
The 19-kDa antigen of Mycobacterium tuberculosis is a major adhesin that binds the mannose receptor...
11
The 19-kDa antigen of Mycobacterium tuberculosis is a major adhesin that binds the mannose receptor of THP-1 monocytic cells and promotes phagocytosis of mycobacteria Hugo Diaz-Silvestre, Patricia Espinosa-Cueto, Alejandro Sanchez-Gonzalez, Miguel A. Esparza-Ceron, Ana Laura Pereira-Suarez, German Bernal-Fernandez, Clara Espitia, Raul Mancilla * Departamento de Inmunologia, Instituto de Investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico, Apartado Postal 70-228, 04510 Mexico, DF, Mexico Received 6 October 2003; received in revised form 11 June 2005; accepted 20 June 2005 Abstract Identification of mycobacterial adhesins is needed to understand better the pathogenesis of tuberculosis and to develop new strategies to fight this infection. In this work, THP-1 monocytic cells were incubated with Mycobacterium tuberculosis culture filtrate proteins labelled with biotin and a dominant 19-kDa adhesin was found. This adhesin was characterized as the glycosylated and acylated 19-kDa antigen (Rv 3763). These findings were confirmed in assays with culture filtrate proteins and cell-wall fractions from a recombinant Mycobacterium smegmatis strain that overexpresses the 19-kDa antigen. Further, fluorescent microspheres coated with recombinant culture filtrate proteins adhere to cells in higher numbers than microspheres coated with native M. smegmatis proteins. The binding of the 19-kDa antigen to cells was inhibited with mannose receptor competitor sugars, Ca 2C chelators and with a monoclonal antibody to the human mannose receptor. Phagocytosis assays showed high-level binding of bacilli to THP-1 cells that was inhibited with a-methyl-mannoside, mannan, EDTA and mAbs to the mannose receptor and to the 19-kDa M. tuberculosis antigen. Immunoprecipitation, cell-surface ELISA and immunostaining confirmed the expression of the mannose receptor by THP-1 cells. In conclusion, here we show that the macrophage mannose receptor, considered a pathogen pattern recognition receptor, may interact with mannose residues of mycobacterial glycoproteins that could promote the phagocytosis of mycobacteria. q 2005 Elsevier Ltd. All rights reserved. Keywords: Mycobacterium tuberculosis; 19-kDa antigen; Mannose receptor; Immunoprecipitation; Macrophages; Phagocytosis 1. Introduction Mycobacterium tuberculosis is an intracellular pathogen that infects preferentially the lungs. The initial event in the pathogenesis of tuberculosis (TB) is the adherence of bacilli to host alveolar macrophages. For this, bacilli interact with a variety of macrophage surface receptors including comp- lement receptors CR1, CR3, and CR4 [1–3] and the mannose receptor [2,4,5]. Receptors for the surfactant protein A [6,7], the CD14 glycoprotein [8] and scavenger receptors [9] have also been implicated in cell infection. Little is known on the role of mycobacterial ligands (adhesins) in the adherence of bacilli to macrophages. LAM, a cell-wall lipoglycan, interacts with macrophages promoting phagocytosis of bacilli [5,10,11]. There are few reports describing myco- bacterial proteins with an adhesin function. Non-invasive E. coli transfected with M. tuberculosis DNA encoding a 50-kDa peptide acquires the ability to invade epithelial cells [12]. It has been reported that macrophages ingest microbe- ads coated with the antigen 85 complex [13] and that the mycobacterial heparin-binding-hemaglutinin binds epi- thelial cells and macrophages [14]. The receptors involved in the binding of these putative adhesins and their participation in the uptake of bacilli by human macrophages are not documented. Microbial Pathogenesis 39 (2005) 97–107 www.elsevier.com/locate/micpath 0882-4010/$ - see front matter q 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.micpath.2005.06.002 * Corresponding author. Tel.: C52 55 5622 3868; fax: C52 55 5622 3369. E-mail address: [email protected] (R. Mancilla).
Transcript of The 19-kDa antigen of Mycobacterium tuberculosis is a major adhesin that binds the mannose receptor...

The 19-kDa antigen of Mycobacterium tuberculosis is a major adhesin that
binds the mannose receptor of THP-1 monocytic cells and promotes
phagocytosis of mycobacteria
Hugo Diaz-Silvestre, Patricia Espinosa-Cueto, Alejandro Sanchez-Gonzalez,
Miguel A. Esparza-Ceron, Ana Laura Pereira-Suarez, German Bernal-Fernandez,
Clara Espitia, Raul Mancilla*
Departamento de Inmunologia, Instituto de Investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico,
Apartado Postal 70-228, 04510 Mexico, DF, Mexico
Received 6 October 2003; received in revised form 11 June 2005; accepted 20 June 2005
Abstract
Identification of mycobacterial adhesins is needed to understand better the pathogenesis of tuberculosis and to develop new strategies to
fight this infection. In this work, THP-1 monocytic cells were incubated with Mycobacterium tuberculosis culture filtrate proteins labelled
with biotin and a dominant 19-kDa adhesin was found. This adhesin was characterized as the glycosylated and acylated 19-kDa antigen (Rv
3763). These findings were confirmed in assays with culture filtrate proteins and cell-wall fractions from a recombinant Mycobacterium
smegmatis strain that overexpresses the 19-kDa antigen. Further, fluorescent microspheres coated with recombinant culture filtrate proteins
adhere to cells in higher numbers than microspheres coated with native M. smegmatis proteins. The binding of the 19-kDa antigen to cells
was inhibited with mannose receptor competitor sugars, Ca2C chelators and with a monoclonal antibody to the human mannose receptor.
Phagocytosis assays showed high-level binding of bacilli to THP-1 cells that was inhibited with a-methyl-mannoside, mannan, EDTA and
mAbs to the mannose receptor and to the 19-kDa M. tuberculosis antigen. Immunoprecipitation, cell-surface ELISA and immunostaining
confirmed the expression of the mannose receptor by THP-1 cells. In conclusion, here we show that the macrophage mannose receptor,
considered a pathogen pattern recognition receptor, may interact with mannose residues of mycobacterial glycoproteins that could promote
the phagocytosis of mycobacteria.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: Mycobacterium tuberculosis; 19-kDa antigen; Mannose receptor; Immunoprecipitation; Macrophages; Phagocytosis
1. Introduction
Mycobacterium tuberculosis is an intracellular pathogen
that infects preferentially the lungs. The initial event in the
pathogenesis of tuberculosis (TB) is the adherence of bacilli
to host alveolar macrophages. For this, bacilli interact with a
variety of macrophage surface receptors including comp-
lement receptors CR1, CR3, and CR4 [1–3] and the mannose
receptor [2,4,5]. Receptors for the surfactant protein A [6,7],
0882-4010/$ - see front matter q 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.micpath.2005.06.002
* Corresponding author. Tel.: C52 55 5622 3868; fax: C52 55 5622
3369.
E-mail address: [email protected] (R. Mancilla).
the CD14 glycoprotein [8] and scavenger receptors [9] have
also been implicated in cell infection. Little is known on the
role of mycobacterial ligands (adhesins) in the adherence of
bacilli to macrophages. LAM, a cell-wall lipoglycan,
interacts with macrophages promoting phagocytosis of
bacilli [5,10,11]. There are few reports describing myco-
bacterial proteins with an adhesin function. Non-invasive
E. coli transfected with M. tuberculosis DNA encoding a
50-kDa peptide acquires the ability to invade epithelial cells
[12]. It has been reported that macrophages ingest microbe-
ads coated with the antigen 85 complex [13] and that the
mycobacterial heparin-binding-hemaglutinin binds epi-
thelial cells and macrophages [14]. The receptors involved
in the binding of these putative adhesins and their
participation in the uptake of bacilli by human macrophages
are not documented.
Microbial Pathogenesis 39 (2005) 97–107
www.elsevier.com/locate/micpath
H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–10798
Considering that the identification and characterization
of mycobacterial adhesins could help to understand better
the pathogenesis of TB and to develop strategies aimed to
block initial events in the infection, we carried out the
present study. Our results show that the 19-kDa antigen of
M. tuberculosis (Rv 3763) binds THP-1 monocytic cells via
the macrophage mannose receptor (MR) promoting the
uptake of mycobacteria. These observations add importance
to the 19-kDa antigen, a mycobacterial moiety with
variegated biological effects including the induction of
immune responses [15–17], macrophage apoptosis [18] and
inhibition of Class II antigen presentation [19].
