
The Cholesterol Absorption Inhibitor Ezetimibe Acts by Blocking the Sterol-Induced Internalization...
12
Cell Metabolism Article The Cholesterol Absorption Inhibitor Ezetimibe Acts by Blocking the Sterol-Induced Internalization of NPC1L1 Liang Ge, 1,3 Jing Wang, 1,3 Wei Qi, 2,3 Hong-Hua Miao, 1 Jian Cao, 1 Yu-Xiu Qu, 1 Bo-Liang Li, 1 and Bao-Liang Song 1, * 1 State Key Laboratory of Molecular Biology, Institute of Biochemistry and Cell Biology 2 Key Laboratory of Nutrition and Metabolism, Institute for Nutritional Sciences Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200031, China 3 These authors contributed equally to this work *Correspondence: [email protected] DOI 10.1016/j.cmet.2008.04.001 SUMMARY Niemann-Pick C1-like 1 (NPC1L1) is a polytopic transmembrane protein that plays a critical role in cholesterol absorption. Ezetimibe, a hypocholestero- lemic drug, has been reported to bind NPC1L1 and block cholesterol absorption. However, the molecu- lar mechanism of NPC1L1-mediated cholesterol uptake and how ezetimibe inhibits this process are poorly defined. Here we find that cholesterol specif- ically promotes the internalization of NPC1L1 and that this process requires microfilaments and the cla- thrin/AP2 complex. Blocking NPC1L1 endocytosis dramatically decreases cholesterol internalization, indicating that NPC1L1 mediates cholesterol uptake via its vesicular endocytosis. Ezetimibe prevents NPC1L1 from incorporating into clathrin-coated ves- icles and thus inhibits cholesterol uptake. Together, our data suggest a model wherein cholesterol is in- ternalized into cells with NPC1L1 through clathrin/ AP2-mediated endocytosis and ezetimibe inhibits cholesterol absorption by blocking the internaliza- tion of NPC1L1. INTRODUCTION Cholesterol is an essential component of most biological mem- branes and is the precursor for synthesis of steroid hormones and bile acids. However, high levels of cholesterol cause severe problems including coronary heart disease (CHD). In developed societies, excessive dietary cholesterol uptake is a major risk factor for CHD. Mammals obtain cholesterol by de novo synthe- sis from acetyl-CoA and by absorption from diet and bile. Although much is known about the enzymology and regulatory pathways of cholesterol biosynthesis (Brown and Goldstein, 1986; Goldstein et al., 2006), far less is understood about the molecular mechanisms by which cholesterol is absorbed. Niemann-Pick C1-like 1 (NPC1L1) was recently identified as a critical player in the intestinal absorption of dietary cholesterol (Altmann et al., 2004). It is highly expressed in small intestine and localized along the brush border in both human and mouse (Davis et al., 2004; Altmann et al., 2004; Sane et al., 2006). How- ever, significant expression of NPC1L1 is also observed in human liver, but not in mouse liver (Davies et al., 2005; Altmann et al., 2004). NPC1L1-deficient mice exhibit a drastic reduction of dietary cholesterol absorption (Davies et al., 2005; Davis et al., 2004; Altmann et al., 2004). Besides mediating intestinal cholesterol uptake, overexpression of NPC1L1 in liver in a trans- genic mouse model dramatically decreases biliary cholesterol concentration and increases plasma cholesterol level, indicating that liver NPC1L1 mediates biliary cholesterol reabsorption (Temel et al., 2007). Taken together, these data indicate that NPC1L1 plays key roles in dietary and biliary cholesterol absorp- tion in both intestine and liver. The NPC1L1 protein contains about 1300 residues with 13 predicted transmembrane domains (Davies et al., 2000). Inter- estingly, the third to seventh transmembrane helices constitute a sterol-sensing domain that is also present in other sterol-regu- lated proteins including NPC1, HMG-CoA reductase, and SCAP (Kuwabara and Labouesse, 2002). NPC1L1 has been reported to localize on the plasma membrane (PM) (Altmann et al., 2004; Iyer et al., 2005) or in intracellular compartments (Davies et al., 2005; Sane et al., 2006). More recently, Yu et al. found that NPC1L1 is transported from the endocytic recycling compartment (ERC) to the PM after cholesterol depletion and that only when localized on the PM can it promote cholesterol uptake (Brown et al., 2007; Yu et al., 2006). Ezetimibe (trademark name Zetia) is a potent cholesterol absorption inhibitor that is being clinically used to treat hyper- cholesterolemia (Davis and Veltri, 2007). NPC1L1 knockout mice show no response to ezetimibe (Altmann et al., 2004), and the phenotypes of NPC1L1 transgenic mice can be normal- ized by ezetimibe (Temel et al., 2007). These findings indicate that NPC1L1 and ezetimibe function in the same pathway. It has been further found that ezetimibe binds to brush-border membranes from wild-type animals but not those of NPC1L1 knockout mice, suggesting that ezetimibe interacts with NPC1L1 and presumably inactivates it (Davis and Veltri, 2007; Garcia-Calvo et al., 2005). However, how ezetimibe inhibits NPC1L1-mediated cholesterol uptake is largely unknown. In the present study, we find that NPC1L1 recycles between ERC and PM: depletion of cholesterol causes the transport of NPC1L1 from ERC to PM, whereas replenishment of cholesterol results in the transportation of NPC1L1 from PM to ERC. Mean- while, cholesterol is internalized together with NPC1L1. Through disruption of microfilaments and knockdown of clathrin/AP2, we 508 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The Cholesterol Absorption Inhibitor Ezetimibe Acts by Blocking the Sterol-Induced Internalization...

Cell Metabolism
Article
The Cholesterol Absorption Inhibitor EzetimibeActs by Blocking the Sterol-InducedInternalization of NPC1L1Liang Ge,1,3 Jing Wang,1,3 Wei Qi,2,3 Hong-Hua Miao,1 Jian Cao,1 Yu-Xiu Qu,1 Bo-Liang Li,1 and Bao-Liang Song1,*1State Key Laboratory of Molecular Biology, Institute of Biochemistry and Cell Biology2Key Laboratory of Nutrition and Metabolism, Institute for Nutritional Sciences
Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200031, China3These authors contributed equally to this work
*Correspondence: [email protected]
DOI 10.1016/j.cmet.2008.04.001
SUMMARY
Niemann-Pick C1-like 1 (NPC1L1) is a polytopictransmembrane protein that plays a critical role incholesterol absorption. Ezetimibe, a hypocholestero-lemic drug, has been reported to bind NPC1L1 andblock cholesterol absorption. However, the molecu-lar mechanism of NPC1L1-mediated cholesteroluptake and how ezetimibe inhibits this process arepoorly defined. Here we find that cholesterol specif-ically promotes the internalization of NPC1L1 andthat this process requires microfilaments and the cla-thrin/AP2 complex. Blocking NPC1L1 endocytosisdramatically decreases cholesterol internalization,indicating that NPC1L1 mediates cholesterol uptakevia its vesicular endocytosis. Ezetimibe preventsNPC1L1 from incorporating into clathrin-coated ves-icles and thus inhibits cholesterol uptake. Together,our data suggest a model wherein cholesterol is in-ternalized into cells with NPC1L1 through clathrin/AP2-mediated endocytosis and ezetimibe inhibitscholesterol absorption by blocking the internaliza-tion of NPC1L1.
INTRODUCTION
Cholesterol is an essential component of most biological mem-
branes and is the precursor for synthesis of steroid hormones
and bile acids. However, high levels of cholesterol cause severe
problems including coronary heart disease (CHD). In developed
societies, excessive dietary cholesterol uptake is a major risk
factor for CHD. Mammals obtain cholesterol by de novo synthe-
sis from acetyl-CoA and by absorption from diet and bile.
Although much is known about the enzymology and regulatory
pathways of cholesterol biosynthesis (Brown and Goldstein,
1986; Goldstein et al., 2006), far less is understood about the
molecular mechanisms by which cholesterol is absorbed.
Niemann-Pick C1-like 1 (NPC1L1) was recently identified as
a critical player in the intestinal absorption of dietary cholesterol
(Altmann et al., 2004). It is highly expressed in small intestine and
localized along the brush border in both human and mouse
(Davis et al., 2004; Altmann et al., 2004; Sane et al., 2006). How-
508 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.
ever, significant expression of NPC1L1 is also observed in
human liver, but not in mouse liver (Davies et al., 2005; Altmann
et al., 2004). NPC1L1-deficient mice exhibit a drastic reduction
of dietary cholesterol absorption (Davies et al., 2005; Davis
et al., 2004; Altmann et al., 2004). Besides mediating intestinal
cholesterol uptake, overexpression of NPC1L1 in liver in a trans-
genic mouse model dramatically decreases biliary cholesterol
concentration and increases plasma cholesterol level, indicating
that liver NPC1L1 mediates biliary cholesterol reabsorption
(Temel et al., 2007). Taken together, these data indicate that
NPC1L1 plays key roles in dietary and biliary cholesterol absorp-
tion in both intestine and liver.
The NPC1L1 protein contains about 1300 residues with 13
predicted transmembrane domains (Davies et al., 2000). Inter-
estingly, the third to seventh transmembrane helices constitute
a sterol-sensing domain that is also present in other sterol-regu-
lated proteins including NPC1, HMG-CoA reductase, and SCAP
(Kuwabara and Labouesse, 2002). NPC1L1 has been reported to
localize on the plasma membrane (PM) (Altmann et al., 2004; Iyer
et al., 2005) or in intracellular compartments (Davies et al., 2005;
Sane et al., 2006). More recently, Yu et al. found that NPC1L1 is
transported from the endocytic recycling compartment (ERC) to
the PM after cholesterol depletion and that only when localized
on the PM can it promote cholesterol uptake (Brown et al.,
2007; Yu et al., 2006).
