. ODELL. 1990. Food habits of bottlenose dolphins in the southeastern United States. Pages 309-328 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
DENIRO, M. J., AND S. EPSTEIN. 1978. Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimica Acta 42:495-506.
DENIRO, M. J., AND S. EPSTEIN. 1981. Influences of diet on the distribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta 45541-35 1.
DENIRO, M., AND S. WEINER. 1988. Organic matter within crystallin aggregates of hydroxyapatite: A new substrate for stable isotopic and possibly other biogeo- chemical analyses of bone. Geochimica et Cosmochimica Acta 52:2415-2423.
DUFFIELD, D. A., S. H. RIDGWAY AND L. H. CORNELL. 1983. Hematology distinguishes coastal and offshore forms of dolphins (Tu~siops). Canadian Journal of Zoology 61:
FOGEL, M. L., AND L. A. CIFUENTES. 1993. Isotope fractionation during primary pro- duction. Pages 73-98 in M. H. Engel and S. A. Macko, eds. Organic geochemistry principles and applications. Plenum Press, New York, NY.
FRANCE, R. L. 1995. Carbon-13 enrichment in benthic compared to planktonic algae: Foodweb implications. Marine Ecology Progress Series 124307-3 12.
FRY, B. 1988. Food web structure on Georges Bank from stable C , N, and S isotopic compositions. Limnology and Oceanography 33:1182-1190.
FRY, B., AND E. B. SHERR. 1984. 613C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contributions in Marine Science 27: 13-47.
1984. Isotopic variability of organic carbon in a phytoplankton-based, temperate estuary. Geochimica et Cosmochimica Acta 48:1089-1098.
HANSSON, S., J. E. HOBBIE, R. ELMGREN, U. LAMON, B. FRY AND S. JOHANSSON. 1997. The stable nitrogen isotope .ratio as a marker of food-web interactions and fish migration. Ecology 78:2249-22 57.
HARE, P. E., M. L. FOGEL, T. W. STAFFORD JR., A. D. MITCHELL. AND T. C. HOERING. 1991. The isotopic composition of carbon and nitrogen in individual amino acids isolated from modern and fossil proteins. Journal of Archaeological Science 18:
HERSH, S. L., AND D. A. DUFFIELD. 1990. Distinction between Northwest Atlantic offshore and coastal bottlenose dolphins based on hemoglobin profile and mor- phometry. Pages 129-139 in S. Leatherwood and R. R. Reeves, eds. The bottle- nose dolphin. Academic Press, San Diego, CA.
HOBSON, K. A., AND W. A. MONTEVECCHI. 1991. Stable isotopic determinations of trophic relationships of great auks. Oecologia 87:528-53 1.
HOBSON, K. A., D. M. SCHELL, D. RENOUF AND E. NOSEWORTHY. 1996. Stable carbon and nitrogen isotopic fractionacion between diet and tissues of captive seals: Im- plications for dietary reconstructions involving marine mammals. Canadian Jour- nal of Fisheries and Aquatic Sciences 53:528-533.
KOHL, D. H., G. SHEARER AND B. COMMONER. 1971. Fertilizer nitrogen: Contributions to nitrate in surface water in a corn belt watershed. Science 174:1331-1334.
MACKO, S. A,, AND N. E. OSTROM. 1994. Pollution studies using stable isotopes. Pages 45-62 in K. Lajtha and R. H. Michener, eds. Stable isotopes in ecology and environmental science. Blackwell Scientific Publications. London.
MACKO, S. A., W. Y. LEE AND P. L. PARKER. 1982. Nitrogen and carbon isotope frac- tionation by two species of marine amphipods: Laboratory and field studies. Jour- nal of Experimental Marine Biology and Ecology 63:145-149.
MACKO, S. A., L. ENTZEROTH AND P. L. PARKER. 1984. Regional differences in nitrogen and carbon isotopes on the continental shelf of the Gulf of Mexico. Naturwissen- schaften 7 1 :374-375.
MACKO, S. A., M. L. F. ESTEP, P. E. HARE AND T. C. HOERING. 1987. Isotopic frac-
930-93 3.
GEARING, J. N., P. J. GEARING, D. T. RUDNICK, A. G. REQUEJO AND M. J. HUTCHINS.
277-292.

WALKER E T A L . : BOTTLENOSE DOLPHIN DIET 349
tionation of nitrogen and carbon in the synthesis of amino acids by microorgan- isms. Chemical Geology 65:79-92.
MCCONNAUGHEY, T., AND C. P. McRou. 1979. Food-web structure and the fractionation of carbon isotopes in the Bering Sea. Marine Biology 53:247-262.
