
Curcumin protects against ischemia/reperfusion injury in rat kidneys
Upload
independentCategory
view
2download
0
Journal of Surgical Research 155, 345–356 (2009)doi:10.1016/j.jss.2008.07.013
RESEARCH REVIEW
Systemic Administration of Interleukin-10 Attenuates Early Ischemic
Response Following Spinal Cord Ischemia Reperfusion Injury in Rats
H. Hakan Oruckaptan, M.D., Ph.D,*,1 Pinar Ozisik, M.D., Ph.D.,* Pergin Atilla, M.D.,† Muruvet Tuncel, M.D.,‡Kamer Kilinc, M.D.,§ Pinar Ozdemir Geyik, Ph.D.,k Nursen Basaran, M.D.,{ Eser Yuksel, M.D.,#
and O. Ekin Ozcan, M.D.*
*Department of Neurosurgery; †Department of Histology and Embryology; ‡Department of Anatomy; §Department of Biochemistry;kDepartment of Biostatistics; {Department of Pharmacology, Division of Toxicology, Hacettepe University School of Medicine,
Ankara, Turkey; and #Baylor College of Medicine, Division of Plastic and Reconstructive Surgery, Houston, Texas
Submitted for publication April 8, 2008
Background. The aim of this experimental study wasto investigate the early effects of interleukin-10 (IL-10)and interleukin-1b antagonist (anti-IL-1b) againstcellular damage, inflammatory reactivity, lipid peroxi-dation (LPO), and myeloperoxidase (MPO) activity in-duced by spinal cord ischemia reperfusion injury (IRI).
Methods. Thirty-two single strain female Albino ratswere divided into four groups: control (sham-oper-ated), IRI-alone, IL-10-treated (100 mg/kg), and anti-IL-1b-treated (1 mg/kg) groups after IRI. IRI was inducedby balloon occlusion of the aorta and simultaneoushypovolemia during occlusion. The animals were sacri-ficed at 24 h. Histopathological and ultrastructuralanalyses, biochemical studies for determination ofLPO and MPO activity and Comet assays (single cellelectrophoresis for detecting DNA single strandbreaks) were performed in all study groups.
Results. Compared with the levels of control (sham-operated) animals, IRI produced a significant increasein the levels of LPO and MPO activity, and prominenttissue damage characterized by leukocyte infiltration,edema and neuronal and glial damage in the affectedspinal cord in 24 h. The administration of IL-10 de-creased LPO and MPO activity, and suppressed initialinflammatory response in the first 24 h. The effects ofanti-IL-1b were limited to decrease in LPO activitywithout considerable evidence of cellular preservation.
Conclusions. These data suggest that systemicadministration of IL-10 attenuates the early ischemicresponse, and may restrict the tissue damage in the
1 To whom correspondence and reprint requests should be ad-dressed at Neurosurgery Department, Hacettepe University, Schoolof Medicine, (Zone 71), Hacettepe, Sihhiye, Ankara, 06100, Turkey.E-mail: [email protected].
345
first 24 h after spinal cord ischemia reperfusion injury.Anti-IL-1b has no considerable effect in this time win-dow. The results of this preliminary study promote fur-ther studies with longer time windows on the effects ofanti-inflammatory cytokines in spinal cord IRI. � 2009
Elsevier Inc. All rights reserved.
Keywords: spinal cord; ischemia; reperfusion injury;cytokine; interleukin-10; interleukin-1b; inflamma-tion; central nervous system; lipid peroxidation; mye-loperoxidase.
INTRODUCTION
Spinal cord ischemia reperfusion injury (IRI) is a dev-astating complication of thoraco-abdominal aortic sur-gery, and can be occasionally seen after surgical orendovascular treatment of spinal vascular lesions. Inthe central nervous system (CNS), any serious decreasein the blood flow causes energy failure and triggersa cascade of events leading to cell damage through am-plification of many pathways set in motion by ischemia.The early accumulation of leukocytes in and around themicrovessels at the ischemic zones has been clearlydemonstrated by many investigators [1–5]. Expressionof proinflammatory cytokines (IL-1, TNF-a, interferon-g) and up-regulation of adhesion receptors play animportant role in the early and late course of ischemiccascade [6–8]. Interleukin-1 family cytokines includethe secreted proinflammatory agonist interleukin-1beta (IL-1b), interleukin-18 (IL-18), and anti-inflamma-tory receptor antagonist interleukin-1 alpha (IL-1a).
0022-4804/09 $36.00� 2009 Elsevier Inc. All rights reserved.

JOURNAL OF SURGICAL RESEARCH: VOL. 155, NO. 2, AUGUST 2009346
Constitutive expression of IL-1 is low in normal brain;however, there is considerable evidence for the in-creased expression of IL-1b and IL-1ra mRNA from vas-cular endothelium, astrocytes, microglia, and neuronsduring the early course of cerebral ischemia [9, 10].The releasing of IL-1 from these cells increases endo-thelial permeability and expression of adhesionmolecules. Increased endothelial permeability and up-regulation of adhesion molecules promote neutrophilmigration from the circulation and phagocytosis [11].Several lines of evidence suggest that IL-1 also inducesthe synthesis of IL-6, arachidonic acid, and nitric oxide(NO), and increases glutamate mediated toxicity in theCNS [9, 10, 12–14].
IL-10 is an anti-inflammatory cytokine, synthesizedin normal brain from astrocytes and microglia. Endog-enous IL-10 expression strongly increases in variousdiseases of the CNS such as trauma, inflammation,and ischemia. Excessive production of IL-10 has alsobeen reported in the heart, liver, and kidney duringthe early course of IRI [15, 16]. Several data in the liter-ature suggest the inhibitory effects of IL-10 over thesynthesis of proinflammatory cytokines and nitric oxide(NO), IF-g expression and glutamate-mediated excito-toxicity in the brain [17–19] . The role of cytokines inIRI has been increasingly investigated in many organssuch as the brain, heart, liver, and kidney. However,there are limited data about the role of these cytokinesin the spinal cord despite its vulnerability to IRI. Ourstudy was therefore initiated to determine the early ef-fects of systemic administration of anti-IL-1b and IL-10in an experimental model of spinal cord IRI. We hypoth-esized that anti-IL-1b and IL-10 might limit the earlytissue damage caused by spinal cord IRI.
MATERIALS AND METHODS
The current experiment was undertaken with the approval of theAnimal Care and Use Committee of Hacettepe University. The animalcare complied with the Guide for the Care and Use of Laboratory An-imals published by the National Institute of Health (NIH publication5377-3, 1996). Thirty-two single strain female Albino rats weighingbetween 200 and 260 g (mean 220 g) were used in this study.
Surgical Procedure
General anesthesia was induced by intraperitoneal (i.p.) adminis-tration of a mixture of ketamine hydrochloride 40 mg/kg (Ketalar;Parke-Davis Eczacibasi, Istanbul, Turkey) and xylazine 5 mg/kg(Rompun 2%; Bayer, Istanbul, Turkey). The core temperature wascontinuously monitored, with a flexible rectal probe, and maintainedbetween 37
�C to 38.5
�C by a thermal pad and a heating lamb through-
out the procedure. A transient aortic balloon occlusion model via theleft common carotid artery (LCCA) catheterization was used for theinduction of lower thoracic and lumbar spinal cord ischemia-reperfu-sion injury as described by Kanellopoulos et al. [20]. The animals werepinned in supine position after the induction of general anesthesia.The left femoral artery (LFA) was cannulated with a 24 G catheterfor the induction of simultaneous hypovolemia and distal blood
pressure monitoring. A midline skin incision over the neckwas made to exposure the left common carotid artery (LCCA).The LCAA was freed from the adjacent jugular vein and vagus nerveunder magnification (Zeiss OPMI 99; Carl Zeiss, Oberkochen,Germany).
Induction of Ischemia Reperfusion Injury (IRI)
Following the systemic administration of heparin (20 U/100 g i.v.),a 1.8 F single lumen balloon catheter (Magellan-Balt, Paris, France)was advanced to the descending aorta (6 cm proximal from insertion)through the LCCA. It was inflated with 0.1 cc physiological saline (SF)and then pulled back gradually up to the origin of the left subclavianartery (LSA). At this step, the intraaortic balloon was re-inflated with0.2 mL SF (total 0.3 cc) and kept for 10 min for temporary occlusion ofthe descending aorta. Simultaneous hypovolemia was induced bywithdrawing a blood volume of 1 cc/100 g weight (mean 2.2 mL per an-imal) through the LFA. As soon as the aortic occlusion was termi-nated, induced hypovolemia was restored with reinfusion ofwithdrawn blood through the LFA. The heparin was neutralizedwith protamine sulphate, and the exposed arteries were tied upwith a 4.0 silk suture before the eventual wound closure.
Description of Groups
Four groups were composed of the following: Group 1: control group(sham-operated, n ¼ 8): All vessels were cannulated as describedabove. They were ligated without performing an aortic occlusion pro-cedure. All animals subjected to the same aesthetic procedures, drugand fluid administrations including heparin and protamine sulphatewith the same doses. Group 2: IRI (n¼ 8): ischemia reperfusion injurywas induced as described above. Group 3: IL-10 treated group (n¼ 8):recombinant rat IL-10 in a dose of 50 mg/kg (RDI-4019; Research Di-agnostics Inc., Flanders, NJ) was intraperitoneally administeredtwice, 2 min and 6 h after IRI (total 100 mg/kg).
Group 4: anti-IL-1b-treated group (n ¼ 8): recombinant anti-inter-leukin-1b in a dose of 0.5 mg/kg (I-3767; Sigma-Aldrich, St. Louis,MO) was intraperitoneally administered twice, 2 min and 6 h afterIRI (total 1 mg/kg).