2. Results
2.1. The 19-kDa antigen of M. tuberculosis is a major
cell-wall located adhesin that binds THP-1 macrophage-
like cells
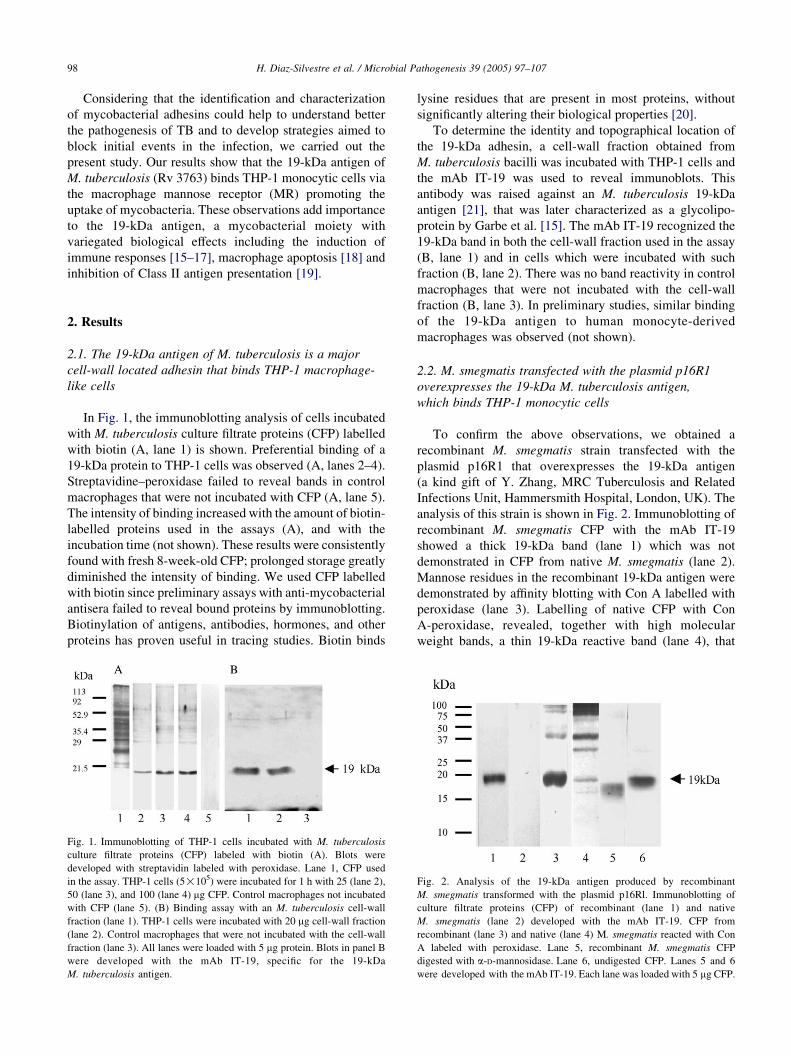
In Fig. 1, the immunoblotting analysis of cells incubated
with M. tuberculosis culture filtrate proteins (CFP) labelled
with biotin (A, lane 1) is shown. Preferential binding of a
19-kDa protein to THP-1 cells was observed (A, lanes 2–4).
Streptavidine–peroxidase failed to reveal bands in control
macrophages that were not incubated with CFP (A, lane 5).
The intensity of binding increased with the amount of biotin-
labelled proteins used in the assays (A), and with the
incubation time (not shown). These results were consistently
found with fresh 8-week-old CFP; prolonged storage greatly
diminished the intensity of binding. We used CFP labelled
with biotin since preliminary assays with anti-mycobacterial
antisera failed to reveal bound proteins by immunoblotting.
Biotinylation of antigens, antibodies, hormones, and other
proteins has proven useful in tracing studies. Biotin binds
Fig. 1. Immunoblotting of THP-1 cells incubated with M. tuberculosis
culture filtrate proteins (CFP) labeled with biotin (A). Blots were
developed with streptavidin labeled with peroxidase. Lane 1, CFP used
in the assay. THP-1 cells (5!105) were incubated for 1 h with 25 (lane 2),
50 (lane 3), and 100 (lane 4) mg CFP. Control macrophages not incubated
with CFP (lane 5). (B) Binding assay with an M. tuberculosis cell-wall
fraction (lane 1). THP-1 cells were incubated with 20 mg cell-wall fraction
(lane 2). Control macrophages that were not incubated with the cell-wall
fraction (lane 3). All lanes were loaded with 5 mg protein. Blots in panel B
were developed with the mAb IT-19, specific for the 19-kDa
M. tuberculosis antigen.
lysine residues that are present in most proteins, without
significantly altering their biological properties [20].
To determine the identity and topographical location of
the 19-kDa adhesin, a cell-wall fraction obtained from
M. tuberculosis bacilli was incubated with THP-1 cells and
the mAb IT-19 was used to reveal immunoblots. This
antibody was raised against an M. tuberculosis 19-kDa
antigen [21], that was later characterized as a glycolipo-
protein by Garbe et al. [15]. The mAb IT-19 recognized the
19-kDa band in both the cell-wall fraction used in the assay
(B, lane 1) and in cells which were incubated with such
fraction (B, lane 2). There was no band reactivity in control
macrophages that were not incubated with the cell-wall
fraction (B, lane 3). In preliminary studies, similar binding
of the 19-kDa antigen to human monocyte-derived
macrophages was observed (not shown).
2.2. M. smegmatis transfected with the plasmid p16R1
overexpresses the 19-kDa M. tuberculosis antigen,
which binds THP-1 monocytic cells
To confirm the above observations, we obtained a
recombinant M. smegmatis strain transfected with the
plasmid p16R1 that overexpresses the 19-kDa antigen
(a kind gift of Y. Zhang, MRC Tuberculosis and Related
Infections Unit, Hammersmith Hospital, London, UK). The
analysis of this strain is shown in Fig. 2. Immunoblotting of
recombinant M. smegmatis CFP with the mAb IT-19
showed a thick 19-kDa band (lane 1) which was not
demonstrated in CFP from native M. smegmatis (lane 2).
Mannose residues in the recombinant 19-kDa antigen were
demonstrated by affinity blotting with Con A labelled with
peroxidase (lane 3). Labelling of native CFP with Con
A-peroxidase, revealed, together with high molecular
weight bands, a thin 19-kDa reactive band (lane 4), that
Fig. 2. Analysis of the 19-kDa antigen produced by recombinant
M. smegmatis transformed with the plasmid p16Rl. Immunoblotting of
culture filtrate proteins (CFP) of recombinant (lane 1) and native
M. smegmatis (lane 2) developed with the mAb IT-19. CFP from
recombinant (lane 3) and native (lane 4) M. smegmatis reacted with Con
A labeled with peroxidase. Lane 5, recombinant M. smegmatis CFP
digested with a-D-mannosidase. Lane 6, undigested CFP. Lanes 5 and 6
were developed with the mAb IT-19. Each lane was loaded with 5 mg CFP.
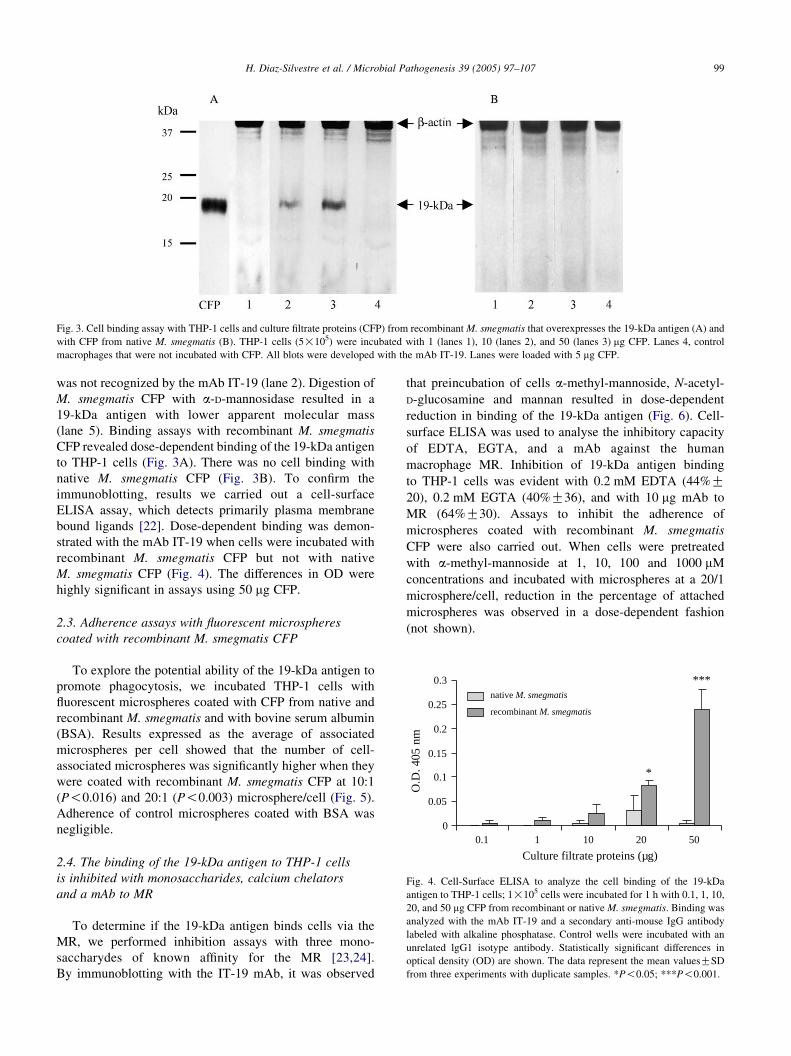
Fig. 3. Cell binding assay with THP-1 cells and culture filtrate proteins (CFP) from recombinant M. smegmatis that overexpresses the 19-kDa antigen (A) and
with CFP from native M. smegmatis (B). THP-1 cells (5!105) were incubated with 1 (lanes 1), 10 (lanes 2), and 50 (lanes 3) mg CFP. Lanes 4, control
macrophages that were not incubated with CFP. All blots were developed with the mAb IT-19. Lanes were loaded with 5 mg CFP.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.1 1 10 20 50
O.D
. 405
nm
***
*
native M. smegmatis
recombinant M. smegmatis
Culture filtrate proteins (µg)
Fig. 4. Cell-Surface ELISA to analyze the cell binding of the 19-kDa
antigen to THP-1 cells; 1!105 cells were incubated for 1 h with 0.1, 1, 10,
20, and 50 mg CFP from recombinant or native M. smegmatis. Binding was
analyzed with the mAb IT-19 and a secondary anti-mouse IgG antibody
labeled with alkaline phosphatase. Control wells were incubated with an
unrelated lgG1 isotype antibody. Statistically significant differences in
optical density (OD) are shown. The data represent the mean valuesGSD
from three experiments with duplicate samples. *P!0.05; ***P!0.001.
