
Cytotoxicity and apoptotic effects of nickel oxide nanoparticles in cultured HeLa cells
Upload
independentCategory
view
2download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Tanshinone IIA, an isolated compound from Salvia miltiorrhiza Bunge, inducesapoptosis in HeLa cells through mitotic arrest
Lingli Zhou a,1,2, W.K. Chan a,1,3, Naihan Xu a, Kang Xiao a, Houwei Luo c, Kathy Qian Luo b, Donald C. Chang a,⁎a Department of Biology, The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong, Chinab Bioengineering Graduate Program, The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong, Chinac Department of Natural Medical Chemistry, China Pharmaceutical University, Nanjing, China
a b s t r a c ta r t i c l e i n f o
Article history:Received 30 November 2007Accepted 4 July 2008
Keywords:Tanshinone IIABcl-2Microtubule-interfering agentsApoptosisMitotic arrestAnti-cancer drug
Aims: Tanshinone IIA (Tan IIA) is a compound isolated from Salvia miltiorrhiza Bunge (Danshen). The aim ofthis study is to investigate the mechanisms of its anti-cancer effect.Main methods: To clearly delineate the cell cycle-dependent effects of Tan IIA, we used either synchronizedcells or single living cell analysis to conduct our studies. Subcellular fractionation, Western blot analysis,immuno-fluorescence staining and FACS analysis were also employed in our study.Key findings: We found that Tan IIA could arrest cancer cells in mitosis by disrupting the mitotic spindle andsubsequently triggered cells to enter apoptosis through the mitochondria-dependent apoptotic pathway.Thus, Tan IIA could selectively kill mitotic cells over interphase cells. In comparison with other existing anti-cancer drugs that cause mitotic arrest by interfering with the microtubule structure (such as vincristine ortaxol), Tan IIA destroyed only the mitotic spindle during the M phase but not the microtubule structure ininterphase cells. Furthermore, Tan IIA could trigger the mitotic arrested cells to enter apoptosis faster thanvincristine or taxol.Significance: Since Tan IIA can selectively induce cancer cells to enter apoptosis through mitotic arrest, it hasthe potential to be developed into an anti-cancer drug.
© 2008 Elsevier Inc. All rights reserved.
Introduction
A basic requirement for anti-cancer drugs is that they should havea strong preference in killing cancer cells over non-cancer cells. Sincecancer cells usually undergo active cell division (mitosis), a usefulapproach in finding anti-cancer drugs is to test whether a compoundcan selectively kill mitotic cells. For example, an important class ofanti-cancer drugs called “microtubule-interfering agents (MIAs)”,including taxol, vincristine and vinblastine, can disrupt the formationof the mitotic spindle and thus blocks the cell cycle progression inmitosis. Prolonged mitotic arrest, in turns, will induce cells to enterapoptosis (Bhalla, 2003; Mollinedo and Gajate, 2003; Wang et al.,2000).
The MIA drugs, however, are not free of undesirable side effects.Many normal cell functions are known to depend on either the
dynamics of microtubules or molecular movement associated withthe microtubule-based motor proteins (Burkhardt, 1998; Karki andHolzbaur, 1999; Levy and Holzbaur, 2006; Murray andWolkoff, 2003).Thus, treatment with MIAwill unavoidably interfere with certain vitalfunctions of the normal cells. A more preferred class of anti-cancerdrugs would be compounds that could induce mitotic arrest butwould not interfere with themicrotubule structure in interphase cells.In order to search for such compounds, we have conducted a seriesof studies on the apoptotic effects of many herbal extracts, par-ticularly those compounds isolated from traditional Chinese medicine(TCM).
Danshen (Salvia miltiorrhiza Bunge) has been widely used in TCMpractice for over a thousand years in the treatment of coronary arterydisease and cerebrovascular diseases. Tanshinone IIA (Tan IIA), anabietane type-diterpene quinine, is amajor extractable component ofDanshen. (Its structure is shown in Fig. 1A). Recently, it was reportedthat Tan IIA could induce cell death in several human tumor cell lines(Meng et al. 2002; Tang et al., 2003a,b; Yoon et al. 1999; Yuan et al.2003, 2004). The mechanism, however, was not clearly understood.Recently, we found that Tan IIA could selectively induce mitotic cellsto enter apoptosis. This finding suggests that Tan IIA (or itsderivatives) may have good potential to be developed into a novelclass of anti-cancer drugs. Thus, we conducted a series of experi-ments to investigate the following questions: (1) Does Tan IIA only
Life Sciences 83 (2008) 394–403
⁎ Corresponding author. Tel.: +852 2358 7326; fax: +852 2358 1559.E-mail address: [email protected] (D.C. Chang).
1 These authors contributed equally to this work.2 Current address: The Endocrinology Department, Peking University People’s
Hospital, Beijing, China.3 Current address: Department of Pediatrics and Adolescent Medicine, The University
of Hong Kong, Hong Kong, China.
0024-3205/$ – see front matter © 2008 Elsevier Inc. All rights reserved.doi:10.1016/j.lfs.2008.07.011
Contents lists available at ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r.com/ locate / l i fesc ie

Author's personal copy
kill mitotic cells but not cells in interphase? (2) What is themechanism of Tan IIA-induced cell death? Is it through apoptosis?If this is the case, is the apoptotic process mediated through themitochondria-dependent pathway? (3) How would the apoptoticprocess induced by Tan IIA differ from that induced by MIAs? DoesTan IIA offer any potential advantages overMIAs? In order to examinewhether the apoptotic effects of Tan IIA are cell cycle-dependent, ourexperiments used mainly synchronized cell population or singleliving cell analysis.