Follow-up
The animals were housed in cages with an ambient temperature ofapproximately 21�C soon after the complete recovery from anesthesia.A 6-point lower extremity walking scale was used for neurologicalevaluation during the experiment [21]. All animals were given a ‘mo-tor deficit score’ (MDS) at the beginning of the procedure (which waszero in all animals), 1 h after the balloon occlusion (early MDS), and24 h later (late MDS), respectively, according to the sum of the follow-ing two main criteria; A: walking with the lower limbs (0 ¼ normal;1 ¼ legs are weak, but walks normally; 2 ¼ ataxic, but able to walkon the toes; 3 ¼ improper steeping, but able to walk on the knuckles;4 ¼ no significant movement in the lower limbs), B: placing and step-ping (0 ¼ normal; 1 ¼ weak; 2 ¼ no stepping, either spasticity of flac-cidity were present) (AþB: between 0-6). Qualifying was always madeby two researchers, one of whom was unaware of the initial surgicaland therapeutic step. The difference between late and early MDSswas recorded as ‘motor recovery score’ (MRS) of each animal.
Obtaining Samples from the Spinal Cord
Twenty-four hours after the procedure, anesthesia was reinduced.The animals were fixed by transcardial perfusion of 100 mL physiolog-ical saline and 200 mL 10% buffered paraformaldehyde. Approxi-mately 20 mm of spinal cord segments between T7 and T13 levelswere obtained for histological, ultrastructural, biochemical, and mo-lecular analyses. All samples were kept in randomly numbered con-tainers and transferred to the blinded observers. Owing to thedistinct rostrocaudal gradient of increasing tissue damage in the

TABLE 1
Ultrastructural Grading System: The Sum of A, B,and C Reflects General Neuron Score, and Was Usedfor the Evaluation of the Gray Matter. D and E WereUsed for the White matter, and F Was Done for the En-dothelium
Pathology Degree Score
A- Intracytoplasmicedema: 25 neuronsfor each samplewere evaluated
Absent 0Light 1Mild 2Severe 3
B- Nucleus: 25 neuronsfor each samplewere evaluated
Normal 0Clumping 1Sparse chromatin 2Nuclear membrane defect 3
C- Mitochondrion:100 mitochondria in25 neurons of eachsample were evaluated
Normal 0Light edema 1Mild edema 2No structure inside 3
D- Axon general score:25 axons in each samplewere evaluated
Normal 0Light edema 1Mild edema 2No structure inside 3
E- Myelin score: 25axons in each samplewere evaluated
Normal 0Cleft between the layers 1Slender myelin 2Abundantly cracked myelin 3
F- Endothelium score:25 endothelial cell ineach sample wereevaluated
Normal 0Light edema 1Mild edema 2Complete disruption
of integrity3
ORUCKAPTAN ET AL.: SYSTEMIC ADMINISTRATION OF INTERLEUKIN-10 347
aortic occlusion model, tissue samples from the same level were as-signed to the identical histological or biochemical study in all animals.The samples from the lowermost level of the spinal cord were used forhistopathological and ultrastructural analyses, while the samplesfrom upper levels were used for the determination of LPO and MPOactivity as well as DNA damage in turn from caudal to rostral direc-tion.
Determination of Malondialdehyde (MDA) Levels
The fresh tissue samples from the similar levels of the harvestedspinal cords were stored at –80�C for assays of MDA. Tissue lipid per-oxidation (LPO), as an indirect indicator of free radical formation, wasmeasured by using the thiobarbituric acid method of Uchiyama andMihara (nmol MDA/g wet tissue) [22].
Determination of Myeloperoxidase (MPO) Activity
Tissue MPO activity was determined by measuring the H2O2-de-pendent oxidation of O-dianisidine as described by Glowick and Ka-plan [23]. The amount of MPO caused an absorbance of 1.0 per minat 410 nm and reflected one unit of enzyme activity (U/g).
Histological Examination
The tissue samples from the similar levels of the harvested spinalcords were fixed in 10% buffered formalin for 4 to 10 d before theywere embedded in paraffin. For each animal, transverse sections in1.6 mm thickness were obtained and stained with hematoxylin and eo-sin (H and E) and Luxol fast blue. The architecture of the white andgray matter, vascular mash, neuronal and glial cell damage and in-flammatory cell accumulation in the spinal cord were analyzed andphotographed under the 340 to 3400 magnifications. Besides thequalitative analysis of each slide, quantitative assessments weredone to increase the reliability of results. The presence of two ormore of the findings including intracellular edema, chromatolysis, nu-clear hyperchromasia, loss of intracytoplasmic organelles, and/or pyk-nosis, even mild or moderate was accepted as sufficient criteria forcellular disintegration. These cells were assigned as ‘damaged’, whilethe remaining were as ‘normal’ during the quantitative analysis oflight microscopic findings. A detailed cell count, by using an ocular mi-crometer (Olympus BH-2; Olympus, Tokyo, Japan), was done in theanterior, middle and posterior one third of the each spinal cord sec-tions separately in order to verify regional differences of cellular dis-integration in the axial plane. Glial cell damage ratio (GDR) wascalculated by dividing the number of disintegrated glial cells by thetotal number of glial cells counted within the entire microscopic sec-tion. Neuronal damage ratio (NDR) was determined in the same man-ner. Inflammatory cell ratio (ICR) was determined by dividing thenumber of inflammatory cells by the total number of cells (neurons,glia, and inflammatory cells) counted within the entire microscopicsection.
Ultrastructural Analysis (Transmission Electron Microscopy)
The fresh tissue samples from the similar levels of the harvestedspinal cords were fixed in 2.5% glutaraldehyde for 24 h, postfixed inOsO4 for 1 h before gradual dehydration with alcohol. The propyl-ene-treated samples were embedded in Araldite CY212. One to twomm semi-thin sections were stained with methylene-blue for light mi-croscopic examination under the 31000 magnification (Nikon Opti-phot, Tokyo, Japan). Ultra-thin sections were collected on coopergrids, stained with uranyl acetate and lead citrate and examined un-der the 35000 and 310,000 magnifications (JEOL JEM 1200 EX, To-kyo, Japan). Besides the qualitative analysis of the samples, anultrastructural grading system was used for quantitative evaluation[24]. Cytoplasmic edema in 25 neurons, nuclear integrity in 25 neu-
rons, 100 mitochondria in 25 neurons, axonal degeneration, myelindamage, and endothelial integrity in 25 structures were evaluatedseparately and scored from 0 to 3 for each sample (Table 1). The re-sults were statistically analyzed for the determination of variationsamong the study groups.
DNA Fragmentation (Comet Assay)
The technique known as the Comet assay (single cell electrophore-sis for detecting DNA single strand breaks) or single cell gel electro-phoresis was used to determine the degree of DNA damage inindividual cells. The fresh tissue samples from the similar levels ofthe harvested spinal cords were kept at 4�C and transferred for Cometassay. The determination of Comet assay was made with basic alka-line technique [25, 26]. In this technique, a suspension of cells was em-bedded in agarose on microscope slides, lysed and subjected toelectrophoresis, and then stained with a fluorescent DNA bindingdye. During alkaline gel electrophoresis, single-stranded DNA frag-ments can migrate out of the nucleus towards the anode and forma comet according to the extent of damage. For visualization of DNAdamage, slides were examined at 10003 magnification usinga 1003 objective (oil immersion) on a fluorescence microscope (Zeiss).Images obtained from randomly selected 100 cells of each animal wereanalyzed. DNA fragmentation was graded in four stages according tothe degree of comet formation; grade 1 (normal or minimal damage0%–25%), grade 2 (mild damage, 26%–50%), grade 3 (moderate dam-age, 51%–75%), grade 4 (serious damage, 76%–100%). Total cell countin each grade was multiplied with a corresponding coefficient (one forgrade 1 and four for grade 4) to determine total weighted score of DNAinjury (TWSD) in each animal. The sums of TWSDs were statisticallyanalyzed for the determination of variations in the study groups.
JOURNAL OF SURGICAL RESEARCH: VOL. 155, NO. 2, AUGUST 2009348
Statistical Analysis
All the data from the experiment were coded, recorded, and ana-lyzed by using SPSS software ver. 11.5 (SPSS Inc, Chicago, IL). Krus-kal-Wallis followed by Mann-Whitney-U tests were used for theanalyses of motor deficit and mean recovery scores (MDS and MRS)of the animals and MDA and MPO levels. For ultrastructural findingsand COMET assay data, Kruskal-Wallis and following KW-Multiplecomparison tests (Tukey HSD) were done to demonstrate the differ-ences among the study groups. Two-way ANOVA and multiple com-parison tests were used for the analyses of glial and neuronaldegeneration and inflammatory cell infiltration. A P value of < 0.05was accepted significant. The data were expressed as the mean value6 SD.
