
Política Estatal Anticorrupción de Sinaloa (PEA), publicada en ...
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/jphotobiol
Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
The action of oxygen on chlorophyll fluorescence quenchingand absorption spectra in pea thylakoid membranes under the
steady-state conditions
Maciej Garstka *, Patrycja Nejman 1, Małgorzata Rosiak
Department of Metabolic Regulation, Institute of Biochemistry, Faculty of Biology, Warsaw University, Miecznikowa 1, PL-02-096 Warszawa, Poland
Received 19 December 2003; received in revised form 21 August 2004; accepted 24 August 2004
Available online 22 October 2004
Abstract
The effect of oxygen concentration on both absorption and chlorophyll fluorescence spectra was investigated in isolated pea thyl-
akoids at weak actinic light under the steady-state conditions. Upon the rise of oxygen concentration from anaerobiosis up to 412 lMa gradual absorbance increase around both 437 and 670 nm was observed, suggesting the disaggregation of LHCII and destacking of
thylakoids. Simultaneously, an increase in oxygen concentration resulted in a decline in the Chl fluorescence at 680 nm to about 60%
of the initial value. The plot of normalized Chl fluorescence quenching, F(�O2)/F(+O2), showed discontinuity above 275 lM O2,
revealing two phases of quenching, at both lower and higher oxygen concentrations. The inhibition of photosystem II by DCMU
or atrazine as well as that of cyt b6f by myxothiazol attenuated the oxygen-induced quenching events observed above 275 lM O2,
but did not modify the first phase of oxygen action. These data imply that the oxygen mediated Chl fluorescence quenching is par-
tially independent on non-cyclic electron flow. The second phase of oxygen-induced decline in Chl fluorescence is diminished in thyl-
akoids with poisoned PSII and cyt b6f activities and treated with rotenone or N-ethylmaleimide to inhibit NAD(P)H-plastoquinone
dehydrogenase. The data suggest that under weak light and high oxygen concentration the Chl fluorescence quenching results from
interactions between oxygen and PSI, cyt b6f and Ndh. On the contrary, inhibition of non-cyclic electron flow by antimycin A or
uncoupling of thylakoids by carbonyl cyanide m-chlorophenyl hydrazone did not modify the steady-state oxygen effect on Chl flu-
orescence quenching. The addition of NADH protected thylakoids against oxygen-induced Chl fluorescence quenching, whereas in
the presence of exogenic duroquinone the decrease in Chl fluorescence to one half of the initial level did not result from the oxygen
effect, probably due to oxygen action as a weak electron acceptor from PQ pool and an insufficient non-photochemical quencher. The
data indicate that mechanism of oxygen-induced Chl fluorescence quenching depends significantly on oxygen concentration and is
related to both structural rearrangement of thylakoids and the direct oxygen reduction by photosynthetic complexes.
� 2004 Elsevier B.V. All rights reserved.
Keywords: Absorption changes; NAD(P)H-plastoquinone dehydrogenase; Oxygen; Pea thylakoids; Steady-state chlorophyll fluorescence quenching;
Plastoquinone pool; PSII inhibition
1. Introduction
Changes in absorption and chlorophyll a (Chl) fluo-
rescence reflect changes in properties and structure of
1011-1344/$ - see front matter � 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jphotobiol.2004.09.001
* Corresponding author. Tel.: +48 22 5543215; fax: +48 22 5543221.
E-mail address: [email protected] (M. Garstka).1 Present address: Department of Pneumology, Medical University
of Warsaw, PL-02-097.
thylakoid membranes. In isolated chloroplasts Chl fluo-
rescence yield depends on: (i) equilibrium between redox
state of the first PSII quinone acceptor (QA) and plasto-
quinone (PQ) pool [1,2], (ii) the extent of thylakoid
membranes stacking [3,4], (iii) redistribution of excita-
tion energy between PS I and PSII [5], (iv) singlet–singletexcitation transfer to carotenoids [6], (v) LHCII aggre-
gation induced by the acidification of thylakoid lumen

80 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
and violaxanthin deepoxidation [7]. In addition, other
quenchers like membrane energization [7] and effect of
donor side of PSII (cf. [2]) have to be taken into consid-
eration. Moreover, the light-induced LHCII aggregation
may occur upon detaching of these complexes from PSII
[8], suggesting that fluorescence quenching takes placeindependently at both PSII and LHCII. It seems likely
that in thylakoid membranes the change in Chl fluores-
cence yield is the common event of stacking, photosys-
tems segregation, transthylakoid pH gradient and
LHCII aggregation [4]. In fact, these phenomena are
closely related to each other, e.g. dissagregation of
LHCII-containing microdomains is associated with
destacking of thylakoid membranes [4,9].The reduction/oxidation ratio of PQ pool effect on
chlorophyll fluorescence depends on: (i) non-cyclic elec-
tron flow from PSII to stromal acceptors as well as
molecular oxygen, (ii) cyclic electron transport around
PSI and (iii) chlororespiration, including dark reduction
of PQ by endogenous reductants and its oxidation by
molecular oxygen [10,11]. The existence of two different
pathways of cyclic electron flow was suggested [12]: (i)an antimycin-sensitive one, requiring probably ferre-
doxin and hypothetical ferredoxin-plastoquinone reduc-
tase (FQR) [13,14], and (ii) a route including the
chloroplast NAD(P)H-plastoquinone dehydrogenase
(Ndh) activity [12,14].
The direct oxygen reduction by PSI in intact chloro-
plasts, the Mehler reaction [15], is closely associated
with superoxide dismutase and ascorbate peroxidaseactivities, which detoxify superoxide and hydrogen
peroxide in sequential reactions producing
monodehydroascorbate (MDA) andwater [15,16]. Ferre-
doxin-dependent reduction of MDA by monodehydro-
ascorbate reductase completes these reactions in
water–water cycle, which is thought to involve 30% of to-
tal non-cyclic electron flow [16]. However, this reaction is
likely favored at extreme electron supply, i.e. at highlight, due to competition for other stromal electron
acceptors, e.g. NADP+ or oxaloacetate [17].
Reduction of oxygen in chloroplasts can be performed
not only at PSI acceptor side but probably also at PQ oxi-
dase [10,11]. It was suggested that oxidation of plastoqu-
inol under both dark and light conditions occurs via
chlororespiration pathway [10,18,19]. However, as chlo-
rorespiration complexes involved in PQ reduction havebeen recognized [11,13,14,20], the nature of enzyme par-
ticipating in PQ oxidation remains under debate
[10,18,21]. This plastid terminal oxidase (PTOX) activity
seems to be linked to both carotene desaturation and
chlororespiration [22,23]. Contribution of cyt b559[21,24] andperoxidase [18] to dark plastoquinol oxidation
has been suggested. Moreover, the non-enzymatic mech-
anism of PQ oxidation has been recently proposed [25].The relationship between a light-induced shrinkage
of chloroplasts, oxygen concentration and light quality
had been observed many years ago [26,27]. Recently,
following oxygen treatment of isolated pea chloroplasts
a decline in excitation of chlorophyll fluorescence
accompanied by an increase in absorbance around
435, 470 and 674 nm was observed [28]. Moreover,
the kinetics of light-induced chlorophyll a fluorescencequenching in aggregates of LHCII was sensitive to the
oxygen presence [29], while the violaxanthin photo-
isomerization at the weak light closely associated with
structural rearrangement of LHCII was detected exclu-
sively in the presence of oxygen [30].
The oxygen concentration in chloroplast suspension
is proportional to O2 partial pressure in atmosphere
and amounts 256–275 lM O2 [31,32], while in thylakoidlipid phase it is 20% higher than in water phase and
amounts to about 300 lMO2 [25,33]. Furthermore, oxy-
gen concentration, depending on photosynthetic activity
and environmental conditions, may control the distribu-
tion of electrons between the cyclic and noncyclic elec-
tron pathways [15,17,31]. Both participation of oxygen
in electron transport [15,17] and structural changes of
thylakoid membranes [15,28,30] have been proved, butthe relationships between these phenomena are far from
the complete understanding. Hence, the aim of the pre-
sent investigation was to study these two events by
measuring of both absorbance and chlorophyll fluores-
cence in isolated thylakoids at various oxygen concen-
trations increasing from anaerobiosis up to 412.5 lMO2 at weak light conditions. The experiments were per-
formed under the steady-state conditions, i.e. when thefinal equilibrium between oxygen concentration and
both PSII-driven reduction of PQ pool and the level of
reduced centers of photosynthetic complexes is estab-
lished. Hence, the fluorescence and absorbance changes
reflect the oxygen effect on membrane components un-
der weak light conditions which limit the rate of photo-
synthesis [15]. In view of the data presented one might
conclude that the oxygen-induced Chl fluorescencequenching is partially independent on non-cyclic elec-
tron flow and may be related to structural rearrange-
ment of thylakoid membranes. The contributions of
PSI, cyt b6f and Ndh to oxygen-induced quenching phe-
nomena under the steady-state conditions at weak light
are discussed.
