
The N-terminal domain of the Flo11 protein from Saccharomyces cerevisiae is an adhesin without...
10
RESEARCH ARTICLE The N-terminal domain of the Flo11 protein from Saccharomyces cerevisiae is an adhesin without mannose-binding activity Katty V.Y. Goossens & Ronnie G. Willaert Laboratory of Structural Biology, Department of Bioengineering Sciences, Vrije Universiteit Brussel, Brussels, Belgium Correspondence: Ronnie G. Willaert, Laboratory of Structural Biology, Department of Bioengineering Sciences, Vrije Universiteit Brussel, Pleinlaan 2, B-1050 Brussel, Belgium. Tel.: +32 2 6291846; fax: +32 2 6291963; e-mail: [email protected] Received 13 September 2011; revised 11 November 2011; accepted 11 November 2011; Final version published online 22 December 2011. DOI: 10.1111/j.1567-1364.2011.00766.x Editor: Jens Nielsen Keywords Saccharomyces cerevisiae; flocculation; Flo11 protein; carbohydrate binding; self- interaction. Abstract The expression of the Flo11 flocculin in Saccharomyces cerevisiae offers the cell a wide range of phenotypes, depending on the strain and the environmental conditions. The most important are pseudohyphae development, invasive growth and flocculation. The mechanism of cellular adhesion mediated by Flo11p is not well understood. Therefore, the N-terminal domain of Flo11p was purified and studied. Although its amino acid sequence shows less similar- ity with the other flocculins, Flo11p belongs to the flocculin family. However, the N-terminal domain contains the ‘Flo11-domain’ (PF10181), but not the mannose-binding PA14 domain, which is present in the other flocculins (Flo1p, Flo5p, Flo9p and Flo10p). Structural and binding properties of the N- terminal domain of Flo11p were studied. It is shown that this domain is O-gly- cosylated and is structurally composed mainly of b-sheets, which is typical for the members of the flocculin family. Furthermore, fluorescence spectroscopy binding studies revealed that N-Flo11p does not bind mannose, which is in contrast to the other Flo proteins. However, surface plasmon resonance analy- sis showed that N-Flo11p self-interacts and explains the cell–cell interaction capacity of FLO11-expressing cells. Introduction Saccharomyces cerevisiae shows different phenotypes depending on its genetic background and its environmen- tal condition. The yeast has been commonly studied as a single-cell organism living freely in suspension. However, in its natural environment, the cells frequently switch over from a planktonic life style to complex multicellular structures such as flocs, filaments, mats and flors (Palkova, 2004). These morphological transitions have been sug- gested as a mechanism for this nonmotile species to permit foraging for nutrients (Gimeno et al., 1992; Iserentant, 1996; Cullen & Sprague, 2000; Dickinson, 2008). For example, haploid cells become adherent and invade the surface of a semi-solid agar medium, so that they do not wash off. This is called invasive growth, and this morphological switch is caused by glucose depletion around the growing cells (Roberts & Fink, 1994; Cullen & Sprague, 2000). Diploid cells display a similar filamentous transition by adhering to each other after cell division, when sensing a low nitrogen concentration. In diploid pseudohyphal growth, the cells adopt an elongated shape and form long filaments that grow out from the colony edge (Gimeno et al., 1992; Kron et al., 1994; Mosch, 2000). Pseudohyphal development of S. cerevisiae has fea- tures in common with dimorphism of pathogenic fungi, where the dimorphic transition is often correlated with pathogenicity. Thus, the molecular models drawn from studies of pseudohyphal development of S. cerevisiae build a valuable basis for studying dimorphism in patho- genic fungi that are experimentally less accessible and therefore offer important insights into the mechanism and control of fungal disease (Mosch, 2000). Yeast cells exhibit these morphological transitions when a class of genes, encoding flocculins, is present in their genome. More specifically, the adhesion of the cells to substrates, the pseudohyphal and the invasive growth are S. cerevisiae phenotypes resulting from the expression of ª 2011 Federation of European Microbiological Societies FEMS Yeast Res 12 (2012) 78–87 Published by Blackwell Publishing Ltd. All rights reserved YEAST RESEARCH
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The N-terminal domain of the Flo11 protein from Saccharomyces cerevisiae is an adhesin without...

R E S EA RCH AR T I C L E
The N-terminal domain of the Flo11 protein fromSaccharomyces cerevisiae is an adhesin without
mannose-binding activity
Katty V.Y. Goossens & Ronnie G. Willaert
Laboratory of Structural Biology, Department of Bioengineering Sciences, Vrije Universiteit Brussel, Brussels, Belgium
Correspondence: Ronnie G. Willaert,
Laboratory of Structural Biology, Department
of Bioengineering Sciences, Vrije Universiteit
Brussel, Pleinlaan 2, B-1050 Brussel, Belgium.
Tel.: +32 2 6291846; fax: +32 2 6291963;
e-mail: [email protected]
Received 13 September 2011; revised 11
November 2011; accepted 11 November
2011; Final version published online 22
December 2011.
DOI: 10.1111/j.1567-1364.2011.00766.x
Editor: Jens Nielsen
Keywords
Saccharomyces cerevisiae; flocculation; Flo11
protein; carbohydrate binding; self-
interaction.
Abstract
The expression of the Flo11 flocculin in Saccharomyces cerevisiae offers the cell
a wide range of phenotypes, depending on the strain and the environmental
conditions. The most important are pseudohyphae development, invasive
growth and flocculation. The mechanism of cellular adhesion mediated by
Flo11p is not well understood. Therefore, the N-terminal domain of Flo11p
was purified and studied. Although its amino acid sequence shows less similar-
ity with the other flocculins, Flo11p belongs to the flocculin family. However,
the N-terminal domain contains the ‘Flo11-domain’ (PF10181), but not the
mannose-binding PA14 domain, which is present in the other flocculins
(Flo1p, Flo5p, Flo9p and Flo10p). Structural and binding properties of the N-
terminal domain of Flo11p were studied. It is shown that this domain is O-gly-
cosylated and is structurally composed mainly of b-sheets, which is typical for
the members of the flocculin family. Furthermore, fluorescence spectroscopy
binding studies revealed that N-Flo11p does not bind mannose, which is in
contrast to the other Flo proteins. However, surface plasmon resonance analy-
sis showed that N-Flo11p self-interacts and explains the cell–cell interaction
capacity of FLO11-expressing cells.
Introduction
Saccharomyces cerevisiae shows different phenotypes
depending on its genetic background and its environmen-
tal condition. The yeast has been commonly studied as a
single-cell organism living freely in suspension. However,
in its natural environment, the cells frequently switch
over from a planktonic life style to complex multicellular
structures such as flocs, filaments, mats and flors (Palkova,
2004). These morphological transitions have been sug-
gested as a mechanism for this nonmotile species to permit
foraging for nutrients (Gimeno et al., 1992; Iserentant,
1996; Cullen & Sprague, 2000; Dickinson, 2008).
