
Novel flavohemoglobins of mycobacteria
9
Research Communication Novel Flavohemoglobins of Mycobacteria Sanjay Gupta 1 , Sudesh Pawaria 1 , Changyuan Lu 2 , Syun-Ru Yeh 2 and Kanak L. Dikshit 1 1 Institute of Microbial Technology, Sector 39 A, CSIR, Chandigarh 160036, India 2 Department of Physiology and Biophysics, Albert Einstein College of Medicine, Bronx, NY 10461 Summary Flavohemoglobins (flavoHbs) constitute a distinct class of chimeric hemoglobins in which a globin domain is coupled with a ferredoxin reductase such as FAD- and NADH-binding modules. Structural features and active site of heme and reduc- tase domains are highly conserved in various flavoHbs. A new class of flavoHbs, displaying crucial differences in functionally conserved regions of heme and reductase domains, have been identified in mycobacteria. Mining of microbial genome data indicated that the occurrence of such flavoHbs might be re- stricted to a small group of microbes unlike conventional fla- voHbs that are widespread among prokaryotes and lower eukaryotes. One of the representative flavoHbs of this class, encoded by Rv0385 gene (MtbFHb) of Mycobacterium tubercu- losis, has been cloned, expressed, and characterized. The ferric and deoxy spectra of MtbFHb displayed a hexacoordinate state indicating that its distal site may be occupied by an intrinsic amino acid or an external ligand and it may not be involved in nitric oxide detoxification. Phylogenetic analysis revealed that mycobacterial flavoHbs constitute a separate cluster distinct from conventional flavoHbs and may have novel function(s). Ó 2011 IUBMB IUBMB Life, 63(5): 337–345, 2011 Keywords flavohemoglobin; mycobacteria; electron-transfer; phylog- eny; nitric-oxide dioxygenase. INTRODUCTION Flavohemoglobins (flavoHbs) are monomeric proteins having a classical 3/3 helical globin domain linked with a flavin ade- nine dinucleotide (FAD)-binding reductase domain that may transfer electrons to the heme iron and allow its rapid re-reduc- tion to maintain it in the ferrous state for the rebinding of oxy- gen (1–3). Rapid electron transfer via redox domain may allow these two-domain globins to carry out diverse redox reactions that may be vital for their native host. Recent upsurge in avail- ability of genomic data from various organisms has indicated that gene encoding for flavoHb may be widely distributed in unicellular organisms, both prokaryotes and lower eukaryotes (1, 2, 4); however, none has been found so far in higher organ- isms suggesting that flavoHbs may be specifically relevant to the cellular metabolism of microbes. Physiological function(s) of flavoHbs has been the subject of intense debate over the last decade. Several putative functions have been proposed for fla- voHbs (5–8). However, accumulating experimental evidences support its role in protection of microorganisms from the delete- rious effects of nitric oxide (NO) by detoxifying it to nitrate via the NO-dioxygenation reaction (1, 2, 7, 8). The hydrophobic distal pocket of flavoHbs has been found much more flexible and expandable than conventional Hbs (3, 9) that allow them to accommodate large apolar molecules including phospholipids and perform other cellular functions. Existence of more than one flavoHb in some bacteria, yeasts, and fungi (10, 11) suggest that these hemoglobins (Hbs) may be involved in diverse cellu- lar functions. FlavoHbs usually function as an integral part of stress response and virulence of several pathogenic bacteria and fungi by maintaining the cell redox homeostasis at the aerobic/anaero- bic interface when cells are exposed to various environmental stresses (7, 12). Although importance of flavoHbs for the para- sitic life has been established in several cases (13, 14), none has been reported from pathogenic or nonpathogenic mycobacteria so far. When genomic data of various mycobacterial species was examined, occurrence of multiple flavoHb encoding genes has been detected in many of them. After amino-acid sequence alignment and comparison of mycobacterial flavoHbs, we identi- fied a novel class of flavoHb, exhibiting unconventional heme and reductase domains, in mycobacteria apart from conventional flavoHb. Occurrence of two distinct classes of flavoHbs in mycobacteria is interesting as well as intriguing. Notably, the new class of flavoHb is present in majority of mycobacterial species and it also coexists along with a conventional flavoHb in Address correspondence to: Kanak L. Dikshit, Institute of Microbial Technology, CSIR, Sector 39A, Chandigarh 160036, India. E-mail: [email protected] Received 17 January 2011; accepted 28 February 2011 ISSN 1521-6543 print/ISSN 1521-6551 online DOI: 10.1002/iub.460 IUBMB Life, 63(5): 337–345, May 2011
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Novel flavohemoglobins of mycobacteria

Research Communication
Novel Flavohemoglobins of Mycobacteria
Sanjay Gupta1, Sudesh Pawaria1, Changyuan Lu2, Syun-Ru Yeh2 and Kanak L. Dikshit11Institute of Microbial Technology, Sector 39 A, CSIR, Chandigarh 160036, India2Department of Physiology and Biophysics, Albert Einstein College of Medicine, Bronx, NY 10461
Summary
Flavohemoglobins (flavoHbs) constitute a distinct class ofchimeric hemoglobins in which a globin domain is coupledwith a ferredoxin reductase such as FAD- and NADH-bindingmodules. Structural features and active site of heme and reduc-tase domains are highly conserved in various flavoHbs. A newclass of flavoHbs, displaying crucial differences in functionallyconserved regions of heme and reductase domains, have beenidentified in mycobacteria. Mining of microbial genome dataindicated that the occurrence of such flavoHbs might be re-stricted to a small group of microbes unlike conventional fla-voHbs that are widespread among prokaryotes and lowereukaryotes. One of the representative flavoHbs of this class,encoded by Rv0385 gene (MtbFHb) of Mycobacterium tubercu-losis, has been cloned, expressed, and characterized. The ferricand deoxy spectra of MtbFHb displayed a hexacoordinate stateindicating that its distal site may be occupied by an intrinsicamino acid or an external ligand and it may not be involved innitric oxide detoxification. Phylogenetic analysis revealed thatmycobacterial flavoHbs constitute a separate cluster distinctfrom conventional flavoHbs and may have novelfunction(s). � 2011 IUBMB
IUBMB Life, 63(5): 337–345, 2011
Keywords flavohemoglobin; mycobacteria; electron-transfer; phylog-
eny; nitric-oxide dioxygenase.
