
The Cotton Centromere Contains a Ty3-gypsy-like LTR Retroelement
Upload
independentCategory
view
1download
0
The Adult Pituitary Contains a Cell PopulationDisplaying Stem/Progenitor Cell and EarlyEmbryonic Characteristics
Jianghai Chen, Nicole Hersmus, Vik Van Duppen, Pieter Caesens, Carl Denef, and Hugo Vankelecom
Laboratory of Cell Pharmacology, Department of Molecular Cell Biology (J.C., N.H., P.C., C.D., H.V.), and Division ofHematology, Department of Pathophysiology (V.V.D.), University of Leuven (K.U.Leuven) School of Medicine, B-3000Leuven, Belgium
A side population (SP) has been identified in a number oftissues, where it typically represents a small population en-riched in stem/progenitor cells. In this study we show that theadult mouse anterior pituitary (AP) also contains a charac-teristic SP displaying verapamil-sensitive Hoechst dye effluxcapacity. A majority of the SP cells express stem cell antigen1 at a high level (Sca1high). Using (semi)quantitative RT-PCRand immunofluorescence, we characterized the Sca1high SP asa population enriched in cells expressing stem/progenitorcell-associated factors and components of the Notch, Wnt, andsonic hedgehog signaling pathways, functional in stem cellhomeostasis as well as in early pituitary embryogenesis. Lhx4,a transcription factor pivotal for early embryonic develop-ment of the AP, was only detected in the Sca1high SP, whereasLhx3, in contrast to Lhx4 not down-regulated after AP devel-opment, was only found in the main population. The Sca1high
SP was depleted from cells expressing phenotypic markers ofdifferentiated AP cells (hormones), but contained a small pro-portion of folliculo-stellate cells. Stem cells of many tissuescan clonally expand to nonadherent spheres in culture. Clonalspheres also developed in AP cell cultures. Spheres showed anexpression pattern resembling that of Sca1high SP cells. More-over, the sphere-initiating cells of the pituitary segregated tothe SP and not to the main population. In conclusion, we showthat the adult pituitary contains a hitherto undescribed pop-ulation of cells with SP phenotype and clonal expansion ca-pacity. These cells express (signaling) molecules generallyfound in stem/progenitor cells and/or operative during pitu-itary early embryonic development. These characteristics aresupportive of a stem/progenitor cell phenotype. (Endocrinol-ogy 146: 3985–3998, 2005)
THE ANTERIOR PITUITARY (AP) is an endocrine organthat dynamically adjusts its hormone output to chang-
ing physiological and pathological conditions. Some of theseadaptations are associated with rapid expansion of specificAP cell populations. It is unknown whether stem/progenitorcells play a role in this expansion. On the other hand, thecellular constitution of the mature gland appears stable un-der basal conditions, implying steady cell turnover, which isestimated to be approximately 1.5%/d in the young adultmale rat (1). Formation of new hormone-producing cellsinvolves mitosis of differentiated cells, but also, and appar-ently to a greater extent, maturation of undifferentiated cells(2, 3). It has repeatedly been suggested that AP stem cells areinvolved in this process of AP cell turnover (1–3) as well as
in regenerative processes after partial hypophysectomy, al-though the restorative capacity of the AP is not unanimouslyaccepted (4, 5). At present, stem/progenitor cells are beingidentified in a growing number of adult tissues. To date, noclear indications have been provided that the adult AP con-tains cells with stem/progenitor cell characteristics.
The search for tissue stem cells is hampered by the lack ofuniversal stem cell phenotypic markers. Two methods, how-ever, have proved valuable for prospective identification andcharacterization of (adult) stem cells in a number of tissues;one is based on dye efflux capacity, and the other on clonalsphere formation. Cells that efficiently exclude the vital dyeHoechst 33342 have originally been identified in bone mar-row and are visualized as a small streak of side population(SP) cells in dual-color flow cytometry (6). Bone marrow SPcells were found to be enriched in the most primitive, long-term repopulating hemopoietic stem cells (HSC). Mean-while, cells with the SP phenotype and endowed with stem/progenitor cell qualities have been documented in a numberof other tissues as well, such as brain, muscle, mammarygland, and testis (7–11). The identification and characteriza-tion of neural stem cells (NSC) has gained fundamental im-petus by the creation of a floating sphere (neurosphere) cul-ture technique (12, 13). NSC brought in suspension culturewith basic fibroblast growth factor (bFGF) and/or epidermalgrowth factor (EGF) divide to form a clonal sphere of cellswith self-renewal capacity. A comparable floating sphereculture format has, meanwhile, been adopted to isolate and
First Published Online June 2, 2005Abbreviations: AMCA, Amino-methylcoumarin acetic acid; AP, an-
terior pituitary; bFGF, basic fibroblast growth factor; BrdU, bromode-oxyuridine; DAPI, 4�,6-diamidino-2-phenylindole; E, embryonic day;EGF, epidermal growth factor; eGFP, enhanced green fluorescent pro-tein; ES, embryonic stem; FACS, fluorescence-activated cell sorter; FS,folliculo-stellate; Fzd, frizzled; �GSU, glycoprotein hormone �-subunit;Hes1, hairy and enhancer of split 1; HSC, hemopoietic stem cell; M-MuLV, Moloney murine leukemia virus; MP, main population; NSC,neural stem cell; PE, phycoerythrin; PI, propidium iodide; POMC, pro-opiomelanocortin; PRL, prolactin; Ptch1, Patched 1; RNase, ribonucle-ase; Sca1, stem cell antigen 1; Shh, sonic hedgehog; SP, side population;WT, wild type.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/05/$15.00/0 Endocrinology 146(9):3985–3998Printed in U.S.A. Copyright © 2005 by The Endocrine Society
doi: 10.1210/en.2005-0185
3985

characterize potential stem/progenitor cells from a numberof other tissues (retina, skin, mammary gland, heart, andpancreas) (14–18).
Despite the lack of universal stem cell markers, certaincellular molecules have been found in at least part of the stemcells isolated, such as stem cell antigen 1 (Sca1), c-Kit(CD117), CD34, prominin-1 (CD133/AC133), Oct-4 (alsocalled Oct-3, NF-A3, or Pou5f1), Nanog, nestin, and Bmi-1 (7,8, 11, 14, 15, 17, 19–27). These factors can be consideredprospective identifiers of candidate stem/progenitor cells. Inaddition to these cellular markers, the signaling pathways ofNotch, Wnt, and sonic hedgehog (Shh) have recently beenimplicated in stem cell renewal and fate determination in avariety of tissues (brain, intestine, bone marrow, mammarygland) (28–36).
Cues for adult stem cell characterization may also be gath-ered from embryonic development (19, 31, 34). The AP de-velops from Rathke’s pouch that starts to invaginate from theoral ectoderm on embryonic d 8.5 (E8.5) in the mouse (37).Morphogenesis and maturation of the gland are driven by anorchestrated action of extrinsic and intrinsic factors. Gradi-ents of these factors leave a locally and temporally preciseimprint of transcription factor expression, instrumental forcell type determination and expansion (37, 38). Among thosesignals, Notch, Wnt, and Shh appear to be essential (37, 39,40). Furthermore, the LIM homeobox transcription factors,Lhx3 (P-Lim) and Lhx4 (Gsh-4), play a crucial role in lineagespecification and expansion of AP cell types during earlyembryonic development (41, 42). Lhx3 expression persistsinto adulthood, whereas the expression of Lhx4 is signifi-cantly down-regulated from E15.5, coinciding with the pe-riod of terminal differentiation of AP cells (41–43).
In the present study we searched for potential stem/pro-genitor cells in the adult AP on the basis of dye efflux andsphere-forming capacity. We identified a population of cellswith SP phenotype that clonally replicate to nonadherentspheres and express candidate stem/progenitor cell mark-ers, signaling molecules known to be involved in stem cellrenewal and fate decision, and pituitary-related embryonicsignals and transcription factors.
Materials and MethodsMaterials
DMEM/F12, B27, N2 supplement, and chemically defined culturemedium (44, 45) were produced by Invitrogen Life Technologies, Inc.(Grand Island, NY), and BSA by Serva (Heidelberg, Germany). Basicfibroblast growth factor (bFGF) and epidermal growth factor (EGF) wereobtained from R&D Systems (Minneapolis, MN). Trypsin (from porcinepancreas) was purchased from ICN Biomedicals (Aurora, OH); Hoechst33342 (bisbenzimide), verapamil, and propidium iodide (PI) from Sig-ma-Aldrich Corp. (St. Louis, MO); and bromodeoxyuridine (BrdU) fromBD Biosciences (San Jose, CA).
All PCR reagents were supplied by Applied Biosystems (AppleraBelgium, Lennik, Belgium). Ribonuclease (RNase) inhibitor (RNasin)was purchased from Promega Corp. (Leiden, The Netherlands), andMoloney murine leukemia virus (M-MuLV) reverse transcriptase,RNase-free deoxyribonuclease I (amplification grade), and oligonucle-otide primers were obtained from Invitrogen Life Technologies, Inc.TriPure RNA isolation reagent was purchased from Roche (Brussels,Belgium), and RiboGreen quantitation reagent from Molecular Probes(Eugene, OR). TaqMan Universal PCR Mix was obtained from AppliedBiosystems; MasterMix Plus for SYBR Green I and TaqMan probes(designed using PrimerXpress, Applied Biosystems) were obtained from
Eurogentec (Seraing, Belgium). Quantitative real-time PCR was run onan ABI PRISM 7700 or ABI PRISM 7000 Sequence Detector (AppliedBiosystems).
The dipeptide �-Ala-Lys-N�-amino-methylcoumarin acetic acid(AMCA) was provided by Dr. K. Bauer (Max Planck Institute, Hannover,Germany). Antibodies used for flow cytometry were R-phycoerythrin(R-PE)-conjugated rat antimouse Sca1, R-PE- and PE-Cy5-conjugated ratantimouse CD45, R-PE- and fluorescein isothiocyanate-conjugated ratantimouse CD117 (c-Kit), and R-PE-conjugated rat antimouse CD34 aswell as the corresponding isotype controls (all from BD Biosciences).Mouse anti-BrdU antibody was obtained from BD Biosciences. Anti-bodies for immunofluorescent analysis of sorted cells included goatanti-Notch1 (Notch1 C-20) from Santa Cruz Biotechnology, Inc. (SantaCruz, CA); rabbit antisera against Lhx3 and Lhx4 (provided by Dr. S.Pfaff, The Salk Institute for Biological Studies, La Jolla, CA); mouseantinestin (Rat-401) from BD Biosciences; rabbit antinestin (provided byDr. R. McKay, National Institutes of Health, National Institute for Neu-rological Disorders and Stroke, Bethesda, MD); and antisera againstpituitary hormones (provided by Dr. A. F. Parlow through the NationalHormone and Pituitary Program, National Institute of Diabetes andDigestive and Kidney Diseases, Torrance, CA). The secondary antibod-ies, Alexa Fluor 555-labeled goat antimouse and Alexa Fluor 488-labeledgoat antirabbit, as well as the nuclear stain, ToPro-3, were purchasedfrom Molecular Probes. Cy3-conjugated donkey antigoat was obtainedfrom Jackson ImmunoResearch Laboratories, Inc. (Soham, UK), and4�,6-diamidino-2-phenylindole (DAPI) was purchased from Sigma-Aldrich Corp. The mounting medium, Vectashield, was provided byVector Laboratories, Inc. (Burlingame, CA).
