
Systematics of Eligmodontia 2007
14
www.elsevier.de/mambio Original investigation Integrative taxonomy, systematics and distribution of the genus Eligmodontia (Rodentia, Cricetidae, Sigmodontinae) in the temperate Monte Desert of Argentina By Cecilia Lanzone, R.A. Ojeda and M.H. Gallardo Grupo de Investigaciones de la Biodiversidad, Instituto Argentino de Investigaciones de las Zonas A ´ ridas, CONICET, Mendoza, Argentina and Instituto de Ecologı ´a y Evolucio ´n, Universidad Austral de Chile, Valdivia, Chile Receipt of Ms. 22.6.2006 Acceptance of Ms. 24.9.2006 Abstract The systematics and distribution of South American sigmodontine rodents a matter of continuous revision and debate. The silky mice, genus Eligmodontia Cuvier, 1837, are among the most specialized murid rodents endemic to South America and its diversification for desert existence is associated with the uplift of the Andes and the early development of arid landscapes. Aiming to clarify the systematics of the species of silky mice occurring in the driest portion of the temperate Monte Desert in Argentina, qualitative and quantitative external and cranial characters, cytogenetics and molecular relationships, were studied. We characterized three karyotypes of Eligmodontia; two of which are described for the first time, and allocated them to previously named species. E. moreni Thomas, 1896 (2n=52 and FN=50), E. typus Cuvier, 1837 (2n=44 and FN=44) and E. marica Thomas, 1918. The later shows the same diploid number of E. typus, but its X— chromosome is not METACENTRIC but ACROCENTRIC . A discriminant analysis of external and cranial data separates E. moreni from E. typus and E. marica. Whereas these last ones show some degree of overlap. The morphological and chromosomal differentiation of Eligmodontia is sustained by DNA distances. Phylogenetic analyses show two major clades. One formed by E. moreni, E. puerulus and E. hirtipes, sharing a high FN and a northern distribution, and THE other formed by E. typus, E. marica and E. morgani with low FN and a central-southern distribution. Two centers of diversification are proposed to explain the evolution of Eligmodontia. r 2006 Deutsche Gesellschaft fu ¨r Sa ¨ugetierkunde. Published by Elsevier GmbH. All rights reserved. Key words: Eligmodontia, sigmodontines, morphometry, karyotypes, phylogeny Introduction The evolutionary history of South American rodents is a topic of continuous revision and debate (Hershkovitz 1962; Steppan 1995, 1998; Smith and Patton 1999; D’ Elı´a 2003; Musser and Carleton 2005). The sigmodon- tines are the largest rodent fauna of South America, with about 377 species (Musser and Carleton 2005), differentiated as a distinct lineage during middle to late Miocene (Reig 1986; Salazar-Bravo et al. 2001; Steppan et al. 2004). Their explosive radiation con- fronts researchers in defining their species ARTICLE IN PRESS 1616-5047/$ - see front matter r 2006 Deutsche Gesellschaft fu ¨r Sa ¨ugetierkunde. Published by Elsevier GmbH. All rights reserved. doi:10.1016/j.mambio.2006.09.001 Mamm. biol. 72 (2007) 5 299–312
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Systematics of Eligmodontia 2007

ARTICLE IN PRESS
www.elsevier.de/mambio
1616-5047/$ - see
doi:1
Original investigation
Integrative taxonomy, systematics and distribution of thegenus Eligmodontia (Rodentia, Cricetidae, Sigmodontinae)in the temperate Monte Desert of Argentina
By Cecilia Lanzone, R.A. Ojeda and M.H. Gallardo
Grupo de Investigaciones de la Biodiversidad, Instituto Argentino de Investigaciones de las Zonas Aridas,CONICET, Mendoza, Argentina and Instituto de Ecologıa y Evolucion, Universidad Austral de Chile, Valdivia, Chile
Receipt of Ms. 22.6.2006Acceptance of Ms. 24.9.2006
Abstract
The systematics and distribution of South American sigmodontine rodents a matter of continuousrevision and debate. The silky mice, genus Eligmodontia Cuvier, 1837, are among the mostspecialized murid rodents endemic to South America and its diversification for desert existence isassociated with the uplift of the Andes and the early development of arid landscapes. Aiming toclarify the systematics of the species of silky mice occurring in the driest portion of the temperateMonte Desert in Argentina, qualitative and quantitative external and cranial characters,cytogenetics and molecular relationships, were studied. We characterized three karyotypes ofEligmodontia; two of which are described for the first time, and allocated them to previously namedspecies. E. moreni Thomas, 1896 (2n=52 and FN=50), E. typus Cuvier, 1837 (2n=44 and FN=44) andE. marica Thomas, 1918. The later shows the same diploid number of E. typus, but its X—chromosome is not METACENTRIC but ACROCENTRIC . A discriminant analysis of external and cranialdata separates E. moreni from E. typus and E. marica. Whereas these last ones show some degree ofoverlap. The morphological and chromosomal differentiation of Eligmodontia is sustained by DNAdistances. Phylogenetic analyses show two major clades. One formed by E. moreni, E. puerulus and E.hirtipes, sharing a high FN and a northern distribution, and THE other formed by E. typus, E. maricaand E. morgani with low FN and a central-southern distribution. Two centers of diversification areproposed to explain the evolution of Eligmodontia.r 2006 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
Key words: Eligmodontia, sigmodontines, morphometry, karyotypes, phylogeny
Introduction
The evolutionary history of South Americanrodents is a topic of continuous revision anddebate (Hershkovitz 1962; Steppan 1995,1998; Smith and Patton 1999; D’ Elıa 2003;Musser and Carleton 2005). The sigmodon-tines are the largest rodent fauna of South
front matter r 2006 Deutsche Gesellschaft fur S
0.1016/j.mambio.2006.09.001
America, with about 377 species (Musser andCarleton 2005), differentiated as a distinctlineage during middle to late Miocene (Reig1986; Salazar-Bravo et al. 2001; Steppanet al. 2004). Their explosive radiation con-fronts researchers in defining their species
augetierkunde. Published by Elsevier GmbH. All rights reserved.