MEAD, J. G. 1975. Preliminary report on the former net fisheries for Tursiops truncatus in the western North Atlantic. Journal of the Fisheries Research Board of Canada
MEAD, J. G., AND C. W. POTTER. 1990. Natural history of bottlenose dolphins along the central Atlantic coast of the United States. Pages 165-195 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
MEAD, J. G., AND C. W. POTTER. 1995. Recognizing two populations of the bottlenose dolphin (Tursiups truncatus) off the Atlantic coast of North America: Morphologic and ecologic considerations. IBI Reports 5:3 1-44.
MERCER, M. C. 1973. Observations on distribution and intraspecific variation in pig- mentation patterns of odotocete Cetacea in the western North Atlantic. Journal of the Fisheries Research Board of Canada 30: 11 11-1 130.
MINAGAWA, M., AND E. WADA. 1984. Stepwise enrichment of 15N along food chains: Further evidence and the relation between 6I5N and animal age. Geochimica et Cosmochimica Acta 48:1135-1140.
NELSON, B. K., M. J. DENIRO, M. J. SCHOENINGER AND D. J. DE PAOLO. 1986. Effects of diagenesis of strontium, carbon, nitrogen and oxygen concentration and isotopic composition of bone. Geochimica et Cosmochimica Acta 50:1941-1949.
OSTROM, P. H., S. A. MACKO, M. H. ENGEL, J. A. SILPER AND D. RUSSEL. 1989. Geo- chemical characterization of high molecular weight material isolated from Late Cretaceous fossils. Advances in Organic Geochemistry 16:1139-1144.
OSTROM, P. H., S. A. MACKO, M. H. ENGEL AND D. A. RUSSELL. 1 9 9 3 ~ . Assessment of trophic structure of Cretaceous communities based on stable nitrogen isotope analyses. Geology 2 1 :491-494.
OSTROM, P. H., J. LIEN AND S. A. MACKO. 19936. Evaluation of the feeding behavior of the Sowerby's beaked whale, Mesoplodun bidens, based on isotopic comparisons among northwestern Atlantic cetaceans. Canadian Journal of Zoology 7 1:858- 861.
RAU, G. H., T. TAKAHASHI AND D. J. DES MARAIS. 1989. Latitudinal variations in plankton P3C: Implications of CO, and productivity in past oceans. Nature 341: 5 1 6-5 1 8.
SACKETT, W. M., W. R. ECKELMANN, M. L. BENDER AND A. W. H. BE. 1965. Temper- ature dependence of carbon isotope composition in marine plankton and sedi- ments. Science 148:235-237.
SCHELL, D. M., S. M. SAUPE AND N. HAUBENSTOCK. 1989. Bowhead whale (Balaena mysticetus) growth and feeding as estimated by 613C techniques. Marine Biology 103:433-443.
SCHOENINGER, M. J., AND M. J. DENIRO. 1984. Nitrogen and carbon isotopic compo- sition of bone collagen from marine and terrestrial animals. Geochimica et Cos- mochimica Acta 48:625-639.
SMITH, R. J., K. A. HOBSON, H. N. KOOPMAN AND D. M. LAVIGNE. 1996. Distinguish- ing between populations of fresh- and salt-water harbor seals (Phoca vitulina) using stable-isotope ratios and fatty acid profiles. Canadian Journal of Fisheries and Aquatic Sciences 53:272-279.
SWEENEY, R. E., E. K. KAHIL AND I. R. KAPLAN. 1980. Characterization of domestic and industrial sewage in Southern California coastal sediments using nitrogen, carbon, sulfur, and uranium tracers. Marine Environmental Research 3:225-243.
TIESZEN, L. L., T. W. BOUTTON, K. G. TESDAHL AND N. A. SLADE. 1983. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for 613C analysis of diet. Oecologia 57:32-37.
32~1155-1162.
TOWNSEND, C. H. 1914. The porpoise in captivity. Zoologica I 16:289-299.

350 MARINE MAMMAL SCIENCE. VOL. 15. NO. 2. 1999
TRUE, F. W. 1885. The bottle-nose dolphin, Tursiops ttrrsio, as seen at Cape May, New Jersey. Science 116:338-339.
TIJROSS, N., M. L. FOGEL AND P. E. HARE. 1988. Variability in the preservation of the isotopic composition of collagen from fossil bone. Geochimica et Cosmochimica Acta 52:929-935.
WALKER, J. L., AND S. A. MACKO. 1999. Dietary studies of marine mammals using stable carbon and nitrogen isotopic ratios of teeth. Marine Mammal Science 15: 314-334.
WONG, W. W., AND W. M. SACKETT. 1978. Fractionation of stable carbon isotopes by marine phytoplankton. Geochimica et Cosmochimica Acta 42: 1809-1 8 15.
Received: 29 July 1996 Accepted: 12 May 1998




