Materials and methods
Chemicals
Crystallized Tanshinone IIA (purified from Danshan extract byDr. Houwei Luo from China Pharmaceutical University, Nanjing,China) was dissolved in dimethyl sulfoxide (DMSO) at 2 mg/ml andthen sterilized by filtration. Anti-cytochrome c mouse monoclonalantibody was purchased from BD Biosciences PharMingen (SanDiego, CA). Thymidine, hydroxyurea, propidium iodide, and othercommon chemicals were all purchased from Sigma-Aldrich (St.Louis, MO). Hoechst 33342 was from Molecular Probes (Eugene,OR). Geneticin was purchased from Invitrogen (Carlsbad, CA). RNase
A was from Roche (Nutley, NJ). Mowiol 4-88 was from Calbiochem(San Diego, CA). Anti-PARP mouse monoclonal antibody was fromSanta Cruz Biotechnology, Inc. (Santa Cruz, CA). JNK inhibitorSP600125 and Cdk1 inhibitor roscovitine were from Calbiochem(San Diego, CA).
Plasmid construction and gene transfection
The original Mcl-1 gene was kindly provided by Dr. Steven W.Edwards from the University of Liverpool (Akgul et al. 2000). Mcl-1was amplified and cloned into pEGFP-C3 vector (Clontech, CA) byprimers: Forward 5′-CCGGAATTCCGATGTTTGGCCTCAAAAGAAACG-3′and Reverse 5′-CGCGGATCCCGCTATCTTATTAGATATGCCAAAC-3′. Thefusion genewas transfected into cell using Lipofectamine™ 2000 fromInvitrogen (Carlsbad, CA) using the standard protocol recommendedby Invitrogen.
Cell culture and synchronization
HeLa cells were cultured in minimum essential medium (MEM)containing 10% fetal bovine serum, 100 U/ml penicillin and 100 μg/mlstreptomycin in 5% CO2 at 37 °C. A stable HeLa cell line expressing YFP-tubulin fusion protein was used to study the effect of Tanshinone IIA
Fig. 1. Effects of Tan IIA on HeLa cells. (A) The molecular structure of Tan IIA. (B) Phase contrast images of HeLa cells under different times (12 h; 16 h; 20 h; 24 h) of Tan IIA (5 μg/ml)treatment. Scale bar, 50 μm. (C) Phase contrast and DNA counterstaining images of HeLa cells under Tan IIA (5 μg/ml) treatment. Scale bar, 10 μm. (D) Tan IIA had a dosage-dependentapoptotic effect on HeLa cells. Cells were treated with various concentrations of Tan IIA for 24 h. The percentage of apoptotic cells was determined based upon cell and nuclearmorphology. Data were from 4 independent experiments.
395L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
on microtubule structure and cell cycle progression. The culturingcondition was the same as normal HeLa cells except 0.5 mg/ml ofGeneticin was added into the culture medium.
Double thymidine block was used to synchronize cells at early Sphase. Briefly, cells were grown in the presence of 2mM thymidine for18 h, washed with PBS 3 times, and grown again in fresh medium
Fig. 2. Tan IIA induces cell death through mitotic arrest. (A) Flow cytometry analysis of Tan IIA-treated HeLa cells. Non-synchronized cells were treated with Tan IIA (5 μg/ml) fordifferent time periods. (B) Ratios of cell population in G2/M over G1 at different times after Tan IIA treatment as determined from FASC analysis. (C, D) Percentage of mitotic (andapoptotic) cells in a synchronized HeLa cell population. Cells were synchronized at early S phase by double thymidine block. Four hours after removing thymidine, cells were treatedwith (D) or without (C) 5 μg/ml of Tan IIA. Data are from 3 independent experiments. (E) Effects of Tan IIA treatment on interphase HeLa cells. Cells were synchronized in early G1phase (see “Materials and methods”). Tan IIA (5 μg/ml) was applied at 3–4 h after releasing from mitotic block. (F) Differential effects of Tan IIA when the drug was added to cellseither in late S phase or in early G1 phase. The mitotic and apoptotic cells were counted at 16 h after Tan IIA treatment. Data were based on 4 independent experiments and shown asmeans±SD. (⁎⁎indicates pb0.01).
396 L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
without thymidine for 9 h. Cells were then treated with 2 mMthymidine for 18 h to block cells at early S phase. To synchronize cellsin early G1 phase, cells were first synchronized at early S phase by2 mM hydroxyurea for 16 h, and then released into fresh medium for10 h. The mitotic cells were shaken off and re-seeded into culturedishes for 3–4 h. At this point most cells were at the early G1 phase.
Determination of mitotic and apoptotic cells
At indicated time points after Tan IIA treatment, cells werecollected by trypsinization and centrifugation. The cell pellet wasthen suspended and fixed with 100 μl of 4% paraformaldehyde for30 min, and nuclei were stained with 100 ng/ml Hoechst 33342 for10 min. Cells were examined under a fluorescence microscope using a40× objective. Cells with smooth surface and intact chromosomeswere categorized as mitotic cells, whereas cells with rough surfaceand fragmented chromosomes were considered as apoptotic cells. Atleast 300 cells were counted for each sample.
FACS analysis
The cell cycle profile was analyzed by flow cytometry after stainingDNA with propidium iodide (PI). At the indicated time points after5 μg/ml of Tan IIA treatment, cells were trypsinized, rinsed with ice-cold PBS, and fixed in 70% ethanol for overnight at −20 °C. After thefixation, cells were washed with ice-cold PBS and treated with 200 μlof RNase A (1 mg/ml) at 37 °C for 30–60 min. Cells were thenincubated with 800 μl of PI staining buffer (0.1 mg/ml PI and 1% TritonX-100 in PBS) at 4 °C in the dark for 30 min. Afterwards, cells wereanalyzed by FACS on a Calibur Flow Cytometer (Becton Dickinson,Franklin Lakes, NJ).
Immuno-fluorescence staining
The cell monolayer on coverslip was fixed with 4% paraformalde-hyde plus 0.1% glutaraldehyde for 15 min at room temperature andpermeabilized in 0.2% Triton X-100 for another 15 min. Non-specificbinding was blocked by incubating cells in 3% BSA for 1 h. Cells werethen incubated with anti-cytochrome c antibody (1:200 dilution) for2 h at room temperature. After washing in PBS 3 times, cells wereincubated with the secondary antibody at a dilution of 1:200 (FITC-conjugated goat anti-mouse antibody) for 1 h. Auto fluorescence ofglutaraldehyde was quenched with a freshly prepared solution of 0.1%NaBH4 for 10min. Then the coverslip was washed andmounted onto aslide using Mowiol 4-88. After solidifying, the sample was ready forobservation under a fluorescence microscope.