Mamm. biol. 72 (2007) 5 � 299–312

ARTICLE IN PRESS
300 C. Lanzone et al.
limits, relationships and contents of themajor tribes (D’ Elıa 2000; Steppan et al.2004). Among sigmodontines, the represen-tatives of the Phyllotini tribe are small-sizedrodents, and primarily distributed along thearid and semiarid landscapes of the Neotro-pical southern cone. Among these, the genusEligmodontia F. Cuvier, 1837, has a wide-spread distribution, ranging from the highAndean plateau of southern Peru, westernBolivia and scrublands of Chile, to the Puna,Monte and Patagonian aridlands biomes ofArgentina. It possesses several anatomical,and physiological adaptations for xeric ex-istence (Mares 1977; Diaz and Ojeda 1999;Diaz 2001).The taxonomy, systematics and distributionof the genus Eligmodontia is confusing sincemost species characterizations are based onfew external and cranial features of fewspecimens. Cabrera (1961) recognizes twospecies, E. puerulus and E. typus. In the firstreview of the genus, Hershkovitz (1962)synonymized all forms within E. typus withtwo subspecies: E. t. typus and E. t. puerulus.More recently, in their account of Neotropi-cal mammals, Redford and Eisenberg (1992)recognized only one species: E. typus, butsuggest the existence of two others. Finally,Braun (1993) and Musser and Carleton(2005) recognize six species (E. hirtipes,E. marica, E. moreni, E. morgani, E. puerulusand E. typus), and four species (E. typus,E. moreni, E. morgani and E. puerulus),respectively, but no diagnostic features areprovided in either cases.The integration of cytogenetic, morphologi-cal and molecular studies has contributed toclarify the limits among Eligmodontia speciesin the southern part of their distributionalrange in Patagonia (Ortells et al. 1989; Keltet al. 1991; Zambelli et al. 1992; Hillyardet al. 1997; Sikes et al. 1997; Tiranti 1997).Their results have shown a high karyotypicand molecular divergence, but rather littlemorphological differentiation between:E. typus and E. morgani (Hillyard et al.1997; Sikes et al. 1997).Having in mind the confusing and contra-dictory taxonomy, systematic relationships,and delimitation of the geographic ranges ofEligmodontia in the lowland Monte Desert
here, we propose to clarify these issues usingcytogenetic, morphological and molecular-based phylogenetic approach.
Material and methods
Animals were collected at eight localities along thenorthern-central portion of the Monte desertbiome. Localities, collected individuals, coordi-nates and provinces are as follows: Cafayate(n=3) 261050/651580/1660m, Salta Province; Cam-po Arenal-Los Nacimientos (n=19), 27108050.3’’/66140042.7’’/ 2139m, Salar de Pipanaco (n=5),27149016.1’’/66114034.9’’/740m, Catamarca Pro-vince; Ischigualasto Reserve (n=3), 30105019’’/6715600’’/1252m, San Juan Province; Pampa delas Salinas (n=1) 321130/661360/420m, San LuisProvince; Telteca Reserve, (n=12), 32123035.5’’/68102046.8’’/520m; 33 km NE of Costa de Araujo(n=1) 321450/681250/520m; Reserve of Nacunan,(n=27), 34102041.1’’/97154033.9’’/565m, MendozaProvince (Fig. 1). Voucher specimens and tissuesamples are housed at the mammal collection of theInstituto Argentino de Zonas Aridas (IADIZA),CONICET, Mendoza.Chromosomal preparations of 71 specimens wereobtained using the standard hypotonic techniquefor bone-marrow (Ford and Hamerton 1956) withsmall modifications. Chromosomes were stainedwith Giemsa (pH=6.8). Ten metaphases spreadswere counted for each specimen. FundamentalNumber of chromosome arms (FN) followedPatton (1967).Five external (total length, tail length, hind footlength, ear length and weight) and 21 standardcranial features were measured using a caliperrounded to the nearest 0.1mm (Sikes et al. 1997;Martin et al. 2001). Only animals with completelyerupted dentition were used for the analyses. Thecranial features used were: condylobasal length,least interorbital breadth, zygomatic breadth,greatest length of skull, basal length, breadth ofbraincase, length of maxillary toothrow, palatallength, bullar length, bullar width, length ofmandible toothrow, greatest length of mandible,diastema length, palatal bridge, breadth of palate(in M1 and in M3), incisive breadth, incisiveforamina, nasal breadth, nasal length, and rostrumwidth. Univariate and multivariate discriminantanalysis (MDA) were implemented using InfoStatand Statistica programs.For the molecular analyses, total DNA from liverwas extracted by the standard Phenol–Chloroformmethod (Sambrook et al. 1989). PCR amplificationof a 687 pb of the cyt b gene was performed usingprimers MVZ 05 and MVZ 16 (Smith and Patton

ARTICLE IN PRESS
Fig. 1. Map of Central Argentina showing the populations of Eligmodontia analyzed. 1: Cafayate, 2: CampoArenal, 3: Pipanaco, 4: Ischigualasto, 5: Pampa de las Salinas, 6: Telteca, 7: Costa de Araujo, 8: Nacunan andtheir type localities, a: Chumbicha, b: Chilecito, c: Buenos Aires.
Systematics of Eligmodontia in the Monte Desert 301
1993) following thermal profiles of 951C denatura-tion (3min), 501C annealing (1min), and 721Cextension (1min) for 35 cycles. Negative controlswere included in all experiments. The PCRproducts were purified with Wizard SV Gel andPCR clean UpSystem (Promega) and sequenced inboth directions.Five individuals of different localities were sequenced(E. typus from Telteca; E. sp. with the karyotype 1from Campo Arenal; E. sp. with the karyotype 2from Campo Arenal and Telteca; E. puerulus
Philippi 1896, from Abra Pampa, Jujuy) and fourEligmodontia sequences from the GeneBank wereincluded to incorporate specimens of all recognizedEligmodontia species: E. typus (Accession Numbers:AF108692), E. morgani (NA: AF108691) and E.
hirtipes (NA: AY341054; AF159289). Sequences ofrelated phyllotine Graomys griseoflavus (AN:AY275117), G. domorum (AN: AF159291) andCalomys laucha (AN: AY033190) obtained fromthe GeneBank were used as outgroup species.Sequence alignment was carried out with Clustal X(Thompson et al. 1997) and corroborated by eye
inspection. Uncorrected and Kimura two parameter(K2p) distances were calculated. Phylogenetic rela-tionships among Eligmodontia species were analysedusing neighbour-joining (NJ), maximum parsimony(MP) and maximum-likelihood (ML) in PAUP4.0B10 (Swofford 2002). Variable nucleotide posi-tions were equally weighted and treated as unorderedcharacters. Data was subject to 1000 bootstrapreplications in the NJ and MP. MP analyses wereused with heuristic search with tree bisectionreconnection (TBR) branch swapping and randomaddition sequence. The distance used in NJ tree wasK2p. ML parameter values were estimated inModeltest 3.04 (Posada and Crandall 1998). Themodel selected by the AIC (minimum theoreticalinformation criterion) was GTR+I+G (�lnL=2718.5781) with the following parameters: per-centage of invariable sites =0.5369; gamma dis-tribution shape parameter =2.0428; assumed basefrequencies A=0.3055, C=0.2740, G=0.1264,T=0.2941; transition to transversion ratio=9.4182, and the support for the nodes wasevaluated for 100 replicates.