For example, haploid cells become adherent and invade
the surface of a semi-solid agar medium, so that they do
not wash off. This is called invasive growth, and this
morphological switch is caused by glucose depletion
around the growing cells (Roberts & Fink, 1994; Cullen &
Sprague, 2000). Diploid cells display a similar filamentous
transition by adhering to each other after cell division,
when sensing a low nitrogen concentration. In diploid
pseudohyphal growth, the cells adopt an elongated shape
and form long filaments that grow out from the colony
edge (Gimeno et al., 1992; Kron et al., 1994; Mosch,
2000). Pseudohyphal development of S. cerevisiae has fea-
tures in common with dimorphism of pathogenic fungi,
where the dimorphic transition is often correlated with
pathogenicity. Thus, the molecular models drawn from
studies of pseudohyphal development of S. cerevisiae
build a valuable basis for studying dimorphism in patho-
genic fungi that are experimentally less accessible and
therefore offer important insights into the mechanism
and control of fungal disease (Mosch, 2000).
Yeast cells exhibit these morphological transitions when
a class of genes, encoding flocculins, is present in their
genome. More specifically, the adhesion of the cells to
substrates, the pseudohyphal and the invasive growth are
S. cerevisiae phenotypes resulting from the expression of
ª 2011 Federation of European Microbiological Societies FEMS Yeast Res 12 (2012) 78–87Published by Blackwell Publishing Ltd. All rights reserved
YEA
ST R
ESEA
RC
H

the FLO11 (also known as MUC1) gene (Lambrechts
et al., 1996; Lo & Dranginis, 1998; Rupp et al., 1999).
Other phenotypes of S. cerevisiae associated with the
FLO11 expression are the formation of mats, complex
colony-like structures on low-density semi-solid medium
(Reynolds & Fink, 2001; Reynolds et al., 2008); the air–liquid interfacial cellular aggregation in the process of
sherry-like wine fermentations, called a flor or velum
(Zara et al., 2005; Ishigami et al., 2006); and the adher-
ence to a range of solid surfaces, such as glass, stainless
steel, agar and plastics, possibly leading to the develop-
ment of biofilms (Reynolds & Fink, 2001; Mortensen
et al., 2007). Biofilm formation is a severe problem in the
medical sector as they offer protection to the cells, for
example, by conferring resistance to antifungal drugs
(Bryers, 2008).
Furthermore, it was observed that the expression of the
FLO11 gene also controls cell–cell adhesion, which leads
to the aggregation of the yeast cells into clumps that sedi-
ment to the bottom of the fermentor (Lo & Dranginis,
1996; Bayly et al., 2005; Douglas et al., 2007). This phe-
nomenon occurs at the end of the beer fermentation,
when almost all fermentable sugars are converted into
ethanol and carbon dioxide and is called flocculation.
Remarkably, the flocculation phenotype caused by
Flo11p was observed only for Saccharomyces cerevisiae
var. diastaticus (Lo & Dranginis, 1996; Bayly et al., 2005;
Douglas et al., 2007), and also microscopic flocs consist-
ing of 6–30 cells could be observed under the microscope
for the laboratory strains BY4741 and BY4742 (S288C)
when Flo11p was overexpressed (Purevdorj-Gage et al.,
2007; Van Mulders et al., 2009). The other phenotypes,
such as invasive growth, pseudohyphal growth, develop-
ment of mats and biofilm formation, are mainly associ-
ated with S. cerevisiae strain Σ1278b, expressing Flo11p
(Lo & Dranginis, 1998; Cullen & Sprague, 2000; Douglas
et al., 2007). However, when overexpressed in S. cerevisiae
strain Σ1278b, a Flo11p-dependent, but calcium-indepen-
dent flocculation behaviour was observed (Guo et al.,
2000), indicating that an identical adhesin expressed in
different yeast strains can exhibit different FLO11-depen-
dent properties. Clearly, other factors must be involved in
these phenomena.
The FLO11 gene in S. cerevisiae encodes the Flo11 pro-
tein, which is a member of the flocculin family. However,
Flo11p is more distantly related to the other flocculins in
this family. Indeed, the predicted protein product of
FLO11 is 37% similar to the product of FLO1 (26% iden-
tical) (Lo & Dranginis, 1996), while the other members
of the flocculin family show a higher degree of similarity
(Jin & Speers, 1998). The FLO11 gene contains 4101 base
pairs and is located on chromosome IX. It encodes a
1367 amino acid cell wall protein assembled by three
major domains: a carboxy (C)-terminal domain possess-
ing a glycosylphosphatidylinositol (GPI) remnant to
anchor the protein in the cell wall, a central domain con-
taining a highly repeated threonine- and serine-rich
sequence and finally an amino (N)-terminal domain
located outside the cell wall (Lambrechts et al., 1996; Lo
& Dranginis, 1996, 1998).
The wide range of phenotypes that are directly attrib-
uted to the FLO11-encoded glycoprotein are based on
adhesion events, and therefore, the binding properties of
Flo11p were investigated in this study. As Flo11p is an
adhesin from the Flo family, where the adhesive proper-
ties are located in the N-terminus of the proteins, only
the N-terminal domain was examined. In the Pfam data-
base, a domain is associated with this part of Flo11p,
which is called the ‘Flo11-domain’ (PF10181), covering
residues 42–195. This domain is conserved in some
related species within the Saccharomycotina (Linder &
Gustafsson, 2008). Moreover, it is suggested that Flo11p
shows a lectin activity that is related to amino acids pres-
ent in the N-terminus, in particular the WQWGT motif
(Bayly et al., 2005; Douglas et al., 2007). The N-terminal
domain of Flo11p was expressed in S. cerevisiae, purified
and studied. The secondary structure of the domain was
evaluated as well as the presence of glycans on the pro-
tein. Our results show that N-Flo11p is not a mannose-
binding protein but can make homotypic interactions.
Materials and methods
Construction of N-FLO11 expression plasmid
The determination of the N-terminal domain of FLO11
was based on the results of Lambrechts et al. (1996). It
was proposed that the N-terminal domain stops where
the tandem repeats start (at aa 210). Two oligonucleo-
tides [Flo11_3, 5′-(TCCTTAGTCAAAAGGTTTCCAACTGCACTAGTTCCAAGA)-3′; and Flo11_4, 5′-(GGA-GATCGGA ATTCGTCAGTGATGGTGATGGTGATGCTT
CGTACCGCCACAATTATTGTCA)-3′] were designed to
amplify the N-terminal domain of Flo11p coding
sequence from the genomic DNA of S. cerevisiae strain
S288C. Primer Flo11_4 contains a sequence for 6 histi-
dines to add a histidine tag to the C-terminal part of the
protein, which will allow purifying the protein using a
Ni-NTA column. The predicted molecular weight of the
expressed protein is 22 kDa. The yeast-Escherichia coli
shuttle vector pYEX-S1 was used for cloning of the N-ter-
minal part of the FLO11 gene. Cloning was performed
using the In-FusionTM method (Clontech, Mountain
View, CA), which is based on the recombination between
identical sequences of the vector and the amplified gene
(Benoit et al., 2006; Zhu et al., 2007). The construction
FEMS Yeast Res 12 (2012) 78–87 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
S. cerevisiae Flo11p does not bind mannose 79

of the expression plasmid was confirmed by a combination
of PCR and DNA sequence analysis (Genetic Service
Facility, VIB, Belgium). In the pYEX-S1 vector, the
cloned gene is regulated by the strong constitutive phos-
phoglycerate kinase (PGK) promoter, and this vector
includes the b-lactamase gene for selection in E. coli and
the yeast-selectable marker URA3. The Flo11p secretion
sequence (aa 1–21) was not included during cloning, as
the pYEX-S1 vector contains the full-length leader
sequence from Kluyveromyces lactis to direct proteins
through the secretory pathway for secretion into the
growth medium. Transformation into E. coli strain DH5awas carried out using a heat shock as described previously
(Sambrook & Russel, 2001). The S. cerevisiae strain
S288C BY4741 was transformed using the lithium acetate
procedure (Ito et al., 1983; Gietz & Schiestl, 2007).