Subcellular fractionation
To isolate the cytosolic fraction and membrane (containingmitochondria) fraction, cells were treated with 50∼100 μl of PBScontaining 0.01% digitonin and inhibitors for proteases and phospha-tases for 2 min at room temperature, followed by immediatecentrifugation at 14,000 rpm for 2 min at room temperature. Thesupernatant was collected as the cytosolic fraction. The cell pelletswere further lysed in NP-40 lysis buffer (50 mM Tris–HCl, pH 8.0,150 mM NaCl, and 1% NP-40) with inhibitors for proteases andphosphatases for 30 min on ice. The lysates were then centrifuged for10 min at 14,000 rpm at 4 °C. The supernatant was collected as themembrane fraction.
Western blotting
HeLa cells were cultured in 60 or 100 mm petri-dishes. Cells atdifferent time points after Tan IIA treatment were collected and lysedin NP-40 lysis buffer (50 mM Tris–HCl, pH 8.0, 150 mM NaCl, and 1%
NP-40) in the presence of protease inhibitors. Whole cell lysates(100 μg/lane) were separated on 12% SDS-PAGE and transferred onto aHybond ECL nitrocellulose membrane (Amersham). After blocking,the membranes were incubated for 3 h at room temperature orovernight at 4 °C with antibodies at a dilution of 1:500 or 1:1000. Themembranes were incubated with horseradish peroxidase-conjugatedsecondary antibody at a dilution of 1:5000 for 1 h and developed usingECL™ Western blotting analysis system.
The real-time living cell imaging system
Living cells grown on a 35-mm round coverslip was mounted ontoa chamber containing a pH-buffered observation medium (Hepes-buffered Dulbecco's MEM with 4 mM glutathione, 1 mM L-ascorbicacid, 0.5 mM DTT, pH 7.4). Cells were then examined under amicroscope equipped with a heating stage maintaining the tempera-ture at 37 °C. Two imaging systems were used for observation: (1) aninverted fluorescence microscope (Axiovert 200M, Zeiss) equippedwith a cooled CCD camera (Diagnostic Instruments) and (2) a confocalmicroscope (Leica TCS SP2) equipped with multiple laser sources.
Results
Tan IIA could induce apoptosis in HeLa cells in a dosage-dependentmanner
The apoptotic effect of Tan IIA on HeLa cells was first determinedby the morphological assay. Cells were treated with or without Tan IIAfor 12 to 24 h and stained with a cell permeable DNA dye (Hoechst33342) and they were then analyzed under the fluorescencemicroscope. We found that many cells became rounded under theTan IIA treatment (Fig. 1B). These rounded cells represented bothmitotic and apoptotic cells. Cells with a smooth surface and intactchromosomes were classified as mitotic cells, whereas cells withrough surface and fragmented chromosomes were identified asapoptotic cells (Fig. 1C). After HeLa cells were treated with differentconcentrations of Tan IIA (1 μg/ml, 3 μg/ml, 5 μg/ml and 10 μg/ml) for24 h, the percentages of apoptotic cells were found to be 5.4%, 50.7%,63.3% and 65.6% respectively (Fig. 1D). A very low percentage of celldeath (1.6%) was detected in the control cells containing noTan IIA but0.5% DMSO. Based on this result, we concluded that Tan IIA (≥3 μg/ml)can significantly induce cell death in HeLa cells.
Tan IIA arrested cells in mitosis first before inducing them to undergoapoptosis
We then conducted a time-lapse imaging measurement. We foundthat many Tan IIA-treated cells became rounded first and finally died afew hours later (see also Fig. 1B). We suspected that Tan IIA mightarrest cells in mitosis before triggering them to undergo apoptosis.Thus, we decided to determine the effect of Tan IIA on cell cycleprogression using a flow cytometry analysis (Fig. 2A). Followingtreatment with 5 μg/ml of Tan IIA, the percentage of cells in G1 phasegradually decreased, while a higher proportion of cells wereaccumulated at G2/M phase (Fig. 2A and B). Sub-G1 population cellsalso started to appear at 12 h of Tan IIA treatment, indicating thatsome of the Tan IIA-treated cells had become apoptotic (Fig. 2A). Thesefindings suggest a possibility that Tan IIA may arrest cells in the G2/Mphase first before inducing them to undergo apoptosis.
To directly determine whether Tan IIA arrests cells in G2 or Mphase, we synchronized HeLa cells at S phase using a doublethymidine block. Tan IIA was applied to the medium 4 h after cellswere released from the thymidine block. Cells were fixed and stainedwith Hoechst 33342 to examine both cellular and nuclear morphol-ogies. The rounded cells with smooth cell membrane and condensedchromosomes were identified as mitotic cells, while rounded cells
397L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
with rough membrane and fragmented chromosomes were classifiedas apoptotic cells. In the absence of Tan IIA, the control cells enteredMphase around 10–12 h after thymidine release and exited frommitosis
after 13 h (Fig. 2C). Practically no apoptotic cells were observed. Incomparison, when Tan IIA (5 μg/ml) was added into the culturemedium, cells could still enter mitosis around 10–12 h after release
398 L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
from double thymidine block (i.e. 6–8 h after Tan IIA treatment). They,however, did not exit the M phase and were arrested in mitosis(Fig. 2D). Then, after about 4 to 6 h of mitotic arrest, the arrested cellsstarted to become apoptotic (Fig. 2D). This finding demonstrated thatTan IIA can indeed arrest cells in mitosis and then induce them toundergo apoptosis.