INTRODUCTION
Flavohemoglobins (flavoHbs) are monomeric proteins having
a classical 3/3 helical globin domain linked with a flavin ade-
nine dinucleotide (FAD)-binding reductase domain that may
transfer electrons to the heme iron and allow its rapid re-reduc-
tion to maintain it in the ferrous state for the rebinding of oxy-
gen (1–3). Rapid electron transfer via redox domain may allow
these two-domain globins to carry out diverse redox reactions
that may be vital for their native host. Recent upsurge in avail-
ability of genomic data from various organisms has indicated
that gene encoding for flavoHb may be widely distributed in
unicellular organisms, both prokaryotes and lower eukaryotes
(1, 2, 4); however, none has been found so far in higher organ-
isms suggesting that flavoHbs may be specifically relevant to
the cellular metabolism of microbes. Physiological function(s)
of flavoHbs has been the subject of intense debate over the last
decade. Several putative functions have been proposed for fla-
voHbs (5–8). However, accumulating experimental evidences
support its role in protection of microorganisms from the delete-
rious effects of nitric oxide (NO) by detoxifying it to nitrate via
the NO-dioxygenation reaction (1, 2, 7, 8). The hydrophobic
distal pocket of flavoHbs has been found much more flexible
and expandable than conventional Hbs (3, 9) that allow them to
accommodate large apolar molecules including phospholipids
and perform other cellular functions. Existence of more than
one flavoHb in some bacteria, yeasts, and fungi (10, 11) suggest
that these hemoglobins (Hbs) may be involved in diverse cellu-
lar functions.
FlavoHbs usually function as an integral part of stress
response and virulence of several pathogenic bacteria and fungi
by maintaining the cell redox homeostasis at the aerobic/anaero-
bic interface when cells are exposed to various environmental
stresses (7, 12). Although importance of flavoHbs for the para-
sitic life has been established in several cases (13, 14), none has
been reported from pathogenic or nonpathogenic mycobacteria
so far. When genomic data of various mycobacterial species was
examined, occurrence of multiple flavoHb encoding genes has
been detected in many of them. After amino-acid sequence
alignment and comparison of mycobacterial flavoHbs, we identi-
fied a novel class of flavoHb, exhibiting unconventional heme
and reductase domains, in mycobacteria apart from conventional
flavoHb. Occurrence of two distinct classes of flavoHbs in
mycobacteria is interesting as well as intriguing. Notably, the
new class of flavoHb is present in majority of mycobacterial
species and it also coexists along with a conventional flavoHb in
Address correspondence to: Kanak L. Dikshit, Institute of Microbial
Technology, CSIR, Sector 39A, Chandigarh 160036, India.
E-mail: [email protected]
Received 17 January 2011; accepted 28 February 2011
ISSN 1521-6543 print/ISSN 1521-6551 online
DOI: 10.1002/iub.460
IUBMB Life, 63(5): 337–345, May 2011

some of the mycobacterial species. At present, nothing is known
about mycobacterial flavoHbs and their role in cellular metabo-
lism. This study provides the first report on a new class of
flavoHbs that have been identified in mycobacteria and may not
be widespread among microbes unlike conventional flavoHbs.
Primary characterization of one of the flavoHbs of this group,
present in Mycobacterium tuberculosis, was done, and phyloge-
netic analysis was conducted to understand functional and
evolutionary correlation between two classes of mycobacterial
flavoHbs.
