
Semantic Work Process Analysis – A Reflexive Stakeholder Articulation Approach

Taphonomic biases in the insect fossil record: shifts in articulationover geologic time
Jered A. Karr and Matthew E. Clapham
Abstract.—Insect taphonomy is a topic that has drawn interest because of its potential biases ondiversity patterns and the ecological information recorded by ancient insect faunas. Other than theonset of common amber fossilization in the Cretaceous, very little is known about long-term trends inthe nature and quality of insect preservation and, as a result, the effects of taphonomic biases are poorlyconstrained. We assembled a database of nearly 7000 Carboniferous–Pliocene insect adpression(compression and impression) species from the primary literature to assess changes in insecttaphonomy over time and test biotic and environmental controls on preservation. We grouped thefossils into 10-Myr bins and scored preservation of holotype specimens as either articulated bodies orisolated wings; articulated specimens with a body implied a generally higher quality of preservation.Paleozoic and Triassic insect holotypes are known overwhelmingly from isolated wings (only 12%articulated bodies), but our database shows a significant increase in the percentage preserved asarticulated bodies, to more than 70%, beginning about 160 Myr ago in the Late Jurassic. This transitioncould reflect variations in the robustness of different insect orders and shifts in the taxonomiccomposition of insect faunas, but all the major orders in the database exhibit significant increases inarticulation. Instead, a shift to increased preservation in lacustrine paleoenvironments, which contain agreater proportion of articulated body fossils, explains most of the trend. The pronounced Late Jurassicincrease in articulation has implications for evolutionary and ecological reconstructions, for example,suggesting that preserved insect diversity may be biased downward in the earlier part of their historywhen articulation was poor.
Jered A. Karr and Matthew E. Clapham.* Department of Earth and Planetary Sciences, University of California,Santa Cruz, California 95064, U.S.A. E-mail: [email protected], [email protected]
* Corresponding authorAccepted: 26 May 2014Supplemental materials deposited at Dryad: doi: 10.5061/dryad.p28c8
Introduction
Insects are the most diverse and successfulanimal group today (Gaston 1991) but under-standing their past evolutionary patterns,diversity, and ecology may be hindered bypreservational biases. Taphonomic processescan change our view of ecologic roles andimportance or can obscure taxonomically-important characters and evolutionary rela-tionships among groups (e.g., the case ofProtorthoptera [Bethoux 2005, 2007]). Laban-deira and Sepkoski (1993) note that theirdiversity curve is likely biased by exceptionalTertiary deposits such as Baltic amber andFlorissant, so understanding insect preserva-tion can also help us elucidate their true pastdiversity trends.
Although their fossil record is richer thangenerally assumed (Labandeira and Sepkoski
1993; Rasnitsyn and Quicke 2002; Grimaldi
and Engel 2005), insects lack mineralized
tissue and their preservation usually requires
extraordinary circumstances, commonly in
Konservat-Lagerstatten. These insect Konser-
vat-Lagerstatten come from varied environ-
ments, particularly lacustrine settings and
amber, but also shallow-marine or lagoonal
deposits and small ponds (Labandeira 1999),
each of which may have been influenced by a
variety of biases. Because amber preservation
became common only in the Early Cretaceous,
has a more ephemeral record than lacustrine
deposits (Labandeira 1999), and was influ-
enced by a different set of taphonomic biases
(Zherikhin et al. 1999; Martınez-Delclos et al.
2004), we will focus only on insects preserved
as adpressions (compressions and impres-
sions) in sedimentary rocks.
� 2015 The Paleontological Society. All rights reserved. 0094-8373/15
Paleobiology, 41(1), 2015, pp. 16–32doi: 10.1017/pab.2014.3

Smith (2012) divided the factors that controlpreservation quality into ‘‘insect inputs’’ and‘‘depositional factors.’’ Factors such as insectsize, morphology, and taxonomic group (‘‘in-sect input’’ variables), or environment type,bathymetry, and energy levels (‘‘depositionalfactors’’) influence if and how well an insect ispreserved in the fossil record, so temporaltrends in their importance may cause system-atic biases in preservation quality. Maximuminsect size has broadly decreased since the latePaleozoic (Clapham and Karr 2012), whiletaxonomic changes in insect assemblages,such as radiations of groups with highpreservation potential like beetles (e.g., Lab-andeira and Sepkoski 1993) might also haveinfluenced levels of preservation. The domi-nant depositional environment of insect Kon-servat-Lagerstatten has also changed, fromCarboniferous delta plain Lagerstatten toEarly Jurassic shallow marine and Creta-ceous–Cenozoic lacustrine deposits (Allisonand Briggs 1991; Briggs 2003), but it isunknown how this shift has affected preser-vation of all animal groups and in particularinsects.
Many studies have assessed the factors thatinfluence the quality of insect preservation,but these studies have largely focused ondetailed taphonomic investigation of singlelocalities with specific environmental condi-tions (Wilson 1980, 1988; McCobb et al. 1998;Ansorge 2003; Coram 2003; Wappler 2003;Henning et al. 2012). We examined long-termtrends in the articulation of insect fossils,using a large database of specimen datacompiled from the primary literature. Al-though insect preservation quality is multifac-eted, we used articulation as a proxy for decaybecause experiments examining the variousstages of insect decomposition found thatisolated wings, even with minute details, oftenrepresent one of the final stages of decompo-sition (Duncan et al. 2003). Using a large dataset of fossil insect localities allows us to poolmultiple sites, removing idiosyncratic, loca-tion-specific taphonomic signals to revealoverarching trends. We tested the effects ofdepositional environment, insect size, mor-phology/taxonomic group, and geologic ageon insect articulation. Understanding the
overall biases in the fossil record of insectswill allow assessment of the ecology andevolution of this group across geologic timeand comparison with trends observed in othergroups of animals and plants.
Insect Taphonomy
In order to use insect fossil assemblages forevolutionary, ecological, or diversity studies, itis important to understand the steps fromliving insect to fossil to published literature.The rate at which an insect passes throughthese steps influences the quality of preserva-tion by altering the time exposed to predation,decomposition, and disarticulation. An insectmust pass through four taphonomic steps,which act as filters, before being preserved asan adpression in the fossil record. First, thebody must be transported to an aquaticenvironment by rafting or aerial deposition(except in the case of aquatic insects, whichalready inhabit the environment). Most insectsare not aquatic so they must next break thesurface tension of water. Martınez-Delclos andMartinell (1993) examined insect deaths inaquatic environments of a variety of taxonom-ic groups and found a large range of outcomesdepending on size and weight, described bythe SM (wing surface area-to-mass) index(Wagner et al. 1996), and wing type or shape.There tends to be a bias against large-wingedor fragile groups such as Lepidoptera andagainst insects with wingspans smaller than 5mm, typically leading to enrichment in heavy,medium-sized insects at the sediment inter-face relative to their life abundance. Archibaldand Makarkin (2006) and Wang et al. (2013)showed that insects with larger SM indices aremore prone to disarticulation, likely due tolonger floating time at the water surface.Larger wings commonly have microstructuresthat influence the wettability of the wings andcan increase floating time as well (Wagner etal. 1996; Rust 1998; Archibald and Makarkin2006). After breaking the water surface, thethird step requires the insect to sink throughthe water column. Temperature and chemicalcomposition affect the rate of sinking, and adensity change at the thermocline (in deeperlakes or the ocean) or halocline (in somesaltwater environments) can prevent sinking,
17TRENDS IN INSECT ARTICULATION