Tan IIA mainly killed mitotic cells but not interphase cells
To examine whether Tan IIA kills cells through mitotic arrest, wecompared the apoptotic effects of Tan IIA on cells at different stages ofthe cell cycle. That is, we tested if Tan IIA selectively kills mitotic cellsbut not cells in interphase. HeLa cells were first synchronized at S orMphase (see “Materials andmethods”). Then, Tan IIAwas added into the
culture medium when cells were either in late S phase (i.e., 4 h afterreleasing from the S phase block) or in early G1 phase (4 h afterreleasing from the mitotic block). The percentage of cells in mitosis orapoptosis was subsequently determined as a function of time (up to16 h). Since the length of the cell cycle of HeLa cell is roughly 20 h, cellstreated with Tan IIA at late S phase would enter the M phase quicklyduring the 16 h experimental periodwhile cells treated with Tan IIA inearly G1 phase would not. Indeed, we observed a much higherpercentage of cells undergoing apoptosis when Tan IIA was applied tocells in late S phase (Fig. 2D and F). Very few cells became apoptoticwhen Tan IIA was applied in early G1 phase (Fig. 2E). (Note: The smallpercentage of mitotic and apoptotic cells found in t=16 h in Fig. 2Ewas probably due to imperfect cell synchronization). These resultssuggest that Tan IIA induces cell death mainly through mitotic arrest.
Fig. 4. Tan IIA induces apoptosis via the mitochondria-dependent pathway. (A) Upper panels: Results of immunostaining showing the distribution of cytochrome c in mitotic HeLacells with or without Tan IIA (5 μg/ml) treatment. Phase images of the same cells are also shown. Scale bar,10 μm. Lower panel:Western blot analysis showing that increasing amountsof cytochrome c were released from mitochondria into cytosol during Tan IIA (5 μg/ml) treatment. HeLa cells were synchronized first at early S phase by double thymidine blockbefore the addition of Tan IIA. (B) Western blot analysis showing that caspase-3 was activated during the course of Tan IIA treatment. (C) Mcl-1 was degraded during the Tan IIAtreatment and over-expression of GFP–Mcl-1 can inhibit the activation of caspase-3. HeLa cells were transfected with GFP–Mcl-1 (or GFP alone) for 24 h before treated with Tan IIA(5 μg/ml). (D) Comparison of Tan IIA-induced apoptosis in non-synchronized cells transfected with GFP–Mcl-1 or GFP alone. Data were from 3 independent experiments. (⁎ indicatespb0.05, ⁎⁎ indicates pb0.01).
Fig. 3. Tan IIA causes mitotic spindle disruption in prometaphase. (A) Comparison of microtubule structures in interphase cells with or without the treatment of taxol (50 nM),vincristine (10 nM) or Tan IIA (5 μg/ml). HeLa cells stably expressing YFP-tubulin fusion protein were treated with different drugs for 6 h. Scale bar, 10 μm. (B) Living cell imagingrecords showing the change of microtubule (MT) structures during G2/M transition in cells treated with taxol, Tan IIA or vincristine. HeLa cells stably expressing YFP-tubulin fusionprotein were treated with different drugs at 4 h after release from the S phase block (by HU treatment). The fluorescence images of YFP-tubulin were recorded using a confocalmicroscope. Scale bar, 10 μm. (C) Percentage of cells arrested at different stages of mitosis. Cells were synchronized first in S phase using HU block. Tan IIA (5 μg/ml) was added at 4 hafter HU release. Most cells were arrested in prometaphase (PM) instead of anaphase (Ana). Data were from 3 independent experiments. (D) Percentage of mitotic arrested cells andmitotic cells with abnormal spindle structures at various times after Tan IIA treatment in synchronized HeLa cells. The experimental protocol was similar to that used in (C). Datawerefrom 3 independent experiments.
399L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
Cells under the Tan IIA treatment were arrested before the metaphase–anaphase transition due to destruction of the mitotic spindle
What could cause Tan IIA to arrest cells in mitosis? One possibilityis that Tan IIA might disrupt the microtubule structure in cells andthus prevent the formation of mitotic spindle. This prompted us tocompare the cellular effects of Tan IIA with other MIAs. We found thatthe effects of Tan IIA on the microtubule structure were different fromthose of MIA drugs, such as taxol or vincristine. First, cells treated withMIAs usually had distorted microtubule structure in the interphase,while the microtubule structure of Tan IIA-treated interphase cellswas normal, i.e., similar to control cells (Fig. 3A). Secondly, when cellswere treated with either taxol or vincristine, they failed to formmitotic spindles when cells entered M phase (Fig. 3B). By contrast,when the Tan IIA-treated cells entered the M phase, mitotic spindlescould still be formed in prometaphase. But soon after that, spindlesstarted to deform and finally disappeared (Fig. 3B). The chromosomesremained condensed but failed to line up at the metaphase plate. As aresult, most Tan IIA-treated cells were arrested before the metaphase–anaphase transition (Fig. 3C). In fact, we have done a quantitativemeasurement and found that, when synchronized S phase cells weretreated with Tan IIA, the percentage of cells arrested in mitosis and thepercentage of cells showing distorted mitotic spindles were highlycorrelated (Fig. 3D). These results indicate that themajor causeofmitoticarrest was due to the disruption ofmitotic spindle by Tan IIA atMphase.
There may be a concern about the dosage effects of Tan IIA onmitotic spindles over those of other microtubule targeting drugs. It ispossible that microtubule binding agents may arrest cells in mitosis atdoses lower than those required to disrupt interphase microtubules.To address this possibility, we tested the effects of higher dosages ofTan IIA (10 and 20 μg/ml) on the microtubule structure in interphasecells. We observed that interphase microtubules were not disruptedwhen the applied Tan IIA concentration was up to 20 μg/ml (data notshown). In contrast, 5 μg/ml Tan IIA was sufficient to disrupt themitotic spindles.
