
Gibberellin Signaling in the Endodermis Controls Arabidopsis Root Meristem Size
Upload
ist-austriaCategory
view
0download
0
The CRYPTOCHROME1-Dependent Response to ExcessLight Is Mediated through the Transcriptional Activators ZINCFINGER PROTEIN EXPRESSED IN INFLORESCENCEMERISTEM LIKE1 and ZML2 in Arabidopsis C W
Jehad Shaikhali,a Juan de Dios Barajas-Lopéz,a Krisztina Ötvös,a,b Dmitry Kremnev,a Ana Sánchez Garcia,a
Vaibhav Srivastava,c Gunnar Wingsle,c Laszlo Bako,a and Åsa Stranda,1
a Umeå Plant Science Centre, Department of Plant Physiology, Umeå University, S-901 87 Umea, Swedenb Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, H-6726 Szeged, HungarycUmeå Plant Science Centre, Department of Forest Genetics and Plant Physiology, Swedish University of Agricultural Sciences,S-901 87 Umea, Sweden
Exposure of plants to light intensities that exceed the electron utilization capacity of the chloroplast has a dramatic impacton nuclear gene expression. The photoreceptor Cryptochrome 1 (cry1) is essential to the induction of genes encodingphotoprotective components in Arabidopsis thaliana. Bioinformatic analysis of the cry1 regulon revealed the putative cis-element CryR1 (GnTCKAG), and here we demonstrate an interaction between CryR1 and the zinc finger GATA-typetranscription factors ZINC FINGER PROTEIN EXPRESSED IN INFLORESCENCE MERISTEM LIKE1 (ZML1) and ZML2. TheZML proteins specifically bind to the CryR1 cis-element as demonstrated in vitro and in vivo, and TCTAG was shown toconstitute the core sequence required for ZML2 binding. In addition, ZML2 activated transcription of the yellow fluorescentprotein reporter gene driven by the CryR1 cis-element in Arabidopsis leaf protoplasts. T-DNA insertion lines for ZML2 and itshomolog ZML1 demonstrated misregulation of several cry1-dependent genes in response to excess light. Furthermore, thezml1 and zml2 T-DNA insertion lines displayed a high irradiance-sensitive phenotype with significant photoinactivation ofphotosystem II (PSII), indicated by reduced maximum quantum efficiency of PSII, and severe photobleaching. Thus, weidentified the ZML2 and ZML1 GATA transcription factors as two essential components of the cry1-mediated photoprotectiveresponse.
INTRODUCTION
Plants can detect almost all facets of light, including direction,intensity, duration, and wavelength using three major classes ofphotoreceptors: the red/far-red light absorbing phytochromes,the blue/UV-A light absorbing cryptochromes and phototropins,and the UV-B sensing UV-B receptors. These photoreceptorsperceive light signals and initiate intracellular signaling pathwaysand large-scale reorganization of the transcriptional program tomodulate plant growth and development (Chen et al., 2004).Arabidopsis thaliana cryptochromes cry1 and cry2 are wellcharacterized and regulate several aspects of growth and de-velopment (Li and Yang, 2007). The more divergent memberof the family, cry3, is localized to the organelles and was re-ported to be involved in single-stranded DNA repair (Kleine et al.,2003; Pokorny et al., 2008). Cry2 undergoes degradation fol-lowing light activation via ubiquitylation and targeting to the
proteasome (Lin et al., 1998; Yu et al., 2007), whereas cry1 isstable in bright light (Ahmad and Cashmore, 1993). Thus, cry1 isresponsible for seedling deetiolation in high fluence rates of bluelight, and cry2 mediates flowering time and deetiolation in re-sponse to lower fluence rates (Ahmad and Cashmore, 1993; Linet al., 1998). It has also been demonstrated that cry1 playsa critical role in the photoprotective response to excess light(Kleine et al., 2007). Cry1 is present in the nucleus, and althoughhaving considerable effects on transcriptional activity, cry1 isnot known to bind DNA (Cashmore et al., 1999; Lin and Shalitin,2003). Some of the cry1 action on the transcription level hasbeen attributed to its interaction with CONSTITUTIVE PHOTO-MORPHOGENIC1 (COP1). The cry1-COP1 interaction is lightdependent and suppresses the activity of COP/DEETIOLATEDproteins, thereby allowing light-activated transcription (Wanget al., 2001; Yang et al., 2001). Cry1 has also been shown to bepresent in the cytosol (Yang et al., 2000). It has been suggestedthat cry1 may be involved in both nuclear and cytosolic events,and it is possible that there are separate functions for nuclearand cytoplasmic cry1 (Wu and Spalding, 2007).Genetic screens have revealed several regulators in the
cryptochrome-mediated light signaling pathway; a blue light–specific Ser/Thr protein phosphatase (PP7) has been isolated(Møller et al., 2003), and SHORT UNDER BLUE1 is a cytoplas-mic calcium binding protein with a major function in cryptochromesignaling but that also modulates phyA-mediated signaling (Guo
1Address correspondence to [email protected] author responsible for distribution of materials integral to the findingspresented in this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Åsa Strand ([email protected]).C Some figures in this article are displayed in color online but in black andwhite in the print edition.WOnline version contains Web-only data.www.plantcell.org/cgi/doi/10.1105/tpc.112.100099
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been
edited and the authors have corrected proofs, but minor changes could be made before the final version is published. Posting this version online
reduces the time to publication by several weeks.
The Plant Cell Preview, www.aspb.org ã 2012 American Society of Plant Biologists. All rights reserved. 1 of 17

et al., 2001). Cry1 inhibits COP1-mediated degradation of aMYB transcription factor, BIT1, to activate blue light–dependentgene expression (Duek and Fankhauser, 2003; Hong et al.,2008). Furthermore, HFR1, a basic helix-loop-helix proteinidentified as a component in the far-red signaling has beendemonstrated to be crucial for regulating global gene expressionin response to cry1-mediated early blue light response (Duekand Fankhauser, 2003). In addition, HYPERSENSITIVE TO REDAND BLUE1, SHORT HYPOCOTYL UNDER BLUE1, and OBFBINDING PROTEIN3 have been shown to act both in red andblue signaling pathways (Kang et al., 2005; Ward et al., 2005;Kang and Ni, 2006). Photoexcited cry2 was found to interactwith CRYPTOCHROME-INTERACTING BASIC-HELIX-LOOP-HELIX1 to regulate transcription and initiation of flowering (Liuet al., 2008). In addition, cry1 has been shown to interact directlywith ZTL/LKP1/ADO1 in yeast two-hybrid assays and in vitropull-down tests (Kiyosue and Wada, 2000; Somers et al., 2000;Jarillo et al., 2001). ZTL/LKP1/ADO1 is a protein that was orig-inally identified in a study of the circadian clock in Arabidopsis,and ZTL/LKP1/ADO1 plays an important role in regulating thecircadian clock and photoperiodic flowering in Arabidopsis(Somers et al., 2000). Thus, cryptochromes are involved in nu-merous aspects of plant growth and development.
Cry1 has also been assigned a role in plant stress responses,and analysis of the high irradiance response of the photore-ceptor mutants phyA, phyB, cry1, and cry2 revealed a functionof cry1 in mediating plant responses to high irradiances (Kleineet al., 2007). In Arabidopsis, cry1 regulates a large number ofgenes in response to excess light, and as a consequence of themisregulation of these genes the cry1 mutant displayed a highirradiance-sensitive phenotype with significant photoinactivationof photosystem II (PSII) (Kleine et al., 2007). Thus, in addition toa role in photomorphogenesis, cry1 is essential to the inductionof photoprotective mechanisms against high light stress (Kleineet al., 2007; Li et al., 2009). Numerous array experiments haveshown that exposure to excess light results in dramatic changesin gene expression in plants (Kimura et al., 2001; Rossel et al.,2002; Richly et al., 2003; Kleine et al., 2007). A significantnumber of these excess light–regulated genes depend on theaction of cry1 for correct regulation and 26 of those genes werealso LONG HYPOCOTYL5 (HY5) dependent (Kleine et al., 2007).The cry1-dependent genes encode components of the protectivemechanisms against light stress, such as several enzymes in thephenylpropanoid pathway, EARLY LIGHT INDUCED PROTEIN1(ELIP1) and ELIP2, GPX7 that encodes a putative glutathioneperoxidase, the glutathione S-transferase ERD9, and a MYBfamily transcription factor (At5g49330). Bioinformatics studiesperformed on the HY5-independent cry1 regulon revealed thattwo previously undescribed cis-elements, CryR1 (GnTCkAG)and CryR2 (ACATAwCT), were enriched in the list of cry1-dependent genes responding to excess light (Kleine et al.,2007). CryR1 was significantly enriched in the promoters ofgenes induced by excess light in the wild type, suggestinginteraction with an activator of gene expression (Kleine et al.,2007).
Although cry1 plays an essential role in mediating plantresponses to high irradiances, HY5 is the only transcriptionfactor responding to cry1 that has been identified to play a role
under these conditions (Kleine et al., 2007). Thus, to discovernuclear components involved in the cry1-mediated signalingpathway in response to excess light, we used a biochemicalapproach and identified interaction between the GATA-typetranscription factor ZINC FINGER PROTEIN EXPRESSED ININFLORESCENCE MERISTEM LIKE2 (ZML2; also known asTIFY2a) (Vanholme et al., 2007) and the CryR1 cis-element. Weconfirmed binding and specificity of ZML2 and its homologZML1 (TIFY2b) to the CryR1 cis-element in vitro and in vivo. Wealso demonstrate a function of ZML2 as a trans-activation fac-tor. T-DNA insertion lines for ZML2 and ZML1 demonstratedan excess light-sensitive phenotype, increased accumulation ofreactive oxygen species (ROS), and misregulation of severalcry1-dependent genes in response to excess light. Thus, weidentified two key regulators, ZML2 and ZML1, of gene ex-pression in the cry1-dependent response to excess light.
RESULTS
Identification of a Transcription Factor Binding to thePutative CryR1 cis-Element
To test whether a nuclear protein(s) recognizes the putativeCryR1 cis-element, oligonucleotides corresponding to the CryR1element (GAAAAAGTTCTAGAATTTTTT) were used in electro-phoretic mobility shift assays (EMSAs). Nuclear protein extractsfor EMSA were prepared from wild-type plants grown undercontrol conditions. When nuclear protein extracts were in-cubated with biotin-labeled CryR1 oligonucleotides, a DNA-protein complex with slower mobility in EMSA was observed(see Supplemental Figure 1 online).To identify the transcription factor(s) responding to the cry1-
mediated signal and interacting with the putative cis-elementCryR1, we used a biochemical approach where a DNA fragmentcontaining the cis-element of interest was tagged with biotin andlinked to microbeads via interaction with streptavidin (Figure 1)(Gabrielsen et al., 1989; Rey et al., 2003). The 218-bp DNApromoter fragment containing the CryR1 putative cis-elementwas amplified from the R2R3 transcription factor (At5g49330)(Figure 2) representing one of the genes responding to excesslight in a cry1-dependent manner. MatInspector analysis for cis-elements within the 218-bp CryR1-containing promoter frag-ment revealed the presence of DOF, MADS box, GT-box, GAGA,L1-box, W-box, heat shock, homeobox, and AS1/AS2 elements(http://www.genomatix.de).Nuclear proteins were prepared from wild-type plants ex-
posed to control conditions and to excess light. The two dif-ferent sample types of nuclear proteins were incubated togetherwith the DNA fragment containing the CryR1 cis-elements, andthe proteins binding to the DNA fragment were isolated andidentified using mass spectrometry. Using this approach, weidentified an interaction between ZML2 (At1g51600) and theCryR1 element (Table 1). ZML2 was identified in both the controlsample and the sample exposed to excess light (Table 1). TheZML2 protein belongs to the GATA-type zinc finger family, andZIM was originally identified in Arabidopsis due to its pro-nounced expression in flowers and flower buds (Nishii et al.,
2 of 17 The Plant Cell

2000). There are two ZIM homologs, ZML1 and ZML2, and thesethree proteins form a separate group when compared with theother typical GATA-type proteins (Teakle et al., 2002). The zincfinger domains of ZIM, ZML1, and ZML2 have 20 residues be-tween the two Cys pairs (C-X2-C-X20-C-X2-C), while those of theother Arabidopsis GATA factors all have 18 residues (Shikataet al., 2004).
EMSAs Confirmed Binding of ZML2 to the CryR1cis-Element
EMSA was performed to confirm the interaction between theisolated protein and the DNA fragment used as bait in the bio-chemical approach. Full-length cDNA of ZML2 was expressedas a His-tagged protein in Escherichia coli, and the ability of thefusion protein to bind specific DNA fragments was examined(Figure 2A). Binding was not detected in the presence of controllysate from E. coli bacterial cells transformed with an emptyexpression vector, pET100D (Figure 2B). The ZML2 protein wasdemonstrated by EMSA to bind to the 218-bp DNA fragmentcontaining the CryR1 cis-element used in the biochemical assay
(Figure 2C). Furthermore, we demonstrated that ZML2 bindsspecifically to the CryR1 cis-element using a 21-bp DNA frag-ment containing the CryR1 element (Figure 2D). To determinethe core sequence that is essential for ZML2 binding of theCryR1 cis-element, mutations were introduced into this element(Figure 2A) and in vitro binding to ZML2 was tested by EMSA.Changing the sequence of the CryR1 cis-element fromGTTCTAG to TGGTGGT (M1) abolished DNA binding of theZML2 protein to the CryR1 target completely. Similarly to M1,M3 in which TCT of the CryR1 element were mutated to GTGand M4 in which the nucleotides AGA were mutated to GTGresulted in loss of DNA binding (Figure 2E). By contrast, muta-tion of AGT to GTG (M2) did not abolish ZML2 binding (Figure2E). Taken together, these results indicate that the CryR1 cis-element is the target motif for the ZML2 transcription factor andthat the core sequence required for binding is TCTAG. Ourresults also validate the method used to trap DNA bindingproteins. This biochemical approach was previously provensuccessful in bacterial and yeast systems (Gabrielsen et al.,1989; Rey et al., 2003), and our results demonstrate that it ispossible to successfully apply this technique also on the morecomplex plant material.Arabidopsis plants overexpressing ZIM (ZIM-ox) were shown
to upregulate a significant number of genes compared with thewild type when grown under control conditions (Shikata et al.,2004). We analyzed 500 bp from the ATG site of the promotersof these genes and found that the CryR1 cis-element wasoverrepresented in the promoters of the genes upregulated inthe ZIM-ox plants (see Supplemental Table 1 online). In 92%of the promoters, the CryR1 cis-element showed 71 to 100%identity to GTTCTAG, which is the sequence of the CryR1cis-element in the promoter of R2R3, and the CryR1 cis-element occurred at least twice in 62% of the promoters (seeSupplemental Table 1 online). This suggests that ZIM and ZMLsrecognize and bind to the same cis-element.
Subcellular Localization of ZML2
ZML2 was isolated as a nuclear protein binding to the CryR1element, and it contains a basic residue-rich region indicatinga putative NLS. The subcellular localization of ZML2 protein wasinvestigated in transiently transfected protoplasts from Arabi-dopsis using confocal laser scanning microscopy. The ZML2:CFP (for cyan fluorescent protein) fusion protein was transfectedinto the protoplasts either separately or with the nuclear-localized bZIP transcription factor, ABI5 (Lopez-Molina et al.,2003; Shaikhali et al., 2008). Detection of ZML2:CFP and ABI5:YFP (for yellow fluorescent protein) fusion proteins demon-strated that both proteins were distributed exclusively to thenucleus (Figure 3). The fluorescence distribution of ZML2:CFP inthe nucleus was consistent with expected nuclear localization ofthis protein and confirms that ZML2 is localized to the nucleus.
Chromatin Immunoprecipitation Analysis Reveals in VivoBinding of ZML2 to the CryR1 Element
To investigate if ZML2 is also able to bind the CryR1 element invivo, chromatin immunoprecipitation (ChIP) assays were performed
Figure 1. Schematic Outline of the Method for Affinity Trapping of DNABinding Proteins.
PCR amplification of a promoter region of interest where primer1 wasmodified with biotin. The biotinylated PCR product was immobilized onstreptavidin-coated magnetic beads. Nuclear proteins from Arabidopsiswere incubated with the DNA fragment linked to the beads to allow in-teraction between the protein(s) and the DNA. To eliminate bindingof nonspecific proteins, several washing steps were performed. DNAbinding proteins were eluted from the complex using high salt buffer.
ZML1/2, Key Components of the Photoprotective Response 3 of 17