H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107 99
was not recognized by the mAb IT-19 (lane 2). Digestion of
M. smegmatis CFP with a-D-mannosidase resulted in a
19-kDa antigen with lower apparent molecular mass
(lane 5). Binding assays with recombinant M. smegmatis
CFP revealed dose-dependent binding of the 19-kDa antigen
to THP-1 cells (Fig. 3A). There was no cell binding with
native M. smegmatis CFP (Fig. 3B). To confirm the
immunoblotting, results we carried out a cell-surface
ELISA assay, which detects primarily plasma membrane
bound ligands [22]. Dose-dependent binding was demon-
strated with the mAb IT-19 when cells were incubated with
recombinant M. smegmatis CFP but not with native
M. smegmatis CFP (Fig. 4). The differences in OD were
highly significant in assays using 50 mg CFP.
2.3. Adherence assays with fluorescent microspheres
coated with recombinant M. smegmatis CFP
To explore the potential ability of the 19-kDa antigen to
promote phagocytosis, we incubated THP-1 cells with
fluorescent microspheres coated with CFP from native and
recombinant M. smegmatis and with bovine serum albumin
(BSA). Results expressed as the average of associated
microspheres per cell showed that the number of cell-
associated microspheres was significantly higher when they
were coated with recombinant M. smegmatis CFP at 10:1
(P!0.016) and 20:1 (P!0.003) microsphere/cell (Fig. 5).
Adherence of control microspheres coated with BSA was
negligible.
2.4. The binding of the 19-kDa antigen to THP-1 cells
is inhibited with monosaccharides, calcium chelators
and a mAb to MR
To determine if the 19-kDa antigen binds cells via the
MR, we performed inhibition assays with three mono-
saccharydes of known affinity for the MR [23,24].
By immunoblotting with the IT-19 mAb, it was observed
that preincubation of cells a-methyl-mannoside, N-acetyl-
D-glucosamine and mannan resulted in dose-dependent
reduction in binding of the 19-kDa antigen (Fig. 6). Cell-
surface ELISA was used to analyse the inhibitory capacity
of EDTA, EGTA, and a mAb against the human
macrophage MR. Inhibition of 19-kDa antigen binding
to THP-1 cells was evident with 0.2 mM EDTA (44%G20), 0.2 mM EGTA (40%G36), and with 10 mg mAb to
MR (64%G30). Assays to inhibit the adherence of
microspheres coated with recombinant M. smegmatis
CFP were also carried out. When cells were pretreated
with a-methyl-mannoside at 1, 10, 100 and 1000 mM
concentrations and incubated with microspheres at a 20/1
microsphere/cell, reduction in the percentage of attached
microspheres was observed in a dose-dependent fashion
(not shown).
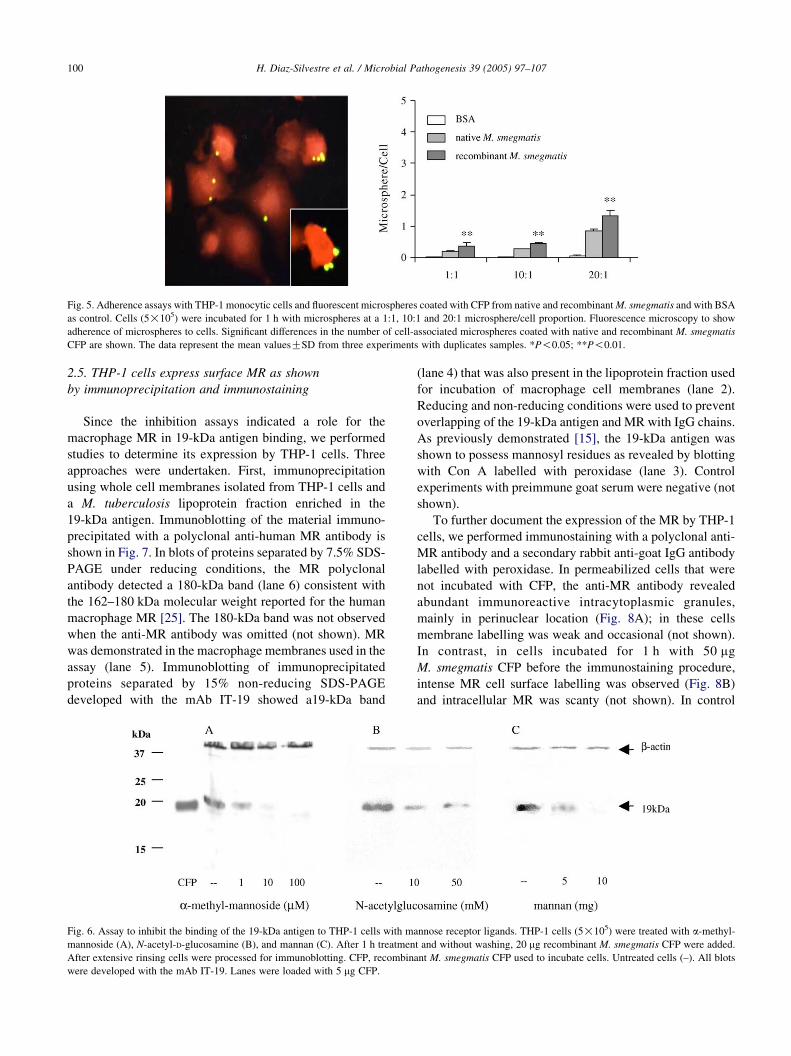
Fig. 5. Adherence assays with THP-1 monocytic cells and fluorescent microspheres coated with CFP from native and recombinant M. smegmatis and with BSA
as control. Cells (5!105) were incubated for 1 h with microspheres at a 1:1, 10:1 and 20:1 microsphere/cell proportion. Fluorescence microscopy to show
adherence of microspheres to cells. Significant differences in the number of cell-associated microspheres coated with native and recombinant M. smegmatis
CFP are shown. The data represent the mean valuesGSD from three experiments with duplicates samples. *P!0.05; **P!0.01.
H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107100
2.5. THP-1 cells express surface MR as shown
by immunoprecipitation and immunostaining
Since the inhibition assays indicated a role for the
macrophage MR in 19-kDa antigen binding, we performed
studies to determine its expression by THP-1 cells. Three
approaches were undertaken. First, immunoprecipitation
using whole cell membranes isolated from THP-1 cells and
a M. tuberculosis lipoprotein fraction enriched in the
19-kDa antigen. Immunoblotting of the material immuno-
precipitated with a polyclonal anti-human MR antibody is
shown in Fig. 7. In blots of proteins separated by 7.5% SDS-
PAGE under reducing conditions, the MR polyclonal
antibody detected a 180-kDa band (lane 6) consistent with
the 162–180 kDa molecular weight reported for the human
macrophage MR [25]. The 180-kDa band was not observed
when the anti-MR antibody was omitted (not shown). MR
was demonstrated in the macrophage membranes used in the
assay (lane 5). Immunoblotting of immunoprecipitated
proteins separated by 15% non-reducing SDS-PAGE
developed with the mAb IT-19 showed a19-kDa band
Fig. 6. Assay to inhibit the binding of the 19-kDa antigen to THP-1 cells with ma
mannoside (A), N-acetyl-D-glucosamine (B), and mannan (C). After 1 h treatmen
After extensive rinsing cells were processed for immunoblotting. CFP, recombin
were developed with the mAb IT-19. Lanes were loaded with 5 mg CFP.
(lane 4) that was also present in the lipoprotein fraction used
for incubation of macrophage cell membranes (lane 2).
Reducing and non-reducing conditions were used to prevent
overlapping of the 19-kDa antigen and MR with IgG chains.