Tan IIA-induced apoptosis was through the mitochondria-dependentpathway
Then, we examined whether Tan IIA induced cell death throughthe mitochondria-dependent apoptotic pathway or not. It is wellknown that when cells enter apoptosis through the mitochondria-dependent pathway, cytochrome c is released frommitochondria intocytosol, where it binds to Apaf-1 and activates caspase-9 (Budihardjoet al., 1999; Green and Reed, 1998; Li et al., 1997; Wang, 2001). Theactivated caspase-9 then activates an executive protease, caspase-3(Budihardjo et al., 1999; Degterev et al., 2003; Li et al., 1997; Riedlet al., 2001; Slee et al., 1999). The activation of caspase-3 then canresult in multiple biochemical and morphological changes inapoptotic cells, including DNA fragmentation, chromatin
Fig. 5. Comparing the apoptotic pathways of Tan IIA with those of MIA. (A) Over-expression of GFP–Mcl-1 protected cells from MIAs-induced apoptosis. HeLa cells were transfectedwith GFP or GFP–Mcl-1. Taxol (50 μM) was added at 12 h after transfection. The percentage of apoptotic cells (in GFP-positive cells) was determined at 0, 12, 18 and 24 h after taxoltreatment. (⁎⁎ indicates pb0.01). (B) The effect of JNK inhibitor on taxol-induced apoptosis. HeLa cells were synchronized at early S phase by double thymidine block. Taxol (50 μM)was added at 8 h after release from S phase block. A JNK inhibitor, SP600125 (50 μM), was added at 8 h after drug application. The cells were collected at the indicated time points andthe apoptotic cells were counted based on DNA staining. Data were from three independent experiments. (⁎ indicates pb0.05). (C) The effects of kinase inhibitors on Tan IIA-inducedapoptosis. HeLa cells were synchronized at early S phase by double thymidine block. Tan IIA was applied at 4 h when cells were released from early S phase block. Cdk1 inhibitorroscovitine (25 μM) or JNK inhibitor SP600125 (20 μM) was added at 10 h after Tan IIA treatment. (⁎ indicates pb0.05). (D) A model summarizing the apoptotic pathway induced byTan IIA in HeLa cells based on our experimental findings.
400 L. Zhou et al. / Life Sciences 83 (2008) 394–403
Author's personal copy
condensation and plasma membrane blebbing (Porter and Janicke,1999; Sebbagh et al., 2001). Thus, we conducted a series ofexperiments to determine: (a) whether or not cytochrome c wasreleased frommitochondria to cytosol following the Tan IIA treatment,and (b) whether caspase-3 was activated during this process.
To detect the subcellular localization of cytochrome c in HeLa cells,cells were fixed and stained with anti-cytochrome c antibody. In anormal mitotic cell, the distribution pattern of cytochrome c appearedto be filamentous, indicating that cytochrome c was localized withinmitochondria (Fig. 4A). In cells treated with 5 μg/ml of Tan IIA, adiffused pattern of cytochrome c was observed, indicating thatcytochrome c was released from mitochondria into cytosol (Fig. 4A).This result was further confirmed using aWestern blot analysis, whichshowed that increasing amounts of cytochrome cwere detected in thecytosolic fraction of the cell extract following the treatment of Tan IIA(Fig. 4A).
To detect the activation of caspase-3, we assayed the cleavage ofthe cellular substrate of caspase-3, PARP, using Western blot analysis(Fig. 4B). At 8 h after the Tan IIA treatment, PARP was cleaved intosmaller products. Activated caspase-3 was also detected at the sametime (Fig. 4B). Taken together, these results suggest that Tan IIAinduces apoptosis in HeLa cells through the mitochondria-dependentpathway.
The signaling pathway of Tan IIA-induced apoptosis involved thedegradation of Mcl-1
Next, we examined if the Tan IIA-induced apoptotic processinvolves any Bcl-2 family proteins, which play very important rolesin regulating the dysfunction of mitochondria during apoptosis(Nijhawan et al., 2003; Reed, 2006; Sharpe et al., 2004). UsingWestern blot analysis, we found that Mcl-1 (an anti-apoptoticmember of the Bcl-2 family proteins) decreased rapidly under theTan IIA treatment (Fig. 4C). To further investigate the role of Mcl-1, wetransfected either GFP–Mcl-1 or GFP alone (as a control) into HeLacells. Tan IIA was added into the medium at 24 h after transfection.Cells were collected at different time points and were assayed for bothcaspase-3 activation and PARP cleavage by Western blotting analysis.We found that over-expression of GFP–Mcl-1 could inhibit theactivation of caspase-3 triggered by Tan IIA, while cells expressingGFP could not (Fig. 4C). Then, by cell counting analysis, we found thatover-expression of GFP–Mcl-1 (but not GFP alone) could significantlyinhibit Tan IIA-induced apoptosis in HeLa cells (Fig. 4D). Takentogether, these results indicate that Mcl-1 plays an important role inthe signaling pathway of Tan IIA-induced apoptosis.
Similarity and difference between the signaling pathways of TanIIA-induced apoptosis and MIAs-induced apoptosis
Currently there is an active interest in studying the signalingpathways of MIA-induced apoptosis, which is still not fully under-stood (Chadebech et al., 2000; Huang et al., 2000; Jordan and Kamath,2007; Wang et al., 1999; Wang et al., 2000). Like Tan IIA, we observedthat MIA-induced apoptosis also depends on the degradation of Mcl-1.First, we observed that Mcl-1 was degraded in taxol-treated cells (datanot shown). Second, we found that over-expression of GFP–Mcl-1could also strongly inhibit taxol-induced cell death in HeLa cells(Fig. 5A). Thus, it appears that there is a certain similarity between theMIAs-induced apoptosis and Tan IIA-induced apoptosis.
Furthermore, we observed that JNK was activated in both taxol-treated and Tan IIA-treated cells (data not shown). A specific JNKinhibitor (SP600125) could suppress both MIA-induced and Tan IIA-induced apoptosis (Fig. 5B and C). Since it is known that inhibiting JNKcan block the G2/M transition (Mingo-Sion et al., 2004), we would liketo exclude this effect and test specifically the effect of JNK on Tan IIA-(orMIA-) induced apoptosis after cells already entered theM phase. To
achieve this goal, we first synchronized HeLa cells at early S phaseusing double thymidine treatment. Then, Tan IIA (5 μg/ml) or taxol(50 nM) was added into the culture medium at 4 h after released fromthe S phase block. Specific kinase inhibitors were applied into themedium at 10 h after double thymidine release, when cells werealready in the M phase. The percentage of apoptotic cells was de-termined at different times following drug treatment. The resultsindicate that inhibition of JNK can modestly suppress apoptosisinduced by either taxol or Tan IIA (Fig. 5B and C). Previously, it hasbeen demonstrated that taxol-induced apoptosis is at least partiallymediated by JNK (Kolomeichuk et al., 2008; Selimovic et al., 2008).