using Arabidopsis protoplasts (Lee et al., 2007). The fusion proteinHA-ZML2 was expressed in Arabidopsis protoplasts under darkand light conditions (Figure 4A). Interestingly, the level of ZML2was much higher in light compared with the dark-grown samples.For ChIP analysis, Arabidopsis protoplasts were transfected withHA-ZML2 fusion protein or a mock control and incubated in lightfor 16 to 24 h. At the end of the incubation time, chromatincomplexes were cross-linked with formaldehyde and the chro-matin was fragmented by sonication and incubated with anti-HA
monoclonal antibody. The immunoprecipitated complexes werecaptured with protein G-coated magnetic beads. PCR analysisperformed on the DNA eluted from the beads revealed that R2R3promoter fragments containing CryR1 element, F2 and F5, wereenriched in the ChIP samples prepared with the anti-HA com-pared with the mock control (Figure 4B). By contrast, no PCRproduct was amplified using primers designed for the lateral root-specific promoter (At2g42430), indicating that these sequences wereabsent from the immunoprecipitated chromatin (Figure 4B). Taken
Figure 2. Characterization of DNA Binding of Recombinant ZML2 Protein.
(A) Nucleotide sequence of the 218-bp fragment containing the CryR1 cis-element from the promoter of R2R3 family transcription factor and the upperstrand sequence of the synthetic double-stranded oligonucleotide containing CryR1 cis-element or its mutagenized variants used as probes.(B) EMSA of lysate from E. coli transformed with pET100D empty expression vector and the 218-bp CryR1 fragment.(C) to (E) EMSA of recombinant ZML2 protein with 218-bp CryR1 fragment (C), CryR1 cis-element (D), and its mutagenized variants (E). Positions offree DNA probes and DNA-protein complexes are indicated by arrows.
Table 1. ZML2 Was Identified to Interact with the CryR1 cis-Element
cis-Element Protein Samples in Which Peptide Was Identified Peptide
CryR1 (GTTCTAG) At1g51600 Control light YTVRKEVALR*ZML2 transcription factor Exposure to excess light
Complete Mascot score result and ion spectra for the ZML2 identification in control sample are available in Supplemental Figure 9 online. The asteriskindicates that the same peptide was also identified in samples exposed to excess light.
4 of 17 The Plant Cell

together, the ChIP assay indicates that ZML2 specifically binds theR2R3 promoter in vivo and most likely to the CryR1 cis-element.
Transactivation Assays Demonstrated That ZML2 ActivatesGene Expression via the CryR1 cis-Element
We demonstrated that ZML2 interacts with the CryR1 element invitro and in vivo by EMSA and ChIP, respectively. CryR1 wassignificantly enriched in the promoters of genes induced byexcess light in the wild type, suggesting interaction with anactivator of gene expression (Kleine et al., 2007). To determinewhether the ZML2 protein could transactivate transcription, weperformed a transactivation assay using protoplasts preparedfrom Arabidopsis leaves. The protoplasts were cotransfectedwith an effector plasmid and a reporter plasmid (Figure 4C). The
effector plasmid carried the ZML2 cDNA driven by cauliflowermosaic virus (CaMV) 35S promoter, and in the reporter plasmid,the YFP gene was linked to the 218-bp fragment of the R2R3promoter containing CryR1 (Figure 4C). An empty effectorplasmid was used as a negative control. A vector containing theCaMV35S:CFP was cotransfected in each experiment as a con-trol for transfection efficiency (Figure 4C). The ZML2 proteintransactivated expression of the YFP reporter gene in Arabidopsisprotoplasts. The YFP expression was eightfold higher comparedwith the empty vector control (Figure 4D). Thus, the putativeCryR1 cis-element identified using bioinformatics (Kleine et al.,2007) was demonstrated to be biologically active. Furthermore,ZML2 functions as a transcriptional activator in the cry1-mediatedresponse to excess light, which is in agreement with the bio-informatic analysis that demonstrated that CryR1 was signifi-cantly enriched in the promoters of genes induced by excesslight in a cry1-dependent manner.
The zml Mutants Showed Impaired Regulation of Genes inthe cry1 Regulon
To demonstrate whether the identified interaction between ZML2and the CryR1 cis-element was biologically significant, we ob-tained T-DNA insertion lines for ZML2; zml2-1 and zml2-2, andZML1; zml1 and ZIM; zim. The presence of the T-DNA insertionswas confirmed by PCR (see Supplemental Figure 2 online). RNAwas isolated from homozygous plants, and quantitative RT-PCRanalysis confirmed the absence of ZML2, ZML1, and ZIM tran-scripts in the T-DNA insertion lines (see Supplemental Figure 2online). Under normal growth conditions, no obvious phenotypecould be observed in the zml2, zml1, and zim mutants. The ex-pression of ZML1 and ZIM in the zml2 mutant and vice versa wasnot altered compared with the wild type or the cry1 mutant (seeSupplemental Figure 3 online). Furthermore, no significant changein expression of the ZML1/2 or ZIM could be detected followingexposure to excess light in the wild type or in the mutant lines (seeSupplemental Figure 3 online).Expression of four marker genes in the cry1-dependent reg-
ulon containing the CryR1 cis-element in their promoters andtwo marker genes in the cry1-independent regulon (Kleine et al.,2007) was investigated in the zml2, zml1, and zim mutants andcompared with the wild type and the cry1 mutant (Figure 5).Under normal growth conditions, the expression of the cry1-dependent genes ELIP2, GPX7, and ERD9 was not significantlyaltered in any of the mutants compared with the wild type (seeSupplemental Figure 4 online). Expression of a gene encodinga MYB family transcription factor, MYB7, was significantly re-pressed in the zim mutant but demonstrated wild-type levelsin the other mutants when grown at control conditions (seeSupplemental Figure 4 online).When exposed to excess white light (1000 µmol m22 s21) for
3 h, the expression of ELIP2 was induced 15-fold compared withcontrol conditions in the wild type and the zim mutant. The cry1,zml2, and zml1 mutants demonstrated significantly impaired in-duction of ELIP2 expression in response to excess light (Figure5A). Expression of GPX7 and ERD9 was induced severalfold inresponse to excess light in the wild type and the zim mutant,whereas the zml2 and zml1 mutant lines demonstrated strongly
Figure 3. Subcellular Localization of ZML2 Fusion Protein in ArabidopsisProtoplasts.
Confocal laser scanning microscope images of ZML2-CFP and ABI5-YFP coexpressed transiently in Arabidopsis protoplasts. Transmittedlight image (A), chlorophyll fluorescence image (B), ZML2-CFP image(C), ABI5-YFP image (D), and merged image of (A) to (D) in (E). CFPsignal is shown in blue, YFP signal is shown in yellow, and the chloro-plast (chlorophyll) autofluorescence is shown in red. Bars = 10 mm.
ZML1/2, Key Components of the Photoprotective Response 5 of 17

attenuated induction of GPX7 and ERD9 similar to the cry1mutant(Figures 5B and 5C). Expression of MYB7 was induced fivefold inthe wild type in response to excess light (Figure 5D), whereas all ofthe mutants demonstrated inability to induce expression of MYB7in response to excess light (Figure 5D). Expression of the cry1-independent genes encoding ribulose bisphosphate carboxylasesmall chain (RBCS) and the light-harvesting protein (LHCB2.4) inresponse to excess light revealed similar expression patterns inthe wild type and the cry1, zml2, zml1, and zimmutants (Figures 5Eand 5F). Under control conditions, the expression of RBCS andLHCB2.4 was not significantly altered in any of the mutants
compared with the wild type (see Supplemental Figure 4 online).The misregulation of cry1-dependent marker genes in both thezml2 and zml1 mutants suggests that the ZML1 and ZML2 tran-scription factors act in concert and are significant components ofthe cry1-mediated response to excess light.
The zml Mutants Displayed Impaired Response toExcess Light
Several ROS, such as hydrogen peroxide (H2O2), singlet oxygen,superoxide, and hydroxyl radicals, are generated in chloroplasts
Figure 4. Binding of ZML2 to CryR1 in Vivo.
(A) Transient expression of HA-ZML2 fusion protein in protoplasts from Arabidopsis cell culture under light (lane 2) and dark (lane 3) conditions. MWT,molecular weight.(B) ChIP assay using Arabidopsis protoplasts expressing HA-ZML2 fusion protein was performed with anti-HA antibody. The positions of the CryR1elements identified in the R2R3 promoter and the fragments analyzed by PCR are represented in the scheme. Genomic DNA obtained from ChIP wasanalyzed by PCR. Immunoprecipitation with IgG antibody was used as a control. Primers for F2 and F5 of the R2R3 promoter and primers for lateralroot-specific promoter (At2g42430) were used for PCR amplification. Input, total input chromatin DNA; HA, DNA precipitated using HA antibody; IgG,DNA precipitated using IgG antibody.(C) Schematic diagram of the effector and reporter constructs used in the transactivation assay. The effector plasmid carried the ZML2 cDNA driven byCaMV 35S promoter. NosT represents the polyadenylation site of nopaline synthetase gene. In the reporter plasmid, the YFP gene was linked to the218-bp CryR1 fragment and a 35S minimal TATA promoter. As a negative control, an empty effector plasmid was used, and as a control for transfectionefficiency, CaMV35S:CFP was cotransfected in each experiment.(D) CryR1 reporter construct was transfected into protoplasts from Arabidopsis leaves with ZML2 effector construct or 35S empty control vector. Tonormalize the transfection efficiency, 35S:CFP:NosT was cotransfected in each experiment. Bars represent means of three independent experiments(6SD) with five protoplasts in each experiment. The asterisk indicates significant difference according to the Student’s t test (P < 0.05).
6 of 17 The Plant Cell

under exposure to excess light (Asada, 2006). Superoxide isoften the first reduced form of oxygen to be generated in planttissues leading to the subsequent formation of H2O2 and hy-droxyl radical. We examined the distribution of superoxide withnitro blue tetrazolium (NBT) staining in 2-week-old Arabidopsisplants of the wild type, cry1, zml2-1, zml2-2, zml1, and zimmutants exposed for 24 h to excess light (1000 µmol m22 s21). Aclear NBT staining was observed following exposure to excesslight, but no difference in superoxide accumulation and NBTstaining could be detected between the wild type and the dif-ferent mutants (see Supplemental Figure 5 online). In addition,we determined H2O2 accumulation in 2-week-old Arabidopsisplants of the wild type, cry1, zml2-1, zml2-2, zml1, and zimmutants following 24 h exposure to excess light (1000 µmol m22
s21). When exposed to excess light, a two- to threefold increasein H2O2 content was observed in the wild type and the zimmutant (Figure 6A). The cry1, zml2, and zml1mutants accumulated
significantly more H2O2 compared with the wild type followingexposure to excess light (Figure 6A).In the wild type, anthocyanin accumulation increased sixfold
after 48 h exposure to 1000 µmol m22 s21 excess light (Figure6B). In the cry1 mutant, the anthocyanin accumulation wasless compared with the wild type (Figure 6B) (Kleine et al.,2007). Reduced anthocyanin levels in cry1-deficient Arabidopsisseedlings have been shown in continuous blue light (Ahmadet al., 1995; Lin et al., 1996) and in continuous white light (Neffand Chory, 1998). In contrast with the cry1 mutant, the zmlmutants accumulated wild-type levels of anthocyanins in re-sponse to excess light (Figure 6B). The anthocyanin accumu-lation pattern in response to excess light in mature plants (seeSupplemental Figure 6 online) was similar to that observed inseedlings (Figure 6B). The cry1-mediated accumulation of an-thocyanin was shown to be mediated via HY5 (Kleine et al.,2007) and is independent of ZML1 and ZML2. The CryR1
Figure 5. Expression of Genes in the cry1 Regulon in the cry1, zml2-1, zml2-2, zml1, and zim Mutant Lines after Exposure to High Light.
Wild-type and the different mutant seedlings were grown for 10 d in continuous white light (100 mmol quanta m22 s21) at 23°C and shifted to excesslight (3 h 1000 mmol quanta m22 s21). Expression of genes from the cry1-dependent regulon ELIP2 (At4g14690) (A), GPX7 (At4g31870) (B), ERD9(At1g10370) (C), and MYB7 (At1g56650) (D) as well as the cry1-independent regulon RBCS (At1g67090) (E) and LHCB2.4 (At3g27690) (F) wereanalyzed using real-time PCR. The expression following exposure to high light was related to the control of each genotype. The results were normalizedto the expression level of At4g36800 encoding a ubiquitin-protein ligase-like protein. The mean (6SD) of at least three biological replicates is shown. WT,the wild type.
ZML1/2, Key Components of the Photoprotective Response 7 of 17