Animals
FVB mice (8-wk-old female; 3-wk-old male and female) were pur-chased from Elevage Janvier (Schaijk, The Netherlands). Green fluores-cent protein (GFP) transgenic mice that ubiquitously express enhancedGFP (eGFP) were provided by Dr. A. Nagy (Mount Sinai Hospital,Samuel Lunenfeld Research Institute, Toronto, Canada), and back-crossed to the FVB genetic background for more than 20 generations. Allanimals were kept in an environment of constant temperature, humidity,and day-night cycle within the animal house facilities and had free accessto water and food (University of Leuven, Leuven, Belgium). All animalexperiments were conducted in accordance with the Endocrine Society’sethical guideline, and were approved by the University of Leuven ethicalcommittee.
Isolation of anterior pituitary cells
Mice were killed by CO2 asphyxiation, followed by decapitation. ForSP analysis and sorting, animals were first perfused with PBS to rinseblood out of the pituitary. The neurointermediate lobe was carefullydiscarded under the stereomicroscope. Anterior lobes (AP) were dis-sociated into single cells using trypsin (2.5%; 15 min) as previouslydescribed in detail (44). Cells were finally resuspended in serum-free,chemically defined culture medium (45).
Flow cytometric analysis of AP cells after incubation withHoechst 33342 and membrane antigen staining
We adapted the original SP identification method of Goodell et al. (6)for optimal resolution of pituitary SP cells. The main difference was themedium used (chemically defined culture medium instead of DMEM).Cell density was adjusted to 1 � 106 cells/ml. Hoechst 33342 was addedto a final concentration of 2.5 �g/ml together with verapamil (50–100�m) in control samples to verify blockage of dye efflux as a confirmationof the SP phenotype. Samples were incubated for 90 min at 37 C, thenfor 20–30 min on ice with R-PE-labeled anti-Sca1, R-PE- or fluoresceinisothiocyanate-labeled anti-c-Kit, R-PE-labeled anti-CD34, and/or R-PE-or PE-Cy5-labeled anti-CD45 (all at 1 �g/106 cells). Cells were finallyresuspended in ice-cold PBS with fetal calf serum (2%) and PI (2 �g/ml)for fluorescence-activated cell sorter (FACS) analysis and sorting. CD45expression within the Sca1high SP was analyzed by flow cytometry ofHoechst-incubated cells stained with anti-Sca1-R-PE and anti-CD45-PE-Cy5 (analyzed in three independent cell isolations). To control whethertrypsinization affected membrane antigen staining, mouse bone marrow
3986 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary

cells were treated with trypsin in an identical way as AP cells and stainedfor Sca1, CD45, c-Kit, and CD34. FACS analysis revealed no differencesbetween untreated and trypsin-treated bone marrow cells (data notshown).
Flow cytometric analysis was carried out with a FACSVantage (BDBiosciences). The SP was visualized using dual-wavelength flow cy-tometry with UV excitation and detection of blue emission with a BP424/44 filter and of red emission with a BP 630/22 filter, resolving thetotal population in a Hoechstlow SP and a Hoechstmid�high non-SP. Cellswere analyzed and sorted within the living (PI�) cell population.
In some experiments the dipeptide �-Ala-Lys-N�-AMCA (100 �m)was added to the cells during incubation with Hoechst as a marker forfolliculo-stellate (FS) cells (46). Cells were analyzed by FACS for SPphenotype (Hoechst exclusion) and FS cell phenotype (dipeptide up-take) using UV excitation.
Immunofluorescent analysis of SP and main population(MP) cells sorted by FACS
SP and MP cells were sorted by FACS in chemically defined mediumand deposited by cytospin on glass slides. Cells were permeabilized withTriton X-100 (0.4% in PBS), blocked with preimmune sera (10% in PBS),incubated with primary antibodies (rabbit antinestin at 1:2000; mouseanti-nestin at 1:100; anti-Notch1 at 1:100; anti-Lhx3 at 1:1000; anti-Lhx4at 1:500; hormone antisera at the dilutions recommended by Dr. A. F.Parlow) for 2 h, and finally with secondary antibody (Alexa-labeledantisera at 1:1000; Cy3-labeled antibody at 1:400) for 1.5 h. Cells werecovered with Vectashield and analyzed using an epifluorescence mi-croscope (DMRB, Leica, Wetzlar, Germany). Negative controls wereperformed by omitting primary antibodies and substituting the primaryantibodies with mouse nonsense IgG or normal serum at comparabledilutions.
Growth and characterization of nonadherent spheres fromAP cells
Mouse AP cells were prepared as described above and seeded in35-mm nontreated petri dishes at a density of 100,000 cells/ml (2 ml/dish) in either DMEM/Ham’s F-12 or chemically defined culture me-dium, both supplemented with B27 (1:50) and bFGF (20 ng/ml; in someexperiments replaced by EGF at 20 ng/ml). Cells were kept at 37 C ina 1.5% CO2 incubator. From d 7–8 in culture, clumps and cells insuspension started to stick to the spheres to eventually form largeraggregates. Spheres could be recovered from these aggregates by treat-ing them at 37 C for 5 min with trypsin-EDTA (0.05% trypsin in DMEMwith 0.037% EDTA, 0.3% BSA, and anorganic salts).
To explore the potential for self-renewal, primary spheres were dis-persed into single cells, and generation of secondary spheres was ex-amined. Primary spheres were picked from 6-d cultures, incubated withtrypsin-EDTA for 15 min at 37 C, and then mechanically dispersed undervisual microscopic control. Trypsin was inhibited by adding the samevolume of 10% fetal calf serum (diluted in chemically defined medium).Cells were spun down immediately, resuspended in sphere culturemedium, analyzed for single cellularity, and cultured at 10,000 cells/ml(2 ml/dish) for secondary sphere generation.
To study whether pituitary spheres were clonal in origin, spherecultures were initiated as described above from a 1:1 mixture of wild-type (WT) and GFP� AP cells isolated from FVB WT and FVB eGFP-expressing transgenic mice, respectively. At different time points inculture, all floating material in a dish (spheres, aggregates, irregular cellclumps, and single cells in suspension) was fixed with paraformalde-hyde (4% in PBS) for 15 min at room temperature. Samples weremounted in Vectashield for epifluorescent analysis or for examinationusing a confocal laser scanning microscope (LSM 510, Zeiss, Zaventem,Belgium). Nuclear counterstain was obtained with DAPI for epifluo-rescence analysis and with ToPro-3 for confocal analysis.
To examine whether spheres developed by cell division as opposedto cellular aggregation, the thymidine analog BrdU (10 �m) was addedto the cells 1 d after seeding. On d 3, all cell material in suspension wasfixed as described above and further analyzed for BrdU incorporationby immunofluorescent staining. Therefore, cells were permeabilizedwith 0.5% saponin in PBS and treated with deoxyribonuclease I (100U/ml) for 30 min at 37 C for BrdU antigen retrieval. Then the cell pool
was successively incubated with normal goat serum (20%; 10 min),anti-BrdU antibody (1:2.5; overnight), and Alexa Fluor 555 goat anti-mouse (1:1000; 1.5 h). Samples were mounted in Vectashield.
For analysis of protein expression by immunofluorescence, spheresand clumps were treated as described for BrdU immunostaining, withomission of the antigen retrieval step, using the antibody dilutionsmentioned above.
The number of cells per pituitary sphere was counted as ToPro-3-stained nuclei in images of consecutive confocal slices and as DAPI-labeled nuclei in epifluorescence images. Diameters were measured onlive culture pictures using analysis tools provided by ImageJ (freelyavailable at http://rsb.info.nih.gov/ij/).
Gene expression analysis by RT-PCR
RT. Sca1high SP cells and MP cells within the living (PI�) population weresorted by FACS in TriPure with glycogen. A typical yield of Sca1high SPcells from a single sort was between 20,000 and 50,000 cells. In someexperiments, cells outside both the SP and MP gates (non-SP�non-MP)were also sorted (see Fig. 1A for sorting gates). Total RNA was isolatedaccording to the manufacturer’s recommendations and dissolved in 8 �lRNase-free H2O. A comparable amount of RNA was obtained from anidentical number of SP and MP cells. The RNA solution was treated withRNase-free deoxyribonuclease I (1 U), quantified with RiboGreen quan-titation reagent following the manufacturer’s protocol (MolecularProbes), and stored at �80 C until additional analysis by RT-PCR.
Pituitary spheres or irregular groups of cells (clumps) of comparablesize were individually picked from cultures, rinsed in PBS (37 C), andtransferred to PCR tubes into 10.2 �l RNase-free H2O for osmotic lysis.The samples were heated at 65 C for 10 min and stored at �80 C untiladditional analysis by RT-PCR. To control the reliability of the singlesphere or clump lysis method for gene expression analysis, RT-PCR wasalso performed on RNA isolated from an identical number of cellsderived from dissociated spheres or clumps. Spheres and clumps werepicked to be pooled within their respective group and were dispersedinto single cells as described above. RNA was isolated from 10,000 cellsof each (a comparable amount was obtained from this same number ofsphere and clump cells) and checked by semiquantitative RT-PCR asdescribed below.
For RT, 2 �l RNA solution from Sca1high SP, from MP or from non-SP�non-MP cells was added to 18 �l of the following RT reactionmixture: 4 �l MgCl2 (25 mm), 2 �l 10� PCR Gold buffer, 0.8 �l dNTPs(100 mm), 1 �l random hexamers (50 �m), 1 �l M-MuLV reverse tran-scriptase (200 U/�l), 1 �l RNasin (40 U/�l), and 8.2 �l RNase-free H2O.For RT of clump or sphere RNA, 9.8 �l RT mixture (without H2O) wasadded to the lysed cell material. RT was performed using the followingtemperature cycle: 10–15 min at 25 C, 50 min at 42 C, and 10 min at 95C. As a positive control for the RT reaction, 10 ng total RNA, purifiedfrom perfused mouse AP, were added to the RT mixture. As a negativecontrol, RNase-free H2O was used.
PCR. PCR was performed on 1 �l of the RT reaction, added to 9 �lof a PCR mixture containing 0.9 �l MgCl2 (25 mm; except for Oct-4amplification, where 0.75 �l was optimal), 1.1 �l 10� PCR Goldbuffer, 0.1 �l dNTPs (100 mm), 0.15 �l AmpliTaq Gold DNA poly-merase (5 U/�l), 6.55 �l RNase-free H2O, and 0.1 �l sense andantisense oligonucleotide primers (100 �m each). The nucleotide se-quences of the PCR primers are presented in Table 1. Primers weredesigned using Vector NTI (InforMax, Invitrogen Life Technologies,Inc.) to span at least one intron except for frizzled (Fzd), which lacksintrons. To control for amplification of the Fzd gene from residualgenomic DNA due to incomplete DNA digestion, PCR on an RTsample run without M-MuLV was performed. The expression of Fzdreceptors was analyzed using PCR primers in consensus sequencesof all known Fzd receptor subtypes (Fzd1–10; primers were calledFzdall; see Table 1). RT-PCR for L19, a ribosomal protein constitu-tively expressed in all cells, was used as an internal control. NoL19-negative sphere or clump samples were observed. PCR condi-tions were optimized using total RNA from perfused mouse APs, andthe final specific parameters used are also detailed in Table 1. Thereproducibility of the PCR was 80 –100% as tested on 10 spheres andclumps in five independent PCRs. The reproducibility of the PCR forhormones has been determined previously as being at least 80% (47).