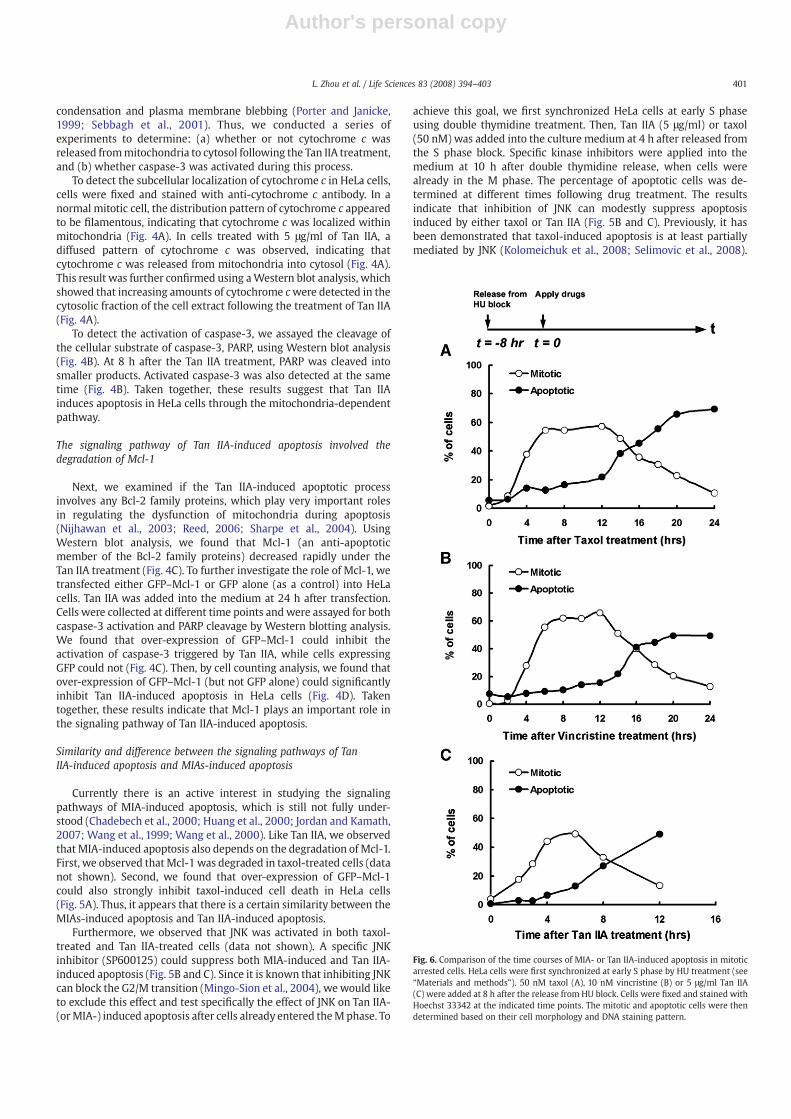
Fig. 6. Comparison of the time courses of MIA- or Tan IIA-induced apoptosis in mitoticarrested cells. HeLa cells were first synchronized at early S phase by HU treatment (see“Materials and methods”). 50 nM taxol (A), 10 nM vincristine (B) or 5 μg/ml Tan IIA(C) were added at 8 h after the release from HU block. Cells were fixed and stained withHoechst 33342 at the indicated time points. The mitotic and apoptotic cells were thendetermined based on their cell morphology and DNA staining pattern.
401L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
Our results are consistent with these findings. Furthermore, ourresults suggest that theremay be parallel apoptotic pathways betweencell death induced by MIA and Tan IIA.
To further compare these apoptotic pathways, we investigated theeffect of Cdk1 (cyclin-dependent kinase 1) in Tan IIA-induced celldeath. It has been reported that Cdk1 plays an important role in MIAs-induced apoptosis (Castedo et al., 2002; Chadebech et al., 2000; Huanget al., 2000; Yu et al., 1998). We found that Cdk1 activity was elevatedin the Tan IIA-treated cells in comparison to control cells (data notshown). Using a protocol similar to that of the JNK experiment, wetested if application of Cdk1 inhibitor (roscovitine, 25 μM) couldsuppress Tan IIA-induced apoptosis. The results are shown in Fig. 5C.Unlike the case of MIAs-induced apoptosis, inhibition of Cdk1 couldnot prevent Tan IIA-induced cell death. Instead, it slightly promotedTan IIA-induced apoptosis. So, there were both similarity anddifference between the apoptotic pathways of MIAs-induced andTan IIA-induced cell death. The signaling pathway of Tan IIA-inducedapoptosis based on our findings is summarized in Fig. 5D.
We have also conducted another study to compare the time courseof mitotic arrest and apoptosis induced either by Tan IIA or by MIAs,such as taxol and vincristine. HeLa cells were synchronized at S phaseby hydroxyurea (HU) treatment, and different drugs were applied 8 hafter release from the S phase block. The percentage of mitotic andapoptotic cells were determined at different time points. As shown inFig. 6A and B, both taxol and vincristine arrested cells in mitosis forabout 12 h before cell death began to occur. By comparison, under theTan IIA treatment, apoptosis occurred only 4 to 6 h after mitotic arrest(Fig. 6C). Thus, Tan IIA can induce cell death in mitotic arrested cellsfaster than MIAs and most likely utilize a slightly different apoptoticmechanism.
Discussion
The results of this study showed that Tan IIA can arrest HeLa cellsin mitosis and then induce them to enter apoptosis. Because of that,Tan IIA has a higher preference in killing cells in mitosis than cells ininterphase. Since most somatic cells in a human body are in thequiescent stages of the cell cycle, Tan IIA should have a preferentialcytotoxic effect on the actively dividing cancer cells over the somaticcells.