element was identified in the HY5-independent regulon of thecry1-regulated genes.
Exposure to excess light resulted in photoinactivation of PSIIas demonstrated by a drop in maximum quantum efficiency ofPSII (Fv/Fm) from 0.836 to 0.639 after 12 h in the wild type (Table2). The cry1 mutant was almost twice as sensitive to excesslight exposure as the wild type, shown by a drop in Fv/Fm from0.837 to 0.374 (Table 2). The zml2 and zml1 mutant lines also
demonstrated a significant increased sensitivity to excess light,measured as lower Fv/Fm compared with the wild type and thezim mutant (Table 2). Exposure to excess light for 48 h resultedin lower chlorophyll content in wild-type and zim seedlings(Figure 6C). However, the chlorophyll content was significantlylower in cry1, zml1, and zml2 mutants compared with the wildtype (Figure 6C). Furthermore, 10-d-old seedlings of the cry1,zml2, and zml1 mutants demonstrated severe photobleachingfollowing exposure to excess light (Figure 7). A similar effect wasobserved with mature plants (see Supplemental Figure 6 online).Thus, the high light–sensitive phenotype in the zml mutantsdemonstrated by the drop in Fv/Fm, the enhanced photo-bleaching, and the higher accumulation of ROS coincides withthe misregulation of GPX7 and ERD9, encoding componentsinvolved in the photoprotective response (Kleine et al., 2007). Insummary, these results support the conclusion that the ZMLtranscription factors are significant components of the cry1-mediated photoprotective response.
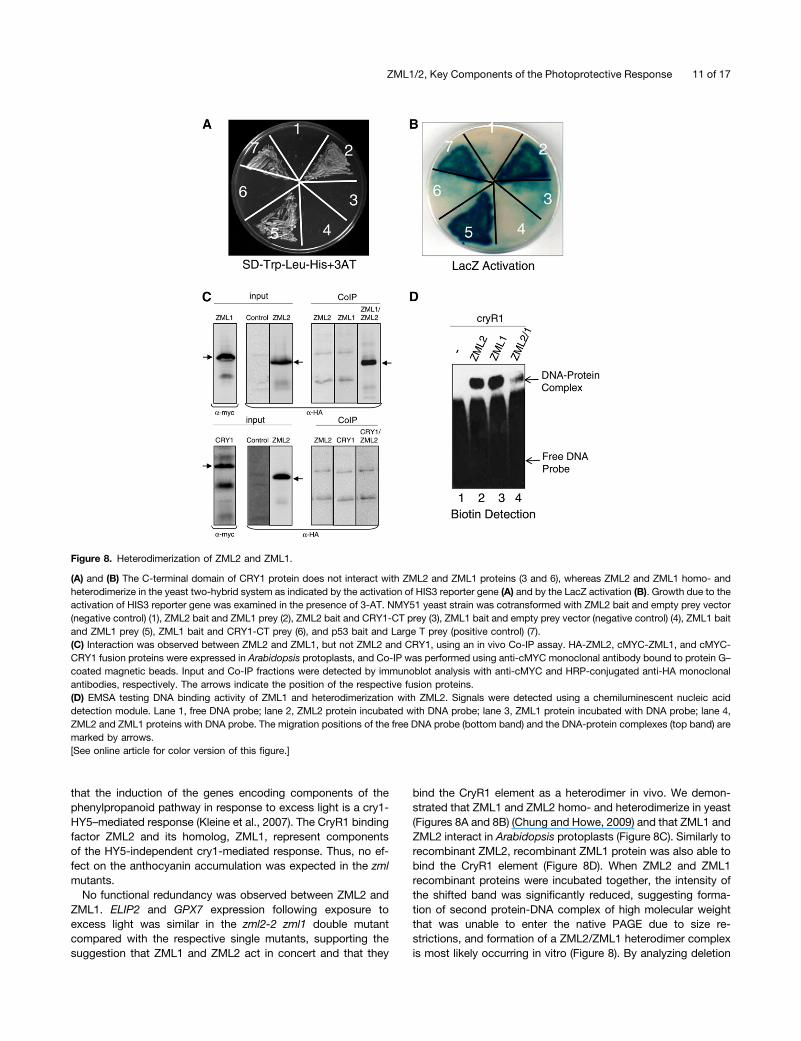
ZML2 and ZML1 Physically Interact in Vivo in the YeastTwo-Hybrid System and in a Coimmunoprecipitation Assay
The misregulation of cry1-dependent genes in both the zml2 andzml1 mutants suggests that there is no redundancy and thatZML1 and ZML2 act in concert in the cry1-mediated response toexcess light. To determine if there is a direct physical interactionbetween ZML2 and ZML1, a yeast two-hybrid assay was per-formed. A fusion protein of the GAL4 DNA binding domain andZML2 (ZML2 bait) activated the transcription of the HIS3 re-porter gene in the presence of GAL4 activation domain andZML1 (ZML1-prey) (Figure 8A) but not with GAL4 activationdomain only (empty prey) (Figure 8A), suggesting a directprotein–protein interaction between ZML2 and ZML1 in yeastcells. ZML1 not only heterodimerizes with ZML2 but also ho-modimerizes with itself when ZML1 bait was cotransformed withZML1 prey (Figure 8A) but not with an empty prey. The HIS3reporter gene activation assay was performed in the presence of7.5 mM 3-aminotriazole (3-AT), a competitive inhibitor of theHIS3 gene. To confirm the observed interactions further, theinteraction was also monitored through LacZ reporter activation,and strong activation was observed for the combinations ZML2bait and ZML1 prey, and ZML1 bait and ZML1 prey (Figure 8B).Whereas it was clear that ZML2 interacted with ZML1, theC-terminal domain of CRY1 (CRY1-CT) was unable to interactwith ZML2 or ZML1 (Figures 8A and 8B). Thus, no directphysical interaction between ZML1/2 and the C terminus ofCRY1 could be detected in yeast. We also confirmed theinteraction between ZML1 and ZML2 using an in vivo coimmu-noprecipitation (Co-IP) assay. HA-ZML2, cMYC-ZML1, andcMYC-CRY1 fusion proteins were expressed in Arabidopsisprotoplasts, and Co-IP was performed using anti-cMYC mono-clonal antibody bound to protein G–coated magnetic beads. Inputand Co-IP fractions were detected by immunoblot analysis withanti-cMYC and horseradish peroxidase (HRP)–conjugated anti-HA monoclonal antibodies, respectively (Figure 8C). From the invivo Co-IP, it is clear that ZML1 and ZML2 can form heterodimersbut that no direct interaction can be detected between ZML2and CRY1 (Figure 8C).
Figure 6. Physiological Characterization of zml2, zml1, and zim T-DNAInsertion Lines in Response to Excess Light.
(A) H2O2 accumulation in control and 24 h high light–exposed 2-week-old seedlings of the wild type, cry1, zml2-1, zml2-2, zml1, and zim mu-tants. The mean (6SD) of three biological replicates is shown. FW, freshweight.(B) Relative anthocyanin content in control and 48 h high light–exposed10-d-old seedlings of the wild type, cry1, zml2-1, zml2-2, zml1, and zimmutants. The mean (6SD) of three biological replicates is shown.(C) Chlorophyll content relative to the control of 48 h high light–exposed10-d-old seedlings of the wild type, cry1, zml2-1, zml2-2, zml1, and zimmutants. The mean (6SD) of three biological replicates is shown. Sig-nificant differences compared with the wild type were assessed ac-cording to Student’s t test: *P < 0.05, **P < 0.01, and ***P < 0.001. Graybars and black bars represent control and high light samples, re-spectively. WT, the wild type.
8 of 17 The Plant Cell

The strong similarity between the DNA binding domains ofZML2 and ZML1 suggests that both proteins could recognizeand interact with the CryR1 binding site. To confirm this hy-pothesis, DNA binding was tested by EMSA (Figure 8D). Simi-larly to recombinant ZML2 (Figure 8D), recombinant ZML1protein was able to bind the CryR1 element (Figure 8D). It is notsurprising that the position of the protein-DNA complexes wasthe same since the size of ZML2 and ZML1 recombinant pro-teins is almost the same (;37 kD); therefore, the size of theprotein-DNA complexes formed should be similar for both pro-teins. To confirm the formation of a heterodimer complex invitro, ZML2 and ZML1 recombinant proteins were incubatedtogether at room temperature prior to the binding reactioncontaining the CryR1 DNA probe. A shifted band was detectedat the same position as that for the DNA-protein complexesformed with the single proteins ZML2 and ZML1, respectively.However, the intensity of the shifted band was significantly re-duced, suggesting formation of second protein-DNA complex ofhigh molecular weight that was unable to enter the native PAGEdue to size restrictions (Figure 8D, lane 4). Thus, formation ofa ZML2/ZML1 heterodimer complex is most likely occurring invitro and the heterodimer also binds to the CryR1 element.
Analysis of Double and Triple Mutants
To test the interaction between cry1 and ZML1/2 genetically,double and triple mutants were generated for the differentgenotypes. Expression of two marker genes, ELIP2 and GPX7,in the cry1-dependent regulon was investigated in the zml2-2zml1, cry1 zml1, and cry1 zml2-2 double mutants and cry1 zml1zml2-2 triple mutant and compared with the wild type and thesingle mutants (Figure 9). The ELIP2 and GPX7 expression fol-lowing exposure to excess light was similar in the zml2-2 zml1double mutant compared with the respective zml2-2 and zml1single mutants (Figure 9). Thus, no enhanced suppression ofexpression could be found in the double mutant compared withthe single mutants, supporting the suggestion that ZML1 andZML2 are genetically linked and act in concert in the cry1-mediated response to excess light. In addition, expression ofELIP2 and GPX7 following exposure to excess light in the cry1zml1 and cry1 zml2-2 double mutants and the cry1 zml1 zml2-2triple mutant was similar to the expression observed in the cry1single mutant (Figure 9). Thus, no significant enhanced sup-pression of expression could be found in the double and triple
mutants compared with the cry1 single mutants, demonstratingthat ZML1/2 and cry1 are components in the same signalingpathway. Furthermore, it is clear that induction of ELIP2 andGPX7 expression in response to excess light is controlled by thecry1-ZML1/2 pathway.
DISCUSSION
The CryR1 cis-element was identified through bioinformaticanalysis and was suggested to be a putative cis-element in-volved in the cry1-dependent response to excess light (Kleineet al., 2007). We identified an interaction between the CryR1 cis-element and the GATA-type zinc finger protein, ZML2 (Table 1),using a biochemical approach previously proven successful inbacterial and yeast systems (Gabrielsen et al., 1989; Rey et al.,2003). ZML2 was demonstrated to be localized exclusively tothe nucleus (Figure 3) and to act as a transcriptional activator ofthe YFP reporter gene driven by the CryR1 cis-element in Arab-idopsis leaf protoplasts (Figure 4). Thus, the putative cis-elementCryR1 was demonstrated to be biologically active, and an in-terplay between cry1, the CryR1 binding transcription factorZML2, and its homolog ZML1 is essential for the induction ofgenes encoding photoprotective components in response to ex-cess light in Arabidopsis.ZIM, ZML2, and ZML1 belong to a group of atypical plant-
specific GATA factors. The plant GATA zinc finger type tran-scription factors have been divided into subfamilies I, II, III, andIV, where group III is also known as the atypical GATA zinc fingertype (Shikata et al., 2004; Manfield et al., 2007). The GATAfactors were first identified as proteins that interacted with GATAmotif (WGATAR) in vertebrates (Martin et al., 1989). In fungi, theGATA (GATAAGG) motif has been identified as the binding sitefor AREA (Kudla et al., 1990), and the sequence motif ATGA-TAAGG was found to be present in the promoter of many LHCand RBCS genes from different plant species (Dean et al., 1985;Grob and Stüber, 1987). In vitro binding studies with GATA1,GATA2, GATA3, and GATA4 from subfamily I, which representsthe majority the plant GATA factors, demonstrated specificity ofthese proteins for the GATA motif (Teakle et al., 2002; Jeong andShih, 2003). However, to the best of our knowledge, no previousreport is available on the binding site of the atypical GATAtranscription factors. Here, we demonstrated that ZML1 andZML2 specifically bind to the CryR1 cis-element in vitro (Figures
Table 2. Maximum Quantum Efficiency of PSII (Fv/Fm)
Genotype Control Fv/Fm 12-h High-Light Fv/Fm
Wild type 0.836 6 0.002 0.693 6 0.083cry1 0.837 6 0.005 0.374 6 0.065*zml2-1 0.835 6 0.003 0.569 6 0.025*zml2-2 0.821 6 0.002 0.556 6 0.033*zml1 0.836 6 0.004 0.397 6 0.109*zim 0.837 6 0.001 0.657 6 0.027
Fv/Fm was estimated before and after exposure to 12 h of white light at 1000 mmol of photons m22 s21 in 5-week-old plants of the wild type and cry1,zml2-1, zml2-2, zml1, and zim mutants. Measurements were made in humidified air. Each point is the mean 6 SD of nine leaves from at least threeindividual plants after 30 min of dark acclimation. The Fv/Fm was significantly different from the wild type in the cry1, zml2, and zml1 mutants followinghigh light exposure (asterisk) according to the Student’s t test (P < 0.05).
ZML1/2, Key Components of the Photoprotective Response 9 of 17