2. Materials and methods
2.1. Plant material and thylakoids preparation
The pea (Pisum sativum L. cv. Baron) plants were
grown in perlite-containing pots in the controlled envi-
ronment at 24 �C/20 �C day/night under 16 h photope-
riod with irriadiance of 80 lmol m�2 s�1. Plants werefertilized with diluted Knop�s solution. Leaves of 14
days old seedlings were homogenized in 330 mM

M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92 81
sorbitol, 20 mM tricine–NaOH (pH 7.5), 15 mM NaCl,
4 mM MgCl2 and 40 mM ascorbate. The homogenate
was filtered and centrifuged at 2000g for 5 min to obtain
chloroplasts. Thylakoids were prepared from chloro-
plasts by osmotic shock in the medium containing 20
mM tricine–NaOH (pH 7.0), 15 mM NaCl and 4 mMMgCl2 and centrifugation at 6000g for 10 min. The thyl-
akoids were resuspended in the medium of 330 mM
sorbitol, 20 mM Hepes–NaOH (pH 7.0), 15 mM NaCl
and 4 mM MgCl2. Following the second centrifugation
at 6000g for 10 min the pellet was resuspended in the
above buffer and kept on ice in dark. Thylakoids were
always freshly prepared before experiments. Chloro-
phyll concentration was quantified after extraction with80% acetone [34].
2.2. Preparation of samples
Samples for spectroscopic assays were prepared in
buffer used for resuspending of thylakoids, but pH was
adjusted to 7.5. All procedures were performed at
25 �C in 4 ml closed cuvettes filled completely with reac-tion mixture. The thylakoids suspension and reagents
were added using Hamilton syringe through 0.5 mm
hole made in teflon plug. Anaerobic conditions were
achieved by the addition of 1 mM glucose, glucose oxi-
dase (18.4 U/ml) and catalase (3600 U/ml) to the nitro-
gen washed-medium 5 min before the thylakoids were
included. The increased concentrations of O2 were
achieved by sequential addition of appropriate amounts(2–8 ll) of 1% H2O2 under dark conditions. A relation-
ship between amount of H2O2 and O2 concentration was
estimated with Clark electrode. In control conditions no
artificial electron acceptors were added. Inhibitors and
electron acceptors were dissolved either in ethanol (anti-
mycin, atrazine, DCMU, DBMIB, myxothiazol, rote-
none) or in water (MV, NEM). No effect of ethanol
on absorbance and fluorescence values was observed atused ethanol concentration (up to 1%). DQ was reduced
to DQH2 following the treatment with NaHB4 immedi-
ately before use [35]. The inhibitors affected significantly
neither glucose oxidase and catalase activities nor the
oxygen concentration.
2.3. Determination of absorbance and fluorescence emis-
sion spectra under the steady-state conditions
Absorption spectra were measured with the use of
D-600 Beckman spectrophotometer whereas fluores-
cence excitation spectra were determined with the Shim-
adzu RF-5301PC spectrofluorometer. The scans were
recorded for each 0.1 and 0.2 nm and the mean accumu-
lation times were 300 and 70 s for absorbance and fluo-
rescence spectra, respectively. Fluorescence emissionspectra were recorded with the excitation and emission
slit set to 3 and 10 nm, respectively. Samples were illumi-
nated with probing light of spectrofluorometer at 435
nm (36 lmol m�2 s�2). Changes of fluorescence were
measured from 630 to 760 nm with magnetical stirring
to prevent thylakoids settling. For absorbance and fluo-
rescence measurements the thylakoids suspensions were
diluted to concentration of 6 lg Chl ml�1.Thylakoids were incubated for 2–8 min with indi-
cated oxygen concentration in darkness, i.e. under con-
ditions sufficient to establish the constant oxygen
concentration. Samples were scanned thrice for each
measurement, i.e. under conditions necessary to attain
the invariable absorbance or fluorescence level. Time
of illumination at relative weak light was sufficient to in-
duce a reversible Chl fluorescence quenching [8,30], butdid not cause the Chl degradation. No significant
changes in oxygen concentration during period of thyl-
akoid illumination were observed in agreement with pre-
vious observations [25,31]. The changes in oxygen
concentration related to both O2 evolution in PSII and
reduction by PSI or plastoquinone pool were negligible
[25]. The first and the second absorbance scans were car-
ried out to eliminate light-dependent changes in theabsorbance amplitude. The third scan, which did not
show significant changes in comparison with the second
scan, was used to estimate the oxygen action. In order to
evaluate fluorescence measurements the second and the
third scans were used to obtain an average maximum
emission spectrum of thylakoids. The differences in the
absorbance or fluorescence between the first scan and
subsequent one did not exceed 1.5% and 1% (at 437and 680 nm), respectively. This procedure made us able
to obtain a steady-state equilibrium between each oxy-
gen concentration and redox state of membrane compo-
nents at weak light illumination.
2.4. Analysis of absorbance and fluorescence changes
The spectrum for anaerobic conditions was taken as areference one in calculations of the difference spectra.
The maxima of fluorescence emission spectra at 680 nm
for anaerobiosis and appropriate oxygen concentration
were utilized for the calculation of the oxygen-induced
quenching curves. Ratio of fluorescence in the absence
and presence of oxygen, F(�O2)/F(+O2), as a function
of the O2 concentration was presented as a normalized
fluorescence quenching, according to the principle ofquenching analysis [2]. Linear segments of the plot were
calculated as the best fitted simple or pair of regression
lines. For samples treated with exogenous chemicals
the F(�O2) values were determined under anaerobic con-
ditions in the presence of appropriate agent.
Inhibitors were added to cuvettes under anaerobic
conditions following the first fluorescence measurement.
Oxygen treatment affected the spectral properties of glu-cose oxidase/catalase in the UV absorbance region,
while fluorescence emission spectra were free of

82 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
significant interferences. The corresponding difference
absorbance enzyme spectra were subtracted from the
thylakoid absorbance ones.
3. Results and discussion
3.1. Effect of oxygen concentration increase on thylakoid
absorbance spectra
As the increase of absorbance after transfer of chlo-
roplasts from anaerobic (6 nM O2) to air-saturation
conditions (�250 lM O2) was recently established [28],
in the present study a gradual elevation of thylakoidabsorption induced by increasing oxygen concentrations
from anaerobiosis to 412.5 lM O2 concentration was
investigated (Fig. 1(a)). Difference spectra of the scans
Fig. 1. Effect of oxygen concentration on difference absorbance spectra (a) an
in isolated pea thylakoids. (a) shows absorbance difference spectra for thylako
were normalized at 540 nm. Equilibrium with desidered oxygen concentratio
shows the absorbance difference spectrum recorded following restoration
concentration. Spectrum was normalized at 400 nm. The spectra are represe
(with anaerobic spectrum taken as the reference one)
yielded positive broad band at 437 with shoulders at
470 and 480 nm and positive peak at 670 nm, while a
small negative band was detected at around 700 nm.
The Chl a difference absorbance bands were shifted to
blue region in comparison with those at 439 and 681nm revealed in the anaerobic spectrum. However, ampli-
tudes of the Soret band transitions at 437 nm rised with
the oxygen concentration increase (Fig. 1(a)). The ab-
sence of negative bands around 436 and 680 nm, charac-
teristic for Chl a irreversible bleaching under strong
illumination at aerobic conditions in both PSII and
PSI reaction centers [36,37], suggests that in the presence
of oxygen (cf. Fig. 1(a)), the absorbance changes are notassociated with degradation of photosynthetic com-
plexes under the steady-state conditions. The small oxy-
gen-induced negative bands around 700 nm recorded for
(a)
(b)
d difference spectrum following restoration of anaerobic conditions (b)
ids sequentially treated with increasing oxygen concentrations. Spectra
n was reached in dark and was followed by scanning of the sample. (b)
of anaerobic conditions versus spectrum recorded at 250 lM O2
ntative for scans obtained for five separate experiments.

M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92 83
increased O2 concentration may be related to oxidation
of PSI reaction center [37]. However, this effect is fully
reversible following oxygen removal (Fig. 1(b)).
Oxygen-induced absorption changes might be effec-
tively reversed upon the decrease in O2 concentrations
from 250 lM to 15 nM following nitrogen and glucose(10 mM) treatment. The amplitude of difference spec-
trum of thylakoids (the second anaerobic–aerobic)
(Fig. 1(b)) was reversed in comparison with that for oxy-
gen-treated thylakoids in the Soret region of scans (Fig.
1(a)). However, significant red shifts at 442 nm were ob-
served. In the reversed spectrum both a wide positive
absorbance band between 520 and 640 nm and a sharp
negative one at 681 nm have appeared, whereas respec-tive symmetrical bands were not detected in the oxygen-
induced spectrum (Fig. 1(a)).