ARTICLE IN PRESS
302 C. Lanzone et al.
Results
Karyotypes
Two major karyotypes differing in numberand morphological features of chromosomes(2n=44 and 2n=52) were found. A thirdkaryotypic variant, coinciding with the2n=44-chromosome, but differing from itby the morphology of the X–chromosomewas detected too. The coinciding 2n=44,NF=44 karyotype with that of E. typus(Ortells et al. 1989; Tiranti 1997) was foundin Costa de Araujo, Pampa de las Salinas,Nacunan, and in one specimen from Telteca.This karyotype is characterized by a large-sized metacentric autosomal pair, whereasthe rest of the elements are acrocentrics ofdecreasing size. The X–chromosome is alarge metacentric, and Y–chromosome is asmall submetacentric (Fig. 2a).A karyotype with 2n=44, NF=44, verysimilar to the one described for E. typus,but with an acrocentric X–chromosome(karyotype 1) was found in all specimensfrom Cafayate and the Salar, and in somefrom Campo Arenal (n=5). Although theX–chromosome of this karyotype and that ofE. typus differ morphologically they have asimilar size (Fig. 2b).A chromosome complement with 2n=52,NF=50 (karyotype 2) was found in allindividuals from Ischigualasto and in somefrom Telteca (n=11), and Campo Arenal(n=14). In this karyotype all autosomes areacrocentric and can be arranged by decreas-ing size. The X–chromosome is a largeacrocentric, and Y a small submetacentric(Fig. 2c). Secondary constrictions were ob-served in pairs 3 and 7 in interstitial position(Fig. 2c).
External and craniometrical analysis
Comparative morphology: A similar skullmorphology was found in specimens ascribedto E. typus and to karyotype 1, while they aremarkedly different from specimens having thekaryotype 2. Diagnoses of major external andcranial characters of the three Eligmodontiakaryomorphs (E. typus, E. sp. with thekaryotype 1 and with the karyotype 2) are asfollows:
E. typus: Externally, this species is larger thanspecimens with karyotype 1 and smaller thanthose with karyotype 2 (Tab. 1). The tail islonger than head and body and slightlybicolored with a dark dorsal surface; notpencilled or with a tip of hairs less than 5mm.The dorsum is brownish and darker thanspecimens with the karyotype 1 and theventer has white or gray hairs at the base.The dorsum and the venter are delineated bya yellowish lateral line. Pelage around themouth is white; with a whitish patch behindeach eye. In the skull (Fig. 3a, b), the nasalsare straight and the zygomatic arcs areparallel and narrower than in those with thekaryotype 2. In the anterior border of maxillathe tubercule is absent or slightly developedand rounded. The posterior margin of palateis straight. The maxillary foramina arelocated in the anterior half of M1 and largerthan in specimens with the karyotype 2;foramen ovale present and rounded; para-pterygoid plate more expanded and flat thanin those with the karyotype 2. The petro-tympanic fossa is elongated along the ante-rior edge of tympanic bullae; bullaemoderately inflated with long bullar tubes.M1 with anteromedian flexus slightly markedor absent; anterolingual style absent orslightly developed and mesostyle absent; nocontact between labial (hypoflexus) andsecondary lingual fold (metaflexus); M3 with-out enamel island; anteriormedian flexidabsent; posterolophid present.Eligmodontia with karyotype 1: Size small(Tab. 1); tail unicolored and pale, equal orsligthly larger than head and body, with nopenciled tip; hind feet smaller with hairy sole;dorsum light brown and greyish with whitehairs at the base; a light lateral colorationwith a diffuse line; area around the mouthwhite; a whitish patch behind each eye. In theskull (Fig. 3c, d) the zygomatic arc is paralleland narrower than in specimens with thekaryotype 2; anterior border of maxilla witha developed and rounded tubercule; max-illary foramina located in the anterior half ofM1 larger than in specimens with thekaryotype 2; foramen ovale present androunded; parapterygoid plate more expandedand flat than in those with the karyotype 2;petrotympanic fossa elongated along the

ARTICLE IN PRESS
Fig. 2. Bone marrow standard Giemsa staining karyotype of (A) Eligmodontia typus from Costa de Araujo; (B)karyotype 1 from the salt basin of Pipanaco; (C) karyotype 2 from Telteca and Campo Arenal.
Systematics of Eligmodontia in the Monte Desert 303
anterior edge of tympanic bullae; bullaemoderately inflated with long bullar tubes.M1 with anteromedian flexus slightly markedor absent; anterolingual style absent orslightly developed and mesostyle absent; nocontact between labial (hypoflexus) andsecondary lingual fold (metaflexus); M3 with-
out enamel island; anteriormedian flexidabsent; posterolophid present.Eligmodontia with the karyotype 2: Sizelarger than the others. Tail longer than headand body, markedly bicolored and moder-ately haired, with dark dorsal surface and apencilled tip of hairs longer than 5mm. Large

ARTICLE IN PRESS
Table 1. Descriptive statistics (mean7SD and range) for external and cranial variables (in mm) and significantlevels for Kruskal–Wallis test.