2 and 8). In addition, the bases TCTAG of the CryR1 cis-elementwere demonstrated to be the core sequence required for ZML2binding (Figure 2E). In a ChIP assay, we demonstrated thatZML2 pulled down two fragments from the R2R3 promoter bothcontaining the CryR1 element (Figure 4B). Furthermore, usingtransient expression in Arabidopsis leaf protoplasts, we dem-onstrated that ZML2 transactivated the R2R3 promoterfragment containing CryR1 cis-element (Figure 4D). Thus, inagreement with the indications from the bioinformatic analysiswhere the CryR1 cis-element was enriched in the promoters ofgenes induced by excess light in the wild type, we could dem-onstrate that the ZML2 protein acts as transcriptional activator.An acidic region in the N terminus of the ZIM protein has beendemonstrated to play a role as a transcriptional activation do-main in transactivation experiments using cultured tobacco(Nicotiana tabacum) Bright Yellow-2 cells (Shikata et al., 2003).The entire amino terminal region (76 amino acids) of ZML2shares 44% similarity with ZIM. Although the sequence itself is
not highly conserved, the ZML2 amino terminal region is rich inacidic amino acids, suggesting a similar role for this region. ZIMwas overexpressed using the CaMV 35S promoter in Arabi-dopsis, and the ZIM-ox plants demonstrated elongated hypo-cotyls and petioles and an upward positioning of the leaves(Shikata et al., 2004). The phenotype was observed under allwavelengths of light and in the presence of inhibitors of bras-sinosteroid and gibberellin biosynthesis (Shikata et al., 2004). Asignificant number of genes were shown to be upregulated inthe ZIM-ox plants, and in 92% of the promoters of those genes,one or two CryR1 cis-elements were found either in exact copiesas it occurs in the promoter of R2R3 or in slight variants (seeSupplemental Table 1 online). This suggests that ZIM, ZML2,and ZML1 recognize and bind to the same cis-element. Inaddition to the GATA domain, the ZIM and ZML proteins arecharacterized by a 36–amino acid domain containing the con-served ZIM/TIFY motif (TIFF/YXG) (Vanholme et al., 2007). TheZIM/TIFY domain has been found in other plant proteins, and thecorresponding genes have been grouped into the plant-specificTIFY family with 18 members in Arabidopsis. According to a sug-gested nomenclature, the ZIM, ZML2, and ZML1 were namedTIFY1, TIFY2a, and TIFY2b, respectively (Vanholme et al., 2007).The TIFY family is very diverse, and ZIM/TIFY1, ZML2/TIFY2a,and ZML1/TIFY2b are the only proteins with a GATA zinc finger(Vanholme et al., 2007). The TIFY domain has been demon-strated to be involved in protein–protein interactions (Chung andHowe, 2009).ZML1 and ZML2 are involved in the cry1-dependent induction
of ELIP2, GPX7, ERD9, and MYB7 expression in response toexcess light, and induction of these genes was significantlyimpaired in the zml2 and zml1 single and zml2-2 zml1 doublemutants (Figures 5 and 9). Analysis of the cry1 zml1 and cry1zml2-2 double mutants and the cry1 zml1 zml2-2 triple mutantconfirmed that cry1 and ZML1/2 are components in the samesignaling pathway since no enhanced suppression of ELIP2 andGPX7 expression could be found in the double and triple mu-tants compared with the cry1 single mutant (Figure 9). Similarlyto the cry1 mutant, the zml mutants displayed a clear excesslight–sensitive phenotype as demonstrated by the drop in Fv/Fm,the enhanced photobleaching, reduced chlorophyll content, andhigher accumulation of ROS compared with the wild type (Table2, Figures 6 and 7; see Supplemental Figure 6 online). Theexcess light–sensitive phenotype of the cry1, zml2, and zml1mutants could possibly be explained by the misregulation ofthe stress-related genes encoding the chloroplastic glutathioneperoxidase7 (GPX7) and the glutathione S-transferases (ERD9)in the mutants. Glutathione peroxidases have been shown tofunction as both ROS transducers and scavengers, and gluta-thione S-transferases have been shown to respond to variousstresses such as high light, cold, and drought (Wagner et al.,2002; Seki et al., 2003; Goulas et al., 2006; Miao et al., 2006).A gpx7 T-DNA insertion mutant demonstrated similar phenotypeto the cry1 and zml mutants with compromised photooxidativestress tolerance and higher H2O2 levels in the leaves followingexposure to excess light, demonstrating an important role forGPX7 during photoprotection (Chang et al., 2009). In contrastwith cry1, the zml2 and zml1 lines accumulated anthocyaninfollowing exposure to excess light (Figure 6). It was demonstrated
Figure 7. The zml2, zml1, and zim T-DNA Insertion Lines Exposed toExcess Light.
Ten-day-old seedlings of the wild type, cry1 mutant, and the zml2-1,zml2-2, zml1, and zim T-DNA insertion lines following 24 h (A) and 48 h(B) exposure to high light (HL; 1000 mmol quanta m22 s21). The severephotobleached phenotype observed in the zml2-1, zml2-2, and zml1 wasreproduced in a second independent experiment. WT, the wild type.
10 of 17 The Plant Cell
that the induction of the genes encoding components of thephenylpropanoid pathway in response to excess light is a cry1-HY5–mediated response (Kleine et al., 2007). The CryR1 bindingfactor ZML2 and its homolog, ZML1, represent componentsof the HY5-independent cry1-mediated response. Thus, no ef-fect on the anthocyanin accumulation was expected in the zmlmutants.
No functional redundancy was observed between ZML2 andZML1. ELIP2 and GPX7 expression following exposure toexcess light was similar in the zml2-2 zml1 double mutantcompared with the respective single mutants, supporting thesuggestion that ZML1 and ZML2 act in concert and that they
bind the CryR1 element as a heterodimer in vivo. We demon-strated that ZML1 and ZML2 homo- and heterodimerize in yeast(Figures 8A and 8B) (Chung and Howe, 2009) and that ZML1 andZML2 interact in Arabidopsis protoplasts (Figure 8C). Similarly torecombinant ZML2, recombinant ZML1 protein was also able tobind the CryR1 element (Figure 8D). When ZML2 and ZML1recombinant proteins were incubated together, the intensity ofthe shifted band was significantly reduced, suggesting forma-tion of second protein-DNA complex of high molecular weightthat was unable to enter the native PAGE due to size re-strictions, and formation of a ZML2/ZML1 heterodimer complexis most likely occurring in vitro (Figure 8). By analyzing deletion
Figure 8. Heterodimerization of ZML2 and ZML1.
(A) and (B) The C-terminal domain of CRY1 protein does not interact with ZML2 and ZML1 proteins (3 and 6), whereas ZML2 and ZML1 homo- andheterodimerize in the yeast two-hybrid system as indicated by the activation of HIS3 reporter gene (A) and by the LacZ activation (B). Growth due to theactivation of HIS3 reporter gene was examined in the presence of 3-AT. NMY51 yeast strain was cotransformed with ZML2 bait and empty prey vector(negative control) (1), ZML2 bait and ZML1 prey (2), ZML2 bait and CRY1-CT prey (3), ZML1 bait and empty prey vector (negative control) (4), ZML1 baitand ZML1 prey (5), ZML1 bait and CRY1-CT prey (6), and p53 bait and Large T prey (positive control) (7).(C) Interaction was observed between ZML2 and ZML1, but not ZML2 and CRY1, using an in vivo Co-IP assay. HA-ZML2, cMYC-ZML1, and cMYC-CRY1 fusion proteins were expressed in Arabidopsis protoplasts, and Co-IP was performed using anti-cMYC monoclonal antibody bound to protein G–coated magnetic beads. Input and Co-IP fractions were detected by immunoblot analysis with anti-cMYC and HRP-conjugated anti-HA monoclonalantibodies, respectively. The arrows indicate the position of the respective fusion proteins.(D) EMSA testing DNA binding activity of ZML1 and heterodimerization with ZML2. Signals were detected using a chemiluminescent nucleic aciddetection module. Lane 1, free DNA probe; lane 2, ZML2 protein incubated with DNA probe; lane 3, ZML1 protein incubated with DNA probe; lane 4,ZML2 and ZML1 proteins with DNA probe. The migration positions of the free DNA probe (bottom band) and the DNA-protein complexes (top band) aremarked by arrows.[See online article for color version of this figure.]
ZML1/2, Key Components of the Photoprotective Response 11 of 17

constructs of the JASMONATE ZIM-domain (JAZ) proteins in theTIFY family, it was demonstrated that the conserved TIFY do-main mediates homo- and heteromeric interactions betweenseveral JAZ proteins in Arabidopsis, and it was suggested thatcombinatorial interactions between various JAZ proteins playa role in generating diverse jasmonate signal outputs (Chungand Howe, 2009). Thus, it is possible that the ZML1 and ZML2interact via the TIFY domain. However, unlike the other GATAproteins or proteins in the TIFY family in Arabidopsis, ZML2 andZML1 also contain a CCT domain (Shikata et al., 2004). The CCTdomain is present in the flowering time protein CONSTANS (CO;Robson et al., 2001; Suárez-López et al., 2001), CO-like protein(Griffiths et al., 2003), and in TIMING OF CAB EXPRESSION1(Strayer et al., 2000). CCT was first identified in CO, and itwas suggested to mediate protein–protein interactions (Robsonet al., 2001). This was supported by the observation that theCCT domain of CO interacted with the Arabidopsis transcription
factor ABI3 in yeast cells (Kurup et al., 2000). Thus, it is alsopossible that ZML2 interacts with ZML1 through the CCTdomain.No direct interaction between the C terminus of CRY1 and
ZML1 or ZML2 could be detected in yeast (Figures 8A and 8B) orbetween the full-length CRY1 and ZML2 in Arabidopsis proto-plasts (Figure 8C). Our results suggest that cry1 and ZML1/2 aregenetically linked and that cry1 action, via some unknowncomponent, stimulates the formation and/or activation of theZML heterodimer required for full expression of genes encodingphotoprotective components (Figure 10). It is also possible thatcry1 action modifies the ZML1/2 protein levels or phosphory-lation status. Expression of the genes containing the CryR1 cis-element in their promoters was not impaired in zml mutant linesor in cry1 under control conditions (see Supplemental Figure 4online), suggesting that cry1 activates the ZML1/2 complex onlyin response to excess light to stimulate induction of genes en-coding photoprotective components (Figure 10). Furthermore,the hypocotyl growth response to blue light was not affected inseedlings of the zml2 and zml1 mutants (see SupplementalFigure 7 online), suggesting that normal blue light response isnot mediated via the ZMLs. The ZIM, ZML1, and ZML2 proteinshomo- and heterodimerize in all possible combinations (Chungand Howe, 2009), and it is possible that different combinationsof the GATA transcription factors are triggered by different cuesand regulate specific gene sets. It is also possible that hetero-dimers could be formed with other members of the TIFY family,such as the PEAPOD1 (PPD1) and PPD2. Similarly to ZIM, PPD1and PPD2 have been shown to regulate leaf size and shape inArabidopsis (Shikata et al., 2004; White, 2006). Further experi-ments will determine the exact interplay between the TIFY pro-teins and their role in gene regulation. However, it is clear thatthe TIFY protein ZML2 and its homolog, ZML1, are essentialcomponents of the cry1-mediated response to excess light.
Figure 9. Double and Triple Mutant Analysis in Response to ExcessLight.
Wild-type (WT) and the different mutant seedlings were grown for 10 d incontinuous white light (100 mmol quanta m22 s21) at 23°C and shifted toexcess light (3 h 1000 mmol quanta m22 s21). Expression of genes fromthe cry1-dependent regulon GPX7 (A) and ELIP2 (B) was analyzed usingreal-time PCR. The expression following exposure to high light was re-lated to the control of each genotype. The results were normalized to theexpression level of At4g36800 encoding a ubiquitin-protein ligase-likeprotein. The mean (6SD) of nine biological replicates is shown.
Figure 10. Working Model for the cry1-Mediated HL Response.
In response to excess light, cry1 activates the ZML complex via someunknown component, possibly by facilitating heterodimerization betweenZML1 and ZML2. The ZML complex binds to the CryR1 cis-element,and expression of genes encoding photoprotective components isinduced.
12 of 17 The Plant Cell