thus promoting decomposition before carcass-es can be deposited on the sediment(Martınez-Delclos et al., 2004). The final stepis burial after the insect settles onto thesediment, influenced by the distance fromshore, water chemistry, and tectonic setting. Instudies of Eocene lakes from British Columbia,Wilson (1980, 1988) noted a trend for morearticulated specimens in offshore compared tonearshore sediment. In another study, Briggset al. (1998) found that insects from deep-water anoxic zones had 95–98% more chitinpreserved than insects from the shalloweroxygenated part in Pliocene lake sedimentsfrom Willershausen, Germany.
After being preserved as a fossil the insectmust still be collected and described, whichmay impart a different set of biases. Due todifferences in sampling intensity and theemphasis of the collector, widely differentratios of taxonomic groups can be collectedat the same locality (Sukacheva and Rasnitsyn2004). The final step after being collected isdescription and entering the published litera-ture, which generates a strong bias towardbetter-preserved material with more diagnos-tic characters that allow the specimen to bereadily identified.
Methods
Using primary literature data from 1560published papers (see supplementary infor-mation), we quantified the articulation, size,taxonomic identity, and depositional environ-ment of Carboniferous-Pliocene insects pre-served as adpression fossils. All collectionsand occurrences, taxonomic names, and sizemeasurements used in this study are housedin the Paleobiology Database (PaleoDB:http://paleobiodb.org); the data used herewere downloaded on 12 May 2014. We scoredeach species on the basis of the holotypespecimen and categorized it as either anexoskeleton (‘‘articulated’’) or wing element(‘‘disarticulated’’). Because holotypes are al-ways more likely to be articulated than thefossil assemblage as a whole, our absolutevalues of articulation will overestimate thearticulation in the entire insect assemblage butthe relative shifts will be robust. All wingelements (elytron, tegmen, forewing, hind-
wing, or wing) were grouped together, where-as an exoskeleton includes any specimen withboth the body and wings. In many cases thewings and/or body are incomplete or frag-mentary, but we did not distinguish fragmen-tation in our analyses. We excluded all specieswhere the holotype body part was unidenti-fied or a nymph/larva and all occurrences ofquestionably identified genera and species orthose in open nomenclature. Because we arealso interested in the effect of size onarticulation we also excluded all occurrenceswithout a wing measurement.
For each locality, we assessed the deposi-tional environment to test its effect on articu-lation. The PaleoDB allows specification of awide range of environments for each collec-tion, making analysis unfeasible due to smallsample size in some cases, so similar environ-ments were grouped together into the follow-ing categories: marine, delta/lagoon, HE (highenergy) terrestrial, shallow lakes, and deeplakes. Because we hypothesized that deeplakes have uniquely favorable conditions forinsect fossilization, for most analyses we usethe term deep lake or non-deep lake (com-bined marine, delta/lagoon, HE terrestrial,shallow lakes) to categorize the environment.The deep lake category includes large lakes(‘‘lacustrine – large’’ in the PaleoDB) such asFlorissant (Veach and Meyer 2008), the YixianFormation (Hethke et al. 2013), and LakeGosiute (Green River Formation; Surdamand Stanley 1980). Because we consider depthto be an important factor, the deep lakecategory also contains several localities as-signed to the PaleoDB ‘‘crater lake’’ environ-ment, with smaller surface area but withsignificant depth, such as Messel (Harms2002; Felder and Harms 2004), the EckfeldMaar (Lutz 2003), Menat (Wappler et al. 2009),and the Randeck Maars (Zeuner 1942). Theshallow lake category includes localities re-ferred to as ponds (Engel and Gross 2008) oroxbow lakes like Madygen (Shcherbakov2008).
Size data were also collected to examine itseffect on articulation. The length of the wing,tegmen (Orthoptera and relatives, Blattodeaand relatives, some Hemiptera), elytron (Co-leoptera), or hemelytron (some Hemiptera)
18
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM

was recorded. In groups with two pairs ofwings (fore and hind wings), the larger of thetwo pairs was used. Sizes were taken frommeasurements directly reported in the sys-tematic description or were measured frompublished illustrations if no size was given inthe text. Many fossil wings are incompletebecause of biostratinomic processes, especiallyin the largest insects, so we used estimates ofcomplete wing size provided in the descrip-tion. After taxonomic filtering and removingamber specimens we have 6915 measuredspecies.
We binned the data into 30 time intervalsequivalent to geological stages or sets ofneighboring stages, starting in the mid Car-boniferous. The interval definitions are thePaleoDB 10-Myr bins, the same as those usedin several recent papers (Alroy et al. 2008).Stage-level time intervals were not usedbecause of the paucity of data and uncertainage assignment of many localities.
Model Construction
We performed multiple logistic regressionanalyses in R (R Development Core Team2012) to estimate the effect of morphology,depositional environment, and size on theprobability of articulation. We log10 trans-formed wing size (measured in mm) tonormalize the size distribution. The morphol-ogy predictor was coded as a binomial factor(beetle or non-beetle) for simplicity; eventhough articulation varies within and amongorders, beetles have the greatest preservationpotential (Smith 2000). It is also difficult, apriori, to predict or assign numerical values tothe articulation of each insect order, but ifmorphology is an important predictor ofarticulation the comparison of beetles to otherinsects should show this. We coded theenvironment predictor as a binomial factoras well, based on whether the specimen camefrom a ‘‘deep lake’’ or a ‘‘non-deep lake’’depositional environment. As with morphol-ogy, it is not possible to transform theenvironmental categories into an orderednumerical variable and there are multiplereasons to believe that deep lakes should havesuperior fossil preservation compared to otherdepositional environments. Because we want-
ed to keep the results interpretable, wenarrowed our pool of models to only combi-nations of these three variables and did notinclude interaction effects. We used Akaikeinformation criterion (AIC) to determine mod-el selection.
We also analyzed the effects of environmentand size on six common clades: Odonatoptera(dragonflies, damselflies and extinct relatives),Blattodea (cockroaches, excluding termites),Orthoptera (grasshoppers and crickets), He-miptera (bugs), Diptera (flies), and Coleoptera(beetles) with separate multiple logistic re-gression models for each group. Finally, weexamined the effect of size alone by perform-ing logistic regression models, on all insectsand on the same six common groups, withsize as the only predictor.
Using the magnitude of the odds ratios fordirect comparison of the relative importanceof all three predictor variables is not possiblebecause ‘‘environment’’ and ‘‘morphology’’ areboth binomial predictors whereas ‘‘size’’ iscontinuous. We converted size to a binomialpredictor comparable to environment andmorphology by using the mean log10 size forthe Cenozoic as ‘‘small’’ (0.83) and the meanfor the Carboniferous as ‘‘large’’ (1.54). Wethen predicted the probability of articulationthrough geologic time on the basis of the eightpossible combinations of the now binomialpredictor variables (size, morphology, envi-ronment). We compared each combinationwith the overall shift in articulation. To dothat, we created a synthetic data set in which‘‘size’’ was replaced with values rangingcontinuously from 1.54 in the oldest bin to0.83 in the youngest bin. ‘‘Environment’’ wasreplaced with values ranging from 0.0 to 0.82(the proportion of insect occurrences occurringin deep lakes in the Carboniferous andCenozoic, respectively) and ‘‘morphology’’was replaced with values from 0.0 to 0.26(the proportion of insect occurrences that areColeoptera in the Carboniferous and Cenozo-ic). We also compared actual variations inarticulation with predicted articulation pro-portion, using the relationship derived fromoverall logistic regression model and theenvironment (proportion deep lake), morphol-
19TRENDS IN INSECT ARTICULATION