Chen et al. • Candidate Stem Cells in Adult Pituitary Endocrinology, September 2005, 146(9):3985–3998 3987

For semiquantitative RT-PCR, the cycle number was varied between35 and 50, as explained below. The identities of the amplified frag-ments were confirmed by sequence analysis (performed by LarkTechnologies, Essex, UK). PCR protocols for hormones have beenoptimized and validated previously in the laboratory (47). In general,PCR cycling consisted of the following temperature steps: 7 min at 95C, followed by 35–50 cycles of denaturation, annealing, and exten-sion, the details of which depend on the specific cDNA to be amplified(described in Table 1). Finally, samples were heated at 72 C for 7 minbefore cooling to 4 C. A positive control sample for PCR performanceconsisting of 10 ng reverse transcribed mouse AP total RNA wasincluded as well as a negative control consisting of 1 �l RNase-freeH2O instead of sample. Spheres and clumps to be compared (from thesame culture) were simultaneously subjected to RT using the same RTmixture and per marker to PCR using the same PCR mixture. Am-plified DNA products were separated by electrophoresis in a 2%agarose gel containing 0.5 �g/ml ethidium bromide.
Semiquantitative RT-PCR. To assess the mRNA expression level semi-quantitatively, a 10-fold dilution series of Sca1high SP, MP, or non-SP�non-MP RNA (from 10 ng/�l to 1 pg/�l) was subjected to RT,and PCR amplification was carried out using different cycle numbers(35, 40, 45, 48). Thus, both template input amounts and amplificationcycles were varied to ensure that the signal taken for comparisonbetween Sca1high SP and MP (and in some experiments non-SP�non-MP) was in the exponential phase of the amplification reaction. Theribosomal protein L19 mRNA was used as the normalization stan-dard, and RNA dilution series of Sca1high SP and MP (and in someexperiments non-SP�non-MP) were first checked on the basis of L19amplification signals. Only exceptionally did the RNA dilution seriesof SP, MP, or non-SP�non-MP have to be adjusted with a maximalfactor of 2. Detection limits (i.e. lowest dilution of RNA showing anamplification signal) in Sca1high SP and MP, obtained under expo-
nential amplification conditions, were used to calculate the SP/MPexpression ratios. An example is given in Fig. 2A for Notch1. It isworth mentioning that comparison of expression between markers isnot meaningful, because RT-PCR sensitivity may vary per marker.For the most reliable comparison between SP and MP (and in someexperiments non-SP�non-MP), RT was carried out simultaneouslyfor the RNA serial dilutions to be compared (i.e. from SP, MP, andnon-SP�non-MP from the same cell sorting) using the same RTmixture, and PCR per marker was performed on these cDNA serialdilutions using the same PCR mixture.
Quantitative real-time RT-PCR. To inspect the reliability of the semiquan-titative method, quantitative real-time RT-PCR was performed for aselection of the markers using the primers shown in Table 1 or otherprimers already published (47). As an additional control, another in-ternal (normalization) standard (18S rRNA instead of L19 mRNA) wasused. 18S rRNA levels were comparable in an identical amount ofSca1high SP and MP total RNA. Real-time RT-PCR for hormones wasperformed using sequence-specific TaqMan probes as described in detailpreviously (47); sequences used for proopiomelanocortin (POMC)mRNA quantitation are given in Table 1. Real-time RT-PCR for glyco-protein hormone �-subunit (�GSU) and markers was carried out in ananalogous way with SYBR Green as quantifying reagent, using Mas-terMix Plus for SYBR Green I instead of TaqMan Universal PCR Mix. Thespecificity of the amplification reactions quantified with SYBR Greenwas verified by melt curve analysis. Each sample was measured intriplicate. Data were analyzed with SDS software (Applied Biosystems).Relative expression values were obtained using the � threshold cycle/standard curve method and were normalized to (relative) 18S rRNAlevels as described previously (47). The normalized expression valueswere used to calculate the SP/MP expression ratios.
FIG. 1. The adult mouse AP contains a SP en-riched in Sca1high-expressing cells. Dual-wavelength FACS analysis reveals the pres-ence of typical SP cells (1.7% of total livingcells) in the adult AP that are Hoechstlow be-cause of efficient Hoechst 33342 efflux capac-ity (A) that can be blocked by verapamil (B).The SP contains cells with high expression ofSca1 (C; only cells gated as SP in A are shown).A large proportion of the SP cells (70%) areSca1high, whereas the majority of the residualnon-SP cells are Sca1neg (D; mean � SEM of 13independent cell isolations are shown).
3988 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary
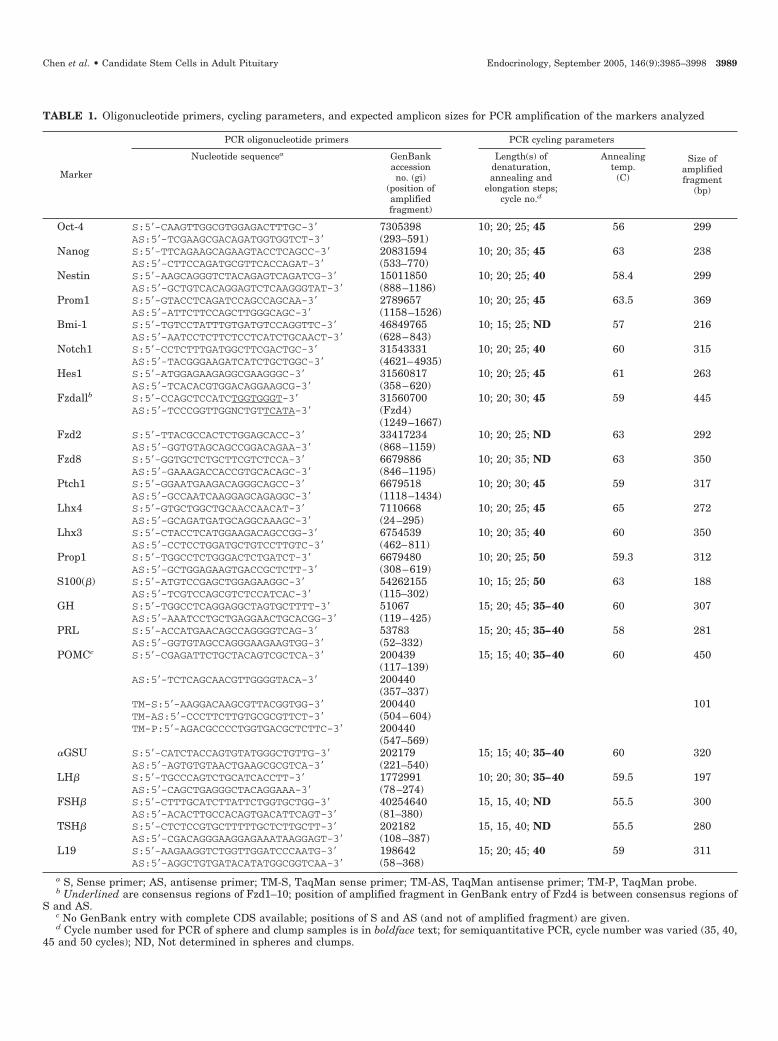
TABLE 1. Oligonucleotide primers, cycling parameters, and expected amplicon sizes for PCR amplification of the markers analyzed
Marker
PCR oligonucleotide primers PCR cycling parameters
Size ofamplifiedfragment
(bp)
Nucleotide sequencea GenBankaccessionno. (gi)
(position ofamplifiedfragment)
Length(s) ofdenaturation,annealing and
elongation steps;cycle no.d
Annealingtemp.
(C)
Oct-4 S:5�-CAAGTTGGCGTGGAGACTTTGC-3� 7305398 10; 20; 25; 45 56 299AS:5�-TCGAAGCGACAGATGGTGGTCT-3� (293–591)
Nanog S:5�-TTCAGAAGCAGAAGTACCTCAGCC-3� 20831594 10; 20; 35; 45 63 238AS:5�-CTTCCAGATGCGTTCACCAGAT-3� (533–770)
Nestin S:5�-AAGCAGGGTCTACAGAGTCAGATCG-3� 15011850 10; 20; 25; 40 58.4 299AS:5�-GCTGTCACAGGAGTCTCAAGGGTAT-3� (888–1186)
Prom1 S:5�-GTACCTCAGATCCAGCCAGCAA-3� 2789657 10; 20; 25; 45 63.5 369AS:5�-ATTCTTCCAGCTTGGGCAGC-3� (1158–1526)
Bmi-1 S:5�-TGTCCTATTTGTGATGTCCAGGTTC-3� 46849765 10; 15; 25; ND 57 216AS:5�-AATCCTCTTCTCCTCATCTGCAACT-3� (628–843)
Notch1 S:5�-CCTCTTTGATGGCTTCGACTGC-3� 31543331 10; 20; 25; 40 60 315AS:5�-TACGGGAAGATCATCTGCTGGC-3� (4621–4935)
Hes1 S:5�-ATGGAGAAGAGGCGAAGGGC-3� 31560817 10; 20; 25; 45 61 263AS:5�-TCACACGTGGACAGGAAGCG-3� (358–620)
Fzdallb S:5�-CCAGCTCCATCTGGTGGGT-3� 31560700 10; 20; 30; 45 59 445AS:5�-TCCCGGTTGGNCTGTTCATA-3� (Fzd4)
(1249–1667)Fzd2 S:5�-TTACGCCACTCTGGAGCACC-3� 33417234 10; 20; 25; ND 63 292
AS:5�-GGTGTAGCAGCCGGACAGAA-3� (868–1159)Fzd8 S:5�-GGTGCTCTGCTTCGTCTCCA-3� 6679886 10; 20; 35; ND 63 350
AS:5�-GAAAGACCACCGTGCACAGC-3� (846–1195)Ptch1 S:5�-GGAATGAAGACAGGGCAGCC-3� 6679518 10; 20; 30; 45 59 317
AS:5�-GCCAATCAAGGAGCAGAGGC-3� (1118–1434)Lhx4 S:5�-GTGCTGGCTGCAACCAACAT-3� 7110668 10; 20; 25; 45 65 272
AS:5�-GCAGATGATGCAGGCAAAGC-3� (24–295)Lhx3 S:5�-CTACCTCATGGAAGACAGCCGG-3� 6754539 10; 20; 35; 40 60 350
AS:5�-CCTCCTGGATGCTGTCCTTGTC-3� (462–811)Prop1 S:5�-TGGCCTCTGGGACTCTGATCT-3� 6679480 10; 20; 25; 50 59.3 312
AS:5�-GCTGGAGAAGTGACCGCTCTT-3� (308–619)S100(�) S:5�-ATGTCCGAGCTGGAGAAGGC-3� 54262155 10; 15; 25; 50 63 188
AS:5�-TCGTCCAGCGTCTCCATCAC-3� (115–302)GH S:5�-TGGCCTCAGGAGGCTAGTGCTTTT-3� 51067 15; 20; 45; 35–40 60 307
AS:5�-AAATCCTGCTGAGGAACTGCACGG-3� (119–425)PRL S:5�-ACCATGAACAGCCAGGGGTCAG-3� 53783 15; 20; 45; 35–40 58 281
AS:5�-GGTGTAGCCAGGGAAGAAGTGG-3� (52–332)POMCc S:5�-CGAGATTCTGCTACAGTCGCTCA-3� 200439 15; 15; 40; 35–40 60 450
(117–139)AS:5�-TCTCAGCAACGTTGGGGTACA-3� 200440
(357–337)TM-S:5�-AAGGACAAGCGTTACGGTGG-3� 200440 101TM-AS:5�-CCCTTCTTGTGCGCGTTCT-3� (504–604)TM-P:5�-AGACGCCCCTGGTGACGCTCTTC-3� 200440
(547–569)�GSU S:5�-CATCTACCAGTGTATGGGCTGTTG-3� 202179 15; 15; 40; 35–40 60 320
AS:5�-AGTGTGTAACTGAAGCGCGTCA-3� (221–540)LH� S:5�-TGCCCAGTCTGCATCACCTT-3� 1772991 10; 20; 30; 35–40 59.5 197
AS:5�-CAGCTGAGGGCTACAGGAAA-3� (78–274)FSH� S:5�-CTTTGCATCTTATTCTGGTGCTGG-3� 40254640 15, 15, 40; ND 55.5 300
AS:5�-ACACTTGCCACAGTGACATTCAGT-3� (81–380)TSH� S:5�-CTCTCCGTGCTTTTTGCTCTTGCTT-3� 202182 15, 15, 40; ND 55.5 280
AS:5�-CGACAGGGAAGGAGAAATAAGGAGT-3� (108–387)L19 S:5�-AAGAAGGTCTGGTTGGATCCCAATG-3� 198642 15; 20; 45; 40 59 311
AS:5�-AGGCTGTGATACATATGGCGGTCAA-3� (58–368)a S, Sense primer; AS, antisense primer; TM-S, TaqMan sense primer; TM-AS, TaqMan antisense primer; TM-P, TaqMan probe.b Underlined are consensus regions of Fzd1–10; position of amplified fragment in GenBank entry of Fzd4 is between consensus regions of
S and AS.c No GenBank entry with complete CDS available; positions of S and AS (and not of amplified fragment) are given.d Cycle number used for PCR of sphere and clump samples is in boldface text; for semiquantitative PCR, cycle number was varied (35, 40,
45 and 50 cycles); ND, Not determined in spheres and clumps.
