
T.C. İSTANBUL ÜNİVERSİTESİ İSTANBUL TIP FAKÜLTESİ ...
90
T.C. İSTANBUL ÜNİVERSİTESİ İSTANBUL TIP FAKÜLTESİ KARDİYOLOJİ ANABİLİM DALI TEZ DANIŞMANI PROF. DR. MELİKE ZEHRA BUĞRA KARDİYOLOJİ ANABİLİM DALI İSTANBUL-2010 “DR. ERHAN TEKER” “ROMATİZMAL KALP KAPAK HASTALIĞININ KAPAKLARDA NEDEN OLDUĞU HASARIN DERECESİ İLE İNTERFERON GAMA 874 T/A POLİMORFİZMİ ARASINDAKİ İLİŞKİNİN DEĞERLENDİRİLMESİ” (TIPTA UZMANLIK TEZİ)
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of T.C. İSTANBUL ÜNİVERSİTESİ İSTANBUL TIP FAKÜLTESİ ...

T.C.
İSTANBUL ÜNİVERSİTESİ
İSTANBUL TIP FAKÜLTESİ
KARDİYOLOJİ ANABİLİM DALI
TEZ DANIŞMANI
PROF. DR. MELİKE ZEHRA BUĞRA
KARDİYOLOJİ ANABİLİM DALI
İSTANBUL-2010
“DR. ERHAN TEKER”
“ROMATİZMAL KALP KAPAK HASTALIĞININ KAPAKLARDA NEDEN OLDUĞU HASARIN DERECESİ İLE İNTERFERON GAMA 874 T/A
POLİMORFİZMİ ARASINDAKİ İLİŞKİNİN DEĞERLENDİRİLMESİ”
(TIPTA UZMANLIK TEZİ)

ii
TEZ ONAYI
(Bu sayfa yerine, başarılı geçen Tez Sınavı sonrası sınav tutanağı ekinde yer alan Tez Onay sayfası gelecektir.)

iii
BEYAN
Bu tez çalışmasının kendi çalışmam olduğunu, tezin planlanmasından yazımına kadar
bütün safhalarda etik dışı davranışımın olmadığını, bu tezdeki bütün bilgileri akademik ve etik
kurallar içinde elde ettiğimi, bu tez çalışmayla elde edilmeyen bütün bilgi ve yorumlara kaynak
gösterdiğimi ve bu kaynakları da kaynaklar listesine aldığımı, yine bu tezin çalışılması ve
yazımı sırasında patent ve telif haklarını ihlal edici bir davranışımın olmadığı beyan ederim.
Dr. Erhan Teker

iv
İTHAF
Tezimi eşim’e ithaf ediyorum

v
TEŞEKKÜR
Kardiyoloji Anabilim Dalındaki uzmanlık eğitimim süresince bilgi, görgü ve
becerilerinden yararlandığım değerli hocalarım Anabilim Dalı Başkanımız Prof. Dr. Kamil
Adalet’e, Prof. Dr. Faruk Erzengin’e, Prof. Dr. Mustafa Özcan’a, Prof. Dr. Nevrez Koylan’a,
Prof. Dr. Sabahattin Umman’a, Prof. Dr. Taner Gören’e, Prof. Dr. Dursun Atılgan’a, Prof. Dr.
Aytaç Öncül’e, Prof. Dr. Berrin Umman’a, Prof. Dr. Fehmi Mercanoğlu’na, Prof. Dr. Hüseyin
Oflaz’a, Doç. Dr. Murat Sezer’e, Doç. Dr. Ahmet Kaya Bilge’ye, emekli hocalarımız Prof. Dr.
Özen Güven’e, Prof. Dr. Ercüment Yılmaz’a ve birlikte çalıştığım Kardiyoloji Anabilim
Dalı’mızda görevli tüm asistan arkadaşlarım, hemşirelerimiz ve personelimize teşekkürlerimi
sunarım.
Moleküler kardiyoloji alanında beni cesaretlendiren ve bu alanda engin bilgilerinden
yararlandığım değerli hocam Prof. Dr. Yılmaz Nişancı’ ya ayrıca teşekkür ederim.
Klinik bilgi ve becerilerimi geliştirmemde büyük emeği olan ve tez çalışmamın her
aşamasında yakın ilgi ve desteğini gördüğüm tez danışmanı ve sorumlusu değerli hocam Prof.
Dr. Melike Zehra Buğra’ya teşekkürü bir borç ve görev bilirim.
Ayrıca çalışmamın genetik kısmında sabırla bana tam destekçi olan eşim Ayşegül
Başak Teker’e ve hem laboratuvar hemde istatistik alanlarında engin bilgisinden yararlandığım
değerli hocam Doç. Dr. Oğuz Öztürk’e teşekkür ederim.
Bu çalışma, İstanbul Üniversitesi Bilimsel Araştırma Projeleri Birimi tarafından
desteklenmiştir. Proje No: 6963

vi
İÇİNDEKİLER
TEZ ONAYI ............................................................................................................... İİ
BEYAN ..................................................................................................................... İİİ
İTHAF ....................................................................................................................... İV
TEŞEKKÜR ................................................................................................................ V
İÇİNDEKİLER .......................................................................................................... Vİ
TABLOLAR LİSTESİ ............................................................................................ Vİİİ
ŞEKİLLER LİSTESİ .................................................................................................. İX
SEMBOLLER / KISALTMALAR LİSTESİ ............................................................... X
ÖZET ........................................................................................................................ Xİİ
ABSTRACT ............................................................................................................ Xİİİ
1. GİRİŞ VE AMAÇ .................................................................................................... 1
2. GENEL BİLGİLER .................................................................................................. 2
Romatizmal Ateş 2
Epidemiyoloji-Tanısı 2
2.1. ROMATİZMAL KALP KAPAK HASTALIĞI ................................................... 4
Romatizmal kalp kapak hastalığının patogenezi 5
OTOİMMUNİTE...................................................................................................... 6
Romatizmal kalp kapak hastalığında otoimmun reaksiyonlar 7
2.1.1. ROMATİZMAL KALP KAPAK HASTALIĞININ GENETİK TEMELLERİ 9
2.1.1.1. İNTERFERON GAMA ............................................................................ 11
İnterferon gama'nın moleküler etki mekanızması 13
İnterferon gama'nın gen ekspresyonunun regülasyonu 14
İnterferon gama geni üzerindeki polimorfizmler 16
3. GEREÇ VE YÖNTEM ........................................................................................... 18
3.1. Seçilen Örneklerin Tanımı ................................................................................. 18
3.1.1. Kullanılan Yöntemle ..................................................................................... 20
Periferik Kandan DNA İzolasyonu.......................................................................... 20
Elde Edilen DNA’nın Konsantrasyon ve Kalitesinin Tayini .................................. 21
3.1.1.1. IFN-gama geni Amplifikasyon Ürünlerinde Gen Polimorfizminin Tespiti: 22
IFN-gama Geninin PZR Yöntemi İle Çoğaltılması ................................................ 22

vii
IFN-gama PZR Sonuçlarının Agaroz Jel Elektroforezi ile Görüntülenmesi............ 23
4. BULGULAR .......................................................................................................... 25
5. TARTIŞMA ........................................................................................................... 36
KAYNAKLAR ........................................................................................................ 411
HAM VERİLER....................................................................................................... 488
FORMLAR .............................................................................................................. 733
ETİK KURUL KARARI .......................................................................................... 744
PATENT HAKKI İZNİ ............................................................................................ 755
TELİF HAKKI İZNİ ................................................................................................ 766
ÖZGEÇMİŞ ............................................................................................................. 777

viii
TABLOLAR LİSTESİ
Tablo 2-1:Güncelleştirilmiş Jones kriterleri ...................................................................3
Tablo 3-1: IFN-gama +874 A/T polimorfizminin gözlendiği bölgenin PZR reaksiyonu
için termal profil şartları .............................................................................................. 23

ix
ŞEKİLLER LİSTESİ
Şekil 3-1: IFN-γ ARMS PZR ürünlerinin %2’lik agaroz jeldeki görüntüsü. ................. 23
Şekil 4-1: Hasta ve kontrol grubu arasında yaş ve cinsiyet dağılımı ............................. 26
Şekil 4-2: Hasta ve kontrol grupları arasında IFN γ 874 T-A genotip dağılımı ........... 26
Şekil 4-3: Hasta ve kontrol grubu arasında T homozigot genotip dağılımı.................... 27
Şekil 4-4: Hasta ve kontrol grubu arasında T allel dağılımı .......................................... 27
Şekil 4-5: Hasta grubunda genotipe göre kapak hasarında ciddiyet dağılımı ................ 28
Şekil 4-6:Hasta grubunda T homozigot genotip ve kapak hasarında ciddiyet dağılımı .. 28
Şekil 4-7: Hasta grubunda T alleli taşıma ve kapak hasarında ciddiyet dağılımı ........... 29
Şekil 4-8: Hasta grubunda IFN γ 874T/A genotip ve romatizmal kapak tutulumunun
darlık veya yetersizlik ağırlıklı olmasına göre dağılımı ................................................ 30
Şekil 4-9: Hasta grubunda T homozigot genotip ve romatizmal kapak tutulumunun
darlık veya yetersizlik ağırlıklı olmasına göre dağılımı ................................................ 30
Şekil 4-10: Hasta grubunda; valvüloplasti öyküsü ve IFN γ 874T/A genotip dağılımı. . 31
Şekil 4-11: Hasta grubunda valvüloplasti öyküsü ve T homozigot genotip dağılımı ..... 31
Şekil 4-12: Hasta grubunda; kapak replasmanı öyküsü ile IFN γ 874T/A genotip
dağılımı. ...................................................................................................................... 32
Şekil 4-13: Hasta grubunda kapak replasmanı öyküsü ile A homozigot genotip
dağılımı ....................................................................................................................... 32
Şekil 4-14: Hasta grubunda kapak replasmanı öyküsü ile T homozigot genotip
dağılımı ....................................................................................................................... 32
Şekil 4-15: Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda IFN γ 874T/A
genotip dağılımı .......................................................................................................... 33
Şekil 4-16: Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda T homozigot
genotip dağılımı .......................................................................................................... 33
Şekil 4-17: Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda T allel
dağılımı ....................................................................................................................... 34
Şekil 4-18: Penisilin proflaksisi-Kapak hasarı .............................................................. 35

x
SEMBOLLER / KISALTMALAR LİSTESİ
RKKH: Romatizmal kalp kapak hastalığı
IFN-γ: İnterferon gama
RA: Romatizmal ateş
ASH: Antijen Sunucu Hücresi
IL-1RA: İnterlökin 1 reseptör antagonistini
IL-1: İnterlökin 1
IL-2: İnterlökin 2
IL-3: İnterlökin 3
IL-4: İnterlökin 4
IL-5: İnterlökin 5
IL-6: İnterlökin 6
IL-10 : interlökin 10
IL-12: İnterlökin 12
IL-13: İnterlökin 13
TNF-α: Tümör nekrozis faktör alfa
TNF-ß: Tümör nekrozis faktör beta
GM-CSF: Granülosit-Monosit koloni uyarıcı faktör
Th1: Yardımcı T Hücresi 1
Th2: Yardımcı T Hücresi 2
CD+4 T: Yardımcı T hücresi
HLA: Human Lökosit Antijeni
MBL: Mannoz bağlayıcı lectin
LTA: lipoteikoik asit
TLR: Tool-Like receptörler
NF-kß: Nükleer faktör kappa beta
IFNGR: İnterferon gama reseptörü
Jak : Janus aktive edici kinazlar
STAT: Sinyal iletici ve transkripsiyon aktivitörü
GAF: IFN γ aktive edici faktör
GAS: IFN γ aktive edici sensör
ISGF: İnterferon Stimüle eden Gen Faktörü
T bet: T Box ailesine ait nükleer faktör
MAF:Makrofaj aktivitör faktör

xi
NFAT: Aktif T hücresinin Nükleer Faktörü
AP-1: Aktivatör protein 1
STAR 4: sinyal iletici aktivitör reseptörü 4
ARMS- PZR:Amplifikasyon Refrakter Mutasyon Sistemi- Polimeraz Zincir Reaksiyonu
ACC/AHA: American College of Cardiology and American Heart Association

xii
ÖZET
Teker,E. (2010). Romatizmal Kalp Kapak Hastalığının Kapaklarda Neden Olduğu Hasarın
Derecesi İle İnterferon gama 874 T/A Polimorfizmi arasındaki ilişkinin değerlendirilmesi
İstanbul Üniversitesi Tıp Fakültesi, Kardiyoloji Anabilim Dalı Tıpta uzmanlık Tezi. İstanbul.
Giriş: Romatizmal kalp kapak hastalığı grup A streptokoksik infeksiyonun otoimmun sekelidir.
Hem romatizmal ateş hem de romatizmal kalp hastalığının, grup A streptoksik infeksiyondan
sonra anormal immun cevap sonucu geliştiği düşünülmektedir. Enfeksiyon ajanının
yokluğunda bile kalp kapaklarında süregelen bir inflamasyon mevcuttur. Fakat kapak hasarının
şiddeti herkesde aynı değildir. Bazı hastalarda genç yaşta fibrozis ve kalsifikasyon nedenli
ciddi kalp kapak tutulumu mevcut iken bazı hastalarda bütün yaşamı boyunca hafif kapak
lezyonları mevcuttur. Çapraz reaktif T hücreleri ve inflamatuvar sitokinler hastalığın hem
tetiğinin çekilmesinde hem de kalp kapaklarında oluşan inflamasyonun sürekliliğinde önemli
role sahiptir. IFN-γ çok fonksiyonlu bir sitokin olup immun sistemin bütün fazlarının
düzenlenmesinde önemli rol oynamaktadır. IFN-γ, RKKH’nın patogenezindeki artmış fibrozis
ve kalsifikasyondan sorumlu olabilir. Buna rağmen, RKKH’ında IFN-γ nın genetik
varyasyonları günümüzde hala araştırılmamıştır. Bu çalışmamın amacı; Türk popülasyonunda
RKKH’daki kalp kapak hasarının şiddeti ve IFN-γ polimorfizmi arasındaki muhtemel ilişkiyi
araştırmaktır. Gereç ve yöntem: IFN-γ 874 T-A polimorfizmi, 152 RKKH ve 151 sağlıklı
kontrollerde ARMS PZR yöntemi kullanılarak tespit edilmiştir. Hasta grubu kapak hasarının
ciddiyetine göre iki gruba ayrılmıştır; ilk grup ciddi kapak hastalığı , kapak replasmanı ve balon
valvüloplasti ile daha önce tedavi görmüş olan şiddetli kapak hasarlı hastalardan oluşmaktadır.
İkinci grup hafif kapak hasarından oluşmaktadır. Sonuçlar: Hasta ve kontrol grubu
karşılaştırıldığında IFN-γ 874 T allel and TT genotipi hasta grubunda önemli olarak artmış
bulundu. (T allel için p=0.002,OR= 2.40 and TT genotip için p=0.018,OR= 2.14). Şiddetli
kapak hasarı olan hasta grubunda IFN-γ 874 TT genotip sıklığın artmış olarak bulundu.
(p=0.009, OR= 3.66) Ayrıca valvüloplasti öyküsü mevcut olan hastalarda da IFN-γ 874 TT
genotip sıklığı artmış olarak bulundu. (p=0.012, OR= 2.75). Dahası , kapak replasmanı öyküsü
olan hastalarda IFN-γ874 AA genotipinde azalma (p=0.01, OR= 3.06) bulundu. Yorum:Veriler
Türk popülasyonunda RKKH ile IFN-γ 874 T/A polimorfizmi arasındaki ilişkiyi ve bu
polimorfizmin şiddetli kapak hasarı ile ilişkisini gösterdi.A allelinin kapak hasarına karşı
koruyucu etkisi görüldü. Anahtar kelimeler: Romatizmal kalp kapak hastalığı , kapak hasarı,
IFN-γ polimorfizmi, genetik yatkınlık.
Bu çalışma, İstanbul Üniversitesi Bilimsel Araştırma Projeleri Birimi tarafından desteklenmiştir.
Proje No: 6963

xiii
ABSTRACT
Introductıon:Rheumatic heart disease (RHD) is an autoimmune sequela of group A
streptococcal infection complicated by rheumatic fever (RF).Both rheumatic fever and
rheumatic heart disease are considered as the results of abnormal immune response after a
Group A streptococcal infection. Persistent inflammatory process occurs in the heart tissue,
even in the absence of the infectious agent, but the severity of sequela on heart valves is not the
same. Some patients have severe valvular damage with fibrosis and calcification at younger
ages, while others have mild lesions for the rest of their lives. Cross reactive T cells and
inflammatory cytokines have a crucial role in both triggering and maintaining inflammation in
the heart. Interferon gamma (IFN-γ) is a multifunctional cytokine and plays an important role in
modulating almost all phases of the immune response.IFN-γ may be responsible for the
increased valvular fibrosis and calcification in the pathogenesis of RHD. However the role of
IFN-γ genetic variant in RHD has not been studied. The aim of this study is to investigate the
possible relationship between the IFN-γ gene polymorphisms and RHD in the Turkish
population.Methods:IFN-γ polymorphisms (positions -874) were determined with ARMS
(Amplification Refractory Mutation System) PCR in 152 RHD Turkish patients and 151
healthy controls. Patients were classified into two groups; first group consisted of patients with
severe valve damage (SVD) and were treated with valve replacement or balloon valvuloplasty,
second group consisted of mild valvular damage (MVL).Results: IFN-γ 874 T allele and TT
genotype were significantly increased in patients compared to healthy controls (for T allel
p=0.002, OR= 2.40 and for TT genotype p=0.018, OR= 2.14, respectively). Patients with SVD
showed increased frequencies of -874 TT (p=0.009, OR= 3.66)and patients with a history of
valvuloplasty showed increased frequencies of -874 TT (p=0.012, OR= 2.75). Moreover
decreased frequencies of INF gama 874 AA genotype (p=0.01, OR= 3.06) were found in
patients with a history of valve replacement .Conclusıon: The data demonstrated that RHD is
associated with IFN-γ 874 T-A polymorphism in the Turkish population and these
polymorphism (IFN-γ 874 T) has a relation with severe valve damage. A allel was found to
have a protective effect against severe valve damage.Keywords: Rheumatic heart disease, valve
damage, IFN-γ polymorphism, genetic susceptibility.
The present work was supported by the Research Fund of Istanbul University. Project No. 6963

1. GİRİŞ VE AMAÇ
Romatizmal Kalp Kapak Hastalığının (RKKH) patogenezinde otoimmun reaksiyonlar
mevcuttur. Bu hastalıkta Streptokokus Pyogenes bakterisinin virulan faktörü olan M proteinine
karşı gelişen konakçı immunitenin, kardiyak proteinler ile çapraz reaksiyona girmesi sonucu
oluşan inflamasyon mevcuttur. Buradaki inflamasyon, M proteini olmaksızın otoimmun ataklar
halinde oluşmakta ve kalp kapaklarında ilerleyici hasarlara neden olmaktadır.
Kalp kapaklarında meydana gelen fibrotik ve inflamatuvar değişiklikler, kapaklarda
yapısal bozukluklar, kalınlaşma ve fonksiyon bozukluklarına neden olmaktadır. Fonksiyon
bozuklukları, klinikte kapak darlığı ve/veya kapak yetersizliği olarak ortaya çıkmaktadır.
RKKH’ ında kapaklardaki otoimmun inflamasyon, geçikmiş tip aşırı duyarlılık
reaksiyonu olup, bu reaksiyonun ana sitokini Interferon gama (IFN-gama) dır. IFN gama,
kapaklarda süregelen inflamasyonun şiddetini, karakterini ve iyileşme sürecindeki fibrotik
prosesleri, inflamatuvar hücrelerde ve fibroblastlarda bulunan reseptörleri aracılığıyla
etkilemektedir.
Akut ateşli romatizma geçirmiş, bunu takiben Romatizmal Kalp Kapak Hastalığı
gelişmiş, klinik takipte etkin Romatizmal Ateş sekonder profilaksisi yapılmış olan hastaların
tümünde romatizmal kapak tutulumunun ciddiyeti aynı düzeyde olmamaktadır. Aynı yaş
grubuna ulaşmış hastaların bir bölümünde kapak replasmanı veya balon valvuloplasti
endikasyonu oluşturacak ciddiyette kapak lezyonları mevcut iken diğerlerinde uzun süreli klinik
izlemeye rağmen aynı derecede kapak tutulumu görülmemektedir. Bu durum otoimmun
inflamasyona genetik zeminde farklı düzeylerde aşırı duyarlılık reaksiyonları olabileceğini akla
getirmektedir ve bu inflamatuar süreçteki ana sitokin INF gamadır.
Bu çalışmanın amacı, RKKH nın merkezinde rol oynayan IFN gama sitokininin genetik
olarak yapımını etkileyen 874 T-A polimorfizmi ile kalp kapaklarında oluşan romatizmal
hasarın ciddiyeti arasındaki ilişkiyi araştırmaktır.

2
2. GENEL BİLGİLER
ROMATİZMAL ATEŞ
Günümüzde; özellikle gelişmiş ülkelerde romatizmal ateş (RA) yirminci yüzyılın erken
ve orta dönemlerine göre daha az sıklıkla görülmektedir. Gelişmiş ülkelerde RA’nın
insidansının hızla azalmış olmasına rağmen dünya nüfusunun yaklaşık üçte ikisini oluşturan
gelişmekte olan ülkelerin bir çoğunda streptokok enfeksiyonları, RA ve romatizmal kalp kapak
hastalığı (RKKH) çok önemli bir halk sağlığı problemi olarak varlığını sürdürmektedir. Bir çok
ülkede RKKH çocuklarda ve 40 yaş altı genç erişkinlerde kardiyak mortalitenin en sık
sebebidir. Bizim ülkemizde de RA insidansı 20 / 100.000 olarak bildirilmiştir (1).
EPİDEMİYOLOJİ
Akut RA’nın epidemiyolojisi , A grubu streptokokların yaptığı üst solunum yolu
enfeksiyonlarınınki ile özdeştir. Streptokoksik farenjit gibi, akut RA da daha çok çocuklarda
ortaya çıkmaktadır ve insidansı 5-15 yaşları arasında en üst seviyeye ulaşmaktadır. İlk atak
erişkinde ortaya çıkmışsa bu çoğunlukla yaşamın 2. dekadının sonu ile 3. dekadının başında
meydana gelir. Ender olarak ilk atak 4. hatta 5. dekadlarda da görülebilmektedir.
Epidemiyolojik risk faktörleri; klasik olarak, bireysel ataklarla ve özellikle kalabalık
ortamlarda düşük yaşam standartlarına sahip bireylerdeki akut romatizmal ateş salgınlarıyla
ilişkilidir. Etkilenmiş bir toplumda yaygın serotiplere karşı genel konakçı immunitesinin
derecesi kadar, organizmanın kendisinin de eşit önemde bir risk faktörü olduğu sonucuna
varılabilir (1).
Çalışmalar, tedavi edilmemiş streptokoksik farenjiti olan kişilerin yaklaşık % 3’ ünde
RA geliştiğini göstermiştir. RA’nın epidemiyolojisi toplumda mevcut olan A grubu streptokok
serotipleri tarafından da etkilenmektedir. Bazı serotiplere sahip streptokokların romatojen
özellik gösterdiğine ilişkin epidemiyolojik kanıtlar vardır. Bu serotipler 1, 3, 5, 6, 18’ dir.
TANI
RA tanısı koydurabilecek özel bir laboratuar testi yoktur. Dolayısıyla tanı klinik olarak
konulmakta fakat klinik mikrobiyoloji ve klinik immünoloji laboratuarlarının desteğini
gerektirmektedir. RA sendromu ile ilişkili bulgu ve semptomların çeşitliliği nedeniyle ilk kez
1944’de Jones, klinisyenlerinin RA tanısı koymasını standardize etmeye yardımcı olacak
kriterleri ortaya koymuştur. Jones kriterlerinin en son modifikasyonu ( güncelleştirilmiş Jones
kriterleri) 1992 de Amerikan Kalp Derneği nin özel bir grubu tarafından yayınlanmıştır

3
(Tablo2- 1).
MAJÖR KRİTERLER : MİNÖR KRİTERLER :
Kardit
Gezici poliatrit
Eritema Marginatum
Sydenham Koresi
Subkutan nodüller
Ateş
Artralji
Akut faz reaktanlarının artışı(ESR,CRP)
Uzamış PR intervali
DAHA ÖNCE KANITLANMIŞ ROMATİZMAL ATEŞ GEÇİRENLERDE :
Bir major kriter + streptokok enfeksiyonu bulgusu
veya
ateş + artralji + akut faz reaktanlarında artış + streptokok enfeksiyonu bulgusu olması yeterlidir.
Tablo 2-1:Güncelleştirilmiş Jones kriterleri
RA hastalarında en sık bulunmaları nedeniyle beş kriter major olarak adlandırılmıştır.
Bunlar kardit, gezici poliartrit, Sydenham koresi, subkutan nodüller ve eritema marginatumdur.
Jones kriterlerinin sağlanabilmesi için iki major veya bir major iki minör kriterlerin yanısıra
geçirilmiş streptokok enfeksiyonu kanıtlarının olması gereklidir (1).
BUNLARA EK OLARAK;
Geçirilmiş A grubu beta hemolitik streptokok bulgusu olması (boğaz kültürü, ASO, boğaz
sürüntüsünde hızlı antijen testi, kızıl geçirme)
TANI İÇİN :
2 Majör + Streptokok enfeksiyonu bulgusu
VEYA
1 Majör +2 Minör +Streptokok enfeksiyonu bulgusu
Korea veya başka bir nedene bağlanamayan kardit tek başına yeterli kabul edilir.