Ezetimibe (trademark name Zetia) is a potent cholesterol
absorption inhibitor that is being clinically used to treat hyper-
cholesterolemia (Davis and Veltri, 2007). NPC1L1 knockout
mice show no response to ezetimibe (Altmann et al., 2004),
and the phenotypes of NPC1L1 transgenic mice can be normal-
ized by ezetimibe (Temel et al., 2007). These findings indicate
that NPC1L1 and ezetimibe function in the same pathway. It
has been further found that ezetimibe binds to brush-border
membranes from wild-type animals but not those of NPC1L1
knockout mice, suggesting that ezetimibe interacts with
NPC1L1 and presumably inactivates it (Davis and Veltri, 2007;
Garcia-Calvo et al., 2005). However, how ezetimibe inhibits
NPC1L1-mediated cholesterol uptake is largely unknown.
In the present study, we find that NPC1L1 recycles between
ERC and PM: depletion of cholesterol causes the transport of
NPC1L1 from ERC to PM, whereas replenishment of cholesterol
results in the transportation of NPC1L1 from PM to ERC. Mean-
while, cholesterol is internalized together with NPC1L1. Through
disruption of microfilaments and knockdown of clathrin/AP2, we
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
demonstrate that the endocytosis of NPC1L1 is dependent
on microfilaments and the clathrin/AP2 complex. Blocking
NPC1L1 endocytosis decreases cholesterol uptake, indicating
that NPC1L1 mediates cholesterol uptake through vesicular
endocytosis. We further find that ezetimibe blocks the internali-
zation of NPC1L1, thereby inhibiting cholesterol uptake. In sum-
mary, our results reveal the mechanism of NPC1L1-mediated
cholesterol uptake and how ezetimibe inhibits the process.
RESULTS
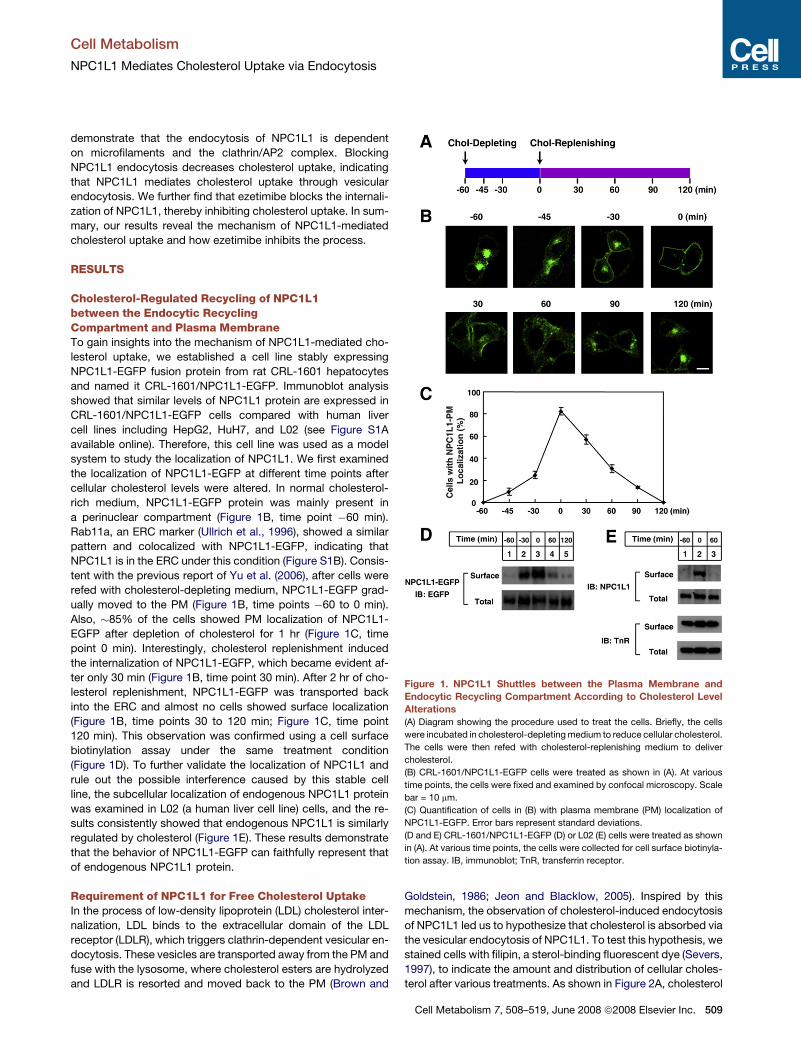
Cholesterol-Regulated Recycling of NPC1L1between the Endocytic RecyclingCompartment and Plasma MembraneTo gain insights into the mechanism of NPC1L1-mediated cho-
lesterol uptake, we established a cell line stably expressing
NPC1L1-EGFP fusion protein from rat CRL-1601 hepatocytes
and named it CRL-1601/NPC1L1-EGFP. Immunoblot analysis
showed that similar levels of NPC1L1 protein are expressed in
CRL-1601/NPC1L1-EGFP cells compared with human liver
cell lines including HepG2, HuH7, and L02 (see Figure S1A
available online). Therefore, this cell line was used as a model
system to study the localization of NPC1L1. We first examined
the localization of NPC1L1-EGFP at different time points after
cellular cholesterol levels were altered. In normal cholesterol-
rich medium, NPC1L1-EGFP protein was mainly present in
a perinuclear compartment (Figure 1B, time point �60 min).
Rab11a, an ERC marker (Ullrich et al., 1996), showed a similar
pattern and colocalized with NPC1L1-EGFP, indicating that
NPC1L1 is in the ERC under this condition (Figure S1B). Consis-
tent with the previous report of Yu et al. (2006), after cells were
refed with cholesterol-depleting medium, NPC1L1-EGFP grad-
ually moved to the PM (Figure 1B, time points �60 to 0 min).
Also, �85% of the cells showed PM localization of NPC1L1-
EGFP after depletion of cholesterol for 1 hr (Figure 1C, time
point 0 min). Interestingly, cholesterol replenishment induced
the internalization of NPC1L1-EGFP, which became evident af-
ter only 30 min (Figure 1B, time point 30 min). After 2 hr of cho-
lesterol replenishment, NPC1L1-EGFP was transported back
into the ERC and almost no cells showed surface localization
(Figure 1B, time points 30 to 120 min; Figure 1C, time point
120 min). This observation was confirmed using a cell surface
biotinylation assay under the same treatment condition
(Figure 1D). To further validate the localization of NPC1L1 and
rule out the possible interference caused by this stable cell
line, the subcellular localization of endogenous NPC1L1 protein
was examined in L02 (a human liver cell line) cells, and the re-
sults consistently showed that endogenous NPC1L1 is similarly
regulated by cholesterol (Figure 1E). These results demonstrate
that the behavior of NPC1L1-EGFP can faithfully represent that
of endogenous NPC1L1 protein.
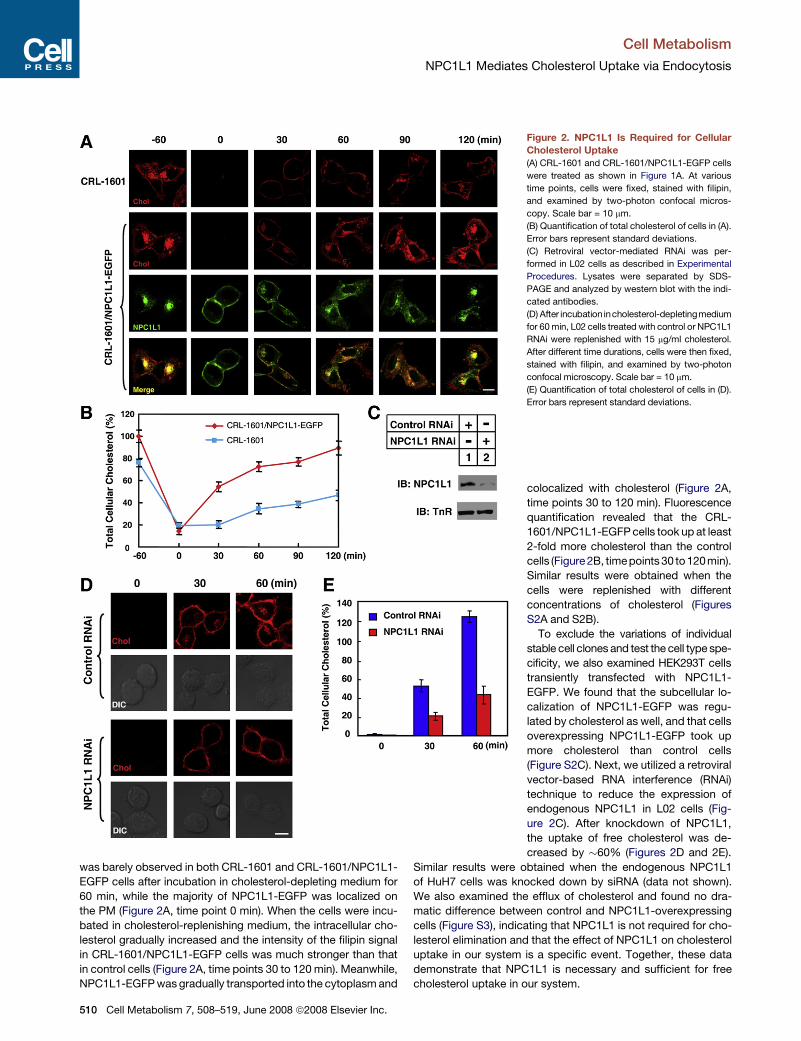
Requirement of NPC1L1 for Free Cholesterol UptakeIn the process of low-density lipoprotein (LDL) cholesterol inter-
nalization, LDL binds to the extracellular domain of the LDL
receptor (LDLR), which triggers clathrin-dependent vesicular en-
docytosis. These vesicles are transported away from the PM and
fuse with the lysosome, where cholesterol esters are hydrolyzed
and LDLR is resorted and moved back to the PM (Brown and
Goldstein, 1986; Jeon and Blacklow, 2005). Inspired by this
mechanism, the observation of cholesterol-induced endocytosis
of NPC1L1 led us to hypothesize that cholesterol is absorbed via
the vesicular endocytosis of NPC1L1. To test this hypothesis, we
stained cells with filipin, a sterol-binding fluorescent dye (Severs,
1997), to indicate the amount and distribution of cellular choles-
terol after various treatments. As shown in Figure 2A, cholesterol
Figure 1. NPC1L1 Shuttles between the Plasma Membrane and
Endocytic Recycling Compartment According to Cholesterol Level
Alterations
(A) Diagram showing the procedure used to treat the cells. Briefly, the cells
were incubated in cholesterol-depleting medium to reduce cellular cholesterol.