Chen et al. • Candidate Stem Cells in Adult Pituitary Endocrinology, September 2005, 146(9):3985–3998 3989

ResultsPituitary contains cells with SP phenotype
AP cells from perfused adult mice were stained withHoechst 33342 and analyzed by dual-wavelength flow cy-tometry. A Hoechstlow SP was detected, comprising 1.7%(mean � sem, 1.74 � 0.05%; n � 14 independent cell isola-tions) of the total number of AP cells (Fig. 1A). The SPphenotype was confirmed by the disappearance of theHoechstlow cell population when verapamil was added dur-ing incubation with Hoechst (Fig. 1B), as has been shown forthe SP of other tissues (6–11). Cytometric analysis also re-vealed that pituitary SP cells were cells of small to mediumsize and of very low granularity, clearly different from thebulk of AP cells. SP cells were also identified in AP fromimmature (3 wk old) mice (2.39 � 0.08% of total cells; n � 5)as well as in immature (2 wk old) and adult, rat, and chickenpituitaries (data not shown).
Adult pituitary SP is enriched in cells expressing Sca1, butnot c-Kit or CD34
SP cells from perfused adult AP were characterized byflow cytometry for the expression of HSC markers reportedto be present on stem/progenitor cells and SP cells fromother tissues (7, 8, 11, 17, 19). A majority (70.87 � 1.33%; n �13) of the adult SP cells expressed Sca1 at a high level(Sca1high SP cells), whereas the remaining SP cells were Sca1negative (Sca1neg; Fig. 1, C and D). In the non-SP cell pop-ulation, the high majority of cells were Sca1neg; 5% (5.20 �0.86%) expressed Sca1 at a low level (Sca1low), and 2.5%(2.42 � 0.26%) at a high level (Fig. 1D). Neither the expressionof c-Kit nor that of CD34 (a HSC marker in man) was detectedin SP cells (n � 3). The pan-hemopoietic marker CD45 waspresent on 5% of the SP cells (5.09 � 0.40%; n � 4) and on1% (1.01 � 0.29%) of the non-SP cells. However, no CD45expression was detected within the Sca1high SP, as analyzedby flow cytometry of Hoechst-incubated cells stained withanti-Sca1-R-PE and anti-CD45-PE-Cy5 (n � 3), thereby ex-cluding the presence of HSC in the pituitary Sca1high SP.Epitopes of the membrane antigens CD45, CD34, c-Kit, andSca1 were not destroyed by trypsin treatment as tested inbone marrow (see Materials and Methods).
FIG. 2. The adult AP Sca1high SP is enriched in cells expressingmRNA of stem/progenitor cell- and embryonic pituitary-related fac-tors and signaling molecules. Semiquantitative RT-PCR was per-formed to compare mRNA expression levels in FACS-sorted Sca1high
SP and MP. A, A representative example is shown. A dilution seriesof Sca1high SP and MP RNA (from 104–100 pg/�l), showing comparableL19 expression, was analyzed by RT-PCR for Notch1 expression. InSca1high SP cells, Notch1 mRNA can be detected in a 100-fold lowerRNA amount than in MP cells, resulting in a Notch1 mRNA SP/MPexpression ratio of 100. In addition to varying the input amount ofRNA, amplification cycle numbers were varied (not shown) to ensurequantification in exponential phase (see Materials and Methods). Band C, Representative PCR signals obtained under exponential am-plification conditions are shown together with the final SP/MP ex-pression ratio. For each factor analyzed, RNA isolated from three tofive independent cell sortings was checked by RT-PCR, performed oneor two times per cell sorting. Higher mRNA levels of stem/progenitor
cell-associated factors as well as of members of signaling systems(Notch, Notch1, and Hes1; Wnt, Fzd8; Shh, Ptch1), operative in stemcell regulation and during pituitary embryogenesis, are found in SPthan in MP (B), except for Fzd2. With respect to pituitary-relatedfactors (C), markers of mature pituitary cell phenotypes (hormones)are expressed at a higher mRNA level in MP than in SP. The FS cellmarker S100 was only detected in SP (and in the residual non-SP�non-MP; data not shown). Lhx4, which is crucial for early em-bryonic pituitary development, is only detected in SP, whereas Lhx3,which, in contrast to Lhx4, is not down-regulated after development,is only found in MP. D, Quantitative real-time RT-PCR was performedto verify the results obtained by semiquantitative RT-PCR. Similardifferences in expression between SP and MP were found. The meansof two independent measurements (two independent cell sortings,each analyzed once) are shown. � and indicate that no amplificationsignal is detected in MP or SP, respectively, even at the maximumconditions used (104 pg/�l RNA and 50 cycles of PCR amplification).M, 100-bp marker; Prom1, prominin-1.
3990 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary

Adult AP Sca1high SP is enriched in cells expressing stem/progenitor cell-associated factors
Sca1high (CD45neg) SP cells as well as MP cells (see Fig. 1A)were sorted by FACS and analyzed by semiquantitative RT-PCR (as an example, see Fig. 2A) for the expression of genesassociated with a stem/progenitor cell phenotype in othertissues, i.e. Oct-4 and Nanog expressed in stem cells of theembryo (ES cells), and nestin, prominin-1 and Bmi-1, mainlyfound in NSC and HSC (14, 15, 20–27). Results are shown inFig. 2B. Oct-4 and Nanog mRNA were detected in SP andwere not detectable in MP, at least under the conditions used(for amounts of RNA and cycle numbers, see Materials andMethods). Semiquantitative RT-PCR revealed a 10-fold higherlevel of nestin, Bmi-1, and prominin-1 mRNA in SP than inMP. Quantitative real-time RT-PCR confirmed these higherexpression levels in SP (Fig. 2D). In accordance, nestin pro-tein expression was predominantly detected in SP cells, asanalyzed by immunofluorescence on sorted SP and MP cells(Fig. 3, A and B).
Adult AP Sca1high SP is enriched in cells expressingmembers of Notch, Shh, and Wnt signaling pathways
Sca1high (CD45neg) SP cells and MP cells were analyzed byRT-PCR for the expression of signaling molecules that arefunctional in stem cell renewal and fate determination andthat also play a key role during pituitary early embryonicdevelopment (28–37, 39, 40). Semiquantitative RT-PCR (Fig.2B) showed 100-fold higher levels of Notch1 mRNA in SPthan in MP, and 100- to 1000-fold higher mRNA values of itsdownstream target hairy and enhancer of split 1 (Hes1). ThemRNA of the Shh receptor Patched1 (Ptch1; also referred toas Ptc1) was 10-fold more abundant in SP than in MP. Ex-pression levels of Notch1, Hes1, and Ptch1 were confirmedby quantitative real-time RT-PCR; SP/MP ratios were of thesame order of magnitude as those obtained by semiquanti-tative RT-PCR (see Fig. 2D: SP/MP � 392 for Notch1, 694 forHes1, and 7 for Ptch1). Similarly, Notch1 protein was presentat a detectable level in (some) cells of the SP, but not in cellsof the MP (Fig. 3, C and D).
The expression of frizzled (Fzd) receptors, implied in ca-nonical Wnt signaling, was analyzed by RT-PCR, first usingPCR primers in consensus sequences of all known Fzd re-ceptor subtypes (Fzd1–10; see Table 1). A 10-fold higher levelof Fzd mRNA expression (FzdAll) was detected in SP vs. MP(Fig. 2B). Sequencing of the amplified fragments revealedpredominant amplification of Fzd1 and Fzd2 from MP, andof Fzd8 from SP. Using sequence-specific PCR primers (seeTable 1), it was found that Fzd8 mRNA was 10-fold higherin SP than in MP, whereas Fzd2 mRNA showed the oppositepicture. Quantitative real-time RT-PCR supported the dif-ferences obtained by semiquantitative RT-PCR (Fig. 2D).
Adult AP Sca1high SP is enriched in cells expressing theearly embryonic pituitary transcription factor Lhx4, but notLhx3 or hormones
Sca1high SP cells as well as MP cells were analyzed byRT-PCR for the expression of transcription factors essentialduring early embryonic development of the AP (37, 38, 41,
42) and of hormones as phenotypic markers of differentiatedAP cells. Lhx4 was only detectable in SP, not in MP, at boththe mRNA and protein levels (Fig. 2C and Fig. 3, E and F).In contrast, Lhx3 (which is not down-regulated after pitu-itary development) was only detected in MP (Fig. 2C and Fig.3, G and H). Prop1, an embryonic transcription factor indis-pensable for development of the Pit-1 lineage (lactotropes,somatotropes, and thyrotropes) (49) was only found in MP(Fig. 2C). The �GSU mRNA level was 10-fold higher in MPthan in SP, prolactin (PRL), LH�, GH, and POMC levels were100-fold higher, and FSH� and TSH� levels were more than1000-fold higher (not detected in SP). Differences of the sameorder of magnitude were found using quantitative real-timeRT-PCR (Fig. 2D). Correspondingly, PRL-immunoreactive
FIG. 3. Expression of stem/progenitor cell- and pituitary-related fac-tors and signaling molecules in SP and MP cells, as analyzed byimmunofluorescence. Sorted SP and MP cells were subjected to im-munofluorescent analysis. Nestin-immunoreactive cells were pre-dominantly found in the SP when compared with the MP (A and B).Notch1 (C and D) and Lhx4 (E and F) were expressed at detectablelevels in some SP cells, but not in MP cells. In contrast, Lhx3-immunoreactive (G and H) and PRL-immunoreactive (I and J) cellswere abundant in the MP, but were depleted from the SP. Theseimmunofluorescent data confirm the mRNA expression profilesshown in Fig. 2. Bar, 50 �m.