4
Akut romatizmal ateşdeki kardit; perikard, miyokard ve endokardı tutan bir pankardittir.
Yayınlanmış serilerin çoğunda, akut romatizmal ateşli hastaların % 40-% 60’ ında sinüs
taşikardisi, mitral yetersizliği üfürümü, S3 gallop, perikardiyal frotman ve kardiyomegali
bulgularından biri veya daha fazlası ile karakterize olan kardit kanıtları vardır (1).
Ekokardiyografinin devreye girmesi mitral kapağındaki gizli bozuklukların açığa çıkarılmasında
yardımcı olmuştur. Hastalarda uzamış PR aralığı ve kalp yetersizliği kanıtları da mevcut
olabilmektedir.
Romatizmal ateşin neden olduğu valvulitin iyileşmesi sırasında kapaklarda gelişen
fibrozis, kalınlaşma ve yapışıklıklar, romatizmal ateşin en ciddi komplikasyonları olan kapak
darlığı ve yetersizlikleri ile sonuçlanmaktadır. En sık mitral kapak, daha sonra aort kapağı
tutulmaktadır. Akut romatizmal ateş sonucu ortaya çıkan izole aort kapağı hastalığı oldukça
seyrektir. Romatizmal ateş sonucu aort kapağı hastalığı olan hastalarda hemen her zaman eş
zamanlı olarak mitral kapağı da etkilenmektedir (1).
2.1. ROMATİZMAL KALP KAPAK HASTALIĞI
RKKH, akut romatizmal pankarditis’in iyileşme döneminde kalp kapaklarının
şekillerinin ve yapısal özelliklerinin bozulması sonucu oluşur. Bu deformasyon sürecinde
kapaklardaki inflamasyonun şiddeti önemlidir. RA’nın meydana getirdiği bu inflamatuvar hasar
kalp kapaklarının fonksiyonlarının bozulmasına (kapak yetersizliği ya da darlığı) yol açar.
Fonksiyonları bozulmuş kapaklar ise, hemodinamik değişikliklere neden olur. Bu hemodinamik
değişiklikler başta kalp ve akciğer olmak üzere diğer dokularda da organik ve fonksiyonel
değişikliklere neden olarak klinik tabloyu oluşturur.
İlk RA atağının başlangıcında ya da seyrinde veya hastalığın iyileşmesinden hemen
sonra tespit edilen ve devam eden aşikar bir kalp kapak hastalığı olabilir. Ancak vakaların bir
kısmında, ilk atak sırasında ya da sonrasında kalp kapaklarında herhangi bir bozukluk tespit
edilememesine rağmen yıllar sonra yeni atak olmaksızın RKKH görülebilir. Bununla beraber,
erişkin çağında kronik RKKH saptanan hastaların yaklaşık % 40’ında daha önce geçirilmiş bir
romatizmal ateş hikayesine rastlanmayabilir. Bütün bunlar, kapak hastalığında üzerinde durulan
otoimmun mekanizmaları destekleyen bulgulardır (1).
Gelişmiş ülkelerde RKKH’nın prevalansı bütün yaş grupları için % 0.7-1 arasında, 45
yaş üstündekilerde ise % 1.3 ‘tür. Gelişmekte olan ülkelerde çocukluk çağında görülen kalp
hastalığının en sık nedeni romatizmal kalp hastalığıdır. Bu ülkelerde 45 yaş üzerindeki toplum
bireylerinde iskemik ve hipertansif kalp hastalığına çok sık rastlanıldığından RKKH bu yaş
grubunda 3.sıraya düşmektedir.

5
RKKH başlıca, kalbin sol tarafındaki kapakları tutar. Bunlardan mitral kapağın tutulma
insidansı % 85, aort kapağının tutulma insidansı ise % 44 dür. Romatizmal triküspit lezyonu
diğerlerine nispeten seyrektir ( % 10-15), pulmoner kapak tutulumu ise çok nadirdir. RA
geçirmiş erişkinlerde en sık görülen valvülopati mitral darlığıdır. Bunu sıklık sırası ile mitral
yetersizliği, aort yetersizliği ve aort darlığı izler. RA ‘nın tek bir kalp kapağını tuttuğu vakalar
olmakla birlikte, birden fazla kapak tutulumu daha sık rastlanan bir bulgudur.
RKKH ‘da tutulmuş olan kalp kapağına göre de bir cinsiyet farkı dikkati çeker. RKKH
daha çok kadınlarda görülür. Bunlardan mitral darlığı olan kadınlar erkeklere oranla 2-9 kat
daha fazladır. Mitral darlığı ve mitral yetersizliğinin beraber olduğu durumlar yine kadınlarda,
erkeklere oranla 3 kat daha fazla görülür. Tek başına mitral yetersizliğinin görülme sıklığı her
iki cinsde de aynıdır. Diğer taraftan romatizmal aort kapak hastalığı erkeklerde kadınlara oranla
1.5 kat ,triküspit darlığı kadınlarda erkeklere oranla 1,5 kat daha fazla görülür. Otoimmunite
nedenli oluşan hastalıkların kadınlarda erkeklere oranla daha fazla görülmesinin RKKH daki bu
tabloya uygun olduğu söylenebilir (1).
ROMATİZMAL KALP KAPAK HASTALIĞININ PATOGENEZİ
RA , insanda moleküler benzerlik nedeniyle oluşan çapraz reaksiyona bağlı otoimmun
hastalığa en çok uyan örnektir. Bu hastalığın tetiği streptokokus Pyogenes bakterisi tarafından
çekilir ve tedavi edilmemiş çocukların % 3-4’ ü etkilenir. Streptokoksik antijenlere karşı verilen
immun cevap , aynı zamanda bireyin kendi doku proteinleri ile çapraz reaksiyonu da başlatır.
Bu protein kalp kapağında laminindir. RKKH romatizmal ateşin en ciddi komplikasyonudur ve
grup A streptokoksik infeksiyondan 4-8 hafta sonra bireylerin % 30-40’ da görülür (2) .
GRUP A STREPTOKOKLAR
Grup A streptokoklar hafif seyirli boğaz ve deri infeksiyonundan ( farenjit, impedigo)
şiddetli hatta hayati tehlike oluşturabilen septik kan , kas, böbrek, karaciğer ( nekrozitan fasciit,
Scarlet ateşi ve sterptokoksik toksik şok sendromu) gibi infeksiyonlara neden olabilir (3) .
Ayrıca; hastalarda immun nedenli post streptokoksik sekel de gelişebilir (Örn: akut romatizmal
ateş ve akut glomerülonefrit). RA; artritis, Sydenham koresi ve karditis gibi manifestasyonlar ile
karşımıza çıkabilir. Coburn 1930’ da Streptokokus pyogenesin boğaz enfeksiyonundan sonra
RA’ya neden olduğunu açıklayan ilk hekimdir (4).
Lancefield, 1941’de streptokoksik bakterileri hücre duvarındaki polisakkarid yapılarına
göre serolojik olarak gruplandırmıştır ( grup A,B,C,F,G ). A grubu streptokoklar, hücre
duvarında tekrarlayan N-asetil ß-D- glukozamin karbonhidratları içerir. Streptokokus Pyogenes
A grubundandır. Ek olarak, grup A streptokoklar M, T, R yüzey proteinleri ve lipoteikoik asit

6
(LTA) de içerir. Bu proteinler, bakterinin boğaz epiteline tutunmasında, dolayısıyla
virulansında rol oynamaktadır. Hücre yüzeyindeki kompleks yapısı özellikle konak
kolonizasyonu, fagositozdan korunma ve konağın immun cevabından korunmada S. pyogenes
virulansını belirler. M proteini virulan faktör olarak özellikle önemlidir (5) .
M proteini, bakterinin hücre duvarında bulunur ve yapısında her biri 450 amino asit
içeren iki polipeptid zincir vardır. Bu proteinin N-terminal parçasında A ve B bölgesi olarak
adlandırılan tekrarlı amino asit dizileri mevcuttur. A bölgesi serolojik adlandırmada kullanılır
ve buna göre 130 adet sınıflandırılmıştır. B tekrarlayıcı bölgesi streptoksik seroloji hakkında
antijenik varyasyonlar oluşturur. Proteinin C-terminal parçası yapısal koruma görevi yapmakta
olup çoklu tekrar bölgeleri içermektedir (6). M proteini bakterinin antijenik yapısında
önemlidir ve konağın kendi dokuları ile çapraz reaksiyon göstermesine neden olur. Konak
dokuda bulunan Miyozin, tropomiyozin, keratin, laminin, vimentin gibi kardiyak proteinler ile
M proteininin α-helikal yapısı arasında yapısal benzerlik vardır ( 7-8-9-10-11). Bu benzerlik
çapraz reaksiyon nedenli otoimmuniteyi başlatarak konak dokusunda post streptokoksik
sekeller oluşturmaktadır.
OTOİMMUNİTE
Otoimmunite, immun toleransı düzenleyen temel mekanizmalardan bir veya daha
fazlasının disfonksiyonunu temsil eder. Otoimmun hastalıkların başlıca özelliği olan doku
hasarı, organizmanın kendi dokularına immunolojik reaksiyonundan kaynaklanır.
Normal süreçte immun toleransın bozulması, otoimmunite gelişmesine eğilimi arttırır.
Genel olarak bu anormal yanıtlar; genellikle bakteriyel uyarılar, viral ekzojen uyarılar veya
immun sistemdeki hücrelerin endojen anormallikleri ile ilişkilidir. Bu nedenle, otoreaktivite
self-antijenlere yanıtsızlığın düzenlenmesinin üstesinden gelecek tarzda immun sistemin
ekzojen stimülasyonundan kaynaklanır. Otoantijene reaktif T ve / veya B hücreleri reseptör
eksprese ederse otoimmunite gelişebilir. Tersine, moleküler benzerlik veya mikrobiyal ürün ile
herhangi bir self-antijen arası çapraz-reaktivite, otoreaktif lenfositlerin aktivasyonuna yol
açabilir. Moleküler benzerlikten kaynaklanan otoreaktivite ve otoimmun hastalığın en iyi
örneklerinden biri RA ve RKKH’ dır. Burada antikorlar; streptokokların M proteinlerine karşı
miyozin, laminin ve diğer matriks proteinleri ile çapraz reaksiyon verir. Bu otoantikorların
kalpte depolanması inflamatuvar bir yanıtı başlatır. T ve / veya B hücre aktivitelerinde, sitokin
dengesinde veya defektif immunregülatuvar döngülerdeki primer değişiklikler otoimmunitenin
oluşumuna katkıda bulunur. TNF-α, IL-10 ve IFN γ gibi sitokinlerin ekspresyon seviyelerindeki
kalıtılmış değişikliklerin, otoimmun hastalığa duyarlılığı arttırdığına dair kanıtlar mevcuttur (35
37-69).

7
ROMATİZMAL KALP KAPAK HASTALIĞINDA OTOİMMUN REAKSİYONLAR
RKKH nın patogenezindeki otoimmun reaksiyon geçikmiş tip aşırı duyarlılık
reaksiyonudur. Oluşumunda kazanılmış immunitenin rolü vardır. Kazanılmış immunite ,
yabancı bir antijen veya patojene verilen antijen-spesifik yanıtlarla karakterizedir ve ani ortaya
çıkan doğal immunite ile karşılaştırıldığında oluşumu birkaç gün veya daha uzun bir süreyi
içerir. Kazanılmış immunitede önemli rol oynayan hücre T hücresi olup ana sitokin IFN γ dır.
T hücreleri nativ protein veya karbonhidrat antijenlerini tanıyamazlar. Bunun yerine T
hücreleri yalnız antijen sunucu hücreler (ASH) tarafından taşınan veya üretilen protein
antijenlerinden türeyen peptid fragmanlarını tanırlar. Bu peptit fragmanları HLA antijenleri
olarak bilinir. RKKH ında rol oynayan HLA antijenleri sınıf II grubundadır.
HLA sınıf II moleküller ile T hücresine antijen tanıtıldıktan sonra T hücrelerinde aktivasyon
sinyalleri nükleus’a iletilir ve IFN γ , IL-2 gibi sitokinlerin gen eksresyonu başlar. IFN γ
ekspresyonu T hücresine otokrin etki yaparak TH1 yönünde farklılaşmasını sağlar. Ayrıca
makrofajlara etki ederek inflamasyonun şiddetini arttırır.
HLA ilişkili otoimmünitede , immün toleranstan kaçan ve external antijenik uyarıya
benzeyen kendi molekülleri ile çapraz reaksiyona girebilen periferik T hücrelerinin aktivasyonu
vardır (12). RKKH ında M proteini ile birçok kalp proteini çapraz reaksiyona girmektedir.
Böylece bu hastalarda moleküler benzerlik nedeniyle otoimmun reaksiyonun tetiği
çekilmektedir (13). Eğer INF-γ gen ekspresyonunda kalıtsal farklar mevcutsa bu sitokinin
miktarı bireyler arasında farklılıklara neden olur ve immün fonksiyon farklılık gösterebilir.
Yardımcı T hücreleri (Th) ürettikleri sitokinlerine göre alt gruplara ayrılır. Aktive Th1
tipi yardımcı T hücreleri; IFN γ,IL-2, IL-3,TNF-ß, GM-CSF; aktive Th2 tipi yardımcı T
hücreleri ise IL-3,-4,-5,-6,-10 ve -13 salgılar. Th1 hücreleri, IFN-γ aracılığı ile çeşitli
patojenlerin intrasellüler alanda öldürülmesi, antijen sunumunun arttırılması ve immün sistemin
yönlendirilmesinde merkezi bir rol oynar.
RA; eklem, santral sinir sistemi ve kalp inflamasyonuna neden olabilir. Akut romatizmal
kardit önce humoral immun cevap ile başlar. Otoantikorlar kalp kapağında ve bazal membranda
reksiyona girerek kalp dokusu içerisine hücresel infiltrasyonu kolaylaştırır (14-15). Romatizmal
kalp hastalığının patognomik bulgusu granülomatöz bir lezyon olan Aschoff nodülleri oluşur.
Burada T ve B hücreleri, makrofajlar, geniş mononükleer hücreler, multinükleer hücreler ve
polimorf nüveli lökositler hem miyokardiyumda hem de endokardiyumda görülür (16).
Miyokardiyum ve endokardiyumdaki bu gelişmeler muhtemelen endotel yoluyla hücresel
infiltrasyonun bir sonucudur. Romatizmal kalp kapak lezyonlarında CD+4 T hücre ağırlıklı

8
mononükleer hücre infiltrasyonunun varlığı romatizmal kalp kapak hastalığının ilk bulgusudur
(17-18).
RKKH’ında kapak lezyonlarının içinde T hücre kolonilerinde % 62 oranında baskın olan
reaktif M protein bölgesi M5(81-103) dir. Bu bölgede 22 amino asit rezidüsü mevcuttur. 3 adet
iç içe geçmiş peptid içermektedir, bunlar, M5( 81-96), M5( 83-103), M5( 91-103)’dir. Bazı M5
epitopları özellikle periferik T hücre kolonileri tarafından tanınabilir (12-13). Bu sonuçlar
RKKH’daki otoimmün reaksiyonların tetiğini çeken streptokoksik antijenlerden birinin M5( 81-
103) bölgesi olduğunu göstermiştir (13).
RKKH’ındaki inflamasyon gelişiminde sitokinlerin rolü büyüktür. Genel olarak Sitokinler,
çok çeşitli hematopoetik ve hematopoetik olmayan hücre tipleri tarafından üretilen çözünebilir
proteinlerdir ve hücresel aktivasyon, büyüme, farklılaşma, fonksiyonel hücre yüzeyi molekül
ekspresyonu ve hücresel efektör fonksiyona yol açan gen aktivasyonu üzerinden etkilerini
gösterirler. Bundan dolayı, sitokinlerin immun yanıtların düzenlenmesinde ve bir çok hastalığın
patogenezinde önemli etkileri vardır.
Streptokoksik antijen ile uyarılmasından sonra proinflamatuar sitokinlerin üretiminde
polimorf nüveli lökosit ve tonsiller mononükleer hücreler arasında farklar görülmüştür. TNF-α,
IL-1 ve IL-2’nin yapımı polimorf nüveli lökositler aracılığıyla arttırılırken, tonsillerdeki
mononükleer hücreler tarafından azaltılır (19). Akut RA ve RKKH olanlarda, IL-2’nin
yapımının artmış olduğu rapor edilmiştir. Bu hastalarda aktif inflamasyon varlığının göstergesi
olan Th hücreleri de görülmüştür.
RKKH olan hastalarda TNF-α plazma seviyeleri artmıştır. Kalp lezyonlarında,
romatizmal kalp hastalığının akut fazında, IL-1,TNF-α ve IL-2 üretimi ile Aschoff nodülünün
progresyonu arasında korelasyon mevcuttur (20). Kalp lezyonlarından alınan mononükleer
hücrelerin inflamasyon sürecinde baskın olarak IFN-γ ve TNF- α sekrete ettiği gösterilmiştir.
Bu sitokinler Th1 sitokinlerdir ve hem akut hem de kronik fazdaki RKKH’da sekrete edilir.
Kalp kapağını infiltre etmiş hücrelerde IL- 4 sekresyonu % 10 ‘dan daha azdır. Miyokardiyumu
infiltre etmiş hücrelerde IL- 4 sekresyonu % 50’ nin üzerindedir. Bu durum, kapak dokusundaki
IL-4 üretimindeki azalmanın kalp kapak hasarı progresyonuna katkıda bulunduğunu
göstermektedir (21). İn vitro analizlerde, miyokardiyumdaki T hücre kolonilerinin IL-4 ve IL-10
ürettikleri, buna karşın kalp kapak kaynaklı T hücrelerinin IL-4 ve IL-10 sitokinlerini
miyokardiyumdaki T hücre kolonilerine göre daha az ürettikleri görülmüştür. Geçikmiş tip aşırı
duyarlılık reaksiyonlarında artmış INF-γ düzeyi ile IL-4 arasında ters ilişki mevcuttur. Farklı
olarak, streptokoksik M5 antijeni ile uyarılmış T hücre kolonileri RKKH vakalarının % 85 ‘inde
T hücrelerinin lokalizasyonuna bakılmaksızın (miyokard ya da valvül) IFN-γ üretir. Bu
romatizmal kalp hastalığındaki iyileşmenin ya da kapak hasarının kalıcı veya ilerleyici
olmasında belirleyicidir (21).

9
2.1.1. ROMATİZMAL KALP KAPAK HASTALIĞININ GENETİK TEMELLERİ
AİLESEL YATKINLIK
RKKH’da ailesel ilişkiyi gösteren birkaç çalışma yapılmıştır ve bu çalışmaların
bazılarında hastalık dağılımının basit resesif geçişli olduğu rapor edilmiştir (22-23). Ancak
diğer çalışmalarda bu ilişki gösterilememiştir (24-25). Ailesel kalıtımla ilgili farklı sonuçlara
rağmen RA ve RKKH’da bireylerin genetik yatkınlığı üzerinde yıllarca durulmuştur. İkizlerde
yapılan çalışmaların sonuçlarına göre, monozigot ikizlerde (% 18.7) dizigot ikizlere (% 2.5)
göre RA ve RKKH görülme riski daha yüksek bulunmuştur (26) .
HLA DOKU ANTİJENLERİ
HLA molekülleri antijen sunan hücrelerde antijenin T hücrelerine tanıtılmasında rol oynayan
moleküllerdir. T hücresi yüzeyindeki reseptörlere bağlanarak inflamatuvar süreci başlatırlar.
Akut RA ve RKKH’da kalp dokularıyla benzer moleküler yapıya sahip olan M proteinin bazı
fraksiyonları ( M5 81-103) inflamatuvar süreçte bazı HLA molekülleri ile T hücrelerine sunulur.
Bu HLA molekülleri topluluklar arasında farklılık gösterebilir. HLA moleküllerindeki bu
farklılık RKKH’daki genetik yatkınlıkla ilişkilendirilebilir.
Bazı çalışmalarda, HLA sınıf I antijenlerden olan hem HLA-A hemde HLA-B ler ile
RA arasında ilişki kurulmasına rağmen, sonuçlar çelişki göstermiştir ( 27-28) . HLA sınıf I
antijenler hücre içerisindeki ya kendi antijenlerini ya da intrasellüler patojen antijenlerini T
hücre reseptörüne sunmada rol oynar. HLA Sınıf II antijenler extrasellüler antijenleri sunarak
kazanılmış immun sistemi aktive eder. M proteini ekstrasellüler antijen olup T hücresine HLA
sınıf II antijenler ile sunulur, dolayısıyla RA ve RKKH’a yatkınlık ile HLA sınıf I antijenler
arasında ilişki yoktur.
Romatizmal kalp hastalığı ile HLA sınıf II antijenleri arasındaki ilişkiyi ilk olarak Ayoub
ve arkadaşları göstermiştir (29). Bu çalışmada Afrika kökenli Amerikalılarda HLA-DR2,
Kafkas kökenli Amerikalılarda HLA-DR4 ilişkisi bulunduğu bildirilmiştir.
Farklı ırklarda yapılan çalışmalarda HLA sınıf II allelleri ile RKKH na yatkınlık arasında
2.3 kattan 13.6 kat a kadar ulaşan kuvvetli bir ilişki mevcuttur. Bu ilişki HLA sınıf II
alellerinin romatizmal kalp hastalığının gelişimindeki önemli rolünü göstermektedir.

10
RKKH’nın patogenezindeki antijen sunumunda HLA sınıf II antijenlerinin önemli rolünün
olmasından dolayı yapılan çalışmalardaki bu korelasyon şaşırtıcı değildir. Bu sonuçlar
RKKH’daki etnik dağılımı açıklamaya katkıda bulunmuştur.
HLA DIŞI MOLEKÜLLER
RKKH oluşumunda hem doğal hem de kazanılmış immunite rol oynar. Bu hastalığın
oluşum süresinde sadece HLA antijenleri değil doğal ve kazanılmış immunitede görevli diğer
moleküllerin de rolü vardır. Bu moleküller genel olarak HLA dışı moleküller olarak adlandırılır.
Bu moleküller Mannoz bağlayıcı lectin (MBL), Tool-Like reseptörler (TLR) ve bazı
sitokinlerdir. Bütün bu moleküller RKKH sürecindeki inflamasyonda rol oynar.
MBL çözünür haldeki patojenleri tanımaya yardımcı olan bir akut faz proteinidir. Bu
molekül birçok patojenin yüzeyinde bulunan şeker moleküllerini tanır ve patojenlere karşı
verilen doğal immunitede rol oynar. RKKH oluşumunda rol oynayan streptokokların
membranında bulunan N-asetil glikozamin‘e bağlanarak bakterinin fagosite edilmesini sağlar.
MBL 10. kromozom üzerinde bulunan MBL2 geni tarafından kodlanır. Bu genin 1.ekzonundaki
mutasyonlar MBL’nin plazma seviyelerine etki eder. Reason ve ark. tarafından yapılan
çalışmada MBL nin yüksek gen ekspresyonu ile korele genotipe sahip bireylerde RKKH
gelişimi riski arasında ilişki bulunmuştur (30) .
TLR patojenleri tanımaya yardımcı olan reseptördür. Patojenler algılandıktan sonra
doğal immüniteyi başlatırlar. Türk popülasyonunda bu reseptörlerdeki polimorfizm ile yapılmış
çalışmalarda RKKH yatkınlık açısından çelişkili sonuçlar rapor edilmiştir. Berdelli ve ark.’nın
yaptığı bu çalışmalarda TLR deki Arg 753 Gln değişimine neden olan TLR2 genindeki
polimorfim ile RA arasında çok kuvvetli ilişki rapor edilmiş fakat bu populasyondaki ikinci
çalışmada bu ilişki gösterilememiştir ( 31-32) .
RKKH sürecinde önemli rolü olan diğer bir molekül ise sitokinlerdir. Sitokinler
inflamatuvar sürecin her aşamasında rol oynar. IL-1 ailesi bu sitokinlerden biridir ve 2.
kromozom tarafından kodlanır. Bu gen proinflamauvar IL-1α ve IL-1ß ve onların inhibitörü IL-
1 reseptör antagonistini (IL-1RA) eksprese eder. IL-1ß ve IL-1 RA varyasyonları ile yapılmış
bir çalışmada RKKH arasında ilişki görülmemiştir ( 33).
TNF-α diğer bir inflamatuvar sitokindir. RKKH da IFN gama ile beraber ekspresyonu
en fazla olan sitokindir. TNF- α gen lokusu MHC sınıf III gen lokusuna yakındır. TNF–α gen
polimorfizmi ile yapılan 3 bağımsız çalışmada RKKH arasında ilişki gösterilmiştir. Brezilya
toplumunda, TNF- α allelleri’nin (-308 A ve – 238 A) RKKH gelişminde ilişkili olduğu
görülmüştür. Bu ilişkinin aort kapağının tutulduğu hastalarda daha kuvvetli olduğu rapor
edilmiştir (34).