Stacking of thylakoids [4,28] and aggregation of iso-
lated LHCII [40] cause a decrease of absorbance in the
Soret region due to an increase in light scattering and
the flattening effect [41]. These scattering changes have
been related to formation of multiaggregates of LHCII
[40,41] and changes in intrinsic thylakoid membranestructure [4]. On the other hand, a weak light-induced
increase in absorption values at 400–520 and 690 nm
is accompanied by a simultaneous decrease in light scat-
tering associated with disaggregation of LHCII com-
plexes [30]. In this case the light-driven isomerization
of violaxanthin under aerobic conditions induces proba-
bly the conformational reorganization of LHCII [30].
An oxygen-induced increase of absorption (cf. Fig.1(a)) is possibly related to this event. The destacking
of thylakoids, also visible as an absorbance increase
[4,28] (cf. Fig. 1(a)), may occur in the aftermath of
light-induced and oxygen-dependent reorganization of
LHCII at trimeric level [9,30]. However, in contrast to
isolated LHCII, in thylakoids the participation of other
membrane components should be also considered. In
pea chloroplasts changes in the excitation spectrum fol-lowing oxygen-treatment are very similar to those in-
duced by Mg2+-depletion [28], suggesting that similarly
to the action of ions, oxygen may induce thylakoids
destacking resulting from a change in the surface mem-
branes potential [3,4,28]. Since oxygen is required for
LHCII kinase activation [42], the influence of oxygen
on LHCII phosphorylation and subsequent thylakoids
destacking is possible in intact chloroplasts.As shown in Fig. 1 following increase or decrease of
oxygen concentration the Chl a absorption bands of pea
thylakoids were reversibly shifted to blue or red, respec-
tively. Furthermore, after oxygen removal the reversion
of bands at 442 and 488 nm to the initial amplitude was
observed (cf. Fig. 1). Similarly, both the red shift of
bands at 680 and 490 nm and the decrease of absorbance
in the Soret region of scan have been observed after theaggregations of LHCII [41]. On the other hand, the
destacking of thylakoid membranes is associated with
the blue shift of bands at 681 and 439 nm [4,28]. Hence,
the red and blue shifts observed at various oxygen con-
centrations suggest changes in both degree of membrane
stacking and aggregation of LHCII complexes. How-
ever, the formed bands (positive at 540–620 and negative
at 681 nm) in the reversed spectrum (cf. Fig. 1(b)) sug-gest that restoration of anaerobic conditions caused
additional changes in thylakoid membranes in compari-
son with those induced by oxygen treatment (cf. Fig.
1(a)). Both increase in steady-state transmittance in the
range from 548 to 575 nm [4] and decrease in absorbance
around 520 nm [28] were observed under cation-depend-
ent moderate thylakoid destacking. Moreover, changes
in absorbance region between 505 and 550 nm are alsoknown to be associated with (i) formation of trans-
thylakoid pH gradient [15,26,39], (ii) electrochromic
shifts of light-harvesting pigments reflecting the trans-
thylakoid electric field [38] and (iii) aggregation of
LHCII [7,39,40]. Since in our experiments the relatively
weak light (36 lmol m�2 s�2) was used, it seems possible
that positive absorbance band between 520 and 640 nm
in reversed spectrum is associated with increasing stack-ing of thylakoid membranes. On the other hand, the
presence of negative band at 680 nm may be related to
positive charge accumulation on thylakoid membranes
[38], suggesting that restoration of anaerobic conditions
also caused changes in membrane surface potential [28].
The results indicate that at weak light and at wide range
of O2 concentrations (from 0.08 to 412.5 lM O2), the
oxygen action is related to substantial, partly reversiblerearrangement of thylakoid membranes, probably
resulting of both disaggregation of LHCII and destack-
ing of thylakoids.
3.2. Relationship between oxygen concentration and Chl
fluorescence quenching
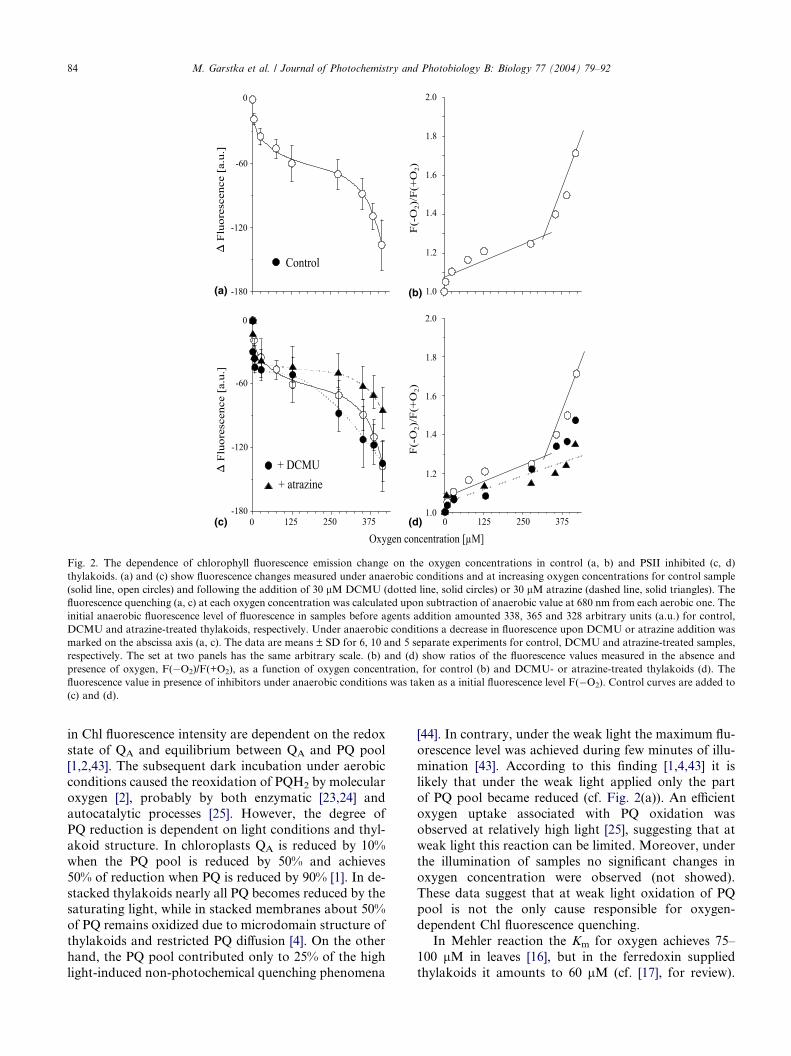
As shown in Fig. 2(a), with increasing oxygen con-centration the maximum fluorescence at 680 nm has de-
creased, while the quenching curve exhibited two phases.
At oxygen concentration equal to the 275 and 412 lMO2, the fluorescence decreased by 15% and 40%, respec-
tively. It should be noted that under the atmospheric
pressure the oxygen concentration at air saturated buffer
in 25 �C corresponds to 256 lM. Above this oxygen con-
centration the normalized fluorescence quenching,F(�O2)/F(+O2), showed the break point and the linear
phase of plot exhibiting slope 6-fold higher than the lin-
ear phase below this point (Fig. 2(b)). A similar decrease
in fluorescence at 680 nm was observed with excitation
at 455, 470 and 490 nm (data not shown).
Two phase dependence of Chl fluorescence on oxygen
concentration (cf. Fig. 2(a)) suggests diverse mecha-
nisms of quenching at low and high oxygen. In the ab-sence of exogenous electron acceptors during
illumination of thylakoids with a weak light, changes
(a) (b)
(c) (d)
Fig. 2. The dependence of chlorophyll fluorescence emission change on the oxygen concentrations in control (a, b) and PSII inhibited (c, d)
thylakoids. (a) and (c) show fluorescence changes measured under anaerobic conditions and at increasing oxygen concentrations for control sample
(solid line, open circles) and following the addition of 30 lM DCMU (dotted line, solid circles) or 30 lM atrazine (dashed line, solid triangles). The
fluorescence quenching (a, c) at each oxygen concentration was calculated upon subtraction of anaerobic value at 680 nm from each aerobic one. The
initial anaerobic fluorescence level of fluorescence in samples before agents addition amounted 338, 365 and 328 arbitrary units (a.u.) for control,
DCMU and atrazine-treated thylakoids, respectively. Under anaerobic conditions a decrease in fluorescence upon DCMU or atrazine addition was
marked on the abscissa axis (a, c). The data are means ± SD for 6, 10 and 5 separate experiments for control, DCMU and atrazine-treated samples,
respectively. The set at two panels has the same arbitrary scale. (b) and (d) show ratios of the fluorescence values measured in the absence and
presence of oxygen, F(�O2)/F(+O2), as a function of oxygen concentration, for control (b) and DCMU- or atrazine-treated thylakoids (d). The
fluorescence value in presence of inhibitors under anaerobic conditions was taken as a initial fluorescence level F(�O2). Control curves are added to
(c) and (d).