Chen et al. • Candidate Stem Cells in Adult Pituitary Endocrinology, September 2005, 146(9):3985–3998 3991

cells were abundantly present in the MP, whereas they weredepleted from the SP (Fig. 3, I and J).
AP Sca1high SP contains cells with FS cell phenotype
Sca1high SP and MP cells were analyzed by semiquantita-tive RT-PCR for the expression of S100 as a marker of AP FScells. S100 mRNA was detected in SP, but not in MP (Fig. 2C).Because there are more FS cells (7%) (50) than SP cells(1.7%) in the adult mouse AP, we searched for the remainderof the FS cells. In addition to the SP and MP, the residualnon-SP�non-MP cell population was sorted (see Fig. 1A forSP and MP sorting gates) and analyzed for S100 expression.S100 mRNA was found in the non-SP�non-MP at the sameexpression level as in the SP (data not shown). To analyzewhether all or only part of the SP cells were FS cells, quan-tification of FS cells within the SP was performed by FACSanalysis of AP cells simultaneously incubated with Hoechstand the dipeptide �-Ala-Lys-N�-AMCA. Uptake of thisdipeptide is a characteristic capacity of FS cells in the AP (46).FACS analysis revealed that dipeptide-positive cells ac-counted for 6% of the total AP cells (5.90 � 0.35%; n � 4), apercentage comparable to the proportion of FS cells in themouse AP (50). Within the SP, only 5.81 � 0.50% (n � 4) ofthe cells were dipeptide positive.
Nonadherent spheres develop in AP cell cultures underneurosphere-like growth conditions
In a complementary approach to prospectively identifyand characterize pituitary stem/progenitor cells, AP cellsfrom 3-wk-old or adult mice were cultured under slightlymodified neurosphere-forming conditions (see Materials andMethods). During stationary culture in serum-free, chemi-cally defined culture medium (45) supplemented with B27and bFGF, free-floating spheres progressively developed intime (Fig. 4). Spheres were bright, smooth-edged, and com-pact and were clearly different from irregular clumps of cellsthat also appeared in the culture (Fig. 4). On d 3 of culture(Fig. 4D), spheres displayed a diameter of 41.6 � 1.0 �m(mean � sem of 39 spheres; range, 30.2–56.4 �m), and con-tained 17 � 2 cells (mean � sem of 10 spheres; range, 9–26cells). The spheres expanded in size as well as in numberduring the next 3 d. After 6 d in culture (Fig. 4F), 30–50spheres were observed per 200,000 AP cells, displaying adiameter of 70.9 � 5.7 �m (mean � sem of 10 spheres; range,45.2–97.0 �m) and containing 42 � 2 cells (mean � sem of 10spheres; range, 32–54 cells). From d 6, spheres did not ob-servably increase in size (Fig. 4, F vs. G), but clumps and cellsin suspension started to attach to the spheres to eventuallyform larger aggregates (diameter, 100–300 �m; Fig. 4H).Spheres could be recovered from these aggregates by me-ticulously controlled trypsinization (see Materials and Meth-ods). Spheres recovered from d 8–10 aggregates showed adiameter of 96.6 � 4.6 �m (mean � sem of five spheres; range,78.4–111.7 �m) and contained an estimated 100–200 cells.
Trials with other media were performed to grow pituitaryspheres. B27 supplement provided essential components,which could not be replaced by N2, another supplementsometimes used to grow neurospheres. bFGF (or, alterna-tively, EGF) was found to be indispensable, and no spheres
were obtained with other growth factors, such as nervegrowth factor. When the chemically defined medium wasreplaced by the classical neurosphere culture mediumDMEM/Ham’s F-12 (also supplemented with B27 and bFGFor EGF), floating spheres appeared, but a cavity almost im-mediately developed in the body of the sphere (data notshown). These hollow spheres transformed into aggregatesmore readily (1–2 d after appearance) than the more compactspheres in chemically defined medium.
Pituitary spheres are clonal in origin and can generatesecondary spheres
Because pituitary cells have a tendency to reassociate, asevident from the eventual formation of larger aggregates inthe sphere cultures as well as from our previous work usingreaggregate pituitary cell cultures (44, 45), it was determinedwhether pituitary spheres originated from one cell. First,low-density cultures (1,000 cells/ml instead of 100,000) werescrutinized, and spheres were also obtained under theseconditions (about one sphere per 4,000 cells). In view of thelow incidence of sphere formation, an approach of growingspheres from single cells cloned by FACS or limiting dilution
FIG. 4. Nonadherent spheres develop in AP cell cultures underneurosphere-like growth conditions. Cells from dispersed APs areseeded in serum-free chemically defined culture medium supple-mented with B27 and bFGF (A; D0, d 0). Bright smooth-edged spheresstart to appear on d 1 (B; arrow), whereas some cells stick to each otherand form clumps with an irregular outline (arrowhead). Spheres growuntil d 6 (C–G). Then, clumps and cells in suspension start to attachto the spheres, eventually forming larger aggregates (H). Bar, 50 �m.
3992 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary
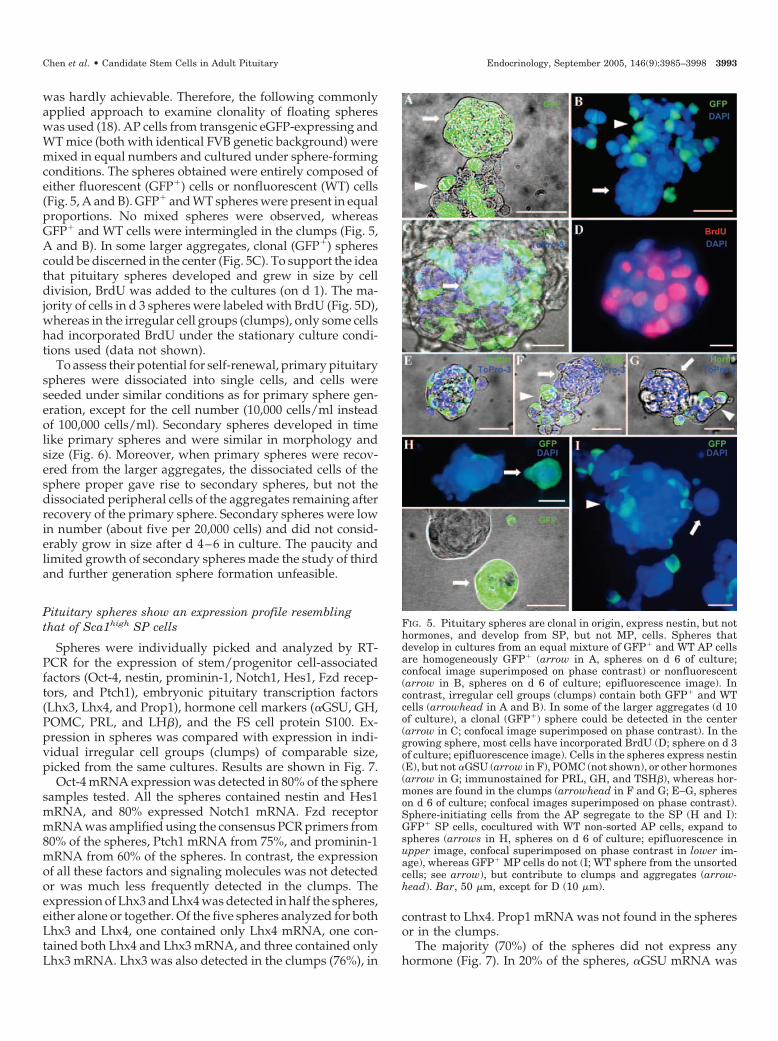
was hardly achievable. Therefore, the following commonlyapplied approach to examine clonality of floating sphereswas used (18). AP cells from transgenic eGFP-expressing andWT mice (both with identical FVB genetic background) weremixed in equal numbers and cultured under sphere-formingconditions. The spheres obtained were entirely composed ofeither fluorescent (GFP�) cells or nonfluorescent (WT) cells(Fig. 5, A and B). GFP� and WT spheres were present in equalproportions. No mixed spheres were observed, whereasGFP� and WT cells were intermingled in the clumps (Fig. 5,A and B). In some larger aggregates, clonal (GFP�) spherescould be discerned in the center (Fig. 5C). To support the ideathat pituitary spheres developed and grew in size by celldivision, BrdU was added to the cultures (on d 1). The ma-jority of cells in d 3 spheres were labeled with BrdU (Fig. 5D),whereas in the irregular cell groups (clumps), only some cellshad incorporated BrdU under the stationary culture condi-tions used (data not shown).
To assess their potential for self-renewal, primary pituitaryspheres were dissociated into single cells, and cells wereseeded under similar conditions as for primary sphere gen-eration, except for the cell number (10,000 cells/ml insteadof 100,000 cells/ml). Secondary spheres developed in timelike primary spheres and were similar in morphology andsize (Fig. 6). Moreover, when primary spheres were recov-ered from the larger aggregates, the dissociated cells of thesphere proper gave rise to secondary spheres, but not thedissociated peripheral cells of the aggregates remaining afterrecovery of the primary sphere. Secondary spheres were lowin number (about five per 20,000 cells) and did not consid-erably grow in size after d 4–6 in culture. The paucity andlimited growth of secondary spheres made the study of thirdand further generation sphere formation unfeasible.
Pituitary spheres show an expression profile resemblingthat of Sca1high SP cells
Spheres were individually picked and analyzed by RT-PCR for the expression of stem/progenitor cell-associatedfactors (Oct-4, nestin, prominin-1, Notch1, Hes1, Fzd recep-tors, and Ptch1), embryonic pituitary transcription factors(Lhx3, Lhx4, and Prop1), hormone cell markers (�GSU, GH,POMC, PRL, and LH�), and the FS cell protein S100. Ex-pression in spheres was compared with expression in indi-vidual irregular cell groups (clumps) of comparable size,picked from the same cultures. Results are shown in Fig. 7.
Oct-4 mRNA expression was detected in 80% of the spheresamples tested. All the spheres contained nestin and Hes1mRNA, and 80% expressed Notch1 mRNA. Fzd receptormRNA was amplified using the consensus PCR primers from80% of the spheres, Ptch1 mRNA from 75%, and prominin-1mRNA from 60% of the spheres. In contrast, the expressionof all these factors and signaling molecules was not detectedor was much less frequently detected in the clumps. Theexpression of Lhx3 and Lhx4 was detected in half the spheres,either alone or together. Of the five spheres analyzed for bothLhx3 and Lhx4, one contained only Lhx4 mRNA, one con-tained both Lhx4 and Lhx3 mRNA, and three contained onlyLhx3 mRNA. Lhx3 was also detected in the clumps (76%), in
contrast to Lhx4. Prop1 mRNA was not found in the spheresor in the clumps.