11
Türk ve Meksikalılarda , TNF- α -308 A alleli RKKH ile ilişkili bulunmuştur ( 35).
Türk populasyonunda yapılan başka bir çalışmada bu ilişkiler görülmemiştir (36).
TNF- α -308 A aleli ile RKKH arasındaki ilişkinin altında yatan mekanizma tam
olarak bilinmemektedir ancak, TNF -308 A alelinin varlığı TNF- α gen ekspresyonunu NF-kß
aracılığı ile arttırdığını gösteren çalışmalar mevcuttur (37) .
2.1.1.1. İNTERFERON GAMA ( INF γ )
1957 yılında, inaktive viruslar ile temas eden hücrelerin en az bir tane olmak üzere
eriyebilme özelliğine sahip bir faktör ürettikleri ve bunun yeni enfekte olmuş hücrelere
uygulandığında viral replikasyonu önlediği keşfedilmiştir. Bu faktöre İnterfer: mani olmak,
engel olmak kelimesinden esinlenerek interferon adı verilmiştir. İnterferonların sınıflandırılması
aminoasit sıralamasına, üretildiği hücreye, antijen için gerekli stimulus gibi kriterlere göre
yapılmaktadır. İnterferonların bir tanesi hemen hemen bütün özellikleriyle diğerlerinden ayrılır.
IFN-gama olarak isimlendirilen bu interferon diğerleri gibi antiviral etki göstermekle beraber en
önemli özelliği immünoregulatör bir molekül olmasıdır.
Tip II interferon; Gamma interferon veya immun IFN diye de adlandırılır. Molekül
ağırlığı 20000-25000 olan tek aktif IFN-γ türü vardır. IFN γ tek bir gen’e bağlı olarak
sentezlenir, kodlayan gen insanlarda 12. kromozom üzerinde bulunur, diğer interferonlar ile
hiçbir benzerlik göstermez. Bağlandıkları reseptörler, tip I interferonun reseptörlerinden
farklıdır. Anti-viral aktivitesi olmakla beraber, bu etkisi Tip I interferonlara göre çok düşüktür.
IFN-gama başlıca CD4+ T (özellikle Th1 grubu) hücreleri ve NK hücreleri tarafından sentez
edilir. Kazanılmış immunitede T hücreleri IFN γ’yı antijenin tanınmasına yanıt olarak salgılar
ve bu da yine IL-12 tarafından arttırılır. Neredeyse bütün hücre tipleri IFN γ için reseptör
taşırlar.
Sitokin’in reseptöre bağlanması STAT’ı aktive eder. Böylelikle IFN γ üretiminden
sorumlu genlerin transkripsiyonunu dolayısıyla da MHC molekülünü ve diğer sitokinleri
kodlayan genlerin transkripsiyonunu stimule eder. STAT1, IFN-γ’nın primer regülatörüdür. IFN
γ Sınıf II MHC proteinleri taşıyan hücrelerde bu proteinlerin miktarını arttırarak CD4+ T
lenfositlere karşı antijen sunumunu yükseltir.
IFN γ bilinen en güçlü makrofaj aktivatörüdür. IFN γ ’ya maruz kalma sonucunda;
makrofajların mikrobisidal aktiviteleri, daha düşük seviyede de sitotoksik kapasiteleri artar. IFN
γ makrofajları etkileyerek; IL-1, IL-6, IL-8, TNF-α gibi sitokinlerin salınmasına yol açar.
Bunun yanısıra proteolitik enzimler, transkripsiyon faktörleri de sentezleyerek özellikle
intrasellüler bakterilerin yol açtığı hastalıkların kontrolünde önemli rol oynar. Th 1 hücrelerin
aktivitelerini arttırarak hücresel immüniteyi arttırır. Th-1 tip sitokinlerin en belirleyici olanıdır.

12
Buna karşılık Th-2 hücrelerinin proliferasyonunu inhibe ederek hümoral immünitenin
baskılanmasına sebep olur. IFN-gama normalde kanda dolaşmaz, fizyolojik olarak otokrin veya
parakrin mekanizmalarla etki eder. IFN γ ’nın immunoregulator mekanizmadaki görevleri şöyle
sıralanabilir:
1. Antijen sunumunun arttırılması.
2. Fagositlerdeki reaktif oksidan (superoksit, hidrojen peroksit ve nitrit oksit) üretiminde
kullanılan enzimlerin sentezini artırarak intrasellüler patojenlerin öldürülmesi.
3. İnfektif ajanların yok edilme kapasitesinin arttırılması
4. Makrofaj aktivitesinin arttırılması ve intrasellüler antimikrobiyal konsantrasyonun
arttırılması.
5. Antijen sunan hücrelerin üzerindeki Sınıf II MHC moleküllerinin sunumunun arttırılması.
6. T hücrelerinin Th1 tipine dönüşümünü arttırıp Tip 2 hücre proliferasyonunun inhibe
edilmesi.
7. Nötrofillerin aktive edilmesi ve NK hücrelerinin sitolitik aktivitesininin arttırılması.
IFN- γ’ NIN MOLEKÜLER ETKİ MEKANİZMASI
İnterferon γ pleiotropik etki gösteren bir sitokindir ve bütün hücrelerde reseptörü
mevcuttur. Tetramerik yapıdaki reseptörlerine (IFNGR1 ve IFNGR2) bağlanır. Reseptör
oligomerizasyonundan sonra Janus tirozin kinazlar ( Jak 1 ve 2) aktive edilir. Takiben STAT-1
fosforilasyonu gerçekleşir. STAT dimerleri IFN-γ aktive edici faktör (GAF) olarak
isimlendirilir. STAT dimeri (GAF) nücleusa transloke olur ve genin GAS (IF-γ aktive edici
sensör) bölgesinin promotör kısmına bağlanarak gen transkripsiyonunu uyarır (38-39). IFN γ
genellikle GAF yolunu kullanmasına rağmen, ISGF ve diğer transkripsiyon faktörlerini de
kullanabilir (40). STAT1 ile etkileşime girebilen moleküller IFN-γ ’nın transkripsiyonel
aktivitesini değiştirebilir (41).
IFN γ diğer sitokinlerin fonksiyonlarını ve ekspresyonunu etkileyebilir. Bundan dolayı,
direkt etkilerine ek olarak indirekt etkilerini de tanımak önemlidir. IFN-γ diğer sitokinler ile
arasında antagonistik ya da sinerjik ilişki nedeniyle immünite ve otoimmunite etkilenebilir.
IFN γ kendi ekspresyonunu arttırabildiği gibi TNF-α ile sinerjik etkileşerek MHC sınıf I
molekülünün ekspresyonunu da arttırabilir (42). Ayrıca; makrofaj ve monositlerden IL-12
yapımını sağlamaktadır (43).
IL-12’de T hücrelerine ve NK hücrelerine IFN γ yaptırmaktadır (44). IFN γ ve IL-6
fibroblastlar ve epitelyum hücreleri üzerine kemokin yapımı için sinerjik etki gösterirler. IL-6
kemokinler aracılığı ile inflamasyon bölgesine nötrofil toplanmasına neden olmaktadır (46).
IFN γ ve IL-4 biribirlerinin fonksiyonlarını ve expresyonunu antagonize edebilirler.

13
DOĞAL VE KAZANILMIŞ İMMUN CEVABIN REGÜLASYONUNDA IFN γ’ NIN
ROLÜ
IFN γ uyarımından sonra, farklılaşmamış T hücresinden STAT-4 ve T bet transkripsiyon
faktörleri aracılığıyla Th-1 ve T hafıza hücreleri farklılaşır. Th-1 hücresinden IFN γ başta
olmak üzere IL-18, IL-12 gibi inflamasyonda önemli rollere sahip sitokin salınımı gerçekleşir
(44) .
B hücrelerinde IFN γ uyarımı T bet transkripsiyon faktörünün expresyonunu artırır. Bu
uyarım immunglobulin dönüşümüne neden olur ve B hücrelerinde Ig-G ekspresyonu artar.
Ayrıca IL-4 etkisini antagonize ederek Ig-E oluşumunu engeller (47-48).
IFN γ MHC sınıf II moleküllerinin yapımını arttırarak antijen sunan hücre ile CD4+ T-
yardımcı hücreleri arasındaki etkileşimi arttırır (49).
ROMATİZMAL KALP KAPAK HASTALIĞI PATOGENEZİNDE ROL OYNAYAN
HÜCRELER İLE INF γ ETKİLEŞİMİ
RKKH ’da histolojik incelemelerde inflamasyon bölgesinde T hücresinden zengin
inflamasyon dikkati çekmektedir. T hücrelerinin alt-grup incelemelerinde, Th-1 tipindeki T
hücreleri çoğunluktadır. Ayrıca inflamatuvar bölgede mononükleer hücreler, NK hücreleri de
mevcuttur.
IFN- γ VE MAKROFAJLAR:
Mononükleer fagositler patojenlere karşı konağın savunulmasında önemli rollere
sahiptir. Makrofajlar hüçre içi patojenleri yok ederek ve T hücrelerine antijen sunarak immun
sistemde rol oynamaktadır. Makrofajlar bu fonksiyonlarını Makrofaj aktivitör faktör (MAF)
olarak adlandırılan ve önemli bir komponentini IFN γ ’nın yaptığı moleküller ile arttırırlar.
Böylece makrofajlarda İnflamatuvar süreçte IFN γ etkileşimi ile hücre içi patojenlerin yok
edilme yeteneği ve IFN γ etkisi ile MHC sınıf II gen ekspresyonu arttırılır (50-51-52) .
RKKH ‘da M proteininin makrofaj içerisinde degrade edilip HLA antijenleri ile T
hücresine sunulması, T hücrelerine M proteininin bazı fraksiyonlarını tanıma özelliği kazandırır.
Bunu takiben M proteinini tanıyabilen T hücreleri, vücutta bulunan M proteinine benzer
moleküller ile çapraz reaksiyona girerek otoimmun kapak hastalığını oluşturur. Bu süreçte IFN
γ, makrofajlarda HLA antijen sunulmasını artırarak RKKH oluşumunu kolaylaştırır.

14
IFN GAMA VE T HÜCRESİ
IFN- γ’nın en önemli yapım yeri T hücreleridir. İnflamatuvar süreçte T hücresinden
salınarak otokrin ve parakrin etki gösterir.
Otokrin etki ; T hücrelerine etki ederek T hücresini Th olarak diferansiye eder ve
inflamatuvar yolu Th-1’ e doğru yönlendirir. Bu yol otoimmün hastalıkların patogenezindeki
yoldur ve Th-1 proliferasyonunu sağlar. Ayrıca yine otokrin etkiyle T hücresinin gelişmini de
sağlamaktadır (53) .
Parakrin etki; T hücresine antijen sunulmasını kolaylaştırır. Antijen sunan hücrede
MHC sınıf II gen ekspresyonunu artırır. Ayrıca B hücresine etki ederek Ig G yapımını
artırmaktadır (54).
RKKH’ da inflamatuvar bölgede Th-1 baskınlığı mevcuttur ve buradaki inflamasyonda
IFN γ ’nın hem doku destrüksiyonunda hem de inflamasyonun şiddetinde rolünün olması
şaşırtıcı değildir.
IFN- γ VE ANTİJEN SUNUCU HÜCRELER
IFN-γ antijen sunan hücrelerde MHC sınıf II moleküllerinin yapımını artırır fakat
antijen sunumunda önemli olan diğer membran reseptör moleküllerinin yapımını inhibe eder (
ör: MHC sınıf I) ( 55). Antijen sunan hücrelerin aktivasyonunda IL-12 molekülü önemlidir.
IFN γ IL-12 yapımını arttırarak bu hücrelerin aktivasyonunda önemli rol oynar (56) .
RKKH’ da kapaklardaki inflamatuvar süreçte antijen sunucu hücreler görev alır ve M-
proteinine benzer moleküllerin ( laminin, vimentin.) T hücrelerine sunulmasını sağlar.
IFN- γ GEN EKSPRESYONUNUN REGÜLASYONU
Ökaryotik hücrelerde gen ekspresyonu genel olarak iki yolla düzenlenir. Bu genlerin
ürünleri ya sürekli eksprese edilirler (örneğin albumin koagülasyon faktörlerinin sentezi gibi) ya
da ihtiyaç halinde ekspresyonları gerçekleşir ( örneğin sitokinler gibi).
RKKH ‘da çapraz reaksiyon nedeniyle T hücrelerinin tetiğini çektiği ve yönettiği
inflamatuar cevap oluşmakta ve oluşan doku hasarının sınırlandırılması yine inflamatuvar
hücrelerdeki IFN γ gen ekspresyonunun regülasyonu ile sağlanmaktadır. İnflamatuvar süreçteki
birbiri içine geçmiş moleküler mekanizmalar dokuya uygun spesifik IFN γ gen ekspresyonunu
oluşturmak için geliştirilmiştir.
IFN γ gen ekspresyonunun kontrolünde epigenetik olaylar, transkripsiyonun indüklenmesi
(transkripsiyon faktörleri), post transkripsiyonel ve post translasyonel modifikasyonlar rol
oynamaktadır.
İnterferon 146 amino asitten oluşan bir proteindir. 12. kromozomun uzun kolunun 14.
segmentinde yer alan IFN γ geni tarafından kodlanır. ( 12q14). Bu gen 4 exon ve 3 introndan

15
oluşmaktadır. Eksonlarda polimorfizm gösterilmemiştir, ancak intronlarda polimorfizm
mevcuttur ( Naylor et all. 1983) .
TRANSKRİPSİYON FAKTÖRLERİ
IFN γ gen lokusunda kromatin yapısının farklı bölgelerinde DNA’ ya bağlanabilen ve
genin transkripsiyonunu regüle eden proteinler mevcuttur. Bu proteinlere transkripsiyon
faktörleri denir. Bu transkripsiyon faktörleri IFN γ ekspresyonunda major regülatör göreve
sahiptir. IFN- gama geninde proksimal promotor bölgeye ve 1. intronik bölgeye bağlanırlar.
Başlıcaları; NF-kß, NFAT,AP-1,T-bet,STAR4 olup bunlar IFN- gama ekspresyonunu regüle
ederler.
NF-Kß AİLESİ
NF-kß DNA’nın transkripsiyonunu kontrol eden protein kompleksidir ve bütün
hücrelerde bulunur. Sitokinler, serbest oksijen radikalleri, ultraviyole ışınları, okside LDL,
bakteriyal ve viral antijenler gibi uyarılar ile hücresel cevaba neden olur. Genel olarak NF-kß;
sitokinlerin gen ekspresyonunda rol oynayarak infeksiyona karşı immun cevabın
regülasyonunda önemli rol oynar.
Memeli hücresinde 5 adet NF-kß proteini bulunur. Bunlar sınıf 1; NF-kß1, NF-kß2 ve sınıf
II; RelA, RelB ve c-Rel proteinleridir. Bu proteinlerin N-terminal uçlarında DNA bağlayan
bölgeleri vardır (57).
IFN γ gen regülasyonundan sorumlu olan nükleer faktör NF-kß1 dir. Bu faktörün IFN γ geni
1. intronik bölgede bağlanma yeri mevcuttur. IFN γ ekspresyonuna direkt etki eder. NF-kß hem doğal hem de kazanılmış immun cevapta gen regülasyonundan sorumlu major
transkripsiyon faktörüdür. T hücresinin gelişimi, farklılaşması ve proliferasyonundan
sorumludur (58).
T-BET
T-Box ailesinin bir üyesidir. IFN γ ekspresyonunu düzenleyen nükleer faktörlerden biridir.
STAT-4’ e benzer. IFN γ geni promotör bölgelerinde birden fazla fonksiyonel T-bet bağlanma
bölgesi gösterilmiştir. ( Lee at Al 2004; Tong et al 2005). IFN-gama geni CNS 22 kb bölgesinde
T-bet bağlanma bölgesi vardır. Tbet eksik farelerde Th1 gelişmi ve IFN γ ekspresyonunda
defektler görülür.

16
STAT4
STAT-4 bir nükleer faktördür. IL-12/ STAT4 yolağı Th1 gelişimi ve IFN γ
ekspresyonunda önemli kritik regülatör özelliğe sahiptir. STAT-4 IFN γ geni promotor bölgeye
tutunur. STAT-4 diğer transkripsiyon faktörlerinin de IFN-γ geni promotör bölgesine
bağlanmasını kolaylaştırarak gen ekspresyonunu düzenler.
INF- γ GEN EKSPRESYONUN NFKB İLE DÜZENLENMESİ
NF-KB ilk olarak B lenfositlerince yapılan Kappa hafif zincirinin gen ekspresyonunun
regülatörü olarak tanımlanmış, fakat daha sonra bir çok farklı hücrede de bulunmuştur. Bir çok
farklı NF kB proteini bulunmasına rağmen, aktif olan heterodimer yapıda olanıdır. Yapısında iki
farklı protein bulunur. Bu proteinler P 65 ( rel A) ve P50 proteinleridir. İmmun uyarı yok
iken NF kB kompleksi sitoplazmada Ik Ba ve Ik B ß ya bağlı halde bulunur. Bu iki inhibitör
protein NF kB kompleksinin nükleus’a geçişini engeller. Hücre uyarıldığında, spesifik kinazlar
IkB proteinlerini fosfatlar. Hızlı olarak degrade olan pretozomlar NF Kb’ dan inhibitör
proteinleri ayırır, böylece serbestleşen NFkb nükleus’a transloke olur ve interferon gama gen
lokusundaki ilk intronun spesifik bölgesine bağlanarak inteferon gama ekspresyonunu başlatır.
NF kb özellikle + 874 T alleline bağlanır. Bu inteferon gama gen ekspresyonunun artışı ile
sonuçlanır. Bireyler arasındaki interferon gama yapabilme yeteneğinden dolayı otoimmun
hastalıklara yatkınlık açısından farklar oluşabilir
IFN- γ GENİ ÜZERİNDEKİ POLİMORFİZMLER
IFN-γ geni’nin ilk intronu CA dizilerinden oluşan mikrosatellik tekrarlamalar
içermektedir. Bu CA tekrarlamalarına göre 6 adet polimorfizm gösterilmiştir. Allel 2 olarak
adlandırılan 12 li CA tekrar dizilerinin varlığı gen ekspresyonunda artış ile yakından ilişkilidir.
(59). Ayrıca bu allel 2 tekrar dizileri ile tek nükleotid polimorfizmi arasında tam bir ilişki
mevcuttur. Bu tek nükleotid polimorfizmi 874 A/T polimorfizmidir. IFN γ’nın bu bölgesi gen
ekspresyonu için gerekli olan transkripsiyon faktörü NFkb’ nın bağlandığı bölgedir. Eğer T aleli
mevcutsa NFkb’nın bu bölgeye bağlanma afinitesi artar. Bu immun uyarı halinde interferon
gama ekspresyonun fazla yapımına neden olur (59-60).
Rossoum ve arkadaşları + 874 A-T polimorfizmi ile transkripsiyon faktörü olan NFkb
arasındaki ilişkiyi ve NF-kB’ nin spesifik gen bağlanma bölgesine bağlandığını göstermişlerdir
(61).

17
IFN-γ +874 A-T polimorfizmi ile romatoid artrit (69), akciğer transplantasyonundan
sonra gelişen akciğer fibrozisi (62), kemik iliği transplantasyonu yapılan hastalarda gelişen akut
greft versus host hastalığı arasında ilişki bulunmuştur ( 70 ).

18
3. GEREÇ VE YÖNTEM
Bu çalışmaya İstanbul Üniversitesi İstanbul Tıp Fakültesi, Kardiyoloji Anabilim Dalı
polikliniğince takip edilen ve ACC/AHA guidelines for the clinical application of
echocardiography kılavuzuna göre transtorasik ekokardiyografi ile RKKH tanısı konulmuş olan
152 hasta ile, ekokardiyografik incelemesi normal sınırlarda bulunmuş olan ve bilinen bir
immunolojik hastalığı olmayan 151 sağlıklı birey dahil edilmiştir.
3.1. Seçilen Örneklerin Tanımı
RKKH’ nın tanısı:
İlk RA hecmesini 15 yaşından önce geçirmiş olan ve “ACC/AHA guidelines for the
clinical application of echocardiography” klavuzuna uygun olarak yapılan transtorasik
ekokardiyografi ile RKKH tanısı konulmuş olan hastalar, semptomları ve ekokardiyografik
bulguları değerlendirilerek kalp kapak hasarı şiddetli veya hafif olmak üzere iki gruba ayrıldı.
RKKH ‘ nın Transtorasik Ekokardiyografik bulguları
Mitral kapaklarında; Fibrotik kalınlaşma
Komissürlerde füzyon
Diastolde ‘ doming ’ hareketi
Kordalarda kısalma, kalınlaşma, füzyon
Kapak hareket amplitüdleri ve açılımında azalma ve kısıtlanma
Kapaklarda kalsifik nodüller
Aort kapaklarında; Kalınlaşma ve komüssürlerde füzyon ve adezyonlar.
Kapakların serbest kenarlarında fibrotik kalınlaşma ve retraksiyonlar,
Kalsifik nodüller
Kapak hareketleri ve açılımında azalma
Kapak hasarının ciddiyetinin belirlenmesi, kapak replasmanı ve mitral balon
valvüloplasti endikasyonları, ACC/AHA 2008 guideline uptade on valvular heart disease
klavuzu esas alınarak değerlendirildi. Aynı kılavuz esas alınarak mitral balon valvüloplasti
yapılacak hastalarda uygun endikasyonlarda transözofageal ekokardiyografi de yapıldı.

19
Ciddi mitral darlığı: Ortalama mitral kapak gradiyentinin 10 mm Hg’nın üzerinde,
pulmoner arter basıncının 50 mm Hg’nın üzerinde, Pressure Half Time (PHT) veya planimetrik
yöntemle ölçülen kapak alanının 1 cm2 den dar olması
Ciddi mitral yetersizliği: Renkli Doppler inceleme ile 3(+) – 4(+) yetersizlik akımı
olması, 7 mm den büyük vena kontrakta ile beraber santral jet akımının sol atriyum alanının %
40 ından fazla olması ya da herhangi uzunlukta olan mitral yetersizliği jetinin sol atriyum
duvarına çarparak eksantrik, türbülan akım oluşturması, sol atriyumda ve sol ventrikülde
genişleme olması
Ciddi aort darlığı: Ortalama aort kapak gradientinin 40 mm hg den fazla olması, kapak
alanının 1 cm2 den dar olması
Ciddi aort yetersizliğİ: Renkli Doppler inceleme ile 3 (+) - 4 (+) aort yetersizliği
olması, santral yetersizlik jetinin sol ventrikül çıkış yolu alanının % 65 inden fazla olması, vena
kontraktanın 6 mm den fazla olması, sol ventrikülde genişleme olması kriterlerine göre
değerlendirildi.
Mitral balon valvüloplasti endikasyonları:
1. Semptomatik olan veya asemptomatik olup pulmoner arter basıncı istirahatte 50
mmHg’nın, egzersiz ile 60 mm Hg’nın üzerinde bulunan
2. Mitral kapak alanı 1.5 cm2’den dar
3. Mitral darlığına mitral yetersizliği eşlik ediyor ise orta derecenin altında mitral
yetersizliği olan
4. Transözofageal ekokardiyografi ile sol atriyum, sol atriyum apendiksi ve diğer kalp
boşlukarında trombüs görülmeyen
5. Wilkins skoru 8’in altında olan hastalar
Wilkins skoru hesaplanması;
• Kapak mobilitesinin derecesi
• Subvalvüler yapıların kalınlığı
• Kapağın kalınlığı
• Kalsifikasyon gibi parametreler, 1’den 4’e kadar puanlanarak, puanların toplamı
olarak değerlendirilir.