Expression and purification of the N-terminal
domain of Flo11p
Transformed S. cerevisiae cells were cultivated in shake
flasks. The cells were grown in yeast extract-peptone-dex-
trose (YPD) medium containing 1% (w/v) yeast extract,
2% (w/v) peptone and 2% (w/v) D-glucose. Overnight
cultures were grown by inoculating 90 mL YPD medium
with one pYEX-S1-containing S. cerevisiae colony. Cul-
tures were shaken and incubated at 30 °C. Subsequently,the overnight cultures were diluted into 2 L of YPD med-
ium and allowed to grow to an OD600nm of 10 by incuba-
tion at 30 °C for 64 h while shaking (110 r.p.m.). The
cells were separated from the medium by centrifugation
(12 000 g, 20 min, 4 °C), and the resulting supernatant
was adjusted to pH 7.2 to protonate the histidines of the
His-tag. The N-terminal domain of Flo11p was captured
from the growth medium by affinity chromatography
using a nickel-nitrilotriacetic acid (Ni-NTA) Sepharose
column (6 mL) and was eluted with 1 M imidazole in
phosphate-buffered saline. To further purify N-Flo11p,
the elution fractions were pooled, concentrated to 2 mL
(using an Amicon centrifugal filter unit with MWCO of
3000 Da; Millipore, Billerica, MA), and subjected to gel
filtration chromatography (Superdex 75 HR 10/30; GE
Healthcare) in 20 mM Tris, pH 7.5 and 150 mM NaCl.
The eluate of the Ni-column and the purified proteins
from gel filtration were analysed by sodium dodecyl sul-
phate-polyacrylamide gel electrophoresis (SDS-PAGE).
The concentration of pure N-Flo11p was estimated from
the absorption at 280 nm.
Detection of (glyco) proteins
After purifying the N-terminal domain of Flo11p, SDS-
PAGE was performed to visualize the protein and to esti-
mate its molecular mass. The gel (15% resolving gel) was
stained with Coomassie staining [1 g L�1 Coomassie Bril-
liant Blue R250, 50% (v/v) methanol, 10% (v/v) acetic
acid] for 30 min and destained with 50% (v/v) methanol
and 10% (v/v) acetic acid. To visualize glycans, the Pro-Q
Emerald 300 Glycoprotein Gel and Blot Stain kit (Invitro-
gen) was used.
Enzymatic deglycosylation
To remove the N-glycans, the protein was treated with
endo-b-N-acetylglucosaminidase H (Endo H; New Eng-
land Biolabs) at a concentration of 100 U lg�1 protein,
in 20 mM Na-acetate buffer (pH 5.0). The mixture was
incubated at 37 °C for 4 h.
Circular dichroism
Circular dichroism (CD) measurements were performed
with a J715 spectropolarimeter (Jasco) in the far-UV (200
–240 nm) region, using a cell with a path length of
0.1 cm. The protein was diluted in 400 lL phosphate buf-
fer (50 mM at pH 7.5) to obtain a final concentration of
0.3 mg mL�1. Data were acquired with a scanning speed
of 50 nm min�1, a 2-s integration time and a 1-nm band-
width at a constant temperature of 25 °C. Six spectra
were measured and averaged. The mean residual elliptic-
ity, [h], was calculated as follows: [h] = (100 9 hm)
(cL)�1, where h is the observed ellipticity, m is the mean
residual weight, c is the concentration (mg mL�1) and L
is the path length (cm). A quantitative estimation of the
secondary structure composition was facilitated by apply-
ing the K2D2 method and using the SELCON3, CONTINLL
and CDSSTR programs (Provencher & Glockner, 1981; And-
rade et al., 1993; Johnson, 1999; Sreerama et al., 1999;
Perez-Iratxeta & Andrade-Navarro, 2008).
Fluorescence spectroscopy
The binding of D-(+)-mannose to N-Flo11p was studied
by fluorescence spectroscopy. The measurements were
performed at 20 °C, using a luminescence spectrometer
(model LS55; PerkinElmer, MA). An excitation wave-
length of 277 nm, a scan speed of 100 nm min�1 and an
excitation and emission slit width of 5 nm were used
during the experiments. The initial sample volume was
500 lL, containing 2 lM N-Flo11p in flocculation buffer
(50 mM sodium acetate, pH 4.5, 100 mM NaCl and
7 mM CaCl2). One hundred millimolars of mannose
from a concentrated stock solution containing 2 M sugar
was added to the protein solution, then the sample was
mixed and the fluorescence was measured immediately.
As a positive control, 100 mM D-(+)-mannose was added
ª 2011 Federation of European Microbiological Societies FEMS Yeast Res 12 (2012) 78–87Published by Blackwell Publishing Ltd. All rights reserved
80 K.V.Y. Goossens & R.G. Willaert
to N-Flo1p, under exactly the same conditions. Emission
spectra were collected from 300 to 450 nm and corrected
relative to a buffer blank. Averages obtained from three
independent experiments were displayed on the graph.
Surface plasmon resonance
The interaction of the N-terminal domain of Flo11p to
another N-Flo11p was studied by surface plasmon reso-
nance (SPR) using a Biacore 3000TM instrument (GE
Healthcare, Upsalla, Sweden) at 25 °C. The covalent
immobilization of N-Flo11p in flow cell 2 (FC 2) on the
CM5 BIAsensor chip was accomplished via amine cou-
pling chemistry, using an amino coupling kit (BIAcore;
GE Healthcare) with 20 mM sodium acetate pH 4.0 as
immobilization buffer. Glycosylated N-Flo11p was immo-
bilized to a response around 95 resonance units (RU).
Flow cell 1 (FC 1) was treated in the same way as FC 2
except that it was blocked immediately after activation
and was used as the reference surface to subtract the RU
as a result of the refractive index of the protein solution
and any instrument noise.
Increasing concentrations ranging from 11 nM to
90 lM of glycosylated N-Flo11p were allowed to flow
across both flow cells FC 1 and FC 2 of the CM5 chip
containing N-Flo11p. The experiment was carried out in
HBS buffer [20 mM Hepes pH 7.4, 150 mM NaCl,
10 mM CaCl2 and 0.005% (v/v) Tween 20]. No binding
was detected in the control flow cells. Complete dissocia-
tion of N-Flo11p was performed with HBS buffer before
starting a new binding cycle. The association time was
3 min, and the dissociation time was 7 min. Additionally,
a zero concentration cycle of N-Flo11p (injection of run-
ning buffer) was run to determine the effect of the buffer.
All measurements were performed at a flow rate of
20 lL min�1 in HBS buffer, and the proteins were dialy-
sed to exactly the same buffer (Spectra/Por® Dialysis
membrane, 3500 Da MWCO; Spectrum Laboratories,
Inc., CA). The buffer was filtered through a 0.45-lm filter
(MF-MilliporeTM membrane filter) and degassed before
usage.