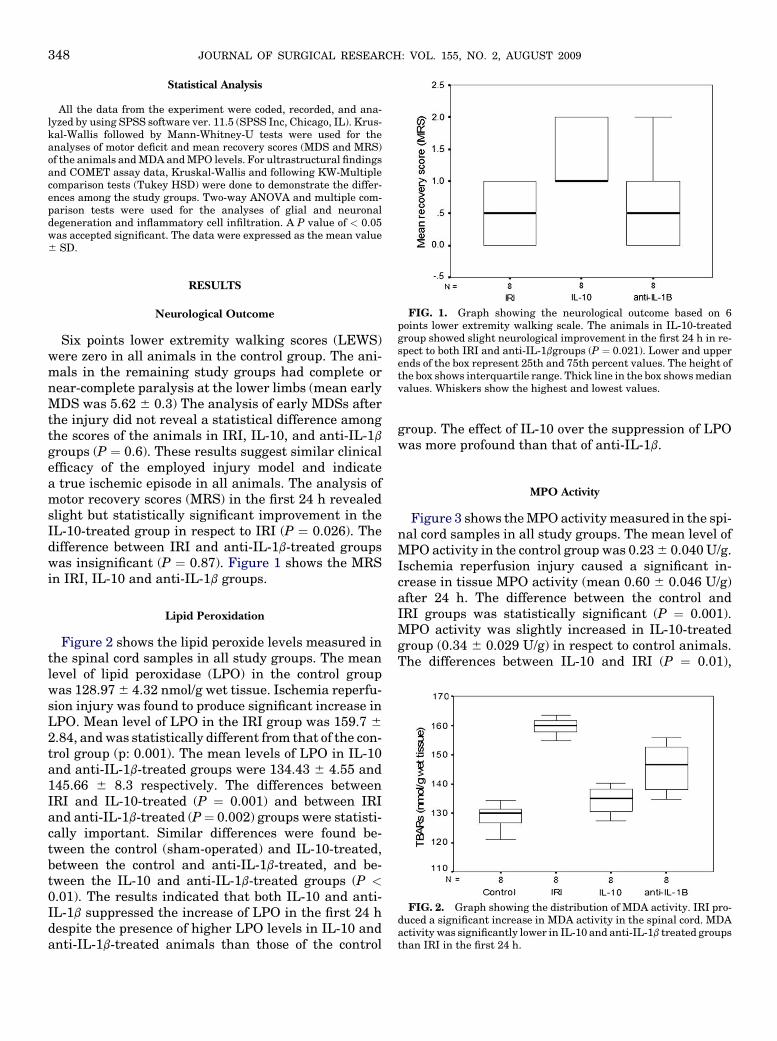
FIG. 1. Graph showing the neurological outcome based on 6points lower extremity walking scale. The animals in IL-10-treatedgroup showed slight neurological improvement in the first 24 h in re-spect to both IRI and anti-IL-1bgroups (P ¼ 0.021). Lower and upperends of the box represent 25th and 75th percent values. The height ofthe box shows interquartile range. Thick line in the box shows medianvalues. Whiskers show the highest and lowest values.
RESULTS
Neurological Outcome
Six points lower extremity walking scores (LEWS)were zero in all animals in the control group. The ani-mals in the remaining study groups had complete ornear-complete paralysis at the lower limbs (mean earlyMDS was 5.62 6 0.3) The analysis of early MDSs afterthe injury did not reveal a statistical difference amongthe scores of the animals in IRI, IL-10, and anti-IL-1b
groups (P ¼ 0.6). These results suggest similar clinicalefficacy of the employed injury model and indicatea true ischemic episode in all animals. The analysis ofmotor recovery scores (MRS) in the first 24 h revealedslight but statistically significant improvement in theIL-10-treated group in respect to IRI (P ¼ 0.026). Thedifference between IRI and anti-IL-1b-treated groupswas insignificant (P ¼ 0.87). Figure 1 shows the MRSin IRI, IL-10 and anti-IL-1b groups.
FIG. 2. Graph showing the distribution of MDA activity. IRI pro-duced a significant increase in MDA activity in the spinal cord. MDAactivity was significantly lower in IL-10 and anti-IL-1b treated groupsthan IRI in the first 24 h.
Lipid Peroxidation
Figure 2 shows the lipid peroxide levels measured inthe spinal cord samples in all study groups. The meanlevel of lipid peroxidase (LPO) in the control groupwas 128.97 6 4.32 nmol/g wet tissue. Ischemia reperfu-sion injury was found to produce significant increase inLPO. Mean level of LPO in the IRI group was 159.7 6
2.84, and was statistically different from that of the con-trol group (p: 0.001). The mean levels of LPO in IL-10and anti-IL-1b-treated groups were 134.43 6 4.55 and145.66 6 8.3 respectively. The differences betweenIRI and IL-10-treated (P ¼ 0.001) and between IRIand anti-IL-1b-treated (P¼ 0.002) groups were statisti-cally important. Similar differences were found be-tween the control (sham-operated) and IL-10-treated,between the control and anti-IL-1b-treated, and be-tween the IL-10 and anti-IL-1b-treated groups (P <0.01). The results indicated that both IL-10 and anti-IL-1b suppressed the increase of LPO in the first 24 hdespite the presence of higher LPO levels in IL-10 andanti-IL-1b-treated animals than those of the control
group. The effect of IL-10 over the suppression of LPOwas more profound than that of anti-IL-1b.
MPO Activity
Figure 3 shows the MPO activity measured in the spi-nal cord samples in all study groups. The mean level ofMPO activity in the control group was 0.23 6 0.040 U/g.Ischemia reperfusion injury caused a significant in-crease in tissue MPO activity (mean 0.60 6 0.046 U/g)after 24 h. The difference between the control andIRI groups was statistically significant (P ¼ 0.001).MPO activity was slightly increased in IL-10-treatedgroup (0.34 6 0.029 U/g) in respect to control animals.The differences between IL-10 and IRI (P ¼ 0.01),
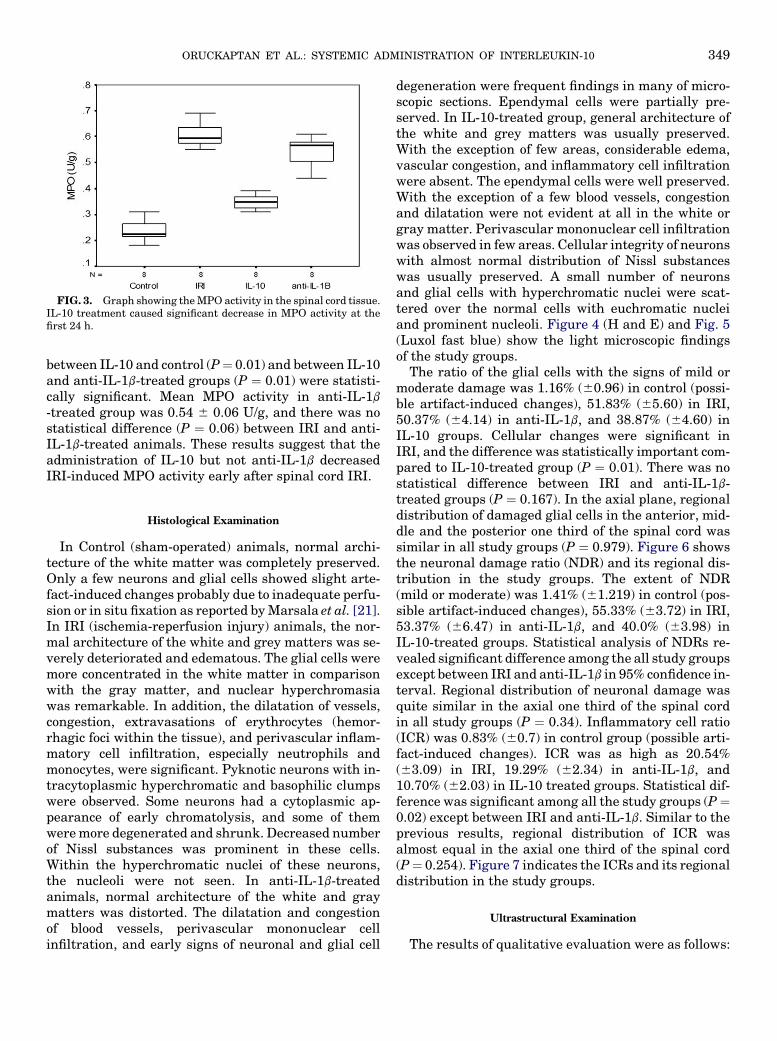
FIG. 3. Graph showing the MPO activity in the spinal cord tissue.IL-10 treatment caused significant decrease in MPO activity at thefirst 24 h.
ORUCKAPTAN ET AL.: SYSTEMIC ADMINISTRATION OF INTERLEUKIN-10 349
between IL-10 and control (P¼ 0.01) and between IL-10and anti-IL-1b-treated groups (P ¼ 0.01) were statisti-cally significant. Mean MPO activity in anti-IL-1b
-treated group was 0.54 6 0.06 U/g, and there was nostatistical difference (P ¼ 0.06) between IRI and anti-IL-1b-treated animals. These results suggest that theadministration of IL-10 but not anti-IL-1b decreasedIRI-induced MPO activity early after spinal cord IRI.
Histological Examination
In Control (sham-operated) animals, normal archi-tecture of the white matter was completely preserved.Only a few neurons and glial cells showed slight arte-fact-induced changes probably due to inadequate perfu-sion or in situ fixation as reported by Marsala et al. [21].In IRI (ischemia-reperfusion injury) animals, the nor-mal architecture of the white and grey matters was se-verely deteriorated and edematous. The glial cells weremore concentrated in the white matter in comparisonwith the gray matter, and nuclear hyperchromasiawas remarkable. In addition, the dilatation of vessels,congestion, extravasations of erythrocytes (hemor-rhagic foci within the tissue), and perivascular inflam-matory cell infiltration, especially neutrophils andmonocytes, were significant. Pyknotic neurons with in-tracytoplasmic hyperchromatic and basophilic clumpswere observed. Some neurons had a cytoplasmic ap-pearance of early chromatolysis, and some of themwere more degenerated and shrunk. Decreased numberof Nissl substances was prominent in these cells.Within the hyperchromatic nuclei of these neurons,the nucleoli were not seen. In anti-IL-1b-treatedanimals, normal architecture of the white and graymatters was distorted. The dilatation and congestionof blood vessels, perivascular mononuclear cellinfiltration, and early signs of neuronal and glial cell
degeneration were frequent findings in many of micro-scopic sections. Ependymal cells were partially pre-served. In IL-10-treated group, general architecture ofthe white and grey matters was usually preserved.With the exception of few areas, considerable edema,vascular congestion, and inflammatory cell infiltrationwere absent. The ependymal cells were well preserved.With the exception of a few blood vessels, congestionand dilatation were not evident at all in the white orgray matter. Perivascular mononuclear cell infiltrationwas observed in few areas. Cellular integrity of neuronswith almost normal distribution of Nissl substanceswas usually preserved. A small number of neuronsand glial cells with hyperchromatic nuclei were scat-tered over the normal cells with euchromatic nucleiand prominent nucleoli. Figure 4 (H and E) and Fig. 5(Luxol fast blue) show the light microscopic findingsof the study groups.