20
Şiddetli kapak hasarı olan birinci grupta; Semptomatik olup, en az bir kalp kapağında
cerrahi ya da invaziv girişim gerektirecek derecede hemodinamik bozukluğa yol açan kapak
darlığı ve / veya yetersizliği olan hastalar bulunmakta idi. Ciddi aort ve mitral kapak fonksiyon
bozukluğunun birlikte olması nedeniyle kapak replasmanı yapılan veya ciddi mitral darlığı
nedeniyle mitral balon valvüloplasti yapılan hastalar da bu gruba dahil edildi.
Hafif kapak hasarı olan ikinci grupta; Semptomu olmayan veya hafif semptomatik
olup transtorasik ekokardiyografi ile RKKH tanısı konulan ancak kapak lezyonunun cerrahi
veya invaziv yöntemlerle tedavi endikasyonu olmayan hastalar mevcuttu.
Proje kapsamında hasta ve/veya kontrol grubuna dahil edilen her birey için EDTA’lı
tüpe 3ml periferik venöz kan alındı. Kit manuelinde belirtilen şekilde DNA izole edildi (Roche
Diagnostic, GmbH, Mannheim, Almanya). İzole edilen örneklerin spektrofotometrik olarak
saflık tayinleri yapıldı ve çalışma zamanına kadar -20 C’de bekletildi. Elde edilen DNA
örneklerinden ARMS-PCR yapılarak IFN-gama genindeki hedef bölge çoğaltıldı. PCR
sonucunda her hasta için hazırlanmış olan iki ayrı tüp (A ve T allel) içindeki ürünler %2’lik
agaroz jelde yürütülerek UV altında görüntülendi ve bireylerin genotipleri belirlendi.
3.1.1. Kullanılan Yöntemler
Periferik Kandan DNA İzolasyonu
Genomik DNA izolasyonu için, alınan tam kan örneğinden kit manuelinde belirtilen şekilde
DNA izole edildi (Roche Diagnostic, GmbH, Mannheim, Almanya). İzolasyon sonrası DNA
miktar tayini 260 nm spektrofotometrik ölçümlerdeki absorbans oranlarına göre belirlendi.
DNA İzolasyonu
Ön hazırlık: Wash Buffer II (mavi kapaklı) e 40ml absolute etanol eklendi.
Wash Buffer I (siyah kapaklı) e 20ml absolute etanol eklendi.
1-Fankon tüpüne 200 µl (minimum) kan , 200 µl Binding buffer ve 40 µl Proteinaz K konularak
karıştırıldı ve 70C’de 10 dakika inkübe edildi .
2- İnkübasyon sonrasında isopropanol (100 µl) alkol eklenerek kısa bir vorteks uygulandı.
3- Collection tüplerinin içine High Pure Filter tüpleri (kolon) yerleştirildi.
4-Her bir örnek kolonlara pipetlendi. (kolonların kapasitesi maksimum 700 µl dir)

21
5-8000g de 1dk santrifüj edildi.
6-Collection tüplerinde biriken sıvı atıldı.
4- 500 µl (sabit miktar) inhibitör removal buffer eklendi.
5-8000g de 1dk santrifüj edildi.
6-Collection tüplerinde biriken sıvı atıldı.
7-500 µl (sabit miktar ) wash buffer eklendi Bu işlem 2 defa tekrarlandı.
8- 8000g de 1dk santrifüj edildi.
9-2dk maximum devirde santrifüj edildi.
10- Collection tüplerinde biriken sıvı atıldı.
11-Kolonlar 1,5ml lik steril, kapaklı, Dnase/RNase Free mikrosantrifüj tüplerine kondu.
12-50-100 µl Elution Buffer eklendi.
13-1dk 8000g de santrifüj edildi.
14-Tüpün içine süzülen elution buffer çekilip tekrar kolona yüklendi ( eğer kolonda DNA
kalmışsa 2 kere bu işlem yapılarak iyice süzülmesi sağlanmış olundu)
15 -1dk 8000g de santrifüj edildi.
16-Kolonlar atıldı.
Elde Edilen DNA’nın Konsantrasyon ve Kalitesinin Tayini
Elde edilen DNA örneklerinde konsantrasyon ve miktar tayini yapmadan önce her
örnekten 20µl alınarak üzeri 380µl 0.5X TE tamponu ile tamamlandı ve bu şekilde 1/20
dilüsyonu sağlandı. Bu dilüsyon örneklerinin daha sonra spektrofotometrede 260 nm ve 280 nm
dalga boylarında OD ölçümleri yapıldı.
Spektrofotometrede 260 nm ve 280 nm dalga boylarında yapılan ölçümlerle DNA’nın
saflığı ve konsantrasyonu belirlendi. 50µg/ml çift iplikçikli DNA içeriğinin 260 nm dalga
boyunda 1 optik densite (OD) verdiği kabul edilmektedir. 260 nm’deki ölçüm değeri aşağıdaki
formüle uygulanarak DNA konsantrasyonu hesaplandı.
DNA Konsantrasyonu: OD260 X 50µg/ml X Sulandırma oranı
DNA örneklerinin saflığı OD260/ OD280 oranı kullanılarak belirlendi. Yeterince iyi
saflıkta kabul edilen DNA’nın OD260/ OD280 değeri yaklaşık 1,8 idi. Ortamda fenol veya protein
mevcutsa bu oran 1,8’den küçük olacaktı. OD260/ OD280 değeri 2’den büyükse ortamda RNA
bulunduğu anlamına geliyor idi.

22
3.1.1.1. IFN- γ geni Amplifikasyon Ürünlerinde Gen Polimorfizminin Tespiti:
IFN- γ Geninin PZR Yöntemi İle Çoğaltılması
IFN- γ geninde +874 A/T polimorfizminin gözlendiği bölgeyi çoğaltmada kullanılan
primer dizileri: SSS
IFN- γ generic primer: 5’-TCAACAAAGCTGATACTCCA- 3’
IFN- γ T allel : 5’-TTCTTACAACACAAAATCAAATCT-3’
IFN- γ A allel : 5’-TTCTTACAACACAAAATCAAATCA-3’
Internal control1 : 5’-GCCTTCCCAACCATTCCCTTA-3’
Internal control2: 5’- TCACGGATTTCTGTTGTGTTTC-3’
DNA örneklerinden IFN- γ +874 gen bölgesinin çoğaltılması için toplam reaksiyon
hacmi 25 µl olacak şekilde, PZR karışımı 0,5 ml’lik steril tüpte her hasta için A ve T allelerine
ait iki tüp hazırlandı. 10 örnek çalışılacak ise pozitif ve negatif kontrol için birer tane ve
pipetleme hataları için de bir tane olmak üzere toplam 13 üzerinden reaksiyon karışımı
hazırlandı. PZR karışımının hazırlanma işlemlerinin hepsi buz üzerinde soğukta ve steril kabin
içerisinde yapıldı.
PZR karışımı :
ddH2O : 14,7 µL
Taq polimeraz tamponu 10x reaksiyon (NaSO4) çözeltisi : 1,5 µL
25mM MgCl2 : 1,5 µL
1mM dNTP (her bir bazdan) : 5,0 µL
Primer 1 (25µg/100µL) : 0,5µl
Primer 2 (25µg/100µL) : 0,5µl
Primer 3 (25µg/100µL) : 0,5µl
Primer 4 (25µg/100µL) : 0,5µl
Primer 5 (25µg/100µL) : 0,5µl
Taq polimeraz enzimi : 0,3 ünite
Taq polimeraz eklendikten sonra hiç vakit kaybetmeden tüp içine konan bileşenlerin
iyice karışması için pipetleme işlemi yapıldı. Örnek sayısı kadar 0,2 ml’lik tüplere 24 µl
reaksiyon karışımı dağıtıldı. Daha sonra her tüpe 1 µl DNA 500 ng eklenerek yeniden pipetleme

23
yapıldı ve daha önceden 96 ºC sıcaklığa çıkarıldı PZR cihazına (Termal Cycler) örnekler
yerleştirildi ve PZR işlemi başlatıldı. IFN-gama promotör gen bölgesi için kullanılan
amplifikasyon koşulu tablo 3-1’ de gösterilmiştir.
Tablo 3-1: IFN- γ +874 A/T polimorfizminin gözlendiği bölgenin PZR reaksiyonu için termal profil şartları
95 ºC 5 dakika
95 ºC
55 ºC
72ºC
45 saniye
45 saniye 35 döngü
45 saniye
72ºC 5 dakika
IFN- γ PZR Sonuçlarının Agaroz Jel Elektroforezi ile Görüntülenmesi
PZR sonrası oluşan amplifikasyon ürünlerinin incelenmesi için %2’lik agaroz jel
hazırlandı. Her PZR tüpünden elde edilen 10 µL amlikon örneği, agaroz jel elektroforezi için
hazırlanmış, %2’lik agaroz jele tatbik edilerek 120 voltluk elektrik akımında yürütüldü ve
polaroid kamera ile UV ışık altında jelin fotoğrafı çekilip PZR’nin başarısı tespit edildi.
Şekil 3-1: IFN-γ ARMS PZR ürünlerinin %2’lik agaroz jeldeki görüntüsü.
Üst kısımda bulunan şekil A alleli alt kısımda bulunan şekilde T alleli göstermektedir.Her ikisi içinde kontrol
bölgeleride çoğaltılmıştır.(1.hasta AT genotipi, 2.hasta AA genotipi, 3.hasta TT genotipi, 4.hasta AT genotipi)
Kontrol bantı A allel
T allel

24
İSTATİSTİKSEL ANALİZ
Hasta ve kontrol grubunda cinsiyet, genotip, kapak hasarın derecesi, valvüloplasti
öyküsü, kapak replasmanı öyküsü, penisilin proflaksisi kullanım öyküsü gibi numerik olmayan
veriler ki -kare testi kullanılarak değerlendirildi. P değerinin 0.05’ten küçük olduğu durumlar
istatistiksel olarak anlamlı kabul edildi.

25
4. BULGULAR
Şiddetli kapak hasarı olan grupta: 97 hasta mevcuttu. Bu hastaların 54 ü daha önceden
valvüloplasti öyküsü olan, 52 hasta kapak replasmanı yapılmış olan hastalardı 9 hastada
valvüloplasti sonrası kapak replasmanı yapılmıştı. Hafif kapak hasarı olan grupta 55 hasta
mevcuttu.
Hasta ve kontrol grupları arasında IFN γ 874 T-A genotip dağılımında istatistiksel fark
bulundu. (Şekil 4-2 P= 0.002) T allelini taşımanın , taşımayanlara göre RKKH yatkınlık
açısından 2.4 kat risk oluşturduğu görüldü (şekil 4-3).
Hasta grubunda kapak hasarının ciddiyeti ile IFN γ 874 T-A genotip dağılımı
incelendiğinde, istatistiksel fark bulundu (şekil 4-5 P=0.001 ). Hasta grubunda T allelini
taşımanın, taşımayanlara göre kapaklardaki hasarın ciddiyeti arttırdığı bulundu. Bu artış T alleli
için 2.08, eğer hasta T homozigot ise 3.66 kat olarak bulundu (Şekil 4-6, 4-7).
RKKH’ında şiddetli kapak tutulumunun bir göstergesi olan valvüloplasti öyküsü ile
IFN γ 874 T-A genotip dağılımı arasında istatistiksel anlamlı fark bulundu (şekil 4-10 P=0.034)
T homozigot bireylerde diğer genotiplere sahip bireylere göre valvüloplasti öyküsü istatistiksel
olarak anlamlı derecede fazla bulundu (şekil 4-11 P= 0.012).
RKKH’ında kapak hasarının şiddetli bir göstergesi olan kapak replasmanı ile IFN γ 874 T-
A genotip dağılımı arasında istatistiksel anlamlı fark bulundu (şekil 4-12 P=0.01 ) A homozigot
bireylerde diğer genotip’lere sahip bireylerle karşılaştırıldığında kapak replasmanı anlamlı
derecede daha az sayıda bulundu ( Şekil 4-13 P= 0.015).
Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda darlığın ciddiyeti ile
IFN γ 874 T-A genotip dağılımı arasında istatistiksel anlamlı fark bulundu (Şekil 4-15 P=0.002)
Bu hastalardan T homozigotlar diğer genotip’lere sahip hastalarla kıyaslandığında darlıkla
seyreden kapakların darlık derecesinin şiddeti anlamlı derecede daha fazla bulundu (Şekil 4-16
P= 0.012).
Hasta grubunda düzenli penisilin profilaksisi kullanımı ile kapak hasarının şiddeti
arasında anlamlı istatistiksel fark bulunmadı.( Şekil 4-18 P= 0. 613 )
26
125 124
27 27
49± 13 49± 13
0
20
40
60
80
100
120
140
Kadın Erkek Yaş
Hasta ve kontrol grubu arasında yaş ve cinsiyet
dağılımı
Hasta
Kontrol
Şekil 4-1: Hasta ve kontrol grubu arasında yaş ve cinsiyet dağılımı
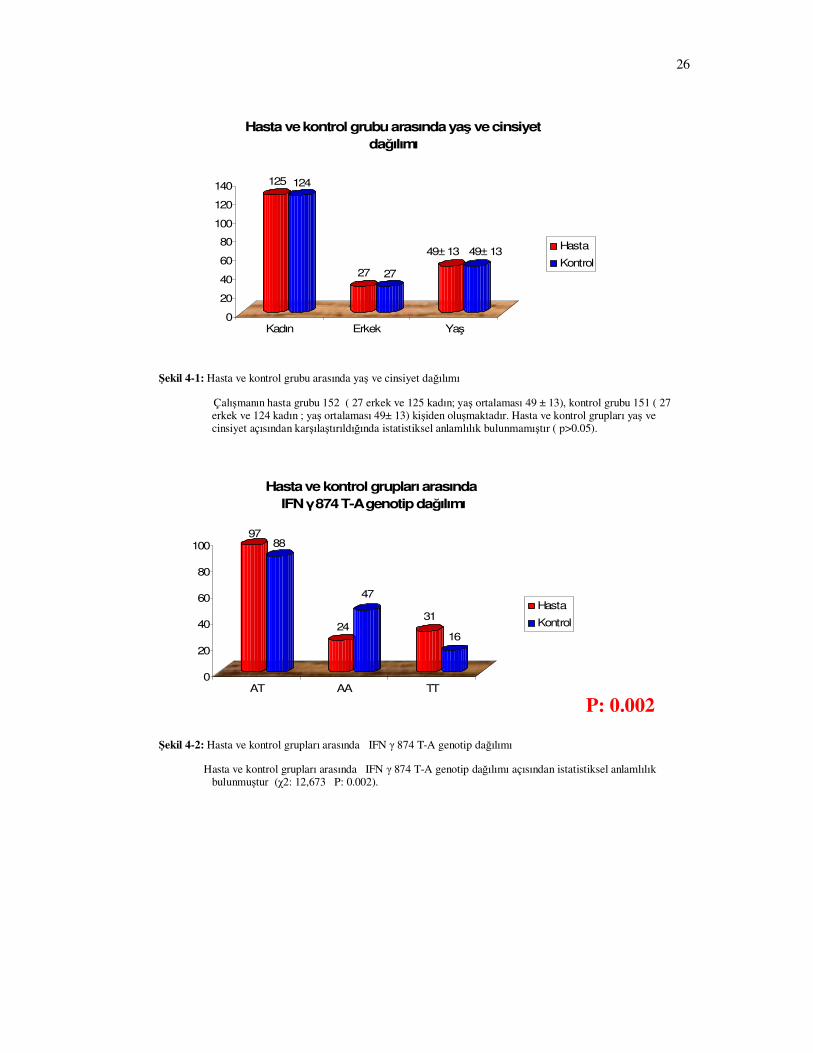
Çalışmanın hasta grubu 152 ( 27 erkek ve 125 kadın; yaş ortalaması 49 ± 13), kontrol grubu 151 ( 27 erkek ve 124 kadın ; yaş ortalaması 49± 13) kişiden oluşmaktadır. Hasta ve kontrol grupları yaş ve cinsiyet açısından karşılaştırıldığında istatistiksel anlamlılık bulunmamıştır ( p>0.05).
9788
24
47
31
16
0
20
40
60
80
100
AT AA TT
Hasta ve kontrol grupları arasında
IFN γ 874 T-A genotip dağılımı
Hasta
Kontrol
P: 0.002
Şekil 4-2: Hasta ve kontrol grupları arasında IFN γ 874 T-A genotip dağılımı
Hasta ve kontrol grupları arasında IFN γ 874 T-A genotip dağılımı açısından istatistiksel anlamlılık bulunmuştur (χ2: 12,673 P: 0.002).

27
121135
3116
0
20
40
60
80
100
120
140
AT/AA TT
Hasta ve kontrol grubu arasında
T homozigot genotip dağılımı
Hasta
Kontrol
P:0.018
Şekil 4-3: Hasta ve kontrol grubu arasında T homozigot genotip dağılımı
T allelini homozigot taşıma açısından hasta ve kontrol grupları karşılaştırıldığında T homozigot bireyler hasta grubunda kontrol grubuna göre istatistiksel olarak daha fazla bulunmuşur (χ2: 5.550 P: 0.018, OR:2.16 (% 95 güven aralığında 1.12-4.14).
128
104
24
47
0
20
40
60
80
100
120
140
T allel(AT-TT) AA
Hasta ve kontrol grubu arasında
T allel dağılımı
Hasta
Kontrol
P: 0.002
Şekil 4-4: Hasta ve kontrol grubu arasında T allel dağılımı
T allelli taşıyanlarda taşımayanlara göre RKKH görülme riski 2.4 kat daha fazladır (χ2: 9.930 P: 0.002, OR:2.40 (% 95 güven aralığında 1.38- 4.20).

28
63
34
816
26
5
0
10
20
30
40
50
60
70
AT AA TT
Hasta grubunda genotipe göre
kapak hasarında ciddiyet dağılımı
Kapak hasarı şiddetli
Kapak hasarı hafif
P:0.001
Şekil 4-5: Hasta grubunda genotipe göre kapak hasarında ciddiyet dağılımı
Hasta grubunda; kapak hasarının derecesine göre genotip dağılımında istatistiksel fark vardır (χ2: 15,111 P: 0.001). T homozigot hastalarda kapak hasarı daha ciddi olma eğilimindedir.
71
50
26
5
0
10
20
30
40
50
60
70
80
AT/AA TT
Hasta grubunda T homozigot genotip ve
kapak hasarında ciddiyet dağılımı
Kapak hasarı şiddetli
Kapak hasarı hafif
P:0.009
Şekil 4-6:Hasta grubunda T homozigot genotip ve kapak hasarında ciddiyet dağılımı
T allelini homozigot taşıyan bireyler AA ve AT genotiplere sahip bireylerle karşılaştırıldığında kapak hasarının şiddeti T homozigot bireylerde, diğerlerine göre istatistiksel olarak anlamlı derecede fazla bulunmuştur (χ2: 6,783 P: 0.009). OR: 3.66 ( % 95 güven aralığında 1.31-10.18).
29
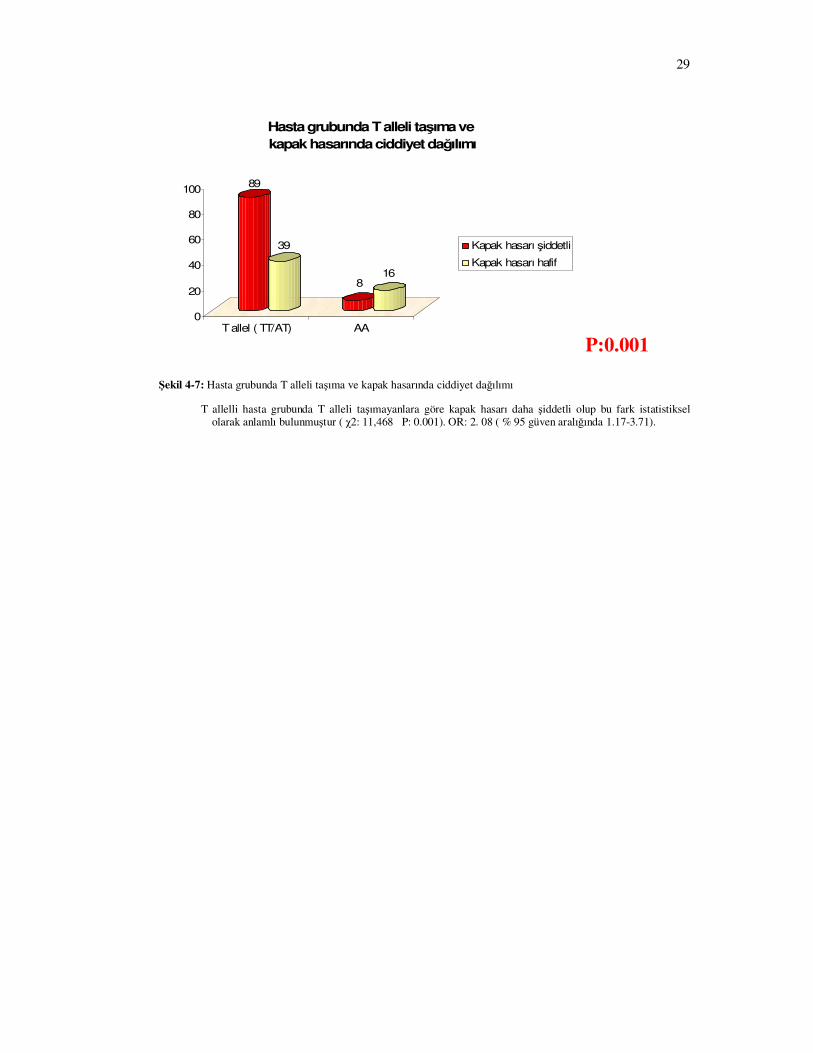
89
39
816
0
20
40
60
80
100
T allel ( TT/AT) AA
Hasta grubunda T alleli taşıma ve
kapak hasarında ciddiyet dağılımı
Kapak hasarı şiddetli
Kapak hasarı hafif
P:0.001
Şekil 4-7: Hasta grubunda T alleli taşıma ve kapak hasarında ciddiyet dağılımı
T allelli hasta grubunda T alleli taşımayanlara göre kapak hasarı daha şiddetli olup bu fark istatistiksel olarak anlamlı bulunmuştur ( χ2: 11,468 P: 0.001). OR: 2. 08 ( % 95 güven aralığında 1.17-3.71).
30
60
37
17
7
27
4
0
10
20
30
40
50
60
AT AA TT
Hasta grubunda IFN γ 874 T-A genotip ve
romatizmal kapak tutulumunun darlık veya
yetersizlik ağırlıklı olmasına göre dağılımı
Darlık
Yetersizlik
P: 0.03
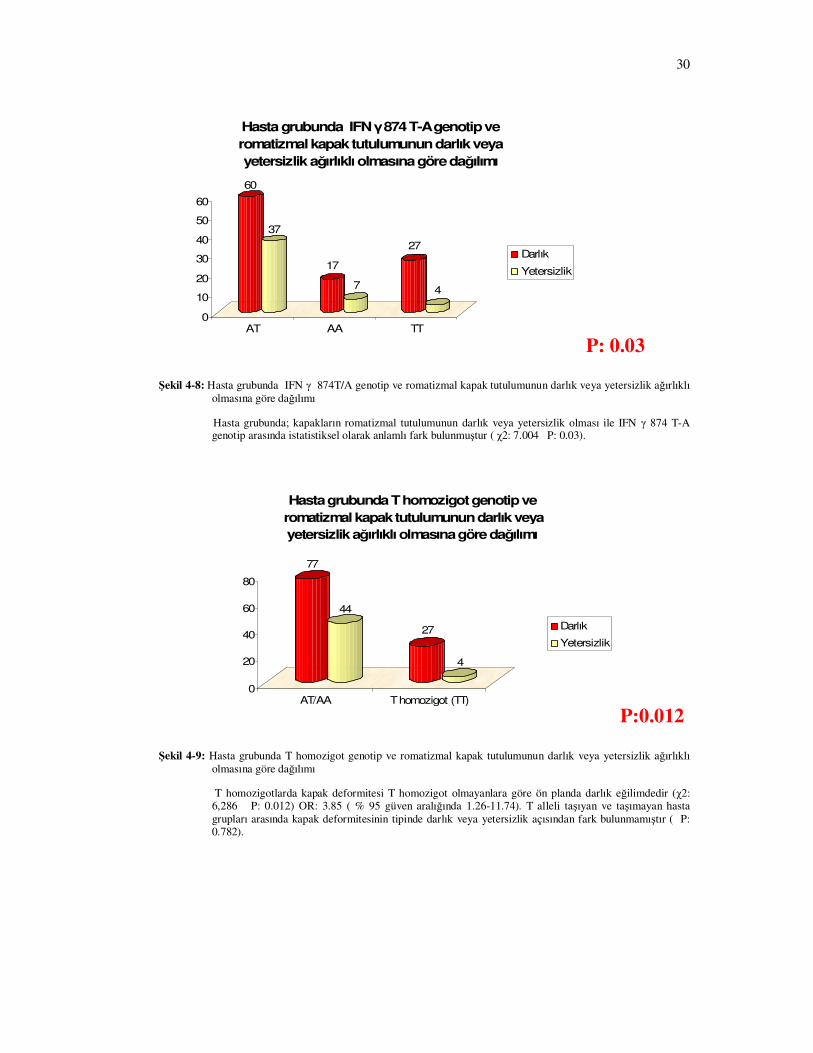
Şekil 4-8: Hasta grubunda IFN γ 874T/A genotip ve romatizmal kapak tutulumunun darlık veya yetersizlik ağırlıklı olmasına göre dağılımı
Hasta grubunda; kapakların romatizmal tutulumunun darlık veya yetersizlik olması ile IFN γ 874 T-A genotip arasında istatistiksel olarak anlamlı fark bulunmuştur ( χ2: 7.004 P: 0.03).
77
44
27
4
0
20
40
60
80
AT/AA T homozigot (TT)
Hasta grubunda T homozigot genotip ve
romatizmal kapak tutulumunun darlık veya
yetersizlik ağırlıklı olmasına göre dağılımı
Darlık
Yetersizlik
P:0.012
Şekil 4-9: Hasta grubunda T homozigot genotip ve romatizmal kapak tutulumunun darlık veya yetersizlik ağırlıklı olmasına göre dağılımı
T homozigotlarda kapak deformitesi T homozigot olmayanlara göre ön planda darlık eğilimdedir (χ2: 6,286 P: 0.012) OR: 3.85 ( % 95 güven aralığında 1.26-11.74). T alleli taşıyan ve taşımayan hasta grupları arasında kapak deformitesinin tipinde darlık veya yetersizlik açısından fark bulunmamıştır ( P: 0.782).
31
31
66
6
18 1714
0
10
20
30
40
50
60
70
AT AA TT
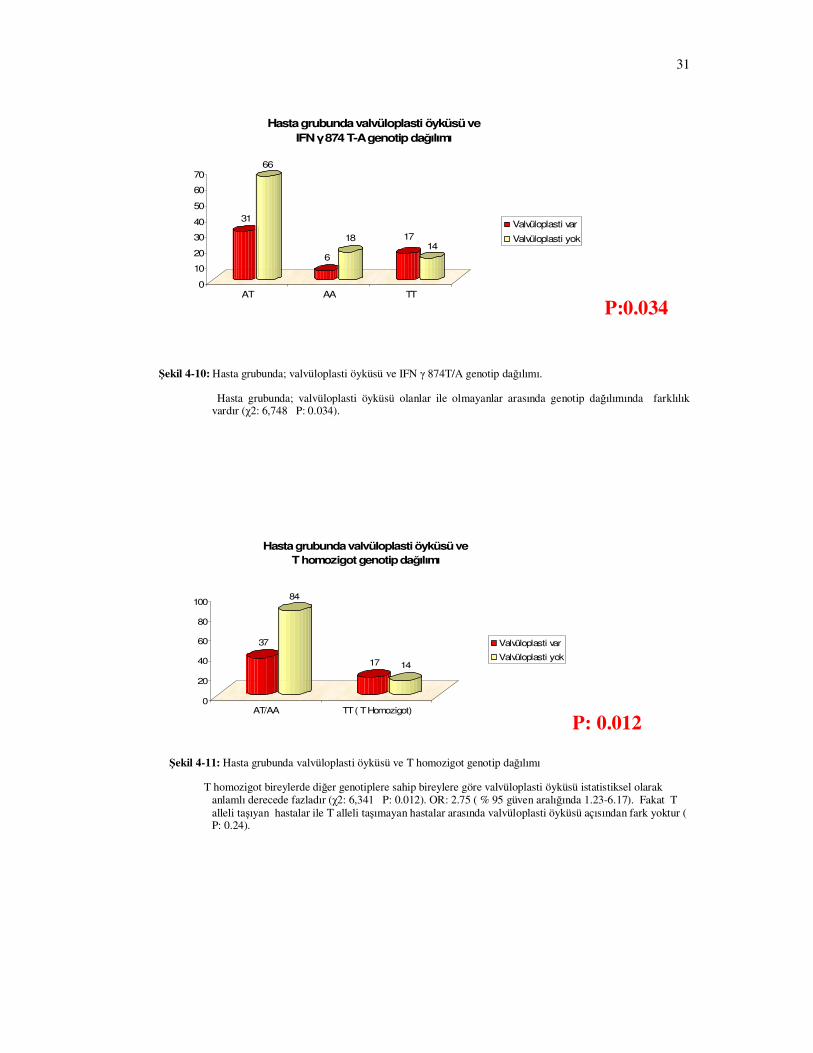
Hasta grubunda valvüloplasti öyküsü ve
IFN γ 874 T-A genotip dağılımı
Valvüloplasti var
Valvüloplasti yok
P:0.034
Şekil 4-10: Hasta grubunda; valvüloplasti öyküsü ve IFN γ 874T/A genotip dağılımı.
Hasta grubunda; valvüloplasti öyküsü olanlar ile olmayanlar arasında genotip dağılımında farklılık vardır (χ2: 6,748 P: 0.034).
37
84
17 14
0
20
40
60
80
100
AT/AA TT ( T Homozigot)
Hasta grubunda valvüloplasti öyküsü ve
T homozigot genotip dağılımı
Valvüloplasti var
Valvüloplasti yok
P: 0.012
Şekil 4-11: Hasta grubunda valvüloplasti öyküsü ve T homozigot genotip dağılımı
T homozigot bireylerde diğer genotiplere sahip bireylere göre valvüloplasti öyküsü istatistiksel olarak anlamlı derecede fazladır (χ2: 6,341 P: 0.012). OR: 2.75 ( % 95 güven aralığında 1.23-6.17). Fakat T alleli taşıyan hastalar ile T alleli taşımayan hastalar arasında valvüloplasti öyküsü açısından fark yoktur ( P: 0.24).