EXPERIMENTAL PROCEDURES
Bioinformatics and Phylogenetic Analysis
BLAST searches within the mycobacterial protein data bank
were done by using the sequences of different flavoHbs, for
example, E. coli, Ralstonia eutropha, and Saccharomyces cere-
visiae. Sequences retrieved from these searches were aligned
using CLUSAL W sequence alignment tool from EMBL-EBI
server. Individual FlavoHb of mycobacteria were identified by
paralogs and ortholog searches using E. coli HMP as primary
template. Multiple sequence alignment and conservation and
divergence within the heme and reductase domains of various
mycobacterial flavoHbs were analyzed. A phylogenetic tree was
built up using the neighbor-joining method (15) focusing on
mycobacterial flavoHbs and their close paralogs/orthologs and
conventional flavoHbs of E. coli, Ralstonia eutropha, Bacillus
subtilis, and Saccharomyces cerevisiae. The evolutionary distan-
ces were computed using the Poisson correction method (16)
and are in the units of the number of amino-acid substitutions
per site. All positions containing gaps and missing data were
eliminated from the dataset (complete deletion option). Phyloge-
netic analyses were conducted in MEGA4 (17).
Bacterial Strains, Plasmids, Gene Cloning,and Protein Purification
E. coli strains, JM 109 and BL21DE3, were used for the
cloning and expression of recombinant proteins. E. coli cells
were grown in Luria Bertani or terrific broth (containing 24 g
of yeast extract, 12 g of Bacto-Tryptone, 12.3 g of K2HPO4, 2.3
g of KH2PO4) at 37 8C at 180 rpm unless mentioned otherwise.
MtbFHb were retrieved from the genomic DNA of M. tubercu-
losis H37Rv and expressed in E. coli using standard polymerase
chain reaction (PCR) techniques. Authenticity of PCR-amplified
gene was checked by nucleotide sequencing. Recombinant
genes were cloned on pET 15b at NdeI –BamHI sites and
expressed in E. coli BL21DE3, cultured in Terrific broth supple-
mented with d-aminolevulinic acid (500 lM) and FeCl3 (20
lM) at 37 8C. Recombinant MtbFHb when expressed in E. coli,
appeared associated with cell membranes and was retrieved af-
ter treating the cell pellet with 1% sarcosyl. Recombinant
MtbFHb was purified from the cell lysate using nickel NTA
column (Quiagen) following manufacturer’s instructions. It
resulted in approximately 70–75% pure preparation of protein
exhibiting distinct reddish pink color. These fractions were fur-
ther purified by ion-exchange column (DEAE-Sepharose CL4B,
Pharmacia), equilibrated with 10-mM TrisHCl (pH 8.0) and
eluted using 0.12 M NaCl gradient. The protein and hemoglobin
profile was monitored at 280 and 414 nm. Absorption and CO
difference spectra of whole cells or purified protein preparation
were recorded using Carry 100 Spectrophotometer as described
previously (18). Gel-filtration chromatography was done on
Superdex 75 column, equilibrated with 50 mM TrisHCl (pH
7.0) and protein was eluted in 0.2 M NaCl after calibration with
standard molecular weight markers (Carbonic anhydrase, 29 kD;
Egg albumin, 40 kD and BSA 66 kD).
NO Consumption and Oxygen Uptake
NO consumption activity of MtbFHb expressing cells was
measured polarographically in a 2-ml reaction chamber with
ISO-NO (World Precision Instruments) essentially as described
previously (19). NO uptake was measured from the slope of
curving traces recorded in the specified concentration of NO
(30 lM) and corrected for background rate of NO decomposi-
tion recorded for the control in the absence of any protein. Ni-
tric oxide dioxygenase (NOD) activity of MtbFHb was deter-
mined following the published procedure (20). Oxygen con-
sumption by cells or cell extract was measured with an oxygen
monitor (Yellow Spring Instrument model 55) at 37 8C in 3 mL
of air saturated 0.1 M potassium phosphate buffer (pH 7.5). The
electrode was calibrated with air equilibrated water. Cell culture
(1 mL) was concentrated, washed with 0.1 M phosphate buffer,
and then added quantitatively to 3-mL air-saturated buffer to
check the oxygen consumption by monitoring the change in the
oxygen concentration of the buffer containing the cells. Three
independent experiments were performed for each set.
RESULTS
Two Distinct Classes of FlavoHbs in Mycobacteria
Computational and sequence analysis of available mycobac-
terial genome and protein data indicated that more than one fla-
voHb may be present in many species of mycobacteria. An
interesting pattern in the occurrence of flavoHb encoding genes
was found in both slow- and fast-growing mycobacteria. Oppor-
tunistic pathogens and fast-growing mycobacteria displayed
presence of two flavoHb encoding genes (Table 1); a conven-
tional flavoHb (type I) similar to other microbial flavoHbs (1,
2) and a new class (type II) showing crucial differences within
the functionally conserved regions of heme and reductase
domains. Interestingly, the new class of type II flavoHb
appeared in majority of mycobacteria including both virulent
and avirulent species. Presence of type II flavoHb in microbial
genome was found restricted to certain bacterial species, mainly
belonging to actinomycetes. Existence of two different flavoHbs
in the same organism suggests that they may be playing differ-
ent functions in cellular metabolism of mycobacteria.