The majority (70%) of the spheres did not express anyhormone (Fig. 7). In 20% of the spheres, �GSU mRNA was
FIG. 5. Pituitary spheres are clonal in origin, express nestin, but nothormones, and develop from SP, but not MP, cells. Spheres thatdevelop in cultures from an equal mixture of GFP� and WT AP cellsare homogeneously GFP� (arrow in A, spheres on d 6 of culture;confocal image superimposed on phase contrast) or nonfluorescent(arrow in B, spheres on d 6 of culture; epifluorescence image). Incontrast, irregular cell groups (clumps) contain both GFP� and WTcells (arrowhead in A and B). In some of the larger aggregates (d 10of culture), a clonal (GFP�) sphere could be detected in the center(arrow in C; confocal image superimposed on phase contrast). In thegrowing sphere, most cells have incorporated BrdU (D; sphere on d 3of culture; epifluorescence image). Cells in the spheres express nestin(E), but not �GSU (arrow in F), POMC (not shown), or other hormones(arrow in G; immunostained for PRL, GH, and TSH�), whereas hor-mones are found in the clumps (arrowhead in F and G; E–G, sphereson d 6 of culture; confocal images superimposed on phase contrast).Sphere-initiating cells from the AP segregate to the SP (H and I):GFP� SP cells, cocultured with WT non-sorted AP cells, expand tospheres (arrows in H, spheres on d 6 of culture; epifluorescence inupper image, confocal superimposed on phase contrast in lower im-age), whereas GFP� MP cells do not (I; WT sphere from the unsortedcells; see arrow), but contribute to clumps and aggregates (arrow-head). Bar, 50 �m, except for D (10 �m).
Chen et al. • Candidate Stem Cells in Adult Pituitary Endocrinology, September 2005, 146(9):3985–3998 3993

detected, alone or in combination with POMC mRNA. Noneof the pituitary spheres tested contained mRNA of PRL orLH� at a detectable level. In contrast, PRL and/or LH�mRNA was detected in 89% and 33% of the clumps, respec-tively. �GSU and/or GH mRNA were amplified from 80%of the clumps, and POMC mRNA was amplified from 100%.S100 mRNA was observed in some clumps (7%), but not inspheres.
Similar differences in mRNA expression pattern betweenspheres and clumps were found when total RNA from anidentical number of sphere cells or clump cells was analyzedby semiquantitative RT-PCR as performed for a selection ofthe markers (see Materials and Methods; data not shown).These control experiments indicate that the single sphere orclump lysis method used above (see Materials and Methods)did not affect RT-PCR outcome, and that unintentional dif-
ferences in cell number (which we tried to avoid by pickingspheres and clumps of similar sizes) were not responsible forthe specific expression profiles obtained.
The expression of nestin and Lhx3 was also detected at theprotein level in the spheres (in almost all and in 70% of thespheres, respectively; Fig. 5E and data not shown). In accor-dance with the mRNA expression pattern, nestin and Lhx3protein were also detected in some or a large part of theclumps, respectively. The expression of �GSU, POMC, andother hormones (combined analysis for PRL, GH, and TSH�)was detected in cell clumps, but not in spheres (Fig. 5, F andG, and data not shown).
Sphere-initiating cells of AP segregate to SP
In view of the largely overlapping expression profile of SPand sphere cells, we examined whether there was a rela-tionship between these cells and, in particular, whether thesphere-initiating cells of the AP displayed an SP phenotypeand purified within the SP. GFP� SP cells as well as GFP� MPcells were isolated by FACS from APs of eGFP transgenicmice and cocultured under sphere-growing conditions withunsorted AP cells from WT mice at a ratio of 10,000/200,000/dish. In the GFP� SP cocultures, spheres (10–20/dish) weredetected that were homogeneously GFP� (Fig. 5H) in addi-tion to the normal number of spheres (n � 30–50) consistingof WT cells only. In the GFP� MP cocultures, no GFP�
spheres were found, but only spheres consisting of WT cells(Fig. 5I). GFP� cells were present in clumps and aggregatesof the GFP� MP cocultures (Fig. 5I), indicating that longerretention of Hoechst dye was not toxic to the MP cells. Also,the viability of SP and MP cells was identical after sorting(90%), as checked by the trypan blue exclusion test.
Discussion
In the study presented here, we show that the adult mouseAP contains a population of cells with verapamil-sensitivedye efflux capacity, a characteristic originally assigned toHSC in bone marrow (6) and subsequently to stem/progen-itor cells in a number of other tissues, such as forebrain,
FIG. 7. Pituitary spheres express mRNA of stem/progenitor cell- and embryonic pituitary-relatedfactors and signaling molecules. Spheres andclumps (irregular groups of attached cells) wereindividually picked after 6–12 d in culture andanalyzed by RT-PCR for mRNA expression of themarkers shown. The expression of stem/progen-itor cell-associated factors and signaling mole-cules was detected in a much higher proportion ofpituitary spheres than clumps. The early embry-onic transcription factor Lhx4 was only detectedin spheres, whereas Lhx3 was found in bothspheres and clumps. In contrast, hormone mRNAexpression was observed in a high number ofclumps and in only a minority of the pituitaryspheres. The total numbers of spheres and clumpsanalyzed per marker are indicated below the his-togram; they were picked from three to six inde-pendent cultures.
FIG. 6. Cells from primary pituitary spheres can generate secondaryspheres. Primary pituitary spheres are dissociated into single cellsand cells seeded in culture under sphere-forming conditions (A; D0,d 0). Secondary spheres develop from these cells and grow until d 4–6(B–D). Bar, 50 �m.
3994 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary

mammary gland, skeletal muscle, and testis (7, 8, 10, 11). Wefound that this so-called SP is enriched in cells that expressstem/progenitor cell-connected factors, such as Sca1, Oct-4,Nanog, nestin, prominin-1, and Bmi-1, and members of theNotch (Notch1 and Hes1), Wnt (Fzd8), and Shh (Ptch1) sig-naling pathways functional in stem cell homeostasis as wellas in pituitary embryogenesis. Moreover, the SP containscells expressing the early embryonic pituitary transcriptionfactor Lhx4, whereas it is depleted from cells expressing Lhx3and from differentiated cells (expressing hormones). Finally,the SP includes cells that can clonally expand to sphericalstructures, which in several other tissues are known to growfrom the resident stem/progenitor cells (12–19). These pitu-itary spheres display an expression profile largely overlap-ping with that of the Sca1high SP cells. Thus, our study de-scribes a hitherto unobserved cell population in the adult APwith characteristics suggestive of an early embryonic andstem/progenitor cell phenotype.
The AP Sca1high SP, characterized in the present study,does not appear to be contaminated with HSC, because nei-ther CD45� nor c-Kit� and CD34� cells were detected withinthis fraction. In fact, this was not expected, because mice wereperfused before harvesting the pituitaries, removing poten-tial HSC present in the intrapituitary blood. Furthermore,pituitary Sca1high SP cells were not able to rescue mice fromlethal irradiation (Vankelecom, H., unpublished observations).
The transcription factors Oct-4 and Nanog, to date onlyreported in the archetype stem cell (ES cell) (22, 23) and insome candidate adult stem/progenitor cells obtained in cul-ture (24), are expressed in the adult pituitary SP. To ourknowledge, this is the first report of Oct-4 and Nanog ex-pression in an adult organ ex vivo. Oct-4 and Nanog play anessential role in keeping ES cells undifferentiated and self-renewing (22, 23). Along the same line, the protooncogeneproduct, Bmi-1, and the membrane antigen, Sca1, have beendemonstrated to be functional in HSC and NSC self-renewal(26, 27, 48). It is tempting to assign similar roles to thesefactors within the context of the adult AP. The function of theintermediate filament protein nestin and of the membraneglycoprotein prominin-1 in stem/progenitor cell biology isas yet not clear.
The adult pituitary SP displays a striking embryonic ex-pression profile. Lhx4 is exclusively found in SP, whereas therelated Lhx3 is only detected within MP cells. Lhx3 and Lhx4play cardinal roles in formation and fate commitment ofRathke’s pouch as well as in AP cell lineage specification,expansion, and terminal differentiation (41, 42). A recentstudy placed Lhx4 upstream of Lhx3 during pituitary de-velopment by showing that Lhx4 is required for the timelyactivation of Lhx3 (51). The same study also identified Lhx4as a critical factor for survival and maintenance of the pu-tative progenitor cells in the embryonic pituitary. Lhx3 re-mains well expressed in the adult gland (41, 42). In contrast,Lhx4 expression shows a marked diminution from E15.5 andis only detected at a very low level in the adult AP (42). Ourfinding of selective Lhx4 expression in the SP indicates thatLhx4-expressing cells of the adult pituitary segregate to theSP. These SP cells may represent enduring early embryoniccells maintained by Lhx4 expression, which is in line with theabove-mentioned role for Lhx4 in embryonic progenitor cell
maintenance (51), or alternatively, cells in the adult pituitarycapable of using the embryonic program. In both cases, thesecells can further differentiate and thus function as sourcecells for new endocrine cells within the adult gland. Lhx3 hasbeen suggested to remain expressed in the differentiatedhormone cells considering its widespread presence in theadult AP (41, 42). Along the same line, Lhx3 has been shownto regulate the expression of Pit-1, �GSU, FSH�, and, insynergy with Pit-1, GH and PRL (41, 52). Our finding of Lhx3expression specifically in the MP is in agreement with Lhx3activity in further differentiated pituitary cells. Prop1 wasnot detected in the SP, but only in the MP. Prop1 is essentialfor development of Pit-1-dependent lineages (49), but hasrecently also been suggested to play a role in the migrationof putative AP progenitors from the perilumenal region ofRathke’s pouch to the developing AP while they differentiate(53). Thus, SP cells in the adult pituitary do not seem toexpress this property.
Other protagonists of pituitary early embryonic develop-ment, similar to Lhx4, are detected at higher levels in SP thanin MP, viz. members of the Notch, Wnt, and Shh pathway,which are, at the same time, key regulators of the balancebetween self-renewal and fate determination in stem cells(28–36). A first striking observation is the markedly higherexpression of Notch1 and one of its downstream effectors(Hes1) by SP cells. The expression of both members suggestsactivated Notch signaling in pituitary SP cells or in at leastsome of them. Notch signaling has only very recently beenimplicated in pituitary embryogenesis based on the detectionof Notch pathway components in a temporally and spatiallyrestricted manner (40). Interestingly, Hes1 expression is re-stricted to the proliferating cells adjacent to the residuallumen of Rathke’s pouch, which are generally regarded asembryonic progenitor cells (40, 53, 54), whereas it is excludedfrom the differentiating cells in the developing AP. In thecentral nervous system, Hes1 is a key component in pre-serving the self-renewal capacity of NSC and repressing theircommitment to a neuronal fate (55). Our finding of Hes1 inthe adult pituitary SP could therefore be consistent with arole in maintaining these cells, or at least some of them, in anundifferentiated phenotype and capable of self-renewing.
Another signaling system recently implicated in pituitaryorganogenesis is the canonical Wnt pathway (37, 56, 57). Ourfinding of differential expression of Fzd receptors in SP andMP, i.e. more Fzd8 in SP and more Fzd2 in MP, suggests adivergent function for Wnt signals in each of these two cellpopulations within the adult AP. Previously, the expressionof Fzd2 (�Fzd10) was detected not only in the early embry-onic pituitary, but also in the adult gland (56) as well as inpituitary cell lines (G7, GHFT1, �TSH, and �T3) that repre-sent different stages of committed or differentiated hormonecell lineages (56). Our finding of higher Fzd2 expression inMP is consistent with Fzd2 being present in committed ordifferentiated cells. Fzd8, on the other hand, has (togetherwith Fzd4, Fzd6, and Fzd7) mainly been associated with astem cell phenotype (9), and our finding of Fzd8 expressionpredominantly in SP seems to be in line with this.