As previously demonstrated [15], the 19-kDa antigen was
shown to possess mannosyl residues as revealed by blotting
with Con A labelled with peroxidase (lane 3). Control
experiments with preimmune goat serum were negative (not
shown).
To further document the expression of the MR by THP-1
cells, we performed immunostaining with a polyclonal anti-
MR antibody and a secondary rabbit anti-goat IgG antibody
labelled with peroxidase. In permeabilized cells that were
not incubated with CFP, the anti-MR antibody revealed
abundant immunoreactive intracytoplasmic granules,
mainly in perinuclear location (Fig. 8A); in these cells
membrane labelling was weak and occasional (not shown).
In contrast, in cells incubated for 1 h with 50 mg
M. smegmatis CFP before the immunostaining procedure,
intense MR cell surface labelling was observed (Fig. 8B)
and intracellular MR was scanty (not shown). In control
nnose receptor ligands. THP-1 cells (5!105) were treated with a-methyl-
t and without washing, 20 mg recombinant M. smegmatis CFP were added.
ant M. smegmatis CFP used to incubate cells. Untreated cells (–). All blots
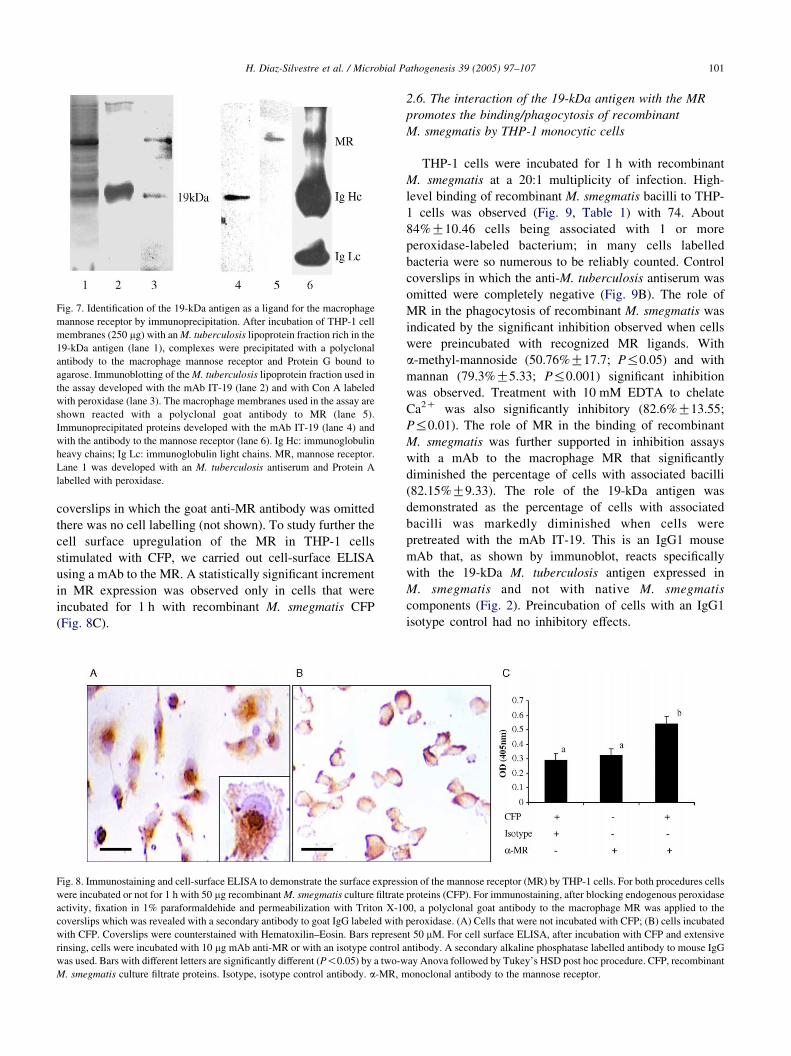
Fig. 7. Identification of the 19-kDa antigen as a ligand for the macrophage
mannose receptor by immunoprecipitation. After incubation of THP-1 cell
membranes (250 mg) with an M. tuberculosis lipoprotein fraction rich in the
19-kDa antigen (lane 1), complexes were precipitated with a polyclonal
antibody to the macrophage mannose receptor and Protein G bound to
agarose. Immunoblotting of the M. tuberculosis lipoprotein fraction used in
the assay developed with the mAb IT-19 (lane 2) and with Con A labeled
with peroxidase (lane 3). The macrophage membranes used in the assay are
shown reacted with a polyclonal goat antibody to MR (lane 5).
Immunoprecipitated proteins developed with the mAb IT-19 (lane 4) and
with the antibody to the mannose receptor (lane 6). Ig Hc: immunoglobulin
heavy chains; Ig Lc: immunoglobulin light chains. MR, mannose receptor.
Lane 1 was developed with an M. tuberculosis antiserum and Protein A
labelled with peroxidase.
H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107 101
coverslips in which the goat anti-MR antibody was omitted
there was no cell labelling (not shown). To study further the
cell surface upregulation of the MR in THP-1 cells
stimulated with CFP, we carried out cell-surface ELISA
using a mAb to the MR. A statistically significant increment
in MR expression was observed only in cells that were
incubated for 1 h with recombinant M. smegmatis CFP
(Fig. 8C).
Fig. 8. Immunostaining and cell-surface ELISA to demonstrate the surface express
were incubated or not for 1 h with 50 mg recombinant M. smegmatis culture filtrate
activity, fixation in 1% paraformaldehide and permeabilization with Triton X-10
coverslips which was revealed with a secondary antibody to goat IgG labeled with
with CFP. Coverslips were counterstained with Hematoxilin–Eosin. Bars represen
rinsing, cells were incubated with 10 mg mAb anti-MR or with an isotype control a
was used. Bars with different letters are significantly different (P!0.05) by a two-w
M. smegmatis culture filtrate proteins. Isotype, isotype control antibody. a-MR, m
2.6. The interaction of the 19-kDa antigen with the MR
promotes the binding/phagocytosis of recombinant
M. smegmatis by THP-1 monocytic cells
THP-1 cells were incubated for 1 h with recombinant
M. smegmatis at a 20:1 multiplicity of infection. High-
level binding of recombinant M. smegmatis bacilli to THP-
1 cells was observed (Fig. 9, Table 1) with 74. About
84%G10.46 cells being associated with 1 or more
peroxidase-labeled bacterium; in many cells labelled
bacteria were so numerous to be reliably counted. Control
coverslips in which the anti-M. tuberculosis antiserum was
omitted were completely negative (Fig. 9B). The role of
MR in the phagocytosis of recombinant M. smegmatis was
indicated by the significant inhibition observed when cells
were preincubated with recognized MR ligands. With
a-methyl-mannoside (50.76%G17.7; P%0.05) and with
mannan (79.3%G5.33; P%0.001) significant inhibition
was observed. Treatment with 10 mM EDTA to chelate
Ca2C was also significantly inhibitory (82.6%G13.55;
P%0.01). The role of MR in the binding of recombinant
M. smegmatis was further supported in inhibition assays
with a mAb to the macrophage MR that significantly
diminished the percentage of cells with associated bacilli
(82.15%G9.33). The role of the 19-kDa antigen was
demonstrated as the percentage of cells with associated
bacilli was markedly diminished when cells were
pretreated with the mAb IT-19. This is an IgG1 mouse
mAb that, as shown by immunoblot, reacts specifically
with the 19-kDa M. tuberculosis antigen expressed in
M. smegmatis and not with native M. smegmatis
components (Fig. 2). Preincubation of cells with an IgG1
isotype control had no inhibitory effects.
ion of the mannose receptor (MR) by THP-1 cells. For both procedures cells
proteins (CFP). For immunostaining, after blocking endogenous peroxidase
0, a polyclonal goat antibody to the macrophage MR was applied to the
peroxidase. (A) Cells that were not incubated with CFP; (B) cells incubated
t 50 mM. For cell surface ELISA, after incubation with CFP and extensive
ntibody. A secondary alkaline phosphatase labelled antibody to mouse IgG
ay Anova followed by Tukey’s HSD post hoc procedure. CFP, recombinant
onoclonal antibody to the mannose receptor.