ogy (proportion beetle), and size (mean size)data for each time bin.
All data files and R scripts for data analysisare available at https://github.com/mclapham/insect_taph.
Results
The proportion of articulated holotypesincreased significantly through time. Beforethe Late Jurassic (Jurassic bin 5, Callovian-Kimmeridgian, ca. 161 Ma), 88.8% of insectholotypes are disarticulated wing elements,whereas 73.4% of holotypes in the LateJurassic and after are preserved as articulatedbodies (Fig. 1). In a comparison of multiplelogistic regression models, the model includ-ing all three variables (environment, morphol-ogy, and size) was the best-supported (Table1), indicating that morphology, environment,and size all significantly affected insect artic-ulation (p , 0.001 for all variables; Table 2).While holding other parameters constant,
beetles were more articulated than insects as
a whole, insects in deep lake environments
were significantly more likely to be articulat-
ed, and specimens with larger wing size were
less likely to be articulated (Figs. 2–4).
Morphological Controls
Because higher-level taxonomic groupings
typically represent distinctive body plans with
FIGURE 1. Phanerozoic trend in insect articulation, based on the proportion of occurrences of species with articulatedholotypes in each 10-Myr bin. Bubble size scales with the number of occurrences in each 10-Myr bin (maximum 1061).Dashed line represents the beginning of the Callovian (Jurassic bin 5).
TABLE 1. Support for models explaining change inarticulation ordered from best supported to leastsupported by Akaike information criterion.
Models AIC D AIC
Morphologyþsizeþenvironment 6366.52 0
Sizeþenvironment 6440.27 73.75Morphologyþenvironment 7110.85 744.33
Environment 7298.21 931.69Morphologyþsize 8511.83 2145.30Size 8656.44 2289.92Morphology 9644.58 3278.06
20
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM
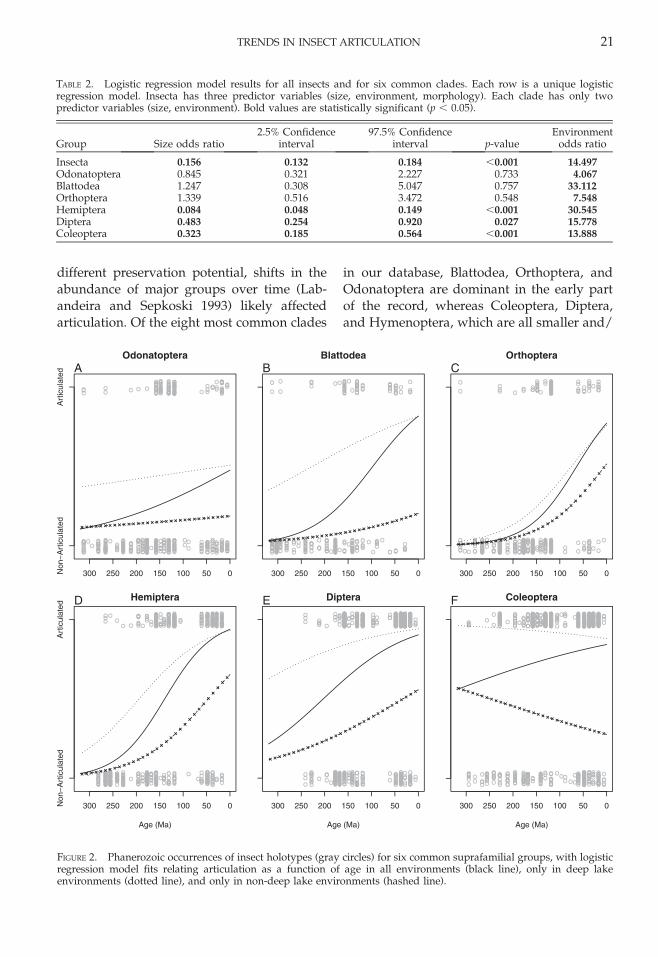
different preservation potential, shifts in theabundance of major groups over time (Lab-andeira and Sepkoski 1993) likely affectedarticulation. Of the eight most common clades
in our database, Blattodea, Orthoptera, andOdonatoptera are dominant in the early partof the record, whereas Coleoptera, Diptera,and Hymenoptera, which are all smaller and/
TABLE 2. Logistic regression model results for all insects and for six common clades. Each row is a unique logisticregression model. Insecta has three predictor variables (size, environment, morphology). Each clade has only twopredictor variables (size, environment). Bold values are statistically significant (p , 0.05).
Group Size odds ratio2.5% Confidence
interval97.5% Confidence
interval p-valueEnvironment
odds ratio
Insecta 0.156 0.132 0.184 ,0.001 14.497Odonatoptera 0.845 0.321 2.227 0.733 4.067Blattodea 1.247 0.308 5.047 0.757 33.112Orthoptera 1.339 0.516 3.472 0.548 7.548Hemiptera 0.084 0.048 0.149 ,0.001 30.545Diptera 0.483 0.254 0.920 0.027 15.778Coleoptera 0.323 0.185 0.564 ,0.001 13.888
FIGURE 2. Phanerozoic occurrences of insect holotypes (gray circles) for six common suprafamilial groups, with logisticregression model fits relating articulation as a function of age in all environments (black line), only in deep lakeenvironments (dotted line), and only in non-deep lake environments (hashed line).
21TRENDS IN INSECT ARTICULATION

