
Symmetric DNA Sites are Functionally Asymmetric Within Flp and Cre Site-specific DNA Recombination...
13
UNCORRECTED PROOF Symmetric DNA Sites are Functionally Asymmetric Within Flp and Cre Site-specific DNA Recombination Synapses Ian Grainge 1 , Shailja Pathania 1 , Alexander Vologodskii 2 Rasika M. Harshey 1 and Makkuni Jayaram 1 * 1 Section of Molecular Genetics and Microbiology and the Institute of Cell and Molecular Biology, University of Texas at Austin, Austin TX 78712, USA 2 Department of Chemistry New York University, 31 Washington Place, New York NY 10003, USA Flp and Cre-mediated recombination on symmetrized FRT and loxP sites, respectively, in circular plasmid substrates yield both DNA inversion and deletion. However, upon sequestering three negative supercoils out- side the recombination complex using the res II– res III synapse formed by Tn3 resolvase and the LER synapse formed by phage Mu transposase in the case of Flp and Cre, respectively, the reactions are channeled towards inversion at the expense of deletion. The inversion product is a trefoil, its unique topology being conferred by the external resolvase or LER synapse. Thus, Flp and Cre assign their symmetrized substrates a strictly antiparallel orientation with respect to strand cleavage and exchange. These conclusions are supported by the product profiles from tethered parallel and antiparallel native FRT sites in dilution and competition assays. Furthermore, the observed recombination bias favoring deletion over inversion in a nicked circular substrate containing two symmetrized FRT sites is consistent with the predictions from Monte Carlo simulations based on antiparallel synapsis of the DNA partners. q 2002 Elsevier Science Ltd. All rights reserved Keywords: symmetric recombination sites; DNA desymmetrization; hybrid recombination synapses; difference topology; Monte Carlo simulation *Corresponding author Introduction The chemical steps of recombination by the inte- grase/tyrosine family recombinases are executed within the context of a tetrameric assembly of the recombinase protein arranged on two relatively short DNA partners of defined sequence. 1–4 The Flp protein, encoded by the 2 m plasmid of Saccharomyces cerevisiae and the Cre protein encoded by the bacteriophage P1 are members of this family. In each system, recombination occurs readily between two copies of the respective 34 bp target sites (the FRT site for Flp and the lox site for Cre), each bound by two monomers of the corre- sponding recombinase. The Flp protein plays a central role in the copy number control of the yeast plasmid. 2,5 The Cre protein is believed to facilitate the segregation of the unit copy P1 phage/plasmid by resolving post-replication dimer genomes into monomers. 6 The strand exchange regions (or spacers) in Flp and Cre are 8 bp and 6 bp long, and are asym- metric in sequence. This asymmetry may, in principle, confer directionality to the recombina- tion target sites and hence to the recombination reaction. A misdirected exchange would be highly disfavored because of the resulting base non- complementarity in the spacer of the recombinant product. Depending on the orientation of the recombination sites, head-to-head or head-to-tail, Flp and Cre mediate inversion and deletion reactions efficiently. When the spacer sequence is artificially symmetrized while retaining homology between partners, recombination can cause either inversion or deletion with roughly equal probability. 7,8 The substrates containing symmetric spacers used in the present study are referred to as symmetrized even when the dyad symmetry between their recombinase-binding elements is not perfect. This nomenclature is justified, since the binding elements do not partake 0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved Present address: I. Grainge, Imperial Cancer Research Fund, Clare Hall Laboratories, Potters Bar, Hertfordshire EN6 3LD, UK. E-mail address of the corresponding author: [email protected] Abbreviations used: TAP, tethered antiparallel; RAP, radioactive product. doi:10.1016/S0022-2836(02)00517-X available online at http://www.idealibrary.com on B w J. Mol. Biol. (2002) 320, 515–527
-
Upload
newcastle-au -
Category
Documents
-
view
4 -
download
0
Transcript of Symmetric DNA Sites are Functionally Asymmetric Within Flp and Cre Site-specific DNA Recombination...

UNCORRECTED PROOF
Symmetric DNA Sites are Functionally AsymmetricWithin Flp and Cre Site-specific DNARecombination Synapses
Ian Grainge1, Shailja Pathania1, Alexander Vologodskii2
Rasika M. Harshey1 and Makkuni Jayaram1*
1Section of Molecular Geneticsand Microbiology and theInstitute of Cell and MolecularBiology, University of Texas atAustin, AustinTX 78712, USA
2Department of ChemistryNew York University, 31Washington Place, New YorkNY 10003, USA
Flp and Cre-mediated recombination on symmetrized FRT and loxP sites,respectively, in circular plasmid substrates yield both DNA inversionand deletion. However, upon sequestering three negative supercoils out-side the recombination complex using the res II–res III synapse formed byTn3 resolvase and the LER synapse formed by phage Mu transposase inthe case of Flp and Cre, respectively, the reactions are channeled towardsinversion at the expense of deletion. The inversion product is a trefoil, itsunique topology being conferred by the external resolvase or LERsynapse. Thus, Flp and Cre assign their symmetrized substrates a strictlyantiparallel orientation with respect to strand cleavage and exchange.These conclusions are supported by the product profiles from tetheredparallel and antiparallel native FRT sites in dilution and competitionassays. Furthermore, the observed recombination bias favoring deletionover inversion in a nicked circular substrate containing two symmetrizedFRT sites is consistent with the predictions from Monte Carlo simulationsbased on antiparallel synapsis of the DNA partners.