The ratio of the glial cells with the signs of mild ormoderate damage was 1.16% (60.96) in control (possi-ble artifact-induced changes), 51.83% (65.60) in IRI,50.37% (64.14) in anti-IL-1b, and 38.87% (64.60) inIL-10 groups. Cellular changes were significant inIRI, and the difference was statistically important com-pared to IL-10-treated group (P ¼ 0.01). There was nostatistical difference between IRI and anti-IL-1b-treated groups (P ¼ 0.167). In the axial plane, regionaldistribution of damaged glial cells in the anterior, mid-dle and the posterior one third of the spinal cord wassimilar in all study groups (P ¼ 0.979). Figure 6 showsthe neuronal damage ratio (NDR) and its regional dis-tribution in the study groups. The extent of NDR(mild or moderate) was 1.41% (61.219) in control (pos-sible artifact-induced changes), 55.33% (63.72) in IRI,53.37% (66.47) in anti-IL-1b, and 40.0% (63.98) inIL-10-treated groups. Statistical analysis of NDRs re-vealed significant difference among the all study groupsexcept between IRI and anti-IL-1b in 95% confidence in-terval. Regional distribution of neuronal damage wasquite similar in the axial one third of the spinal cordin all study groups (P ¼ 0.34). Inflammatory cell ratio(ICR) was 0.83% (60.7) in control group (possible arti-fact-induced changes). ICR was as high as 20.54%(63.09) in IRI, 19.29% (62.34) in anti-IL-1b, and10.70% (62.03) in IL-10 treated groups. Statistical dif-ference was significant among all the study groups (P¼0.02) except between IRI and anti-IL-1b. Similar to theprevious results, regional distribution of ICR wasalmost equal in the axial one third of the spinal cord(P¼ 0.254). Figure 7 indicates the ICRs and its regionaldistribution in the study groups.
Ultrastructural Examination
The results of qualitative evaluation were as follows:
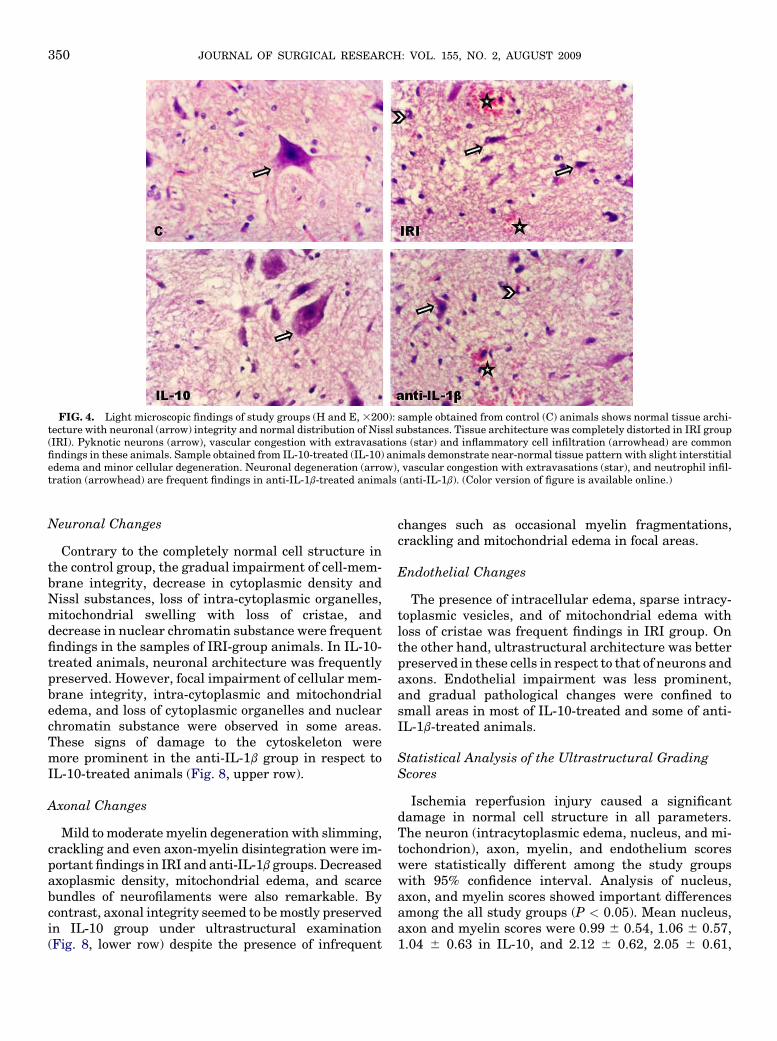
FIG. 4. Light microscopic findings of study groups (H and E, 3200): sample obtained from control (C) animals shows normal tissue archi-tecture with neuronal (arrow) integrity and normal distribution of Nissl substances. Tissue architecture was completely distorted in IRI group(IRI). Pyknotic neurons (arrow), vascular congestion with extravasations (star) and inflammatory cell infiltration (arrowhead) are commonfindings in these animals. Sample obtained from IL-10-treated (IL-10) animals demonstrate near-normal tissue pattern with slight interstitialedema and minor cellular degeneration. Neuronal degeneration (arrow), vascular congestion with extravasations (star), and neutrophil infil-tration (arrowhead) are frequent findings in anti-IL-1b-treated animals (anti-IL-1b). (Color version of figure is available online.)
JOURNAL OF SURGICAL RESEARCH: VOL. 155, NO. 2, AUGUST 2009350
Neuronal Changes
Contrary to the completely normal cell structure inthe control group, the gradual impairment of cell-mem-brane integrity, decrease in cytoplasmic density andNissl substances, loss of intra-cytoplasmic organelles,mitochondrial swelling with loss of cristae, anddecrease in nuclear chromatin substance were frequentfindings in the samples of IRI-group animals. In IL-10-treated animals, neuronal architecture was frequentlypreserved. However, focal impairment of cellular mem-brane integrity, intra-cytoplasmic and mitochondrialedema, and loss of cytoplasmic organelles and nuclearchromatin substance were observed in some areas.These signs of damage to the cytoskeleton weremore prominent in the anti-IL-1b group in respect toIL-10-treated animals (Fig. 8, upper row).
Axonal Changes
Mild to moderate myelin degeneration with slimming,crackling and even axon-myelin disintegration were im-portant findings in IRI and anti-IL-1b groups. Decreasedaxoplasmic density, mitochondrial edema, and scarcebundles of neurofilaments were also remarkable. Bycontrast, axonal integrity seemed to be mostly preservedin IL-10 group under ultrastructural examination(Fig. 8, lower row) despite the presence of infrequent
changes such as occasional myelin fragmentations,crackling and mitochondrial edema in focal areas.
Endothelial Changes
The presence of intracellular edema, sparse intracy-toplasmic vesicles, and of mitochondrial edema withloss of cristae was frequent findings in IRI group. Onthe other hand, ultrastructural architecture was betterpreserved in these cells in respect to that of neurons andaxons. Endothelial impairment was less prominent,and gradual pathological changes were confined tosmall areas in most of IL-10-treated and some of anti-IL-1b-treated animals.