or more robust taxa commonly requiring bodyfeatures for identification, become dominantin the late Mesozoic and Cenozoic (Laban-deira and Sepkoski 1993). This shift likelycontributed to increased articulation in youn-ger collections, but was not the principalfactor because all well-sampled taxonomicgroups independently increase in articulationthrough time (Fig. 2).
Odonatoptera.—Odonatoptera (total-groupOdonata and extinct relatives like Meganisop-tera) includes large insects with high SM indexwings that are spread away from the body atrest (except in Zygoptera). Their wing vena-tion is complex and distinctive for taxonomicidentification. Only 26.9% of Odonatoptera
species have been described from articulatedbodies, but overall articulation increasedsignificantly in younger collections. Articula-tion did not change significantly withinenvironments (Fig. 2A).
Blattodea.—Blattodea (cockroaches, exclud-ing termites in our analyses) have forewingsthat are hardened into tegmina, which aremore resistant to degradation than bodies orhindwings. Blattodea holotypes are mostcommonly an isolated tegmen in Paleozoicdeposits, but in the Mesozoic and Cenozoicthey are more commonly articulated bodies(Fig. 2B). The high diversity of Paleozoicspecies influences the mean percent articula-tion, and only 22.6% of species have been
TABLE 2. Extended.
2.5% Confidenceinterval
97.5% Confidenceinterval p-value
Morphologyodds ratio
2.5% Confidenceinterval
97.5% Confidenceinterval p-value
12.783 16.442 ,0.001 2.186 1.826 2.617 ,0.0012.638 6.270 ,0.001
15.875 69.067 ,0.0014.387 12.984 ,0.001
19.678 47.414 ,0.00111.241 22.147 ,0.00110.043 19.206 ,0.001
FIGURE 3. Phanerozoic insect holotype occurrences (gray circles) with logistic regression model fits relating articulationas a function of age in all environments (black line), and in only deep lake environments (dotted line), and in only non-deep lake environments (hashed line). Points representing articulated and non-articulated insect specimens have beenrandomly jittered for visibility.
22
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM

described from articulated bodies. As inOdonatoptera, articulation did not changesignificantly over time within environments.
Orthoptera.—Orthoptera (crickets and grass-hoppers) can be quite large (the suborderTitanoptera reached wing lengths of 180 mmduring the Triassic) and have sclerotizedforewings like Blattodea. Overall articulationis even lower than in Blattodea (15.8% ofspecies holotypes are articulated) and in-creased significantly in younger deposits.Within-environment trends are more difficultto reconstruct because there are no deep lakeoccurrences of Orthoptera before the Jurassic,but articulation likely increased significantlyboth in non-deep lakes and in deep lakes (Fig.2C).
Hemiptera.—Hemiptera (true bugs) are onethe most heterogeneous orders, with somefamilies consisting of small robust speciessimilar to Coleoptera (such as some Heterop-tera) and other families consisting of large-winged fragile groups more similar to Odo-natoptera. Overall articulation increased con-siderably, with some contribution from theshift to deep lake preservation, but with alarger contribution from changes in articula-tion within both deep lakes and non-deeplakes (Fig. 2D).
Diptera.—Diptera (flies) are typically smallinsects with simple wing venation, makingdescription from isolated wings challenging.They are generally so small that it can bedifficult for them to break surface tension(Martınez-Delclos and Martinell 1993), butthey are well articulated (75.5% of speciesdescribed from complete bodies). Articulationincreased slightly in both non-deep lakes anddeep lakes. Increased preservation in deeplakes accounts for a greater proportion of theoverall trend toward increased articulation inDiptera (Fig. 2E).
Coleoptera.—Coleoptera (beetles) are wellarticulated (77% of species holotypes arecomplete bodies) and increase in abundancetoward the modern (7.1% of occurrencesbefore the Late Jurassic and 22.1% after inour database). Although overall articulationincreased significantly, due to the shift towardpreservation in deep lakes, within-environ-ment articulation apparently decreased bothin deep lakes and, to a greater degree, in non-deep lakes (Fig. 2F).
Environment
The predicted probability of a specimenbeing articulated increased through time inboth non-deep lake localities from ~10% to
FIGURE 4. Occurrences of insect holotypes for all sizes (gray circles) with a logistic regression model fit relatingarticulation as a function of log10 wing length.
23TRENDS IN INSECT ARTICULATION

~50%, and from ~45% to ~90% in deep lakeenvironments, but preservation in deep lakessignificantly increased the likelihood of artic-ulation in all six clades (Table 2, Fig. 3). Deeplake environments also became more commonover time: only 6% of pre-Late Jurassic insectoccurrences are from deep lakes, comparedwith 79.4% of younger occurrences. Thetaxonomic composition of deep lake andnon-deep lake assemblages likely also influ-enced articulation. Orthoptera and Odonatop-tera, which have low articulation, aregenerally strong fliers and are nearly equallycommon in the three main environments,including marine settings far from theiroriginal habitats. In contrast, Coleoptera,which commonly are articulated, are generallypoor fliers and are found in much higherpercentages (73%) in deep lakes (Table 3).
Wing Size
Articulation has a clear negative relation-ship with wing length in our database (Fig. 4).Maximum insect size has decreased over time(Clapham and Karr 2012) but, because othermorphological factors also vary among orders,it is important to look at the trends withineach clade to assess the effects of size alone. Ofthe six best-sampled clades only Hemipteraand Coleoptera have a strong negative rela-tionship between size and articulation (Table2, Fig. 5). Larger Diptera are significantly lesslikely to be articulated after accounting forenvironment of preservation (Table 2). Oddsratios in Orthoptera and Blattodea indicatethat increases in size are positively (althoughnot significantly) associated with increasedarticulation, whereas Odonatoptera exhibit nosignificant relationship (Table 2). There is astrong trend of clades with large wing size(and likely body size) being less articulatedthan clades with small wing size, implying
that intra-clade decreases in size had a smallereffect on articulation than the replacement oflarger-bodied clades by typically smaller ones.
Relative Importance of Size, Morphology, andEnvironment
The odds of an insect preserved in a deeplake being articulated are 1350% higher thanthe odds for an insect in a non-deep lakeenvironment. By comparison, the odds ofarticulation for beetles are nearly 119% higherthan the odds for non-beetles, implying thatchanges in environment have a greater effecton insect articulation than differences in insectmorphology. For every tenfold increase in sizethe odds of articulation decrease by nearly85%. Deep lake environments (bars in columnA of Fig. 6) consistently have greater articu-lation, regardless of taxonomy or size, imply-ing that although size and taxon group affectarticulation, environment is the largest con-tributor.
We modeled the effects of environment,size, and morphology by creating a simulateddata set and compared the results to theobserved trends (Fig. 7). Using our logisticregression model on this idealized databasewe were able to closely simulate the change inarticulation over time in most time bins.Articulation predicted only from the propor-tion of deep-lake occurrences, the proportionof Coleoptera occurrences, and the averagesize does not always correspond to actualvalues, most notably during some time binsdominated by deltaic or lagoonal deposits(e.g., Solnhofen, ca. 150 Ma). Although everylocality has a unique set of taphonomiccharacteristics, the good agreement betweenactual and predicted articulation implies thatchanges in depositional environment, beetleabundance, and size can explain much of the
TABLE 3. Proportion of holotype occurrences per paleoenvironment group for six common insect clades.
Environment Deep lake Shallow lake HE terrestrial Delta/coastal Marine
Odonatoptera 0.39 0.21 0.04 0.21 0.15Blattodea 0.28 0.28 0.02 0.28 0.14Orthoptera 0.32 0.28 0.03 0.16 0.22Hemiptera 0.55 0.17 0.02 0.15 0.11Diptera 0.74 0.09 0.06 0.04 0.07Coleoptera 0.73 0.14 0.03 0.05 0.05
24
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM

variation in insect articulation in the fossil
record.
Discussion
Morphological Controls
Insects are a very disparate group with
major morphological differences among or-
ders. This structural variability likely influ-
ences overall articulation, due to changes in
the relative diversity of insect orders through
time (Labandeira and Sepkoski 1993). In
addition, articulation proportions based on
published holotypes depend not only on
taphonomic biases during fossilization (relat-
ed to robustness, size, and SM index), but also
on conventional taxonomic practices for spe-
cies description in a particular group.
Three general patterns of articulation over
time are evident among the six suprafamilial
groups we analyzed: (1) Each environment
had a consistent level of articulation over time,
but overall articulation increased (observed in
Odonatoptera and to some extent in Blatto-
dea). (2) Overall articulation increased, as did
articulation in both deep lake and non-deep
lake environments (observed in Hemiptera,
Orthoptera and Diptera). (3) Articulation
decreased in both of these environments but
overall articulation increased (observed only
in Coleoptera).
FIGURE 5. Occurrences of insect holotypes for all sizes (gray circles) for six common suprafamilial groups with logisticregression model fit relating articulation as a function of log10 wing length in all environments (black line), in only deeplake environments (dotted line), and in only non-deep lake environments (hashed line).
25TRENDS IN INSECT ARTICULATION