338 GUPTA ET AL.

Structural Characteristics of Type II flavoHbs ofMycobacteria: Unusual Features of Hemeand Reductase Domains
Sequence comparison of mycobacterial type I and type II
flavoHbs with other bacterial flavoHbs (Fig. 1A) indicated
that structural features for adopting a three over three globin
fold and signature sequences of typical microbial globins, for
example, B10-Tyr, CD1-Phe, E7-Gln, F8-His (3, 21), are
present in type II flavoHbs, but there are several differences
within the functionally conserved regions of their heme and
reductase domains. The most notable differences within the
globin domain of type II flavoHb is the lack of conserved
hydrogen bonding and disruption of tetrainteractions between
HisF8-GluH23-TyrG5 and contact between TyrG5 and
TyrH12 within the proximal site due to mutation at GluH23
and TyrH12 residues. These observations indicated that the
peroxidase such as catalytic site, present in conventional fla-
voHbs (22), is absent in this class of flavoHbs. The reductase
domain of type II flavoHbs of mycobacteria is also modified
within the cofactor binding sites, for example, the RXYS
motif of the FAD binding site and GXGXXP motif of the
nicotinamide adenine dinucleotide (NADH) binding sites (23,
24). A RKY/F sequence motif, known as high affinity lipid-
binding motif (25), appeared conserved within the proximal
site of heme in type II flavoHbs. Interestingly, mycobacterial
type II flavoHbs exhibited very high (>70%) overall sequence
conservation among them but displayed less than 25% homol-
ogy with conventional (type I) flavoHbs. These unusual struc-
tural features indicated that type II flavoHbs of mycobacteria
might be structurally and functionally different from conven-
tional flavoHbs.
Cloning, Expression, and Characterization of Type IIFlavoHb from M. tuberculosis
To gain an insight into the primary characteristics of type II
flavoHbs, one of its representatives, encoded by Rv0385 gene
in Mycobacterium tuberculosis, was cloned and expressed in E.
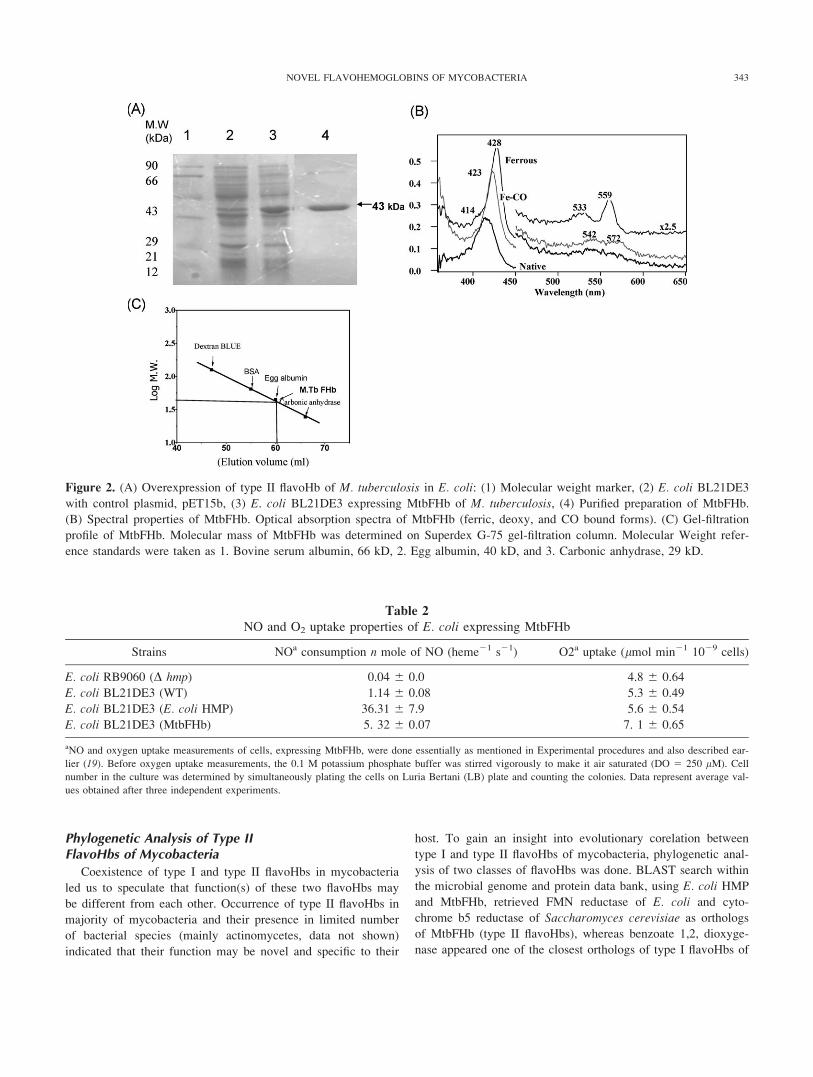
coli. SDS-PAGE analysis confirmed the presence of a 43-kDa
band (Fig. 2A) corresponding to the expected size of M. tubercu-
losis flavoHb (MtbFHb) protein. Gel filtration analysis of
MtbFHb substantiated that it is a monomeric protein of 43.5 kDa
(Fig. 2C). Absolute spectra of MtbFHb indicated that protein pre-
dominantly exists in the ferric state. The absorption spectra of
the ferric species exhibits Soret and visible bands at 414 and
536/570 nm, respectively (Fig. 2B), suggesting a hexacoordinated
low-spin (6CLS) heme with an intrinsic amino acid residue or
exogenous ligand bound to the distal site of the heme. The
absorption spectrum of the ferrous species shows Soret and visi-
ble bands at 428 and 533/559 nm, respectively, substantiating the
6CLS configuration of heme, consistent with the presence of a
sixth ligand. Exposure of the ferrous protein to CO caused the
absorption bands to shift to 423 and 542/572 nm, respectively,
typical for CO-bound heme, indicating that the distal ligand is
displaced by the CO. This is in sharp variance with conventional
flavoHbs that exist in penatcoordinated high spin state (22).