84 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
in Chl fluorescence intensity are dependent on the redox
state of QA and equilibrium between QA and PQ pool
[1,2,43]. The subsequent dark incubation under aerobicconditions caused the reoxidation of PQH2 by molecular
oxygen [2], probably by both enzymatic [23,24] and
autocatalytic processes [25]. However, the degree of
PQ reduction is dependent on light conditions and thyl-
akoid structure. In chloroplasts QA is reduced by 10%
when the PQ pool is reduced by 50% and achieves
50% of reduction when PQ is reduced by 90% [1]. In de-
stacked thylakoids nearly all PQ becomes reduced by thesaturating light, while in stacked membranes about 50%
of PQ remains oxidized due to microdomain structure of
thylakoids and restricted PQ diffusion [4]. On the other
hand, the PQ pool contributed only to 25% of the high
light-induced non-photochemical quenching phenomena
[44]. In contrary, under the weak light the maximum flu-
orescence level was achieved during few minutes of illu-
mination [43]. According to this finding [1,4,43] it islikely that under the weak light applied only the part
of PQ pool became reduced (cf. Fig. 2(a)). An efficient
oxygen uptake associated with PQ oxidation was
observed at relatively high light [25], suggesting that at
weak light this reaction can be limited. Moreover, under
the illumination of samples no significant changes in
oxygen concentration were observed (not showed).
These data suggest that at weak light oxidation of PQpool is not the only cause responsible for oxygen-
dependent Chl fluorescence quenching.
In Mehler reaction the Km for oxygen achieves 75–
100 lM in leaves [16], but in the ferredoxin supplied
thylakoids it amounts to 60 lM (cf. [17], for review).

M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92 85
The affinity of ferredoxin-free PSI preparation to oxy-
gen is high (Km is equal to 3–10 lM) (cf. [17], for re-
view), so under these conditions (cf. Fig. 2(a)) oxygen
reduction is not limited in isolated thylakoids. However,
the break point in normalized fluorescence quenching
(cf. Fig. 2(b)) is considerably above the cited Km values,suggesting that under the weak light the oxygen-induced
decrease in Chl fluorescence does not derive solely from
oxygen reduction by PSI.
It has been proposed that in chloroplasts the oxygen-
dependent decrease in Chl excitation [28] as well as exci-
tation and emission in LHCII aggregates [30] are related
to structural reorganization of thylakoid membranes and
LHCII trimer, respectively. These effects were observedfollowing the increase in oxygen concentration from 4.7
nM to 272 lMO2. Thylakoid membranes have a high
oxygen permeability [33], but the apoprotein of LHCII
may act as a barrier against O2 transport, protecting
Chl and xanthophylls associated with these complexes
[45]. On the contrary, in isolated LHCII the light-induced
decrease in fluorescence is more pronounced at increased
oxygen concentration, indicating that oxygen presenceenables light-induced isomerization of violoxanthin
followed by structural reorganization of LHCII trimer
[30]. Hence, the weak light-induced dissagregation of
aggregated LHCII [30] facilitates oxygen penetration to
complexes resulting in a chlorophyll fluorescence
quenching.
3.3. Effect of PSII inhibitors on oxygen-induced Chl
fluorescence quenching
To evaluate the contribution of PSII and PQ pool to
oxygen-induced fluorescence quenching we have per-
formed experiments in the presence of atrazine and
DCMU, i.e. under conditions when inhibitors block
Q�A oxidation by QB and prevent reduction of the PQ
pool [2,3]. The addition of inhibitors in the dark underanaerobic conditions followed by the period of continu-
ous illumination caused a decrease in maximum fluores-
cence level by about 5% and 8% for atrazine and
DCMU, respectively (Fig. 2(c)). DCMU quenching
action has been ascribed to a non-photochemical
quenching by oxidized PQ pool and might result from
the binding of plastoquinone molecule to the site other
than QB [2].As shown in Fig. 2(c), at saturated concentrations (30
lM) both atrazine and DCMU diminished the overall
oxygen action by about 30 and 50%, respectively. In
view of the normalized fluorescence quenching (Fig.
2(d)) it is clear that the second phase of steady-state
quenching, emerging above 275 lM oxygen concentra-
tion, is affected by atrazine/DCMU treatment. In thyl-
akoids with inhibited PSII (Fig. 2(d)), oxygenconcentration above 275 lM poorly influenced the fluo-
rescence quenching in contrast to quenching processes
observed in control thylakoids, in which electron dona-
tion to PQ pool and PSI was not prohibited (cf. Fig.
2(b)). These data imply the contribution of factors lo-
cated outside PSII to the second phase of Chl fluores-
cence quenching.
The PSII reaction center poisoned with atrazine/DCMU exists in Z+ P680 Phe Q�
A state and cannot acts
as fluorescence quencher [3]. In this case the Chl fluores-
cence yield may be regulated by structural changes in
thylakoid membranes related to the stacking phenome-
non [3] or Chl quenching in LHCII aggregates [7,8].
The hypothetical DCMU-insensitive oxidation of PSII
endogenous plastoquinone different from QA [2] and/
or PSII oxidation by PQ pool at low potential cyt b559[21] may contribute to oxygen-induced Chl fluorescence
quenching phenomenon. Moreover, at high oxygen con-
centration the difference between DCMU and atrazine
action (cf. Fig. 2(d)) may be related to alternative elec-
tron leakage from PSII [2,21]. The small oxygen uptake
observed in chloroplasts, thylakoids and isolated PSII
particles treated with DCMU [19,25] excludes participa-
tion of QB site in oxygen reduction and suggests that QA
is probably the site of electron donation to oxygen inside
PSII complex. The oxygen may react directly with both
QA producing superoxide [46] and non-haem iron cou-
pled with QA and QB co-factors [47]. Hence, in atra-
zine/DCMU-treated thylakoids oxygen-induced Chl a
fluorescence quenching at low oxygen concentration is
probably due to oxidation of P680 by alternative path-
ways [2,47] and/or structural rearrangements of thylak-oids (cf. Fig. 1(a)) [3,4,28,30].
3.4. Chl fluorescence quenching in view of competition
between exogenous quinones and oxygen
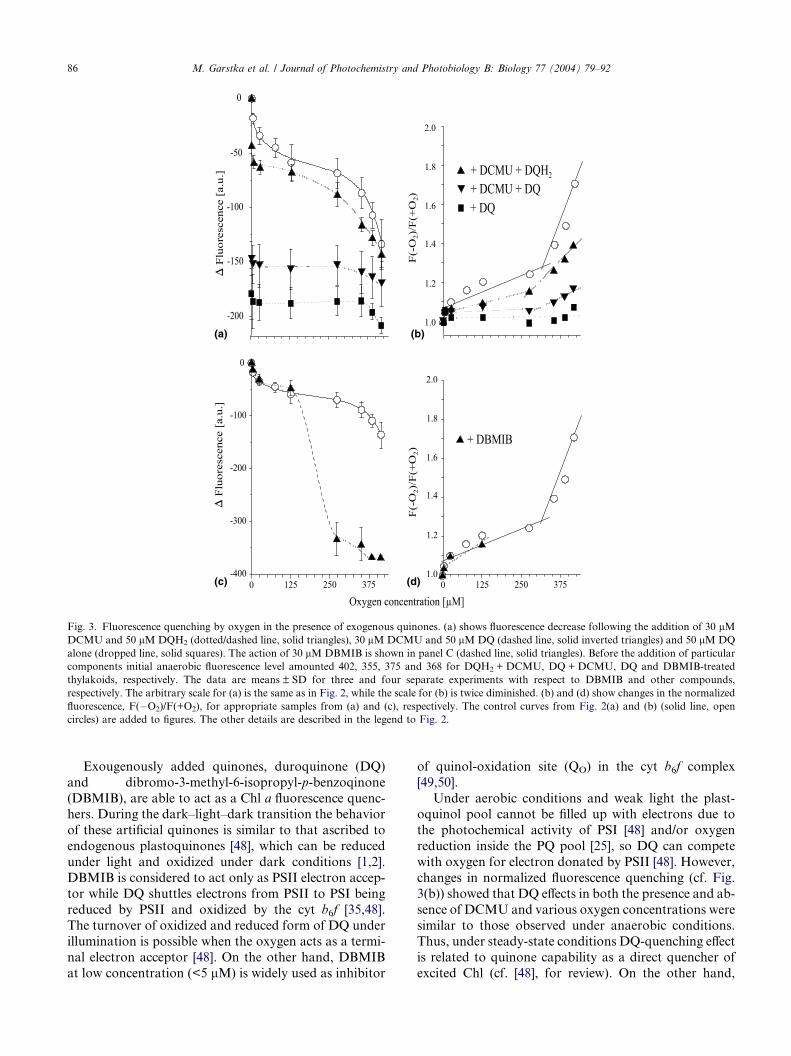
The addition of DQ in the presence of DCMU under
anaerobic condition caused decrease in fluorescence to
about 45% of its initial value. Moreover, the fluores-cence values were practically insensitive to increasing
oxygen concentrations (Fig. 3(a)). Upon the addition
of DQ alone, a decrease in Chl fluorescence under anaer-
obic conditions was higher (Fig. 3(a)) than overall oxy-
gen-induced fluorescence quenching (cf. Fig. 2(a)). In
presence or absence of DCMU the DQ action was sim-
ilar, indicating that the effect of 30 lM duroquinone on
fluorescence yield is independent on QA oxidation state.As shown in Fig. 3(c) DBMIB did not affect oxygen-
induced quenching up to 125 lM O2 (Figs. 3(c) and (d)),
indicating that inhibition of electron flow from PSII to
cyt b6f is negligible compared to oxygen action on Chl
fluorescence quenching. However, above 125 lM O2
concentration a sudden decline of Chl fluorescence was
observed. It should be noted that this effect is twice more
pronounced than the overall oxygen action in the ab-sence of electron acceptor (cf. Fig. 2(a)), and leads to
complete quenching of the Chl fluorescence.