The cells were then refed with cholesterol-replenishing medium to deliver
cholesterol.
(B) CRL-1601/NPC1L1-EGFP cells were treated as shown in (A). At various
time points, the cells were fixed and examined by confocal microscopy. Scale
bar = 10 mm.
(C) Quantification of cells in (B) with plasma membrane (PM) localization of
NPC1L1-EGFP. Error bars represent standard deviations.
(D and E) CRL-1601/NPC1L1-EGFP (D) or L02 (E) cells were treated as shown
in (A). At various time points, the cells were collected for cell surface biotinyla-
tion assay. IB, immunoblot; TnR, transferrin receptor.
Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc. 509
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
was barely observed in both CRL-1601 and CRL-1601/NPC1L1-
EGFP cells after incubation in cholesterol-depleting medium for
60 min, while the majority of NPC1L1-EGFP was localized on
the PM (Figure 2A, time point 0 min). When the cells were incu-
bated in cholesterol-replenishing medium, the intracellular cho-
lesterol gradually increased and the intensity of the filipin signal
in CRL-1601/NPC1L1-EGFP cells was much stronger than that
in control cells (Figure 2A, time points 30 to 120 min). Meanwhile,
NPC1L1-EGFP was gradually transported into the cytoplasm and
Figure 2. NPC1L1 Is Required for Cellular
Cholesterol Uptake
(A) CRL-1601 and CRL-1601/NPC1L1-EGFP cells
were treated as shown in Figure 1A. At various
time points, cells were fixed, stained with filipin,
and examined by two-photon confocal micros-
copy. Scale bar = 10 mm.
(B) Quantification of total cholesterol of cells in (A).
Error bars represent standard deviations.
(C) Retroviral vector-mediated RNAi was per-
formed in L02 cells as described in Experimental
Procedures. Lysates were separated by SDS-
PAGE and analyzed by western blot with the indi-
cated antibodies.
(D) After incubation incholesterol-depletingmedium
for 60 min, L02 cells treated with control or NPC1L1
RNAi were replenished with 15 mg/ml cholesterol.
After different time durations, cells were then fixed,
stained with filipin, and examined by two-photon
confocal microscopy. Scale bar = 10 mm.
(E) Quantification of total cholesterol of cells in (D).
Error bars represent standard deviations.
colocalized with cholesterol (Figure 2A,
time points 30 to 120 min). Fluorescence
quantification revealed that the CRL-
1601/NPC1L1-EGFP cells took up at least
2-fold more cholesterol than the control
cells (Figure 2B, timepoints 30 to 120min).
Similar results were obtained when the
cells were replenished with different
concentrations of cholesterol (Figures
S2A and S2B).
To exclude the variations of individual
stable cell clones and test the cell type spe-
cificity, we also examined HEK293T cells
transiently transfected with NPC1L1-
EGFP. We found that the subcellular lo-
calization of NPC1L1-EGFP was regu-
lated by cholesterol as well, and that cells
overexpressing NPC1L1-EGFP took up
more cholesterol than control cells
(Figure S2C). Next, we utilized a retroviral
vector-based RNA interference (RNAi)
technique to reduce the expression of
endogenous NPC1L1 in L02 cells (Fig-
ure 2C). After knockdown of NPC1L1,
the uptake of free cholesterol was de-
creased by �60% (Figures 2D and 2E).
Similar results were obtained when the endogenous NPC1L1
of HuH7 cells was knocked down by siRNA (data not shown).
We also examined the efflux of cholesterol and found no dra-
matic difference between control and NPC1L1-overexpressing
cells (Figure S3), indicating that NPC1L1 is not required for cho-
lesterol elimination and that the effect of NPC1L1 on cholesterol
uptake in our system is a specific event. Together, these data
demonstrate that NPC1L1 is necessary and sufficient for free
cholesterol uptake in our system.
510 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
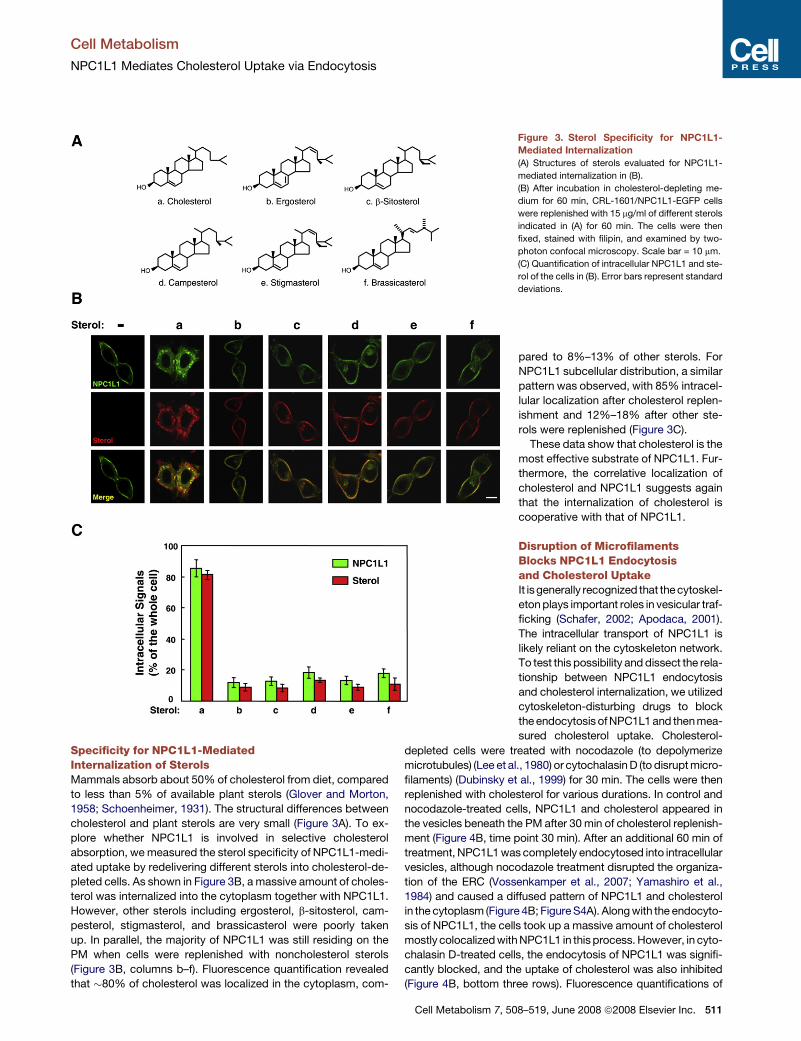
Specificity for NPC1L1-MediatedInternalization of SterolsMammals absorb about 50% of cholesterol from diet, compared
to less than 5% of available plant sterols (Glover and Morton,
1958; Schoenheimer, 1931). The structural differences between
cholesterol and plant sterols are very small (Figure 3A). To ex-
plore whether NPC1L1 is involved in selective cholesterol
absorption, we measured the sterol specificity of NPC1L1-medi-
ated uptake by redelivering different sterols into cholesterol-de-
pleted cells. As shown in Figure 3B, a massive amount of choles-
terol was internalized into the cytoplasm together with NPC1L1.
However, other sterols including ergosterol, b-sitosterol, cam-
pesterol, stigmasterol, and brassicasterol were poorly taken
up. In parallel, the majority of NPC1L1 was still residing on the
PM when cells were replenished with noncholesterol sterols
(Figure 3B, columns b–f). Fluorescence quantification revealed
that �80% of cholesterol was localized in the cytoplasm, com-
Figure 3. Sterol Specificity for NPC1L1-
Mediated Internalization
(A) Structures of sterols evaluated for NPC1L1-
mediated internalization in (B).
(B) After incubation in cholesterol-depleting me-
dium for 60 min, CRL-1601/NPC1L1-EGFP cells
were replenished with 15 mg/ml of different sterols
indicated in (A) for 60 min. The cells were then
fixed, stained with filipin, and examined by two-
photon confocal microscopy. Scale bar = 10 mm.
(C) Quantification of intracellular NPC1L1 and ste-
rol of the cells in (B). Error bars represent standard
deviations.
pared to 8%–13% of other sterols. For
NPC1L1 subcellular distribution, a similar
pattern was observed, with 85% intracel-
lular localization after cholesterol replen-
ishment and 12%–18% after other ste-
rols were replenished (Figure 3C).
These data show that cholesterol is the
most effective substrate of NPC1L1. Fur-
thermore, the correlative localization of
cholesterol and NPC1L1 suggests again
that the internalization of cholesterol is
cooperative with that of NPC1L1.