Statistical Analysis of the Ultrastructural GradingScores
Ischemia reperfusion injury caused a significantdamage in normal cell structure in all parameters.The neuron (intracytoplasmic edema, nucleus, and mi-tochondrion), axon, myelin, and endothelium scoreswere statistically different among the study groupswith 95% confidence interval. Analysis of nucleus,axon, and myelin scores showed important differencesamong the all study groups (P < 0.05). Mean nucleus,axon and myelin scores were 0.99 6 0.54, 1.06 6 0.57,1.04 6 0.63 in IL-10, and 2.12 6 0.62, 2.05 6 0.61,
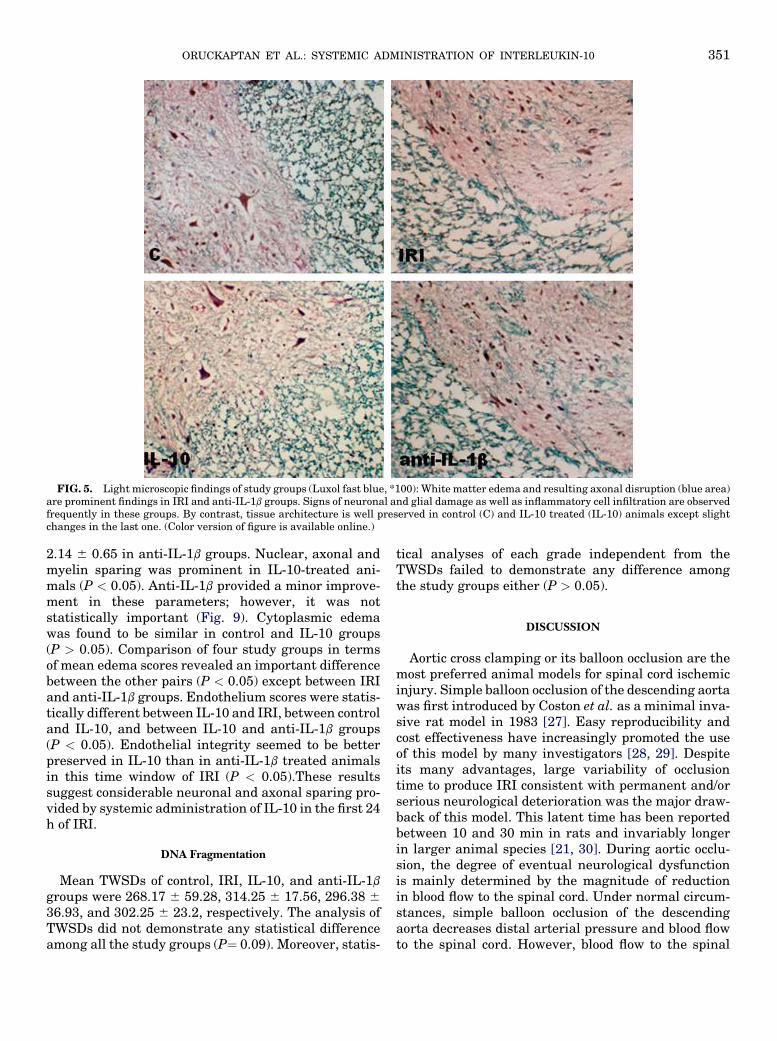
FIG. 5. Light microscopic findings of study groups (Luxol fast blue, *100): White matter edema and resulting axonal disruption (blue area)are prominent findings in IRI and anti-IL-1b groups. Signs of neuronal and glial damage as well as inflammatory cell infiltration are observedfrequently in these groups. By contrast, tissue architecture is well preserved in control (C) and IL-10 treated (IL-10) animals except slightchanges in the last one. (Color version of figure is available online.)
ORUCKAPTAN ET AL.: SYSTEMIC ADMINISTRATION OF INTERLEUKIN-10 351
2.14 6 0.65 in anti-IL-1b groups. Nuclear, axonal andmyelin sparing was prominent in IL-10-treated ani-mals (P < 0.05). Anti-IL-1b provided a minor improve-ment in these parameters; however, it was notstatistically important (Fig. 9). Cytoplasmic edemawas found to be similar in control and IL-10 groups(P > 0.05). Comparison of four study groups in termsof mean edema scores revealed an important differencebetween the other pairs (P < 0.05) except between IRIand anti-IL-1b groups. Endothelium scores were statis-tically different between IL-10 and IRI, between controland IL-10, and between IL-10 and anti-IL-1b groups(P < 0.05). Endothelial integrity seemed to be betterpreserved in IL-10 than in anti-IL-1b treated animalsin this time window of IRI (P < 0.05).These resultssuggest considerable neuronal and axonal sparing pro-vided by systemic administration of IL-10 in the first 24h of IRI.
DNA Fragmentation
Mean TWSDs of control, IRI, IL-10, and anti-IL-1b
groups were 268.17 6 59.28, 314.25 6 17.56, 296.38 6
36.93, and 302.25 6 23.2, respectively. The analysis ofTWSDs did not demonstrate any statistical differenceamong all the study groups (P¼ 0.09). Moreover, statis-
tical analyses of each grade independent from theTWSDs failed to demonstrate any difference amongthe study groups either (P > 0.05).
DISCUSSION
Aortic cross clamping or its balloon occlusion are themost preferred animal models for spinal cord ischemicinjury. Simple balloon occlusion of the descending aortawas first introduced by Coston et al. as a minimal inva-sive rat model in 1983 [27]. Easy reproducibility andcost effectiveness have increasingly promoted the useof this model by many investigators [28, 29]. Despiteits many advantages, large variability of occlusiontime to produce IRI consistent with permanent and/orserious neurological deterioration was the major draw-back of this model. This latent time has been reportedbetween 10 and 30 min in rats and invariably longerin larger animal species [21, 30]. During aortic occlu-sion, the degree of eventual neurological dysfunctionis mainly determined by the magnitude of reductionin blood flow to the spinal cord. Under normal circum-stances, simple balloon occlusion of the descendingaorta decreases distal arterial pressure and blood flowto the spinal cord. However, blood flow to the spinal
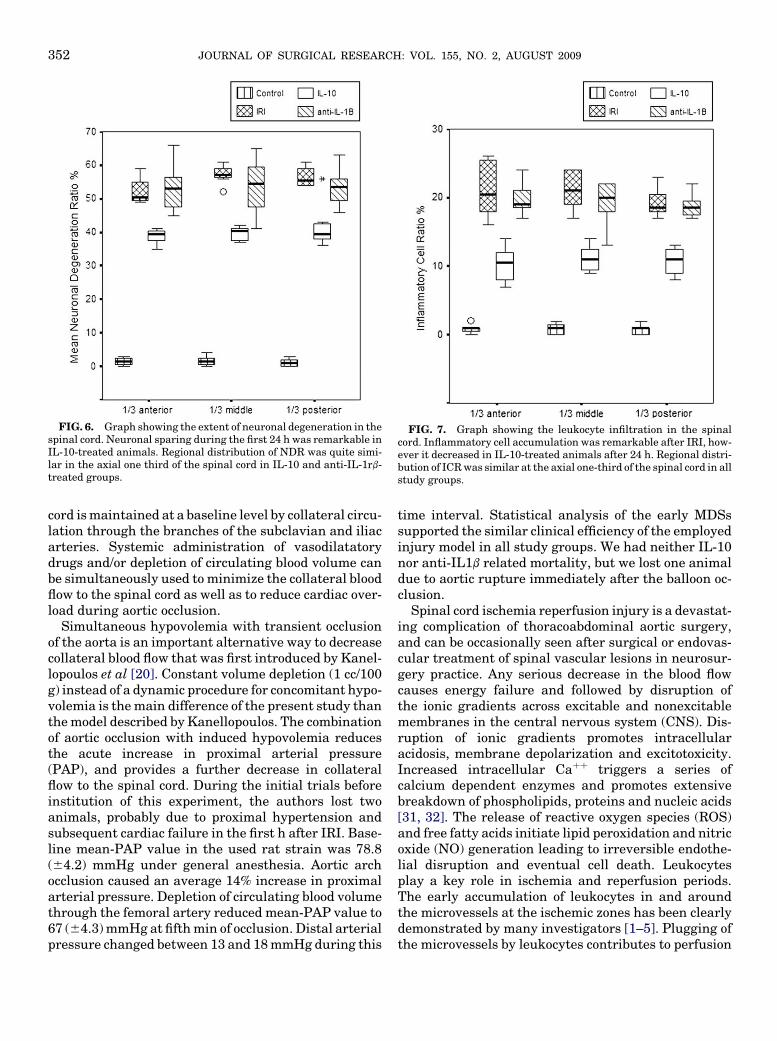
FIG. 6. Graph showing the extent of neuronal degeneration in thespinal cord. Neuronal sparing during the first 24 h was remarkable inIL-10-treated animals. Regional distribution of NDR was quite simi-lar in the axial one third of the spinal cord in IL-10 and anti-IL-1rb-treated groups.
FIG. 7. Graph showing the leukocyte infiltration in the spinalcord. Inflammatory cell accumulation was remarkable after IRI, how-ever it decreased in IL-10-treated animals after 24 h. Regional distri-bution of ICR was similar at the axial one-third of the spinal cord in allstudy groups.
JOURNAL OF SURGICAL RESEARCH: VOL. 155, NO. 2, AUGUST 2009352
cord is maintained at a baseline level by collateral circu-lation through the branches of the subclavian and iliacarteries. Systemic administration of vasodilatatorydrugs and/or depletion of circulating blood volume canbe simultaneously used to minimize the collateral bloodflow to the spinal cord as well as to reduce cardiac over-load during aortic occlusion.
Simultaneous hypovolemia with transient occlusionof the aorta is an important alternative way to decreasecollateral blood flow that was first introduced by Kanel-lopoulos et al [20]. Constant volume depletion (1 cc/100g) instead of a dynamic procedure for concomitant hypo-volemia is the main difference of the present study thanthe model described by Kanellopoulos. The combinationof aortic occlusion with induced hypovolemia reducesthe acute increase in proximal arterial pressure(PAP), and provides a further decrease in collateralflow to the spinal cord. During the initial trials beforeinstitution of this experiment, the authors lost twoanimals, probably due to proximal hypertension andsubsequent cardiac failure in the first h after IRI. Base-line mean-PAP value in the used rat strain was 78.8(64.2) mmHg under general anesthesia. Aortic archocclusion caused an average 14% increase in proximalarterial pressure. Depletion of circulating blood volumethrough the femoral artery reduced mean-PAP value to67 (64.3) mmHg at fifth min of occlusion. Distal arterialpressure changed between 13 and 18 mmHg during this
time interval. Statistical analysis of the early MDSssupported the similar clinical efficiency of the employedinjury model in all study groups. We had neither IL-10nor anti-IL1b related mortality, but we lost one animaldue to aortic rupture immediately after the balloon oc-clusion.