(a) (b)
(c) (d)
Fig. 3. Fluorescence quenching by oxygen in the presence of exogenous quinones. (a) shows fluorescence decrease following the addition of 30 lMDCMU and 50 lM DQH2 (dotted/dashed line, solid triangles), 30 lM DCMU and 50 lM DQ (dashed line, solid inverted triangles) and 50 lM DQ
alone (dropped line, solid squares). The action of 30 lM DBMIB is shown in panel C (dashed line, solid triangles). Before the addition of particular
components initial anaerobic fluorescence level amounted 402, 355, 375 and 368 for DQH2 + DCMU, DQ + DCMU, DQ and DBMIB-treated
thylakoids, respectively. The data are means ± SD for three and four separate experiments with respect to DBMIB and other compounds,
respectively. The arbitrary scale for (a) is the same as in Fig. 2, while the scale for (b) is twice diminished. (b) and (d) show changes in the normalized
fluorescence, F(�O2)/F(+O2), for appropriate samples from (a) and (c), respectively. The control curves from Fig. 2(a) and (b) (solid line, open
circles) are added to figures. The other details are described in the legend to Fig. 2.
86 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
Exougenously added quinones, duroquinone (DQ)
and dibromo-3-methyl-6-isopropyl-p-benzoqinone
(DBMIB), are able to act as a Chl a fluorescence quenc-hers. During the dark–light–dark transition the behavior
of these artificial quinones is similar to that ascribed to
endogenous plastoquinones [48], which can be reduced
under light and oxidized under dark conditions [1,2].
DBMIB is considered to act only as PSII electron accep-
tor while DQ shuttles electrons from PSII to PSI being
reduced by PSII and oxidized by the cyt b6f [35,48].
The turnover of oxidized and reduced form of DQ underillumination is possible when the oxygen acts as a termi-
nal electron acceptor [48]. On the other hand, DBMIB
at low concentration (<5 lM) is widely used as inhibitor
of quinol-oxidation site (QO) in the cyt b6f complex
[49,50].
Under aerobic conditions and weak light the plast-oquinol pool cannot be filled up with electrons due to
the photochemical activity of PSI [48] and/or oxygen
reduction inside the PQ pool [25], so DQ can compete
with oxygen for electron donated by PSII [48]. However,
changes in normalized fluorescence quenching (cf. Fig.
3(b)) showed that DQ effects in both the presence and ab-
sence of DCMU and various oxygen concentrations were
similar to those observed under anaerobic conditions.Thus, under steady-state conditions DQ-quenching effect
is related to quinone capability as a direct quencher of
excited Chl (cf. [48], for review). On the other hand,

M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92 87
above 350 lMO2, the oxygen effects on Chl fluorescence
were poorly visible in the presence of DQ, indicating that
DQ prevents the oxygen-induced quenching action due
to higher affinity for excited Chl molecules in comparison
with that for oxygen. Exogenous duroquinol (DQH2) is
an artificial electron donor to PQ pool [35,43] and cantransfer electrons to PSI under illumination [48]. How-
ever, as shown in Fig. 3(b) the effect of simultaneously
added DCMU and DQH2 on oxygen-induced fluores-
cence quenching was not significantly different from that
of DCMU alone (cf. Fig. 1(b)). Thus, it is likely that
reduction of membrane components by DQH2 is rather
of little importance for steady-state oxygen effect in
poisoned thylakoids.A significant effect of DBMIB on Chl fluorescence
may be associated with diaphorase activity of FNR,
which transfers electrons from PQ pool to DBMIB
being subsequently reoxidised by O2 with Km amounted
to 16 lM [49]. Since the role of DBMIB in quenching
phenomena is determined by its photoreduction [48],
the reoxidation of DBMIB by oxygen may prevent the
exhaustion of the quinone and increase the DBMIB effi-cacy on Chl fluorescence quenching. On the other hand,
DBMIB did not affect the fluorescence under anaerobic
conditions (cf. Fig. 3(c)), suggesting that a direct reduc-
tion of DBMIB at QB site of PSII can be ruled out [51].
Thus, it is likely that oxygen above 125 lM plays a sec-
ondary role in Chl fluorescence quenching, essentially
increasing the DBMIB action. Below 125 lM oxygen,
the Chl fluorescence quenching does not result fromcompetition between DBMIB and oxygen.
3.5. Oxygen effect on uncoupled thylakoids
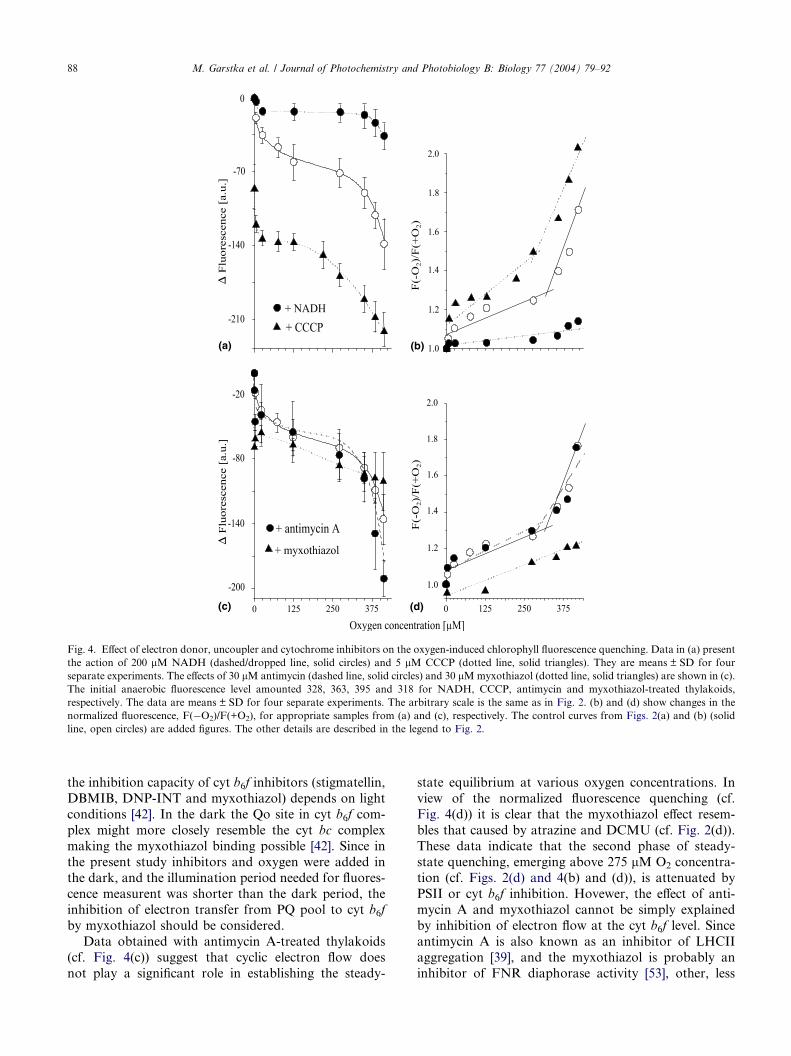
As shown in Fig. 4(a) in a presence of 5 lM CCCP, a
lipid soluble proton carrier, Chl fluorescence was de-
creased by about 20% under anaerobic conditions, indi-
cating the CCCP as efficient Chl fluorescence quencher.Sequential addition of oxygen to uncoupled thylakoids
augmented the decline of the fluorescence quenching
by 40% (Fig. 4(a)) resulting in a decrease in the overall
fluorescence to 40% of the initial level. However, the
plot of oxygen-induced normalized fluorescence quench-
ing with the breakpoint above 275 lM O2 (Fig. 4(b)),
was not significantly different from the control one (cf.