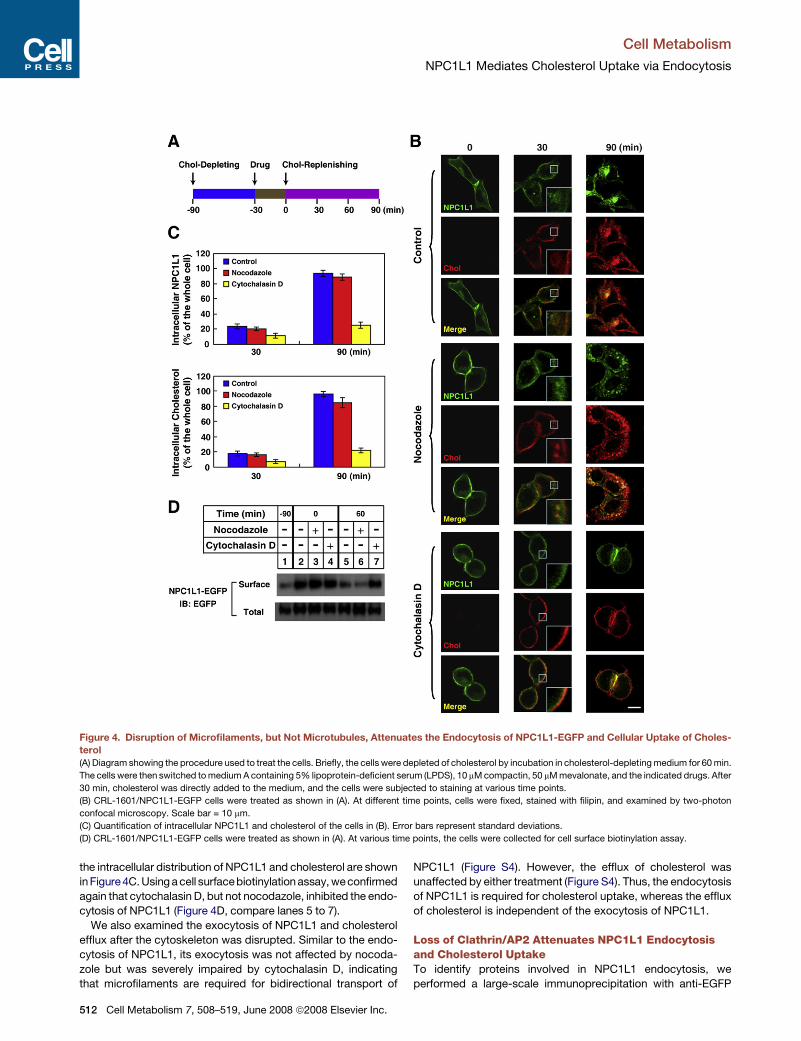
Disruption of MicrofilamentsBlocks NPC1L1 Endocytosisand Cholesterol UptakeIt isgenerally recognized that the cytoskel-
eton plays important roles in vesicular traf-
ficking (Schafer, 2002; Apodaca, 2001).
The intracellular transport of NPC1L1 is
likely reliant on the cytoskeleton network.
To test this possibility and dissect the rela-
tionship between NPC1L1 endocytosis
and cholesterol internalization, we utilized
cytoskeleton-disturbing drugs to block
the endocytosis of NPC1L1 and then mea-
sured cholesterol uptake. Cholesterol-
depleted cells were treated with nocodazole (to depolymerize
microtubules) (Lee et al., 1980) or cytochalasin D (to disrupt micro-
filaments) (Dubinsky et al., 1999) for 30 min. The cells were then
replenished with cholesterol for various durations. In control and
nocodazole-treated cells, NPC1L1 and cholesterol appeared in
the vesicles beneath the PM after 30 min of cholesterol replenish-
ment (Figure 4B, time point 30 min). After an additional 60 min of
treatment, NPC1L1 was completely endocytosed into intracellular
vesicles, although nocodazole treatment disrupted the organiza-
tion of the ERC (Vossenkamper et al., 2007; Yamashiro et al.,
1984) and caused a diffused pattern of NPC1L1 and cholesterol
in the cytoplasm (Figure 4B; Figure S4A). Along with the endocyto-
sis of NPC1L1, the cells took up a massive amount of cholesterol
mostly colocalized with NPC1L1 in this process. However, in cyto-
chalasin D-treated cells, the endocytosis of NPC1L1 was signifi-
cantly blocked, and the uptake of cholesterol was also inhibited
(Figure 4B, bottom three rows). Fluorescence quantifications of
Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc. 511
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
Figure 4. Disruption of Microfilaments, but Not Microtubules, Attenuates the Endocytosis of NPC1L1-EGFP and Cellular Uptake of Choles-
terol
(A) Diagram showing the procedure used to treat the cells. Briefly, the cells were depleted of cholesterol by incubation in cholesterol-depleting medium for 60 min.
The cells were then switched to medium A containing 5% lipoprotein-deficient serum (LPDS), 10 mM compactin, 50 mM mevalonate, and the indicated drugs. After
30 min, cholesterol was directly added to the medium, and the cells were subjected to staining at various time points.
(B) CRL-1601/NPC1L1-EGFP cells were treated as shown in (A). At different time points, cells were fixed, stained with filipin, and examined by two-photon
confocal microscopy. Scale bar = 10 mm.
(C) Quantification of intracellular NPC1L1 and cholesterol of the cells in (B). Error bars represent standard deviations.
(D) CRL-1601/NPC1L1-EGFP cells were treated as shown in (A). At various time points, the cells were collected for cell surface biotinylation assay.
the intracellular distribution of NPC1L1 and cholesterol are shown
in Figure 4C. Using a cell surfacebiotinylation assay, weconfirmed
again that cytochalasin D, but not nocodazole, inhibited the endo-
cytosis of NPC1L1 (Figure 4D, compare lanes 5 to 7).
We also examined the exocytosis of NPC1L1 and cholesterol
efflux after the cytoskeleton was disrupted. Similar to the endo-
cytosis of NPC1L1, its exocytosis was not affected by nocoda-
zole but was severely impaired by cytochalasin D, indicating
that microfilaments are required for bidirectional transport of
512 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.
NPC1L1 (Figure S4). However, the efflux of cholesterol was
unaffected by either treatment (Figure S4). Thus, the endocytosis
of NPC1L1 is required for cholesterol uptake, whereas the efflux
of cholesterol is independent of the exocytosis of NPC1L1.
Loss of Clathrin/AP2 Attenuates NPC1L1 Endocytosisand Cholesterol UptakeTo identify proteins involved in NPC1L1 endocytosis, we
performed a large-scale immunoprecipitation with anti-EGFP
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
agarose from detergent lysates of the membrane fraction of
CRL-1601/NPC1L1-EGFP cells. Specific bands were identified
by tandem mass spectrometry (Figure S5), one of which was
the m2 subunit of the AP2 complex. The AP2 adaptor complex,
which consists of a, b2, m2, and s2 subunits, mainly functions
to select cargo on the PM for inclusion into clathrin-coated
vesicles in the endocytic pathway (Owen et al., 2004). Coimmu-
noprecipitation experiments confirmed that NPC1L1-EGFP, m2,
and clathrin heavy chain (CHC) are present in the same protein
complex (Figure S5C). These data indicate that clathrin-medi-
ated endocytosis is involved in the transport of NPC1L1 protein.
This hypothesis was next addressed by diminishing CHC or m2
through RNAi.
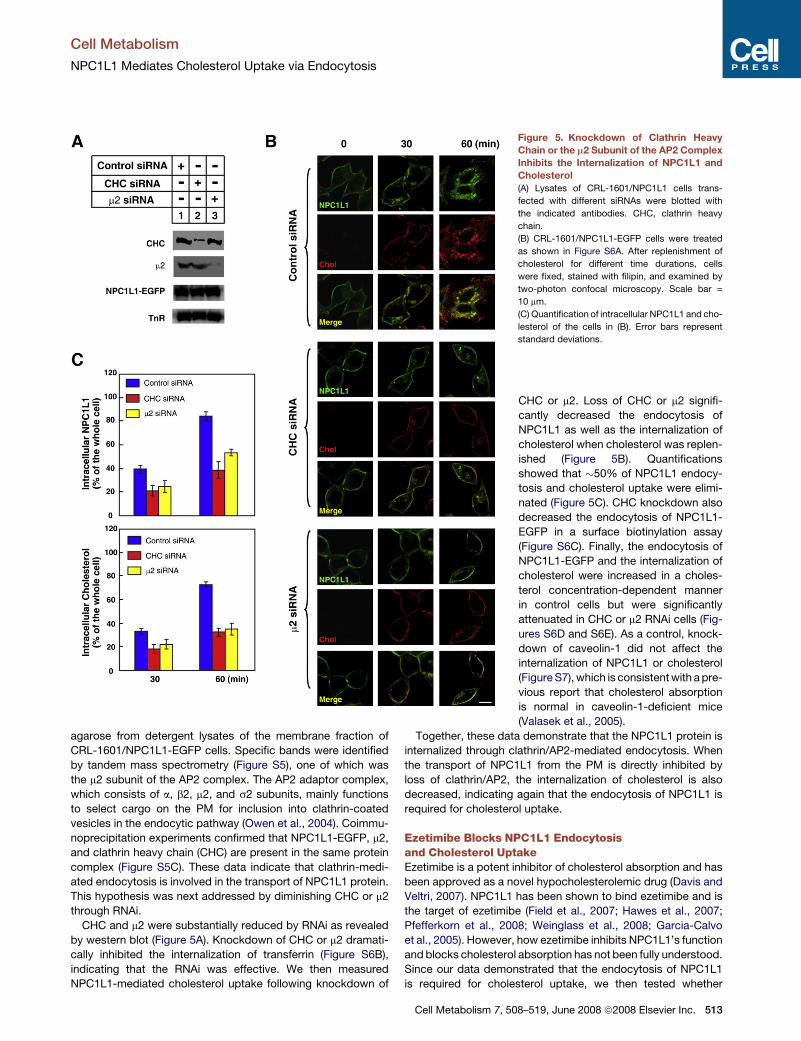
CHC and m2 were substantially reduced by RNAi as revealed
by western blot (Figure 5A). Knockdown of CHC or m2 dramati-
cally inhibited the internalization of transferrin (Figure S6B),
indicating that the RNAi was effective. We then measured
NPC1L1-mediated cholesterol uptake following knockdown of
Figure 5. Knockdown of Clathrin Heavy
Chain or the m2 Subunit of the AP2 Complex
Inhibits the Internalization of NPC1L1 and
Cholesterol
(A) Lysates of CRL-1601/NPC1L1 cells trans-
fected with different siRNAs were blotted with
the indicated antibodies. CHC, clathrin heavy
chain.