The results were analysed with the BIAevaluation soft-
ware version 4.1 (GE Healthcare). Binding was observed
by measuring the increase in RU when injecting different
concentrations of the analyte and after subtracting the
background response.
Results
Expression and purification of N-Flo11p
N-Flo11p was captured from the growth medium using a
nickel column and was eluted with imidazole. The eluate
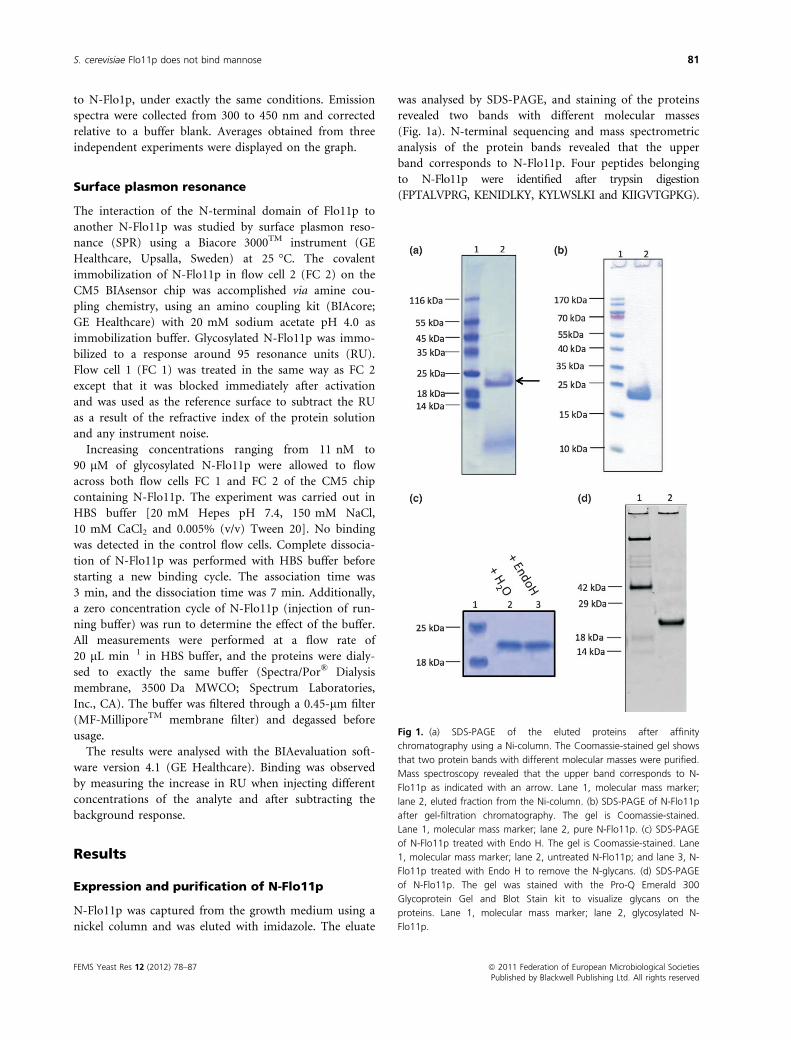
was analysed by SDS-PAGE, and staining of the proteins
revealed two bands with different molecular masses
(Fig. 1a). N-terminal sequencing and mass spectrometric
analysis of the protein bands revealed that the upper
band corresponds to N-Flo11p. Four peptides belonging
to N-Flo11p were identified after trypsin digestion
(FPTALVPRG, KENIDLKY, KYLWSLKI and KIIGVTGPKG).
Fig 1. (a) SDS-PAGE of the eluted proteins after affinity
chromatography using a Ni-column. The Coomassie-stained gel shows
that two protein bands with different molecular masses were purified.
Mass spectroscopy revealed that the upper band corresponds to N-
Flo11p as indicated with an arrow. Lane 1, molecular mass marker;
lane 2, eluted fraction from the Ni-column. (b) SDS-PAGE of N-Flo11p
after gel-filtration chromatography. The gel is Coomassie-stained.
Lane 1, molecular mass marker; lane 2, pure N-Flo11p. (c) SDS-PAGE
of N-Flo11p treated with Endo H. The gel is Coomassie-stained. Lane
1, molecular mass marker; lane 2, untreated N-Flo11p; and lane 3, N-
Flo11p treated with Endo H to remove the N-glycans. (d) SDS-PAGE
of N-Flo11p. The gel was stained with the Pro-Q Emerald 300
Glycoprotein Gel and Blot Stain kit to visualize glycans on the
proteins. Lane 1, molecular mass marker; lane 2, glycosylated N-
Flo11p.
FEMS Yeast Res 12 (2012) 78–87 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
S. cerevisiae Flo11p does not bind mannose 81
To further purify N-Flo11p, the elution fractions were
subjected to gel filtration chromatography. The purified
protein was visualized with SDS-PAGE (Fig. 1b), and the
concentration was estimated from the absorption at
280 nm. The yield obtained with this purification proto-
col was about 0.5 mg L�1 culture.
The N-terminal domain of Flo11p
is O-glycosylated
The calculated molecular mass of the N-terminal domain
of Flo11p without glycosylation is 22 kDa. On the SDS
gel (Fig. 1b), it is observed that N-Flo11p corresponds to
this molecular mass, which indicates that the protein is
not N-glycosylated. This observation is in accordance
with the prediction of N-glycosylation sites based on the
amino acid sequence of the N-terminal domain of Flo11p
(data not shown). To confirm the absence of N-glycans
on N-Flo11p, the protein was treated with Endo H and
visualized with SDS-PAGE (Fig. 1c). No decrease in
molecular mass of N-Flo11p is observed. Hence, it is con-
cluded that no N-glycans are present on the N-terminal
domain of Flo11p.
O-glycosylation often occurs in a region of the protein
that contains a high proportion of serine and threonine,
with neighbouring proline residues (Lehle & Bause,
1984). In the amino acid sequence of the N-terminal
domain of Flo11p, several threonine (18), serine (14) and
proline (7) residues are found. To verify whether
N-Flo11p contains O-glycans, the SDS gel containing
N-Flo11p was stained with the Pro-Q Emerald 300 Glyco-
protein Gel and Blot Stain kit (Fig. 1d). This staining
procedure allows the visualization of glycans. As
N-Flo11p can be visualized on the SDS gel, it is con-
cluded that N-Flo11p is O-glycosylated.