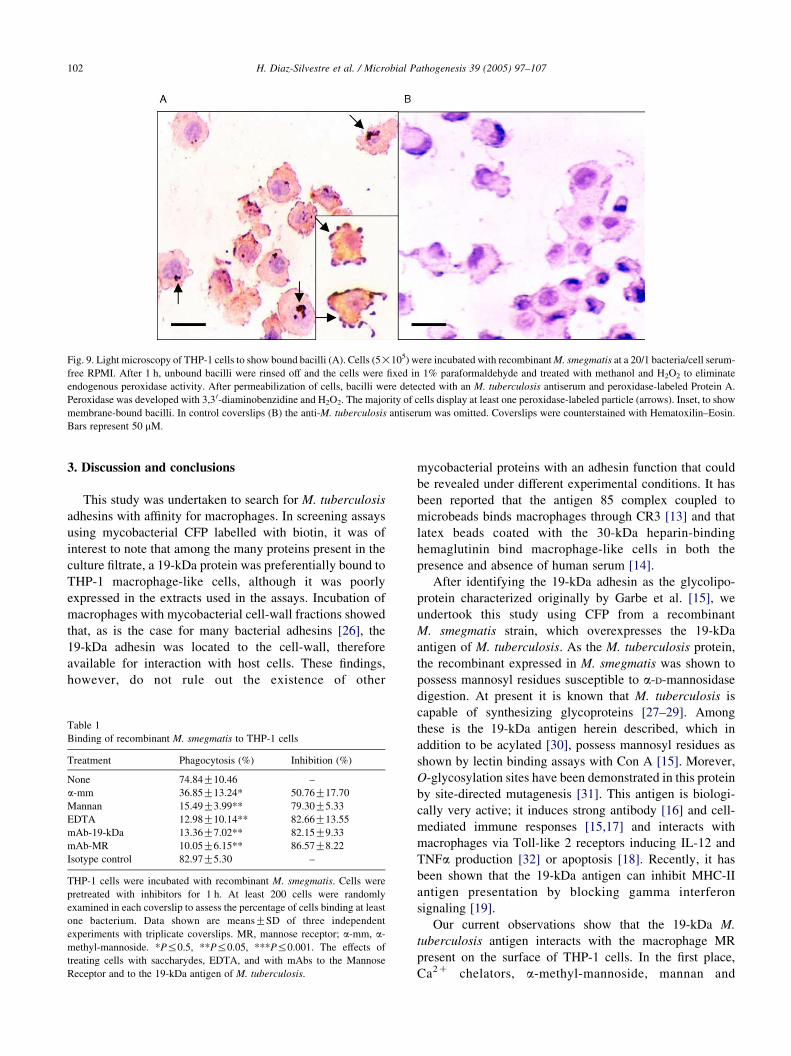
Fig. 9. Light microscopy of THP-1 cells to show bound bacilli (A). Cells (5!105) were incubated with recombinant M. smegmatis at a 20/1 bacteria/cell serum-
free RPMI. After 1 h, unbound bacilli were rinsed off and the cells were fixed in 1% paraformaldehyde and treated with methanol and H2O2 to eliminate
endogenous peroxidase activity. After permeabilization of cells, bacilli were detected with an M. tuberculosis antiserum and peroxidase-labeled Protein A.
Peroxidase was developed with 3,3 0-diaminobenzidine and H2O2. The majority of cells display at least one peroxidase-labeled particle (arrows). Inset, to show
membrane-bound bacilli. In control coverslips (B) the anti-M. tuberculosis antiserum was omitted. Coverslips were counterstained with Hematoxilin–Eosin.
Bars represent 50 mM.
H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107102
3. Discussion and conclusions
This study was undertaken to search for M. tuberculosis
adhesins with affinity for macrophages. In screening assays
using mycobacterial CFP labelled with biotin, it was of
interest to note that among the many proteins present in the
culture filtrate, a 19-kDa protein was preferentially bound to
THP-1 macrophage-like cells, although it was poorly
expressed in the extracts used in the assays. Incubation of
macrophages with mycobacterial cell-wall fractions showed
that, as is the case for many bacterial adhesins [26], the
19-kDa adhesin was located to the cell-wall, therefore
available for interaction with host cells. These findings,
however, do not rule out the existence of other
Table 1
Binding of recombinant M. smegmatis to THP-1 cells
Treatment Phagocytosis (%) Inhibition (%)
None 74.84G10.46 –
a-mm 36.85G13.24* 50.76G17.70
Mannan 15.49G3.99** 79.30G5.33
EDTA 12.98G10.14** 82.66G13.55
mAb-19-kDa 13.36G7.02** 82.15G9.33
mAb-MR 10.05G6.15** 86.57G8.22
Isotype control 82.97G5.30 –
THP-1 cells were incubated with recombinant M. smegmatis. Cells were
pretreated with inhibitors for 1 h. At least 200 cells were randomly
examined in each coverslip to assess the percentage of cells binding at least
one bacterium. Data shown are meansGSD of three independent
experiments with triplicate coverslips. MR, mannose receptor; a-mm, a-
methyl-mannoside. *P%0.5, **P%0.05, ***P%0.001. The effects of
treating cells with saccharydes, EDTA, and with mAbs to the Mannose
Receptor and to the 19-kDa antigen of M. tuberculosis.
mycobacterial proteins with an adhesin function that could
be revealed under different experimental conditions. It has
been reported that the antigen 85 complex coupled to
microbeads binds macrophages through CR3 [13] and that
latex beads coated with the 30-kDa heparin-binding
hemaglutinin bind macrophage-like cells in both the
presence and absence of human serum [14].
After identifying the 19-kDa adhesin as the glycolipo-
protein characterized originally by Garbe et al. [15], we
undertook this study using CFP from a recombinant
M. smegmatis strain, which overexpresses the 19-kDa
antigen of M. tuberculosis. As the M. tuberculosis protein,
the recombinant expressed in M. smegmatis was shown to
possess mannosyl residues susceptible to a-D-mannosidase
digestion. At present it is known that M. tuberculosis is
capable of synthesizing glycoproteins [27–29]. Among
these is the 19-kDa antigen herein described, which in
addition to be acylated [30], possess mannosyl residues as
shown by lectin binding assays with Con A [15]. Morever,
O-glycosylation sites have been demonstrated in this protein
by site-directed mutagenesis [31]. This antigen is biologi-
cally very active; it induces strong antibody [16] and cell-
mediated immune responses [15,17] and interacts with
macrophages via Toll-like 2 receptors inducing IL-12 and
TNFa production [32] or apoptosis [18]. Recently, it has
been shown that the 19-kDa antigen can inhibit MHC-II
antigen presentation by blocking gamma interferon
signaling [19].
Our current observations show that the 19-kDa M.
tuberculosis antigen interacts with the macrophage MR
present on the surface of THP-1 cells. In the first place,
Ca2C chelators, a-methyl-mannoside, mannan and

H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107 103
N-acetyl-D-glucosamine decreased the binding of the
19-kDa antigen to cells. Moreover, a similar effect was
observed with a mAb to the human macrophage MR.
Results of the phagocytosis assays support the above notion.
Indeed, the percentage of THP-1 cells with associated
bacilli was markedly reduced in inhibition assays with
mannan and a-methyl-mannoside, recognized MR ligands
[23,24], and with EDTA. The phagocytosis of recombinant
M. smegmatis was due to the interaction of the 19-kDa
antigen with the MR on THP-1 cells; this was demonstrated
by the marked reduction in the percentage of cells with
bound bacilli observed in inhibition assays with mAbs to the
19-kDa antigen and to the MR.
To our knowledge, there are no previous reports of
M. tuberculosis glycolipoproteins with adhesin function
capable of interacting with the MR that promote phagocy-
tosis of mycobacteria. Although it has been observed that
M. tuberculosis can be ingested by the interaction of MR
with mannosyl residues of LAM, an abundant cell-wall
located mycobacterial lipoglycan [5]. In addition to its role
in homeostasis of glycoproteins [33], the MR is considered a
pattern recognition receptor important in the innate host
response to pathogens [34] that is involved in the
phagocytosis of a variety of microbes [35–38].
As far as we know, the expression of MR on THP-1
monocytes has not yet been documented although some
suggestive observations have been made. It has been
proposed that THP-1 cells ingest Malassezia furfur [39]
and M. tuberculosis [40] via MR since uptake is inhibited by
preincubation with mannan. Another study showed that
LAM and other MR ligands induce the production of
metalloproteinase-9 by THP-1 cells that was abrogated with
an mAb to MR [41]. In this study we have obtained direct
evidences of the expression of MR by THP-1 monocytic
cells. First, immunoprecipitation with a lipoprotein fraction
from M. tuberculosis and THP-1 whole cell membranes
resulted in coprecipitation of both MR and 19-kDa antigen.
Secondly, immunostaining with and anti MR antibody
revealed strong cell surface labelling and intracellular MR
deposits. Finally, the surface expression of the MR on THP-
1 cells was also demonstrated by cell-surface ELISA, a
method that detects primarily cell membrane exposed cell
molecules [22]. It is of interest that cell surface expression
of the MR requires prior stimulation with recombinant
M. smegmatis culture filtrate proteins. These findings
propose that mycobacterial products may upregulate the
surface expression of the MR in cells of the monocytic-
macrophagic lineage which would be of great interest in the
pathogenesis of mycobacterial infections. Whether
recycling or other mechanism are involved in this
phenomenon require research that is not within the scope
of the present study. It is worth to mention that cells
expressing surface MR or with ingested bacilli often
displayed abundant well-spread cytoplasm suggesting they
had been differentiated into macrophages by the addition of
mycobacterial culture filtrate proteins or bacilli. In relation
to our current findings are two recent studies which
demonstrate surface up-regulation of the human macro-
phage MR by 1 h exposure of cells to surfactant proteins A
or D [42,43]. In a study it is proposed that the newly
expressed MR comes from preformed intracellular pools
and that de novo protein synthesis is not required for this
phenomenon to occur.