(B) CRL-1601/NPC1L1-EGFP cells were treated
as shown in Figure S6A. After replenishment of
cholesterol for different time durations, cells
were fixed, stained with filipin, and examined by
two-photon confocal microscopy. Scale bar =
10 mm.
(C) Quantification of intracellular NPC1L1 and cho-
lesterol of the cells in (B). Error bars represent
standard deviations.
CHC or m2. Loss of CHC or m2 signifi-
cantly decreased the endocytosis of
NPC1L1 as well as the internalization of
cholesterol when cholesterol was replen-
ished (Figure 5B). Quantifications
showed that �50% of NPC1L1 endocy-
tosis and cholesterol uptake were elimi-
nated (Figure 5C). CHC knockdown also
decreased the endocytosis of NPC1L1-
EGFP in a surface biotinylation assay
(Figure S6C). Finally, the endocytosis of
NPC1L1-EGFP and the internalization of
cholesterol were increased in a choles-
terol concentration-dependent manner
in control cells but were significantly
attenuated in CHC or m2 RNAi cells (Fig-
ures S6D and S6E). As a control, knock-
down of caveolin-1 did not affect the
internalization of NPC1L1 or cholesterol
(Figure S7), which is consistent with a pre-
vious report that cholesterol absorption
is normal in caveolin-1-deficient mice
(Valasek et al., 2005).
Together, these data demonstrate that the NPC1L1 protein is
internalized through clathrin/AP2-mediated endocytosis. When
the transport of NPC1L1 from the PM is directly inhibited by
loss of clathrin/AP2, the internalization of cholesterol is also
decreased, indicating again that the endocytosis of NPC1L1 is
required for cholesterol uptake.
Ezetimibe Blocks NPC1L1 Endocytosisand Cholesterol UptakeEzetimibe is a potent inhibitor of cholesterol absorption and has
been approved as a novel hypocholesterolemic drug (Davis and
Veltri, 2007). NPC1L1 has been shown to bind ezetimibe and is
the target of ezetimibe (Field et al., 2007; Hawes et al., 2007;
Pfefferkorn et al., 2008; Weinglass et al., 2008; Garcia-Calvo
et al., 2005). However, how ezetimibe inhibits NPC1L1’s function
and blocks cholesterol absorption has not been fully understood.
Since our data demonstrated that the endocytosis of NPC1L1
is required for cholesterol uptake, we then tested whether
Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc. 513
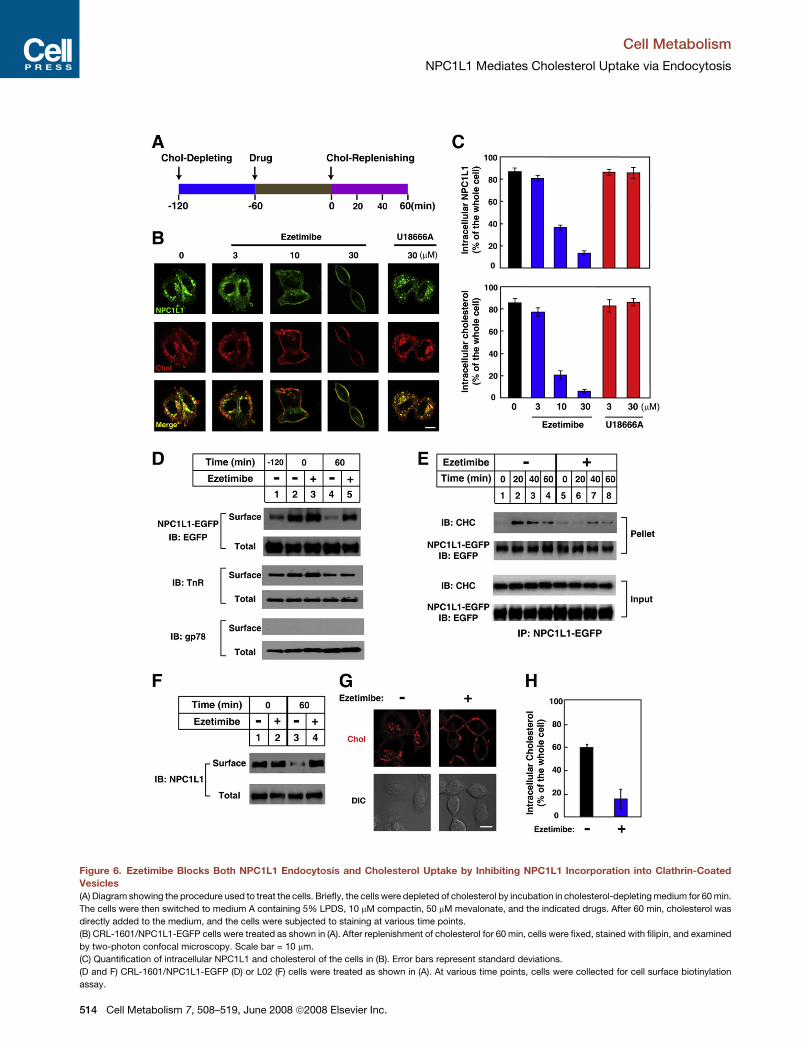
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
Figure 6. Ezetimibe Blocks Both NPC1L1 Endocytosis and Cholesterol Uptake by Inhibiting NPC1L1 Incorporation into Clathrin-Coated
Vesicles
(A) Diagram showing the procedure used to treat the cells. Briefly, the cells were depleted of cholesterol by incubation in cholesterol-depleting medium for 60 min.
The cells were then switched to medium A containing 5% LPDS, 10 mM compactin, 50 mM mevalonate, and the indicated drugs. After 60 min, cholesterol was
directly added to the medium, and the cells were subjected to staining at various time points.
(B) CRL-1601/NPC1L1-EGFP cells were treated as shown in (A). After replenishment of cholesterol for 60 min, cells were fixed, stained with filipin, and examined
by two-photon confocal microscopy. Scale bar = 10 mm.
(C) Quantification of intracellular NPC1L1 and cholesterol of the cells in (B). Error bars represent standard deviations.
(D and F) CRL-1601/NPC1L1-EGFP (D) or L02 (F) cells were treated as shown in (A). At various time points, cells were collected for cell surface biotinylation
assay.
514 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.

Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
ezetimibe inhibits cholesterol uptake by blocking the endocyto-
sis of NPC1L1. Cholesterol-depleted cells were treated with
various concentrations of ezetimibe followed by cholesterol re-
plenishment. Notably, ezetimibe inhibited the endocytosis of
NPC1L1-EGFP in a concentration-dependent manner (Figures
6B and 6C). The internalization of cholesterol was blocked as
well (Figures 6B and 6C). As a control, U18666A, a compound
that interferes with LDL-derived cholesterol transport from endo-
somes/lysosomes to the ER likely by inhibiting NPC1 (Neufeld
et al., 1999), had no effect on NPC1L1-mediated cholesterol
absorption. The results of a cell surface biotinylation assay
confirmed that ezetimibe inhibited the endocytosis of NPC1L1-
EGFP but not that of the transferrin receptor (Figure 6D, compare
lanes 4 and 5), suggesting that the effect of ezetimibe on
NPC1L1 transport is specific.
We showed earlier that the CHC protein can be coimmunopre-
cipitated together with NPC1L1-EGFP protein when cholesterol
is replenished (Figure S5C). We next performed a time-course
study using the coimmunoprecipitation assay and found that
cholesterol promoted the NPC1L1-clathrin complex formation
(Figure 6E). The peak of the NPC1L1-clathrin interaction was
observed after 20 min of cholesterol replenishment, and the in-
teraction then decreased (Figure 6E). This phenomenon can be
explained by the fact that CHC is only involved in the early stage
of vesicular endocytosis and dissociates from cargo vesicles
before the vesicles can fuse with the distal endocytic compart-
ment. However, this cholesterol-induced NPC1L1-CHC com-
plex was dramatically reduced in the presence of ezetimibe
(Figure 6E). Similarly, ezetimibe inhibited the endocytosis of
endogenous NPC1L1 as well as the internalization of cholesterol
in human L02 hepatocytes (Figures 6F and 6H). Similar effects
were observed in HuH7 cells (data not shown). On the other
hand, ezetimibe did not show any significant effect on the exocy-
tosis of NPC1L1, the efflux of cholesterol (Figure S8), or the
uptake of fluorescently labeled LDL (DiI-LDL) (Figure S9).
In summary, these data indicate that ezetimibe specifically
blocks the incorporation of NPC1L1 into clathrin-coated vesi-
cles, thereby inhibiting cholesterol absorption.
DISCUSSION
In this study, we observed cholesterol-regulated NPC1L1 shut-
tling: depletion of cholesterol causes the transport of NPC1L1
from ERC to PM, whereas replenishment of cholesterol results
in the internalization of NPC1L1 (Figure 1). Furthermore, the
endocytosis of NPC1L1 requires microfilaments and the cla-
thrin/AP2 complex (Figure 4; Figure 5; Figure S6). More impor-
tantly, we propose that NPC1L1 mediates cellular cholesterol
uptake through vesicular endocytosis, which is supported by
five lines of evidence:
(1) NPC1L1 is required for free cholesterol uptake (Figure 2;
Figure S2).
(2) Replenishment of cholesterol, but not plant sterols,
promotes the endocytosis of NPC1L1; meanwhile, cho-
lesterol is internalized more efficiently than plant sterols
(Figure 3).
(3) Disruption of microfilaments blocks the endocytosis of
NPC1L1 as well as the uptake of cholesterol (Figure 4).
(4) siRNA-mediated knockdown of the clathrin/AP2 complex
attenuates the endocytosis of NPC1L1 and decreases
cholesterol uptake (Figure 5; Figure S6).
(5) A cholesterol absorption inhibitor, ezetimibe, blocks
NPC1L1 endocytosis and cholesterol internalization
(Figure 6).