METHODS
Plant Material and Growth Conditions
Seeds of Arabidopsis thaliana Columbia-0 wild type, cry1-304 (Ahmadand Cashmore, 1993), zml2-1 (GK448C11), zml2-2 (GK833C07), zml1(SALK_069271), and zim (SALK_144513) were used. The T-DNA insertionlines obtained from the European Arabidopsis Stock Center were grownon soil at 23°C (16 h light 100 mmol quanta m22 s21) and 18°C (8 h dark) at60% relative humidity. For aseptic growth, seeds were sterilized for15 min with 75% (v/v) ethanol, 0.01% Triton X-100, washed three timeswith 95% ethanol, and plated on 0.27% phytoagar plates containing 13Murashige and Skoog salt mixture including vitamins (Duchefa) and 2%Suc. The plates were maintained in darkness at 4°C for 2 d for stratifi-cation and then placed for 10 d in continuous light (100 mmol quanta m22
s21). For high light treatment, either 10-d-old seedlings or 4-week-oldplants were subjected to 3 h 1000 mmol quanta m22 s21 (metal halideHQI-T 400-W day light bulbs; Orsam).
Nuclear Protein Extraction
Control and 3 h high light–treated leaves of 4-week-old Arabidopsisplants were used for nuclear protein extractions using the CelLytic PN-Plant nuclei isolation/extraction kit (Sigma-Aldrich) according to themanufacturer’s instructions. The plant tissue was ground in liquid ni-trogen, homogenized in nuclei extraction buffer, and filtered throughnylon net, and the pellets were collected by centrifugation. For cellmembrane lysis, Triton X-100 was added to 0.3% final concentration.Nuclear proteins were extracted from crude nuclei in protein extractionbuffer.
DNA-Affinity Trapping of DNA Binding Proteins
DNA promoter fragments of the R2R3 transcription factor (At5g49330)containing CryR1 was used to isolate proteins binding to the cis-elementsin these promoter fragments. The promoter region was 21060 to 21278of R2R3. Biotinylated DNA promoter fragments were generated by PCRusing the following primers: CryR1-F, biotin-59-CTTCTTTAACTCGT-TAAATC-39; CryR1-R, 59-TTTATGGTCCAGAGACCAGT-39. BiotinylatedDNA promoter fragment was immobilized on Dynabeads M-280 strep-tavidin (Invitrogen) according to the manufacturer’s instructions in which2 mg beads were immobilized using 23 binding buffer (10 mM Tris HCl,pH 7.5, 1 mM EDTA, and 2 M NaCl). Protein binding to the DNA wasperformed as described by Gabrielsen et al. (1989) with some mod-ifications. Incubation (15min) at 25°C was performed after the beads wereresuspended in protein binding buffer (20 mM Tris HCl, pH 8.0, 1 mMEDTA, 10% glycerol, 100 mM NaCl, 0.05% Triton X-100, and 1 mM DTT)and mixed with Arabidopsis nuclear protein extracts. The Dynabeadswere washed three times with protein binding buffer before proteins wereeluted in elution buffer (20 mM Tris HCl, pH 8.0, 1 mM EDTA, 10%glycerol, 1 M NaCl, 0.05% Triton X-100, and 1 mM DTT).
Protein Digestion, Mass Spectrometry, and Data Analysis
Total proteins in solution were incubated at 95°C for 15 min in presence of0.1 MNH4HCO3 and 10mMDTT, cooled to room temperature, mixed with8 M urea, and incubated for 1 h. Subsequently, alkylation reaction wasdone at 37°C for 30 min in dark after adding 55 mM iodoacetamide. Ureaconcentration was reduced to 0.8 M by diluting with 0.1 M NH4HCO3.Trypsin was added at 1:50 enzyme-to-substrate ratio, and digestion wasperformed overnight at 37°C. The resulting peptides were lyophilized,resuspended in 1% trifluoroacetic acid, and desalted using a Poros 50reverse-phase R2 microcolumn (PerSeptive Biosystems) as described byYanamandra et al. (2009).
The desalted tryptic peptides were separated by reversed-phaseultraperformance liquid chromatography using a nanoACQUITY UPLCsystem (Waters) prior to mass spectrometry analysis. Each sample(peptides) was concentrated on a C18 trap column (Symmetry 180 µm 3
20 mm 5 µm; Waters) and washed with 5% acetonitrile and 0.1% formicacid at 15 µL/min for 1 min. The samples were eluted from the trap columnand separated on a C18 analytical column (75 µm 3 100 mm 1.7 µm;Waters) at 350 nL/min using 0.1% formic acid as solvent A and 0.1%formic acid in acetonitrile as solvent B, in a gradient. The followinggradients were used: linear from 0 to 40% B in 25 min, linear from 40 to80% B in 1 min, isocratic at 80% B in 1 min, linear from 80 to 5% B in1 min, and isocratic at 5% B for 7 min. The eluting peptides were sprayedinto the mass spectrometer (Q-Tof Ultima; Waters) with the capillaryvoltage set to 2.6 kV and cone voltage to 40 V. The instrument wasoperated in data-dependent mode as described (Srivastava et al., 2009)without any further changes.
ProteinLynx Global Server software (V2.2.5) was used to convert rawdata to peak lists for database searching. Proteins were identified bya local version of Mascot search program (V2.1.04; Matrix ScienceLimited) using Arabidopsis protein database from The Arabidopsis In-formation Resource (version 9.0; July 19, 2009; 33,410 sequences). Thefollowing settings were used for the database search: trypsin-specificdigestion with two missed cleavage allowed, carbamidomethylated Cysset as fixedmodification, oxidizedMet in variablemode, peptide toleranceof 80 ppm, and fragment tolerance of 0.1 D. Peptides with Mascot ionscores exceeding the threshold for statistical significance of P < 0.05 wereselected and processed manually to validate their significance (seeSupplemental Figure 9 online).
Expression and Purification of Recombinant Proteins
The full-length open reading frame of ZML2 and ZML1 were PCRamplified using the primers ZML2-F1 (59-CACCATGGATGACCTACAT-GGAA-39), ZML2-R (59-TCACTGTGAGTTGCTTATGTCATT-39), ZML1-F(59-CACCATGGATGATCTTCATGG-39), and ZML1-R (59-TCACTGTG-TGTTGCTAA-39) (see Supplemental Table 2 online). The ZML2 and ZML1PCR products were cloned into the pET100D TOPO vector according tothe manufacturer’s instructions (Invitrogen). After 5 h induction with 1 mMisopropylthio-b-galactoside, Escherichia coli (BL21)–expressed proteinswere affinity purified on Ni2+-NTA agarose resin (Qiagen).
EMSAs
To generate a 218-bp CryR1-containing fragment, the promoter region21060 to 21278 of R2R3 factor gene family was used for PCR ampli-fication using the primers (cryR1-F: 59-CTTCTTTAACTCGTTAAATC-39;cryR1-R: 59-TTTATGGTCCAGAGACCAGT-39). To generate G-box cis-el-ement, CryR1 cis-element, and its mutant variants, forward primers cryR1cis-F (59-TCAACTGACACGTGGCATAAC-39), cryR1cis-F (59-GAAAAAAGTT-CTAGAATTTTTT-39), cryR1-M1-F (59-GAAAAGTGGTGGTAATTTTTT-39),cryR1-M2-F (59-GAAAAGTGTCTAGAATTTTTT-39), cryR1-M3-F (59-GAAA-AAGTGTGAGAATTTTTT-39), and cryR1-M4-F (59-GAAAAAGTTCTGTGA-TTTTTT-39) were annealed with their complementary oligonucleotidesat room temperature after they were incubated at 70°C for 5 min.
DNAprobeswere labeled using theAlkPhos direct labeling anddetectionsystem (Amersham) according to the supplier’s instructions with somemodifications. Reaction buffer (10 mL) was added to 10 mL DNA (ng/mLdiluted DNA) after the DNA was cooled on ice for 5 min, 2 mL labelingreagent was added to the reaction followed by the addition of 10 mL ofcross-linker, and the reactions were incubated for 30 min at 37°C. DNA–protein interactions were performed in 25-mL reactions that contained2.5 mL 103 binding buffer (100 mM Tris HCl, 250 mM KCl, and 10 mMDTT), 1mg poly dI.dC (Sigma-Aldrich), 2.5% glycerol, 0.05% Triton X-100,5 mM MgCl2, 10 mM EDTA, 20 ng DNA, and 2.5 mg protein and were
ZML1/2, Key Components of the Photoprotective Response 13 of 17

then incubated at room temperature for 30 min. The reactions were run on6%native PAGE in 0.53 Tris-borate-EDTA at 100 V. Gels were transferredto a positively charged nylon membrane (Amersham) using a wet transfercell (Bio-Rad), and the DNAwas cross-linked to the membrane using a UVlinker (Spectroline). Membranes were blocked for 1 h at room temperaturein blocking buffer containing 2%blocking reagent (AlkPhos direct labelingand detection system; Amersham) and 1% milk powder in 13 TBS.Membranes were washed four times for 7 min before the DNA was de-tected with CDPstar (AlkPhos direct labeling and detection system).
Biotin Labeling and Detection
PCR products and double-stranded oligonucleotides were labeled at their39 end with biotin-14-dCTP using biotin-labeling kit (Pierce) accordingto the supplier’s instructions. Biotin detection was performed using theChemiluminescent Nucleic Acid Detection Module (Pierce) according tothe supplier’s instructions.
Subcellular Localization and Transactivation Assay
To study the subcellular localization of ZML2 protein, full-length codingsequence, lacking the stop codon, of ZML2 was fused upstream of CFP inthe 35S:CFP vector to produce the fluorescence fusion ZML2:CFP.ZML2:CFP and ABI5:YFP (Shaikhali et al., 2008) were transfected andcoexpressed transiently in protoplasts isolated from Arabidopsis leaves(see Supplemental Table 2 online). After 16 to 24 h incubation of thetransfected cells, fluorescence was visualized using a SP2 confocal laserscanning system equipped with an inverted microscope (Leica) and 363water immersion objective (numerical aperture of 0.75). Representativeimages were taken at 433 and 514 nm for the specific emission of CFP andYFP, respectively. In transactivation assays, effector plasmids used wereconstructed using full-length cDNAs of ZML2 that were cloned into NcoIand EcoRI sites of 35S:CFP-NosT (Seidel et al., 2005), replacing the CFPgene, and the construct was designated 35S:ZML2 (see SupplementalTable 2 online). The empty vector control was constructed after 35S:CFP-NosT vector was digested with NcoI-EcoRI and religated. To constructreporter plasmids, the 35S promoter of 35S:YFP-NosT (Seidel et al., 2005)was replaced with 35S minimal TATA promoter in BamHI-NcoI sites, andthen the 218-bp cryR1-containing fragments were cloned into the HindIIIsite upstream of the 35Sminimal TATA promoter. Isolation and transfectionof Arabidopsis mesophyll protoplasts was performed as described (Seidelet al., 2004). CFP andYFP fluorescencewas quantified using ImageJ (http://rsbweb.nih.gov/ij/). Ten different regions displaying fluorescence signalwere selected for quantification, andYFPbrightnesswas normalized toCFPsignal. 35S-CFP-NosT was transfected in all experiments to serve as aninternal control to normalize the transfection efficiency.
ChIP Assays
For ChIP assays, protoplasts were prepared as described above, andChIP assays were performed as described by Lee et al. (2007). Proto-plasts were transfected with 5 mg of ZML2 cDNA fused to HA tag (HA-ZML2) construct or with water and incubated for 16 to 24 h at roomtemperature. The expression of HA-ZML2 fusion protein was assessed byimmunoblots using protoplast protein extracts. Protoplasts were fixedwith formaldehyde, and chromatin was isolated and sheared by soni-cation to obtain fragments of sizes between 100 and 1500 bp. Anti-HAmonoclonal antibody bound to protein G–coated magnetic beads (Dy-nabeads Protein G Immunoprecipitation kit; Invitrogen) were used toimmunoprecipitate the genomic DNA fragments. Semiquantitative PCRwas performed with immunoprecipitated genomic DNA using primer pairscorresponding to fragments of R2R3 promoter (F2 and F5). Primersspecific for lateral root promoter (At2g42430) were used to amplifya negative control.
Genotyping of T-DNA Insertion Lines
Two mutant alleles of zml2 (GabiKat line 448C11 and GabiKat line833C07), one mutant allele of zml1 (Salk_069271), and one mutant alleleof zim (Salk_144513) were obtained from the European Arabidopsis StockCenter. Individual plants were screened for T-DNA insertion by PCR usinggene-specific primers and primers anchored in the T-DNA borders.Homozygous plants were identified by the absence of the PCR productsobtained from gene-specific primers, which compose the T-DNA insertionsite (see Supplemental Figure 8 online). Gene-specific primers usedfor genotyping were GK448C11-F (59-GTGGTAGTGAACAACAAGGA-GATC-39) and GK448C11-R (59-AGTATGTCAGGAACTCGCAGT-39) forzml2-1 (448C11), GK833C07-F (59-ACATGAGCATGGAACCTATACG-39)and GK833C07-R (59-CTACAGAACCTGAGGCGATTCA-39) for zml2-2(833C07), salk069271-F (59-CCTCGTATCATGTGAAGAATGG-39) andsalk069271-R (59-CATGTTCACAACCATTTGACG-39) for zml1 (069271),and salk144513-F (59-CAGGCTCTTTTTGTGTTCCTG-39) and salk144513-R (59-CCGATGGCTCGAATTACTTC-39) for zim (144513). Primers for theT-DNA left borders were o8409 (59-ATATTGACCATCATACTCATTGC-39)for the Gabi T-DNA and LBa1 (59-TGGTTCACGTAGTGGGCCATCG-39) forthe Salk T-DNA.
RNA Isolation, cDNA Synthesis, and Real-Time PCR
A plant RNA mini kit (EZNA) was used for total RNA isolation according tothemanufacturer’s instructions. Onemicrogram of total RNAwas used forcDNA synthesis using the iScript cDNA synthesis kit (Bio-Rad) accordingto themanufacturer’s instructions. The primers (see Supplemental Table 2online) used for real-time PCR analysis were designed to flank intron sitesto detect amplification of genomic DNA. Real-time PCR was performed in20-mL reactions containing 2 mL cDNA (1:10 dilution) using iQSYBERGreen Supermix (Bio-Rad). Two-step thermal cycling protocol consistedof an initial step at 95°C for 3min followed by 40 cycles of 10 s at 95°C and30 s at 60°C before performing a melt curve 60°C to 95°C was performedusing the CFX96 Real-Time system (C1000 Thermal Cycler; Bio-Rad). Allreactions were performed in triplicate, and the relative transcript abun-dance of each tested gene was normalized to the expression level ofubiquitin (At4g36800). The data were analyzed using LinRegPCR software(Pfaffl, 2001; Kindgren et al., 2012).
Maximal PSII Quantum Yield
In vivo chlorophyll a fluorescence of single leaves wasmeasured using theDual-PAM-100 measuring system (Walz) in ambient air at room tem-perature. Plants were dark adapted for 30min, andminimum fluorescence(F0) excited by weak measuring light at open PSII centers was measured(settings: measuring light, 10). Then, saturating pulses (0.6 s) of white light(3000 mmol photons m22 s21) were used to determine the maximumfluorescence (Fm) at closed PSII centers, and the ratio maximum quantumyield of PSII (Fv/Fm = (Fm 2 F0)/Fm) was calculated.
Chlorophyll and Anthocyanin Measurements
The chlorophyll and anthocyanin measurements were performed asdescribed (Porra et al., 1989; Neff and Chory, 1998). Ten-day-oldseedlings or 5-week-old plants were harvested, weighed, and ground inliquid nitrogen to fine powder. For the chlorophyll measurements, totalchlorophyll was extracted in 80% acetone and calculated according toPorra et al. (1989). For the anthocyanin measurement, the pigments wereextracted in 1% HCl in methanol. Water was added and the chlorophyllwas extracted with an equal volume of chloroform. The anthocyaninquantity was determined by spectrophotometric measurement of theaqueous phase at A530 to A657 and normalized to the total fresh weightused in each sample.
14 of 17 The Plant Cell