N-Flo11p is mainly composed of b-sheets
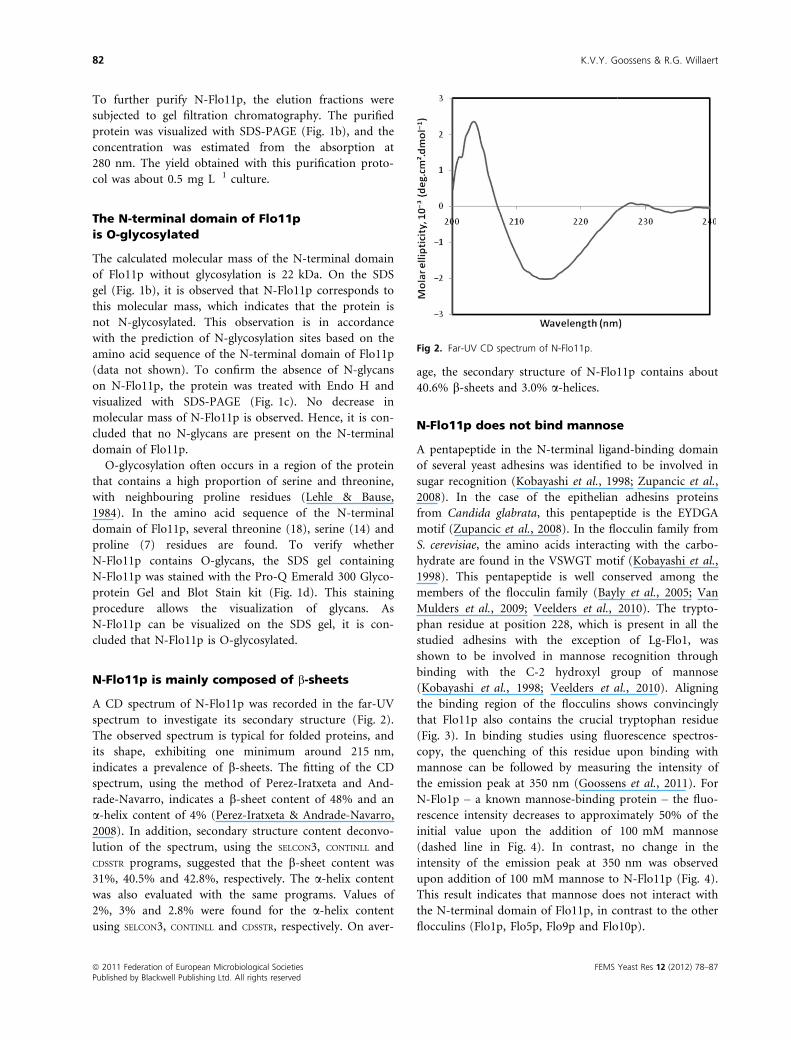
A CD spectrum of N-Flo11p was recorded in the far-UV
spectrum to investigate its secondary structure (Fig. 2).
The observed spectrum is typical for folded proteins, and
its shape, exhibiting one minimum around 215 nm,
indicates a prevalence of b-sheets. The fitting of the CD
spectrum, using the method of Perez-Iratxeta and And-
rade-Navarro, indicates a b-sheet content of 48% and an
a-helix content of 4% (Perez-Iratxeta & Andrade-Navarro,
2008). In addition, secondary structure content deconvo-
lution of the spectrum, using the SELCON3, CONTINLL and
CDSSTR programs, suggested that the b-sheet content was
31%, 40.5% and 42.8%, respectively. The a-helix content
was also evaluated with the same programs. Values of
2%, 3% and 2.8% were found for the a-helix content
using SELCON3, CONTINLL and CDSSTR, respectively. On aver-
age, the secondary structure of N-Flo11p contains about
40.6% b-sheets and 3.0% a-helices.
N-Flo11p does not bind mannose
A pentapeptide in the N-terminal ligand-binding domain
of several yeast adhesins was identified to be involved in
sugar recognition (Kobayashi et al., 1998; Zupancic et al.,
2008). In the case of the epithelian adhesins proteins
from Candida glabrata, this pentapeptide is the EYDGA
motif (Zupancic et al., 2008). In the flocculin family from
S. cerevisiae, the amino acids interacting with the carbo-
hydrate are found in the VSWGT motif (Kobayashi et al.,
1998). This pentapeptide is well conserved among the
members of the flocculin family (Bayly et al., 2005; Van
Mulders et al., 2009; Veelders et al., 2010). The trypto-
phan residue at position 228, which is present in all the
studied adhesins with the exception of Lg-Flo1, was
shown to be involved in mannose recognition through
binding with the C-2 hydroxyl group of mannose
(Kobayashi et al., 1998; Veelders et al., 2010). Aligning
the binding region of the flocculins shows convincingly
that Flo11p also contains the crucial tryptophan residue
(Fig. 3). In binding studies using fluorescence spectros-
copy, the quenching of this residue upon binding with
mannose can be followed by measuring the intensity of
the emission peak at 350 nm (Goossens et al., 2011). For
N-Flo1p – a known mannose-binding protein – the fluo-
rescence intensity decreases to approximately 50% of the
initial value upon the addition of 100 mM mannose
(dashed line in Fig. 4). In contrast, no change in the
intensity of the emission peak at 350 nm was observed
upon addition of 100 mM mannose to N-Flo11p (Fig. 4).
This result indicates that mannose does not interact with
the N-terminal domain of Flo11p, in contrast to the other
flocculins (Flo1p, Flo5p, Flo9p and Flo10p).
Fig 2. Far-UV CD spectrum of N-Flo11p.
ª 2011 Federation of European Microbiological Societies FEMS Yeast Res 12 (2012) 78–87Published by Blackwell Publishing Ltd. All rights reserved
82 K.V.Y. Goossens & R.G. Willaert
N-Flo11p molecules self-interact
Recently, a homotypic adhesive mechanism has been
detected for Flo11p (Douglas et al., 2007). The whole
Flo11 protein was purified and covalently attached to
microscopic beads. The Flo11p-coated beads were able to
bind S. cerevisiae var. diastaticus expressing Flo11p, but
not to cells that do not express Flo11p. Therefore, Flo11p
itself was considered as an important adhesive target on
the cell wall for Flo11p, if not the only target (Douglas
et al., 2007).
In our study, we have assessed the homotypic interac-
tion of the N-terminal domain of Flo11p. SPR was used
to determine qualitatively the binding between N-Flo11
proteins. Therefore, N-Flo11p was coupled to a BIAsensor
chip, and increasing concentrations of N-Flo11p (11 nM,
88 nM, 350 nM, 5.6 lM, 11.2 lM, 22.5 lM, 45 lM and
90 lM) were flown over that chip allowing the proteins
to interact. The clear concentration-dependent increase in
signal indicates that N-Flo11p is able to bind to N-
Flo11p, which is called a homotypic interaction (Fig. 5).
Discussion
The flocculin encoded by the FLO11 gene is responsible
for a wide variety of phenotypes exhibited by S. cerevisiae
cells expressing this Flo11p. These include haploid inva-
sive growth (Roberts & Fink, 1994; Cullen & Sprague,
2000), development of pseudohyphae (Gimeno et al.,
1992; Kron et al., 1994; Mosch, 2000), biofilm formation
(Reynolds & Fink, 2001), mat formation (Reynolds &
Fink, 2001; Reynolds et al., 2008), flor formation (Zara
et al., 2005; Ishigami et al., 2006), adhesion to surfaces
(Lo & Dranginis, 1998; Reynolds & Fink, 2001; Morten-
sen et al., 2007) and flocculation (Lo & Dranginis, 1996;
Bayly et al., 2005). These cellular adhesion phenomena
are mediated by Flo11p, but the mechanism is not well
understood at the molecular level and very little is known
about the structure and function of the protein. There-
fore, in this study, this versatile flocculin was purified
and studied.