Fig. 2(a)).Both the electron flow toward oxygen and cyclic elec-
tron transport around PSI, which can occur in isolated
thylakoids in the absence of artificial acceptors, increase
the transmembrane proton gradient [15,17]. The DpHplays a regulatory role in photosynthetic electron trans-
port at the level of PQ oxidation or the cyt b6f complex
[15,17]. FCCP, acting as a lipid soluble uncoupler, may
increase the proton availability within membranes, andenhance the dismutation of superoxide generated during
O2 reduction [47]. Moreover, the stimulation of oxygen
uptake by the methylamine and gramicidin D in the ab-
sence of artificial acceptors was observed [25]. The data
presented (cf. Fig. 4(a)) do not exclude the contribution
of oxygen to DpH formation [15,17], but obviously sug-
gest that under the weak light conditions uncoupling of
thylakoids did not influence the oxygen-induced fluores-cence quenching.
3.6. Attenuation of oxygen action in the presence of
NADH
As shown in Fig. 4(a), during the steady-state condi-
tions NADH did not influence the fluorescence level in
pea thylakoids under anaerobic conditions, while it effi-ciently attenuated the oxygen effect on Chl fluorescence
and almost completely prevented the quenching below
275 lM O2 (Fig. 4(b)). Moreover, at oxygen concentra-
tions increased up to 412.5 lM O2 overall NADH utili-
zation (measured at 340 nm) did not exceed 10%. These
data are in agreement with no NADH oxidation in the
absence of exogenous acceptors in thylakoids under aer-
obic conditions [52].The NAD(P)H-dependent reduction of PQ pool
may be catalyzed by Ndh [11,20] and/or rotenone-in-
sensitive Ndh [43], whereas a direct participation of
FNR in PQ reduction is not clearly established
[49,52]. The addition of NADH to chloroplasts evokes
a fluorescence rise and reduces the PQ pool due to
chloroplastic dehydrogenase(s) activities [43]. This phe-
nomenom is more pronounced under anaerobic condi-tions suggesting that in the presence of oxygen a
significant part of electrons entering PQ pool is used
for O2 reduction [43]. Thus, the attenuating effect of
NADH on oxygen-induced Chl a fluorescence quench-
ing in isolated thylakoid membranes probably requires
the Ndh activity [43,52]. The successful prevention of
oxygen-induced Chl fluorescence quenching by NADH
under the weak light is understood in view of theobservation that in isolated intact chloroplasts oxygen
is a weak electron acceptor [17].
3.7. Effect of cytochrome inhibitors on oxygen-induced
Chl fluorescence quenching
As shown in Fig. 4(c), under anaerobic condition
both 30 lM antimycin A and 30 lM myxothiazol de-creased the Chl fluorescence level by 8% and 20%,
respectively. Antimycin A did not influence the oxy-
gen-induced Chl fluorescence decline, whereas the
myxothiazol diminished the overall oxygen action by
30% (Fig. 4(d)). Antimycin A is an efficient inhibitor
of cyclic electron flow at FQR site [13,14]. Both antimy-
cin A and myxothiazol inhibit the cyt bc pathway in the
mitochondrial respiration [14,50], while myxothiazol isgenerally considered as inefficient inhibitor of the cyt
b6f complex [50]. Recently, it has been reported that
(a) (b)
(c) (d)
Fig. 4. Effect of electron donor, uncoupler and cytochrome inhibitors on the oxygen-induced chlorophyll fluorescence quenching. Data in (a) present
the action of 200 lM NADH (dashed/dropped line, solid circles) and 5 lM CCCP (dotted line, solid triangles). They are means ± SD for four
separate experiments. The effects of 30 lM antimycin (dashed line, solid circles) and 30 lMmyxothiazol (dotted line, solid triangles) are shown in (c).
The initial anaerobic fluorescence level amounted 328, 363, 395 and 318 for NADH, CCCP, antimycin and myxothiazol-treated thylakoids,
respectively. The data are means ± SD for four separate experiments. The arbitrary scale is the same as in Fig. 2. (b) and (d) show changes in the
normalized fluorescence, F(�O2)/F(+O2), for appropriate samples from (a) and (c), respectively. The control curves from Figs. 2(a) and (b) (solid
line, open circles) are added figures. The other details are described in the legend to Fig. 2.
88 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
the inhibition capacity of cyt b6f inhibitors (stigmatellin,
DBMIB, DNP-INT and myxothiazol) depends on light
conditions [42]. In the dark the Qo site in cyt b6f com-
plex might more closely resemble the cyt bc complex
making the myxothiazol binding possible [42]. Since in
the present study inhibitors and oxygen were added inthe dark, and the illumination period needed for fluores-
cence measurent was shorter than the dark period, the
inhibition of electron transfer from PQ pool to cyt b6f
by myxothiazol should be considered.
Data obtained with antimycin A-treated thylakoids
(cf. Fig. 4(c)) suggest that cyclic electron flow does
not play a significant role in establishing the steady-
state equilibrium at various oxygen concentrations. In
view of the normalized fluorescence quenching (cf.
Fig. 4(d)) it is clear that the myxothiazol effect resem-
bles that caused by atrazine and DCMU (cf. Fig. 2(d)).
These data indicate that the second phase of steady-
state quenching, emerging above 275 lM O2 concentra-tion (cf. Figs. 2(d) and 4(b) and (d)), is attenuated by
PSII or cyt b6f inhibition. Hovewer, the effect of anti-
mycin A and myxothiazol cannot be simply explained
by inhibition of electron flow at the cyt b6f level. Since
antimycin A is also known as an inhibitor of LHCII
aggregation [39], and the myxothiazol is probably an
inhibitor of FNR diaphorase activity [53], other, less

M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92 89
specific, sites of inhibition may be responsible for these
phenomena.
3.8. Effect of inhibitors of chlororespiration pathways on
oxygen-induced Chl fluorescence quenching
As shown in Fig. 5(a), N-ethylmaleimide (NEM), an
inhibitor of sulfhydryl groups [56] and a potent inhibitor
of FNR and chloroplastic dehydrogenases [43] caused a
decrease in Chl fluorescence quenching at rather low (60
lM) concentration compared to that reported for inhi-
bition of NAD(P)H oxidation by FNR (3mM) or Ndh
(0.5mM) [43]. NEM did not influence the first phase of
Chl fluorescence quenching, whereas above 275 lM O2
the effect of oxygen was strongly diminished (Fig.
5(b)). The addition of 60 lM NEM in the presence of
30 lM DCMU under the steady-state conditions (Fig.
5A) attenuated the oxygen action in the same way as
NEM or DCMU alone (cf. Fig. 2(c)), indicating that
these inhibitors did not influence the Chl fluorescence
quenching evoked by oxygen concentration increased
up to 275 lM (cf. Fig. 2(b)). In illuminated thylakoids
(a) (b
(c) (d
Fig. 5. Influence of Ndh inhibitors on the oxygen-induced chlorophyll fluores
line, solid triangles) or 30 lM DCMU and 60 lM NEM (dashed line, solid
solid triangles). The initial anaerobic fluorescence level was 374, 383, 381 for
The data are means ± SD for four separate experiments. The arbitrary scale
fluorescence, F(�O2)/F(+O2), for appropriate samples from (a) and (c), res
circles) are added to figures. The other details are described in the legend to
NEM at milimolar concentrations is very effective elec-
tron acceptor from PQ pool [56]. However, at applied
low NEM concentration the acceptor properties of this
inhibitor are visible probably only under anaerobic con-
ditions, when NEM decreases the fluorescence level by
6% (Fig. 5(a)).As shown in Fig. 5(c) in the presence of 60 lM rote-
none, the inhibitor of mitochondrial complex I and chlo-
roplastic Ndh [43], oxygen caused the overall Chl
fluorescence decrease only by 15% in comparison with
40% estimated in control samples (cf Fig. 2(a)). Moreo-
ver, above 275 lM oxygen concentration the Chl fluo-
rescence quenching was completely diminished by
rotenone (Fig. 5(d)), suggesting the importance of Ndhcomplex in oxygen-induced quenching under the steady-
state conditions.
The effect of oxygen on the relationship between Chl
fluorescence and PQ pool oxidation has been studied
using the propyl gallate and SHAM, inhibitors of alter-
native oxidase [10,23]. Although the plastid terminal
oxidase (PTOX) is probably insensitive to SHAM and
sensitive to propyl gallate [10,23], these two inhibitors
)
)
cence quenching. Data in (a) present the action of 60 lMNEM (dotted
circles). The action of 60 lM rotenone is presented in (c) (dashed line,
NEN, NEM + DCMU and rotenone-treated thylakoids, respectively.
is the same as in Fig. 2. (b) and (d) show changes in the normalized
pectively. The control curves from Figs. 2(a) and (b) (solid line, open
Fig. 2.