H2O2 Measurement
Two-week-old plants were used to measure H2O2 in control and high lighttreatment. Leaf powder (50 to 200 mg) was homogenized in 1 mL of 0.2 MHClO4, incubated on ice for 5 min, and then centrifuged at 10,000g for10min at 4°C. The acidic supernatant was neutralized to pH 7.0 to 8.0 with0.2 M NH4OH, pH 9.5, and briefly centrifuged at 3000g for 2 min tosediment the insoluble material. Quantification of H2O2 in the extracts wasperformed using the Amplex Red Hydrogen Peroxide-Peroxidase Assay kit(Molecular Probes) according to the manufacturer’s instructions. Fluores-cence was measured with a plate reader (Spectra Max Gemini; MolecularDevices) using excitation at 530 nm and fluorescence detection at 590 nm.The concentration of H2O2 was calculated using a standard curve.
ROS (Superoxide) Staining
Two-week-old plants were exposed to high light for 12 h. For superoxidestaining, the plants were incubated for 1.5 h in 0.1% NBT at roomtemperature before they were destained in ethanol.
Yeast Two-Hybrid System
The yeast two-hybrid was performed with the DUALhybrid system(Biotech) according to the manufacturer’s instructions. Full-length codingsequence of ZML2 and ZML1 genes was cloned into pLexA-N vectorto generate fusion proteins with LexA DNA binding domain (bait) (seeSupplemental Table 2 online). ZML1 full-length coding sequence andCRY1 C-terminal domain coding sequence (CRY1-CT) were cloned intopGAD-HA vector to generate fusions of the prey protein with the GAL4activation domain. NMY51 yeast strain was cotransformed with bait andprey vectors and the transformants were selected on selective media SD-Trp-Leu-HIS and containing 7.5 mM 3-AT. b-Galactosidase overlay assayto detect LacZ activation was performed with an overlay buffer containing0.5 M potassium phosphate, pH 7.0, 6% dimethylformamide, 0.1% SDS,50 mL/100 mL b-mercaptoethanol, 5 mg/mL low melting agarose, and0.05% X-Gal (Fermentas).
Co-IP
To generate expression constructs for Co-IP assays, full-length codingsequence of ZML2 was cloned into BamHI-EcoRI sites in pRT104_3HAvector, and the construct was designated HA-ZML2 (see SupplementalTable 2 online). To generate MYC-ZML1 and MYC-CRY1 constructs, full-length coding sequences of ZML1 andCRY1were cloned into EcoRI-KpnIand SmaI-EcoRI sites in pRT104_3MYC vector, respectively. Proteinsfrom protoplasts transformed with expression constructs were extractedin 300 mL immunoprecipitation buffer (25 mM Tris- HCl, pH 7.8, 75 mMNaCl, 10 mM MgCl2, 2 mM DTT, 5 mM EGTA, 0.2% Triton X-100, 10%glycerol, and 0.2 mM PMSF) for 40 min at 4°C. Protein extracts wereincubated with 5 µg of anti-cMYC monoclonal antibody (Bio-Site) boundto protein G–coated magnetic beads (Dynabeads Protein G Immuno-precipitation kit; Invitrogen) for 1 h at 4°C. Subsequent washing stepswere performed according to kit recommendations (Invitrogen), and targetantigen was eluted with SDS-PAGE sample loading buffer. To detect thefusion proteins tagged with cMYC, immunoblots were detected with anti-cMYC chicken IgY fraction (Invitrogen) and rabbit HRP-anti-chicken IgY(H+L) (Invitrogen), respectively. HA-tagged proteins were detected withanti-HA peroxidase (Roche).
Accession Numbers
Sequence data from this article can be found in the Arabidopsis GenomeInitiative or GenBank/EMBL databases under the following accessionnumbers: ZML2, At1g51600; ZML1, At3g21175; ZIM, At4g24470; ELIP2,
At4g14690; GPX7, At4g31870; ERD9, At1g10370; MYB7, At1g56650;RBCS, At1g67090; and LHCB2.4, At3g27690.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. Interaction of Synthetic Double-StrandedOligonucleotide Containing CryR1 cis-Element with Nuclear ProteinExtracts.
Supplemental Figure 2. Genotyping zml2, zml1, and zim T-DNAInsertion Lines.
Supplemental Figure 3. Expression Analysis of ZML2, ZML1, and ZIMin Response to HL and in the Different Genotypes.
Supplemental Figure 4. Real-Time Analysis of cry1-Dependent and-Independent Genes in the cry1, zml2-1, zml2-2, zml1, and zim MutantLines under Control Conditions.
Supplemental Figure 5. Superoxide Accumulation of zml2, zml1, andzim T-DNA Insertion Lines in Response to Excess Light.
Supplemental Figure 6. The zml2, zml1, and zim T-DNA InsertionLines Exposed to Excess Light.
Supplemental Figure 7. Hypocotyl Growth Response to Blue Light.
Supplemental Figure 8. Genotyping of zml2, zml1, and cry1 Doubleand Triple Mutants.
Supplemental Figure 9. Mascot Score Result and Ion Spectra for theZML2 Identification.
Supplemental Table 1. Occurrence of the CryR1 cis-Element in 500bp of the Promoters of Genes Induced in ZIM-ox Plants.
Supplemental Table 2. Primers Used in Real-Time PCR Analysis andfor Generation of Constructs.
ACKNOWLEDGMENTS
We thank Thorsten Seidel for providing the YFP and CFP vectors. Thiswork was supported by grants from the Swedish Research Foundation,Vetenskapsrådet, and the FFL2 grant from the Foundation for StrategicResearch (Å.S.). Å.S. is a Royal Swedish Academy of Sciences ResearchFellow supported by a grant from the Knut and Alice WallenbergFoundation.
AUTHOR CONTRIBUTIONS
Å.S. and J.S. designed the research. J.S., J.B.-L., K.Ö., D.K., A.S.G.,V.S., and Å.S. performed the experiments and analyzed data. G.W., K.Ö.,and L.B. contributed new analytic and experimental tools and analyzeddata. All authors contributed to writing the article.
Received May 1, 2012; revised June 5, 2012; accepted June 19, 2012;published July 10, 2012.
REFERENCES
Ahmad, M., and Cashmore, A.R. (1993). HY4 gene of A. thalianaencodes a protein with characteristics of a blue-light photoreceptor.Nature 366: 162–166.
ZML1/2, Key Components of the Photoprotective Response 15 of 17

Asada, K. (2006). Production and scavenging of reactive oxygenspecies in chloroplasts and their functions. Plant Physiol. 141:391–396.
Cashmore, A.R., Jarillo, J.A., Wu, Y.J., and Liu, D. (1999). Crypto-chromes: Blue light receptors for plants and animals. Science 284:760–765.
Chang, C.C., Slesak, I., Jordá, L., Sotnikov, A., Melzer, M.,Miszalski, Z., Mullineaux, P.M., Parker, J.E., Karpinska, B., andKarpinski, S. (2009). Arabidopsis chloroplastic glutathione perox-idases play a role in cross talk between photooxidative stress andimmune responses. Plant Physiol. 150: 670–683.
Chen, M., Chory, J., and Fankhauser, C. (2004). Light signal trans-duction in higher plants. Annu. Rev. Genet. 38: 87–117.
Chung, H.S., and Howe, G.A. (2009). A critical role for the TIFY motifin repression of jasmonate signaling by a stabilized splice variant ofthe JASMONATE ZIM-domain protein JAZ10 in Arabidopsis. PlantCell 21: 131–145.
Dean, C., Elzen, P.V., Tamaki, S., Dunsmuir, P., and Bedbrook, J.(1985). Differential expression of the eight genes of the petunia ri-bulose bisphosphate carboxylase small subunit multi-gene family.EMBO J. 4: 3055–3061.
Duek, P.D., and Fankhauser, C. (2003). HFR1, a putative bHLHtranscription factor, mediates both phytochrome A and crypto-chrome signalling. Plant J. 34: 827–836.
Gabrielsen, O.S., Hornes, E., Korsnes, L., Ruet, A., and Oyen, T.B.(1989). Magnetic DNA affinity purification of yeast transcriptionfactor tau—A new purification principle for the ultrarapid isolation ofnear homogeneous factor. Nucleic Acids Res. 17: 6253–6267.
Goulas, E., Schubert, M., Kieselbach, T., Kleczkowski, L.A.,Gardeström, P., Schröder, W.P., and Hurry, V. (2006). The chlo-roplast lumen and stromal proteomes of Arabidopsis thaliana showdifferential sensitivity to short- and long-term exposure to lowtemperature. Plant J. 47: 720–734.
Griffiths, S., Dunford, R.P., Coupland, G., and Laurie, D.A. (2003).The evolution of CONSTANS-like gene families in barley, rice, andArabidopsis. Plant Physiol. 131: 1855–1867.
Grob, U., and Stüber, K. (1987). Discrimination of phytochrome de-pendent light inducible from non-light inducible plant genes. Pre-diction of a common light-responsive element (LRE) in phytochromedependent light inducible plant genes. Nucleic Acids Res. 15:9957–9973.
Guo, H., Mockler, T., Duong, H., and Lin, C. (2001). SUB1, an Arab-idopsis Ca2+-binding protein involved in cryptochrome and phyto-chrome coaction. Science 291: 487–490.
Hong, S.H., Kim, H.J., Ryu, J.S., Choi, H., Jeong, S., Shin, J., Choi,G., and Nam, H.G. (2008). CRY1 inhibits COP1-mediated degra-dation of BIT1, a MYB transcription factor, to activate blue light-dependent gene expression in Arabidopsis. Plant J. 55: 361–371.
Jarillo, J.A., Capel, J., Tang, R.H., Yang, H.Q., Alonso, J.M., Ecker, J.R.,and Cashmore, A.R. (2001). An Arabidopsis circadian clock componentinteracts with both CRY1 and phyB. Nature 410: 487–490.
Jeong, M.J., and Shih, M.C. (2003). Interaction of a GATA factor withcis-acting elements involved in light regulation of nuclear genesencoding chloroplast glyceraldehyde-3-phosphate dehydrogenasein Arabidopsis. Biochem. Biophys. Res. Commun. 300: 555–562.Erratum. Biochem. Biophys. Res. Commun. 333: 1385.
Kang, X., Chong, J., and Ni, M. (2005). HYPERSENSITIVE TO REDAND BLUE 1, a ZZ-type zinc finger protein, regulates phytochromeB-mediated red and cryptochrome-mediated blue light responses.Plant Cell 17: 822–835.
Kang, X., and Ni, M. (2006). Arabidopsis SHORT HYPOCOTYL UN-DER BLUE1 contains SPX and EXS domains and acts in crypto-chrome signaling. Plant Cell 18: 921–934.
Kimura, M., Yoshizumi, T., Manabe, K., Yamamoto, Y.Y., andMatsui, M. (2001). Arabidopsis transcriptional regulation by lightstress via hydrogen peroxide-dependent and -independent path-ways. Genes Cells 6: 607–617.
Kindgren, P., Kremnev, D., Blanco, N.E., de Dios Barajas López, J.,Fernández, A.P., Tellgren-Roth, C., Small, I., and Strand, A.(2012). The plastid redox insensitive 2 mutant of Arabidopsis isimpaired in PEP activity and high light-dependent plastid redoxsignalling to the nucleus. Plant J. 70: 279–291.
Kiyosue, T., and Wada, M. (2000). LKP1 (LOV kelch protein 1): Afactor involved in the regulation of flowering time in arabidopsis.Plant J. 23: 807–815.
Kleine, T., Kindgren, P., Benedict, C., Hendrickson, L., and Strand,A. (2007). Genome-wide gene expression analysis reveals a criticalrole for CRYPTOCHROME1 in the response of Arabidopsis to highirradiance. Plant Physiol. 144: 1391–1406.
Kleine, T., Lockhart, P., and Batschauer, A. (2003). An Arabidopsisprotein closely related to Synechocystis cryptochrome is targetedto organelles. Plant J. 35: 93–103.
Kudla, B., Caddick, M.X., Langdon, T., Martinez-Rossi, N.M.,Bennett, C.F., Sibley, S., Davies, R.W., and Arst, H.N. Jr. (1990).The regulatory gene areA mediating nitrogen metabolite repressionin Aspergillus nidulans. Mutations affecting specificity of gene ac-tivation alter a loop residue of a putative zinc finger. EMBO J. 9:1355–1364.
Kurup, S., Jones, H.D., and Holdsworth, M.J. (2000). Interactions ofthe developmental regulator ABI3 with proteins identified from de-veloping Arabidopsis seeds. Plant J. 21: 143–155.
Lee, J.H., Yoo, S.J., Park, S.H., Hwang, I., Lee, J.S., and Ahn, J.H.(2007). Role of SVP in the control of flowering time by ambienttemperature in Arabidopsis. Genes Dev. 21: 397–402.
Li, Q.H., and Yang, H.Q. (2007). Cryptochrome signaling in plants.Photochem. Photobiol. 83: 94–101.
Li, Z., Wakao, S., Fischer, B.B., and Niyogi, K.K. (2009). Sensing andresponding to excess light. Annu. Rev. Plant Biol. 60: 239–260.
Lin, C., and Shalitin, D. (2003). Cryptochrome structure and signaltransduction. Annu. Rev. Plant Biol. 54: 469–496.
Lin, C., Yang, H., Guo, H., Mockler, T., Chen, J., and Cashmore,A.R. (1998). Enhancement of blue-light sensitivity of Arabidopsisseedlings by a blue light receptor cryptochrome 2. Proc. Natl. Acad.Sci. USA 95: 2686–2690.
Liu, H., Yu, X., Li, K., Klejnot, J., Yang, H., Lisiero, D., and Lin, C.(2008). Photoexcited CRY2 interacts with CIB1 to regulate tran-scription and floral initiation in Arabidopsis. Science 322:1535–1539.
Lopez-Molina, L., Mongrand, S., Kinoshita, N., and Chua, N.H.(2003). AFP is a novel negative regulator of ABA signaling thatpromotes ABI5 protein degradation. Genes Dev. 17: 410–418.
Manfield, I.W., Devlin, P.F., Jen, C.H., Westhead, D.R., andGilmartin, P.M. (2007). Conservation, convergence, and divergenceof light-responsive, circadian-regulated, and tissue-specific ex-pression patterns during evolution of the Arabidopsis GATA genefamily. Plant Physiol. 143: 941–958.
Miao, Y., Lv, D., Wang, P., Wang, X.C., Chen, J., Miao, C., andSong, C.P. (2006). An Arabidopsis glutathione peroxidase functionsas both a redox transducer and a scavenger in abscisic acid anddrought stress responses. Plant Cell 18: 2749–2766.
Møller, S.G., Kim, Y.S., Kunkel, T., and Chua, N.H. (2003). PP7 isa positive regulator of blue light signaling in Arabidopsis. Plant Cell15: 1111–1119.
Neff, M.M., and Chory, J. (1998). Genetic interactions betweenphytochrome A, phytochrome B, and cryptochrome 1 during Arab-idopsis development. Plant Physiol. 118: 27–35.
16 of 17 The Plant Cell