E. typus Karyotype 1 Karyotype 2(N=26) (N=14) (N=28)
VARIABLE Mean7SD Mean7SD Mean7SD P
Total length 178.70724.80 170.0078.00 185.71710.34 0,0002(83.00–222.00) (159.00–183.00) (168.00–202.00)
Tail length 94.67713.68 90.3677.38 106.3978.14 o0,0001(72.00–110.00) (80.00–104.00) (93.00–120.00)
Hind-foot length 22.0171.78 21.8771.53 24.1870.90 o0,0001(16.00–25.00) (19.00–24.00) (22.10–26.00)
Ear length 17.8372.10 17.0771.07 19.4371.27 o0,0001(11.00–23.00) (15.00–19.00) (16.00–21.50)
Weight 17.6573.65 18.3673.87 17.6373.31 0,8646(12.50–26.86) (13.50–26.00) (12.50–24.50)
Condylobasal length 21.0970.68 20.9970.61 21.9570.86 0,0004(19.90–22.70) (20.00–22.40) (20.3–23.4)
Least interorbital breadth 3.7970.17 3.8970.16 4.2270.16 o0,0001(3.50–4.30) (3.60–4.10) (3.90–4.60)
Zygomatic breadth 12.1770.39 12.1870.32 12.7470.56 0,0003(11.20–12.90) (11.80–12.90) (11.60–13.90)
Greatest length of skull 23.8570.94 23.9070.77 24.9470.84 0,0001(21.00–25.70) (22.50–25.30) (23.30–26.40)
Basal length 19.9172.07 19.1770.60 19.9970.81 0,0067(17.30–29.50) (18–20.4) (18.20–21.30)
Breadth of braincase 11.2370.26 11.3370.27 11.7370.29 o0,0001(10.60–11.70) (10.9–11.9) (11.20–12.20)
Length maxillary tooth row 3.7870.14 3.6170.14 3.9770.16 o0,0001(3.50–4.00) (3.40–3.80) (3.70–4.30)
Palatal length 11.8170.50 11.8170.38 12.2570.51 0,0059(11.10–13.20) (11.00–12.40) (11.20–13.00)
Bullar width 4.7770.14 4.7370.15 5.4070.16 o0,0001(4.50–5.00) (4.50–5.00) (5.10–5.60)
Bullar length 6.2170.31 5.4370.21 6.2770.20 o0,0001(5.50–6.70) (5.10–5.80) (5.90–6.60)
Length mandible tooth row 3.7270.13 3.5670.13 3.8970.15 o0,0001(3.40–4.00) (3.20–3.80) (3.60–4.30)
Greatest length mandible 11.8470.46 11.7070.47 12.3370.65 0,0027(11.10–12.90) (11.00–12.60) (11.20–13.20)
Diastema length 5.5570.26 5.5870.29 5.5370.35 0,8387(5.10–6.30) (5.20–6.20) (4.90–6.10)
Palatal bridge 4.9870.28 4.6970.39 5.2770.28 o0,0001(4.40–5.80) (4.20–5.60) (4.60–6.00)
Length palate in M1 3.0370.15 2.9170.15 2.9470.17 0,0220(2.60–3.30) (2.70–3.20) (2.60–3.30)
Length palate in M3 2.7270.15 2.6070.15 2.6770.18 0,0620(2.40–3.00) (2.20–2.90) (2.40–3.10)
Incisive width 1.4170.12 1.5670.10 1.5870.10 o0,0001(1.20–1.60) (1.30–1.70) (1.40–1.80)
Incisive foramina 5.0070.25 4.8370.28 4.9470.39 0,0843(4.40–5.40) (4.40–5.30) (4.20–5.70)
Nasal width 2.1270.13 2.2570.18 2.2170.14 0,0148
(1.80–2.40) (1.90–2.60) (1.90–2.50)Nasal length 9.1070.65 9.1170.47 8.9970.45 0,8159
(7.70–10.30) (8.30–10.00) (8.00–9.60)Rostral width 3.7870.14 3.7670.18 3.9170.19 0,0121
(3.60–4.20) (3.40–4.00) (3.60–4.30)
304 C. Lanzone et al.

ARTICLE IN PRESS
Systematics of Eligmodontia in the Monte Desert 305
hind feet with hairs. The coloration isbrownish in the dorsum and the venter iswhite to the base of the hairs. Dorsum andventer are delineated by a yellowish lateralline. The area around the mouth is white andhas a whitish patch behind each eye. Inter-orbital region of the skull is slightly divergentposteriorly and with edges angled for ap-proximately half its length; nasals straight upto the middle of their length and thenexpanded to their anterior margin; zygomaticarcs parallel or slightly convergent anteriorly,narrow and slender; deep zygomatic notchwith ovate internal border; anterior border ofmaxilla with a developed and roundedtubercule; posterior margin of palaterounded; small maxillary foramina locatedin the anterior half of M1; a tear shapedforamen ovale present; pterygoid plate ex-panded; petrotympanic fossa narrow; bullaeinflated with bullar tubes short (Fig. 3e, f).M1 with anteromedian flexus marked toslightly marked; anterolingual and mesostylepresent; labial fold (hypoflexus) in contactwith secondary lingual fold (metaflexus); M3
with enamel island; with anteriormedianflexid variable; condyloid process narrowerthan the angular process. The presence of awide petrotympanic fissure (middle lacerateforamen) and a protuberant stapedial spineof auditory bulla in specimens with thekaryotype 1 and E. typus contrast with thereduction of both structures in specimenswith the karyotype 2 (Fig. 3b, d, f).Kruskal–Wallis test was not significant fordifferences between males and females (uni-variate analyses) in E. typus and in specimenswith the karyotype 1. Males and females withthe karyotype 2 are differentiated in theirtotal length and tail length; no significantdifferences were detected in other external orcranial variables. The three karyotypic formsshowed significant differences in external andcranial variables (Tab. 1), however, there is agreat degree of overlap in measurementsamong the three groups. MDA of externaland cranial measurements, showed a clearseparation of specimens with karyotype 2 andsome degree of overlap in E. typus andspecimens with the karyotype 1 (Fig. 4).Because of differences in sample size, priorprobabilities were set equal to sizes of
samples. This analysis accurately assigned98.6% of individuals to the corrected speciesin a post hoc comparison. The remainingerror (1.4%) is explained by a specimenmisclassified as E. typus. In a step-wisediscriminant function analysis, the variableswhich most contributed to the discriminationwere: least interorbital breadth, greatestlength of skull, length of maxillary toothrow, bullar length, bullar width, breadth ofpalate in M1, incisive foramina and nasallength.