90 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
did not affect the overall oxygen action (data not
shown). The PTOX displays the relatively low affinity
for oxygen [23], which may explain the insignificant ef-
fect of inhibitors on oxygen-induced Chl fluorescence
quenching under steady-state conditions.
Recently, it has been proposed that the oxidationof PQ pool by molecular oxygen may be performed
by: (i) autocatalytic reactions inside thylakoid mem-
branes [25], (ii) oxidation at low potential site of cyt
b559 [21] and (iii) reaction catalyzed by PTOX
[10,23]. Reduction of oxygen by reactions sequence
involving Ndh complex, peroxidase, superoxide dismu-
tase and Mehler reaction was proposed by Casano
et al. [18] in view of in vitro experiments with purifiedenzymes. Molecular interactions between FNR and
Ndh [54] as well as FNR and cyt b6f [55] suggest
the complex relationship between PQ pool and mem-
brane complexes. Hence, these findings and presented
results (cf. Figs. 4 and 5) imply participation of cyt
b6f, PSI and Ndh complex in Chl fluorescence quench-
ing at weak light and oxygen concentration above
275 lM.
4. Conclusions
The results presented in this investigation indicate
that:
(i) The structural changes in thylakoids membrane arecorrelated to oxygen concentration. At weak light
oxygen induces both disaggregation of LHCII and
destacking of thylakoids (cf. Fig. 1), probably due
to oxygen-dependent and light-driven structural
rearrangement of LHCII complexes [8,30]. The
oxygen-induced thylakoids reorganization does
not seem to be the only component responsible
for Chl fluorescence decrease.(ii) The Chl fluorescence quenching is related to oxygen
concentration and reveals two phases with the
break point above 275 lM oxygen concentration
(cf. Figs. 2(a) and (b) and 4(b) and (d)).
(iii) The oxygen-induced decrease in Chl fluorescence
appearing at low oxygen concentration does not
relate to both redox state of QA (cf. Figs. 2(c) and
(d)) and electron flow from PSII to cyt b6f (cf. Figs.4(c) and (d)).
(iv) At high oxygen concentration (above 275 lM O2)
Chl fluorescence decrease seems to be associated
with oxygen reduction by PSI, cyt b6f and/or Ndh
complex (cf. Figs. 4(c) and (d) and 5).
(v) Uncoupling of thylakoids by CCCP does not result
in modification of the steady-state oxygen effect on
Chl fluorescence quenching (cf. Figs. 4(a) and (b)),while oxygen action may be modulated by electron
donors and acceptors, NADH (cf. Fig. 4(a)), DQ
(cf. Fig. 3(a)) and DBMIB (cf. Fig. 3(b)), exhibiting
a higher affinity to membrane complexes than
oxygen.
5. Abbreviations
CCCP carbonyl cyanide m-chlorophenyl hydra-
zone
Chl chlorophyll
DBMIB 2,5-dibromomethyl-6-isopropyl-p-benzo-
quinone
DCMU 3-(3,4-dichlorphenyl)1,1-dimethylurea
DNP-INT dinitrophenylether of 2-iodoo-4-nitrothy-mol
DQ duroquinone
FNR ferredoxin-NADP+-reductase
FQR ferredoxin-plastoquinone reductase
LHCII light harvesting chlorophyll a/b protein
complex II
Ndh NAD(P)H-plastoquinone dehydrogenase
NEM N-ethylmaleimidePQ plastoquinone
PSI, PSII photosystem I, II
SHAM salicylhydroxamic acid
Acknowledgements
This work was partly supported by the grants of Pol-
ish Ministry of Scientific Research and Information
Technology (No. 1455/15 and No. 3 P04C 109 23).
The authors are grateful to Professor J. Bryła for a crit-
ical reading of manuscript. We thank to Dr. H.Grubek-
Jaworska for making available the absorption spectro-
photometer. The assistance of Mrs. A. Dro _zak in
experiments with oxidase inhibitors is acknowledged.
References
[1] Q.J. Groom, D.M. Kramer, A.R. Crofts, D.R. Ort, The non-
photochemical reduction of plastoquinone in leaves, Photosynth.
Res. 36 (1993) 205–215.
[2] J. Kurreck, R. Schodel, G. Renger, Investigation of the plasto-
quinone pool size and fluorescence quenching in thylakoid
membranes and Photosystem II (PSII) membrane fragments,
Photosynth. Res. 63 (2000) 171–182.
[3] A. Jajoo, S. Bharti, Govindjee, Inorganic anions induce state
changes in spinach thylakoid membranes, FEBS Lett. 434 (1998)
193–196.
[4] H. Kirchhoff, S. Horstmann, E. Weis, Control of the photosyn-
thetic electron transport by PQ diffusion microdomains in
thylakoids of higher plants, Biochim. Biophys. Acta 1459 (2000)
148–168.
[5] Ch. Lunde, P.E. Jensen, A. Haldrup, J. Knoetzel, H.V. Scheller,
The PSI-H subunit of photosystem I is essential for state
transitions in plant photosynthesis, Nature 408 (2000) 613–615.

M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92 91
[6] A.J. Young, H.A. Frank, Energy transfer reactions involving
carotenoids: quenching of chlorophyll fluorescence, J. Photochem.
Photobiol. B 36 (1996) 3–15.
[7] P. Horton, A.V. Ruban, R.G. Walters, Regulation of light
harvesting in green plants. Indication by nonphotochemical
quenching of chlorophyll fluorescence, Plant Physiol. 106 (1994)
415–420.
[8] W.I. Gruszecki, W. Grudzinski, M. Matuła, P. Kernen, Z. Krupa,
Light-induced excitation quenching and structural transition in
light-harvesting complex II, Photosynth. Res. 59 (1999) 175–185.
[9] G. Garab, L. Mustardy, Role of LHCII-containing macrodo-
mains in the structure, function and dynamics of grana, Aust. J.
Plant Physiol. 26 (1999) 649–658.
[10] L. Cournac, K. Redding, J. Ravenel, D. Rumeau, E.-M. Josse,
M. Kuntz, G. Peltier, Electron flow between Photosystem II and
oxygen in chloroplasts of Photosytem I-deficient algae is mediated
by a quinol oxidase involved in chlororespiration, J. Biol. Chem.
275 (2000) 17256–17262.
[11] P.J. Nixon, Chlororespiration, Philos. Trans. R. Soc. Lond. B 355
(2000) 1541–1547.
[12] D.S. Bendall, R.S. Manasse, Cyclic photophosphorylation and
electron transport, Biochim. Biophys. Acta 1229 (1995) 23–38.
[13] T. Endo, T. Shikanai, F. Sato, K. Asada, NAD(P)H dehydrog-
enase-dependent, antimycin A-sensitive electron donation to
plastoquinone in tobacco chloroplasts, Plant Cell Physiol. 39
(1998) 1226–1231.
[14] T. Joet, L. Cournac, E.M. Horvath, P. Medgyesy, G. Peltier,
Increased sensitivity to Antimycin A induced by inactivation of
the chloroplasts ndhB gene. Evidence for participation of the
NADH-dehydrogenase complex to cyclic electron flow around
Photosystem I, Plant Physiol. 125 (2001) 1919–1929.
[15] U. Heber, Irrungen, Wirrungen? The Mehler reaction in relation
to cyclic electron transport in C3 plants, Photosynth. Res. 73
(2002) 223–231.
[16] K. Asada, The water–water cycle in chloroplasts: scavenging of
active oxygen and dissipation of excess photons, Annu. Rev.
Plant. Physiol. Plant. Mol. Biol. 50 (1999) 601–639.
[17] J.E. Backhausen, C. Kitzmann, P. Horton, R. Scheibe, Electron
acceptors in isolated intact spinach chloroplasts act hierarchically
to prevent over-reduction and competition for electrons, Pho-
tosynth. Res. 64 (2000) 1–13.
[18] L.M. Casano, J.M. Zapata, M. Martin, B. Sabater, Chlororespi-
ration and poising of cyclic electron transport, J. Biol. Chem. 275
(2000) 942–948.
[19] W.I. Gruszecki, K.P. Bader, G.H. Schmid, Light-induced oxygen
uptake in tobacco chloroplasts explained in terms of chlorores-
piratory activity, Biochim. Biophys. Acta 1188 (1994) 335–338.
[20] L.A. Sazanov, P.A. Burrows, P.J. Nixon, The plastid ndh genes
code for an NADH-specific dehydrogenase: isolation of a complex
I analogue from pea thylakoid membranes, Proc. Natl. Acad. Sci.
USA 95 (1998) 1319–1324.
[21] J. Kruk, K. Strzałka, Redox changes of cytochrome b559 in the
presence of plastoquinone, J. Biol. Chem. 276 (2001) 86–91.