Pfaffl, M.W. (2001). A new mathematical model for relative quantifi-cation in real-time RT-PCR. Nucleic Acids Res. 29: e45.
Pokorny, R., Klar, T., Hennecke, U., Carell, T., Batschauer, A., andEssen, L.O. (2008). Recognition and repair of UV lesions in loopstructures of duplex DNA by DASH-type cryptochrome. Proc. Natl.Acad. Sci. USA 105: 21023–21027.
Porra, R.J., Thompson, W.A., and Kriedemann, P.E. (1989). De-termination of accurate extinction coefficients and simultaneousequations for assaying chlorophyll a and b extracted with four dif-ferent solvents: Verification of the concentration of chlorophyllstandards by atomic absorption spectroscopy. Biochim. Biophys.Acta 975: 384–394.
Rey, D.A., Pühler, A., and Kalinowski, J. (2003). The putative tran-scriptional repressor McbR, member of the TetR-family, is involvedin the regulation of the metabolic network directing the synthesis ofsulfur containing amino acids in Corynebacterium glutamicum. J.Biotechnol. 103: 51–65.
Richly, E., Dietzmann, A., Biehl, A., Kurth, J., Laloi, C., Apel, K.,Salamini, F., and Leister, D. (2003). Covariations in the nuclearchloroplast transcriptome reveal a regulatory master-switch. EMBORep. 4: 491–498.
Robson, F., Costa, M.M., Hepworth, S.R., Vizir, I., Piñeiro, M.,Reeves, P.H., Putterill, J., and Coupland, G. (2001). Functionalimportance of conserved domains in the flowering-time gene CON-STANS demonstrated by analysis of mutant alleles and transgenic plants.Plant J. 28: 619–631.
Rossel, J.B., Wilson, I.W., and Pogson, B.J. (2002). Global changesin gene expression in response to high light in Arabidopsis. PlantPhysiol. 130: 1109–1120.
Seidel, T., Golldack, D., and Dietz, K.J. (2005). Mapping of C-terminiof V-ATPase subunits by in vivo-FRET measurements. FEBS Lett.579: 4374–4382.
Seidel, T., Kluge, C., Hanitzsch, M., Ross, J., Sauer, M., Dietz, K.J.,and Golldack, D. (2004). Colocalization and FRET-analysis ofsubunits c and a of the vacuolar H+-ATPase in living plant cells. J.Biotechnol. 112: 165–175.
Seki, M., Kamei, A., Yamaguchi-Shinozaki, K., and Shinozaki, K.(2003). Molecular responses to drought, salinity and frost: Commonand different paths for plant protection. Curr. Opin. Biotechnol. 14:194–199.
Shaikhali, J., Heiber, I., Seidel, T., Ströher, E., Hiltscher, H.,Birkmann, S., Dietz, K.J., and Baier, M. (2008). The redox-sensitive transcription factor Rap2.4a controls nuclear expressionof 2-Cys peroxiredoxin A and other chloroplast antioxidant en-zymes. BMC Plant Biol. 8: 48.
Shikata, M., Matsuda, Y., Ando, K., Nishii, A., Takemura, M.,Yokota, A., and Kohchi, T. (2004). Characterization of ArabidopsisZIM, a member of a novel plant-specific GATA factor gene family. J.Exp. Bot. 55: 631–639.
Shikata, M., Takemura, M., Yokota, A., and Kohchi, T. (2003).Arabidopsis ZIM, a plant-specific GATA factor, can function as
a transcriptional activator. Biosci. Biotechnol. Biochem. 67:2495–2497.
Somers, D.E., Schultz, T.F., Milnamow, M., and Kay, S.A. (2000).ZEITLUPE encodes a novel clock-associated PAS protein fromArabidopsis. Cell 101: 319–329.
Srivastava, V., Srivastava, M.K., Chibani, K., Nilsson, R., Rouhier,N., Melzer, M., and Wingsle, G. (2009). Alternative splicing studiesof the reactive oxygen species gene network in Populus revealtwo isoforms of high-isoelectric-point superoxide dismutase. PlantPhysiol. 149: 1848–1859.
Strayer, C., Oyama, T., Schultz, T.F., Raman, R., Somers, D.E.,Más, P., Panda, S., Kreps, J.A., and Kay, S.A. (2000). Cloning ofthe Arabidopsis clock gene TOC1, an autoregulatory responseregulator homolog. Science 289: 768–771.
Suárez-López, P., Wheatley, K., Robson, F., Onouchi, H., Valverde,F., and Coupland, G. (2001). CONSTANS mediates between thecircadian clock and the control of flowering in Arabidopsis. Nature410: 1116–1120.
Teakle, G.R., Manfield, I.W., Graham, J.F., and Gilmartin, P.M.(2002). Arabidopsis thaliana GATA factors: Organisation, expressionand DNA-binding characteristics. Plant Mol. Biol. 50: 43–57.
Vanholme, B., Grunewald, W., Bateman, A., Kohchi, T., andGheysen, G. (2007). The tify family previously known as ZIM. TrendsPlant Sci. 12: 239–244.
Wagner, U., Edwards, R., Dixon, D.P., and Mauch, F. (2002).Probing the diversity of the Arabidopsis glutathione S-transferasegene family. Plant Mol. Biol. 49: 515–532.
Wang, H., Ma, L.G., Li, J.M., Zhao, H.Y., and Deng, X.W. (2001).Direct interaction of Arabidopsis cryptochromes with COP1 in lightcontrol development. Science 294: 154–158.
Ward, J.M., Cufr, C.A., Denzel, M.A., and Neff, M.M. (2005). The Doftranscription factor OBP3 modulates phytochrome and crypto-chrome signaling in Arabidopsis. Plant Cell 17: 475–485.
White, D.W. (2006). PEAPOD regulates lamina size and curvature inArabidopsis. Proc. Natl. Acad. Sci. USA 103: 13238–13243.
Wu, G., and Spalding, E.P. (2007). Separate functions for nuclear andcytoplasmic cryptochrome 1 during photomorphogenesis of Arab-idopsis seedlings. Proc. Natl. Acad. Sci. USA 104: 18813–18818.
Yanamandra, K. et al. (2009). Amyloid formation by the pro-inflammatory S100A8/A9 proteins in the ageing prostate. PLoS ONE4: e5562.
Yang, H.Q., Tang, R.H., and Cashmore, A.R. (2001). The signalingmechanism of Arabidopsis CRY1 involves direct interaction withCOP1. Plant Cell 13: 2573–2587.
Yang, H.Q., Wu, Y.J., Tang, R.H., Liu, D., Liu, Y., and Cashmore,A.R. (2000). The C termini of Arabidopsis cryptochromes mediatea constitutive light response. Cell 103: 815–827.
Yu, X., Klejnot, J., Zhao, X., Shalitin, D., Maymon, M., Yang, H.,Lee, J., Liu, X., Lopez, J., and Lin, C. (2007). Arabidopsis crypto-chrome 2 completes its posttranslational life cycle in the nucleus.Plant Cell 19: 3146–3156.
ZML1/2, Key Components of the Photoprotective Response 17 of 17

1 2!
Free DNA Probe!
DNA-Protein!Complex !
Supplemental Figure 1. Interaction of synthetic double-stranded oligonucleotide containing CryR1 cis-element with nuclear protein extracts.!EMSA was performed with synthetic double-stranded oligonucleotide labeled with biotin. Lane 1: free DNA probe. Lane 2: 25 µg of nuclear proteins from crude-nuclei protein extracts from Arabidopsis leaves incubated with 0.2 µmol DNA probe. The migration positions of the free DNA probe (lower band) and the DNA-protein complexes (upper band) are marked by arrows.!
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

B!
5’UTR 3’UTR
ATG
GK-448C11 (zml2-1)! GK-833C07 (zml2-2)!
ZML2!At1g51600!
ZML1!At3g21175!
5’UTR 3’UTR
ATG
Salk_069271 (zml1)!
ATG
5’UTR
Salk_144513 (zim)!
ZIM!At4g24470!
3’UTR
A!
zml2
-1!
wt! wt!
zml2
-1!
LB T-DNA!GKLB+P2!
Gene specific! P1+P2!
zml2
-2!
wt
wt
zml2
-2!
LB T-DNA!GKLB+P3!
Gene Specific! P3+P4!
zml1!
wt!
zml1!
wt!
LB T-DNA!SalkLB+P5!
Gene Specific! P5+P6!
wt!
zim!
wt!
zim!
LB T-DNA!SalkLB+P7!
Gene Specific! P7+P8!
P1! P2! P3! P4!
P5! P6!
P7! P8!
ZML2! ZML1! ZIM!
ACTIN!
C!
Supplemental Figure 2. Genotyping zml2, zml1 and zim T-DNA insertion lines. !(A) Genomic structures of the ZML2, ZML1 and ZIM genes and positions of the Gabi Kat and Salk T-DNA insertions. Exons are indicated by black boxes and introns by lines. Gray boxes indicate untranslated regions (5ʼ or 3ʼ UTR). (B) PCR with genomic DNA from wild type and T-DNA insertion plants. Primers used are indicated on the genomic structure schemes (C) Quantitative RT-PCR with gene specific primers was performed with cDNAs from wild type and zml2-1, zml2-2, zml1 and zim mutants. ACTIN2 (At5g09810) was used as a reference gene.!
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

Supplemental Figure 3. Expression analysis of ZML2, ZML1 and ZIM in response to HL and in the different genotypes. Expression of ZML2, ZML1 and ZIM in control (B-F) and following HL exposure (A,C,E,G) in wild type (A) and the mutant lines (B-G). The fold induction after HL exposure related to the control of each gene is presented. The results were normalized to the expression level of At4g36800 encoding a ubiquitin-protein ligase-like protein. The mean ± SD of three biological replicates is shown. !
A!
Rel
ativ
e ex
pres
sion!
0.0!
0.5!
1.0!
1.5!
0.0!
0.5!
1.0!
1.5!
2.0!
2.5!
3.0!
Rel
ativ
e ex
pres
sion!
WT! cry1! zml1! zim!
ZML2!
0.0!
0.5!
1.0!
1.5!
2.0!
2.5!
Rel
ativ
e ex
pres
sion!
WT! cry1! zml2! zim!
ZML1!
0!
0.5!
1!
1.5!
2!
2.5!
Rel
ativ
e ex
pres
sion!
WT! cry1! zml2! zml1!
ZIM!
0.0!
0.5!
1.0!
1.5!
Rel
ativ
e ex
pres
sion!
WT! cry1! zml1! zim!
ZML2!
0.0!
0.5!
1.0!
1.5!
Rel
ativ
e ex
pres
sion!
WT! cry1! zml2! zim!
ZML1!
0!
0.5!
1!
1.5!
Rel
ativ
e ex
pres
sion!
WT! cry1! zml2! zml1!
ZIM!
B! C!
D! E!
F! G!
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!
ZML2! ZML1!
ZIM!
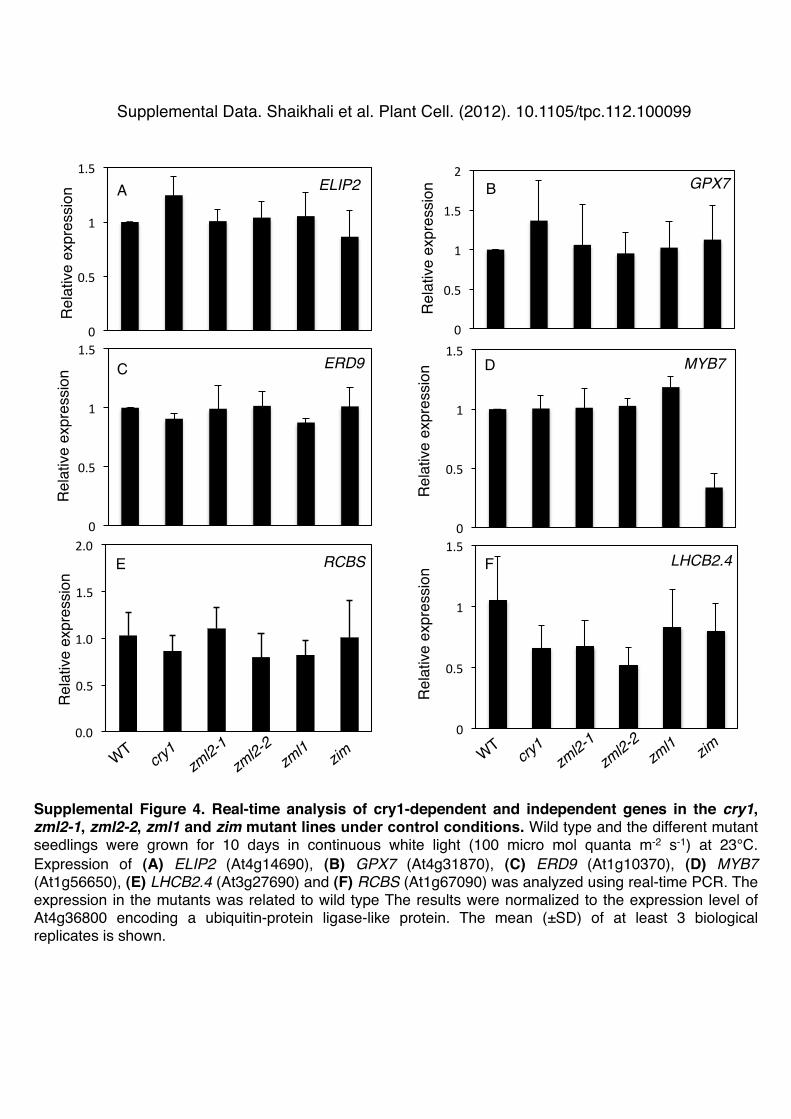
0.0
0.5
1.0
1.5
2.0
0
0.5
1
1.5
0
0.5
1
1.5 ERD9!
ELIP2! GPX7!
Rel
ativ
e ex
pres
sion!
Rel
ativ
e ex
pres
sion!
0
0.5
1
1.5 MYB7!
Rel
ativ
e ex
pres
sion!
A!
C!
E!
Supplemental Figure 4. Real-time analysis of cry1-dependent and independent genes in the cry1, zml2-1, zml2-2, zml1 and zim mutant lines under control conditions. Wild type and the different mutant seedlings were grown for 10 days in continuous white light (100 micro mol quanta m-2 s-1) at 23°C. Expression of (A) ELIP2 (At4g14690), (B) GPX7 (At4g31870), (C) ERD9 (At1g10370), (D) MYB7 (At1g56650), (E) LHCB2.4 (At3g27690) and (F) RCBS (At1g67090) was analyzed using real-time PCR. The expression in the mutants was related to wild type The results were normalized to the expression level of At4g36800 encoding a ubiquitin-protein ligase-like protein. The mean (±SD) of at least 3 biological replicates is shown. !
Rel
ativ
e ex
pres
sion!
Rel
ativ
e ex
pres
sion!
Rel
ativ
e ex
pres
sion!
B!
D!
F!
0
0.5
1
1.5 LHCB2.4!RCBS!
0
0.5
1
1.5
2
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