Consistent within-environment articulationtrends in Odonatoptera (and Blattodea) arebest explained by the conservative morpholo-gy of those groups. Because their overall bodyplan has changed little over their evolutionaryhistory, taphonomic controls on articulationwere similar in the Carboniferous or theCenozoic. Both groups are more consistentlyarticulated in deep lakes, and the overallincrease in articulation resulted from thepost-Jurassic increase in the number of deeplake deposits (Fig. 2A,B).
A shift to greater preservation in deep lakesalso significantly enhanced articulation inHemiptera, Orthoptera, and Diptera (Fig.2C–E), but in addition those three groupsexhibited considerable within-environmentincreases in articulation. Changing articula-tion in deep lakes and non-deep lakes likelyreflects evolutionary trends within the cladesthat resulted in morphologically distinct fam-
ilies changing in proportional abundances. Forexample, Heteroptera, a suborder withinHemiptera that often contains smaller, beetle-like species with folded tegmina and lowerSM indices, is in general more articulated, andconstitutes a larger proportion of occurrenceslater in the fossil record, than ‘‘Homoptera,’’ aparaphyletic assemblage of generally large-winged and poorly sclerotized Hemiptera.Similarly, Cenozoic Orthoptera are largelycomposed of Acrididae (short-horned grass-hoppers) and Tettigoniidae (katydids), twolineages that been observed in actualisticexperiments to have an unusual response todrowning, which might increase sinking rate(Martınez-Delclos and Martinell 1993). Extinctlineages may not have had this response.There is also a trend of wing reduction inOrthoptera (Sharov 1968), which would de-crease their SM index and promote sinking.The patterns in Diptera possibly reflect a shift
FIGURE 6. Occurrences of insect holotypes in the Phanerozoic with logistic regression model fits relating articulation as afunction of tested parameters. Dashed lines represent different regression models holding different combinations of thethree predictors (environment, morphology, size) constant. Bars indicate deep lake environments in column A, beetles incolumn B, and small size in column C. The thick line represents the predicted articulation where values of the threepredictors are simulated to represent the actual fossil record as they change through time. Size changes from 1.54 to 0.83(log10 mm), morphology (proportion of beetles) changes from 0.0 to 0.26, and environment (proportion of deep lakes)changes from 0.0 to 0.82.
26
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM

from more fragile, gracile dipteran groupssuch as Tipulomorpha to more stout-bodied,robust groups such as Muscomorpha.
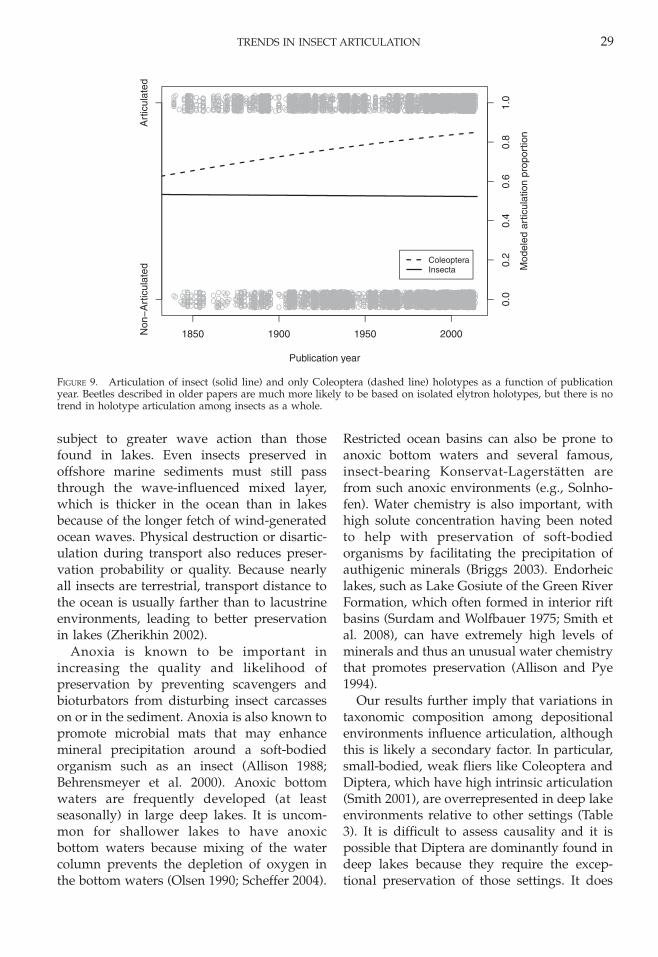
Coleoptera have a higher preservationpotential than other insect orders in bothmodern, actualistic studies and lab experi-ments (Smith 2000; Smith et al. 2006),primarily because their forewings have beenhighly modified into hard sheaths calledelytra. Our results support these findings, asColeoptera is one of the most highly articu-lated groups in our data. In contrast to theother five well-sampled clades, Coleopteraexhibit a highly unusual pattern where theprobability of articulation actually decreasedboth in deep lakes and, especially, in non-deep lakes (Fig. 2F). This trend does notreflect changes in beetle body plans, whichhave remained stable over most of theirevolutionary history, or size, which also hasnot changed substantially. Instead, decreasedarticulation is most likely an artifact oftaxonomic practices. Earlier paleoentomolo-gists, especially in the nineteenth century,described many species based on isolatedelytra from Cenozoic localities. Mesozoic and
Paleozoic isolated elytra typically have littletaxonomic value (even at the family orsuborder level) and are rarely named bymodern paleoentomologists (Arnoldi et al.1977; Ponomarenko 2002), often being placedin morphotypes instead (Papier et al. 2005;Martin 2010). More than 81% of beetle speciesdescribed before 1950 were from Cenozoiclocalities, compared to fewer than 12.5% ofspecies described after 1950. Restricting theanalysis to species described after 1950reduces the effects of outdated taxonomicpractices and indicates no significant shift inarticulation within environments (Fig. 8).Articulation of holotypes does not changesignificantly with year of publication in othertaxonomic groups (Fig. 9), indicating thatshifting taxonomic practices only affect Cole-optera (supplementary figure 1). When out-dated species are excluded, Coleoptera havea pattern of articulation similar to that ofother morphologically stable groups (Odona-toptera and Blattodea), in which the shift togreater deep lake preservation primarilycaused the increase in articulated holotypes.
FIGURE 7. Comparison of actual and modeled insect articulation for bins with at least 100 occurrences. Actualarticulation proportions (open circles) are calculated from holotype specimens in each time bin. Modeled articulationproportions (filled circles) are predicted from the logistic regression model using the proportion of deep-lakeoccurrences, proportion of Coleoptera, and average size in each time bin.
27TRENDS IN INSECT ARTICULATION