From our results, it appears that there are both similarities anddifferences between the apoptotic pathways of MIAs- and Tan IIA-induced cell death. First, both MIAs and Tan IIA-induced cell death aremainly through mitotic arrest. Second, both MIAs and Tan IIA-inducedapoptosis are mediated by the mitochondria-dependent pathway, asevident from cytochrome c release, caspase-3 activation and degrada-tion of Mcl-1. Third, like MIAs, Tan IIA-induced apoptosis is partiallymediated by JNK. However, unlike the MIAs-induced cell death, Cdk1does not appear to play a positive role in Tan IIA-induced apoptosis.
In comparison with the MIAs-based anti-cancer drugs, Tan IIAappears to have some potential advantages. First, unlike the MIAs, TanIIA does not interfere with the microtubule structure in the interphasecells (Fig. 3A), and thus should have less undesirable side effects.(Note: This is most likely because Tan IIA does not interfere with themicrotubule itself, instead, it appears to interact with certain MAPs(microtubule-associated protein) that are required formaintaining thestructure of the mitotic spindle). In addition, the required treatmenttime for Tan IIA could be shorter than the MIAs-based chemother-apeutic drugs. According to the results shown in Fig. 6, when HeLacells were treated with either taxol or vincristine, very little cell deathwas observed for the first 12 h following mitotic arrest. In contrast,under the Tan IIA treatment, cancer cells died 4 to 6 h after mitoticarrest. This finding suggests that Tan IIA can act faster in killingmitoticarrested cells than taxol or vincristine. On the other hand, there is alsoa disadvantage for Tan IIA. The concentration of Tan IIA required forkilling cancer cells appears to be much higher than that of MIAs.
Traditional Chinese medicine (TCM) is a very rich potential sourcefor finding new drugs. Many of these TCM plants had been used inhuman for many centuries, and thus, their toxicity was thoroughlytested. Tan IIA is purified from Danshen (Salvia miltiorrhiza Bunge), amedical plant of TCM widely used in the treatment of coronary arterydisease and cerebrovascular diseases over a thousand years in China,and it is expected to be relatively safe in applying to patients. Ourfindings that Tan IIA kills cancer cells through mitotic arrest can makeit an attractive drug lead for developing a novel class of anti-cancerdrugs. At this point, there are still some limitations in using Tan IIA forcancer treatment, which include: (a) The required dosage is stillrelatively high in comparison to the MIAs drugs. (b) The solubility ofTan IIA is not very good in the aqueous medium (such as the bloodserum). (c) The dosage of Tan IIA we tested so far can only kill about60–70% of HeLa cells. Thus, further chemical modification of Tan IIA isrequired to increase its solubility and apoptotic potency to generate afully useful anti-cancer drug. Alternatively, Tan IIA could be used as asupplementary anti-cancer drug in combination with other anti-cancer treatments.
Conclusion
Tan IIA is an extract from a traditional Chinese medicine that hadbeen used on patients for thousands of years.We found that treatmentof Tan IIA can arrest cancer cells in mitosis. Then, it can trigger themitotic arrested cells to undergo apoptosis through the mitochondria-dependent pathway. The mechanism of mitotic arrest was due to thedestruction of the mitotic spindle by Tan IIA. But unlike the existinganti-cancer drugs based on microtubule-interfering action, such asvincristine and taxol, Tan IIA destroyed only the mitotic spindle individing cells but did not affect microtubule instability in theinterphase cells. Furthermore, Tan IIA induced the mitotic arrestedcells to enter apoptosis faster than vincristine and taxol. Based onthese findings, we suggest that Tan IIA could be a candidate for a newclass of anti-cancer drugs.
Acknowledgments
We thank Dr. S. W. Edwards for providing us the Mcl-1 gene. Wealso thank Ms. Vivian Yu for her technical assistance. This work wassupported by the Research Grants Council of Hong Kong (HKUST6466/05M, N_HKUST616/05 and 660207).
References
Akgul, C., Moulding, D.A., White, M.R., Edwards, S.W., 2000. In vivo localisation andstability of human Mcl-1 using green fluorescent protein (GFP) fusion proteins.FEBS Letters 478 (1–2), 72–76.
Bhalla, K.N., 2003. Microtubule-targeted anticancer agents and apoptosis. Oncogene22 (56), 9075–9086.
Budihardjo, I., Oliver, H., Lutter, M., Luo, X., Wang, X., 1999. Biochemical pathways ofcaspase activation during apoptosis. Annual Review of Cell and DevelopmentalBiology 15, 269–290.
Burkhardt, J.K., 1998. The role of microtubule-based motor proteins in maintainingthe structure and function of the Golgi complex. Biochimica et Biophysica Acta1404 (1–2), 113–126.
Castedo, M., Perfettini, J.L., Roumier, T., Kroemer, G., 2002. Cyclin-dependent kinase-1:linking apoptosis to cell cycle and mitotic catastrophe. Cell Death and Differentia-tion 9 (12), 1287–1293.
Chadebech, P., Truchet, I., Brichese, L., Valette, A., 2000. Up-regulation of cdc2 proteinduring paclitaxel-induced apoptosis. International Journal of Cancer 87 (6),779–786.
Degterev, A., Boyce, M., Yuan, J., 2003. A decade of caspases. Oncogene 22 (53),8543–8567.
Green, D.R., Reed, J.C., 1998. Mitochondria and apoptosis. Science 281 (5381),1309–1312.
Huang, T.S., Shu, C.H., Chao, Y., Chen, S.N., Chen, L.L., 2000. Activation of MAD 2checkprotein and persistence of cyclin B1/CDC 2 activity associate with paclitaxel-induced apoptosis in human nasopharyngeal carcinoma cells. Apoptosis 5 (3),235–241.
Jordan, M.A., Kamath, K., 2007. How domicrotubule-targeted drugs work? An overview.Current Cancer Drug Targets 7 (8), 730–742.
402 L. Zhou et al. / Life Sciences 83 (2008) 394–403

Author's personal copy
Karki, S., Holzbaur, E.L., 1999. Cytoplasmic dynein and dynactin in cell division andintracellular transport. Current Opinion in Cell Biology 11 (1), 45–53.
Kolomeichuk, S.N., Terrano, D.T., Lyle, C.S., Sabapathy, K., Chambers, T.C., 2008. Distinctsignaling pathways of microtubule inhibitors—vinblastine and Taxol induce JNK-dependent cell death but through AP-1-dependent and AP-1-independentmechanisms, respectively. The FEBS Journal 275 (8), 1889–1899.