In summary, the current observations lend further
emphasis to the importance of the 19-kDa antigen as a
mycobacterial moiety of great biological activity. These
result also show, that as other intracellular pathogens, M.
tuberculosis is armed with panoply of resources to
accomplish the crucial step of invading host cells.
4. Materials and methods
4.1. Mycobacteria and mycobacterial protein preparations
M. tuberculosis strain H37Rv (ATCC 27294), obtained
from the American Type Culture Collection (Manassas,
VA), was grown for 6–8 weeks in Proskauer and Beck
synthetic medium. M. smegmatis strain (mc2155) and its
recombinant counterpart transformed by electroporation
with the plasmid p16R1 which contains a 1.8 kb SmaI
fragment that includes the structural gene encoding the
19-kDa M. tuberculosis antigen were kindly donated by
Y. Zhang (MRC Tuberculosis and Related Infections Unit,
Hammersmith Hospital, London, UK). M. smegmatis,
native and recombinant were grown for 4–5 days in
Middlebrook 7H9 medium (BBL, Becton-Dickinson,
Cockeysville, MD) supplemented with 2% glucose and
hygromycin B (50 mg/ml). CFP from M. tuberculosis and
M. smegmatis were obtained by (NH4)2SO4 precipitation of
the culture medium after elimination of bacilli by filtration
[27]. CFP were labelled with biotin as described [44]. A
cell-wall protein fraction was obtained from M. tuberculosis
bacilli as follows. After killing with 2% sodium azide,
whole bacilli were rinsed with PBS and sonicated in ice (5!60 s pulses). After sonication the cell-wall fraction was
recovered by centrifugation (20 min at 20,000 rpm). Protein
concentration was determined by the Lowry protein assay
(Bio-Rad Laboratories, Hercules, and CA).
4.2. Characterization of the recombinant M. smegmatis
19 kDa antigen
To identify the 19-kDa antigen in mycobacterial CFP,
after SDS-PAGE, nitrocellulose membranes or PVDF
membranes (Bio-Rad) were incubated for 1 h with the
mAb IT-19 diluted 1:1000 (a kind gift from the TB Research
Material and Vaccine Testing Contract, Colorado State
University). IT-19 recognizes the 19-kDa M. tuberculosis
antigen [15,21]. The presence of mannose residues was
analysed by affinity blotting with Con A labelled with
peroxidase (Zymed Laboratories, San Francisco, CA) as

H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107104
described elsewhere [27,28]. With the same purpose, CFP
from recombinant M. smegmatis were digested with a-D-
mannosidase as described previously [28]. Briefly, 10 mg
CFP were digested for 1 h at 37 8C with jack bean a-D-
mannosidase (Sigma Chemical Co., St Louis, MO) in 0.1 M
sodium acetate buffer (pH 5), using 15 U of enzyme/mg
CFP. Digestion was stopped with sample buffer and proteins
were boiled and separated by SDS-PAGE to carry out
immunoblotting analysis.
4.3. Culture of THP-1 monocytic cells and binding assays
with mycobacterial CFP
The monocytic human cell line THP-1 was obtained
from ATCC (TIB-202) and cultured in RPMI 1640
medium supplemented with 10% fetal bovine serum
(FBS), 1 mM penicillin–streptomycin, 1 mM non-essential
aminoacids, 100 mM piruvic acid, and 1 mM L-glutamine
(Gibco BRL Products, Rockville, MD). Cell viability was
assessed by trypan blue exclusion. THP-1 cells (5!105)
were incubated with different amounts of M. tuberculosis
CFP labelled with biotin or without labelling at 37 8C,
with 5% CO2 for 1 h. Similarly, THP-1 cells were
incubated with unlabeled CFP obtained from recombinant
and native M. smegmatis 7-days-old cultures. Incubation
was carried out in RPMI 1640 with 10% FBS, 0.02%
NaN3, 0.006 PMSF, and 0.25 mM EDTA. After incubation,
the medium was carefully retired and wells were rinsed
several times by gentle pippeting with PBS and by 5 min
centrifugation in PBS at 1500 rpm in the cold. Cells were
suspended in sample buffer (19 mM EDTA, 1% glycerol,
0.1% SDS, 0.5 mM Tris–HCI pH 6.8, and 2-mercap-
toethanol) and resolved by 15% SDS-PAGE. After transfer
to nitrocellulose and blocking with 3% BSA in PBS,
membranes were incubated for 1 h with the IT-19 mAb
diluted 1:1000. Thereafter, a secondary peroxidase-labeled
goat antibody to mouse IgG was applied diluted 1:2000
(Zymed). Peroxidase was developed with 3,3 0-diamino-
benzidine and H2O2 or with a chemiluminiscence kit
(Pierce Biotechnology, Rockford, IL). For control, a mAb
to mouse b-actin was used in some immunoblots
(Chemicon International, Temecula, CA).
4.4. Cell-surface ELISA to study binding of the 19-kDa
antigen to THP-1 cells
To quantitate the binding of the 19-kDa antigen a cell-
surface ELISA was carried out [22]. Briefly, after blocking
wells for 2 h with 3% BSA in PBS, 1!105 THP-1 cells
were placed in each well. After 2 h, 1, 10, 20 or 50 mg CFP
from native or recombinant M. smegmatis were added. After
1 h incubation at 37 8C, wells were rinsed three times with
PBS containing 1% FBS and 0.1% NaN3.Thereafter, cells
were incubated with mAb IT-19 diluted 1:100 in PBS or
with an lgG1 isotype control antibody (Dako Corporation,
Carpinteria, CA) for 1 h. After rinsing, cells were incubated
with an alkaline phosphatase-conjugated goat anti-mouse
IgG (Dako) diluted 1:1000 for 1 h at 37 8C. After rinsing
extensively, 200 ml of a 1.5 mg/ml p-nitrophenyl phosphate
solution (Sigma) were added to the wells. The reaction was
stopped with 25 ml, 3 M NaOH. Optical densities at 405 nm
were read in an ELISA reader (Bio-Tek Instruments).
4.5. Adherence assays with fluorescent microspheres
coated with mycobacterial CFP
To determine whether the 19-kDa antigen may promote
phagocytosis, assays with microspheres were performed.
Fluoresceinated polystyrene 1 mm microspheres (Molecular
Probes, Inc., Eugene, OR) were coated with various
amounts of CFP from native or recombinant M. smegmatis
and with BSA, following manufacturer’s protocols. Briefly,
1.0810!1010 microspheres/ml were mixed with 500 mg
CFP from native and recombinant M. smegmatis in 1 ml
MES Buffer [2-(N-morpholino)ethanesulfonic acid (pH
6.0)]. Then, 1-ethyl-3-(3-dimethylaminopropyl)-carbodii-
mide (Molecular Probes) was added at 2.5 mg/ml final
concentration. After overnight incubation at room tempera-
ture, glycine (100 mM) was added to stop the reaction.
Microspheres were rinsed three times with PBS and kept a
4 8C in PBS containing 1% BSA to block possible
remaining active sites. Adherence assays were carried out
as follows. THP-1 cells (5!105) were removed with a
scrapper, counted, rinsed, resuspended in RPMI 1640
containing 10% FBS and placed on acid-washed glass
coverslips. Microspheres were added to cells and after 1 h
incubation at 37 8C in 5% CO2, cells were rinsed in PBS to
remove non-adherent microspheres and fixed in 10%
paraformaldehide for 10 min. To estimate the number of
adherent microspheres, coverslips were mounted on a slide
and at least 200 randomly chosen cells were examined in a
Nikon fluorescence microscope.
4.6. Assays to inhibit the binding of the 19-kDa antigen
and of microspheres to THP-1 cells
To determine if the 19-kDa antigen binds cells through
its mannose residues, inhibition assays with mannose
competitors were performed. When immunoblotting was
used to assess binding, THP-1 cells (5!105) were treated at
37 8C with 1, 10, 100 and 1000 mM a-methyl-mannoside,
10 and 50 mM N-acetyl-D-glucosamine and 5 and 10 mg
mannan. After 1 h treatment 50 mg CFP were added to cells.
After three rinses with PBS, cells were processed for
immunoblotting as described above. The binding of the
19-kDa antigen was demonstrated as described above. A
more quantitative Cell-Surface ELISA assay was used to
analyse the effects of pretreating cells with a-methyl-
mannoside, calcium chelators and a mAb to MR; 96-well
flat bottom ELISA plates were blocked as described above
and 1!105 cells were placed in each well for 2 h. Then, 1,
10, 100 and 1000 mM a-methyl-mannoside, 2 mM EDTA,

H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107 105
2 mM EGTA and 10 mg mAb to the MR were added to cells.
At the same time, 20 mg recombinant M. smegmatis CFP
were added for 1 h. Plates were rinsed three times at
1500 rpm with PBS containing 1% FBS and 0.1% NaN3.
Then, the IT-19 mAb diluted 1:100 in 3% BSA-PBS was
added to the wells. After 1 h incubation and three PBS
rinses, bound antibody was revealed with a goat anti-mouse
IgG antibody labelled with alkaline phosphatase as
described above.