Molecular data
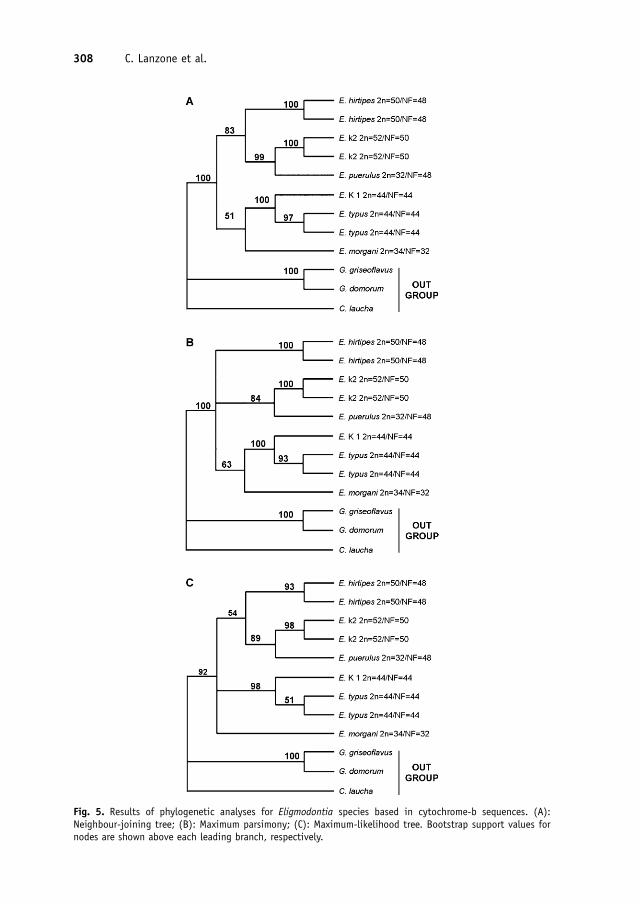
Low sequence divergence in individuals hav-ing the same karyotype, but large interkar-yotypic differences, was found. Distancebetween the 52 chromosome karyotype fromTelteca and Campo Arenal was 1.5% and thedistance between E. typus (2n=44) fromTelteca and Rio Negro was 1%. E. typusdiffered between 8.6% and 9.3% from thosewith 2n=52 (uncorrected), and 9.3% and10% (K2p), whereas specimens with thekaryotypes 1 and 2 were 9.6% and 11%(uncorrected and K2p, respectively). On thecontrary, the divergence between E. typus2n=44 and the specimen with the karyotype1 (2n=44) was 2.4% (uncorrected) and 2.5%(K2p).Analyses using NJ, MP and ML retrieved amonophyletic genus Eligmodontia. Specimenswith identical karyotype are consistentlygrouped together (Fig. 5). All phylogenetictrees resulted in a similar topology, althoughcomplementaries non-resolved polytomiesare recovered in the last two methods, whichinvolved E. hirtipes and E. morgani, respec-tively. Two major clades are found in the NJtree (Fig. 5a). One composed of speciesmostly distributed in the north, with the((E. puerulus+specimens with 2n=52)+E. hirtipes) relationship consistent with ahigh support. Another clade that includes thecentral-southern species ((E. typus+speci-mens with karyotype 1)+E. morgani) isrecovered with a low support (Fig. 5a).The data set of the MP analyses has 234variable characters of which 163 are infor-mative. Only one most parsimonious treeresulted from this analysis (length=405,

ARTICLE IN PRESS
Fig. 3. Ventral cranial views of species of Eligmodontia and detail of auditory bulla. (A, B) E. typus; (C, D)karyotype 1; (E, F) karyotype 2. ptf=petrotympanic fossa, wid=wide interbullar distance, nid=narrow interbullardistance, sts=stapedial spine, bt=bullar tube.
306 C. Lanzone et al.
CI=0.7, RI=0.7). The close relationshipbetween E. typus and that with the karyotype1 is supported by high bootstrap value, whileE. morgani is joined externally to this cladewith a lower support (63%). E. puerulus and
specimens with the karyotype 2 are againrecovered as sister taxa, but specimens ofE. hirtipes form a basal polytomy (Fig. 5b).The results of the ML analysis (Fig. 5c)corroborate the close relationship of

ARTICLE IN PRESS
Fig. 4. Plot of canonical discriminant analysis using all cranial and external characters in the three Eligmodontiakaryotypes.
Systematics of Eligmodontia in the Monte Desert 307
E. puerulus and specimens with karyotype 2;while E. hirtipes appears sister to this group,like in the NJ tree, but with a lower support.The clade formed by specimens with 2n=44are again recovered, but E. morgani is joinedat the base of the Eligmodontia tree as a non-resolved polytomy.
Discussion
Despite the rather little morphological differ-entiation between sigmodontines, cytogeneticstudies have been important in clarifying thespecies limits (Pearson and Patton 1976;Fagundes et al. 2000; Spotorno et al. 2001).In Eligmodontia, four basic karyotypes havebeen described and can be assigned to thefollowing species: E. typus, 2n=43–44,NF=44 (Ortells et al. 1989; Zambelli et al.1992; Tiranti 1997); E. morgani,2n=32–33–34, NF=32 (Ortells et al. 1989;Kelt et al. 1991; Zambelli et al. 1992; Tiranti1997); E. hirtipes, 2n=50, NF=48 (Pearsonand Patton 1976; Ortells et al. 1989; Kelt etal. 1991; Spotorno et al. 1994) andE. puerulus with 2n=32–34, NF=48 (Spot-
orno et al. 2001; Lanzone et al. unpubl. data).The intraspecific variation in these cytotypesis due to Robertsonian translocations (Ortellset al. 1989; Zambelli et al. 1992; Lanzoneet al. unpubl. data) which is the mostfrequent chromosomal rearrangement in ro-dents (Patton and Sherwood 1983).We suggest that the karyotype with 2n=52(NF=50), from Mendoza, San Juan andCatamarca, should correspond to E. moreniThomas, 1896 since its type locality isChilecito, La Rioja, Argentina (Thomas1896), approximately 100 km from our sam-pling site in Ischigualasto. The telocentrickaryotype of E. moreni is reported here forthe first time and corresponds to the highestdiploid and fundamental numbers recordedin Eligmodontia. The high molecular andmorphological distinctiveness of these speci-mens as reported for other well differentiatedsigmodontines (Hillyard et al. 1997; Steppan1995, 1998; Smith and Patton 1993, 1999;D’ Elıa 2003), confirms its full status as aspecies.The karyotypes with 2n=44, NF=44 fromTelteca, Costa de Araujo and Nacunan
ARTICLE IN PRESS
Fig. 5. Results of phylogenetic analyses for Eligmodontia species based in cytochrome-b sequences. (A):Neighbour-joining tree; (B): Maximum parsimony; (C): Maximum-likelihood tree. Bootstrap support values fornodes are shown above each leading branch, respectively.