Our data show that ezetimibe inhibits cholesterol internaliza-
tion by blocking the endocytosis of NPC1L1 (Figure 6). Ezetimibe
does so through preventing the association between NPC1L1
and clathrin/AP2, which is stimulated by cholesterol
(Figure 6E). Interestingly, ezetimibe does not impede the inter-
nalization of the transferrin receptor or LDL receptor. These
results are consistent with the previous finding that ezetimibe
binds to NPC1L1 and is a highly selective inhibitor of cholesterol
absorption (Garcia-Calvo et al., 2005; Davis et al., 2004). How
ezetimibe inhibits NPC1L1-clathrin complex formation is still
unknown. To answer this question, we need to understand the
endocytosis process of NPC1L1 and how cholesterol initiates
this process. An interesting feature revealed by the sequence
of the NPC1L1 protein is the presence of a sterol-sensing do-
main (SSD). The human genome encodes six SSD-containing
proteins: HMGCR, SCAP, NPC1, Patched, TRC8, and NPC1L1
(Kuwabara and Labouesse, 2002). It has been demonstrated
that the SSDs of SCAP and NPC1 can directly bind cholesterol
(Radhakrishnan et al., 2004; Ohgami et al., 2004). Moreover,
the binding of cholesterol causes a conformational change in
SCAP that leads to the association of SCAP with an endoplasmic
reticulum protein called Insig (Brown et al., 2002; Goldstein et al.,
2006). Conceivably, it is likely that cholesterol directly binds the
SSD of NPC1L1, causes the conformational change of NPC1L1,
and promotes the interaction between NPC1L1 and the clathrin/
AP2 complex. If this is the case, ezetimibe may compete with
cholesterol for binding NPC1L1. Another possibility is that the in-
teraction between ezetimibe and NPC1L1 may inhibit the choles-
terol-induced conformational change of the NPC1L1 protein. It is
also possible that ezetimibe interferes with the local distribution
of cholesterol in the plasma membrane, which impedes choles-
terol binding to NPC1L1. Further experiments will be required to
test these possibilities.
Humans obtain cholesterol from de novo synthesis as well as
through dietary and biliary cholesterol absorption. Compared
with the abundantly available inhibitors of cholesterol
(E) CRL-1601/NPC1L1-EGFP cells were treated as shown in (A). After replenishment with cholesterol for 0, 20, 40, and 60 min, cells were collected and sub-
jected to coimmunoprecipitation with anti-EGFP antibodies. The immunoprecipitated proteins were separated by SDS-PAGE and analyzed by western blot
with the indicated antibodies.
(G) L02 cells were first treated with cholesterol-depleting medium in the absence or presence of 30 mM ezetimibe for 60 min. After being switched to cho-
lesterol-replenishing medium for an additional 60 min, cells were washed, fixed, stained with filipin, and examined by two-photon confocal microscopy. Scale
bar = 10 mm.
(H) Quantification of intracellular cholesterol of the cells in (G). Error bars represent standard deviations.
Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc. 515
Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
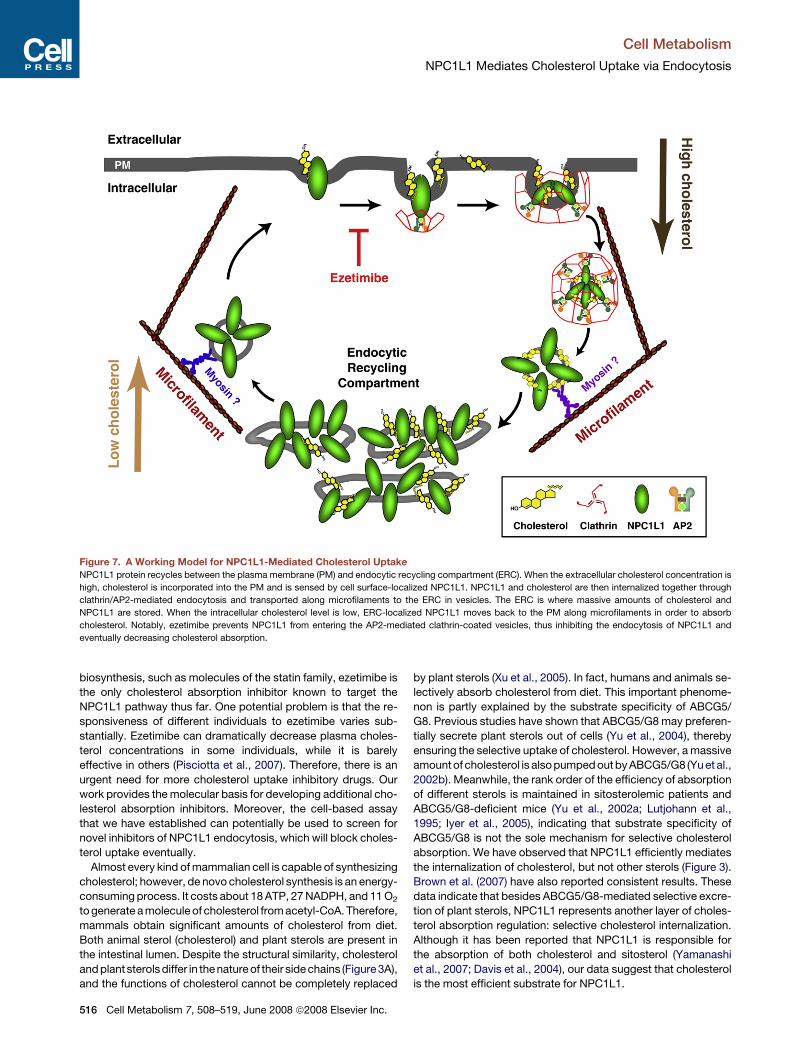
Figure 7. A Working Model for NPC1L1-Mediated Cholesterol Uptake
NPC1L1 protein recycles between the plasma membrane (PM) and endocytic recycling compartment (ERC). When the extracellular cholesterol concentration is
high, cholesterol is incorporated into the PM and is sensed by cell surface-localized NPC1L1. NPC1L1 and cholesterol are then internalized together through
clathrin/AP2-mediated endocytosis and transported along microfilaments to the ERC in vesicles. The ERC is where massive amounts of cholesterol and
NPC1L1 are stored. When the intracellular cholesterol level is low, ERC-localized NPC1L1 moves back to the PM along microfilaments in order to absorb
cholesterol. Notably, ezetimibe prevents NPC1L1 from entering the AP2-mediated clathrin-coated vesicles, thus inhibiting the endocytosis of NPC1L1 and
eventually decreasing cholesterol absorption.
biosynthesis, such as molecules of the statin family, ezetimibe is
the only cholesterol absorption inhibitor known to target the
NPC1L1 pathway thus far. One potential problem is that the re-
sponsiveness of different individuals to ezetimibe varies sub-
stantially. Ezetimibe can dramatically decrease plasma choles-
terol concentrations in some individuals, while it is barely
effective in others (Pisciotta et al., 2007). Therefore, there is an
urgent need for more cholesterol uptake inhibitory drugs. Our
work provides the molecular basis for developing additional cho-
lesterol absorption inhibitors. Moreover, the cell-based assay
that we have established can potentially be used to screen for
novel inhibitors of NPC1L1 endocytosis, which will block choles-
terol uptake eventually.
Almost every kind of mammalian cell is capable of synthesizing
cholesterol; however, de novo cholesterol synthesis is an energy-
consuming process. It costs about 18 ATP, 27 NADPH, and 11 O2
to generate a molecule of cholesterol from acetyl-CoA. Therefore,
mammals obtain significant amounts of cholesterol from diet.
Both animal sterol (cholesterol) and plant sterols are present in
the intestinal lumen. Despite the structural similarity, cholesterol
and plant sterols differ in the nature of their side chains (Figure 3A),
and the functions of cholesterol cannot be completely replaced
516 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.
by plant sterols (Xu et al., 2005). In fact, humans and animals se-
lectively absorb cholesterol from diet. This important phenome-
non is partly explained by the substrate specificity of ABCG5/
G8. Previous studies have shown that ABCG5/G8 may preferen-
tially secrete plant sterols out of cells (Yu et al., 2004), thereby
ensuring the selective uptake of cholesterol. However, a massive
amount of cholesterol is also pumped out by ABCG5/G8 (Yu et al.,
2002b). Meanwhile, the rank order of the efficiency of absorption
of different sterols is maintained in sitosterolemic patients and
ABCG5/G8-deficient mice (Yu et al., 2002a; Lutjohann et al.,
1995; Iyer et al., 2005), indicating that substrate specificity of
ABCG5/G8 is not the sole mechanism for selective cholesterol
absorption. We have observed that NPC1L1 efficiently mediates
the internalization of cholesterol, but not other sterols (Figure 3).
Brown et al. (2007) have also reported consistent results. These
data indicate that besides ABCG5/G8-mediated selective excre-
tion of plant sterols, NPC1L1 represents another layer of choles-
terol absorption regulation: selective cholesterol internalization.
Although it has been reported that NPC1L1 is responsible for
the absorption of both cholesterol and sitosterol (Yamanashi
et al., 2007; Davis et al., 2004), our data suggest that cholesterol
is the most efficient substrate for NPC1L1.

Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
According to our results, we propose a model illustrating the
pathway of dietary or biliary cholesterol absorption, depicted in
Figure 7. After being incorporated into the PM of enterocytes
or hepatocytes, free cholesterol is then sensed by PM-localized
NPC1L1, and they are internalized together through clathrin/
AP2-mediated endocytosis. These NPC1L1-positive and cho-
lesterol-rich vesicles move along microfilaments to the ERC,
where massive amounts of cholesterol and NPC1L1 are stored.
When intracellular cholesterol levels are low, NPC1L1 moves
back to the PM along the microfilaments in order to absorb
cholesterol. Notably, ezetimibe prevents NPC1L1 from entering
the AP2-mediated clathrin-coated vesicles, thus inhibiting the
endocytosis of NPC1L1 and eventually decreasing cholesterol
absorption.