Wing Size
Changes in the median size of insects
through time could influence articulation and
it is known that the size of insects has changed
over time (Dudley 1998; Okajima 2008; Clap-
ham and Karr 2012). Large insects have
greater wing area relative to body mass on
average (high SM index value) and will float
on the water surface longer than smaller-
winged, heavier-bodied insects (low SM index
value) (Archibald and Makarkin 2006). Larger
insects may also be subjected to more intense
predation (Blanckenhorn 2000; Chown and
Gaston 2010), which might favor their disar-
ticulation and/or removal from the deathassemblage.
The median wing size of insects in ourdatabase during the Carboniferous is morethan 30 mm but decreases to only 7 mmduring the Cenozoic, although this trend is notuniform across all orders. Odonatoptera winglength decreased from a median of 140 mm inthe Carboniferous to a median of 35 mmduring the Cenozoic, whereas Coleopteramedian size remained remarkably stable ataround 5 mm. The overall trend of decreasingmedian wing size seen in insects is influencedby two factors. The first is a shift from large-winged groups such as Palaeodictyoptera andMegasecoptera to small-winged orders suchas Diptera and Hymenoptera. Because thedegree of articulation varies widely amonggroups, with smaller fossils typically beingmore articulated, this taxonomic replacementis the major mechanism by which size hasaffected articulation. The second cause is adecrease in median size of both Orthopteraand Odonatoptera, two large winged groupsthat are found abundantly throughout theinsect fossil record. It is less clear whether thissize decrease also drives the size-articulationrelationship, because the logistic regressionresults are inconsistent when orders are testedindividually. Although small specimens weresignificantly more articulated in Odonatop-tera, Hemiptera, and Diptera, the reverse wastrue in Orthoptera and Blattodea, even afteraccounting for environment (Table 2).
Environmental Controls
Our results indicate that insects haveconsistently been more articulated when bur-ied in deep lakes over their entire evolutionaryhistory, in agreement with detailed studies ofspecific localities (Wilson 1980; Smith andMoe-Hoffman 2007; Henning et al. 2012).Deep lakes favor better preservation througha variety of physical and chemical mecha-nisms.
Deep lake floors are calm, low-energyenvironments, which are unlikely to physical-ly disarticulate dead insects, whereas carcass-es in smaller lakes are subjected to higherenergy due to mixing of the water column.Insect carcasses in marine environments are
FIGURE 8. Phanerozoic occurrences of Coleoptera (graycircles) for all holotypes (A) and for only those describedin the literature after 1950 (B) with logistic regressionmodel fits relating articulation as a function of age in allenvironments (black line), in only deep lake environments(dotted line), and in only non-deep lake environments(hashed line).
28
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM
subject to greater wave action than thosefound in lakes. Even insects preserved inoffshore marine sediments must still passthrough the wave-influenced mixed layer,which is thicker in the ocean than in lakesbecause of the longer fetch of wind-generatedocean waves. Physical destruction or disartic-ulation during transport also reduces preser-vation probability or quality. Because nearlyall insects are terrestrial, transport distance tothe ocean is usually farther than to lacustrineenvironments, leading to better preservationin lakes (Zherikhin 2002).
Anoxia is known to be important inincreasing the quality and likelihood ofpreservation by preventing scavengers andbioturbators from disturbing insect carcasseson or in the sediment. Anoxia is also known topromote microbial mats that may enhancemineral precipitation around a soft-bodiedorganism such as an insect (Allison 1988;Behrensmeyer et al. 2000). Anoxic bottomwaters are frequently developed (at leastseasonally) in large deep lakes. It is uncom-mon for shallower lakes to have anoxicbottom waters because mixing of the watercolumn prevents the depletion of oxygen inthe bottom waters (Olsen 1990; Scheffer 2004).
Restricted ocean basins can also be prone toanoxic bottom waters and several famous,insect-bearing Konservat-Lagerstatten arefrom such anoxic environments (e.g., Solnho-fen). Water chemistry is also important, withhigh solute concentration having been notedto help with preservation of soft-bodiedorganisms by facilitating the precipitation ofauthigenic minerals (Briggs 2003). Endorheiclakes, such as Lake Gosiute of the Green RiverFormation, which often formed in interior riftbasins (Surdam and Wolfbauer 1975; Smith etal. 2008), can have extremely high levels ofminerals and thus an unusual water chemistrythat promotes preservation (Allison and Pye1994).
Our results further imply that variations intaxonomic composition among depositionalenvironments influence articulation, althoughthis is likely a secondary factor. In particular,small-bodied, weak fliers like Coleoptera andDiptera, which have high intrinsic articulation(Smith 2001), are overrepresented in deep lakeenvironments relative to other settings (Table3). It is difficult to assess causality and it ispossible that Diptera are dominantly found indeep lakes because they require the excep-tional preservation of those settings. It does
FIGURE 9. Articulation of insect (solid line) and only Coleoptera (dashed line) holotypes as a function of publicationyear. Beetles described in older papers are much more likely to be based on isolated elytron holotypes, but there is notrend in holotype articulation among insects as a whole.
29TRENDS IN INSECT ARTICULATION