32
33
64
3
2116 15
0
10
20
30
40
50
60
70
AT AA TT
Hasta grubunda kapak replasmanı öyküsü ile
IFN γ 874 T-A genotip dağılımı
Kapak replasmanı var
Kapak replasmanı yok
P: 0.01
Şekil 4-12: Hasta grubunda; kapak replasmanı öyküsü ile IFN γ 874T/A genotip dağılımı.
Hasta grubunda; kapak replasmanı yapılmış olanlar ile kapak replasmanı yapılmamış olanlar arasında genotip dağılımında anlamlı fark bulunmuştur ( χ2: 9,199 P: 0.01).
49
79
3
21
0
10
20
3040
50
60
70
80
TT/AT AA (A Homozigot)
Hasta grubunda kapak replasmanı öyküsü ile
A homozigot genotip dağılımı
Kapak replasmanı var
Kapak replasmanı yok
P: 0.01
Şekil 4-13: Hasta grubunda kapak replasmanı öyküsü ile A homozigot genotip dağılımı
A homozigot bireylerde diğer genotip’lere sahip bireylerle karşılaştırıldığında kapak replasmanı anlamlı derecede daha az sayıda yapılmıştır (χ2: 5,969 P: 0.015). OR: 3.06 ( % 95 güven aralığında 1.03-9.02).
36
85
16 15
0
20
40
60
80
100
AA/AT TT
Hasta grubunda kapak replasmanı öyküsü ile
T homozigot genotip dağılımı
Kapak replasmanı var
Kapak replasmanı yok
P:0.022
Şekil 4-144: Hasta grubunda kapak replasmanı öyküsü ile T homozigot genotip dağılımı
T homozigot bireyler diğer genotip’lere sahip bireylerle karşılaştırıldığında kapak replasmanı anlamlı derecede daha fazla sayıda hastada yapılmıştır (χ2: 5,240 P: 0.022). OR: 2.51 ( % 95 güven aralığında 1.12-5.63).
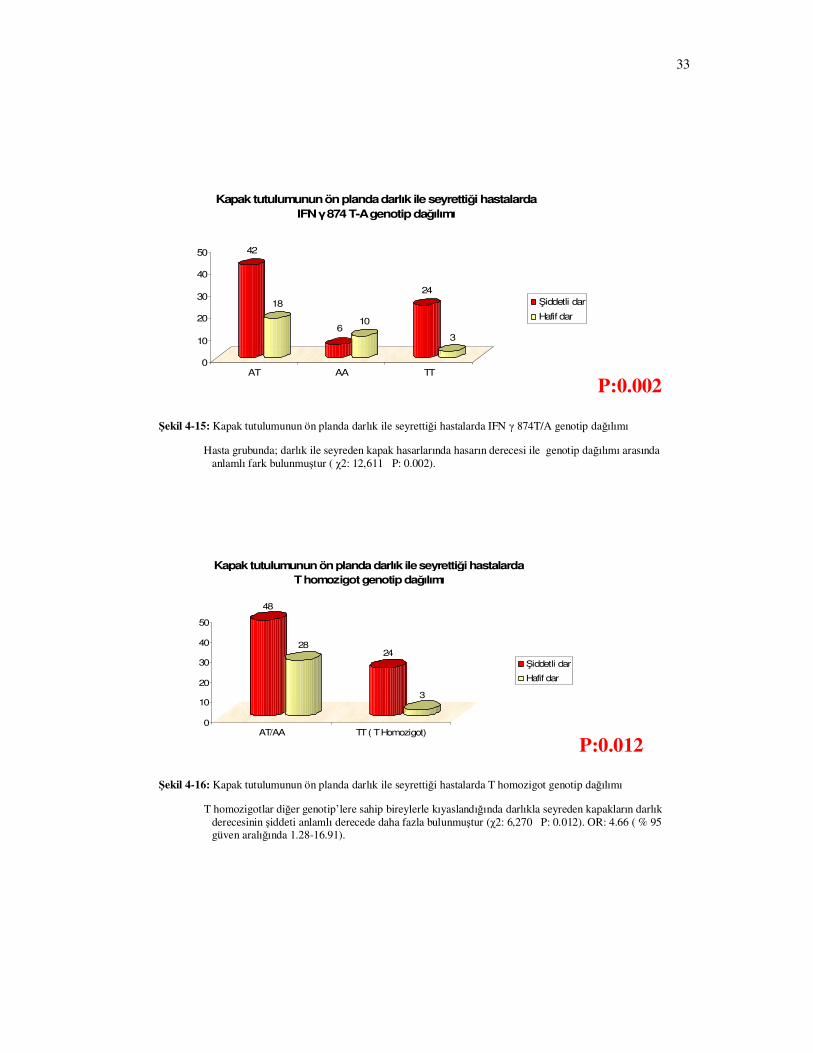
33
42
18
610
24
3
0
10
20
30
40
50
AT AA TT
Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda
IFN γ 874 T-A genotip dağılımı
Şiddetli dar
Hafif dar
P:0.002
Şekil 4-15: Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda IFN γ 874T/A genotip dağılımı
Hasta grubunda; darlık ile seyreden kapak hasarlarında hasarın derecesi ile genotip dağılımı arasında anlamlı fark bulunmuştur ( χ2: 12,611 P: 0.002).
48
2824
3
0
10
20
30
40
50
AT/AA TT ( T Homozigot)
Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda
T homozigot genotip dağılımı
Şiddetli dar
Hafif dar
P:0.012
Şekil 4-16: Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda T homozigot genotip dağılımı
T homozigotlar diğer genotip’lere sahip bireylerle kıyaslandığında darlıkla seyreden kapakların darlık derecesinin şiddeti anlamlı derecede daha fazla bulunmuştur (χ2: 6,270 P: 0.012). OR: 4.66 ( % 95 güven aralığında 1.28-16.91).
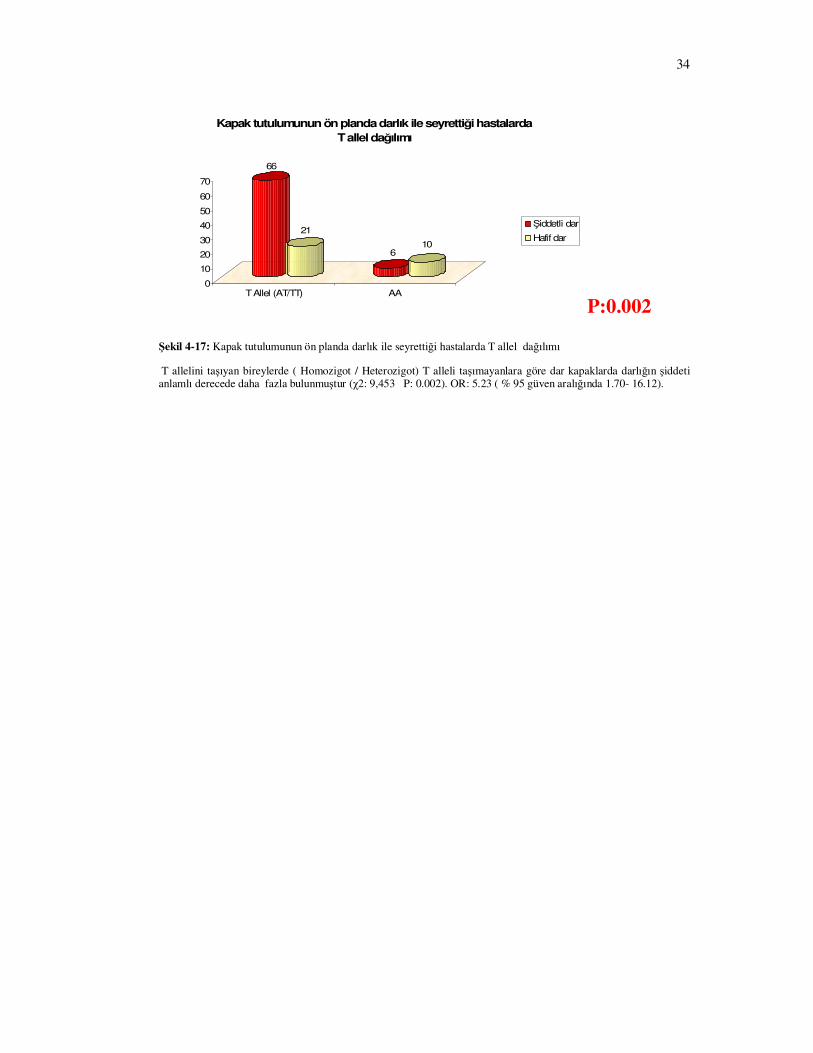
34
66
21
610
0
10
20
30
40
50
60
70
T Allel (AT/TT) AA
Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda
T allel dağılımı
Şiddetli dar
Hafif dar
P:0.002
Şekil 4-17: Kapak tutulumunun ön planda darlık ile seyrettiği hastalarda T allel dağılımı
T allelini taşıyan bireylerde ( Homozigot / Heterozigot) T alleli taşımayanlara göre dar kapaklarda darlığın şiddeti anlamlı derecede daha fazla bulunmuştur (χ2: 9,453 P: 0.002). OR: 5.23 ( % 95 güven aralığında 1.70- 16.12).
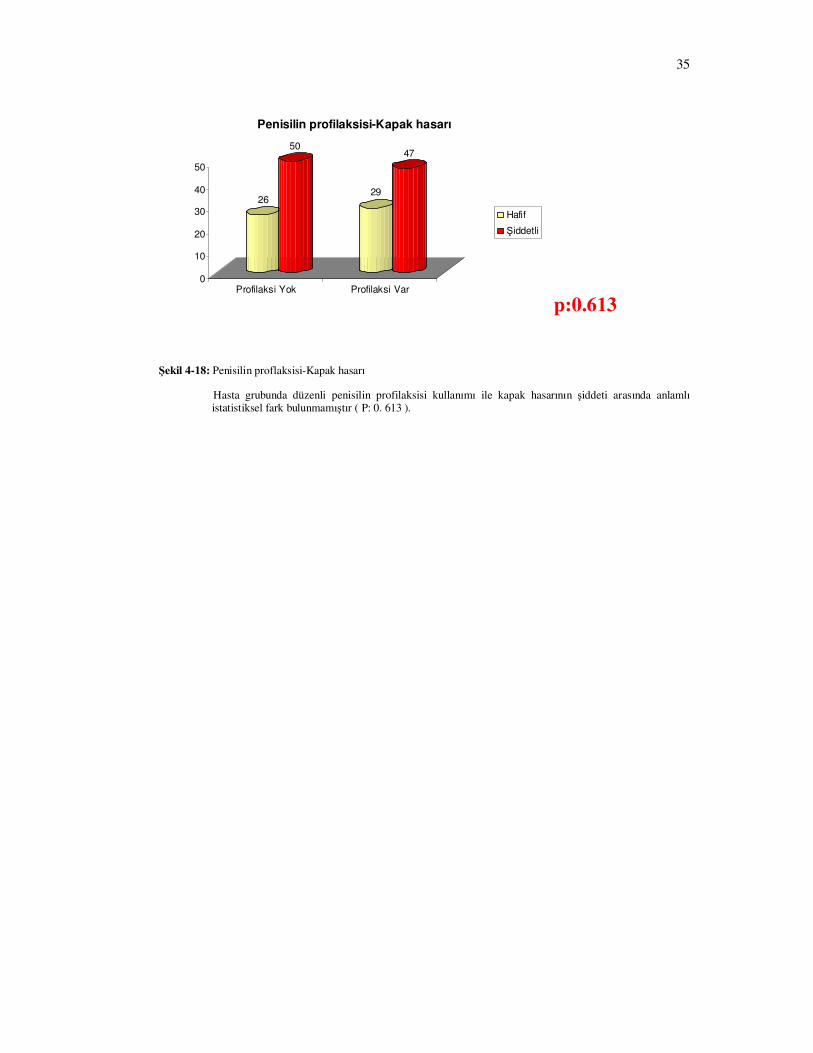
35
26
50
29
47
0
10
20
30
40
50
Profilaksi Yok Profilaksi Var
Penisilin profilaksisi-Kapak hasarı
Hafif
Şiddetli
p:0.613
Şekil 4-18: Penisilin proflaksisi-Kapak hasarı
Hasta grubunda düzenli penisilin profilaksisi kullanımı ile kapak hasarının şiddeti arasında anlamlı istatistiksel fark bulunmamıştır ( P: 0. 613 ).

36
5. TARTIŞMA
Romatizmal ateş (RA) , akut streptokoksik farenjitli hastaların yalnız % 3 ‘ünde
görülür. Bu hastaların yarısında ilk atak sırasında kardit görülebilmesine rağmen tekrarlayan
ataklar sırasında bu risk artmaktadır (63, 64,65). RA, değişik coğrafik bölgeler ve farklı etnik
gruplar incelendiğinde toplumlarda farklı sıklıkta görülmektedir. Ülkelerin gelişmişlik durumu
ile hastalığın görülme sıklığı arasında ters orantı mevcuttur. Günümüzde gelişmiş toplumlarda
RA görülme sıklığı önemli ölçüde azalmıştır (1).
Akut romatizmal ateş ve Romatizmal kalp kapak hastalığının (RKKH) patogenezi
kompleks olup, hem çevresel hem de genetik faktörler etyolojide rol oynamaktadır. Hastalığın
fizyopatolojisi net olarak anlaşılamamasına rağmen çapraz reaksiyon sonucu gelişen otoimmun
hastalık üzerinde durulmaktadır.
A grubu streptokoklar M proteinlerine göre 100 den daha fazla subtipe ayrılabilirler ve
bunlardan bazıları romatojen M proteinine sahiptir (örneğin M3-5). Streptokoksik toksinlerin
yanı sıra süperantijen özellik gösteren M proteini, B ve T hücreleri ile ilişkili otoimmun
aktiviteyi, oluşan otoantikorlar kalp dokularında ,özellikle de kapaklarda inflamatuvar süreci
başlatır. İnflamatuar süreçte sadece humoral immun sistem elemanları rol oynamamaktadır.
Etkilenen organların histolojik incelenmesinden elde edilen verilere göre lezyon etrafında; T
hücre kolonileri, makrofajlar, monositler görülmüştür. Bu hücreler lezyonun oluşumunda ve
progresyonunda ürettikleri sitokinler ile önemli bir role sahiptirler (66).
Genetik olarak hassas bireylerde hedef dokular ile çapraz reaksiyona giren
streptokoksik antijenlere karşı aşırı immun cevap( humoral ve hücresel) oluşması durumunu
destekleyen veriler mevcuttur (66).
Romatizmal kalp kapak hastalığı (RKKH) romatojen A grubu streptokokus Pyogenes
enfeksiyonu ile tetiğin çekildiği otoimmun reaksiyonlar sonucu kalp kapaklarında oluşan
geçikmiş sekel olarak değerlendirilmektedir ve bu hastalığa genetik yatkınlıkla ilgili pek çok
çalışma yapılmıştır (10,11,12,13).
Romatizmal kalp hastalığı ile HLA sınıf II antijenleri arasındaki ilişkiyi ilk olarak Ayoub
ve arkadaşları göstermiştir (29). Bu çalımada Afrika kökenli Amerikalılarda HLA-DR2, Kafkas
kökenli Amerikalılarda HLA-DR4 ilişkisi mevcuttur.
Farklı ırklarda yapılan çalışmalarda HLA sınıf II allelleri ile RKKH na yatkınlık arasında
2.3 kattan 13.6 kat a kadar ulaşan kuvvetli bir ilişki mevcuttur. Bu ilişki HLA sınıf II
alellerinin romatizmal kalp hastalığının gelişimindeki önemli rolünü göstermektedir.

37
RKKH’nın patogenezindeki antijen sunumunda HLA sınıf II antijenlerinin önemli rolünün
olmasından dolayı yapılan çalışmalardaki bu korelasyon şaşırtıcı değildir. Bu sonuçlar
RKKH’daki etnik dağılımı açıklamaya katkıda bulunmuştur.
Sitokinler; inflamasyonda rol oynayan önemli proteinlerdir. RKKH ile pek çok sitokin
ilişkisi üzerine çalışmalar yapılmıştır. TNF-α üzerinde en fazla çalışılan sitokindir ve onu
kodlayan gen dizisindeki bazı polimorfik durumlar bu sitokinin inflamasyonda fazla eksprese
edilmesine neden olmaktadır. RKKH’da kapakların histokimyasal incelenmesinde artmış TNF-
α ve INF-γ miktarı dikkati çekmektedir. TNF-α 308 A alleli ile artmış TNF-α yapımı arasında
ilişki gösterilmiştir. TNF-α ile Meksikalı, Brezilyalı ve Türk ırklarında RKKH ‘ya yatkınlık
ilişkisi de yapılan çalışmalarda ortaya konulmuştur.
INF-γ; proinflamatuvar bir sitokindir. Kronik inflamatuvar süreçte
immunmodülasyonun primer sitokinidir. T hücrelerine NK hücreleri tarafından sekrete edilir.
Parakrin ve otokrin etki ile inflamasyonun yönünü tayin eder (TH1 veya TH2). Romatizmal
kalp kapak hastalarında kapaklardan elde edilen T hücrelerinin immun uyarıya artmış INF-γ
sekresyonu yaptığı bildirilmiştir. INF-γ 12 . kromozomun q kolunda 4 ekzonlu bir gen
tarafından kodlanır. 146 amino asitten oluşur. Bu gen dizisinde bilinen iki adet polimorfizm
vardır. 874 T/A polimorfizmi genin 1. intronik parçasında olup gen ekspresyonunda NF kb
bağlanmasını etkileyerek gen ekspresyonunun düzenlenmesine etki etmektedir. İmmun uyarı
halinde T alleline sahip bireylerde INF-γ ekspresyonu A alleline sahip bireylere göre daha
fazladır (59, 60).
Kalp lezyonlarında, Aschoff nodülleri patognomik bir bulgu olarak önemlidir. Bu
nodüller, miyokardiyal intertisyumun temel olarak endokardiyumunda , subendokardiyumunda
ve perivasküler bölgede yer alan granülomatöz lezyonlardır. Bu nodüller hastalığın seyri
süresince farklı evrede bulunabilirler. Fraser ve arkadaşlarının yaptığı bir çalışmada IL1, TNF-
a,IL2 ve IFN- γ’ nın akut romatizmal ateşli hastalarda kapak lezyonlarında Aschoff nodül
progresyonu ve evresiyle olan ilişkisini göstermiştir. Buna göre 1 ve 2. evre de monosit ve
makrofajlardan , IL1 ve TNF α’ a sekrete edilir. 3.evrede T lenfositlerden IL2 ve IFN δ sekrete
edilir. Bu sonuçlar romatizmal ateşte kalp lezyonlarında proinflamatuar sitokinlerin rol
oynadığını göstermektedir (67). Romatizmal kalp kapak hastalarının kalp lezyonları, kalp ve
streptokok M proteinlerini tanıyan T hücre topluluklarını içerir. Yapılan in situ çalışmalarda bu
T hücrelerinin IFN δ, TNF α, IL4, IL 10 gibi bir çok sitokin ihtiva ettiği gösterilmiştir. Ayrıca
ex vivo çalışmalarda bu hücre toplulukları streptokoksik M proteinine maruz bırakıldığında yine
IFN δ, TNF α, IL4, IL 10 gibi bir çok sitokini ürettiği gösterilmiştir (67).
Sallakcı ve arkadaşlarının Türk popülasyonunda yapmış olduğu bir çalışmada INF-γ 874
T-A polimorfizmi ile akciğer tüberkülozu ve INF-γ cevabı arasındaki ilişki gösterilmiş olup