The members of the flocculin family are composed of
three domains, and the binding activity is located in the
N-terminal part (Bony et al., 1997; Kobayashi et al.,
1998; Goossens et al., 2011). Other adhesins from both
pro- and eukaryotic origin also possess their binding
Fig 4. Emission spectrum of N-Flo11p compared with the N-Flo11p
spectrum with the addition of mannose from a stock solution to a
final concentration of 100 mM. Averages of three independent
experiments are shown. As a positive control, the decrease of the
fluorescence signal upon addition of 100 mM mannose to a known
mannose-binding protein (N-Flo1p) is shown in the graph.
Fig 5. SPR sensorgram of N-Flo11p–N-Flo11p interactions N-Flo11p
was covalently coupled to a CM5 chip at 95 relative units (RU), and
an N-Flo11p solution with the indicated concentrations was injected.
A concentration-dependent interaction between immobilized N-
Flo11p and injected N-Flo11p at different concentrations (11 nM,
88 nM, 350 nM, 56 lM, 112 lM, 225 lM, 45 lM and 90 lM) was
observed.
Fig 3. Multiple sequence alignment of the amino acid sequences
involved in sugar recognition of the Flo1, Flo5, Flo9, Flo10 and Flo11
adhesins from Saccharomyces cerevisiae. The region corresponding to
the pentapeptide, containing the amino acids that interact with the
carbohydrate (Kobayashi et al., 1998; Zupancic et al., 2008), is
marked as a grey box. The conserved tryptophan residue is outlined
in red.
FEMS Yeast Res 12 (2012) 78–87 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
S. cerevisiae Flo11p does not bind mannose 83

domain at the distal tip of the protein. For example,
FimH is the lectin domain at the tip of type 1 pili from
E. coli and recognizes terminal mannose units of uropla-
kin Ia, a membrane glycoprotein that is abundantly
expressed on superficial epithelial umbrella cells of the
urinary tract (Zhou et al., 2001); the N-terminal part of
Als1p from Candida albicans interacts with extracellular
matrix proteins, epithelial and endothelial cells (Loza
et al., 2004; Sheppard et al., 2004; Donohue et al., 2011).
Also, it was suggested that Flo11p shows a lectin activity
that is related to amino acids present in the N-terminus
(Bayly et al., 2005; Douglas et al., 2007). For these rea-
sons, only the N-terminal domain of Flo11p was cloned,
expressed and purified from S. cerevisiae to study its
structural and binding properties.
In this study, the purified N-Flo11p was analysed by
SDS-PAGE, and it was found that this domain is deco-
rated with O-glycans and does not contain N-glycans.
Moreover, the secondary structure was evaluated by CD.
By analysing the spectrum, we calculated that the sec-
ondary structure of N-Flo11p contains about 40.6% b-sheets and 3.0% a-helices. These values are comparable
with the percentages found for N-Flo1p (37.7% b-sheetsand 6.5% a-helices) (Goossens et al., 2011). The preva-
lence of b-sheets in the N-terminal domain is typical for
flocculins. This part of most of the flocculins has been
assigned to a new domain family (PF07691) in Pfam
(Finn et al., 2010) (http://pfam.sanger.ac.uk), called the
PA14 domain (Rigden et al., 2004). The PA14 domain is
a conserved domain that has been discovered in a wide
variety of bacterial and eukaryotic proteins, which
include many glycosidases, glycosyltransferases, proteases,
amidases, bacterial toxins, such as anthrax protective
antigen (PA), and also yeast adhesins. Most of the exper-
imentally characterized PA14-containing proteins are
involved in carbohydrate binding and/or metabolism.
Therefore, the N-terminal domains of the member pro-
teins of the flocculin family were considered as one of
the many PA14 domain variants (Rigden et al., 2004).
The anthrax toxin PA also contains a PA14 domain, and
the crystal structure of this toxin revealed that this
domain consists of a series of antiparallel b-strands(Petosa et al., 1997). Although a PA14 domain was not
predicted for Flo11p (Goossens & Willaert, 2010), the
secondary fold of the N-terminal part of the protein is
also dominated by b-sheets.The fact that the Flo11 flocculin does not contain this
PA14 domain indicates that no lectin activity was pre-
dicted. Indeed, by fluorescence spectroscopy, it was
shown that N-Flo11p is not able to bind mannose
(Fig. 4). Glycan array screening for N-Flo11p confirmed
this result (Consortium for Functional Glycomics, www.
functionalglycomics.org, request 2078 and 1224, supplying
investigator: Lars-Oliver Essen, Department of Chemistry/
Biochemistry, Philipps-Universitat Marburg, Germany).
The Flo11 protein was tested for its interaction with 465
different glycan structures (version 4.1) printed on an
array (request 2078) and 377 glycan structures (version
3.1) (request 1224), but no conclusive data were obtained
for glycan interaction analysis of N-Flo11p with and with-
out the GST tag, N-Flo11p with a His-tag and after
dimerization of N-Flo11p. No binding to mannose or to
any other glycans from the array was detected. The detec-
tion was based on fluorescence as the adhesin was directly
labelled with the fluorophore ‘Alexa 488’. Other members
of the flocculin family, however, are able to bind man-
nose, and their N-terminal domains correspond to the
PA14 domain (Rigden et al., 2004). Indeed, a clear lectin
activity was demonstrated for N-Flo1p and N-Flo5p
(Veelders et al., 2010; Goossens et al., 2011).
In contrast, Douglas et al. (2007) indirectly detected
mannose-binding activity for Flo11p. The whole Flo11
protein was purified from the growth medium after over-
expression in S. cerevisiae var. diastaticus or in S. cerevisi-
ae Σ1278b. It was observed that beads coated with
Flo11p were able to bind S. cerevisiae var. diastaticus cells
expressing Flo11p. This interaction was inhibited by the
addition of a high (1 M) concentration of mannose,
which suggests that Flo11p is a member of the Flo1-type
flocculins and functions as a lectin in S. cerevisiae var.
diastaticus (Douglas et al., 2007). Moreover, by studying
the interaction between the coated beads and yeast cells,
it was put forward that Flo11p shows a homotypic inter-
action (Douglas et al., 2007). Indeed, the beads could
only interact with yeast cells that express Flo11p, suggest-
ing that Flo11p itself is an important part of the adhesive
target on the cell wall, if not the only target.
In this study, the homotypic binding of Flo11 proteins
was confirmed by SPR as a clear and concentration-
dependent interaction of N-Flo11p molecules was
observed (Fig. 5). The mechanism of this interaction can-
not be based on a lectin–carbohydrate interaction. The
protein–protein interaction could be based on electro-
static and hydrophobic interactions, and influenced by
the presence of the O-glycans (Fig. 1). Also, yeast cells
expressing Flo11p have a highly hydrophobic character
(Reynolds & Fink, 2001; Van Mulders et al., 2009),
although the link between the hydrophobicity of the cell
wall and the presence of expressed Flo11p on the cell wall
is not yet clear. Recently, it was suggested that the whole
Flo11 protein has a hydrophilic character (Karunanithi
et al., 2010).
Recently, it has been demonstrated that many adhesins
from budding yeasts contain amyloid-forming sequences
that can play a role in cell–cell interactions (Otoo et al.,
2008; Alsteens et al., 2010; Frank et al., 2010; Nobbs
ª 2011 Federation of European Microbiological Societies FEMS Yeast Res 12 (2012) 78–87Published by Blackwell Publishing Ltd. All rights reserved
84 K.V.Y. Goossens & R.G. Willaert

et al., 2010; Ramsook et al., 2010; Garcia et al., 2011).