These observations indicated that MtbFHb and presumably other
mycobacterial type II flavoHbs may be structurally and function-
ally distinct from conventional type I flavoHbs.
Table 1
Occurrence and distributions of flavoHbs in mycobacteria
Mycobacterium Species No. of flavoHbs Status
FlavoHbs
Type I (E. coli type) Type II (M. tuberculosis type)
M. laprae 0a Virulent – –
M. tuberculosis (H37 Rv) 1 Virulent – Yes (Rv0385)
M. BovisAF2122/97 1 Virulent – Yes (BCG_0393)
M. abcesseus 1 Virulent – Yes (MAB_4269)
M. paratuberculosis 1 Virulent – Yes (MAP_3851)
M. avium 1 Virulent – Yes (MAV_4795)
M. marinum 1 Virulent – Yes (MMAR_0644)
M. ulcerans 1 Virulent – Yes (MUL_128)
M. gilvum 2 Avirulent Yes (Mflv_4884) Yes (Mflv_0255)
M. smegmatis 2 Avirulent Yes (MSMEG_1336) Yes (MSMEG_0719)
M. vanvallani 2 Avirulent Yes (Mvan_1541) Yes (Mvan_639)
M. sp. JLS 2 Avirulent Yes (Mjls_0944) Yes (Mjls_457)
M. sp. MCS 2 Avirulent Yes (Mmcs0916) Yes (Mmcs_470)
M. sp. KMS 2 Avirulent Yes (Mkms_0933) Yes (Mkms_481)
aPseudogene.
FlavoHbs were identified in different species of mycobacteria using E. coli flavoHb as template followed by its paralogs and orthologs searches. Based on
sequence alignment and comparison, two different classes of flavoHbs were identified. Type I designates for flavoHbs similar to flavoHbs of bacteria and
yeasts. Type II designates for the new class of flavoHbs present in M. tuberculosis and other mycobacteria.
339NOVEL FLAVOHEMOGLOBINS OF MYCOBACTERIA

Figure 1. (A) Structure-based sequence alignment of mycobacterial flavohemoglobins (type I and type II) with E. coli and Ralsto-
nia flavoHbs. Conserved residues in heme and reductase domains of flavoHbs are highlighted in light gray, and the residues, which
are different from conserved and present in MtbFHb (type II), are shown in dark gray. FAD and NADPH binding motifs, present
in conventional flavoHbs, are shown in vertical boxes. E. c, Escherichia coli; R. e, Ralstonia eutropha; M. v, Mycobacterium van-
vallani; M. g, Mycobacterium gilvum; M. j, Mycobacterium ssp. jls; M. s, Mycobacterium smegmatis; M. u, Mycobacterium ulcer-
ans; M. m, Mycobacterium marinum; M. p, Mycobacterium paratuberculosis; M. av, Mycobacterium avium; M. tb, Mycobacterium
tuberculosis; M. a, Mycobacterium absessceu. Type I, denotes protein sequence resembling with conventional E. coli type flavoHbs
and type II, sequence designates for the new class of flavoHbs identified in M. tuberculosis and other mycobacteria. (B) Phyloge-
netic analysis of mycobacterial flavoHbs. The phylogenetic analysis was performed by using MEGA4 (17) program. Mycobacterial
genome was searched extensively for the homologs/paralogs and orthologs for flaoHbs and then the first blast hit of different ge-
nome was selected and analyzed by sequence alignment and comparisons for making the evolutionary tree. Mycobacterial type II
flavoHbs make separate cluster from type I FlavoHbs that form cluster along with conventional flavoHbs of bacteria and yeasts.
[Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]

As protection against NO and reactive nitrogen species has
been found one of the major physiological functions of bacterial
and yeast flavoHbs (type I), we tested the NO and oxygen
metabolizing properties of MtbFHb by comparing the NO and
oxygen uptake of MtbFHb overexpressing cells of E. coli.
MtbFHb expressing cells did not show any significant NO
uptake when compared with isogenic cells expressing E. coli
HMP (Table 2) but displayed moderately improved respiratory
activities. NOD activity of MtbFHb was estimated as 12 lM21
s21, which was several folds lower than the HMP of E. coli and
trHbN of M. tuberculosis (26). Overall observations, thus, sug-
gested significant differences in structural and functional proper-
ties of type II flavoHb of M. tuberculosis (MtbFHb) when com-
pared with conventional type I flavoHbs.
Figure 1. Continued.
341NOVEL FLAVOHEMOGLOBINS OF MYCOBACTERIA

Figure 1. Continued.
342 GUPTA ET AL.
Phylogenetic Analysis of Type IIFlavoHbs of Mycobacteria
Coexistence of type I and type II flavoHbs in mycobacteria
led us to speculate that function(s) of these two flavoHbs may
be different from each other. Occurrence of type II flavoHbs in
majority of mycobacteria and their presence in limited number
of bacterial species (mainly actinomycetes, data not shown)
indicated that their function may be novel and specific to their
host. To gain an insight into evolutionary corelation between
type I and type II flavoHbs of mycobacteria, phylogenetic anal-
ysis of two classes of flavoHbs was done. BLAST search within
the microbial genome and protein data bank, using E. coli HMP
and MtbFHb, retrieved FMN reductase of E. coli and cyto-
chrome b5 reductase of Saccharomyces cerevisiae as orthologs
of MtbFHb (type II flavoHbs), whereas benzoate 1,2, dioxyge-
nase appeared one of the closest orthologs of type I flavoHbs of
Figure 2. (A) Overexpression of type II flavoHb of M. tuberculosis in E. coli: (1) Molecular weight marker, (2) E. coli BL21DE3
with control plasmid, pET15b, (3) E. coli BL21DE3 expressing MtbFHb of M. tuberculosis, (4) Purified preparation of MtbFHb.
(B) Spectral properties of MtbFHb. Optical absorption spectra of MtbFHb (ferric, deoxy, and CO bound forms). (C) Gel-filtration
profile of MtbFHb. Molecular mass of MtbFHb was determined on Superdex G-75 gel-filtration column. Molecular Weight refer-
ence standards were taken as 1. Bovine serum albumin, 66 kD, 2. Egg albumin, 40 kD, and 3. Carbonic anhydrase, 29 kD.
Table 2
NO and O2 uptake properties of E. coli expressing MtbFHb
Strains NOa consumption n mole of NO (heme21 s21) O2a uptake (lmol min21 1029 cells)
E. coli RB9060 (D hmp) 0.04 6 0.0 4.8 6 0.64
E. coli BL21DE3 (WT) 1.14 6 0.08 5.3 6 0.49
E. coli BL21DE3 (E. coli HMP) 36.31 6 7.9 5.6 6 0.54
E. coli BL21DE3 (MtbFHb) 5. 32 6 0.07 7. 1 6 0.65
aNO and oxygen uptake measurements of cells, expressing MtbFHb, were done essentially as mentioned in Experimental procedures and also described ear-
lier (19). Before oxygen uptake measurements, the 0.1 M potassium phosphate buffer was stirred vigorously to make it air saturated (DO 5 250 lM). Cell
number in the culture was determined by simultaneously plating the cells on Luria Bertani (LB) plate and counting the colonies. Data represent average val-
ues obtained after three independent experiments.
343NOVEL FLAVOHEMOGLOBINS OF MYCOBACTERIA

mycobacteria. Therefore, a phylogenetic tree was developed by
focusing on type I, type II flavoHbs of mycobacteria and their
first orthologs present in different groups (Fig. 1B). Topology
of evolutionary tree, thus, developed, separated type II flavoHbs
of mycobacteria from type I flavoHbs that formed a separate
group along with conventional flavoHbs of bacteria and yeasts.
Phylogenetically, type II flavoHbs appeared related to electron-
transfer proteins such as FMN-reductase of E. coli and cyto-
chrome b5reductase of Sacchromyces cerevisiae, whereas type I
flavoHbs of mycobacteria and other conventional flavoHbs
exhibited phylogenetic closeness with dioxygenases. Overall
structure of phylogenetic tree and duplication nodes suggest an
ancestral duplication and diversion of type I and type II myco-
bacterial flavoHbs.
DISCUSSION
FlavoHbs are widely spread among bacteria, yeast, and fungi
(1, 2) and represent a unique example of multidomain protein,
where two domains having different functional properties, inter-
act and perform entirely new function(s) by carrying out diverse
redox reactions. Heme and reductase domains of flavoHbs dis-
play significant sequence conservation within their cofactor bind-
ing sites (3, 21). A novel flavoHb, displaying crucial differences
within the functionally conserved regions of heme and reductase
domain, has been identified in many species of mycobacteria.