38
INF-γ üretiminde azalma ile sonuçlanan AA genotipli bireylerde tüberküloz hastalığının
progresyonu kötü seyrettiği rapor edilmiştir (68).
Awad ve arkadaşlarının yapmış olduğu bir çalışmada akciğer transplantasyonundan sonra
allograft fibrozis ile INF-γ 874 T-A polimorfizmi arasındaki ilişki gösterlimiştir. T alleline sahip
hastalarda allogreft fibrozisinde artış tespit edilmiştir (62).
Khani-Hanjani ve arkadaşlarının yapmış olduğu bir çalışmada otoimmun hastalık olan
Romatoid Artrit ile INF-γ 874 T-A polimorfizmi arasında ilişki gösterilmiştir (69).
Cavet ve arkadaşlarının kemik iliği transplantasyonu yapılmış olan hastalarda yapmış
oldukları bir çalışmada akut greft reddi ile INF-γ 874 T-A polimorfizmi arasında ilişki
gösterilmiştir (70).
RKKH nın patogenezinde fibrozis ile seyreden otoimmun inflamasyon mevcuttur. INF-γ
874 T-A polimorfiziminde T alleli bulunan bireylerde inflamasyonun abartılı seyretmesi nedenli
fibrotik mekanizmaların hızlı ve aşırı olması, kapakların erken ve şiddetli deformitesine zemin
hazırlıyor olabilir.
Bu çalışmada, RKKH grubunda INF-γ miktarını etkileyen INF-γ 874 T-A
polimorfizminin kapak hasarının şiddeti üzerine etkili olabileceği düşünülerek, RKKH
grubunda INF-γ 874 T-A polimorfizmi çalışılmıştır. Elde ettiğimiz bulgularda; T homozigot
bireylerin RKKH na yatkın olduğu görülmüştür. Hasta grupları incelendiğinde T allelini
taşımanın RKKH nda kapak lezyonlarının şiddetini arttırıcı yönde etkide olduğu görülmüştür.
Bu etki, hastaların T homozigot olması durumunda daha da belirginleşmektedir. Ayrıca T
allelini taşımanın kapakların erken deformitesi ve bu deformitenin darlık şeklinde olması ,
replasman veya balon valvüloplasti gerektirecek kritik düzeylere ulaşması açısından risk
oluşturduğu görülmüştür. Bir başka açıdan bakmak gerekise A alleli taşımak RKKH’na karşı
hem kapak tutulumu açısından hem de kapak hasarının şiddeti açısından koruyucu etkiye
sahiptir denilebilir.
Çalışmamızda ki-kare analizi ile yapılan istatistiksel incelemede INF-γ 874 T/A
polimorfizmine ait genotip dağılımlarının hasta ve kontrol grupları arasında istatistiksel olarak
anlamlı derecede farklı olduğu belirlenmiştir. T alleli taşıyan ya da T homozigot bireylerin
RKKH ‘ya genetik olarak daha yatkın olduğu sonucu çıkarılabilir. Başka bir açıdan bakmak
gerekirse A alleli ya da AA genotipi’ne sahip bireylerin RKKH’ya karşı koruyucu bir etkiye
sahip oldukları söylenebilir
Hasta grubu kapak hasarına göre incelendiğinde; INF-γ 874 T alleli yada T homozigot
genotip’e sahip bireylerde kapak hasarı diğer genotiplere göre daha şiddetli bulunmuştur. ( T
alleli için P= 0.001, T homozigot için P= 0.009)

39
Türk ırkında yapılmış olan çalışmada T alleli ile artmış INF-γ miktarı
ilişkilendirilmiştir. Bu ilişki farklı ırklarda da gösterilmiştir. RKKH’ da T alleli ya da T
homozigot genotip’e sahip bireylerde kapaklarda hasarın fazla olması kapak düzeyinde
inflamasyona yön veren ve inflamasyonun şiddetini belirleyen INF-γ düzeyi yüksekliğinin,
genetik yatkınlıktan kaynaklandığını düşündürmüştür. Bu yönü ile çalışmamızın sonuçları
şaşırtıcı değildir. Çalışmamızda kapak tutulumu ciddiyeti ile T aleline sahip bireyler
incelendiğinde , T alleli sahip bireylerde ciddi kapak hasarı T alleline sahip olmayanlara göre
2.08 kat daha fazla bulunmuştur. Ayrıca T homozigot bireylerde bu artış 3.66 kat olarak
bulunmuştur.
Kapak hasarının şiddetinin bir göstergesi olan valvüloplasti öyküsünün genotip ile
ilişkisi değerlendirildiğinde valvüloplasti öyküsü olanlar ile olmayanların genotip
dağılımlarında fark bulunmuştur (P= 0.034). TT genotip’e sahip bireylerde valvüloplastinin
diğer genotiplere sahip olan bireylere göre 2.75 kat daha fazla yapıldığı görülmüştür (P=
0.012). Ayrıca AA genotip’e sahip bireylerde valvüloplasti öyküsü diğer genotiplere göre
anlamlı derecede az bulunmuştur. Bu sonuç A allel ya da AA genotipinin RKKH’ da valvül
hasarında koruyucu etkisi olabileceği şeklinde yorumlanmıştır.
Kapak replasmanı yapılmış hasta grubunun genotip dağılımı da incelendiğinde; kapak
replasmanı yapılmış olan hastalar şiddetli kapak hasarının bir göstergesi olarak
değerlendirilmiştir. Kapak replasmanı yapılan ve kapak replasmanı olmayan RKKH olan
bireylerde genotip dağılımında fark bulunmuştur (P=0.01). A homozigot bireylerde diğer
genotiplere sahip olanlara göre kapak replasman sıklığı az bulundu (P: 0.01). Yine bu sonuç
valvüloplasti grubundaki gibi A allelinin RKKH’da kapak hasarının şiddetini korumaya eğilimli
olduğu şeklinde yorumlanmıştır.
RKKH’lı erişkin yaş grubu içinde genellikle kapak hasarı darlık şeklinde görülmektedir.
Yapmış olduğumuz çalışmada darlık ile seyreden kapak hasarında darlığın derecesi ile genotip
dağılımı incelendiğinde; darlığı şiddetli ve hafif olan hastalar arasında T homozigot ya da T
alleline sahip olmanın daralmış kapaklarda darlık şiddetini arttırıcı etkisi olduğu bulunmuştur
( T homozigot için P= 0.012, T allel için P= 0.002).
Çalışmamızda günümüzde RKKH nın progresyonunu engellemek ya da yavaşlatmak
için uygulanan bir tedavi yaklaşımı olan penisilin profilaksisi de değerlendirilmiştir. Kapak
hasarının şiddeti ile düzenli penisilin profilaksisi kullanan bireyler karşılaştırıldığında
istatistiksel olarak anlamlı bir fark bulunamamıştır ( P= 0.613). RKKH nın patofizyolojisinde
otoimmun inflamasyon mevcuttur. Otoimmun reaktivasyonlarda tekrar antijenik uyarıya
gereksinim yoktur ( Streptokoksik M proteini). Dolayısıyla tekrar antijenik bir uyarı
engellemek için kullandığımız penisilin profilaksisinin kapak hasarının progresyonu
engellenlediği bilgiside tartışmalı hale gelmektedir.

40
Sonuç olarak; Bu çalışma; INF-γ ‘nın ekspresyonunu etkileyen 874 T/A polimorfizmi
ile RKKH arasındaki ilişkiyi araştırmak için yapılmıştır. Çalışmamızda INF-γ ‘nın ekspresyon
seviyesindeki değişikliklerin hem RKKH’a yatkınlığa hem de kapak hasarının şiddetine olan
etkisini araştırmayı amaçladık.
INF-γ; kronik inflamatuvar süreçte immune modülasyonun primer sitokinidir ve
romatojen A grubu streptokokus Pyogenes enfeksiyonu ile tetiği çekilen otoimmun
reaksiyonların sonucu oluşan RKKH’a yatkınlıktaki aday genlerdendir.
Bulgularımız INF-γ 874 T alleli ya da T homozigot genotip’e sahip olmanın kapak
hasarını arttırdığı yönündedir. Kapak hasarının darlık yönünde şiddetini gösteren valvüloplasti
öyküsünün genotip ile ilişkisi değerlendirildiğinde de TT genotip’e sahip olmanın valvüloplasti
riskini 2.75 kat arttırdığı gösterilmiştir. Ayrıca A alleline sahip olmanın da kapak hasarı
açısından koruyucu olduğunu düşündürmüştür.
Yapmış olduğumuz çalışmada bazı sınırlılıklar da vardır. Bunlardan bazıları şöyledir;
RKKH patogenezinde birden fazla patofizyolojik mekanizmanın varlığı nedeniyle sadece INF-
γ ‘nın rolünü kabul etmek doğru olmayacaktır. Hasta ve kontrol popülasyonumuz genetik bir
çalışma için sınırlı sayılacak bir katılımdadır. RA hecmesi sayısı hastalar tarafından net olarak
belirtilemediğinden, incelenen parametreler arasına dahil edilememiştir. Teknik sınırlılıklar
nedeniyle RKKH ile INF-γ geni üzerindeki diğer polimorfizmler incelenememiştir. Yine
teknik sınırlılıklar nedeniyle INF-γ nın kantitatif miktar tayini yapılamamıştır.
Romatizmal ateş ve RKKH’nın halen ciddi bir toplum sağlığı sorunu olmaya devam ettiği
ülkemizde, INF-γ gen varyantları ile RKKH ilişkisinin daha kapsamlı ve ileri teknolojik
yöntemlerle araştırılmasının, hasta takibi, tedavi ve profilaksi açısından yol gösterici
olabileceği ve araştırmaya değer olduğu düşünülmüştür.

41
KAYNAKLAR
1) Braunwald, zipes DP, Libby P (eds). Heart Disease: A Textbook of Cardiovascular Medicine
8th ed.
2) Da Silva, C.H. (1999) Rheumatic fever: a multicenter study in the state of Sao Paulo.
Pediatric Committee—Sao Paulo Pediatric Rheumatology Society. Rev Hosp Clin Fac Med Sao
Paulo 54, 85-90, PubMed: 10668278
3) Cunningham, M.W. (2000) Pathogenesis of group A streptococcal infections. Clin Microbiol
Rev 13, 470-511, PubMed: 10885988
4) Taranta, A. (1999) A history of rheumatic fever. In Rheumatic Fever (Narula, J. et al., eds),
pp. 1-40, American Registry Pathology
5) Tylewska, S.K., Fischetti, V.A. and Gibbons, R.J. (1988) Binding of Streptococcus pyogenes
and M protein to epithelial cells differs from that of lipoteichoic acid. Curr Microbiol 16, 209-
216
6) Fischetti, V.A. (1991) Streptococcal M protein. Sci Am 264, 58-65, PubMed: 1857955
7) Manjula, B.N. and Fischetti, V.A. (1986) Sequence homology of group A streptococcal Pep
M5 protein with other coiled-coil proteins. Biochem Biophys Res Commun 140, 684-690,
PubMed: 3535794
8) Dale, J.B. and Beachey, E.H. (1985) Epitopes of streptococcal M proteins shared with
cardiac myosin. J Exp Med 162, 583-591, PubMed: 2410530
9) Fenderson, P.G., Fischetti, V.A. and Cunningham, M.W. (1989) Tropomyosin shares
immunologic epitopes with group A streptococcal M proteins. J Immunol 142, 2475-2481,
PubMed: 2466896
10) Gulizia, J.M., Cunningham, M.W. and McManus, B.M. (1991) Immunoreactivity of
antistreptococcal monoclonal antibodies to human heart valves. Evidence for multiple cross-
reactive epitopes. Am J Pathol 138, 285-301, PubMed: 1704188

42
11) Guilherme, L. et al. (1995) Human heartinfiltrating T-cell clones from rheumatic heart
disease patients recognize both streptococcal and cardiac proteins. Circulation 92, 415-420,
PubMed: 7634457
12) Manjula, B.N. et al. (1984) The complete amino acid sequence of a biologically active 197-
residuefragment of M protein isolated from type 5 group A streptococci. J Biol Chem 259,
3686-3693, PubMed: 6368549
13) Guilherme, L. et al. (2001) T-cell reactivity against streptococcal antigens in the periphery
mirrors reactivity of heart-infiltrating T lymphocytes in rheumatic heart disease patients. Infect
Immun 69, 5345-5351, PubMed: 11500404
14) Galvin, J.E. et al. (2000) Cytotoxic mAb from rheumatic carditis recognizes heart valves
and laminin. J Clin Invest 106, 217-224, PubMed: 10903337
15) Roberts, S. et al. (2001) Pathogenic mechanisms in rheumatic carditis: focus on valvular
endothelium. J Infect Dis 183, 507-511, PubMed: 11133385
16) Unny, S.K. and Middlebrooks, B.L. (1983) Streptococcal rheumatic carditis. Microbiol Rev
47, 97-120, PubMed: 6343829
17) Raizada, V. et al. (1983) Tissue distribution of lymphocytes in rheumatic heart valves as
defined by monoclonal anti-T cell antibodies. Am J Med 74, 90-96, PubMed: 6336893
18) Kemeny, E. et al. (1989) Identification of mononuclear cells and T cell subsets in rheumatic
valvulitis. Clin Immunol Immunopathol 52, 225-237, PubMed: 2786783
19) Miller, L.C. et al. (1989) Cytokines and immunoglobulin in rheumatic heart disease:
production by blood and tonsillar mononuclear cells. J Rheumatol 16, 1436-1442, PubMed:
2600943
20) Fraser, W.J. et al. (1997) Rheumatic Aschoff nodules revisited. II: Cytokine expression
corroborates recently proposed sequential stages. Histopathology 31, 460-464, PubMed:
9416487

43
21) Guilherme, L. et al. (2004) Rheumatic heart disease: proinflammatory cytokines play a role
in the progression and maintenance of valvular lesions. Am J Pathol 165, 1583-1591, PubMed:
15509528
22) Wilson MG, Schweitzer MD. Pattern of hereditary susceptibility in rheumatic fever.
Circulation 1954;10:699–704.
23) Gray FG, Quinn RW, Quinn JP. A long term survey of rheumatic and non-rheumatic
families; with particular reference to environment and heredity. Am J Med 1952;13:400–12.
24) Uchida IA. Possible genetic factors in the etiology of rheumatic fever. Am J Hum Genet
1953;5:61–9.
25) Stevenson AC, Cheeseman EA. Heredity and rheumatic fever; a study of 462 families
ascertained by an affected child and 51 families ascertained by an affected mother. Ann Eugen
1953;17:177–210.
26) Spagnuolo M, Taranta A. Rheumatic fever in siblings. Similarity of its clinical
manifestations. N Engl J Med 1968;278:183–8.
27) Falk JA, Fleischman JL, Zabriskie JB, Falk RE. A study of HL-A antigen phenotype in
rheumatic fever and rheumatic heart disease patients. Tissue Antigens 1973;3:173–8.
28) Matsumori A, Hirose K, Wakabayashi A et al. HLA in hypertrophic cardiomyopathy and
rheumatic heart disease. Jpn Circ J 1979;43:445–9.
29) Ayoub EM, Barrett DJ, Maclaren NK, Krischer JP. Association of class II human
histocompatibility leukocyte antigens with rheumatic fever. J Clin Invest 1986;77:2019–26.
30) Messias Reason IJ, Schafranski MD, Jensenius JC, Steffensen R. The association between
mannose-binding lectin gene polymorphism and rheumatic heart disease. Hum Immunol
2006;67:991
31) Berdeli A, Celik HA, Ozyurek R, Dogrusoz B, Aydin HH. TLR-2 gene Arg753Gln
polymorphism is strongly associated with acute rheumatic fever in children. J Mol Med
2005;83:535–41.

44
32) Duzgun N, Duman T, Haydardedeoglu FE, Tutkak H. The lack of genetic association of the
Toll-like receptor 2 (TLR2) Arg753Gln and Arg677Trp polymorphisms with rheumatic heart
disease. Clin Rheumatol 2007;26:915–9.
33) Chou HT, Tsai CH, Chen WC, Tsai FJ. Lack of association of genetic polymorphisms in the
interleukin-1beta, interleukin-1 receptor antagonist, interleukin-4, and interleukin-10 genes with
risk of rheumatic heart disease in Taiwan Chinese. Int Heart J 2005; 46:397–406.
34) Ramasawmy R, Fae KC, Spina G et al. Association of polymorphisms within the promoter
region of the tumor necrosis factoralpha with clinical outcomes of rheumatic fever. Mol
Immunol 2007;44:1873–8.
35) Sallakci N, Akcurin G, Koksoy S et al. TNF-alpha G-308A polymorphism is associated
with rheumatic fever and correlates with increased TNF-alpha production. J Autoimmun
2005;25:50–4.
36) Berdeli A, Tabel Y, Celik HA, Ozyurek R, Dogrusoz B, Aydin HH. Lack of association
between TNFalpha gene polymorphism at position -308 and risk of acute rheumatic fever in
Turkish patients. Scand J Rheumatol 2006;35:44–7.
37) Kroeger KM, Carville KS, Abraham LJ. The )308 tumor necrosis factor-alpha promoter
polymorphism effects transcription. Mol Immunol 1997;34:391–9.
38) Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-gamma. Annu
Rev Immunol 1997;15:749–795.
39) de Veer MJ, et al. Functional classification of interferon-stimulated genes identified using
microarrays. J Leukoc Biol 2001;69: 912–920.
40) Schroder K, Hertzog PJ, Ravasi T, Hume DA. Interferon-gamma: an overview of signals,
mechanisms and functions. J Leukoc Biol 2004;75:163–189.
41) Ramana CV, Gil MP, Schreiber RD, Stark GR. Stat1-dependent and -independent pathways
in IFN-gamma-dependent signaling. Trends Immunol 2002;23:96–101.

45
42) Johnson DR, Pober JS. Tumor necrosis factor and immune interferon synergistically
increase transcription of HLA class I heavy- and light-chain genes in vascular endothelium.
Proc Natl Acad Sci USA 1990;87: 5183–5187.
43) Yoshida A, Koide Y, Uchijima M, Yoshida TO. IFN-gamma induces IL-12 mRNA
expression by a murine macrophage cell line, J774. Biochem Biophys Res Commun
1994;198:857–861.
44) Trinchieri G, Pflanz S, Kastelein RA. The IL-12 family of heterodimeric cytokines. New
players in the regulation of T cell responses. Immunity 2003;19:641–644.
45) Struyf S, et al. Synergistic induction of MCP-1 and -2 by IL-1beta and interferons in
fibroblasts and epithelial cells. J Leukoc Biol 1998;63: 364–372.
46) McLoughlin RM, et al. Interplay between IFN-gamma and IL-6 signaling governs
neutrophil trafficking and apoptosis during acute inflammation. J Clin Invest 2003;112:598–
607.
47) Glimcher LH, Murphy KM. Lineage commitment in the immune system: the
T helper lymphocyte grows up. Genes Dev 2000;14:1693–1711.
48) Peng SL, Szabo SJ, Glimcher LH. T-bet regulates IgG class switching and pathogenic
autoantibody production. Proc Natl Acad Sci USA 2002;99:5545–5550.
49) Mach B, Steimle V, Martinez-Soria E, Reith W. Regulation of MHC class II genes: lessons
from a disease. Annu Rev Immunol 1996;14:301–331.
50) Buchmu¨ ller Y, Mauel J. Studies on the mechanisms of macrophage activation. II. Parasite
destruction in macrophages activated by supernates from concanavalin A-stimulated
lymphocytes. J Exp Med 1979;150:359–70.
51) Nathan C, Murray HW, Wiebe ME, Rubin BY. Identification of interferon-gama as the
lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity.
J Exp Med 1983;158:670–81.

46
52) Steeg P, Moore RN, Johnson HM, Oppenheim JJ. Regulation of murine macrophage Ia
antigen expression by a lymphokine with immune interferon activity. J Exp Med
1982;156:1780–93.
53) Pernis A, Gupta, Gollob KJ, Garfein E, Coffman RL, Schindler C, et al. Lack of interferon g
receptor b chain and the prevention of interferon gama signalling in TH1 cells. Science
1995;269:245
54) Rabin EM, Mond JJ, Ohara J, Paul WE. Interferon-gamma inhibits the action of B cell
stimulatory factor (BSF)-1 on resting B cells. J Immunol 1986;137:1573–6.
55) RongcunY,MaesH,Corsi M,Dellner F,WenT, Kiessling R. Interferon gamma impairs the
ability of monocyte-derived dendritic cells to present tumour-specific and allo-specific antigens
and reduces their expression of CD1A, CD80 AND CD4. Cytokine 1998;10:747–55.
56) Mosca PJ, Hobeika AC, Clay TM, Nair SK, Thomas EK, Morse MA, et al. A subset of
human monocyte-derived dendritic cells expresses high levels of interleukin-12 in response to
combined CD40 ligand and interferon-gamma treatment. Blood 2000;96:3499–504.
57) Nabel GJ, Verma IM (November 1993). "Proposed NF-κB/IκB family nomenclature".
Genes Dev. 7 (11): 2063.)
58) Livolsi A, Busuttil V, Imbert V, Abraham RT, Peyron JF (March 2001). "Tyrosine
phosphorylation-dependent activation of NF-κB. Requirement for p56 LCK and ZAP-70 protein
tyrosine kinases". Eur. J. Biochem. 268 )
59) Pravica V, Asderakis A, Perrey C, Hajeer A, Sinnott PJ,Hutchinson IV: In vitro production
of IFN-g correlateswith CA repeat polymorphism in the human IFN-g gene. Eur J Immunogenet
26:1, 1999.
60)Pravica, V., Perrey, C., Stevens, A., Lee, J.-H., Hutchinson, I. V. A single nucleotide
polymorphism in the first intron of the human IFN-gamma gene: absolute correlation with a
polymorphic CA microsatellite marker of high IFN-gamma production. Hum. Immun. 61: 863-
866, 2000. [PubMed: 11053629]

47
61) Rossouw, M., Nel, H. J., Cooke, G. S., van Helden, P. D., Hoal, E. G. Association between
tuberculosis and a polymorphic NF-kappa-B binding site in the interferon gamma gene. Lancet
361: 1871-1872, 2003. [PubMed: 12788577]
62) Awad M, Pravica V, Perrey C, El Gamel A, Yonan N,Sinnott PJ, Hutchinson IV: CA repeat
allele polymorphismin the first intron of the human interferon-g gene is associated with lung
allograft fibrosis. Hum Immunol60:343, 1999.
63) Stollerman GH. Rheumatic fever. Lancet. 1997;349:935–942
64) DiSciascio G, Taranta A. Rheumatic fever in children. Am Heart J. 1980;99:635–658
65) Dajani AS, Bisno AL, Chung KJ, Durack DT, Gerber MA, Kaplan EL, Millard HD,
Randolph MF, Shulman ST, J. Immunol., October 15, 2005; 175(8): 5448 – 5456
66) Luiza Guilherme, Kellen Faé, Sandra E. Oshiro and Jorge Kalil: Molecular pathogenesis of
rheumatic fever and rheumatic heart disease. expert reviews in molecular medicine 2005 Dec
8;7(28):1-15
67) Fraser WJ, Haffejee Z, Jankelow D, Wadee A, Cooper K: Rheumatic Aschoff nodules
revisited. II: Cytokine expression corroborates recently proposed sequential stages.
Histopathology 1997 Nov;31(5):460-4.
68)Sallakci N, Coskun M, Berber Z, Gürkan F, Kocamaz H, Uysal G, Bhuju S, Yavuzer U,
Singh M, Yeğin O: Interferon-gamma gene+874T-A polymorphism is associated with
tuberculosis and gamma interferon response.Tuberculosis (Edinb). 2007 May;87(3):225-30.
Epub 2007 Feb 2
69) Khani-Hanjani A, Lacaille D, Hoar D, Chalmers A, Horsman D, Anderson M, Balshaw R,
Keown PA. Association between dinucleotide repeat in non-coding region of interferon-gamma
gene and susceptibility to, and severity of, rheumatoid arthritis. Lancet. 2000 Sep
2;356(9232):820-5.
70) Dickinson AM, Cavet J, Cullup H, Wang XN, Jarvis M, Sviland L, Middleton PG. )
Predicting outcome in hematological stem cell transplantation. Arch Immunol Ther Exp
(Warsz). 2002;50(6):371-8. Review

48
HAM VERİLER
Genotip( Hasta)
Frequency Percent
Valid
Percent
Cumulative
Percent
Valid AT 97 32,0 63,8 63,8
AA 24 7,9 15,8 79,6
TT 31 10,2 20,4 100,0
Total 152 50,2 100,0
Missing System 151 49,8
Total 303 100,0
Genotip (kontrol)
Frequency Percent
Valid
Percent
Cumulative
Percent
Valid AT 88 29,0 58,3 58,3
AA 47 15,5 31,1 89,4
TT 16 5,3 10,6 100,0
Total 151 49,8 100,0
Missing System 152 50,2
Total 303 100,0
Statistics
Yas
Yaş ( Hasta
Grubu)
Yaş (
Kontrol
Grubu)
N Valid 302 152 151
Missing 1 151 152
Mean 49,58 49,7697 49,3907
Std. Deviation 13,164 13,42404 12,89340
Minimum 21 21,00 21,00
Maximum 81 81,00 81,00

49
Genotip * Hasta- Kontrol
Crosstab
Hasta- Kontrol Total
Kontrol Hasta Kontrol
Genotip AT Count 88 97 185
Expected Count 92,2 92,8 185,0
% within Genotip 47,6% 52,4% 100,0%
% within Hasta-
Kontrol 58,3% 63,8% 61,1%
AA Count 47 24 71
Expected Count 35,4 35,6 71,0
% within Genotip 66,2% 33,8% 100,0%
% within Hasta-
Kontrol 31,1% 15,8% 23,4%
TT Count 16 31 47
Expected Count 23,4 23,6 47,0
% within Genotip 34,0% 66,0% 100,0%
% within Hasta-
Kontrol 10,6% 20,4% 15,5%
Total Count 151 152 303
Expected Count 151,0 152,0 303,0
% within Genotip 49,8% 50,2% 100,0%
% within Hasta-
Kontrol 100,0% 100,0% 100,0%
Chi-Square Tests
Value Df
Asymp. Sig.
(2-sided)
Pearson Chi-Square 12,673(a) 2 ,002
Likelihood Ratio 12,894 2 ,002
Linear-by-Linear
Association ,246 1 ,620
N of Valid Cases 303