These adhesins contain sequences with high b-aggregationpotential; VVSTTV and VTTAVTTTVV were indicated
for the full Flo11p (Ramsook et al., 2010) (these
sequences are, however, not present in the N-terminal
part of Flo11p). Experimental results indicate that amy-
loid formation leads to the bundling of adhesins into
nanodomains, which can greatly increase the intercellular
binding strength by increasing the avidity (Douglas et al.,
2007; Ramsook et al., 2010) and, consequently, can rein-
force the Flo11p homotypic interactions between cells. It
has been shown that an applied force can induce the for-
mation of these nanodomains (Alsteens et al., 2010) and
that they can be regulated by exogenous peptides (Garcia
et al., 2011). Experimental results also suggest that amy-
loids are formed between adhesion molecules on contact-
ing cells (Rauceo et al., 2004; Ramsook et al., 2010). Such
intercellular amyloids would also strengthen the protein–protein intercellular adhesive bonds.
In conclusion, this study describes for the first time the
purification and subsequent characterization of the N-ter-
minal domain of Flo11p, a flocculin exhibiting a plethora
of phenotypes. SPR analysis showed a homotypic interac-
tion. However, in contrast to the other Flo proteins, N-
Flo11p does not bind mannose. The exact mechanism of
this protein–protein interaction and the involvement of
the O-glycans have still to be revealed.
Acknowledgements
We thank Prof. J. E. Edwards (Division of Infectious Dis-
eases, Medicine Deptartment, Harbor-UCLA, CA, USA)
for providing us with the pYEX-S1 vector. We also
acknowledge Msc. Catherine Stassen and Prof. Dr. Bart
Devreese (L-Probe, Ghent University) for the mass spec-
trometry analysis. This work was supported by the Euro-
pean Space Agency (Prodex program), the Institute for
the promotion of Innovation by Science and Technology
in Flanders (IWT) and the Research Council of the VUB.
References
Alsteens D, Garcia MC, Lipke PN & Dufrene YF (2010) Force-
induced formation and propagation of adhesion
nanodomains in living fungal cells. P Natl Acad Sci USA
107: 20744–20749.Andrade MA, Chacon P, Merelo JJ & Moran F (1993)
Evaluation of secondary structure of proteins from UV
circular dichroism spectra using an unsupervised learning
neural network. Protein Eng 6: 383–390.Bayly JC, Douglas LM, Pretorius IS, Bauer FF & Dranginis AM
(2005) Characteristics of Flo11-dependent flocculation in
Saccharomyces cerevisiae. FEMS Yeast Res 5: 1151–1156.
Benoit RM, Wilhelm RN, Scherer-Becker D & Ostermeier C
(2006) An improved method for fast, robust, and seamless
integration of DNA fragments into multiple plasmids.
Protein Expr Purif 45: 66–71.Bony M, Thines-Sempoux D, Barre P & Blondin B (1997)
Localization and cell surface anchoring of the Saccharomyces
cerevisiae flocculation protein Flo1p. J Bacteriol 179: 4929–4936.
Bryers JD (2008) Medical biofilms. Biotechnol Bioeng 100: 1–18.
Cullen PJ & Sprague GF Jr (2000) Glucose depletion causes
haploid invasive growth in yeast. P Natl Acad Sci USA 97:
13619–13624.Dickinson JR (2008) Filament formation in Saccharomyces
cerevisiae – a review. Folia Microbiol (Praha) 53: 3–14.Donohue DS, Ielasi F, Goossens KY & Willaert RG (2011) The
Als1 protein from Candida albicans specifically binds
fucose-containing glycans. Mol Microbiol 80: 1667–1679.Douglas LM, Li L, Yang Y & Dranginis AM (2007) Expression
and characterization of the flocculin Flo11/Muc1, a
Saccharomyces cerevisiae mannoprotein with homotypic
properties of adhesion. Eukaryot Cell 6: 2214–2221.Finn RD, Mistry J, Tate J et al. (2010) The Pfam protein
families database. Nucleic Acids Res 38: D211–D222.Frank AT, Ramsook C, Otoo HN, Tan C, Soybelman G,
Rauceo JM, Gaur NK, Klotz SA & Lipke PN (2010)
Structure and function of glycosylated tandem repeats
from Candida albicans Als adhesins. Eukaryot Cell 9: 405–414.
Garcia MC, Lee JT, Ramsook CB, Alsteens D, Dufrene YF &
Lipke PN (2011) A role for amyloid in cell aggregation and
biofilm formation. PLoS ONE 6: e17632.
Gietz RD & Schiestl RH (2007) High-efficiency yeast
transformation using the LiAc/SS carrier DNA/PEG method.
Nat Protoc 2: 31–34.Gimeno CJ, Ljungdahl PO, Styles CA & Fink GR (1992)
Unipolar cell divisions in the yeast S. cerevisiae lead to
filamentous growth: regulation by starvation and RAS. Cell
68: 1077–1090.Goossens K & Willaert R (2010) Flocculation protein structure
and cell-cell adhesion mechanism in Saccharomyces
cerevisiae. Biotechnol Lett 32: 1571–1585.Goossens KV, Stassen C, Stals I, Donohue DS, Devreese B, De
Greve H & Willaert RG (2011) The N-terminal domain of
the Flo1 flocculation protein from Saccharomyces cerevisiae
binds specifically to mannose carbohydrates. Eukaryot Cell
10: 110–117.Guo B, Styles CA, Feng Q & Fink GR (2000) A Saccharomyces
gene family involved in invasive growth, cell-cell adhesion,
and mating. P Natl Acad Sci USA 97: 12158–12163.Iserentant D (1996) Practical aspects of yeast flocculation.
Cerevisiae 21: 30–33.Ishigami M, Nakagawa Y, Hayakawa M & Iimura Y (2006)
FLO11 is the primary factor in flor formation caused by cell
surface hydrophobicity in wild-type flor yeast. Biosci
Biotechnol Biochem 70: 660–666.
FEMS Yeast Res 12 (2012) 78–87 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
S. cerevisiae Flo11p does not bind mannose 85

Ito H, Fukuda Y, Murata K & Kimura A (1983)
Transformation of intact yeast cells treated with alkali
cations. J Bacteriol 153: 163–168.Jin Y-L & Speers RA (1998) Flocculation of Saccharomyces
cerevisiae. Food Res Int 31: 421–440.Johnson WC (1999) Analyzing protein circular dichroism
spectra for accurate secondary structures. Proteins 35: 307–312.
Karunanithi S, Vadaie N, Chavel CA, Birkaya B, Joshi J, Grell
L & Cullen PJ (2010) Shedding of the mucin-like flocculin
Flo11p reveals a new aspect of fungal adhesion regulation.