Spinal cord ischemia reperfusion injury is a devastat-ing complication of thoracoabdominal aortic surgery,and can be occasionally seen after surgical or endovas-cular treatment of spinal vascular lesions in neurosur-gery practice. Any serious decrease in the blood flowcauses energy failure and followed by disruption ofthe ionic gradients across excitable and nonexcitablemembranes in the central nervous system (CNS). Dis-ruption of ionic gradients promotes intracellularacidosis, membrane depolarization and excitotoxicity.Increased intracellular Caþþ triggers a series ofcalcium dependent enzymes and promotes extensivebreakdown of phospholipids, proteins and nucleic acids[31, 32]. The release of reactive oxygen species (ROS)and free fatty acids initiate lipid peroxidation and nitricoxide (NO) generation leading to irreversible endothe-lial disruption and eventual cell death. Leukocytesplay a key role in ischemia and reperfusion periods.The early accumulation of leukocytes in and aroundthe microvessels at the ischemic zones has been clearlydemonstrated by many investigators [1–5]. Plugging ofthe microvessels by leukocytes contributes to perfusion
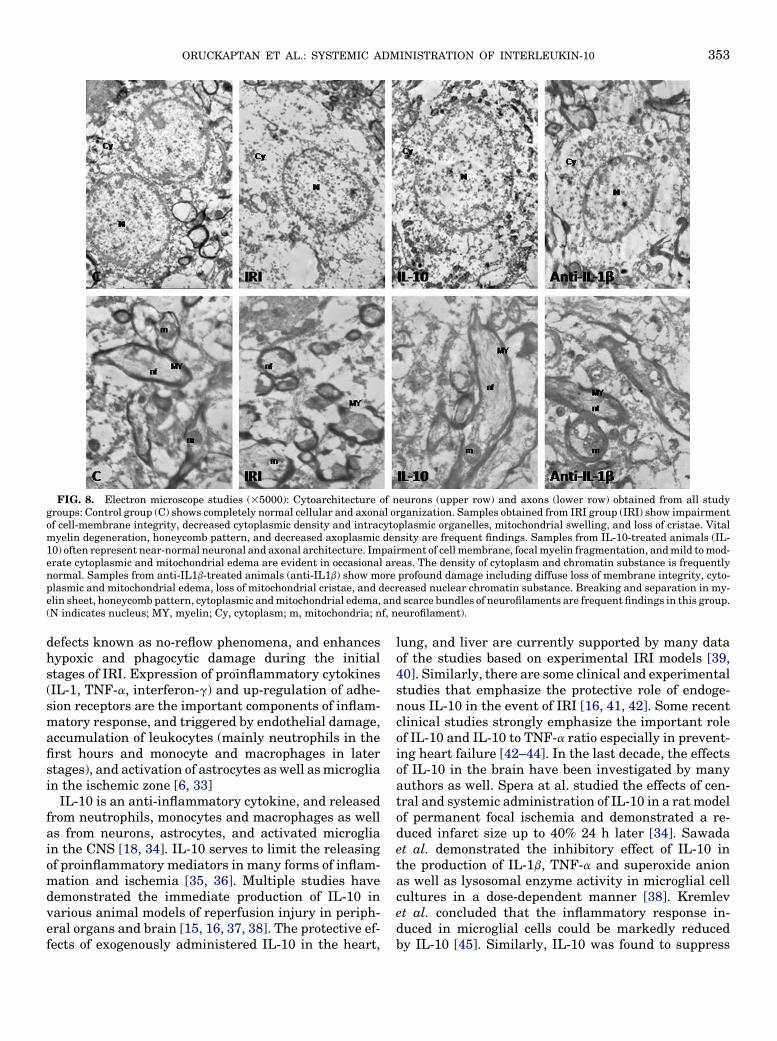
FIG. 8. Electron microscope studies (35000): Cytoarchitecture of neurons (upper row) and axons (lower row) obtained from all studygroups: Control group (C) shows completely normal cellular and axonal organization. Samples obtained from IRI group (IRI) show impairmentof cell-membrane integrity, decreased cytoplasmic density and intracytoplasmic organelles, mitochondrial swelling, and loss of cristae. Vitalmyelin degeneration, honeycomb pattern, and decreased axoplasmic density are frequent findings. Samples from IL-10-treated animals (IL-10) often represent near-normal neuronal and axonal architecture. Impairment of cell membrane, focal myelin fragmentation, and mild to mod-erate cytoplasmic and mitochondrial edema are evident in occasional areas. The density of cytoplasm and chromatin substance is frequentlynormal. Samples from anti-IL1b-treated animals (anti-IL1b) show more profound damage including diffuse loss of membrane integrity, cyto-plasmic and mitochondrial edema, loss of mitochondrial cristae, and decreased nuclear chromatin substance. Breaking and separation in my-elin sheet, honeycomb pattern, cytoplasmic and mitochondrial edema, and scarce bundles of neurofilaments are frequent findings in this group.(N indicates nucleus; MY, myelin; Cy, cytoplasm; m, mitochondria; nf, neurofilament).
ORUCKAPTAN ET AL.: SYSTEMIC ADMINISTRATION OF INTERLEUKIN-10 353
defects known as no-reflow phenomena, and enhanceshypoxic and phagocytic damage during the initialstages of IRI. Expression of proinflammatory cytokines(IL-1, TNF-a, interferon-g) and up-regulation of adhe-sion receptors are the important components of inflam-matory response, and triggered by endothelial damage,accumulation of leukocytes (mainly neutrophils in thefirst hours and monocyte and macrophages in laterstages), and activation of astrocytes as well as microgliain the ischemic zone [6, 33]
IL-10 is an anti-inflammatory cytokine, and releasedfrom neutrophils, monocytes and macrophages as wellas from neurons, astrocytes, and activated microgliain the CNS [18, 34]. IL-10 serves to limit the releasingof proinflammatory mediators in many forms of inflam-mation and ischemia [35, 36]. Multiple studies havedemonstrated the immediate production of IL-10 invarious animal models of reperfusion injury in periph-eral organs and brain [15, 16, 37, 38]. The protective ef-fects of exogenously administered IL-10 in the heart,
lung, and liver are currently supported by many dataof the studies based on experimental IRI models [39,40]. Similarly, there are some clinical and experimentalstudies that emphasize the protective role of endoge-nous IL-10 in the event of IRI [16, 41, 42]. Some recentclinical studies strongly emphasize the important roleof IL-10 and IL-10 to TNF-a ratio especially in prevent-ing heart failure [42–44]. In the last decade, the effectsof IL-10 in the brain have been investigated by manyauthors as well. Spera at al. studied the effects of cen-tral and systemic administration of IL-10 in a rat modelof permanent focal ischemia and demonstrated a re-duced infarct size up to 40% 24 h later [34]. Sawadaet al. demonstrated the inhibitory effect of IL-10 inthe production of IL-1b, TNF-a and superoxide anionas well as lysosomal enzyme activity in microglial cellcultures in a dose-dependent manner [38]. Kremlevet al. concluded that the inflammatory response in-duced in microglial cells could be markedly reducedby IL-10 [45]. Similarly, IL-10 was found to suppress
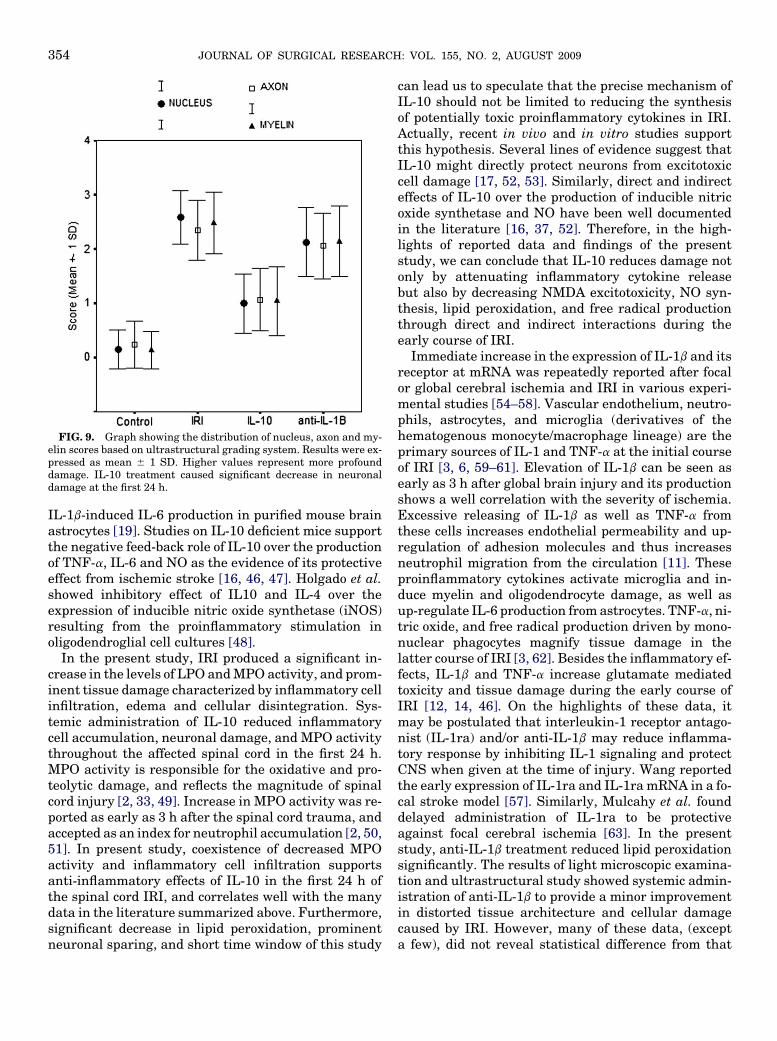
FIG. 9. Graph showing the distribution of nucleus, axon and my-elin scores based on ultrastructural grading system. Results were ex-pressed as mean 6 1 SD. Higher values represent more profounddamage. IL-10 treatment caused significant decrease in neuronaldamage at the first 24 h.