seem plausible, however, that if marinesettings have fewer weakly flying taxa be-cause of their greater distance from theoriginal continental habitat, then reducedarticulation may be attributable to that lackof intrinsically well-preserved taxa.
Conclusion
The fossil record of insects has changed overgeological time to become dominated byarticulated holotype specimens, with a notableshift in the Late Jurassic. Although holotypesare more articulated than an average speci-men, the timing and direction of the shift inarticulation is a robust signal. This shift can beexplained by three factors. The first is adecrease in the average size of insect speci-mens. Overall, larger insects are less articulat-ed than smaller insects, even after accountingfor environment and taxonomic position. Thistrend probably reflects taxonomic shifts fromlarger to smaller groups and is related toshape, especially the ratio of wing to bodysize, rather than size itself. The second factorinvolves shifts in dominant taxonomic groups.Archaic groups, which lack wing-foldingmechanisms and have large wings relative totheir body size, were much more common inthe Carboniferous and Permian, whereas amore modern fauna dominated by Coleoptera,Diptera, and Hymenoptera did not becomeestablished until the Jurassic. Coleoptera,Hymenoptera, and Diptera (especially Cole-optera) all have characteristics that shouldlead to better preservation and their fossils areon average much more articulated. Increasedarticulation, even within single environments,occurs in clades like Hemiptera, Orthoptera,and Diptera also because of changes intaxonomic composition. In contrast, morpho-logically conservative groups like Odonatop-tera, Blattodea, and Coleoptera do not exhibitincreased articulation over time within envi-ronments. Although these results confirm theimportance of insect morphology, especiallyfactors such as the SM index relating wing sizeto body mass, all groups display overallincreases in articulation with time, implyingthat taxonomic shifts cannot completely ex-plain trends in insect articulation.
Our analysis suggests that the depositionalenvironment in which an insect is preserved isthe most important control on articulation.Insect fossils from deep lakes have higherrates of articulation than those from the otherpaleoenvironments and there was a dramaticshift in the Late Jurassic from deltaic, marine,and smaller lake localities to deep and/orlarge lakes. That shift was the primary reasonfor the substantial increase in articulation, alsoin the Late Jurassic. It could affect patterns ininsect diversity, evolution, and ecology thatare preserved in the fossil record, for exampleby biasing diversity in older parts of the insectrecord where preservation quality is poorer.Accounting for differences in depositionalsetting is essential to truly understanding thefossil record of insects, and likely other groupsincluding birds and plants that are also foundlargely in Konservat-Lagerstatten.
Acknowledgments
We thank C. Labandeira and an anonymousreviewer for helpful comments. This is Paleo-biology Database publication 202.
Literature Cited
Allison, P. A. 1988. The role of anoxia in the decay and
mineralization of proteinaceous macro-fossils. Paleobiology
14:139–154.
Allison, P. A., and D. E. G. Briggs. 1991. Taphonomy of
nonmineralized tissues. Pp. 25–70 in P. A. Allison and D. E. G.
Briggs, eds. Taphonomy: releasing the data locked in the fossil
record. Plenum, New York.
Allison, P. A., and K. Pye. 1994. Early diagenetic mineralization
and fossil preservation in modern carbonate concretions. Palaios
9:561–575.
Alroy, J., M. Aberhan, D. J. Bottjer, M. Foote, F. T. Fursich, P. J.
Harries, A. J. W. Hendy, S. M. Holland, L. C. Ivany, W. Kiessling,
M. A. Kosnik, C. R. Marshall, A. J. McGowan, A. I. Miller, T. D.
Olszewski, M. E. Patzkowsky, S. E. Peters, L. Villier, P. J. Wagner,
N. Bonuso, P. S. Borkow, B. Brenneis, M. E. Clapham, L. M. Fall,
C. A. Ferguson, V. L. Hanson, A. Z. Krug, K. M. Layou, E. H.
Leckey, S. Nurnberg, C. M. Powers, J. A. Sessa, C. Simpson, A.
Tomasovych, and C. C. Visaggi. 2008. Phanerozoic trends in the
global diversity of marine invertebrates. Science 321:97–100.
Ansorge, J. 2003. Insects from the Lower Toarcian of middle
Europe and England. Acta Zoologica Cracoviensia 46:291–310.
Archibald, S. B., and V. N. Makarkin. 2006. Tertiary giant
lacewings (Neuroptera: Polystoechotidae): revision and descrip-
tion of new taxa from Western North America and Denmark.
Journal of Systematic Palaeontology 4:119–155.
Arnoldi, L. V, V. V Zherikhin, L. M. Nikritin, and A. G.
Ponomarenko. 1977. Mezozoiskie zhestkokryiye [Mesozoic
Coleoptera]. Trudy Paleontologicheskogo Instituta 161:1–204.
Behrensmeyer, A. K., S. M. Kidwell, and R. A. Gastaldo. 2000.
Taphonomy and paleobiology. Paleobiology 26:103–147.
Bethoux, O. 2005. Cnemidolestodea (Insecta): an ancient order
reinstated. Journal of Systematic Palaeontology 3:403–408.
30
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM

———. 2007. Emptying the Paleozoic wastebasket for insects:
members of a Carboniferous ‘‘protorthopterous family’’ as-
signed to natural groups. Alavesia 1:41–48.
Blanckenhorn, W. U. 2000. The evolution of body size: what keeps
organisms small? Quarterly Review of Biology 75:385–407.
Briggs, D. E. G. 2003. The role of decay and mineralization in the
preservation of soft-bodied fossils. Annual Review of Earth and
Planetary Sciences 31:275–301.
Briggs, D. E. G., B. A. Stankiewicz, D. Meischner, A. Bierstedt, and
R. P. Evershed. 1998. Taphonomy of arthropod cuticles from
Pliocene lake sediments, Willershausen, Germany. Palaios
13:386–394.
Chown, S. L., and K. J. Gaston. 2010. Body size variation in insects:
a macroecological perspective. Biological Reviews of the Cam-
bridge Philosophical Society 85:139–169.
Clapham, M. E., and J. A. Karr. 2012. Environmental and biotic
controls on the evolutionary history of insect body size.
Proceedings of the National Academy of Sciences USA
109:10,927–10,930.
Coram, R. A. 2003. Taphonomy and ecology of Purbeck fossil
insects. Acta Zoologica Cracoviensia 46:311–318.
Dudley, R. 1998. Atmospheric oxygen, giant Paleozoic insects and
the evolution of aerial locomotor performance. Journal of
Experimental Biology 201:1043–1050.
Duncan, I. J., F. Titchener, and D. E. G. Briggs. 2003. Decay and
disarticulation of the cockroach: implications for preservation of
the blattoids of Writhlington (Upper Carboniferous), UK.
Palaios 18:256–265.
Engel, M. S., and M. Gross. 2008. The Pannonian insect fauna of
Styria: a preliminary overview. Austrian Journal of Earth
Sciences 101:52–59.
Felder, M., and F.-J. Harms. 2004. Lithologie und genetische
interpretationder vulkano-sedimentaren ablagerungen aus der
Grube Messel anhand der Forschungsbohrung Messel 2001 und
weiterer Bohrungen. Courier Forschungsinstitut Senckenberg
252:151–203.
Gaston, K. J. 1991. The magnitude of global insect species richness.
Conservation Biology 5:283–296.
Grimaldi, D. A., and M. S. Engel. 2005. Evolution of the insects.
Cambridge University Press, New York.
Harms, F.-J. 2002. Steine erzahlen Geschichte(n): Ursache fur die
Entstehung des Messel-Sees gefunden. Natur und Museum
132:1–4.
Henning, J. T., D. M. Smith, C. R. Nufio, and H. W. Meyer. 2012.
Depositional setting and fossil insect preservation: a study of the
Late Eocene Florissant Formation, Colorado. Palaios 27:481–488.
Hethke, M., F. T. Fursich, B. Jiang, and Y. Pan. 2013. Seasonal to
sub-seasonal palaeoenvironmental changes in Lake Sihetun
(Lower Cretaceous Yixian Formation, NE China). International
Journal of Earth Sciences 102:351–378.
Labandeira, C. C. 1999. Insects and other hexapods. Encyclopedia
of Paleontology 1:603–624.
Labandeira, C. C., and J. J. Sepkoski Jr. 1993. Insect diversity in the
fossil record. Science 261:310–315.
Lutz, H. 2003. The Eckfeld Maar (Vulkaneifel, Germany): a key
locality for the understanding of the Eocene paleoenvironment
of the western Rhenish shield. Geıologie de la France 1:63–67.
Martin, S. K. 2010. Early Jurassic coleopterans from the Mintaja
insect locality, Western Australia. Acta Geologica Sinica 84:925–
953.
Martınez-Delclos, X., and J. Martinell. 1993. Insect taphonomy
experiments: their application to the Cretaceous outcrops of
lithographic limestones from Spain. Kaupia: Darmstadter
Beitrage zur Naturgeschichte 2:133–144.
Martıınez-Delclos, X., D. E. G. Briggs, and E. Penalver. 2004.
Taphonomy of insects in carbonates and amber. Palaeogeog-
raphy, Palaeoclimatology, Palaeoecology 203:19–64.
McCobb, L. M. E., I. J. Duncan, E. A. Jarzembowski, B. A.
Stankiewicz, M. A. Wills, and D. E. G. Briggs. 1998. Taphonomy
of the insects from the Insect Bed (Bembridge Marls), late
Eocene, Isle of Wight, England. Geological Magazine 135:553–
563.
Okajima, R. 2008. The controlling factors limiting maximum body
size of insects. Lethaia 41:423–430.
Olsen, P. E. 1990. Tectonic, climatic, and biotic modulations of
lacustrine ecosystems-examples from Newark Supergroup of
Eastern North America. In B. Katz, ed. Lacustrine basin
exploration: case studies and modern analogs. American
Association Petroleum Geologists Memoir 50: 209–224
Papier, F., A. Nel, L. Grauvogel-Stamm, and J.-C. Gall. 2005. La
diversite des Coleoptera (Insecta) du Trias dans le nord-est de la
France. Geodiversitas 27:181–199.
Ponomarenko, A. G. 2002. Superorder Scarabaeidea Laicharting,
1781. Order Co1eoptera Linne, 1758. The beetles. Pp. 164–176 in
A. P. Rasnitsyn and D. L. J. Quicke, eds. History of insects.
Kluwer Academic, Dordrecht.
R Development Core Team. 2012. R: a language and environment
for statistical computing. R Foundation for Statistical Comput-
ing, Vienna.
Rasnitsyn, A. P., and D. L. J. Quicke. 2002. History of insects.
Kluwer Academic, Dordrecht.
Rust, J. 1998. Biostratinomie von insekten aus der Fur-Formation
von Danemark (Moler, oberes Paleozan/unteres Eozan).
Palaontologische Zeitschrift 72:41–58.
Scheffer, M. 2004. Ecology of shallow lakes: population and
community biology series. Springer, Berlin.
Sharov, A. G. 1968. Filogeniya ortopteroidnykh nasekomykh.
Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR
118:1–216.
Shcherbakov, D. E. 2008. Madygen, Triassic Lagerstatte number
one, before and after Sharov. Alavesia 2:113–124.
Smith, D. M. 2000. Beetle taphonomy in a recent ephemeral lake,
southeastern Arizona. Palaios 15:152–160.
———. 2001. Taphonomic bias and insect diversity: a lesson from
the beetles and flies. PaleoBios 21:117.
———. 2012. Exceptional preservation of insects in lacustrine
environments. Palaios 27:346–353.
Smith, D. M., and A. P. Moe-Hoffman. 2007. Taphonomy of
Diptera in lacustrine environments: a case study from Florissant
Fossil Beds, Colorado. Palaios 22:623–629.
Smith, D. M., A. Cook, and C. R. Nufio. 2006. How physical
characteristics of beetles affect their fossil preservation. Palaios
21:305–310.
Smith, M. E., A. R. Carroll, and B. S. Singer. 2008. Synoptic
reconstruction of a major ancient lake system: Eocene Green
River Formation, western United States. Geological Society of
America Bulletin 120:54–84.
Sukacheva, I. D., and A. P. Rasnitsyn. 2004. Jurassic insects
(Insecta) from the Sai-Sagul Locality (Kyrgyzstan, Southern
Fergana). Paleontological Journal 38:182–186.
Surdam, R. C., and K. O. Stanley. 1980. The stratigraphic and
sedimentologic framework of the Green River Formation,
Wyoming. Pp. 205–222 in A. Harrison, ed. Stratigraphy of
Wyoming. Wyoming Geological Association Guidebook, 31st
annual field conference, Jackson Hole, Wyoming, September 6–
10, 1980. Wyoming Geological Association, Casper.
Surdam, R. C., and C. A. Wolfbauer. 1975. Green River Formation,
Wyoming: a playa-lake complex. Geological Society America
Bulletin 86:335–345.
Veach, S. W., and H. W. Meyer. 2008. History of paleontology at
the Florissant Fossil Beds, Colorado. In H. W. Meyer and D. M.
Smith, eds. Paleontology of the Upper Eocene Florissant
Formation, Colorado. Geological Society of America Special
Paper 435:1–18.
31TRENDS IN INSECT ARTICULATION

Wagner, T., C. Neinhuis, and W. Barthlott. 1996. Wettability and
contaminability of insect wings as a function of their surface
sculptures. Acta Zoologica 77:213–225.
Wang, B., H. Zhang, E. A. Jarzembowski, Y. Fang, and D. Zheng.
2013. Taphonomic variability of fossil insects: a biostratinomic
study of Palaeontinidae and Tettigarctidae (Insecta: Hemiptera)
from the Jurassic Daohugou Lagerstatte. Palaios 28:233–242.
Wappler, T. 2003. Systematik, Phylogenie, Taphonomie und
Palaookologie der Insekten aus dem Mittel-Eozan des Eckfelder
Maares, Vulkaneifel. Clausthaler Geowissenschaften 2:1–126.
Wappler, T., E. D. Currano, P. Wilf, J. Rust, and C. C. Labandeira.
2009. No post-Cretaceous ecosystem depression in European
forests? Rich insect-feeding damage on diverse middle Palae-
ocene plants, Menat, France. Proceedings of the Royal Society of
London B 276:4271–4277.
Wilson, M. V. H. 1980. Eocene lake environments: depth and
distance-from-shore variation in fish, insect, and plant assem-
blages. Palaeogeography, Palaeoclimatology, Palaeoecology
32:21–44.
———. 1988. Reconstruction of ancient lake environments using
both autochthonous and allochthonous fossils. Palaeogeogra-
phy, Palaeoclimatology, Palaeoecology 62:609–623.
Zeuner, F. E. 1942. Two new fossil butterflies of the family
Pieridae. Annals and Magazine of Natural History 9:409–416.
Zherikhin, V. V. 2002. Pattern of insect burial and conservation. Pp.
17–62 in A. P. Rasnitsyn and D. L. J. Quicke, eds. History of
Insects. Kluwer Academic, Dordrecht.
Zherikhin, V. V, M. B. Mostovski, P. Vrsansky, V. A. Blagoderov,
and E. D. Lukashevich. 1999. The unique Lower Cretaceous
locality Baissa and other contemporaneous fossil insect sites in
North and West Transbaikalia. Pp. 185–191 in P. Vrsansky, ed.
Proceedings of the First International Palaeoentomological
Conference, Moscow 1998. AMBA Projects International, Bra-
tislava.
32
JERED A. KARR AND MATTHEW E. CLAPHAM
JERED A. KARR AND MATTHEW E. CLAPHAM
Copyright © 2022 FDOKUMEN




