[22] P. Carol, M.A. Kuntz, Plastid terminal oxidase comes to light:
implications for carotenoid biosynthesis and chlororespiration,
Trends Plant Sci. 6 (2001) 31–36.
[23] P. Bennoun, The present model for chlororespiration, Photosynth.
Res. 73 (2002) 273–277.
[24] J. Kruk, K. Strzałka, Dark reoxidation of the plastoquinone-pool
is mediated by low-potential form of cytochrome b559 in spinach
thylakoids, Photosynth. Res. 62 (1999) 273–279.
[25] S.A. Khorobrykh, B.N. Ivanow, Oxygen reduction in a plasto-
quinone pool of isolated pea thylakoids, Photosynth. Res. 71
(2002) 209–219.
[26] U. Heber, Conformational changes of chloroplasts induced by
illumination of leaves in vivo, Biochim. Biophys. Acta 180 (1969)
302–319.
[27] G. Krause, The high-energy state of the thylakoid system as
indicated by chlorophyll fluorescence and chloroplast shrinkage,
Biochim. Biophys. Acta 292 (1973) 715–728.
[28] M. Garstka, A. Jagielski, Peroxidative reactions attenuate oxygen
effect on spectroscopic properties of isolated chloroplasts, J.
Photochem. Photobiol. B 64 (2001) 82–92.
[29] V. Barzda, M. Vengris, L. Valkunas, R. van Grondelle, H. van
Amerongen, Generation of fluorescence quenchers from the triplet
state of chlorophylls in the major Light-Harvesting Complex II
from green plants, Biochemistry 39 (2000) 10468–40477.
[30] W. Grudzinski, Z. Krupa, M. Garstka, W. Maksymiec, T.E.
Swartz, W.I. Gruszecki, Conformational rearrangements in light-
harvesting complex II accompanying light-induced chlorophyll a
fluorescence quenching, Biochim. Biophys. Acta 1554 (2002) 108–
117.
[31] A. Ligeza, A. Wisniewska, W.K. Subczynski, A.N. Tikhonov,
Oxygen production and consumption by chloroplasts in situ and
in vitro as studied with microscopic spin label probes, Biochim.
Biophys. Acta 1186 (1994) 201–208.
[32] H.M. Steiger, E. Beck, R. Beck, Oxygen concentration in isolated
chloroplasts during photosynthesis, Plant Physiol. 60 (1977) 903–
906.
[33] A. Ligeza, A.N. Tikhonov, J.S. Hyde, W.K. Subczynski, Oxygen
permeability of thylakoid membranes: electron paramagnetic
resonance spin labeling study, Biochim. Biophys. Acta 1365
(1998) 453–463.
[34] D.I. Arnon, Copper enzyme in isolated chloroplasts, Polyphe-
nolooxidase in beta vulgaris, Plant. Physiol. 24 (1949) 1–5.
[35] C. White, C.R.K. Chain, R. Malkin, Duroquinol as an electron
donor for chloroplast electron transfer reaction, Biochim. Bio-
phys. Acta 502 (1978) 127–137.
[36] N.P. Mishra, Ch. Francke, H.J. van Gorkom, D.F. Ghanotakis,
Destructive role of singlet oxygen during aerobic illumination of
the Photosystem II core complex, Biochim. Biophys. Acta 1186
(1994) 81–90.
[37] S. Rajagopal, N.G. Bukhov, R. Carpentier, Changes in the
structure of chlorophyll–protein complexes and excitation energy
during photoinhibitory treatment of isolated photosystem I
submembrane particles, J. Photochem. Photobiol. B 62 (2002)
194–200.
[38] P. Siffel, I. Hunalova, K. Rohacek, Light-induced quenching
of chlorophyll fluorescence at 77 K in leaves, chloroplasts
and Photosystem II particles, Photosynth. Res. 65 (2000)
219–229.
[39] A.V. Ruban, D. Rees, A. Pascal, P. Horton, Mechanism of DpH-
dependent dissipation of absorbed excitation energy by photo-
synthetic membranes. II. The relationship between LHCII aggre-
gation in vitro and qE in isolated thylakoids, Biochim. Biophys.
Acta 1102 (1992) 39–44.
[40] A.V. Ruban, F. Calkoen, S.L.S. Kwa, R. van Grondelle, P.
Horton, J.P. Dekker, Characterisation of LHC II in the aggre-
gated state by linear and circular dichroism spectroscopy,
Biochim. Biophys. Acta 1321 (1997) 61–70.
[41] V. Barzda, G. Garab, V. Gulbinas, L. Valkunas, Evidence for
long-range excitation energy migration in macroaggregates of the
chlorophyll a/b light-harvesting antenna complexes, Biochim.
Biophys. Acta 1273 (1996) 231–236.
[42] C.-X. Hou, E. Rintamaki, E.-M. Aro, Ascorbate-mediated LHCII
protein phosphorylation – LHCII kinase regulation in light and in
darkness, Biochemistry 42 (2003) 5828–5836.
[43] S. Corneille, L. Cournac, G. Guedeney, M. Havaux, G. Peltier,
Reduction of the plastoquinone pool by exogenous NADH and
NADPH in higher plant chloroplasts. Characterization of a
NAD(P)H-plastoquinone oxidoreductase activity, Biochim. Bio-
phys. Acta 1363 (1998) 59–69.
[44] B. Yaakoubd, R. Andersen, Y. Desjardins, G. Samson, Contri-
bution of the free oxidized and QB-bound plastoquinone mole-

92 M. Garstka et al. / Journal of Photochemistry and Photobiology B: Biology 77 (2004) 79–92
cules to the thermal phase of chlorophyll-a fluorescence, Pho-
tosynth. Res. 74 (2002) 251–257.
[45] D. Siefermann-Harms, A. Angerhofer, Evidence for an O2-barrier
in the light-harvesting chlorophyll-a/b–protein complex LHCII,
Photosynth. Res. 55 (1998) 83–94.
[46] R.E. Cleland, S.C. Grace, Voltametric detection of superoxide
production by photosystem II, FEBS Lett. 457 (1999) 348–352.
[47] J.H.A. Nugent, Photoreducible high spin iron electron paramag-
netic resonance signals in dark-adapted Photosystem II: are they
oxidised non-haem iron formed from interaction of oxygen with
PSII electron acceptors? Biochim. Biophys. Acta 1504 (2001) 288–
298.
[48] N.G. Bukhov, S. Govindachary, E.A. Egorova, R. Carpentier,
Interaction of exogenous quinones with membranes of higher
plant chloroplasts: modulation of quinone capacities as photo-
chemical and non-photochemical quenchers of energy in Photo-
system II during light–dark transitions, Biochim. Biophys Acta
1604 (2003) 115–123.
[49] M. Bojko, St. Wie�ckowski, NADPH and ferredoxin:NADP+
oxidoreductase-dependent reduction of quinones and their reox-
idation, Phytochemistry 50 (1999) 203–208.
[50] T.-X. Lee, S.U. Metzger, Y.S. Cho, J. Whitmarsh, T. Kallas,
Modification of inhibitor binding sites in the cytochrome bf
complex by directed mutagenesis of cytochrome b6 in Synecho-
coccus sp. PCC 7002, Biochim. Biophys. Acta 1504 (2001) 235–
247.
[51] K. Satoh, H. Koike, T. Ichimura, S. Katoh, Binding affinities of
benzoquinones to the QB site of Photosystem II in Synechococcus
oxygen-evolving preparation, Biochim. Biophys. Acta 1102 (1992)
45–52.
[52] H.B. Teicher, H.V. Scheller, The NAD(P)H dehydrogenase in
barley thylakoids is photoactivable and uses NADPH as well as
NADH, Plant Physiol. 117 (1998) 525–532.
[53] J. Andrews, H. Schmidt, E. Heinz, Interference of electron
transport inhibitors with desaturation of monogalactosyl diacyl-
glycerol in intact chloroplasts, Arch. Biochem. Biophys. 270
(1989) 611–622.
[54] G. Guedeney, S. Corneille, S. Cuine, G. Peltier, Evidence
for an association of ndhB, ndhJ gene products and
ferredoxin-NADP-reductase as components of chloroplastic
NAD(P)H dehydrogenase complex, FEBS Lett. 378 (1996)
277–280.
[55] H. Zhang, J.P. Whitelegge, W.A. Cramer, Ferredoxin:NADP+
oxidoreductase is a subunit of the chloroplast cytochrome b6f
complex, J. Biol. Chem. 279 (2001) 38159–38165.
[56] B. Ivanov, A. Ignat�ev, S. Khorobrykh, Maleimides stimulate
oxygen reduction in illuminated thylakoids, FEBS Lett. 532 (2002)
193–197.
Copyright © 2022 FDOKUMEN




