308 C. Lanzone et al.

ARTICLE IN PRESS
Systematics of Eligmodontia in the Monte Desert 309
correspond to E. typus Cuvier, 1837 (Ortellset al. 1989; Zambelli et al. 1992; Hillyardet al. 1997; Sikes et al. 1997; Tiranti 1997).This distinctive species is the only one in thegenus that has a biarmed X-chromosomemorphology, suggesting that it is a derivedcharacter.The 2n=44, NF=44 karyotype with anancestral X-chromosome is reported herefor the first time. We suggest that thiskaryotype should be assigned to E. maricaThomas, 1918, since its type locality inChumbicha (Catamarca province), is 80 kmfrom our collecting locality (Pipanaco).Nevertheless, the morphological overlap andlimited molecular distinctiveness of E. typusand E. marica, which are in the upper limit ofgenetic distance observed for intraspecificvariation in Eligmodontia from Patagonia(Hillyard et al. 1997), render doubt about itsspecific status. However, the karyotypicdifferentiation between E. typus andE. marica might result in sterility of femalehybrids (King 1993). In fact, the hetero-morphic pairing of X-chromosomes in fe-males is likely to produce duplications anddeficiencies in the absence of preventingmechanisms to chiasma formation (King1993).The close relationship among specimens with2n=44 karyotypes is confirmed by chromo-somal, morphological and molecular ana-lyses. The molecular data show that a sisterspecies to this group is the Patagonian silkymouse E. morgani. Although, their relation-ships have a low support, they are recoveredin the NJ and MP analyses. These three taxashare also a low FN and a central southerndistribution, which sustain their close rela-tionship compared to other Eligmodontiaspecies.In the other group, molecular data indicatethat E. moreni is more related to E. puerulus,and that relationship is recovered in all treeswith high bootstrap. However, this lastspecies has a highly derived karyotype (Spot-orno et al. 2001; Lanzone et al. unpubl. data).On the other hand, the Andean species,E. hirtipes is karyotypically more similar toE. moreni, which appears to have an extrapair of acrocentric autosomes (Spotorno etal. 1994; Lanzone and Ojeda 2005). The high
FN shared among these three northwesterntaxa supports their close phylogenetic rela-tionship.Several studies have dealt with sigmodontinerelationships at the generic level (Smith andPatton 1993, 1999; D’ Elıa 2003; Steppanet al. 2004, and references therein), butanalyses on intrageneric relationships arerather scarce (Steppan 1998). We propose aphylogenetic tree for the silky mice Eligmo-dontia, with two major clades (lineages)representing the ‘‘northern’’ and ‘‘central-southern’’ species. Major clades in our treeare in agreement with previous analysis(Spotorno et al. 2001), but differ with theproposition made by Braun (1993) whoconsiders E. marica as a highly differentiated,early offshoot in the phylogeny of the group.Our results indicate the opposite, and suggesta recent origin as supported by our data sets(i.e. low morphological, karyotypical andmolecular divergence). Moreover, the rela-tionship between E. moreni and E. morgani,as well as between E. hirtipes and E. puerulusas proposed by Braun (1983) is not supportedeither by molecular or karyotypic data.Mapping karyotypes data onto the phylo-geny of Eligmodontia indicates convergenceto low diploid numbers in both clades, withRobertsonian fusion as the major forcedirecting the karyotypic evolution (Gardnerand Patton 1976; Pearson and Patton 1976)in the northern clade. Nevertheless, no suchcorrelation exists in the southern one, wherekaryotypes cannot be derived by invokingRobertsonian rearrangement only. Eventhough relevant banding patterns are notknown yet, the difference between thesekaryotypes strongly suggest a complex arrayof chromosomal rearrangements, involvingpericentric inversions and or tandem fusions(Ortells et al. 1989) in the southern species.Convergent evolution to reduced diploidnumber in different intrageneric lines was ob-served in other related phyllotines (Steppan1998; Salazar-Bravo et al. 2001).The Miocene–Pliocene uplift of the Andes,followed by the Quaternary climatic events,provided a topographically diverse scenariofor the evolution of desert adapted rodents inthe high Puna desert and Andean rockyfoothills (Mares 1975; Reig 1986). The

ARTICLE IN PRESS
310 C. Lanzone et al.
evolution of phyllotines based on chromoso-mal changes suggests that high diploidnumbers, as found in species of the Andeanplateau is the ancestral karyotype (Pearsonand Patton 1976; Spotorno et al. 2001).Accordingly, phylogenetically derived spe-cies, with low diploid numbers and mostlybiarmed karyotypes should be distributedin the lowlands. Nevertheless, we foundhigh and low diploid numbers in both, theAndean plateau and lowlands, complicatingthe classical scheme of chromosomal evolu-tion in the phyllotine rodents (Bonvicinoet al. 2003 and literature cited there; Oldset al. 1987).Regarding the origin of Eligmodontia, Reig(1986) proposed a differentiation of the genusfrom an ancestral, Andean stock of Calomys,to the desert lowlands about 11.6myr (Sala-zar-Bravo et al. 2001). Our results expandReig’s (1986) proposition since they suggestthat arid lowlands acted as a second diversi-fication center. A similar proposition of morethan one divergent lines in the basal radiationof genera was suggested for other species ofsigmodontines (Salazar-Bravo et al. 2001;Steppan 1998).Biogeographically, the north-central portionof the Monte biome is a rain shadow desertsurrounded by Andean and preAndeanmountain ranges. These arid landscapescompose the scenario where the speciationand adaptation processes of the lowland
species of Eligmodontia took place. Inthis region we recorded two areas ofsympatry between E. moreni and E. marica(Campo Arenal) and E. typus and E. moreni(Telteca Reserve). Other contact zones inEligmodontia have been reported for thePatagonian populations of E. typus and E.morgani (Zambelli et al. 1992; Hillyard et al.1997; Sikes et al. 1997). Whether the mechan-isms of coexistence of these sympatric popu-lations are mediated through habitatselection or food partitioning deservesfurther studies and is beyond the scope ofthis research.