JOURNAL OF SURGICAL RESEARCH: VOL. 155, NO. 2, AUGUST 2009354
IL-1b-induced IL-6 production in purified mouse brainastrocytes [19]. Studies on IL-10 deficient mice supportthe negative feed-back role of IL-10 over the productionof TNF-a, IL-6 and NO as the evidence of its protectiveeffect from ischemic stroke [16, 46, 47]. Holgado et al.showed inhibitory effect of IL10 and IL-4 over theexpression of inducible nitric oxide synthetase (iNOS)resulting from the proinflammatory stimulation inoligodendroglial cell cultures [48].
In the present study, IRI produced a significant in-crease in the levels of LPO and MPO activity, and prom-inent tissue damage characterized by inflammatory cellinfiltration, edema and cellular disintegration. Sys-temic administration of IL-10 reduced inflammatorycell accumulation, neuronal damage, and MPO activitythroughout the affected spinal cord in the first 24 h.MPO activity is responsible for the oxidative and pro-teolytic damage, and reflects the magnitude of spinalcord injury [2, 33, 49]. Increase in MPO activity was re-ported as early as 3 h after the spinal cord trauma, andaccepted as an index for neutrophil accumulation [2, 50,51]. In present study, coexistence of decreased MPOactivity and inflammatory cell infiltration supportsanti-inflammatory effects of IL-10 in the first 24 h ofthe spinal cord IRI, and correlates well with the manydata in the literature summarized above. Furthermore,significant decrease in lipid peroxidation, prominentneuronal sparing, and short time window of this study
can lead us to speculate that the precise mechanism ofIL-10 should not be limited to reducing the synthesisof potentially toxic proinflammatory cytokines in IRI.Actually, recent in vivo and in vitro studies supportthis hypothesis. Several lines of evidence suggest thatIL-10 might directly protect neurons from excitotoxiccell damage [17, 52, 53]. Similarly, direct and indirecteffects of IL-10 over the production of inducible nitricoxide synthetase and NO have been well documentedin the literature [16, 37, 52]. Therefore, in the high-lights of reported data and findings of the presentstudy, we can conclude that IL-10 reduces damage notonly by attenuating inflammatory cytokine releasebut also by decreasing NMDA excitotoxicity, NO syn-thesis, lipid peroxidation, and free radical productionthrough direct and indirect interactions during theearly course of IRI.
Immediate increase in the expression of IL-1b and itsreceptor at mRNA was repeatedly reported after focalor global cerebral ischemia and IRI in various experi-mental studies [54–58]. Vascular endothelium, neutro-phils, astrocytes, and microglia (derivatives of thehematogenous monocyte/macrophage lineage) are theprimary sources of IL-1 and TNF-a at the initial courseof IRI [3, 6, 59–61]. Elevation of IL-1b can be seen asearly as 3 h after global brain injury and its productionshows a well correlation with the severity of ischemia.Excessive releasing of IL-1b as well as TNF-a fromthese cells increases endothelial permeability and up-regulation of adhesion molecules and thus increasesneutrophil migration from the circulation [11]. Theseproinflammatory cytokines activate microglia and in-duce myelin and oligodendrocyte damage, as well asup-regulate IL-6 production from astrocytes. TNF-a, ni-tric oxide, and free radical production driven by mono-nuclear phagocytes magnify tissue damage in thelatter course of IRI [3, 62]. Besides the inflammatory ef-fects, IL-1b and TNF-a increase glutamate mediatedtoxicity and tissue damage during the early course ofIRI [12, 14, 46]. On the highlights of these data, itmay be postulated that interleukin-1 receptor antago-nist (IL-1ra) and/or anti-IL-1b may reduce inflamma-tory response by inhibiting IL-1 signaling and protectCNS when given at the time of injury. Wang reportedthe early expression of IL-1ra and IL-1ra mRNA in a fo-cal stroke model [57]. Similarly, Mulcahy et al. founddelayed administration of IL-1ra to be protectiveagainst focal cerebral ischemia [63]. In the presentstudy, anti-IL-1b treatment reduced lipid peroxidationsignificantly. The results of light microscopic examina-tion and ultrastructural study showed systemic admin-istration of anti-IL-1b to provide a minor improvementin distorted tissue architecture and cellular damagecaused by IRI. However, many of these data, (excepta few), did not reveal statistical difference from that

ORUCKAPTAN ET AL.: SYSTEMIC ADMINISTRATION OF INTERLEUKIN-10 355
of the IRI group. In our opinion, these unfavorable re-sults might be due to time window of this study. Despitesome evidence about the effects of IL-1b on glutamate-mediated neurotoxicity, data in the literature supportsits major role on inducing the inflammatory cascadethat is amplified with mononuclear phagocytes and ac-tivated microglia in the latter course of IRI. Our studyfocused on the extent of injury in the first 24 h anddid not reflect delayed inflammatory cascade. Inade-quate therapeutic concentration of drug in the givendose and/or its insufficient penetration into the CNSmay be other, but less likely, explanations of the failureof anti-IL-1b treatment in the present study. The role ofthese factors on the failure of anti-IL-1b treatmentneeds to be clarified with further experimental studiesdesigned in extended time windows.
Another finding of the present study is the similarityof DNA damage among the study groups. This may bedue to the time window of the presented study or attrib-utable to technical errors during harvesting or trans-porting the samples for analysis. Comet assays weredone in the uppermost segments of spinal cord samplesnear to mid-thoracic level in all animals. This may be animportant factor because of the distinct rostrocaudalgradient of increasing tissue damage in the aortic occlu-sion model. Small increase in mean MRS of IL-10-treated group may be important; however, the shorttime window in this preliminary study strongly re-stricts its reliability for a scientific conclusion.
CONCLUSION
The role of cytokines in IRI has been increasingly in-vestigated in the heart, liver, and kidney as well as thebrain; however, studies in the spinal cord were quitelimited despite its high vulnerability. Data from thepresent study suggest that IL-10 may play an impor-tant role in spinal cord at the early stages of IRI, andits systemic administration may attenuate tissue dam-age through direct and indirect interactions. Althoughthis is only a preliminary study and reflects the damageat the first 24 h, the results are promising for further ex-perimental studies on the effects of anti-inflammatorycytokines especially focused on the later stages of spinalcord ischemia reperfusion injury.
REFERENCES
1. Clark RK, Lee EV, White RF, et al. Reperfusion following focalstroke hastens inflammation and resolution of ischemic injuredtissue. Brain Res Bull 1994;35:387.
2. Fleming JC, Norenberg MD, Ramsay DA, et al. The cellular in-flammatory response in human spinal cords after injury. Brain2006;129:3249.
3. Jean WC, Spellman SR, Nussbaum ES, et al. Reperfusion injuryafter focal cerebral ischemia: the role of inflammation and thetherapeutic horizon. Neurosurgery 1998;43:138225.
4. Kochanek PM, Hallenbeck JM. Polymorphonuclear leukocytesand monocytes/macrophages in the pathogenesis of cerebral is-chemia and stroke. Stroke 1992;23:1367.
5. Matsuo Y, Onodera H, Shiga Y, et al. Correlation between mye-loperoxidase-quantified neutrophil accumulation and ischemicbrain injury in rat: Effects of neutrophil depletion. Stroke1994;25:1469.
6. Barone FC, Feuerstein GZ. Inflammatory mediators and stroke:New opportunities for novel therapeutics. J Cereb Blood FlowMetab 1999;19:819.
7. Schoning B, Elepfandt P, Lanksch WR, et al. Continuous infu-sion of proinflammatory cytokines into the brain to study braincytokine induced local and systemic immune effects. Brain ResProt 1999;4:217.
8. Seekamp A, Jochum M, Ziegler M, et al. Cytokines and adhesionmolecules in elective and accidental trauma-related ischemia/re-perfusion. J Trauma 1998;44:874.
9. Schielke GP, Yang GY, Shivers BD, et al. Reduced ischemicbrain injury in interleukin-1b converting enzyme-deficientmice. J Cereb Blood Flow Metab 1998;18:180.
10. Yang G-Y, Liu X-H, Kadoya C, et al. Attenuation of ischemic in-flammatory response in mouse brain using an adenoviral vectoro induce overexpression of interleukin-1 receptor antagonist.J Cereb Blood Flow Metab 1998;18:840.
11. Wang CX, Shuaib A. Involvement of inflammatory cytokines inthe central nervous system injury. Progress Neurobiol 2002;67:161.
12. Griffin WST, Sheng JG, Gentleman SM, et al. Microglial inter-leukin -1a expression in human head injury: Correlation withneuronal and neuritic b-amyloid precursor protein expression.Neurosci Lett 1994;176:133.
13. Hu ZY, Wong SC, He YY, et al. Expression of IL-1b, TNF, and IL-6 mRNAs in the ischemic cerebral cortex. Soc Neurosci Abstr1994;20:615.
14. Liao S-L, Chen C-J. Differential effects of cytokines and redoxpotential on glutamate uptake in rat cortical glial cultures. Neu-rosci Lett 2001;299:113.
15. Lopez-Neblina Toledo-Pereyra LH, Toledo AH, et al. Ryanodinereceptor antagonism protects the ischemic liver and modulatesTNF-alpha and IL-10. J Surg Res 2007;140:121.
16. Yang Z, Zingarelli B, Szabo C. Crucial role of endogenous inter-leukin-10 production in myocardial ischemia/reperfusion injury.Circulation 2000;101:1019.