Assays to block the adherence of microspheres coated
with recombinant M. smegmatis CFP to cells were also
performed. THP-1 cells (1!105) were preincubated with 1,
10, 100 and 1000 mM a-methyl-mannoside. After 1 h,
microspheres were added at a 20:1 microsphere/cell. After
extensive rinsing with PBS, coverslips were analysed by
fluorescence microscopy to count attached microspheres in
at least 200 randomly chosen cells.
4.7. Immunoprecipitation of the mannose macrophage
receptor and the 19-kDa M. tuberculosis antigen
A cell-membrane fraction was obtained following a
reported method [45]. Briefly, THP-1 cells were homogen-
ized by vortex in a lysis buffer (50 mM Tris–HCI, 150 mM
NaCI and 1 mM EDTA, pH 8) containing a cocktail of
protease inhibitors (pepstatin, leupeptin, aprotinin, quimos-
tatin, antipain and PMSF). After elimination of nuclei, cell
membranes were isolated by centrifugation at 32,000 rpm
for 1 h. The pellet was resuspended in the lysis buffer and
membrane proteins were extracted with 0.1% Triton X-100
(Sigma). Membrane proteins (250 mg) were incubated with
50 mg of a M. tuberculosis lipoprotein fraction enriched in
the 19-kDa antigen (kindly donated by TB Research
Material and Vaccine Testing Contract, Colorado State
University), with shaking, overnight at 4 8C. The incubation
buffer contained 2 mM CaCl2 and MgCl2. Immuno-
precipitation was carried out with a goat anti-MR polyclonal
antibody bound to Protein G-agarose microbeads for 3 h at
4 8C. The MR antibody was a kind gift from P. Stahl
(Washington University, St Louis, MO). Before immuno-
precipitation, microbeads were precleaned by incubation
with macrophage membranes and the lipoprotein fraction
for 1 h at 4 8C. After rinsing, microbeads were resuspended
in sample buffer and boiled. Microbeads were eliminated by
centrifugation at 12,000 rpm for 5 min and proteins were
separated by 7.5% SDS-PAGE to detect the mannose
receptor and at 15% to detect the 19-kDa antigen. After
transfer to nitrocellulose, strips were incubated for 3 h at
room temperature with the polyclonal MR antibody diluted
1:200 in PBS-Tween. After rinsing, a rabbit anti-goat IgG
antibody labelled with peroxidase was applied (Zymed).
The 19-kDa antigen was detected with the mAb IT-19 as
described before. A chemiluminescence kit was used to
reveal the blots (Pierce Biotechnology). Control immuno-
precipitation experiments with preimmune goat serum were
carried out.
4.8. Immunostaining procedure to demonstrate the mannose
receptor on THP-1 monocytic cells
THP-1 cells cultured on glass coverslips were incubated
for 1 h with 50 mg recombinant M. smegmatis CFP or with
culture medium alone. Cells were washed extensively with
PBS and fixed with 1% paraformaldehide at room
temperature for 30 min. After washing, cells were treated
with 3% BSA in PBS to inhibit non-specific binding.
Endogenous peroxidase activity was eliminated by treating
cells for 30 min with 5% methanol and 0.1% H2O2 and
permeabilized with Triton X-100 as described above. Then,
the goat polyclonal MR antibody diluted 1:100 in PBS was
applied to the coverslips. After rinsing three times with
PBS, a secondary peroxidase-labeled rabbit antibody to goat
IgG (Zymed) was applied diluted 1:1000. Peroxidase was
revealed with 3,3 0-diaminobenzidine and H2O2. In control
slides, the MR antibody was omitted. Coverslips were
mounted on glass slides, counterstained with Haematoxylin-
Eosin and examined in a light microscope.
4.9. Cell-surface ELISA to study the surface expression
of the mannose receptor on THP-1 cells
To quantitate MR expression a cell-surface ELISA was
carried out [22]. After blocking with BSA-PBS as described
above, 1!105 THP-1 cells were placed in each well. After
2 h 50 mg CFP from recombinant M. smegmatis were added.
After 1 h incubation at 37 8C, wells were rinsed three times
with PBS containing 1% FBS and 0.1% NaN3.Thereafter,
cells were incubated with 10 mg mAb to the MR (BD
Pharmingen) diluted 1:100 in PBS or with an lgG1 isotype
control antibody (Dako Corporation, Carpinteria, CA) for
1 h. After rinsing, cells were incubated with an alkaline
phosphatase-conjugated goat anti-mouse IgG (Dako)
diluted 1:1000 for 1 h at 37 8C. After rinsing extensively
alkaline phosphatase was developed as described above and
optical densities at 405 nm were read in an ELISA reader
(Bio-Tek Instruments).
4.10. Phagocytosis assays with recombinant M. smegmatis
and THP-1 cells
THP-1 cells plated on glass coverslips were cultured in
RPMI-1640 with 10% FBS. Cells were washed and
resuspended in RPMI-1640 in the absence of FBS. An
aliquot of recombinant M. smegmatis was thawed, rinsed
three times with PBS at 12,000 rpm for 20 min. Cells were
incubated with recombinant M. smegmatis at a 20/1
bacteria/cell. Previously, the bacterial suspension was
passed 10 times through a 25-gauge needle attached to a
1 ml syringe to eliminate clumps. After 1 h infection, cells
were washed extensively with PBS, fixed with 1%
paraformaldehide and treated with methanol and H2O2 to
eliminate endogenous peroxidase. Then, cells were permea-
bilized with 0.5% Triton X-100, 1% BSA in PBS.

H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107106
After washing twice with PBS, cells were incubated for 1 h
with a rabbit antiserum to M. tuberculosis H37Rv, diluted 1:
50 in PBS. This antiserum cross-reacts extensively with
recombinant M. smegmatis CFP as seen by immunoblot (not
shown). After extensive washing with PBS, Protein A
labelled with peroxidase diluted 1:1000 in PBS was applied
to the coverslips. Peroxidase was developed as above. In
inhibition assays, cells were preincubated for 1 h with
mannan (5 mg), 100 mM a-methyl-mannoside and with
EDTA (5 mg). Inihibition assays with a mAb to the human
MR (20 mg; BD Pharmingen) and the IT-19 mAb (1:100
dilution) that recognizes the M. tuberculosis 19-kDa antigen
were also performed. Cells were preincubated with mAbs
(both IgG1 isotype) for 1 h, and without removal of the
mAb the bacilli were added. Control isotype experiments
were carried out with an unrelated IgG1 mAb diluted 1:50
(Dako). Each experiment was performed in triplicate and
results are expressed as the meanGSD.
4.11. Statistical analysis
The t Student test was used to make comparisons
between groups. ELISA data were analyzed by a two-way
ANOVA with significant differences evaluated by a Tukey’s
HSD procedure. Data are reported as meanGSEM.
Significance was set at P!0.05.
Acknowledgements
We would like to thank Ismael Ramirez for skilful
technical assistance and to Dr Imelda Lopez for reviewing
the manuscript. This work was supported by grant
IN233201 from DGAPA, UNAM.
References
[1] Schlesinger LS, Bellinger-Kawahara CG, Payne NR, Horwitz MA.
Phagocytosis of Mycobacterium tuberculosis is mediated by human
monocyte complement receptors and complement component C3.
J Immunol 1990;144:2771–80.
[2] Schlesinger LS. Macrophage phagocytosis of virulent but not
attenuated strains of Mycobacterium tuberculosis is mediated by
mannose receptors in addition to complement receptors. J Immunol
1993;150:2920–30.
[3] Hirsch CS, Ellner JJ, Rusell GD, Rich EA. Complement receptor-
mediated uptake and tumor necrosis factor alpha mediated growth
inhibition of M. tuberculosis by human alveolar macrophages.
J Immunol 1993;152:743–53.
[4] Schlesinger LS. Role of mononuclear phagocytes in Mycobacterium
tuberculosis pathogenesis. J Invest Med 1993;44:312–23.
[5] Kang BK, Schlesinger LS. Characterization of mannose receptor-
dependent phagocytosis mediated by Mycobacterium tuberculosis
lipoarabinomannan. Infect Immun 1993;66:2769–77.
[6] Gaynor CD, McCormack FX, Voelker DR, McGowan SE,
Schlesinger LS. Pulmonary surfactant protein A mediates enhanced
phagocytosis of Mycobacterium tuberculosis by a direct interaction
with human macrophages. J Immunol 1995;155:5343–51.
[7] Chroneos ZC, Abdolrasulnia R, Whitsett JA, Rice WR, Shepherd VL.
Purification of a cell-surface receptor for surfactant protein A. J Biol
Chem 1996;271:16375–83.
[8] Peterson PK, Gekker G, Hu S, Sheng WS, Anderson WR, Ulevitch RJ,
et al. CD14 receptor-mediated uptake of nonopsonized Mycobacter-
ium tuberculosis by human microglia. Infect Immun 1995;63:
1598–602.