Occurrence of this unconventional class (type II) of flavoHbs
appeared limited to few microbes, mainly belonging to actinomy-
cetes group. Coexistence of type I and type II flavoHbs in several
mycobacterial species suggest that they may be playing different
functions in cellular metabolism of their host.
Type II flavoHbs may constitute a distinct class within the
flavoHb family and may have novel function(s). Functionally
conserved regions of heme and redox domains of type I fla-
voHbs are modified in type II flavoHbs of mycobacteria sug-
gesting that the interactions of heme and redox domain and
mode of redox reactions are different in these proteins. This is
supported by the observation that type II flavoHb of M. tubercu-
losis exists in hexacoordinate state and lacks NO metabolizing
activity unlike majority of type I flavoHbs. When MtbFHb
(type II) was expressed in E. coli, it appeared strongly associ-
ated with cell membranes and bioinformatics analysis indicated
that its orthologs in E. coli and S. cerevisiae are FMN reductase
and Cytochrome b5 reductse, respectively. Structural and func-
tional properties of type II flavoHbs are not obvious at present;
however, their occurrence in limited group of microbes suggests
that they may have novel function(s) in the cellular metabolism
of their host.
To understand the need for two different classes of flavoHbs
and their evolutionary corelation, phylogenetic analysis of
mycobacterial flavoHbs were conducted. Topology of the evolu-
tionary tree, developed after taking into consideration the paral-
ogs and orthologs of type I and type II flavoHbs of mycobacte-
ria, indicated that type II flavoHbs form a separate cluster from
type I flavoHbs and may be functionally different. However,
both may have originated from a common ancestor and evolved
after duplication and diversion. Although precise origin and
functional roles of mycobacterial flavoHbs remains to be eluci-
dated, respective spacing of mycobacterial type I and type II
flavoHbs and their paralogs suggest that type II flavoHbs may
be phylogenetically more related to electron-transferring pro-
teins (FMN reductases, cytochrome b5 reductases, etc.), whereas
type I flavoHbs are closer to dioxygenases.
The diversity in the number and type of flavoHb-encoding
genes in the genome of mycobacteria might reflect the differ-
ence in requirement of these proteins in individual species.
Type II flavoHbs, thus, may be required for the cellular metabo-
lism of both pathogenic and nonpathogenic mycobacteria as it
has been found in majority of analyzed mycobacterial genome
(except M. leprae, where it has been identified as pseudogene).
Phylogenetic studies on invertebrate globins (27) suggested that
flavoHbs and flavoHb like single-domain Hbs (FHb/SDSgb)
family diverged much more in their structure-function than trun-
cated and globin-coupled sensors. Unconventional flavoHbs of
mycobacteria may add another dimension on diversification of
microbial two-domain globins.
ACKNOWLEDGEMENTS
The authors thank financial support from Council of Scientific
and Industrial Research under the SIP10 grant and NSF grant
095635 (SRY) for carrying out this work.
REFERENCES1. Wu, G., Wainwright, L. M., and Poole, R. K. (2003) Microbial globins.
Adv. Microb. Physiol. 47, 255–310.2. Frey, A. D. and Kallio, P. T. (2003) Bacterial hemoglobins and flavohe-
moglobins: versatile proteins and their impact on microbiology and bio-
technology. FEMS Microbiol. Rev. 27, 525–545.
3. Bonamore, A. and Boffi, A. (2008). Flavohemoglobin: structure and
reactivity. IUBMB Life 60, 19–28.
4. Kobayashi, G., Nakamura, T., Ohmachi, H., Matsuoka, A., Ochiai, T.,
and Shikama, K. (2002) Yeast flavohemolgobin from Candida norve-gensis: its structure, spectral and stability properties. J. Biol. Chem. 277,
42540–42458.
5. Gardner, P. R., Gardner, A. M., Martin, L. A., and Salzman, A. L.
(1998) Nitric oxide dioxygenase: an enzymatic function for flavohemo-
globin Proc. Natl. Acad. Sci. USA 95, 10378–10383.
6. Farres, J., Rechsteiner, M. P., Herold, S., Frey, A. D., and Kallio, P. T.
(2005) Ligand binding properties of bacterial hemoglobins and flavohe-
moglobins. Biochemistry 44, 4125–4134.
7. Gilberthorpe, N. J. and Poole, R. K. (2008) Nitric oxide homeostasis in
Salmonella typhimurium: roles of respiratory nitrate reductase and flavo-
hemoglobin. J. Biol. Chem. 283, 11146–111454.8. Membrillo-Hernandez, J., Coopamah, M. D., Anjum, M. F., Stevanin,
T. M., Kelly, A., Hughes, M. N., and Poole, R. K. (1999) The flavohe-
moglobin of Escherichia coli confers resistance to a nitrosating agent, a
nitric oxide releaser,’’ and paraquat and is essential for transcriptional
responses to oxidative stress. J. Biol. Chem. 274, 748–754.