17. Barbieri I, Brusa R, Basudev H, et al. Neuroprotective action ofinterleukin-10 in mouse primary neuronal cultures. J NeurosciAbstr 1998;568:13.
18. Moore KW, O’Garra A, de Wall Malefyt R, et al. Interleukin-10.Annu Rev Immunol 1993;11:165.
19. Pousset F, Cremona S, Dantzer R, et al. Interleukin-4 and inter-leukin-10 regulate IL-1b induced mouse primary astrocyte acti-vation. Glia 1999;26:12.
20. Kanellopoulos GK, Kato H, Hsu CY, et al. Spinal cord ischemicinjury: Development of a new model in the rat. Stroke 1997;28:2532.
21. Marsala M, Yaksh TY. Transient spinal ischemia in the rat:characterization of behavioral and histopathological conse-quences as a function of the duration of aortic occlusion. J CerebBlood Flow Metab 1994;14:526.
22. Uchiyama M, Mihara M. Determination of malonaldehyde pre-cursor in tissues by thiobarbituric acid test. Annal Biochem1978;86:271.
23. Glowick SP, Kaplan SD. Method in Enzymology. New York:Academic Press, 1995. 769-782.

JOURNAL OF SURGICAL RESEARCH: VOL. 155, NO. 2, AUGUST 2009356
24. Kırkalı Z, Esen AA, Hayran M, et al. The effect of extracorporealelectromagnetic shock waves on the morphology and contractil-ity of rabbit ureter. J Urol 1995;154:1939.
25. Anderson D, Yu TW, Phillips BJ, et al. The effects of various an-tioxidants and other modifying agents on oxygen-radical gener-ated DNA damage in human lymphocytes in the COMET assay.Mutation Res 1994;307:261.
26. Singh NP, McCoy MT, Tice RR, et al. Simple technique for quan-titation of low levels of DNA damage in individual cells. Exp CellRes 1988;175:184.
27. Coston A, Laville M, Baud P, et al. Aortic occlusion by a ballooncatheter: A method to induce hind limb rigidity in rats. PhysiolBehav 1983;30:967.
28. Abraham VS, Swain JA, Forgash AJ, et al. Ischemic precondi-tioning protects against paraplegia after transient occlusion inthe rat. Ann Thorac Surg 2000;69:475.
29. Zivin JA, DeGirolami U. Spinal cord infarction: A highly repro-ducible stroke model. Stroke 1980;11:200.
30. Taira M, Marsala M. Effect of proximal arterial perfusion pres-sure on function, spinal cord blood flow, and histopathologicalchanges after increasing intervals of aortic occlusion in the rat.Stroke 1996;27:1850.
31. Siesjo BK. Calcium-mediated processes in neuronal degenera-tion. Ann NY Acad Sci 1994;747:140.
32. Simpson RK, Robertson CS, Goodman JC. Spinal cord ischemiainduced elevation of amino acids: Extracellular measurementwith microdialysis. Neurochem Res 1990;15:635.
33. Barone FC, Hillegass LM, Price WJ, et al. Polymorphonuclearleukocyte infiltration into cerebral focal ischemic tissue: Myelo-peroxidase activity assay and histologic verification. J NeurosciRes 1991;29:336.
34. Spera PA, Ellison JA, Feuerstein GZ, et al. IL-10 reduces ratbrain injury following focal stroke. Neurosci Lett 1998;251:189.
35. Feuerstein GZ, Wang X, Barone FC. Inflammatory mediatorsand brain injury: The role of cytokines and chemokines in strokeand CNS disease. In: Ginsberg MD, Bogousslavsky J, Eds. Cere-brovascular disease: Pathophysiology, diagnosis and manage-ment. Massachusetts: Blackwell Science, 1998:507–31.
36. Kelly A, Lynch A, Vereker E, et al. The anti-inflammatory cyto-kine, interleukin (IL)-10 blocks the inhibitory effect of IL-1b onlong term potentiation. J Biol Chem 2001;276:45564.
37. Molina-Holgado E, Vela JM, Aravelo-Martin A, et al. LPS/INFg
cytotoxicity in oligodendroglial cells: role of nitric oxide and pro-tection by the anti-inflammatory cytokine IL-10. Eur L Neurosci2001;13:493.
38. Sawada M, Suzumura A, Hosoya H, et al. Interleukin-10 inhibitsboth production of cytokines and expression of cytokine recep-tors in microglia. J Neurochem 1999;72:1466.
39. Dinant S, VetelainenRL Florquin S, et al. IL-10 attenuates he-patic I/R injury and promotes hepatocyte proliferation. J SurgRes 2007;141:176.
40. Hayward R, Nossuli TO, Scalia R, et al. Cardioprotective effect ofinterleukin-10 in murine myocardial ischemia-reperfusion. EurJ Pharmacol 1997;334:157.
41. Farivar AS, Krishnadasan B, Naidu BV, et al. Endogenous inter-leukin-4 and interleukin-10 regulate experimental lung ische-mia reperfusion injury. Ann Thorac Surg 2003;76:253.
42. Kaur K, Sharma AK, Signal PK. Significance of changes in TNF-a and IL-10 levels in the progression of heart failure subsequentto myocardial infarction. Am J Physiol Heart Circ Physiol 2006;291:106.
43. Heeschen C, Dimmeler S, Hamm CW, et al. Serum level of theanti-inflammatory cytokine interleukin-10 is an important prog-nostic determinant in patients with acute coronary syndromes.Circulation 2003;107:2109.
44. Hong M, Wei W, Hu Y, et al. Plasma levels of the anti-inflamma-tory cytokine IL-10 and inflammatory cytokine IL-6 in patientswith unstable angina. J Huazhong Univ Sci Technolog Med Sci2005;25:639.
45. Kremlev SG, Palmer C. Interleukin-10 inhibits endotoxin-induced proinflammatory cytokines in microglial cell cultures.J Neuroimmunol 2005;162:71.
46. Agnello D, Villa P, Ghezzi P. Increased tumor necrosis factor andinterleukin-6 production in the central nervous system of inter-leukin-10-deficient mouse. Brain Res 2000;869:241.
47. Frenkel D, Huang Z, Maron R, et al. Neuroprotection by IL-10-producing CD4þ T cells following ischemic stroke. J Neurol Sci2005;233:125.
48. Holgado EM, Vela JM, Aravelo-Martin A, et al. LPS/INFg cyto-toxicity in oligodendroglial cells: Role of nitric oxide and protec-tion by the anti-inflammatory cytokine IL-10. Eur L Neurosci2001;13:493.
49. Xu J, Hsu CY, Liu TH, et al. Leukotriene B4 release and poly-morphonuclear cell infiltration in spinal cord injury. J Neuro-chem 1990;55:907.
50. Allan G, Bhattachherjee P, Brook CD, et al. Myeloperoxidase ac-tivity as a quantitative marker of polymorphonuclear leukocyteaccumulation into an experimental myocardial infarct: the effectof ibufen on infarct size and polymorphonuclear leukocyte accu-mulation. J Cardiovasc Pharmacol 1985;7:1154.
51. Taoka Y, Okajima K. Role of leukocytes in spinal cord injury inthe rat. J Neurotrauma 2000;17:219.
52. Bachis A, Colangelo AM, Vicini S, et al. Interleukin-10 preventsglutamate-mediated cerebellar granule cell death by blockingcaspase-3-like activity. J Neurosci 2001;21:3104.
53. Balasingham V, Yong VW. Attenuation of astroglial reactivityby interleukin-10. J Neurosci 1996;16:2945.
54. Bonmann E, Suschek C, Spranger M, et al. The dominant role ofexogenous or endogenous interleukin-1b on expression and ac-tivity of inducible nitric oxide synthase in rat microvascularbrain endothelial cells. Neurosci Lett 1997;230:109.
55. Liu T, McDonnell PC, Young PR, et al. Interleukin-1b mRNA ex-pression in ischemic rat cortex. Stroke 1993;24:1746.
56. Sairanen TR, Linsberg TJ, Brenner M, et al. Global forebrain is-chemia results in differential cellular expression of interleukin-1b (IL-1b) and its receptor at mRNA and protein level. J CerebBlood Flow Metab 1997;17:1107.
57. Wang X-K, Barone FC, Aiyar NV, et al. Increased interleukin-1receptor and interleukin 1 receptor antagonist gene expressionafter focal stroke. Stroke 1997;28:155.
58. Zhang Z, Choop M, Goussev A, et al. Cerebral vessels express in-terleukin-1b after focal cerebral ischemia. Brain Research 1998;784:210.
59. Berthea JR, Nagashima HN, Acosta MC, et al. Systemically ad-ministered Interleukin-10 reduces tumor necrosis factor-alphaproduction and significantly improves functional recovery fol-lowing traumatic spinal cord injury in rats. J Neurotrauma1999;16:851.
60. Dinarello CA. Biologic basis for interleukine-1 in disease. Blood1996;87:2095.
61. Lafortune L, Nalbantoglu J, Antle JP. Expression of tumor ne-crosis factor-alpha (TNF-a) and interleukin 6 (IL-6) mRNA inadult human astrocytes: Comparison with adult microglia andfoetal astrocytes. J Neuropathol Exp Neurol 1996;55:515.
62. Tracey KJ, Cerami A. Tumor necrosis factor, other cytokines anddisease. Ann Rev Cell Biol 1993;9:317.
63. Mulcahy NJ, Ross J, Rothwell NJ, et al. Delayed administrationof interleukin-1 receptor antagonist protects against transientcerebral ischemia in the rat. Br J Pharmacol 2003;140:471.
Copyright © 2022 FDOKUMEN




