[9] Zimmerii S, Edwards S, Ernst JD. Selective receptor blockade during
phagocytosis does not alter the survival and growth of Mycobacterium
tuberculosis in human macrophages. Am J Respir Cell Mol Biol 1996;
15:760–70.
[10] Schlesinger LS, Hull SR, Kaufman TM. Binding of the terminal
mannosyl units of lipoarabinomannan from a virulent strain of
Mycobacterium tuberculosis to human macrophages. J Immunol
1994;152:4070–9.
[11] Stokes RW, Speert DP. Lipoarabinomannan inhibits nonopsonic
binding of Mycobacterium tuberculosis to murine macrophages.
J Immunol 1995;155:1361–9.
[12] Arruda S, Bomfim G, Knights R, Huima-Byron T, Riley LW. Cloning
of an M. tuberculosis DNA fragment associated with entry and
survival inside cells. Science 1993;26:1454–7.
[13] Hetland G, Wiker HG. Antigen 85C on Mycobacterium bovis, BCG
and Mycobacterium tuberculosis promotes monocyte-CR3-mediated
uptake of microbeads coated with mycobacterial products. Immu-
nology 1994;82:445–9.
[14] Mueller-Ortiz SL, Wanger AR, Norris SJ. Mycobacterial protein
HbhA binds human complement component C3. Infect Immun 2001;
69:7501–11.
[15] Garbe TR, Harris D, Vordermeier M, Lathigra R, Ivanyi J, Young DB.
Expression of the Mycobacterium tuberculosis 19-kilodalton antigen
in Mycobacterium smegmatis: immunological analysis and evidence
of glycosylation. Infect Immun 1993;61:260–7.
[16] Franco J, Camarena JJ, Nogueira JM, Blanquer R, Ruiz MJ, Marin J.
Serological response (Western blot) to fractions of Mycobacterium
tuberculosis sonicate antigen in tuberculosis patients and contacts. Int
J Tuberc Lung Dis 2001;5:958–62.
[17] Faith A, Moreno C, Lathigra R, Roman E, Fernandez M, Brett S, et al.
Analysis of human T-cell epitopes in the 19,000 MW antigen of
Mycobacterium tuberculosis: influence of HLADR. Immunology
1991;74:1–7.
[18] Lopez M, Sly LM, Luu Y, Young D, Cooper H, Reiner NE. The 19-
kDa Mycobacterium tuberculosis protein induces macrophage
apoptosis through Toll-like receptor-2. J Immunol 2003;170:2409–16.
[19] Fulton SA, Reba SM, Pai RK, Pennini M, Torres M, Harding CV,
Boom WH. Inhibition of major histocompatibility complex II
expression and antigen processing in murine alveolar macrophages
by Mycobacterium bovis/BCG and the 19-kilodalton mycobacterial
lipoprotein. Infect Immun 2004;72(4):2101–10.
[20] Bayer EA, Wilcheck M. Protein biotinylation. Methods Enzymol
1994;184:138–60.
[21] Coates AR, Hewitt J, Alien BW, Ivanyi J, Mitchison DA. Antigenic
diversity of Mycobacterium tuberculosis and Mycobacterium bovis
detected by means of monoclonal antibodies. Lancet 1981;2:167–9.
[22] Grunow R, D’Apuzzo M, Wyss-Coray T, Frutig K, Pichler WJ. A cell
surface ELISA for the screening of monoclonal antibodies to antigens
on viable cells in suspension. J Immunol Methods 1994;171:93–102.
[23] Kery V, Krepinsky JJ, Warren CD, Capek P, Stahl PD. Ligand
recognition by purified human mannose receptor. Arch Biochem
Biophys 1992;298:49–55.
[24] Mullin NP, Hitchen PG, Taylor ME. Mechanism of Ca2C and
monosaccharide binding to a C-type carbohydrate-recognition domain
of the macrophage mannose receptor. J Biol Chem 1997;272:
5668–81.
[25] Wileman TE, Lennartz MR, Stahl PD. Identification of the
macrophage mannose receptor as a 175-kilodalton membrane protein.
Proc Natl Acad Sci USA 1986;83:2501–5.

H. Diaz-Silvestre et al. / Microbial Pathogenesis 39 (2005) 97–107 107
[26] Klemm P, Schembri MA. Bacterial adhesins: function and structure.
Int J Med Microbiol 2000;290:27–35.
[27] Espitia C, Mancilla R. Identification, isolation and partial character-
ization of Mycobacterium tuberculosis glycoprotein antigens. Clin
Exp Immunol 1989;77:378–83.
[28] Espitia C, Espinosa R, Saavedra R, Mancilla R, Remain F,
Laqueyrerie A, et al. Antigenic and structural similarities between
Mycobacterium tuberculosis 50- to 55-kilodalton and Mycobacterium
bovis BCG 45- to 47-kilodalton antigens. Infect Immun 1995;63:
580–4.
[29] Dobos KM, Swiderek K, Khoo KH, Brennan PJ, Belisle JT. Evidence
for glycosylation sites on the 45-kilodalton glycoprotein of
Mycobacterium tuberculosis. Infect Immun 1986;63:2846–53.
[30] Young DB, Garbe TR. Lipoprotein antigens of Mycobacterium
tuberculosis. Res Microbiol 1986;142:55–65.
[31] Herrmann JL, O’Gaora P, Gallagher A, Thole JE, Young DB.
Bacterial glycoproteins: a link between glycosylation and proteolytic
cleavage of a 19-kilodalton antigen from Mycobacterium tubercu-
losis. Eur Mol Biochem Org J 1996;15:3547–54.
[32] Brightbill HD, Libraty DH, Krutzik SR, Yang RB, Belisle JT,
Bleharski JR, et al. Host defense mechanisms triggered by microbial
lipoproteins through Toll-like receptors. Science 1999;285:732–6.
[33] Lee SJ, Evers S, Roeder D, Parlow AF, Risteli J, Risteli L, et al.
Mannose receptor-mediated regulation of serum glycoprotein homeo-
stasis. Science 1995;295:1898–901.
[34] Stahl PD, Ezekowitz RA. The mannose receptor is a pattern
recognition receptor involved in host defense. Curr Opin Immunol
1995;10:50–5.
[35] Marodi L, Schreiber S, Anderson DC, MacDermott RP, Korchak HM,
Johnston Jr RB. Enhancement of macrophage candidacidal activity by
interferon-gamma. Increased phagocytosis, killing, and calcium
signal mediated by a decreased number of mannose receptors.
J Clin Invest 1993;91:2596–601.
[36] O’Riordan DM, Standing JE, Limper AH. Pneumocystis carinii
glycoprotein A binds macrophage mannose receptors. Infect Immun
1995;63:779–84.
[37] Cinco M, Cini B, Murgia R, Presani G, Prodan M, Perticarari S.
Evidence of involvement of the mannose receptor in adhesion of
Borrelia burgdorferi to monocyte/macrophages. Infect Immun 1995;
69:2743–7.
[38] Chakraborty P, Ghosh D, Basu MK. Modulation of macrophage
mannose receptor affects the uptake of virulent and avirulent
Leishmania donovani promastigotes. J Parasitol 1995;87:1023–7.
[39] Suzuki T, Ohno N, Ohshima Y, Yadomae T. Soluble mannan and
beta-glucan inhibit the uptake of Malassezia furfur by human
monocytic cell line, THP-1. Fed Eur Mol Soc Immunol Med
Microbiol 1998;21:223–30.
[40] Stokes RW, Doxsee D. The receptor-mediated uptake, survival,
replication, and drug sensitivity of Mycobacterium tuberculosis
within the macrophage-like cell line THP-1: a comparison with
human monocyte-derived macrophages. Cell Immunol 1999;197:1–9.
[41] Rivera-Marrero CA, Schuyler W, Roser S, Ritzenthaler JD,
Newburn SA, Roman J. M. tuberculosis induction of matrix
metalloproteinase-9: the role of mannose and receptor-mediated
mechanisms. Am J Physiol Lung Cell Mol Physiol 2002;282:
L546–L55.
[42] Beharka AA, Gaynor CD, Kang BK, Voelker DR, McCormack FX,
Schlesinger LS. Pulmonary surfactant protein A up-regulates activity
of the mannose receptor, a pattern recognition receptor expressed on
human macrophages. J Immunol 2002;169:3565–73.
[43] Kudo K, Sano H, Takahashi H, Kuronuma K, Yokota S, Fujii N, et al.
Pulmonary collectins enhance phagocytosis of Mycobacterium avium
through increased activity of mannose receptor. J Immunol 2004;172:
7592–602.
[44] Anderson LJ, Coombs RA, Tsuo C, Hierhalzer JC. Use of biotin–
avidin system to study specificity of antibodies against respiratory
syncitial virus. J Clin Microbiol 1984;19:934–6.
[45] Yoshida H, Kondratenko N, Green S, Steinberg D, Quehenberger O.
Identification of the lectin-like receptor for oxidized low-density
lipoprotein in human macrophages and its potential role as a
scavenger receptor. Biochem J 1998;334:9–13.




