A final signaling pathway analyzed in the present studyand implicated in pituitary embryonic development is thatmediated by Shh. Shh is initially expressed uniformly
Chen et al. • Candidate Stem Cells in Adult Pituitary Endocrinology, September 2005, 146(9):3985–3998 3995

throughout the oral ectoderm, but its expression is excludedfrom the nascent Rathke’s pouch on E9.0–9.5 (37, 39), cre-ating a molecular boundary within the continuous ectoderm.Its receptor, Ptch1 (which silences signaling of Smoothenedin the absence of Shh), remains expressed in the invaginatingpouch, indicating that cells of the nascent pituitary are re-ceiving a Shh signal, because Ptch1 is a direct transcriptionaltarget of Shh. Using genetically modified mice, it was con-firmed that Shh signaling from the expression boundarywithin the oral epithelium is required for proliferation, celltype determination, and, in cooperation with FGF8, properexpression of Lhx3 during pituitary organogenesis (39). Wemeasured higher mRNA levels of Ptch1 receptor in SP thanin MP. The Ptch1-expressing SP cells in the adult pituitarymay thus also represent cells of embryonic phenotype thatreceive Shh signals.
Taken together, identification in the adult pituitary of sig-naling systems that are functional during embryonic devel-opment of the gland is in line with the growing idea thatembryonic morphogens and signals persist in adult tissues tomaintain life-long cellular replacement, and that cells withpreserved embryonic characteristics may function as stem/progenitor cells in the adult tissue (19, 31, 34). In view of thisconcept, it is tempting to hypothesize that SP cells, at leastsome of them, represent stem/progenitor cells in the adultpituitary maintained or triggered by (local) embryonic sig-nals to recapitulate development. To substantiate this hy-pothesis, it will be of great interest to define how the ex-pression of the particular factors identified is distributedover individual SP cells. Alternative hypotheses, however,should not be overlooked. SP cells, or at least some of them,may correspond to cells that have dedifferentiated and arecapable of generating new hormone-producing cells using aprogram mimicking the embryonic process (58). Dediffer-entiated cells may represent a flexible pool of reserve cellsfrom which specific cell types can develop according to theendocrine needs of the organism. On the other hand, SP cellsmay embody intermediates in transdifferentiation eventsthat may occur via an immature (uncommitted) state (58).Such phenotypic switches between different cell types arenot uncommon in the pituitary (59–61).
The AP SP also contains FS cells, because S100 mRNA isdetected in the SP, and some SP cells (6%) have the capacityto take up the dipeptide �-Ala-Lys-N�-AMCA, a character-istic property of FS cells in the AP (46). Because FS cellscomprise approximately 7% of the mouse AP cells (50), andthe SP comprises only 1.7%, it is clear that only a smallsubpopulation of FS cells purifies with the SP. Some inves-tigators have in the past suggested a stem cell role for FS cells(62), but convincing evidence is still lacking. The remainingS100 mRNA-expressing cells are found in the cell populationoutside the SP and MP sorting gates in the FACS plot (thenon-SP�non-MP).
In a complementary approach toward stem/progenitorcell identification, we were able to obtain clonal spheres incell cultures from mouse AP. Sphere cultures have beeninstrumental in the identification and characterization ofstem/progenitor cells from a number of tissues (12–19). In-terestingly, sphere-initiating cells display an SP phenotypeand segregate to this Hoechstlow population, as has also been
reported for neurosphere-forming cells (9, 10). Moreover,pituitary spheres exhibit an expression profile largely incommon with that of Sca1high SP cells: a high percentage ofthe pituitary spheres express factors known to be present orfunctional in stem/progenitor cells and embryonic pituitarycells, whereas only a limited number express markers ofmature pituitary cells (hormones). Again, the expression ofboth Notch1 and Hes1 is very striking and suggests that theNotch signaling pathway is activated in spheres. There arealso some differences, however, between SP and the sphereexpression profile. S100 mRNA is not found in spheres,whereas FS cells are present in the SP, although at low num-ber. The opposite is true for Lhx3, which is detected inspheres, but not in SP. The expression of Lhx3 in pituitaryspheres may be associated with the generation of furtherdifferentiated cells within the sphere. As demonstrated inneurospheres and mammospheres (clonal spheres from themammary gland) (10, 16, 35), it is not inconceivable that thepituitary sphere is composed of one or only a limited numberof sphere-initiating, self-renewing cells and, for the remain-der, of further differentiated, more restricted cells with noself-renewal, but proliferative capacity. During progressionalong the differentiation path, Lhx4 expression may decreaseas occurring during embryonic development, potentially ex-plaining why Lhx4 is not detectable anymore in somespheres. Along the same line, the expression in some spheresof �GSU and POMC, the two earliest markers of embryonicAP cell differentiation (43, 63), may be a first sign of (early)differentiation within the sphere. Again, it will be of greatinterest to identify the expression at the individual cell levelin the sphere to substantiate this hypothesis.
About 0.02% of the AP cells possess sphere-forming ca-pacity, comparable to the proportion of sphere-initiatingcells in pancreas (18) and skin (19), but smaller in numberthan their counterparts in mammary gland (16). In the pi-tuitary spheres obtained, cell proliferation appears to ceaseat a certain time point in culture. A similar halt in spheregrowth has been reported for pancreatic spheres (18). Thereason is unclear at present, but may relate to the lack ofessential cell-cell contacts with other pituitary cells (support-ive niche cells?) and/or of crucial juxtacrine growth factorsfrom these cells. One argument in favor of this hypothesis isthat when other AP cells are attached to the spheres, as in thelarger aggregates, the spheres recovered are bigger and con-tain more cells. The number of cells in pituitary spheres iscomparable to the number reported in skin-derived spheresand mammospheres (16, 19, 35), but lower than that in pan-creatic spheres and neurospheres (10, 18). Only a few sec-ondary spheres were generated from primary sphere cells.The generation of limited numbers of (small) secondaryspheres has also been reported for pancreatic spheres (18).The reasons for this may be similar to those for limitedprimary sphere expansion.
In conclusion, we show in this study that the mouse APcontains a population of cells with SP characteristics, asfound in other tissues. This population is enriched in cellsthat express markers and signaling molecules indicative ofstem/progenitor cells and contains cells that generate clonalspheres with comparable expression profiles. Furthermore,at least some of these cells express members of signaling
3996 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary

systems (Notch, Wnt, and Shh) and transcription factors(Lhx4) that are essential during pituitary embryogenesis,suggesting that they represent cells with preserved embry-onic nature that may function as source cells for new hor-mone-producing cells in the adult gland. Additional elabo-rate studies should provide insight into the potential of thesecells, or at least some of them, to differentiate into multiplelineages of the AP to lend further support to such a stem/progenitor cell phenotype.
Acknowledgments
We thank Dr. A. Nagy (Mount Sinai Hospital, Samuel LunenfeldResearch Institute, Toronto, Canada) for providing the eGFP transgenicmice, Dr. K. Bauer (Max Planck Institute, Hannover, Germany) for thegenerous gift of the dipeptide �-Ala-Lys-N�-AMCA, Dr. R. McKay (Na-tional Institutes of Health, Bethesda, MD) for the polyclonal antinestinantiserum, Dr. S. Pfaff (The Salk Institute for Biological Studies, La Jolla,CA) for the Lhx3 and Lhx4 antibodies, and Dr. A. F. Parlow (Harbor-University of California-Los Angeles Medical Center, Torrance, CA) forthe hormone antisera. We highly appreciate the sharing by K. U. Leuvenresearch groups of the ABI PRISM 7700 (Dr. G. Carmeliet), the ABIPRISM 7000 (Drs. P. Marynen and G. Froyen), the confocal laser scanningmicroscope (Dr. P. Carmeliet), and the FACSVantage (Dr. M. Delforge).M. Boussemaere is acknowledged for her skilful help with mouse per-fusion and pituitary cell dispersion, and Dr. O. Krylyshkina for experthelp with immunostaining.
Received February 11, 2005. Accepted May 23, 2005.Address all correspondence and requests for reprints to: Dr. Hugo
Vankelecom, Laboratory of Cell Pharmacology, Department of Molec-ular Cell Biology, University of Leuven (K.U.Leuven), CampusGasthuisberg (O&N), Herestraat 49, B-3000 Leuven, Belgium. E-mail:[email protected].
This work was supported by grants from the Fund for ScientificResearch-Flanders, Belgium (F.W.O.-Vlaanderen), and the Flemish Min-istry of Science Policy (Concerted Research Actions).