Supplemental Figure 5. Superoxide accumulation of zml2, zml1 and zim T-DNA insertion lines in response to excess light. !NBT staining indicating superoxide radical production in HL-exposed 3-week-old plants of wild type, cry1, zml2-1, zml2-2, zml1, and zim mutants. !
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

Supplemental Figure 6. The zml2, zml1 and zim T-DNA insertion lines exposed to excess light. 5-week-old plants of wild type, cry1 mutant and the zml2-1, zml2-2, zml1 and zim T-DNA insertion lines following 24 h exposure to excess light, HL (1000 micro mol quanta m-2 s-1). Photobleached leaves are indicated by arrows.!
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

0!
2!
4!
6!
8!
10!
wt! cry1! zml2-1! zml2-2! zml1! zim!
Hyp
ocot
yl L
engt
h (m
m)!
Supplemental Figure 7. Hypocotyl growth response to blue light.!Hypocotyl growth response of 4–day-old seedlings grown in blue light (10 micro mol quanta m-2 s-1). The mean (±SD) of at least 20 seedlings is shown for each genotype.!
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

A!
B!
C!
D!
0 2 4 6 8 10
Hyp
ocot
yl L
engt
h (m
m)!
WT!
cry1
zml2
-2!
WT!
cry1
zml2
-2!
Gene Specific! P3+P4!
LB T-DNA!GKLB+P4!
WT!
cry1
zml1!
WT!
cry1
zml1!
Gene Specific! P5+P6!
LB T-DNA!SalkLB+P5! 0
2 4 6 8 10
Hyp
ocot
yl L
engt
h (m
m)!
WT!
zml2
-2 z
ml1!
WT!
Gene Specific! P3+P4!
LB T-DNA!GKLB+P4!
WT!
WT!
Gene Specific! P5+P6!
LB T-DNA!SalkLB+P5!
WT!
cry1
zml2
-2zm
l1!
WT!
Gene Specific! P3+P4!
LB T-DNA!GKLB+P4!
WT!
WT!
Gene Specific! P5+P6!
LB T-DNA!SalkLB+P5!
0 2 4 6 8 10
Hyp
ocot
yl L
engt
h (m
m)!
Supplemental Figure 8. Genotyping of zml2, zml1 and cry1 double and triple mutants. PCR with genomic DNA from wild type and T-DNA insertion plants. Primers used are indicated on the genomic structure schemes (Figure S2). Hypocotyl length of 4–day-old seedlings grown in blue light (10 micro mol quanta m-2 s-1). The mean (±SD) of at least 20 seedlings is shown for each genotype. Genotyping (A) zml2-2zml1 (B) cry1zml2-2 (C) cry1zml1 (D) cry1zml2-2zml1!
zml2
-2 z
ml1!
zml2
-2 z
ml1!
zml2
-2 z
ml1!
cry1
zml2
-2zm
l1!
cry1
zml2
-2zm
l1!
cry1
zml2
-2zm
l1!
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!

Supplemental Figure 9. Mascot Score Result and ion spectra for the ZML2 identification !
Supplemental Data. Shaikhali et al. Plant Cell. (2012). 10.1105/tpc.112.100099!
Mascot Search Results: Peptide View
http://bio-mascot1.chem.umu.se/...2F20100111%2FF066271.dat&query=1&hit=1§ion=5&ave_thresh=32&tick1=0&tick_int=100&range=1300&index=AT1G51600.1&px=1[2010-01-11 16:03:15]
Mascot Search ResultsPeptide View
MS/MS Fragmentation of YTVRKEVALRFound in AT1G51600.1, Symbols: ZML2, TIFY2A | TIFY2A; sequence-specific DNA binding / transcription factor/ zinc ion binding |chr1:19133176-19135252 FORWARD
Match to Query 1: 1233.653823 from(412.225217,3+)Data file CryR1Control_412.22.dta
Click mouse within plot area to zoom in by factor of two about that pointOr, to Da
Monoisotopic mass of neutral peptide Mr(calc): 1233.7193Fixed modifications: Carbamidomethyl (C)Ions Score: 38 Expect: 0.013 Matches (Bold Red): 12/92 fragment ions using 20 most intense peaks
# b b++ b* b*++ b0 b0++ Seq. y y++ y* y*++ y0 y0++ #1 164.0706 82.5389 Y 102 265.1183 133.0628 247.1077 124.0575 T 1071.6633 536.3353 1054.6368 527.8220 1053.6527 527.3300 93 364.1867 182.5970 346.1761 173.5917 V 970.6156 485.8115 953.5891 477.2982 952.6051 476.8062 84 520.2878 260.6475 503.2613 252.1343 502.2772 251.6423 R 871.5472 436.2772 854.5207 427.7640 853.5366 427.2720 75 648.3828 324.6950 631.3562 316.1817 630.3722 315.6897 K 715.4461 358.2267 698.4196 349.7134 697.4355 349.2214 66 777.4254 389.2163 760.3988 380.7030 759.4148 380.2110 E 587.3511 294.1792 570.3246 285.6659 569.3406 285.1739 57 876.4938 438.7505 859.4672 430.2373 858.4832 429.7452 V 458.3085 229.6579 441.2820 221.1446 48 947.5309 474.2691 930.5043 465.7558 929.5203 465.2638 A 359.2401 180.1237 342.2136 171.6104 39 1060.6150 530.8111 1043.5884 522.2978 1042.6044 521.8058 L 288.2030 144.6051 271.1765 136.0919 2
10 R 175.1190 88.0631 158.0924 79.5498 1
NCBI BLAST search of YTVRKEVALR(Parameters: blastp, nr protein database, expect=20000, no filter, PAM30)Other BLAST web gateways
All matches to this query
Score Mr(calc): Delta Sequence38.1 1233.7193 -0.0655 YTVRKEVALR22.7 1233.6387 0.0151 IDMKTGDAVLR21.8 1233.6942 -0.0404 IGTSRRFEIR12.0 1233.6829 -0.0291 EFDLSKRAIR12.0 1233.7445 -0.0907 ELVGPALVPALR12.0 1233.5924 0.0614 GCSALWNSALR12.0 1233.7054 -0.0516 HELRRPSALR12.0 1233.6465 0.0073 RELSFNEALR12.0 1233.6618 -0.0080 SGWERFLALR12.0 1233.6686 -0.0147 SVMTGIMRALR
Mascot: http://www.matrixscience.com/

Supplemental Table 1: Occurrence of the CryR1 cis-element in 500 bp of the promoters of the genes induced in ZIM-ox plants (Shikata et al. 2004). Accession no. Occurrence of
the CryR1 cis-element
% identity with CryR1 consensus motif (GnTCKAG)
% identity with CryR1 as it occurs in the promoter of R2R3 (GTTCTAG)
At1g10550 x2 60 71 At1g01680 x1 80 71 At2g23680 x1 80 71 At1g15010 x2 60 57 At1g72240 x1 80 57 At5g64750 x2 80 85 At1g68690 x2 60 71 At1g75930 x2 60 71 At1g30900 x2 60 71 At1g05340 x2 80 85 At2g04450 x1 100 100 At3g48520 x2 80 71 At3g55840 x1 80 71 At1g02340 x2 80 85 At1g24530 x1 100 85 At2g34930 x2 60 71 At5g63230 x2 60 71 At3g57260 x2 80 85 At2g45680 x1 80 85 At2g41480 x1 80 85 At1g30720 x2 80 85 At3g45970 x1 80 85 At2g27690 x2 80 85 At4g14130 x2 100 100 At2g30540 x1 80 85 At2g15180 x2 60 71

Supplemental Table 2: Primers used in Real-Time PCR analysis and for generation of constructs Accession no.
Gene Identity
Forward Primer (5’ to 3’) Reverse Primer (5’ to 3’)
At4g36800 Ubiquitin like protein
CTGTTCACGGAACCCAATTC GGAAAAAGGTCTGACCGACA
At1g51600 ZML2 GTAGCTTTGAGGATGCAACGC
GAGATCTCTTGATGCTGAGC
At3g21175 ZML1 TAGGGTACTGGGTCTATCTG ACTGTATAGCGAATAGTCTT At4g22470 ZIM TAATGAAACCGGATGATGGT
GG AGATTTCTATCTTCTGATTC
At1g68690 pPK GCAAGATCAGATTCAGCATT CAACCAAATCCACCTTCACC At2g41480 pPRX ACTCTACTAGGTGCACATAC CATCGAATTTTGAGGGGCTA At4g14690 pCAB CAAGCCTAAGGTGAGTACGA TCCTAGAAACCACCCGACAC At4g31870 GPX7 ATTCGACAAGGTTGATGTGA TTGTGGGAGGGTATCTCTCG At1g10370 ERD9 CGACGAAAAGTGGTTTGTCG TCTCAACCAAGCCAAGAAGC At1g56650 MYB7 TGTAAGAGCTGGGCTAAACC
G CTATTCCCTAGAAGCCTATG
Protein expression and purification in Ecoli: ZML2-F1: 5’- CACCATGGATGACCTACATGGAA -3’ ZML2-R: 5’- TCACTGTGAGTTGCTTATGTCATT -3’ ZML1-F: 5’- CACCATGGATGATCTTCATGG -3’ ZML1-R 5’- TCACTGTGTGTTGCTAA -3’ Localization: BamHI-ZML2-F: 5’-AAAAGGATCCATGGATGACCTACATGG-3’ KpnI-ZML2-R: 5’-AAAAGGTACCCTGTGAGTTGCTTATGT-3’ Transactivation assay: Effector construct NcoI-ZML2-F: 5’-AAAACCATGGATGGATGACCTACATGG-3’ EcoRI-ZML2-R: 5’-AAAAGAATTCTCACTGTGAGTTGCTTA-3’ Reporter construct HindIII-R2R3cryR1-F: 5’-AAAAAAGCTTCTTCTTTAACTCG-3’ HindIII-R2R3cryR1-R: 5’-AAAAAAGCTTATGGTCCAGAGACCAG-3’ ChIP Fragments: F2 and F5 cryR1-F2: 5’- TCTTCTTTAACTCGTTAAATCT -3’ cryR1-R2: 5’- AGTTCGAATAAAGAGAGA -3’ cryR1-F5: 5’-TGACCAATAGATGATATACA-3’ cryR1-R5: 5’-AACCATTGATGGAAATGGTCT-3’ Y2H-Primer for Bait constructs: EcoRI-ZML2-F: 5-AAAA GAATTC ATGGATGACCTACATGG-3 KpnI-ZML2-R: 5-AAAA GGTACC TCACTGTGAGTTGCTTA-3 EcoRI-ZML1-F: 5-AAAA GAATTC ATGGATGATCTTCATGG-3

KpnI-ZML1-R: 5-AAAA GGTACC TCACTGTGTGTTGCTTA-3 Y2H-Primer for prey constructs: SmaI-ZML1-F: 5-AAAA CCCGGG ATGGATGATCTTCATGG-3 XhoI -ZML1-R: 5-AAAA CTCGAG TCACTGTGTGTTGCTTA-3 SmaI-cry1-CT-F: 5’-AAAA CCCGGG ATGTGGCAACTAGAAGCTGC-3’ XhoI -cry1-R: 5-AAAA CTCGAG TTACCCGGTTTGTGAAAG-3 CoIP-Primer for HA constructs: BamHI-ZML2-F: 5’-AAAAGGATCCATGGATGACCTACATGG-3’ EcoRI-ZML2-R: 5’-AAAAGAATTCTCACTGTGAGTTGCTTA-3’ CoIP-Primer for myc constructs: EcoRI-ZML1-F: 5-AAAA GAATTC ATGGATGATCTTCATGG-3 KpnI-ZML1-R: 5-AAAA GGTACC TCACTGTGTGTTGCTTA-3 SmaI-cry1-F: 5-AAAA CCCGGG ATGTCTGGTTCTGTATC-3 EcoRI-cry1-R: 5-AAAA GAATTC TTACCCGGTTTGTGAAAG-3

DOI 10.1105/tpc.112.100099; originally published online July 10, 2012;Plant Cell
Vaibhav Srivastava, Gunnar Wingsle, Laszlo Bako and Åsa StrandJehad Shaikhali, Juan de Dios Barajas-Lopéz, Krisztina Ötvös, Dmitry Kremnev, Ana Sánchez Garcia,
ArabidopsisMERISTEM LIKE1 and ZML2 in Transcriptional Activators ZINC FINGER PROTEIN EXPRESSED IN INFLORESCENCE
The CRYPTOCHROME1-Dependent Response to Excess Light Is Mediated through the
This information is current as of July 13, 2012
Supplemental Data http://www.plantcell.org/content/suppl/2012/07/03/tpc.112.100099.DC1.html
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists
Copyright © 2022 FDOKUMEN







![Regeneration Of Three Sweet Potato [Ipomoea Batatas(L.)] Accessions in Ghana via Meristem And Nodal culture](https://static.fdokumen.com/doc/165x107/631af948d43f4e176304af45/regeneration-of-three-sweet-potato-ipomoea-batatasl-accessions-in-ghana-via.jpg)