Curr Biol 20: 1389–1395.Kobayashi O, Hayashi N, Kuroki R & Sone H (1998) Region
of Flo1 proteins responsible for sugar recognition. J
Bacteriol 180: 6503–6510.Kron SJ, Styles CA & Fink GR (1994) Symmetric cell division
in pseudohyphae of the yeast Saccharomyces cerevisiae. Mol
Biol Cell 5: 1003–1022.Lambrechts MG, Bauer FF, Marmur J & Pretorius IS (1996)
Muc1, a mucin-like protein that is regulated by Mss10, is
critical for pseudohyphal differentiation in yeast. P Natl
Acad Sci USA 93: 8419–8424.Lehle L & Bause E (1984) Primary structural requirements for
N- and O-glycosylation of yeast mannoproteins. Biochim
Biophys Acta 799: 246–251.Linder T & Gustafsson CM (2008) Molecular phylogenetics of
ascomycotal adhesins–a novel family of putative cell-surface
adhesive proteins in fission yeasts. Fungal Genet Biol 45:
485–497.Lo WS & Dranginis AM (1996) FLO11, a yeast gene related to
the STA genes, encodes a novel cell surface flocculin. J
Bacteriol 178: 7144–7151.Lo WS & Dranginis AM (1998) The cell surface flocculin
Flo11 is required for pseudohyphae formation and invasion
by Saccharomyces cerevisiae. Mol Biol Cell 9: 161–171.Loza L, Fu Y, Ibrahim AS, Sheppard DC, Filler SG & Edwards
JE Jr (2004) Functional analysis of the Candida albicans
ALS1 gene product. Yeast 21: 473–482.Mortensen HD, Dupont K, Jespersen L, Willats WG &
Arneborg N (2007) Identification of amino acids involved in
the Flo11p-mediated adhesion of Saccharomyces cerevisiae to
a polystyrene surface using phage display with competitive
elution. J Appl Microbiol 103: 1041–1047.Mosch HU (2000) Pseudohyphal development of
Saccharomyces cerevisiae. Contrib Microbiol 5: 185–200.Nobbs AH, Vickerman MM & Jenkinson HF (2010)
Heterologous expression of Candida albicans cell wall-
associated adhesins in Saccharomyces cerevisiae reveals
differential specificities in adherence and biofilm formation
and in binding oral Streptococcus gordonii. Eukaryot Cell 9:
1622–1634.Otoo HN, Lee KG, Qiu W & Lipke PN (2008) Candida
albicans Als adhesins have conserved amyloid-forming
sequences. Eukaryot Cell 7: 776–782.Palkova Z (2004) Multicellular microorganisms: laboratory
versus nature. EMBO Rep 5: 470–476.
Perez-Iratxeta C & Andrade-Navarro MA (2008) K2D2:
estimation of protein secondary structure from circular
dichroism spectra. BMC Struct Biol 8: 25.
Petosa C, Collier RJ, Klimpel KR, Leppla SH & Liddington RC
(1997) Crystal structure of the anthrax toxin protective
antigen. Nature 385: 833–838.Provencher SW & Glockner J (1981) Estimation of globular
protein secondary structure from circular dichroism.
Biochemistry 20: 33–37.Purevdorj-Gage B, Orr ME, Stoodley P, Sheehan KB & Hyman
LE (2007) The role of FLO11 in Saccharomyces cerevisiae
biofilm development in a laboratory based flow-cell system.
FEMS Yeast Res 7: 372–379.Ramsook C, Tan C, Garcia MC et al. (2010) Yeast cell
adhesion molecules have functional amyloid-forming
sequences. Eukaryot Cell 9: 393–404.Rauceo JM, Gaur NK, Lee KG, Edwards JE, Klotz SA & Lipke
PN (2004) Global cell surface conformational shift mediated
by a Candida albicans adhesin. Infect Immun 72: 4948–4955.Reynolds TB & Fink GR (2001) Bakers’ yeast, a model for
fungal biofilm formation. Science 291: 878–881.Reynolds TB, Jansen A, Peng X & Fink GR (2008) Mat
formation in Saccharomyces cerevisiae requires nutrient and
pH gradients. Eukaryot Cell 7: 122–130.Rigden DJ, Mello LV & Galperin MY (2004) The PA14
domain, a conserved all-beta domain in bacterial toxins,
enzymes, adhesins and signaling molecules. Trends Biochem
Sci 29: 335–339.Roberts RL & Fink GR (1994) Elements of a single MAP
kinase cascade in Saccharomyces cerevisiae mediate two
developmental programs in the same cell type: mating and
invasive growth. Genes Dev 8: 2974–2985.Rupp S, Summers E, Lo HJ, Madhani H & Fink G (1999)
MAP kinase and cAMP filamentation signaling pathways
converge on the unusually large promoter of the yeast
FLO11 gene. EMBO J 18: 1257–1269.Sambrook J & Russel DW (2001) Preparation and
transformation of competent E. coli using calcium chloride.
Molecular Cloning: A Laboratory Manual (Sambrook J &
Russel DW, eds), pp. 1116–1118. Cold Spring Harbor
Laboratory Press, New York.
Sheppard DC, Yeaman MR, Welch WH, Phan QT, Fu Y,
Ibrahim AS, Filler SG, Zhang M, Waring AJ & Edwards JE Jr
(2004) Functional and structural diversity in the Als protein
family of Candida albicans. J Biol Chem 279: 30480–30489.Sreerama N, Venyaminov SY & Woody RW (1999) Estimation
of the number of alpha-helical and beta-strand segments in
proteins using circular dichroism spectroscopy. Protein Sci
8: 370–380.Van Mulders SE, Christianen E, Saerens SM, Daenen L,
Verbelen PJ, Willaert R, Verstrepen KJ & Delvaux FR (2009)
Phenotypic diversity of Flo protein family-mediated
adhesion in Saccharomyces cerevisiae. FEMS Yeast Res 9:
178–190.Veelders M, Bruckner S, Ott D, Unverzagt C, Mosch HU &
Essen LO (2010) Structural basis of flocculin-mediated
ª 2011 Federation of European Microbiological Societies FEMS Yeast Res 12 (2012) 78–87Published by Blackwell Publishing Ltd. All rights reserved
86 K.V.Y. Goossens & R.G. Willaert

social behavior in yeast. P Natl Acad Sci USA 107: 22511–22516.
Zara S, Bakalinsky AT, Zara G, Pirino G, Demontis MA &
Budroni M (2005) FLO11-based model for air-liquid
interfacial biofilm formation by Saccharomyces cerevisiae.
Appl Environ Microbiol 71: 2934–2939.Zhou G, Mo WJ, Sebbel P, Min G, Neubert TA, Glockshuber
R, Wu XR, Sun TT & Kong XP (2001) Uroplakin Ia is the
urothelial receptor for uropathogenic Escherichia coli:
evidence from in vitro FimH binding. J Cell Sci 114: 4095–4103.
Zhu B, Cai G, Hall EO & Freeman GJ (2007) In-fusion
assembly: seamless engineering of multidomain fusion
proteins, modular vectors, and mutations. BioTechniques 43:
354–359.Zupancic ML, Frieman M, Smith D, Alvarez RA, Cummings
RD & Cormack BP (2008) Glycan microarray analysis of
Candida glabrata adhesin ligand specificity. Mol Microbiol
68: 547–559.
FEMS Yeast Res 12 (2012) 78–87 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
S. cerevisiae Flo11p does not bind mannose 87

![[123doc vn] - tim-hieu-nam-men-saccharomyces-cerevisiae](https://static.fdokumen.com/doc/165x107/6345cd51f474639c9b0502af/123doc-vn-tim-hieu-nam-men-saccharomyces-cerevisiae.jpg)


