50
T homozigot (Hasta-Kontrol) * Hasta- Kontrol
Crosstab
Hasta- Kontrol Total
Kontrol Hasta Kontrol
T homozigot
(Hasta-
Kontrol)
AA/A
T
Count 135 121 256
Expected Count 127,6 128,4 256,0
% within T homozigot
(Hasta-Kontrol) 52,7% 47,3% 100,0%
% within Hasta-
Kontrol 89,4% 79,6% 84,5%
TT Count 16 31 47
Expected Count 23,4 23,6 47,0
% within T homozigot
(Hasta-Kontrol) 34,0% 66,0% 100,0%
% within Hasta-
Kontrol 10,6% 20,4% 15,5%
Total Count 151 152 303
Expected Count 151,0 152,0 303,0
% within T homozigot
(Hasta-Kontrol) 49,8% 50,2% 100,0%
% within Hasta-
Kontrol 100,0% 100,0% 100,0%
Chi-Square Tests
Value Df
Asymp. Sig.
(2-sided)
Exact Sig.
(2-sided)
Exact Sig.
(1-sided)
Pearson Chi-Square 5,550(b) 1 ,018
Continuity
Correction(a) 4,827 1 ,028
Likelihood Ratio 5,635 1 ,018
Fisher's Exact Test ,026 ,014
Linear-by-Linear
Association 5,531 1 ,019
N of Valid Cases 303
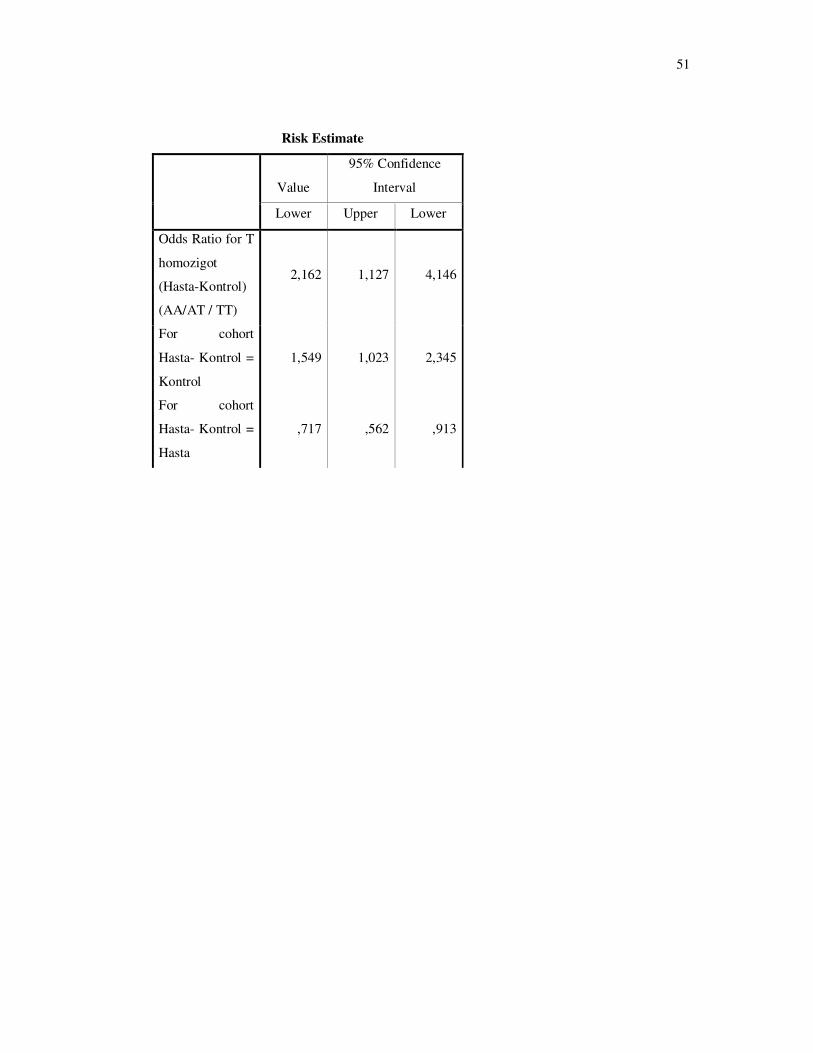
51
Risk Estimate
Value
95% Confidence
Interval
Lower Upper Lower
Odds Ratio for T
homozigot
(Hasta-Kontrol)
(AA/AT / TT)
2,162 1,127 4,146
For cohort
Hasta- Kontrol =
Kontrol
1,549 1,023 2,345
For cohort
Hasta- Kontrol =
Hasta
,717 ,562 ,913
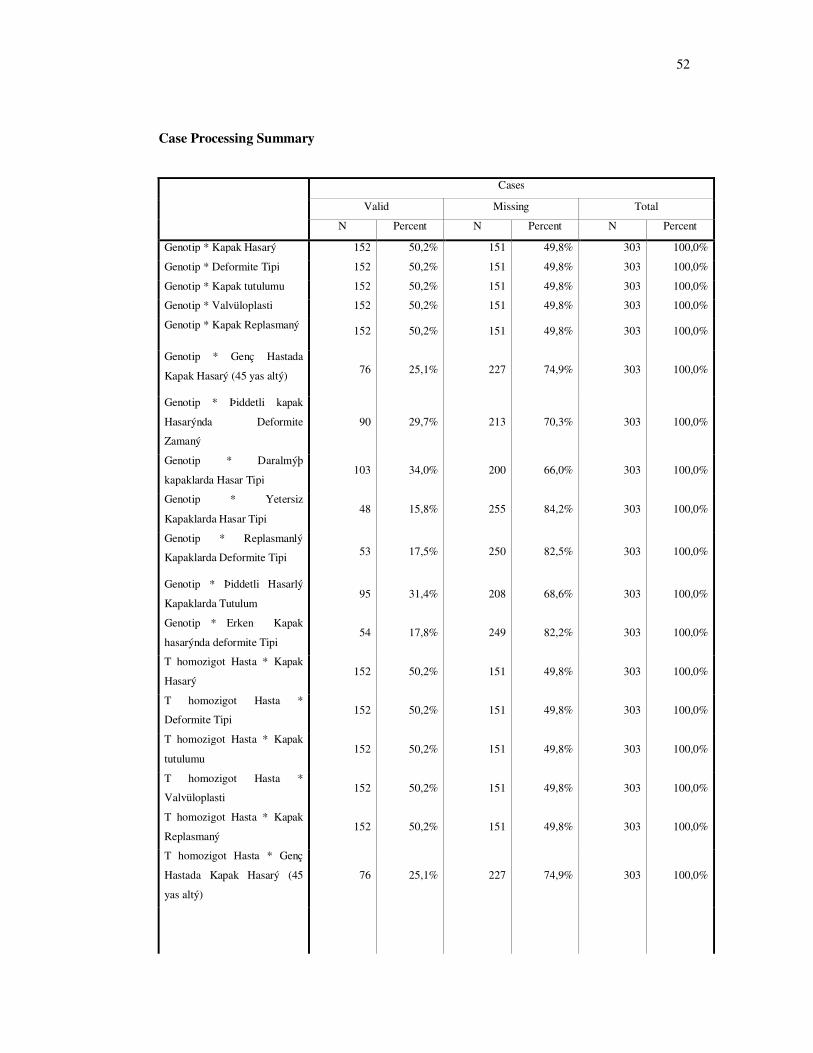
52
Case Processing Summary
Cases
Valid Missing Total
N Percent N Percent N Percent
Genotip * Kapak Hasarý 152 50,2% 151 49,8% 303 100,0%
Genotip * Deformite Tipi 152 50,2% 151 49,8% 303 100,0%
Genotip * Kapak tutulumu 152 50,2% 151 49,8% 303 100,0%
Genotip * Valvüloplasti 152 50,2% 151 49,8% 303 100,0%
Genotip * Kapak Replasmaný 152 50,2% 151 49,8% 303 100,0%
Genotip * Genç Hastada
Kapak Hasarý (45 yas altý) 76 25,1% 227 74,9% 303 100,0%
Genotip * Þiddetli kapak
Hasarýnda Deformite
Zamaný
90 29,7% 213 70,3% 303 100,0%
Genotip * Daralmýþ
kapaklarda Hasar Tipi 103 34,0% 200 66,0% 303 100,0%
Genotip * Yetersiz
Kapaklarda Hasar Tipi 48 15,8% 255 84,2% 303 100,0%
Genotip * Replasmanlý
Kapaklarda Deformite Tipi 53 17,5% 250 82,5% 303 100,0%
Genotip * Þiddetli Hasarlý
Kapaklarda Tutulum 95 31,4% 208 68,6% 303 100,0%
Genotip * Erken Kapak
hasarýnda deformite Tipi 54 17,8% 249 82,2% 303 100,0%
T homozigot Hasta * Kapak
Hasarý 152 50,2% 151 49,8% 303 100,0%
T homozigot Hasta *
Deformite Tipi 152 50,2% 151 49,8% 303 100,0%
T homozigot Hasta * Kapak
tutulumu 152 50,2% 151 49,8% 303 100,0%
T homozigot Hasta *
Valvüloplasti 152 50,2% 151 49,8% 303 100,0%
T homozigot Hasta * Kapak
Replasmaný 152 50,2% 151 49,8% 303 100,0%
T homozigot Hasta * Genç
Hastada Kapak Hasarý (45
yas altý)
76 25,1% 227 74,9% 303 100,0%

53
T homozigot Hasta *
Daralmýþ kapaklarda Hasar
Tipi
103 34,0% 200 66,0% 303 100,0%
T homozigot Hasta * Yetersiz
Kapaklarda Hasar Tipi 48 15,8% 255 84,2% 303 100,0%
T homozigot Hasta *
Replasmanlý Kapaklarda
Deformite Tipi
53 17,5% 250 82,5% 303 100,0%
T homozigot Hasta * Þiddetli
Hasarlý Kapaklarda Tutulum 95 31,4% 208 68,6% 303 100,0%
T homozigot Hasta * Erken
Kapak hasarýnda deformite
Tipi
54 17,8% 249 82,2% 303 100,0%
Genotip * Kapak Hasarı
Crosstab
Kapak Hasarı Total
Hafif şiddetli
Genotip AT Count 34 63 97
Expected Count 35,1 61,9 97,0
% within Genotip 35,1% 64,9% 100,0%
% within Kapak Hasarý 61,8% 64,9% 63,8%
AA Count 16 8 24
Expected Count 8,7 15,3 24,0
% within Genotip 66,7% 33,3% 100,0%
% within Kapak Hasarý 29,1% 8,2% 15,8%
TT Count 5 26 31
Expected Count 11,2 19,8 31,0
% within Genotip 16,1% 83,9% 100,0%
% within Kapak Hasarý 9,1% 26,8% 20,4%
Total Count 55 97 152
Expected Count 55,0 97,0 152,0
% within Genotip 36,2% 63,8% 100,0%
% within Kapak Hasarý 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Pearson Chi-Square 15,111(a) 2 ,001
Likelihood Ratio 15,348 2 ,000
Linear-by-Linear
Association 1,135 1 ,287
N of Valid Cases 152
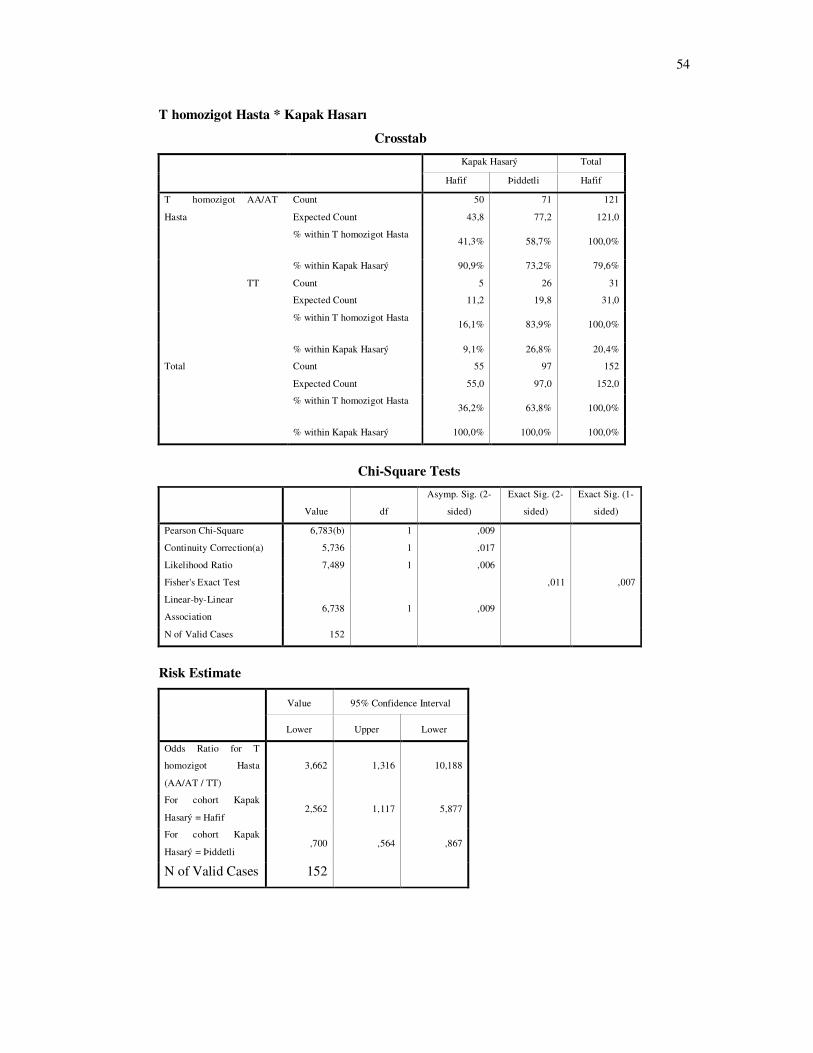
54
T homozigot Hasta * Kapak Hasarı
Crosstab
Kapak Hasarý Total
Hafif Þiddetli Hafif
T homozigot
Hasta
AA/AT Count 50 71 121
Expected Count 43,8 77,2 121,0
% within T homozigot Hasta 41,3% 58,7% 100,0%
% within Kapak Hasarý 90,9% 73,2% 79,6%
TT Count 5 26 31
Expected Count 11,2 19,8 31,0
% within T homozigot Hasta 16,1% 83,9% 100,0%
% within Kapak Hasarý 9,1% 26,8% 20,4%
Total Count 55 97 152
Expected Count 55,0 97,0 152,0
% within T homozigot Hasta 36,2% 63,8% 100,0%
% within Kapak Hasarý 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Exact Sig. (2-
sided)
Exact Sig. (1-
sided)
Pearson Chi-Square 6,783(b) 1 ,009
Continuity Correction(a) 5,736 1 ,017
Likelihood Ratio 7,489 1 ,006
Fisher's Exact Test ,011 ,007
Linear-by-Linear
Association 6,738 1 ,009
N of Valid Cases 152
Risk Estimate
Value 95% Confidence Interval
Lower Upper Lower
Odds Ratio for T
homozigot Hasta
(AA/AT / TT)
3,662 1,316 10,188
For cohort Kapak
Hasarý = Hafif 2,562 1,117 5,877
For cohort Kapak
Hasarý = Þiddetli ,700 ,564 ,867
N of Valid Cases 152

55
Genotip * Deformite Tipi
Crosstab
Deformite Tipi Total
Yetersizlik Darlýk Yetersizlik
Genotip AT Count 37 60 97
Expected Count 30,6 66,4 97,0
% within Genotip 38,1% 61,9% 100,0%
% within Deformite Tipi 77,1% 57,7% 63,8%
AA Count 7 17 24
Expected Count 7,6 16,4 24,0
% within Genotip 29,2% 70,8% 100,0%
% within Deformite Tipi 14,6% 16,3% 15,8%
TT Count 4 27 31
Expected Count 9,8 21,2 31,0
% within Genotip 12,9% 87,1% 100,0%
% within Deformite Tipi 8,3% 26,0% 20,4%
Total Count 48 104 152
Expected Count 48,0 104,0 152,0
% within Genotip 31,6% 68,4% 100,0%
% within Deformite Tipi 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Pearson Chi-Square 7,004(a) 2 ,030
Likelihood Ratio 7,810 2 ,020
Linear-by-Linear
Association 6,841 1 ,009
N of Valid Cases 152

56
T homozigot Hasta * Deformite Tipi
Crosstab
Deformite Tipi Total
Yetersizlik Darlýk Yetersizlik
T homozigot
Hasta
AA/AT Count 44 77 121
Expected Count 38,2 82,8 121,0
% within T homozigot Hasta 36,4% 63,6% 100,0%
% within Deformite Tipi 91,7% 74,0% 79,6%
TT Count 4 27 31
Expected Count 9,8 21,2 31,0
% within T homozigot Hasta 12,9% 87,1% 100,0%
% within Deformite Tipi 8,3% 26,0% 20,4%
Total Count 48 104 152
Expected Count 48,0 104,0 152,0
% within T homozigot Hasta 31,6% 68,4% 100,0%
% within Deformite Tipi 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Exact Sig. (2-
sided)
Exact Sig. (1-
sided)
Pearson Chi-Square 6,286(b) 1 ,012
Continuity Correction(a) 5,247 1 ,022
Likelihood Ratio 7,123 1 ,008
Fisher's Exact Test ,016 ,008
Linear-by-Linear
Association 6,245 1 ,012
N of Valid Cases 152
Risk Estimate
Value 95% Confidence Interval
Lower Upper Lower
Odds Ratio for T homozigot
Hasta (AA/AT / TT) 3,857 1,267 11,745
For cohort Deformite Tipi =
Yetersizlik 2,818 1,096 7,247
For cohort Deformite Tipi =
Darlýk ,731 ,604 ,884
N of Valid Cases 152

57
Genotip * Valvüloplasti
Crosstab
Valvüloplasti Total
Valvüloplasti Yok Valvüloplasti Var
Genotip AT Count 66 31 97
Expected Count 62,5 34,5 97,0
% within Genotip 68,0% 32,0% 100,0%
% within Valvüloplasti 67,3% 57,4% 63,8%
AA Count 18 6 24
Expected Count 15,5 8,5 24,0
% within Genotip 75,0% 25,0% 100,0%
% within Valvüloplasti 18,4% 11,1% 15,8%
TT Count 14 17 31
Expected Count 20,0 11,0 31,0
% within Genotip 45,2% 54,8% 100,0%
% within Valvüloplasti 14,3% 31,5% 20,4%
Total Count 98 54 152
Expected Count 98,0 54,0 152,0
% within Genotip 64,5% 35,5% 100,0%
% within Valvüloplasti 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Pearson Chi-Square 6,748(a) 2 ,034
Likelihood Ratio 6,567 2 ,037
Linear-by-Linear
Association 3,897 1 ,048
N of Valid Cases 152
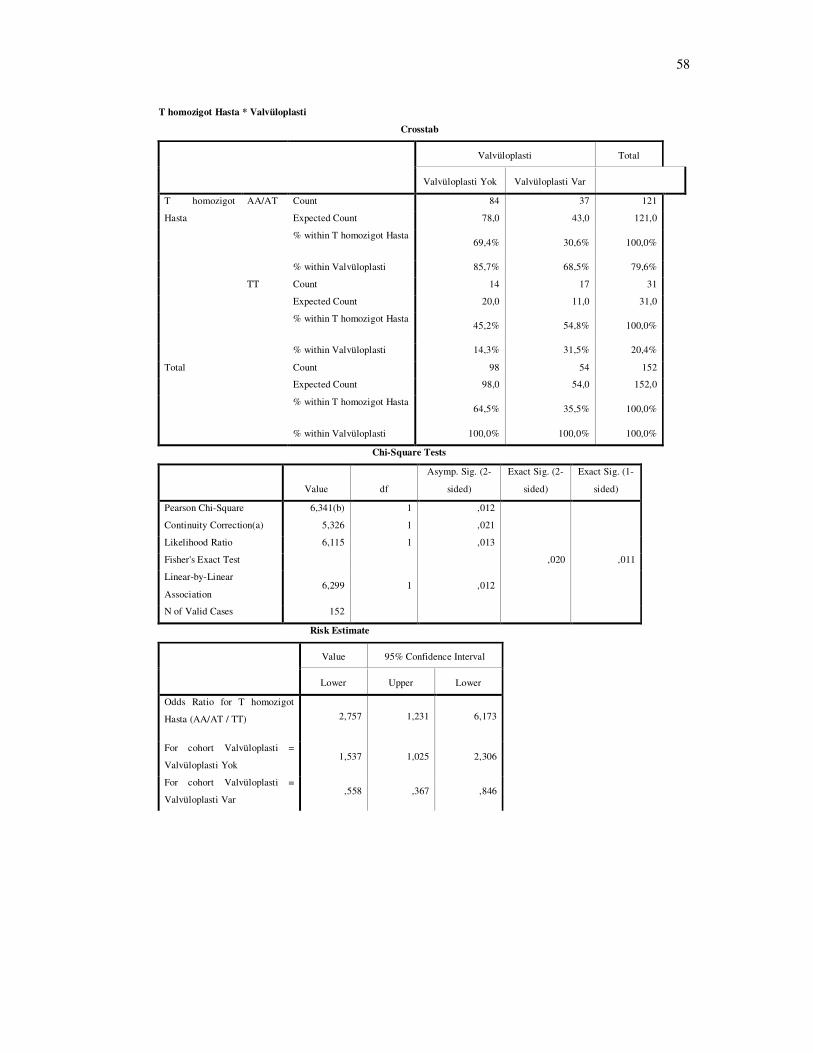
58
T homozigot Hasta * Valvüloplasti
Crosstab
Valvüloplasti Total
Valvüloplasti Yok Valvüloplasti Var
T homozigot
Hasta
AA/AT Count 84 37 121
Expected Count 78,0 43,0 121,0
% within T homozigot Hasta 69,4% 30,6% 100,0%
% within Valvüloplasti 85,7% 68,5% 79,6%
TT Count 14 17 31
Expected Count 20,0 11,0 31,0
% within T homozigot Hasta 45,2% 54,8% 100,0%
% within Valvüloplasti 14,3% 31,5% 20,4%
Total Count 98 54 152
Expected Count 98,0 54,0 152,0
% within T homozigot Hasta 64,5% 35,5% 100,0%
% within Valvüloplasti 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Exact Sig. (2-
sided)
Exact Sig. (1-
sided)
Pearson Chi-Square 6,341(b) 1 ,012
Continuity Correction(a) 5,326 1 ,021
Likelihood Ratio 6,115 1 ,013
Fisher's Exact Test ,020 ,011
Linear-by-Linear
Association 6,299 1 ,012
N of Valid Cases 152
Risk Estimate
Value 95% Confidence Interval
Lower Upper Lower
Odds Ratio for T homozigot
Hasta (AA/AT / TT) 2,757 1,231 6,173
For cohort Valvüloplasti =
Valvüloplasti Yok 1,537 1,025 2,306
For cohort Valvüloplasti =
Valvüloplasti Var ,558 ,367 ,846