Levy, J.R., Holzbaur, E.L., 2006. Cytoplasmic dynein/dynactin function and dysfunctionin motor neurons. International Journal of Developmental Neuroscience 24 (2–3),103–111.
Li, P., Nijhawan, D., Budihardjo, I., Srinivasula, S.M., Ahmad, M., Alnemri, E.S., Wang, X.,1997. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complexinitiates an apoptotic protease cascade. Cell 91 (4), 479–489.
Meng, W., Yang, Y., Deng, C., Liu, T., Jia, Y., 2002. Study on the relationship between NB4cell apoptosis induced by tanshinone IIA and the cell mitochondrial transmem-brane potential. Zhonghua Xue Ye Xue Za Zhi 23 (6), 297–300.
Mingo-Sion, A.M., Marietta, P.M., Koller, E., Wolf, D.M., Van Den Berg, C.L., 2004.Inhibition of JNK reduces G2/M transit independent of p53, leading to endor-eduplication, decreased proliferation, and apoptosis in breast cancer cells.Oncogene 23 (2), 596–604.
Mollinedo, F., Gajate, C., 2003. Microtubules, microtubule-interfering agents andapoptosis. Apoptosis 8 (5), 413–450.
Murray, J.W., Wolkoff, A.W., 2003. Roles of the cytoskeleton and motor proteins inendocytic sorting. Advanced Drug Delivery Reviews 55 (11), 1385–1403.
Nijhawan, D., Fang, M., Traer, E., Zhong, Q., Gao, W., Du, F., Wang, X., 2003. Elimination ofMcl-1 is required for the initiation of apoptosis following ultraviolet irradiation.Genes & Development 17 (12), 1475–1486.
Porter, A.G., Janicke, R.U., 1999. Emerging roles of caspase-3 in apoptosis. Cell Death andDifferentiation 6 (2), 99–104.
Reed, J.C., 2006. Proapoptotic multidomain Bcl-2/Bax-family proteins: mechanisms,physiological roles, and therapeutic opportunities. Cell Death and Differentiation13 (8), 1378–1386.
Riedl, S.J., Renatus, M., Schwarzenbacher, R., Zhou, Q., Sun, C., Fesik, S.W., Liddington,R.C., Salvesen, G.S., 2001. Structural basis for the inhibition of caspase-3 by XIAP.Cell 104 (5), 791–800.
Sebbagh, M., Renvoize, C., Hamelin, J., Riche, N., Bertoglio, J., Breard, J., 2001. Caspase-3-mediated cleavage of ROCK I induces MLC phosphorylation and apoptoticmembrane blebbing. Nature Cell Biology 3 (4), 346–352.
Selimovic, D., Hassan, M., Haikel, Y., Hengge, U.R., 2008. Taxol-induced mitochondrialstress in melanoma cells is mediated by activation of c-Jun N-terminal kinase (JNK)and p38 pathways via uncoupling protein 2. Cellular Signalling 20 (2), 311–322.
Sharpe, J.C., Arnoult, D., Youle, R.J., 2004. Control of mitochondrial permeability by Bcl-2family members. Biochimica et Biophysica Acta 1644 (2–3), 107–113.
Slee, E.A., Harte, M.T., Kluck, R.M., Wolf, B.B., Casiano, C.A., Newmeyer, D.D., Wang, H.G.,Reed, J.C., Nicholson, D.W., Alnemri, E.S., Green, D.R., Martin, S.J., 1999. Ordering thecytochrome c-initiated caspase cascade: hierarchical activation of caspases-2, -3,-6, -7, -8, and -10 in a caspase-9-dependent manner. The Journal of Cell Biology144 (2), 281–292.
Tang, Z., Tang, Y., Fu, L., 2003a. Growth inhibition and apoptosis induction in humanhepatoma cells by tanshinone II A. J Huazhong Univ Sci Technolog Med Sci 23 (2),166–168, 172.
Tang, Z.Z., Tang, Y., Fu, L.B., 2003b. Effect of tanshinone IIA on the growth behavior ofhuman hepatoma cell line BEL-7402 in vitro and its mechanism. Di Yi Jun Yi Da XueXue Bao 23 (6), 595–597, 601.
Wang, X., 2001. The expanding role of mitochondria in apoptosis. Genes & Development15 (22), 2922–2933.
Wang, L.G., Liu, X.M., Kreis, W., Budman, D.R., 1999. The effect of antimicrotubule agentson signal transduction pathways of apoptosis: a review. Cancer Chemotherapy andPharmacology 44 (5), 355–361.
Wang, T.H., Wang, H.S., Soong, Y.K., 2000. Paclitaxel-induced cell death: where the cellcycle and apoptosis come together. Cancer 88 (11), 2619–2628.
Yoon, Y., Kim, Y.O., Jeon, W.K., Park, H.J., Sung, H.J., 1999. Tanshinone IIA isolated fromSalvia miltiorrhiza BUNGE induced apoptosis in HL60 human premyelocyticleukemia cell line. Journal of Ethnopharmacology 68 (1–3), 121–127.
Yu, D., Jing, T., Liu, B., Yao, J., Tan, M., McDonnell, T.J., Hung, M.C., 1998. Overexpression ofErbB2 blocks Taxol-induced apoptosis by upregulation of p21Cip1, which inhibitsp34Cdc2 kinase. Mol Cell 2 (5), 581–591.
Yuan, S.L., Wang, X.J., Wei, Y.Q., 2003. Anticancer effect of tanshinone and itsmechanisms. Ai Zheng 22 (12), 1363–1366.
Yuan, S.L., Wei, Y.Q., Wang, X.J., Xiao, F., Li, S.F., Zhang, J., 2004. Growth inhibition andapoptosis induction of tanshinone II-A on human hepatocellular carcinoma cells.World Journal of Gastroenterology 10 (14), 2024–2028.
403L. Zhou et al. / Life Sciences 83 (2008) 394–403
Copyright © 2022 FDOKUMEN




