EXPERIMENTAL PROCEDURES
A complete description of primary antibodies and other methods are available
in the Supplemental Experimental Procedures.
Materials and Plasmids
We obtained horseradish peroxidase-conjugated donkey anti-mouse and
anti-rabbit IgG from Jackson ImmunoResearch Laboratories; filipin, nocoda-
zole, and cytochalasin D from Sigma; Alexa Fluor 555 donkey anti-rabbit IgG
and iron-loaded transferrin-rhodamine conjugate from Invitrogen; sulfosucci-
nimidyl 6-(biotinamido) hexanoate and NeutrAvidin agarose from Pierce;
methyl-b-cyclodextrin (CDX) from Cyclodextrin Technologies Development,
Inc.; all sterols from Steraloids, Inc.; and other reagents from previously
described sources (Cao et al., 2007). The purity of cholesterol was above
99% as measured by gas chromatography/mass spectrometry (GC/MS) in
the F.-J. Nan laboratory (Shanghai Institute of Materia Medica, Shanghai,
China). Lipoprotein-deficient serum (LPDS, density > 1.215 g/ml) was pre-
pared from newborn calf serum by ultracentrifugation (Goldstein et al., 1983).
The coding region of human NPC1L1 was amplified from human liver cDNA
by standard PCR and cloned into pEGFP-N1 vector. The resulting plasmid
encodes full-length human NPC1L1 fused with EGFP at the COOH terminus.
The coding region of Rab11a was amplified from HEK293 cell cDNA and
cloned into pDsRed-monomer-C1 vector. The resulting construct encodes
DsRed-Rab11a.
Cell Culture
CRL-1601 (a McArdle RH7777 rat hepatoma cell line), L02 (a human liver cell
line), and HuH7 (a human hepatoma cell line) cells were grown in monolayer
at 37�C in 5% CO2. The cells were maintained in medium A (Dulbecco’s
modified Eagle’s medium containing 100 units/ml penicillin and 100 mg/ml
streptomycin sulfate) supplemented with 10% FBS. Cholesterol-depleting
medium was medium A supplemented with 5% LPDS, 10 mM compactin,
50 mM mevalonate, and 1.5% CDX. Cholesterol-replenishing medium con-
tained medium A supplemented with 5% LPDS, 10 mM compactin, 50 mM
mevalonate, and different concentrations of cholesterol/CDX. The sterol/
CDX inclusion complexes were prepared as described previously (Brown
et al., 2002).
Cell Surface Biotinylation Assay
After being washed twice with ice-cold PBS and treated with 1mg/ml sulfosuc-
cinimidyl 6-(biotinamido) hexanoate in PBS for 40 min at 4�C, cells were
washed with ice-cold buffer A (20 mM Tris-HCl [pH 8.0] and 150 mM NaCl)
and incubated in buffer A for 15 min at room temperature. Cells were then col-
lected by scraping and lysed in buffer B (10 mM Tris-HCl [pH 8.0], 150 mM
NaCl, and 1% NP-40). Each sample of cell lysates was incubated with
100 ml 50% (v/v) NeutrAvidin agarose and rotated overnight at 4�C. The aga-
rose beads were then washed three times with buffer B. Biotinylated proteins
binding to the beads were eluted by incubation with SDS-PAGE sample buffer
at 37�C for 30 min and analyzed by SDS-PAGE followed by western blot.
RNA Interference
Duplexes of siRNA were synthesized by Genepharma (Shanghai, China). The
siRNA targeting rat clathrin heavy chain (CHC) was described previously
(Radulescu et al., 2007). The sequences of siRNAs targeting rat AP2 m2 and
caveolin-1 were 50-AAGGCATGAAGGAATCACAGA-30 and 50-AAGCAAGTG
TACGACGCGCAC-30, respectively. The siRNA against vesicular stomatitis
virus G protein (VSV-G), used as a control, was described previously (Song
et al., 2005). Transfection of siRNAs was carried out as described previously
(Sever et al., 2003).
Immunofluorescence Microscopy
Cells were fixed with 4% paraformaldehyde in PBS and permeabilized with
0.2% Triton X-100 in PBS. Cells were then blocked with 1% BSA in PBS
and labeled with primary antibodies and fluorescent secondary antibodies
as described in the figure legends. Confocal immunofluorescence microscopy
was performed using a Leica TCS SP2 laser scanning microscope. In each
experiment, images of the same channel were acquired at identical laser
output, gain, and offset.
Filipin Staining
A fresh 5 mg/ml stock solution of filipin was prepared in ethanol. After fixation
as described earlier, cells were washed twice with PBS and stained with 50 mg/
ml filipin in the dark for 30 min at room temperature followed by three washes
with PBS. Filipin signals of stained cells were analyzed with a Zeiss LSM 510
confocal microscope equipped with a two-photon laser using an excitatory
wavelength of 720 nm. Red pseudocolor was assigned to show the filipin sig-
nal. In each experiment, images were acquired at identical laser output, gain,
and offset.
Fluorescence Quantification
For quantification of intracellular NPC1L1 or cholesterol abundance, two
circles, one outlining the whole cell and the other beneath the plasma mem-
brane, were drawn manually. After subtraction of the background, the fluores-
cence intensities in each circle were measured by Image-Pro Plus 5.02 and
regarded as whole-cell and intracellular fluorescence intensity, respectively.
The intensity of each cell was arbitrarily defined as 1, against which the intra-
cellular intensity was normalized. For each time point, 50 cells were randomly
selected and calculated. The data shown in the figures are representative of
four or more independent experiments.
For quantification of total cellular cholesterol, a similar method was em-
ployed except that the intracellular circles were omitted. The total cholesterol
of control cells at time point �60 min was defined as 1, against which the total
cellular cholesterol at other time points was normalized.
For quantification of cells with NPC1L1-EGFP PM localization, �100 cells in
each treatment were randomly selected and counted. The results are pre-
sented as the percentage of cells with NPC1L1-EGFP PM localization. Three
independent experiments were performed, and one representative is pre-
sented in the figures.
SUPPLEMENTAL DATA
Supplemental Data include Supplemental Experimental Procedures, Supple-
mental References, and nine figures and can be found with this article online
at http://www.cellmetabolism.org/cgi/content/full/7/6/508/DC1/.
ACKNOWLEDGMENTS
B.-L. Song is deeply grateful to R.A. DeBose-Boyd, J. Goldstein, and M. Brown
(University of Texas Southwestern Medical Center at Dallas) for training. We
thank S.-Z. Pan and Q. Li for their excellent technical assistance, K. Liao
and G. Wang for providing materials, F.-J. Nan for GC/MS analysis, and
D.-S. Li for critical reading of the manuscript. This work was supported by grants
from the Ministry of Science and Technology of China (2006CB910600), the
National Natural Science Foundation of China (30670432 and 90713025), the
Chinese Academy of Sciences (KSCX2-YW-R-109), and the Shanghai Science
and Technology Committee (06QA14057).
Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc. 517

Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
Received: December 16, 2007
Revised: March 6, 2008
Accepted: April 1, 2008
Published: June 3, 2008
REFERENCES
Altmann, S.W., Davis, H.R., Jr., Zhu, L.J., Yao, X., Hoos, L.M., Tetzloff, G., Iyer,
S.P., Maguire, M., Golovko, A., Zeng, M., et al. (2004). Niemann-Pick C1 Like 1
protein is critical for intestinal cholesterol absorption. Science 303,
1201–1204.
Apodaca, G. (2001). Endocytic traffic in polarized epithelial cells: role of the
actin and microtubule cytoskeleton. Traffic 2, 149–159.
Brown, A.J., Sun, L., Feramisco, J.D., Brown, M.S., and Goldstein, J.L. (2002).
Cholesterol addition to ER membranes alters conformation of SCAP, the
SREBP escort protein that regulates cholesterol metabolism. Mol. Cell 10,
237–245.
Brown, J.M., Rudel, L.L., and Yu, L. (2007). NPC1L1 (Niemann-Pick C1-like 1)
mediates sterol-specific unidirectional transport of non-esterified cholesterol
in McArdle-RH7777 hepatoma cells. Biochem. J. 406, 273–283.
Brown, M.S., and Goldstein, J.L. (1986). A receptor-mediated pathway for
cholesterol homeostasis. Science 232, 34–47.
Cao, J., Wang, J., Qi, W., Miao, H.H., Wang, J., Ge, L., Bose-Boyd, R.A., Tang,
J.J., Li, B.L., and Song, B.L. (2007). Ufd1 is a cofactor of gp78 and plays a key
role in cholesterol metabolism by regulating the stability of HMG-CoA reduc-
tase. Cell Metab. 6, 115–128.
Davies, J.P., Levy, B., and Ioannou, Y.A. (2000). Evidence for a Niemann-pick
C (NPC) gene family: identification and characterization of NPC1L1. Genomics
65, 137–145.
Davies, J.P., Scott, C., Oishi, K., Liapis, A., and Ioannou, Y.A. (2005). Inactiva-
tion of NPC1L1 causes multiple lipid transport defects and protects against
diet-induced hypercholesterolemia. J. Biol. Chem. 280, 12710–12720.
Davis, H.R., and Veltri, E.P. (2007). Zetia: inhibition of Niemann-Pick C1 Like 1
(NPC1L1) to reduce intestinal cholesterol absorption and treat hyperlipidemia.
J. Atheroscler. Thromb. 14, 99–108.
Davis, H.R., Jr., Zhu, L.J., Hoos, L.M., Tetzloff, G., Maguire, M., Liu, J., Yao, X.,
Iyer, S.P., Lam, M.H., Lund, E.G., et al. (2004). Niemann-Pick C1 Like 1
(NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key
modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 279,
33586–33592.
Dubinsky, W.P., Mayorga-Wark, O., and Schultz, S.G. (1999). Volume regula-
tory responses of basolateral membrane vesicles from Necturus enterocytes:
role of the cytoskeleton. Proc. Natl. Acad. Sci. USA 96, 9421–9426.