9. D’Angelo, P., Lucarelli, D., della Longa, S., Benfatto, M., Hazemann, J.
L., Feis, A., Smulevich, G., IIari, A., Bonamore, A., and Boffi, A.
344 GUPTA ET AL.

(2004) Unusual heme iron-lipid acyl chain coordination in Escherichiacoli flavohemoglonin. Biophys. J. 86, 3882–3892.
10. Zhou, S., Fushinobu, S., Nakanishi, Y., Kim, S. W., Wakagi, T., and
Shoun, H. (2009) Cloning and characterization of two flavohemoglo-
bins from Aspergillus oryzae. Biochem. Biophys. Res. Commun. 381,
7–11.
11. Iijima, M., Shimizu, H., Tanaka, Y., and Urushihara, H. (2000) Identifi-
cation and characterization of two flavohmeoglobin genes in Dictyoste-lium discoideum. Cell. Struct. Funct. 25, 47–55.
12. Bang, I. S., Liu, L. Vazquez-Torres, A., Crouch, M. L., Stamler, J. S.,
and Fang, F. C. (2006) Maintenance of nitric oxide and redox homeo-
stasis by the Salmonella flavohemoglobin hmp. J. Biol. Chem. 281,
28039–280472.
13. Stevanin, T. M., Poole, R. K. Demoncheaux, E. A., and Read, R. C.
(2002) Flavohemoglobin Hmp protects Salmonella enterica serovar
Typhimurium from nitric-oxide related killing by human macrophages.
Infect. Immun. 70, 4399–4405.
14. Favey, S., Labesse, G., Vouille, V., and Boccara, M. (1995) Flavohae-
moglobin HmpX: a new pathogenicity determinant in Erwinia chrysan-themi strain 3937. Microbiology 141, 863–871.
15. Saitou, N. and Nei, M. (1987) The neighbour-joining method: a new
method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–
425.
16. Zuckerkandl, E. and Pauling, L. (1965) Evolutionary divergence and
convergence in proteins. In Evolving Genes and Proteins, (Bryson, V.
and Vogel, H. J.). pp. 97–166, Academic Press, New York.
17. Tamura, K., Dudley, J., Nei, M., and Kumar, S. (2007) MEGA4: molec-
ular evolutionary genetics analysis (MEGA) software version 4.0. Mol.
Biol. Evol. 24, 1596–1599.
18. Dikshit, K. L. and Webster, D. A. (1988) Cloning, characterization and
expression of the bacterial globin gene from Vitreoscilla in Escherichia
coli. Gene 70, 377–386.
19. Pathania, R., Navani, N. K., Gardner, A. M., Gardner, P. R., and Dik-
shit, K. L. (2002) Nitric oxide scavenging and detoxification by the
Mycobacterium tuberculosis haemoglobin, HbN in Escherichia coli.
Mol. Microbiol. 45, 1303–1314.
20. Gardner, P. R., Gardner, A. M., Martin, L. A., Dou, Y., Li, T., Olson, J.
S., Zhou, H., and Riggs, A. F. (2000) Nitric oxide dioxygenase activity
and function of flavohemoglobins. J. Biol. Chem. 275, 31581–31587.
21. Ermler, U., Siddiqui, R. A., Cramm, R., and Friedrich, B. (1995) Crys-
tal structure of the flavoheemoglobin from Alcaligenes eutrophus at 1.
75 A resolution. EMBO J. 14, 6067–6077.
22. Mukai, M., Mills, C. E., Poole, R. K., and Yeh, S. R. (2001) Flavohe-
moglobin, a globin with a peroxidase like catalytic site. J. Biol. Chem.276, 7272–7277.
23. Dym, O. and Eisenberg, D. (2001) Sequence-structure analysis of FAD
containing proteins. Protein Sci. 10, 1712–1728.
24. Kimura, S., Nishida, H., and Iyanagi, T. (2001) Effects of flavin-binding
motif amino acid mutations in the NADH-cytochrome b5 reductase, cata-
ytic domain on protein stability and catalysis. J. Biochem. 30, 481–490.
25. Hunt, C. (2005) Specific protein-lipid interactions in membrane proteins.
Biochem. Soc. Trans. 33, 938–942.26. Gardner, P. R. (2005) Nitric oxide dioxygenase function and mechanism
of flavohemoglobin, myoglobin and their associated reductases. J. Inorg.
Biochem. 99, 247–266.27. Vinogradov, S. N. and Moens, L. (2008) Diversity of globin function:
enzymatic, transport, storage and sensing. J. Biol. Chem. 283, 8773–
8777.
345NOVEL FLAVOHEMOGLOBINS OF MYCOBACTERIA




