Acknowlegements
Our thanks to Steve Lougheed for hisassistance with the molecular analyses andto Agustina Ojeda for her critical commentsand suggestions to the first draft of our ms; toSoledad Albanese, Daniela Rodriguez, Mari-ana Dacar and Silvia Brengio for hercooperation in the field and in the laboratory;Marisa Rosi for help with cranial photo-graphs, and to Nelly Horak for her assistancewith the English version. The map oflocalities was produced by Daniel Duenasat the CRICYT. This study was partiallyfunded by CONICET PIP 2884 and AgenciaSecyt 11768 to RAO in Argentina, and FNC1010727 to MHG in Chile.
Zusammenfassung
Integrative Taxonomie, Systematik und Verbreitung der Gattung Eligmodontia (Rodentia,Cricetidae, Sigmodontinae) in der gemaXigten Monte-Wuste in Argentinien
Systematik und Verbreitung der sigmodontinen Nager Sudamerikas sind Gegenstand standigerRevisionen und Kontroversen. Die Hochland-Wustenmause der Gattung Eligmodontia Cuvier, 1837gehoren zu den am meisten spezialisierten Muriden unter den endemischen Nagern Sudamerikas.Ihre vielfaltige Anpassung an das Wustenleben ist mit der Anhebung der Anden und der fruhenBildung arider Landschaften korreliert. Um die Systematik der Arten von Hochland-Wustenmausen,die den trockensten Teil der Monte-Wuste Argentiniens besiedeln, zu klaren, wurden qualitative undquantitative Korper- und SchadelmaXe, Chromosomen und genetische Beziehungen untersucht. Wirfanden drei verschiedene Karyotypen von Eligmodontia, zwei davon erstmals hier beschrieben, undbeziehen sie auf bereits beschriebene Taxa: E. moreni Thomas, 1896 (2n=52 und FN=50), E. typusCuvier, 1837 (2n=44 und FN=44) und E. marica Thomas, 1918. Die letzte Form hat dieselbe 2n wieE. typus, aber ihr X-Chromosom ist metazentrisch statt akrozentrisch. In einer Diskriminanzanalyseexterner und kranialer Daten wird E. moreni von E. typus und E. marica getrennt, wahrend die letzten

ARTICLE IN PRESS
Systematics of Eligmodontia in the Monte Desert 311
beiden Arten Uberlappungen zeigen. Die morphologische und zytologische Differenzierung vonEligmodontia wird durch DNA Distanzen unterstutzt. Phylogenetische Analysen zeigen zwei groXeGruppen. Eine wird von E. moreni, E. puerulus und E. hirtipes gebildet, die eine hohe FN-Zahl undeine nordliche Verbreitung aufweisen. Die andere Gruppe umfasst E. typus, E. marica und E. morganimit niedriger FN-Zahl und einer zentralen bis sudlichen Verbreitung. Zwei Artbildungszentrenwerden vorgeschlagen, um die Evolution von Eligmodontia zu erklaren.r 2006 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
References
Bonvicino, C. B.; Lima, J. F. S.; Almeida, F. C.(2003): A new species of Calomys Water-house (Rodentia, Sigmodontinae) from theCerrado of Central Brazil. Rev. Bras. Zool.20, 301–307.
Braun, J. K. (1993): Systematic Relationships ofthe Tribe Phyllotini (Muridae:Sigmodontinae)of South America. Spec. Publ. Norman: Okla-homa. Mus. Nat. Hist. Pp. 1–50.
Cabrera, A. (1961): Catalogo de los Mamıferos deAmerica del Sur. Parte II. Rev. Mus. Arg. Cs.Nat. ‘‘Bernardino Rivadavia’’, Cs. Zool 4 (2),309–732.
Cuvier, F. (1837): Ann. Sci. Nat,. Paris 2 (7), 168.D’ Elıa, G. (2000): Comments on recent advances
in understanding sigmodontine phylogeny andevolution. Mastozool. Neotr 7, 47–54.
D’ Elıa, G. (2003): Phylogenetics of Sigmodontinae(Rodentia, Muroidea, Cricetidae), with specialreference to the akodont group, and withadditional comments on historical biogeogra-phy. Cladistics 19, 307–323.
Diaz, G. B. (2001): Ecofisiologıa de pequenosmamıferos de las tierras aridas de Argentina:Adaptaciones renales. Diss. Thesis. Mendoza,Argentina: University of Cuyo.
Diaz, G. B.; Ojeda, R. A. (1999): Kidney structureof Argentine desert rodents. J. Arid Env. 41,453–461.
Fagundes, V.; Sato, Y.; Silva, M. J. J.; Rodrigues,F.; Yonenaga-Yassuda, Y. (2000): A newspecies of Calomys (Rodentia, Sigmodontinae)from Central Brazil identified by its karyotype.Hereditas 133, 195–200.
Ford, C. E.; Hamerton, J. L. (1956): A colchicine,hypotonic citrate, squash sequence for mam-malian chromosomes. S. Technol 31, 247–251.
Gardner, A. L.; Patton, J. L. (1976): Karyotypicvariation in Oryzomyine rodents (Cricetinae)with comments on chromosomal evolution inthe Neotropical complex. Occas. Pap. Mus.Zool. Louisiana State Univ. 49, 1–48.
Hershkovitz, P. (1962): Evolution of NeotropicalCricetine Rodents (Muridae) with special re-ference to the Phyllotine group. Field. Zool 46,1–524.
Hillyard, J. R.; Phillips, C. J.; Birney, E. C.;Monjeau, J. A.; Sikes, R. (1997): Mito-chondrial DNA analysis and zoogeographyof two species of silky desert mice, Eligmodon-
tia, in Patagonia. Z. Saugetierkunde 62,
281–292.Kelt, D. A.; Palma, R. E.; Gallardo, M. H.; Cook,
J. A. (1991): Chromosomal multiformity inEligmodontia (Muridae, Sigmodontinae), andverification of the status of E. morgani. Z.Saugetierkunde 56, 352–358.
King, M. (1993): Species Evolution. The role ofchromosome change. Cambridge: CambridgeUniversity Press.
Lanzone, C.; Ojeda, R. A. (2005): Citotaxonomıa ydistribucion del genero Eligmodontia (Roden-tia, Cricetidae, Sigmodontinae). Mastozool.Neotr. 12, 73–77.
Mares, M. A. (1975): South American mammalzoogeography: Evidence from convergent evo-lution in desert rodents. Proc. Nat. Acad. Sci.72, 1702–1706.