Field, F.J., Watt, K., and Mathur, S.N. (2007). Ezetimibe interferes with choles-
terol trafficking from the plasma membrane to the endoplasmic reticulum in
CaCo-2 cells. J. Lipid Res. 48, 1735–1745.
Garcia-Calvo, M., Lisnock, J., Bull, H.G., Hawes, B.E., Burnett, D.A., Braun,
M.P., Crona, J.H., Davis, H.R., Jr., Dean, D.C., Detmers, P.A., et al. (2005).
The target of ezetimibe is Niemann-Pick C1-Like 1 (NPC1L1). Proc. Natl.
Acad. Sci. USA 102, 8132–8137.
Glover, J., and Morton, R.A. (1958). The absorption and metabolism of sterols.
Br. Med. Bull. 14, 226–233.
Goldstein, J.L., Basu, S.K., and Brown, M.S. (1983). Receptor-mediated
endocytosis of low-density lipoprotein in cultured cells. Methods Enzymol.
98, 241–260.
Goldstein, J.L., Bose-Boyd, R.A., and Brown, M.S. (2006). Protein sensors for
membrane sterols. Cell 124, 35–46.
Hawes, B.E., O’neill, K.A., Yao, X., Crona, J.H., Davis, H.R., Jr., Graziano,
M.P., and Altmann, S.W. (2007). In vivo responsiveness to ezetimibe correlates
with niemann-pick C1 like-1 (NPC1L1) binding affinity: Comparison of multiple
species NPC1L1 orthologs. Mol. Pharmacol. 71, 19–29.
Iyer, S.P., Yao, X., Crona, J.H., Hoos, L.M., Tetzloff, G., Davis, H.R., Jr.,
Graziano, M.P., and Altmann, S.W. (2005). Characterization of the putative
518 Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc.
native and recombinant rat sterol transporter Niemann-Pick C1 Like 1
(NPC1L1) protein. Biochim. Biophys. Acta 1722, 282–292.
Jeon, H., and Blacklow, S.C. (2005). Structure and physiologic function of the
low-density lipoprotein receptor. Annu. Rev. Biochem. 74, 535–562.
Kuwabara, P.E., and Labouesse, M. (2002). The sterol-sensing domain:
multiple families, a unique role? Trends Genet. 18, 193–201.
Lee, J.C., Field, D.J., and Lee, L.L. (1980). Effects of nocodazole on structures
of calf brain tubulin. Biochemistry 19, 6209–6215.
Lutjohann, D., Bjorkhem, I., Beil, U.F., and von Bergmann, K. (1995). Sterol
absorption and sterol balance in phytosterolemia evaluated by deuterium-
labeled sterols: effect of sitostanol treatment. J. Lipid Res. 36, 1763–1773.
Neufeld, E.B., Wastney, M., Patel, S., Suresh, S., Cooney, A.M., Dwyer, N.K.,
Roff, C.F., Ohno, K., Morris, J.A., Carstea, E.D., et al. (1999). The Niemann-
Pick C1 protein resides in a vesicular compartment linked to retrograde
transport of multiple lysosomal cargo. J. Biol. Chem. 274, 9627–9635.
Ohgami, N., Ko, D.C., Thomas, M., Scott, M.P., Chang, C.C., and Chang, T.Y.
(2004). Binding between the Niemann-Pick C1 protein and a photoactivatable
cholesterol analog requires a functional sterol-sensing domain. Proc. Natl.
Acad. Sci. USA 101, 12473–12478.
Owen, D.J., Collins, B.M., and Evans, P.R. (2004). Adaptors for clathrin coats:
structure and function. Annu. Rev. Cell Dev. Biol. 20, 153–191.
Pfefferkorn, J.A., Larsen, S.D., Van, H.C., Sorenson, R., Barton, T., Winters, T.,
Auerbach, B., Wu, C., Wolfram, T.J., Cai, H., et al. (2008). Substituted oxazo-
lidinones as novel NPC1L1 ligands for the inhibition of cholesterol absorption.
Bioorg. Med. Chem. Lett. 18, 546–553.
Pisciotta, L., Fasano, T., Bellocchio, A., Bocchi, L., Sallo, R., Fresa, R., Colan-
geli, I., Cantafora, A., Calandra, S., and Bertolini, S. (2007). Effect of ezetimibe
coadministered with statins in genotype-confirmed heterozygous FH patients.
Atherosclerosis 194, e116–e122.
Radhakrishnan, A., Sun, L.P., Kwon, H.J., Brown, M.S., and Goldstein, J.L.
(2004). Direct binding of cholesterol to the purified membrane region of
SCAP: mechanism for a sterol-sensing domain. Mol. Cell 15, 259–268.
Radulescu, A.E., Siddhanta, A., and Shields, D. (2007). A role for clathrin in
reassembly of the Golgi apparatus. Mol. Biol. Cell 18, 94–105.
Sane, A.T., Sinnett, D., Delvin, E., Bendayan, M., Marcil, V., Menard, D.,
Beaulieu, J.F., and Levy, E. (2006). Localization and role of NPC1L1 in choles-
terol absorption in human intestine. J. Lipid Res. 47, 2112–2120.
Schafer, D.A. (2002). Coupling actin dynamics and membrane dynamics
during endocytosis. Curr. Opin. Cell Biol. 14, 76–81.
Schoenheimer, R. (1931). New contributions in sterol metabolism. Science 74,
579–584.
Sever, N., Song, B.L., Yabe, D., Goldstein, J.L., Brown, M.S., and Bose-Boyd,
R.A. (2003). Insig-dependent ubiquitination and degradation of mammalian 3-
hydroxy-3-methylglutaryl-CoA reductase stimulated by sterols and geranyl-
geraniol. J. Biol. Chem. 278, 52479–52490.
Severs, N.J. (1997). Cholesterol cytochemistry in cell biology and disease.
Subcell. Biochem. 28, 477–505.
Song, B.L., Sever, N., and Bose-Boyd, R.A. (2005). Gp78, a membrane-
anchored ubiquitin ligase, associates with Insig-1 and couples sterol-regu-
lated ubiquitination to degradation of HMG CoA reductase. Mol. Cell 19,
829–840.
Temel, R.E., Tang, W., Ma, Y., Rudel, L.L., Willingham, M.C., Ioannou, Y.A.,
Davies, J.P., Nilsson, L.M., and Yu, L. (2007). Hepatic Niemann-Pick C1-like
1 regulates biliary cholesterol concentration and is a target of ezetimibe.
J. Clin. Invest. 117, 1968–1978.
Ullrich, O., Reinsch, S., Urbe, S., Zerial, M., and Parton, R.G. (1996). Rab11
regulates recycling through the pericentriolar recycling endosome. J. Cell
Biol. 135, 913–924.
Valasek, M.A., Weng, J., Shaul, P.W., Anderson, R.G., and Repa, J.J. (2005).
Caveolin-1 is not required for murine intestinal cholesterol transport. J. Biol.
Chem. 280, 28103–28109.
Vossenkamper, A., Nedvetsky, P.I., Wiesner, B., Furkert, J., Rosenthal, W.,
and Klussmann, E. (2007). Microtubules are needed for the perinuclear

Cell Metabolism
NPC1L1 Mediates Cholesterol Uptake via Endocytosis
positioning of aquaporin-2 after its endocytic retrieval in renal principal cells.
Am. J. Physiol. Cell Physiol. 293, C1129–C1138.
Weinglass, A.B., Kohler, M.G., Nketiah, E.O., Liu, J., Schmalhofer, W.,
Thomas, A., Williams, B., Beers, L., Smith, L., Hafey, M., et al. (2008).
Madin-Darby canine kidney II cells: a pharmacologically validated system for
NPC1L1-mediated cholesterol uptake. Mol. Pharmacol. 73, 1072–1084.
Xu, F., Rychnovsky, S.D., Belani, J.D., Hobbs, H.H., Cohen, J.C., and Rawson,
R.B. (2005). Dual roles for cholesterol in mammalian cells. Proc. Natl. Acad.
Sci. USA 102, 14551–14556.
Yamanashi, Y., Takada, T., and Suzuki, H. (2007). Niemann-Pick C1-like 1
overexpression facilitates ezetimibe-sensitive cholesterol and beta-sitosterol
uptake in CaCo-2 cells. J. Pharmacol. Exp. Ther. 320, 559–564.
Yamashiro, D.J., Tycko, B., Fluss, S.R., and Maxfield, F.R. (1984). Segregation
of transferrin to a mildly acidic (pH 6.5) para-Golgi compartment in the
recycling pathway. Cell 37, 789–800.
Yu, L., Hammer, R.E., Li-Hawkins, J., Von, B.K., Lutjohann, D., Cohen, J.C.,
and Hobbs, H.H. (2002a). Disruption of Abcg5 and Abcg8 in mice reveals their
crucial role in biliary cholesterol secretion. Proc. Natl. Acad. Sci. USA 99,
16237–16242.
Yu, L., Li-Hawkins, J., Hammer, R.E., Berge, K.E., Horton, J.D., Cohen, J.C.,
and Hobbs, H.H. (2002b). Overexpression of ABCG5 and ABCG8 promotes
biliary cholesterol secretion and reduces fractional absorption of dietary
cholesterol. J. Clin. Invest. 110, 671–680.
Yu, L., von Bergmann, K., Lutjohann, D., Hobbs, H.H., and Cohen, J.C. (2004).
Selective sterol accumulation in ABCG5/ABCG8-deficient mice. J. Lipid Res.
45, 301–307.
Yu, L., Bharadwaj, S., Brown, J.M., Ma, Y., Du, W., Davis, M.A., Michaely, P.,
Liu, P., Willingham, M.C., and Rudel, L.L. (2006). Cholesterol-regulated
translocation of NPC1L1 to the cell surface facilitates free cholesterol uptake.
J. Biol. Chem. 281, 6616–6624.
Cell Metabolism 7, 508–519, June 2008 ª2008 Elsevier Inc. 519




