References
1. Nolan LA, Kavanagh E, Lightman SL, Levy A 1998 Anterior pituitary cellpopulation control: basal cell turnover and the effects of adrenalectomy anddexamethasone treatment. J Neuroendocrinol 10:207–215
2. Taniguchi Y, Yasutaka S, Kominami R, Shinohara H 2002 Proliferation anddifferentiation of rat anterior pituitary cells. Anat Embryol 206:1–11
3. Carbajo-Perez E, Watanabe YG 1990 Cellular proliferation in the anteriorpituitary of the rat during the postnatal period. Cell Tissue Res 261:333–338
4. Landolt AM 1973 Regeneration of human pituitary. J Neurosurg 39:35–415. Saeger W, Warnecke H 1980 Ultrastructural examination of the regeneration
of the rat adenohypophysis after partial hypophysectomy. Virchows Arch APathol Anat Histopathol 387:279–288
6. Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC 1996 Isolation andfunctional properties of murine hematopoietic stem cells that are replicatingin vivo. J Exp Med 183:1797–1806
7. Gussoni E, Soneoka Y, Strickland CD, Buzney EA, Khan MK, Flint AF,Kunkel LM, Mulligan RC 1999 Dystrophin expression in the mdx mouserestored by stem cell transplantation. Nature 401:390–394
8. Welm BE, Tepera SB, Venezia T, Graubert TA, Rosen JM, Goodell MA 2002Sca-1(pos) cells in the mouse mammary gland represent an enriched progen-itor cell population. Dev Biol 245:42–56
9. Ivanova NB, Dimos JT, Schaniel C, Hackney JA, Moore KA, Lemischka IR2002 A stem cell molecular signature. Science 298:601–604
10. Kim M, Morshead CM 2003 Distinct populations of forebrain neural stem andprogenitor cells can be isolated using side-population analysis. J Neurosci23:10703–10709
11. Falciatori I, Borsellino G, Haliassos N, Boitani C, Corallini S, Battistini L,Bernardi G, Stefanini M, Vicini E 2004 Identification and enrichment ofspermatogonial stem cells displaying side-population phenotype in immaturemouse testis. FASEB J 18:376–378
12. Reynolds BA, Weiss S 1992 Generation of neurons and astrocytes from iso-lated cells of the adult mammalian central-nervous-system. Science 255:1707–1710
13. Gritti A, Parati EA, Cova L, Frolichsthal P, Galii R, Wanke E, Faravelli L,Morassutti DJ, Roisen F, Nickel DD, Vescovi AL 1996 Multipotential stem
cells from the adult mouse brain proliferate and self-renew in response to basicfibroblast growth factor. J Neurosci 16:1091–1100
14. Tropepe V, Coles BLK, Chiasson BJ, Horsford DJ, Elia AJ, McInnes RR, vander Kooy D 2000 Retinal stem cells in the adult mammalian eye. Science287:2032–2036
15. Toma JG, Akhavan M, Fernandes KJL, Barnabe-Heider F, Sadikot A, KaplanDR, Miller FD 2001 Isolation of multipotent adult stem cells from the dermisof mammalian skin. Nat Cell Biol 3:778–784
16. Dontu G, Abdallah WM, Foley JM, Jackson KW, Clarke MF, Kawamura MJ,Wicha MS 2003 In vitro propagation and transcriptional profiling of humanmammary stem/progenitor cells. Genes Dev 17:1253–1270
17. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, Kasa-hara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B,Anversa P 2003 Adult cardiac stem cells are multipotent and support myo-cardial regeneration. Cell 114:763–776
18. Seaberg RM, Smukler SR, Kieffer TJ, Enikolopov G, Asghar Z, Wheeler MB,Korbutt G, van der Kooy D 2004 Clonal identification of multipotent pre-cursors from adult mouse pancreas that generate neural and pancreatic lin-eages. Nat Biotechnol 22:1115–1124
19. Fernandes KJL, McKenzie IA, Mill P, Smith KM, Akhavan M, Barnabe-Heider F, Biernaskie J, Junek A, Kobayashi NR, Toma JG, Kaplan DR,Labosky PA, Rafuse V, Hui CC, Miller FD 2004 A dermal niche for multi-potent adult skin-derived precursor cells. Nat Cell Biol 6:1082–1093
20. Yin AH, Miraglia S, Zanjani ED, AlmeidaPorada G, Ogawa M, Leary AG,Olweus J, Kearney J, Buck DW 1997 AC133, a novel marker for humanhematopoietic stem and progenitor cells. Blood 90:5002–5012
21. Uchida N, Buck D, He D, Reitsma M, Masek M, Phan T, Tsukamoto A, GageF, Weissman I 2000 Direct isolation of human central nervous system stemcells. Proc Natl Acad Sci USA 97:14720–14725
22. Niwa H, Miyazaki J, Smith AG 2000 Quantitative expression of Oct-3/4defines differentiation, dedifferentiation or self-renewal of ES cells. Nat Genet24:372–376
23. Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, Takahashi K,Maruyama M, Maeda M, Yamanaka S 2003 The homeoprotein Nanog isrequired for maintenance of pluripotency in mouse epiblast and ES cells. Cell113:631–642
24. Jiang YH, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du JB, Aldrich S,Lisberg A, Low WC, Largaespada DA, Verfaillie CM 2002 Pluripotency ofmesenchymal stem cells derived from adult marrow. Nature 418:41–49
25. Lendahl U, Zimmerman LB, Mckay RDG 1990 CNS stem cells express a newclass of intermediate filament protein. Cell 60:585–595
26. Park IK, Qian DL, Kiel M, Becker MW, Pihalja M, Weissman IL, MorrisonSJ, Clarke MF 2003 Bmi-1 is required for maintenance of adult self-renewinghaematopoietic stem cells. Nature 423:302–305
27. Molofsky AV, Pardal R, Iwashita T, Park IK, Clarke MF, Morrison SJ 2003Bmi-1 dependence distinguishes neural stem cell self-renewal from progenitorproliferation. Nature 425:962–967
28. Gaiano N, Fishell G 2002 The role of Notch in promoting glial and neural stemcell fates. Annu Rev Neurosci 25:471–490
29. Ho KS, Scott MP 2002 Sonic hedgehog in the nervous system: functions,modifications and mechanisms. Curr Opin Neurobiol 12:57–63
30. Hitoshi S, Alexson T, Tropepe V, Donoviel D, Elia AJ, Nye JS, Conlon RA,Mak TW, Bernstein A, van der Kooy D 2002 Notch pathway molecules areessential for the maintenance, but not the generation, of mammalian neuralstem cells. Genes Dev 16:846–858
31. de Santa Barbara P, Van den Brink GR, Roberts DJ 2003 Development anddifferentiation of the intestinal epithelium. Cell Mol Life Sci 60:1322–1332
32. Pinto D, Gregorieff A, Begthel H, Clevers H 2003 Canonical Wnt signals areessential for homeostasis of the intestinal epithelium. Genes Dev 17:1709–1713
33. Reya T, Duncan AW, Ailles L, Domen J, Scherer DC, Willert K, Hintz L,Nusse R, Weissman IL 2003 A role for Wnt signalling in self-renewal ofhaematopoietic stem cells. Nature 423:409–414
34. Alvarez-Buylla A, Lim DA 2004 For the long run: maintaining germinal nichesin the adult brain. Neuron 41:683–686
35. Dontu G, Jackson KW, McNicholas E, Kawamura MJ, Abdallah WM, WichaMS 2004 Role of Notch signaling in cell-fate determination of human mam-mary stem/progenitor cells. Breast Cancer Res 6:R605–R615
36. Liu BY, McDermott SP, Khwaja SS, Alexander CM 2004 The transformingactivity of Wnt effectors correlates with their ability to induce the accumulationof mammary progenitor cells. Proc Natl Acad Sci USA 101:4158–4163
37. Treier M, Gleiberman AS, O’Connell SM, Szeto DP, McMahon JA, McMa-hon AP, Rosenfeld MG 1998 Multistep signaling requirements for pituitaryorganogenesis in vivo. Genes Dev 12:1691–1704
38. Rosenfeld MG, Briata P, Dasen J, Gleiberman AS, Kioussi C, Lin CJ,O’Connell SM, Ryan A, Szeto DP, Treier M 2000 Multistep signaling andtranscriptional requirements for pituitary organogenesis in vivo. Recent ProgHorm Res 55:1–14
39. Treier M, O’Connell S, Gleiberman A, Price J, Szeto DP, Burgess R, ChuangPT, McMahon AP, Rosenfeld MG 2001 Hedgehog signaling is required forpituitary gland development. Development 128:377–386
40. Raetzman LT, Ross SA, Cook S, Dunwoodie SL, Camper SA, Thomas PQ
Chen et al. • Candidate Stem Cells in Adult Pituitary Endocrinology, September 2005, 146(9):3985–3998 3997

2004 Developmental regulation of Notch signaling genes in the embryonicpituitary: Prop1 deficiency affects Notch2 expression. Dev Biol 265:329–340
41. Sheng HZ, Zhadanov AB, Mosinger B, Fujii T, Bertuzzi S, Grinberg A, LeeEJ, Huang SP, Mahon KA, Westphal H 1996 Specification of pituitary celllineages by the lim homeobox gene lhx3. Science 272:1004–1007
42. Sheng HZ, Moriyama K, Yamashita T, Li H, Potter SS, Mahon KA, WestphalH 1997 Multistep control of pituitary organogenesis. Science 278:1809–1812
43. Japon MA, Rubinstein M, Low MJ 1994 In situ hybridization analysis ofanterior pituitary hormone gene expression during fetal mouse development.J Histochem Cytochem 42:1117–1125
44. Denef C, Maertens P, Allaerts W, Mignon A, Robberecht W, Swennen L,Carmeliet P 1989 Cell-to-cell communication in peptide target cells of anteriorpituitary. Methods Enzymol 168:47–71
45. Langouche L, Hersmus N, Papageorgiou A, Vankelecom H, Denef C 2004Melanocortin peptides stimulate prolactin gene expression and prolactin ac-cumulation in rat pituitary aggregate cell cultures. J Neuroendocrinol 16:695–703
46. Otto C, Dieck T, Bauer K 1996 Dipeptide uptake by adenohypophyseal fol-liculo-stellate cells. Am J Physiol 271:C210–C217
47. Vankelecom H, Seuntjens E, Hauspie A, Denef C 2003 Targeted ablation ofgonadotrophs in transgenic mice depresses prolactin but not growth hormonegene expression at birth as measured by quantitative mRNA detection.J Biomed Sci 10:805–812
48. Ito CY, Li CYJ, Bernstein A, Dick JE, Stanford WL 2003 Hematopoietic stemcell and progenitor defects in Sca-1/Ly-6A-null mice. Blood 101:517–523
49. Sornson MW, Wu W, Dasen JS, Flynn SE, Norman DJ, O’Connell SM,Gukovsky I, Carriere C, Ryan AK, Miller AP, Zuo L, Gleiberman AS,Andersen B, Beamer WG, Rosenfeld MG 1996 Pituitary lineage determinationby the Prophet of Pit-1 homeodomain factor defective in Ames dwarfism.Nature 384:327–333
50. Vankelecom H, Matthys P, Van Damme J, Heremans H, Billiau A, Denef C1993 Immunocytochemical evidence that S-100-positive cells of the mouseanterior pituitary contain interleukin-6 immunoreactivity. J Histochem Cyto-chem 41:151–156
51. Raetzman LT, Ward R, Camper SA 2002 Lhx4 and Prop1 are required for cell
survival and expansion of the pituitary primordia. Development 129:4229–4239
52. Bach I, Rhodes SJ, Pearse RV, Heinzel T, Gloss B, Scully KM, SawchenkoPE, Rosenfeld MG 1995 P-Lim, a LIM homeodomain factor, is expressedduring pituitary organ and cell commitment and synergizes with Pit-1. ProcNatl Acad Sci USA 92:2720–2724
53. Ward RD, Raetzman LT, Suh H, Stone BM, Nasonkin IO, Camper SA 2005Role of PROP1 in pituitary gland growth. Mol Endocrinol 19:698–710
54. Ikeda H, Yoshimoto T 1991 Developmental changes in proliferative activityof cells of the murine Rathke’s pouch. Cell Tissue Res 263:41–47
55. Nakamura Y, Sakakibara S, Miyata T, Ogawa M, Shimazaki T, Weiss S,Kageyama R, Okano H 2000 The bHLH gene Hes1 as a repressor of theneuronal commitment of CNS stem cells. J Neurosci 20:283–293
56. Douglas KR, Brinkmeier ML, Kennell JA, Eswara P, Harrison TA, Patri-anakos AI, Sprecher BS, Potok MA, Lyons RH, MacDougald OA, Camper SA2001 Identification of members of the Wnt signaling pathway in the embryonicpituitary gland. Mamm Genome 12:843–851
57. Brinkmeier ML, Potok MA, Cha KB, Gridley T, Stifani S, Meeldijk J, CleversH, Camper SA 2003 TCF and Groucho-related genes influence pituitarygrowth and development. Mol Endocrinol 17:2152–2161
58. Lewis R 2004 Dedifferentiation: more than reversing fate. Pinning down theorigins of a slippery cell state. Scientist 18:20
59. Kineman RD, Faught WJ, Frawley LS 1992 Steroids can modulate transdif-ferentiation of prolactin and growth hormone cells in bovine pituitary cultures.Endocrinology 130:3289–3294
60. Vidal S, Horvath E, Kovacs K, Cohen SM, Lloyd RV, Scheithauer BW 2000Transdifferentiation of somatotrophs to thyrotrophs in the pituitary of patientswith protracted primary hypothyroidism. Virchows Arch 436:43–51
61. Childs GV 2002 Development of gonadotropes may involve cyclic transdif-ferentiation of growth hormone cells. Arch Physiol Biochem 110:42–49
62. Horvath E, Kovacs K 2002 Folliculo-stellate cells of the human pituitary: a typeof adult stem cell? Ultrastruct Pathol 26:219–228
63. Simmons DM, Voss JW, Ingraham HA, Holloway JM, Broide RS, RosenfeldMG, Swanson LW 1990 Pituitary cell phenotypes involve cell-specific Pit-1mRNA translation and synergistic interactions with other classes of transcrip-tion factors. Genes Dev 4:695–711
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
3998 Endocrinology, September 2005, 146(9):3985–3998 Chen et al. • Candidate Stem Cells in Adult Pituitary
Copyright © 2022 FDOKUMEN




