Mares, M. A. (1977): Water economy and saltbalance in a South American desert rodent,Eligmodontia typus. Comp. Biochem. Physiol.56, 325–332.
Martin, R. E.; Pine, R. H.; Deblase, A. F. (2001): AManual of Mammalogy with Keys to Familiesof the World. New York: McGraw-Hill.
Musser, G. G; Carleton, M. D. (2005): MammalSpecies of the World. A Taxonomic andGeographic Reference. Baltimore: The JohnsHopkins University Press, D. E. Wilson and D.M. Reeder.
Olds, A.; Anderson, S.; Yates, T. L. (1987): Noteson Bolivian mammals 3: A revised diagnosis ofAndalgalomys (Rodentia, Muridae) and thedescription of a new subspecies. Am. Mus.Nat. Hist. 2890, 1–17.
Ortells, M. O.; Reig, O. A.; Wainberg, R. L.;Hurtado De Catalfo, G. E.; Gentile De Fronza,T. M. L. (1989): Cytogenetics and karyosyste-matics of Phyllotine rodens (Cricetidae, Sigmo-dontinae). II. Chromosome multiformity andautosomal polymorphism in Eligmodontia. Z.Saugetierkunde 54, 129–140.

ARTICLE IN PRESS
312 C. Lanzone et al.
Patton, J. L. (1967): Chromosome studies in certainpocket mouse, genus Perognathus (Rodentia,Heteromyidae). J. Mammalogy 48, 27–37.
Patton, J. L.; Sherwood, S. W. (1983): Chromo-some evolution and speciation in rodents.Annu. Rev. Ecol. Syst. 14, 139–158.
Pearson, O. P.; Patton, J. L. (1976): Relationshipamong South American phyllotine rodentsbased on chromosomal analysis. J. Mammal-ogy 57, 339–350.
Posada, D.; Crandall, K. A. (1998): Modeltest:Testing the model of DNA substitution. Bioin-formatics 14, 817–818.
Redford, K. H., Eisenberg, J. F. (1992); Mammalsof the Neotropics. Vol. 2. The Southern Cone.Chile, Argentina, Uruguay, Paraguay. Chicago:University Press.
Reig, O. A. (1986): Diversity patterns and differ-entiation of high Andean rodents. In: HighAltitude Tropical Biogeography. Ed. by F.Vuilleumier, M. Monasterio. Oxford: Univer-sity Press. Pp. 404–438.
Salazar-Bravo, J.; Dragoo, J. W.; Tinnin, D. S.;Yetes, T. L. (2001): Phylogeny and evolution ofthe Neotropical rodent genus Calomys: Infer-ences from mitochondrial DNA sequence data.Mol. Phyl. Evol. 20, 173–184.
Sambrook, E.; Fritsch, F.; Maniatis, T. (1989):Molecular Cloning. New York: Cold SpringHarbor, Laboratory Press.
Sikes, R. S.; Monjeau, E. C.; Phillips, C. J.;Hillyard, J. R. (1997): Morphological versuschromosomal and molecular divergence in twospecies of Eligmodontia. Z. Saugetierkunde 62,
265–280.Smith, M. F.; Patton, J. L. (1993): The diversifica-
tion of South American murid rodents: evi-dence from mitochondrial DNA sequence datafor the akodontine tribe. Biol. J. Linn. Soc. 50,149–177.
Smith, M. F.; Patton, J. L. (1999): Phylogeneticrelationships and the radiation of sigmodontinerodents in South America: evidence fromcytochrome b. J. Mamm. Evol. 6, 89–128.
Spotorno, A. E.; Sufan-Catalan, J.; Walker, L. I.(1994): Cytogenetic diversity and evolution ofAndean species of Eligmodontia (Rodentia,Muridae). Z. Saugetierkunde 59, 299–308.
Spotorno, A. E.; Walker, L. I.; Flores, S. V.;Yevenes, M.; Marın, J. C.; Zuleta, C. (2001):Evolucion de los filotinos (Rodentia, Muridae)
en los Andes del Sur. Rev. Chil. Hist. Nat. 74,151–166.
Steppan, S. J. (1995): Revision of the tribePhyllotini (Rodentia: Sigmodontinae), with aphylogenetic hypothesis for the Sigmodontinae.Field Zool. 80, 1–112.
Steppan, S. J. (1998): Phylogenetic relationshipsand species limits within Phyllotis (Rodentia:Sigmodontinae): Concordance between mtDNAsequence and morphology. J. Mammalogy 79,
573–593.Steppan, S. J.; Adkins, R. M.; Anderson, J. (2004):
Phylogeny and divergence-data estimates ofrapid radiations in muroid rodents based onmultiple nuclear genes. Syst. Biol. 53, 533–553.
Swofford, D. L. (2002): PAUP*. PhylogeneticAnalysis Using Parsimony (*and other meth-ods), 4. Sunderland, Massachusetts: SinauerAssociates.
Thomas, O. (1896): On the genera of rodents: anattempt to bring up to date the currentarrangement of the Order. Proc. Zool. Soc.London 1896, 1012–1028.
Thomas, O. (1918): A new species of Eligmodontia
from Catamarca. Annu. Mag. Nat. Hist. 9,
482–484.Thompson, J. D.; Gibson, T. J.; Plewniak, F.;
Jeanmougin, F.; Higgins, D. G. (1997): TheClustal X windows interface: flexible strategiesfor multiple sequence alignment aided byquality analysis tools. Nucleic Acids Res 24,
4876–4882.Tiranti, S. I. (1997): Cytogenetic of silky desert
mice Eligmodontia spp. (Rodentia, Sigmodon-tinae) in central Argentina. Z. Saugetierkunde62, 37–42.
Zambelli, A.; Dyzenchauz, F.; Ramos, A.; DeRosa, N.; Wainberg, R.; Wilson, A. C. (1992):Cytogenetics and karyosystematic of phyllotinerodents (Cricetidae, Sigmodontinae). III. Newdata on the distribution and variability ofkaryomorphs of the genus Eligmodontia. Z.Saugetierkunde 57, 155–162.
Authors’ addresses:
Cecilia Lanzone and Ricardo Ojeda, GIB, IADIZA,CONICET, CC 507, 5500 Mendoza, Argentina(e-mail: [email protected])Milton H. Gallardo, Instituto de Ecologıa y Evolu-cion, Universidad Austral de Chile, Casilla 567,Valdivia, Chile




