59
Genotip * Kapak Replasmanı
Crosstab
Kapak Replasmanı Total
Replasman Yok Replasman Var
Genotip AT Count 64 33 97
Expected Count 63,8 33,2 97,0
% within Genotip 66,0% 34,0% 100,0%
% within Kapak
Replasmaný 64,0% 63,5% 63,8%
AA Count 21 3 24
Expected Count 15,8 8,2 24,0
% within Genotip 87,5% 12,5% 100,0%
% within Kapak
Replasmaný 21,0% 5,8% 15,8%
TT Count 15 16 31
Expected Count 20,4 10,6 31,0
% within Genotip 48,4% 51,6% 100,0%
% within Kapak
Replasmaný 15,0% 30,8% 20,4%
Total Count 100 52 152
Expected Count 100,0 52,0 152,0
% within Genotip 65,8% 34,2% 100,0%
% within Kapak
Replasmaný 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Pearson Chi-Square 9,199(a) 2 ,010
Likelihood Ratio 9,881 2 ,007
Linear-by-Linear
Association 1,383 1 ,240
N of Valid Cases 152
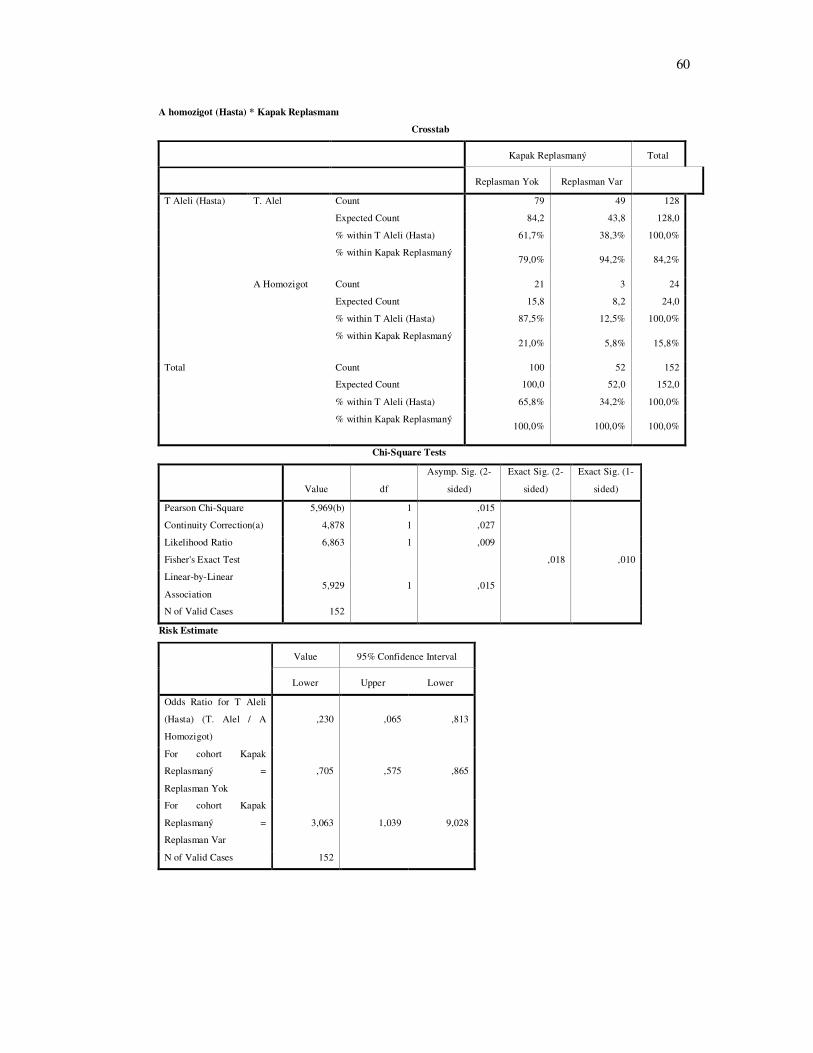
60
A homozigot (Hasta) * Kapak Replasmanı
Crosstab
Kapak Replasmaný Total
Replasman Yok Replasman Var
T Aleli (Hasta) T. Alel Count 79 49 128
Expected Count 84,2 43,8 128,0
% within T Aleli (Hasta) 61,7% 38,3% 100,0%
% within Kapak Replasmaný 79,0% 94,2% 84,2%
A Homozigot Count 21 3 24
Expected Count 15,8 8,2 24,0
% within T Aleli (Hasta) 87,5% 12,5% 100,0%
% within Kapak Replasmaný 21,0% 5,8% 15,8%
Total Count 100 52 152
Expected Count 100,0 52,0 152,0
% within T Aleli (Hasta) 65,8% 34,2% 100,0%
% within Kapak Replasmaný 100,0% 100,0% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Exact Sig. (2-
sided)
Exact Sig. (1-
sided)
Pearson Chi-Square 5,969(b) 1 ,015
Continuity Correction(a) 4,878 1 ,027
Likelihood Ratio 6,863 1 ,009
Fisher's Exact Test ,018 ,010
Linear-by-Linear
Association 5,929 1 ,015
N of Valid Cases 152
Risk Estimate
Value 95% Confidence Interval
Lower Upper Lower
Odds Ratio for T Aleli
(Hasta) (T. Alel / A
Homozigot)
,230 ,065 ,813
For cohort Kapak
Replasmaný =
Replasman Yok
,705 ,575 ,865
For cohort Kapak
Replasmaný =
Replasman Var
3,063 1,039 9,028
N of Valid Cases 152

61
Genotip * Daralmış kapaklarda Hasar Tipi
Crosstab
Daralmýþ kapaklarda Hasar
Tipi Total
Hafif Şiddetli
Genotip AT Count 18 42 60
Expected Count 18,1 41,9 60,0
% within Genotip 30,0% 70,0% 100,0%
% within Daralmýþ
kapaklarda Hasar Tipi 58,1% 58,3% 58,3%
AA Count 10 6 16
Expected Count 4,8 11,2 16,0
% within Genotip 62,5% 37,5% 100,0%
% within Daralmýþ
kapaklarda Hasar Tipi 32,3% 8,3% 15,5%
TT Count 3 24 27
Expected Count 8,1 18,9 27,0
% within Genotip 11,1% 88,9% 100,0%
% within Daralmýþ
kapaklarda Hasar Tipi 9,7% 33,3% 26,2%
Total Count 31 72 103
Expected Count 31,0 72,0 103,0
% within Genotip 30,1% 69,9% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Pearson Chi-Square 12,611(a) 2 ,002
Likelihood Ratio 12,696 2 ,002
Linear-by-Linear
Association 1,582 1 ,208
N of Valid Cases 103

62
T homozigot Hasta * Daralmış kapaklarda Hasar Tipi
Crosstab
Daralmýþ kapaklarda Hasar
Tipi Total
Hafif Þiddetli Hafif
T homozigot
Hasta
AA/AT Count 28 48 76
Expected Count 22,9 53,1 76,0
% within T homozigot Hasta 36,8% 63,2% 100,0%
% within Daralmýþ
kapaklarda Hasar Tipi 90,3% 66,7% 73,8%
TT Count 3 24 27
Expected Count 8,1 18,9 27,0
% within T homozigot Hasta 11,1% 88,9% 100,0%
% within Daralmýþ
kapaklarda Hasar Tipi 9,7% 33,3% 26,2%
Total Count 31 72 103
Expected Count 31,0 72,0 103,0
% within T homozigot Hasta 30,1% 69,9% 100,0%
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Exact Sig. (2-
sided)
Exact Sig. (1-
sided)
Pearson Chi-Square 6,270(b) 1 ,012
Continuity Correction(a) 5,106 1 ,024
Likelihood Ratio 7,137 1 ,008
Fisher's Exact Test ,014 ,009
Linear-by-Linear
Association 6,209 1 ,013
N of Valid Cases 103
Risk Estimate
Value 95% Confidence Interval
Lower Upper Lower
Odds Ratio for T
homozigot Hasta (AA/AT /
TT)
4,667 1,288 16,911
For cohort Daralmýþ
kapaklarda Hasar Tipi =
Hafif
3,316 1,096 10,028
For cohort Daralmýþ
kapaklarda Hasar Tipi =
Þiddetli
,711 ,572 ,883
N of Valid Cases 103
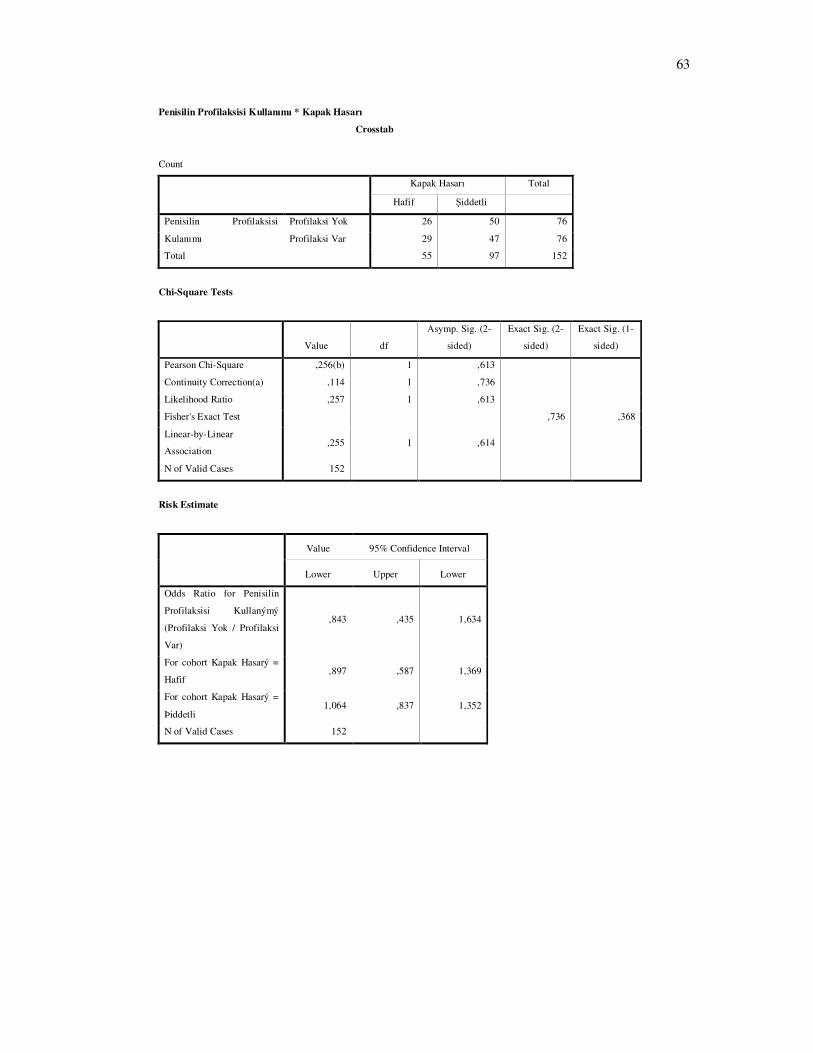
63
Penisilin Profilaksisi Kullanımı * Kapak Hasarı
Crosstab
Count
Kapak Hasarı Total
Hafif Şiddetli
Penisilin Profilaksisi
Kulanımı
Profilaksi Yok 26 50 76
Profilaksi Var 29 47 76
Total 55 97 152
Chi-Square Tests
Value df
Asymp. Sig. (2-
sided)
Exact Sig. (2-
sided)
Exact Sig. (1-
sided)
Pearson Chi-Square ,256(b) 1 ,613
Continuity Correction(a) ,114 1 ,736
Likelihood Ratio ,257 1 ,613
Fisher's Exact Test ,736 ,368
Linear-by-Linear
Association ,255 1 ,614
N of Valid Cases 152
Risk Estimate
Value 95% Confidence Interval
Lower Upper Lower
Odds Ratio for Penisilin
Profilaksisi Kullanýmý
(Profilaksi Yok / Profilaksi
Var)
,843 ,435 1,634
For cohort Kapak Hasarý =
Hafif ,897 ,587 1,369
For cohort Kapak Hasarý =
Þiddetli 1,064 ,837 1,352
N of Valid Cases 152

64
Hasta
No
Gru
p
cins
iyet
Yaş
Gen
oti
p
Kap
ak h
asar
dere
ce
si
Valv
ülo
pla
st
öyk
üs
ü
Kap
ak
rep
lasm
an
öykü
sü
Pro
fila
ksi
1 Hasta Kadın 70 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
2 Hasta Kadın 64 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
3 Hasta Kadın 65 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
4 Hasta Kadın 43 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
5 Hasta Kadın 45 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
6 Hasta Kadın 29 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
7 Hasta Kadın 49 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
8 Hasta Kadın 40 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
9 Hasta Kadın 56 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
10 Hasta Kadın 60 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
11 Hasta Kadın 65 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
12 Hasta Kadın 49 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
13 Hasta Kadın 28 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
14 Hasta Kadın 61 AA Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
15 Hasta Kadın 64 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
16 Hasta Kadın 48 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
17 Hasta Kadın 50 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
18 Hasta Kadın 67 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
19 Hasta Kadın 72 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
20 Hasta Erkek 40 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
21 Hasta Kadın 32 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
22 Hasta Kadın 52 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
23 Hasta Kadın 34 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
24 Hasta Kadın 22 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
25 Hasta Kadın 51 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
26 Hasta Kadın 27 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
27 Hasta Kadın 61 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
28 Hasta Kadın 68 AA Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
29 Hasta Kadın 35 AA Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
30 Hasta Erkek 68 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
31 Hasta Kadın 46 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
32 Hasta Erkek 44 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
33 Hasta Erkek 37 AA Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
34 Hasta Kadın 46 AA Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
35 Hasta Erkek 50 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
36 Hasta Kadın 52 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Var
37 Hasta Kadın 46 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
38 Hasta Kadın 53 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
39 Hasta Kadın 65 TT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
40 Hasta Kadın 61 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
41 Hasta Erkek 40 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
42 Hasta Kadın 50 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
43 Hasta Kadın 28 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
44 Hasta Kadın 60 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok

65
45 Hasta Kadın 34 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
46 Hasta Kadın 54 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
47 Hasta Erkek 70 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
48 Hasta Erkek 54 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
49 Hasta Kadın 79 AA Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
50 Hasta Kadın 55 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
51 Hasta Kadın 52 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
52 Hasta Kadın 39 TT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Yok
53 Hasta Erkek 55 AT Şiddetli Valvüloplasti Var Replasman Yok Profilaksi Yok
54 Hasta Kadın 51 AT Şiddetli Valvüloplasti Var Replasman Var Profilaksi Var
55 Hasta Erkek 69 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
56 Hasta Kadın 45 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
57 Hasta Kadın 63 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
58 Hasta Kadın 61 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
59 Hasta Kadın 60 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
60 Hasta Kadın 67 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
61 Hasta Kadın 50 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
62 Hasta Kadın 59 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
63 Hasta Kadın 49 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
64 Hasta Erkek 40 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
65 Hasta Kadın 50 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
66 Hasta Erkek 50 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
67 Hasta Kadın 35 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
68 Hasta Kadın 36 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
69 Hasta Kadın 50 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
70 Hasta Erkek 79 TT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
71 Hasta Erkek 37 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
72 Hasta Erkek 49 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
73 Hasta Kadın 56 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
74 Hasta Kadın 27 AT Şiddetli Valvüloplasti Replasman Var Profilaksi Var

66
Yok
75 Hasta Erkek 51 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
76 Hasta Kadın 40 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
77 Hasta Kadın 61 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
78 Hasta Erkek 47 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Var
79 Hasta Erkek 63 AT Şiddetli
Valvüloplasti
Yok Replasman Var Profilaksi Yok
80 Hasta Erkek 60 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
81 Hasta Kadın 72 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
82 Hasta Kadın 53 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
83 Hasta Kadın 34 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
84 Hasta Kadın 43 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
85 Hasta Kadın 36 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
86 Hasta Erkek 57 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
87 Hasta Kadın 54 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
88 Hasta Kadın 60 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
89 Hasta Kadın 48 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
90 Hasta Kadın 52 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
91 Hasta Erkek 56 TT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
92 Hasta Kadın 64 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
93 Hasta Kadın 27 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
94 Hasta Kadın 74 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
95 Hasta Kadın 60 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
96 Hasta Kadın 36 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
97 Hasta Kadın 66 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
98 Hasta Kadın 58 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var

67
99 Hasta Erkek 70 TT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
100 Hasta Kadın 81 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
101 Hasta Kadın 65 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
102 Hasta Kadın 56 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
103 Hasta Kadın 38 TT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
104 Hasta Kadın 46 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
105 Hasta Kadın 55 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
106 Hasta Kadın 50 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
107 Hasta Kadın 31 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
108 Hasta Kadın 47 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
109 Hasta Kadın 25 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
110 Hasta Kadın 58 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
111 Hasta Kadın 51 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
112 Hasta Kadın 47 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
113 Hasta Kadın 36 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
114 Hasta Kadın 27 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
115 Hasta Kadın 27 TT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
116 Hasta Kadın 56 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
117 Hasta Kadın 50 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
118 Hasta Kadın 31 TT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
119 Hasta Kadın 57 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
120 Hasta Kadın 44 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
121 Hasta Erkek 42 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
122 Hasta Kadın 78 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
123 Hasta Erkek 61 AA Hafif Valvüloplasti Replasman Yok Profilaksi Yok

68
Yok
124 Hasta Kadın 34 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
125 Hasta Kadın 77 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
126 Hasta Kadın 40 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
127 Hasta Kadın 52 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
128 Hasta Kadın 58 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
129 Hasta Kadın 38 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
130 Hasta Kadın 51 AA Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
131 Hasta Kadın 40 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
132 Hasta Kadın 46 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
133 Hasta Kadın 44 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
134 Hasta Kadın 21 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
135 Hasta Kadın 55 TT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
136 Hasta Kadın 24 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
137 Hasta Kadın 33 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
138 Hasta Kadın 36 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
139 Hasta Kadın 49 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
140 Hasta Kadın 52 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
141 Hasta Kadın 65 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
142 Hasta Erkek 57 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
143 Hasta Kadın 47 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Var
144 Hasta Erkek 49 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
145 Hasta Kadın 31 AT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
146 Hasta Kadın 52 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
147 Hasta Kadın 34 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok

69
148 Hasta Erkek 64 AA Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
149 Hasta Kadın 44 TT Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
150 Hasta Kadın 31 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
151 Hasta Kadın 40 AA Hafif
Valvüloplasti
Yok Replasman Yok Profilaksi Yok
152 Hasta Kadın 32 AT Şiddetli
Valvüloplasti
Yok Replasman Yok Profilaksi Var
153 Kontrol Kadın 66 AA . . . .
154 Kontrol Kadın 64 AT . . . .
155 Kontrol Kadın 65 AT . . . .
156 Kontrol Kadın 43 AT . . . .
157 Kontrol Kadın 45 AT . . . .
158 Kontrol Kadın 29 AT . . . .
159 Kontrol Kadın 49 AA . . . .
160 Kontrol Kadın 40 AT . . . .
161 Kontrol Kadın 56 AT . . . .
162 Kontrol Kadın 60 AT . . . .
163 Kontrol Kadın 65 AA . . . .
164 Kontrol Kadın 49 AA . . . .
165 Kontrol Kadın 28 AA . . . .
166 Kontrol Kadın 62 AT . . . .
167 Kontrol Kadın 64 TT . . . .
168 Kontrol Kadın 48 TT . . . .
169 Kontrol Kadın 50 AT . . . .
170 Kontrol Kadın 67 TT . . . .
171 Kontrol Kadın 72 AT . . . .
172 Kontrol Erkek 40 AT . . . .
173 Kontrol Kadın 31 AA . . . .
174 Kontrol Kadın 51 AT . . . .
175 Kontrol Kadın 33 AT . . . .
176 Kontrol Kadın 22 AA . . . .
177 Kontrol Kadın 52 AT . . . .
178 Kontrol Kadın 28 AA . . . .
179 Kontrol Kadın 60 AT . . . .
180 Kontrol Kadın 67 TT . . . .
181 Kontrol Kadın 35 AT . . . .
182 Kontrol Erkek 61 AA . . . .
183 Kontrol Kadın 46 AA . . . .
184 Kontrol Erkek 44 TT . . . .
185 Kontrol Erkek 37 TT . . . .
186 Kontrol Kadın 45 AT . . . .
187 Kontrol Erkek 50 AT . . . .
188 Kontrol Kadın 53 AT . . . .
189 Kontrol Kadın 46 AA . . . .
190 Kontrol Kadın 53 AT . . . .
191 Kontrol Kadın 64 AT . . . .
70
192 Kontrol Kadın 62 AT . . . .
193 Kontrol Erkek 40 AT . . . .
194 Kontrol Kadın 50 TT . . . .
195 Kontrol Kadın 29 AA . . . .
196 Kontrol Kadın 60 AT . . . .
197 Kontrol Kadın 31 AA . . . .
198 Kontrol Kadın 34 AA . . . .
199 Kontrol Erkek 73 AA . . . .
200 Kontrol Erkek 53 AA . . . .
201 Kontrol Kadın 73 AT . . . .
202 Kontrol Kadın 55 TT . . . .
203 Kontrol Kadın 52 AA . . . .
204 Kontrol Kadın 40 AA . . . .
205 Kontrol Erkek 55 AT . . . .
206 Kontrol Kadın 50 AT . . . .
207 Kontrol Erkek 69 AA . . . .
208 Kontrol Kadın 45 AT . . . .
209 Kontrol Kadın 63 AT . . . .
210 Kontrol Kadın 60 AT . . . .
211 Kontrol Kadın 59 AA . . . .
212 Kontrol Kadın 67 AA . . . .
213 Kontrol Kadın 50 AT . . . .
214 Kontrol Kadın 59 AT . . . .
215 Kontrol Kadın 49 AT . . . .
216 Kontrol Erkek 40 AA . . . .
217 Kontrol Kadın . AT . . . .
218 Kontrol Erkek 53 TT . . . .
219 Kontrol Kadın 38 AT . . . .
220 Kontrol Kadın 37 AT . . . .
221 Kontrol Kadın 50 AT . . . .
222 Kontrol Erkek 81 TT . . . .
223 Kontrol Erkek 38 TT . . . .
224 Kontrol Erkek 48 AT . . . .
225 Kontrol Kadın 56 AT . . . .
226 Kontrol Kadın 27 AT . . . .
227 Kontrol Erkek 56 AA . . . .
228 Kontrol Kadın 40 AA . . . .
229 Kontrol Kadın 57 AT . . . .
230 Kontrol Erkek 44 AA . . . .
231 Kontrol Erkek 60 AT . . . .
232 Kontrol Erkek 58 TT . . . .
233 Kontrol Kadın 70 AA . . . .
234 Kontrol Kadın 53 AT . . . .
235 Kontrol Kadın 30 AT . . . .
236 Kontrol Kadın 42 AA . . . .
237 Kontrol Kadın 39 AT . . . .
238 Kontrol Erkek 54 AA . . . .
239 Kontrol Kadın 52 AT . . . .
240 Kontrol Kadın 58 AT . . . .
71
241 Kontrol Kadın 48 AA . . . .
242 Kontrol Kadın 52 AT . . . .
243 Kontrol Erkek 56 AA . . . .
244 Kontrol Kadın 65 AA . . . .
245 Kontrol Kadın 26 TT . . . .
246 Kontrol Kadın 72 AA . . . .
247 Kontrol Kadın 57 AA . . . .
248 Kontrol Kadın 40 AA . . . .
249 Kontrol Kadın 63 AA . . . .
250 Kontrol Kadın 58 AA . . . .
251 Kontrol Erkek 70 AT . . . .
252 Kontrol Kadın 81 TT . . . .
253 Kontrol Kadın 57 AA . . . .
254 Kontrol Kadın 57 AT . . . .
255 Kontrol Kadın 40 AT . . . .
256 Kontrol Kadın 45 AT . . . .
257 Kontrol Kadın 58 AA . . . .
258 Kontrol Kadın 50 AT . . . .
259 Kontrol Kadın 30 AT . . . .
260 Kontrol Kadın 47 AT . . . .
261 Kontrol Kadın 25 AT . . . .
262 Kontrol Kadın 58 TT . . . .
263 Kontrol Kadın 50 AT . . . .
264 Kontrol Kadın 47 AT . . . .
265 Kontrol Kadın 40 AA . . . .
266 Kontrol Kadın 28 AT . . . .
267 Kontrol Kadın 27 AT . . . .
268 Kontrol Kadın 53 AT . . . .
269 Kontrol Kadın 50 AT . . . .
270 Kontrol Kadın 31 AA . . . .
271 Kontrol Kadın 54 AT . . . .
272 Kontrol Kadın 43 AT . . . .
273 Kontrol Erkek 44 AT . . . .
274 Kontrol Kadın 78 AT . . . .
275 Kontrol Erkek 61 AT . . . .
276 Kontrol Kadın 34 AT . . . .
277 Kontrol Kadın 77 AA . . . .
278 Kontrol Kadın 40 AT . . . .
279 Kontrol Kadın 50 AT . . . .
280 Kontrol Kadın 49 AT . . . .
281 Kontrol Kadın 41 AT . . . .
282 Kontrol Kadın 50 AT . . . .
283 Kontrol Kadın 41 AT . . . .
284 Kontrol Kadın 45 AA . . . .
285 Kontrol Kadın 44 AT . . . .
286 Kontrol Kadın 21 AT . . . .
287 Kontrol Kadın 55 AT . . . .
288 Kontrol Kadın 25 TT . . . .
289 Kontrol Kadın 33 AT . . . .

72
290 Kontrol Kadın 42 AA . . . .
291 Kontrol Kadın 48 AT . . . .
292 Kontrol Kadın 49 AT . . . .
293 Kontrol Kadın 48 AT . . . .
294 Kontrol Erkek 57 AT . . . .
295 Kontrol Kadın 48 AT . . . .
296 Kontrol Erkek 43 AA . . . .
297 Kontrol Kadın 44 AT . . . .
298 Kontrol Kadın 48 AA . . . .
299 Kontrol Kadın 42 AT . . . .
300 Kontrol Erkek 64 AT . . . .
301 Kontrol Kadın 31 AA . . . .
302 Kontrol Kadın 47 AA . . . .
303 Kontrol Kadın 32 AT . . . .

73
FORMLAR

74
ETİK KURUL KARARI

75
PATENT HAKKI İZNİ

76
TELİF HAKKI İZNİ

77
ÖZGEÇMİŞ
Kişisel Bilgiler
Adı Erhan Soyadı TEKER
Doğ.Yeri Giresun Doğ.Tar. 19/ 03/ 1981
Uyruğu TC TC Kim No 11231289850
Email [email protected] Tel 0535 569 25 33
Eğitim Düzeyi
Mezun olduğu okullar:
1987-1992 Giresun Cumhuriyet İlkokulu
1992-1995 Giresun Kanuni Sultan Süleyman Orta Okulu
1995-1998 Giresun Lisesi
2000-2005 İstanbul Üniversitesi Cerrahpaşa Tıp Fakültesi
Yabancı Dilleri
Okuduğunu Anlama*
Konuşma* Yazma* KPDS/ÜDS
Puanı (Diğer) Puanı
İngilizce Çok iyi iyi orta
*Çok iyi, iyi, orta, zayıf olarak değerlendirin
Yayınları/Tebligleri Sertifikaları/Ödülleri
Semptomsuz miyokard iskemisi veya infarktüsünün erken tanısında ve
tedavisinin yönlendirilmesinde çok kesitli bilgisayarlı tomografik koroner
anjiografinin yeri ve önemi




