q 2002 Elsevier Science Ltd. All rights reserved
Keywords: symmetric recombination sites; DNA desymmetrization; hybridrecombination synapses; difference topology; Monte Carlo simulation*Corresponding author
Introduction
The chemical steps of recombination by the inte-grase/tyrosine family recombinases are executedwithin the context of a tetrameric assembly of therecombinase protein arranged on two relativelyshort DNA partners of defined sequence.1 – 4 TheFlp protein, encoded by the 2 m plasmid ofSaccharomyces cerevisiae and the Cre proteinencoded by the bacteriophage P1 are members ofthis family. In each system, recombination occursreadily between two copies of the respective 34 bptarget sites (the FRT site for Flp and the lox site forCre), each bound by two monomers of the corre-sponding recombinase. The Flp protein plays acentral role in the copy number control of theyeast plasmid.2,5 The Cre protein is believed to
facilitate the segregation of the unit copy P1phage/plasmid by resolving post-replicationdimer genomes into monomers.6
The strand exchange regions (or spacers) in Flpand Cre are 8 bp and 6 bp long, and are asym-metric in sequence. This asymmetry may, inprinciple, confer directionality to the recombina-tion target sites and hence to the recombinationreaction. A misdirected exchange would be highlydisfavored because of the resulting base non-complementarity in the spacer of the recombinantproduct. Depending on the orientation of therecombination sites, head-to-head or head-to-tail,Flp and Cre mediate inversion and deletionreactions efficiently. When the spacer sequence isartificially symmetrized while retaininghomology between partners, recombination cancause either inversion or deletion with roughlyequal probability.7,8 The substrates containingsymmetric spacers used in the present study arereferred to as symmetrized even when the dyadsymmetry between their recombinase-bindingelements is not perfect. This nomenclature isjustified, since the binding elements do not partake
0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved
Present address: I. Grainge, Imperial Cancer ResearchFund, Clare Hall Laboratories, Potters Bar, HertfordshireEN6 3LD, UK.
E-mail address of the corresponding author:[email protected]
Abbreviations used: TAP, tethered antiparallel; RAP,radioactive product.
doi:10.1016/S0022-2836(02)00517-X available online at http://www.idealibrary.com onBw
J. Mol. Biol. (2002) 320, 515–527

UNCORRECTED PROOF
in the exchange reaction and are neutral in deter-mining the directionality of recombination.
Biochemical, topological and structural datafrom Flp and Cre are consistent with the recom-bination reaction being initiated by an anti-parallel arrangement of the recombinationpartners and proceeding through a roughlysquare planar antiparallel Holliday junction.9 – 13
However, analyses using electron microscopy,atomic force microscopy and proximity effectson ligation of DNA ends have suggested thepossibility of parallel synapse during integrase/tyrosine family recombination.14,15 The morerecently solved Flp–DNA crystal structure hasnot clarified the issue because of the unexpectedcleavage of all four scissile phosphodiesters inthe synaptic complex.9 The experiments reportedhere attempt to provide a definitive resolutionof the ambiguity in synapse geometry by usinga combination of substrate designs and analyticalmethods.
By fixing the number of DNA crossings externalto the recombination synapse in two differentways, we show that Flp and Cre treat symmetrizedrecombination sites within their respectivesynapses as exclusively antiparallel. This con-clusion is supported by recombination outcomesin tethered molecules at different substratedilutions or in the presence of a competitor sub-strate. Additional support is provided by theagreement between theory and experiment for theobserved direction of recombination when sym-metrized sites react in supercoiled versus nickedcircular molecules.
Results
Experimental strategies
Because of the conformational flexibility of thedouble helix in the hundreds to thousands ofbase-pairs range, recombination products obtainedfrom supercoiled plasmid substrates often displaya distribution of conformational complexity. As aresult, it is difficult to obtain from the productensemble useful information on the conformationalstate or dynamics at the reaction center. We haveused two approaches to overcome this difficulty.In the first, we have restricted the possible DNApaths between two recombination sites either byarranging a specific DNA–protein complex intheir vicinity or by linking them through a veryshort DNA tether. Second, we used computersimulation to account quantitatively for all possibleconformations of the DNA segments connectingspecific sites in the synaptic complex. We couldthen test recombination models by computing thedistribution of the reaction products for a particu-lar recombination mechanism and comparing itwith the corresponding experimentally measureddistribution.
A detailed explanation of the rationale for thedifference topology analysis employed in some ofthe experiments has been given by Grainge et al.and Pathania et al.10,16 Note that, as discussed byGrainge et al.,10 the recombination synapse isinferred to be parallel or antiparallel based onwhether the act of strand exchange introduces aDNA crossing or not.
Recombination in symmetrized FRT sites:predicted effects of trapping three supercoilnodes outside the Flp synapse
In the symmetrized FRT site (FRTsym), the T-Aand G-C base-pair at the fifth and seventh spacerpositions from the left were changed to G-C andA-T, respectively (Figure 1, top). In plasmid pFRT-Sym (Figure 1, bottom), a copy of FRTsym wascloned proximal to each of a duplicated set of theTn3 resolvase accessory sites res II and res IIIarranged in a head-to-tail orientation. The overallorganization of pFRT-Sym, containing two FRT–res II–res III cassettes, was the same as that of theres/FRT hybrid plasmids described by Graingeet al.10 The cross-over points within the FRT sitesdivide pFRT-Sym into two DNA domains, shownin Figure 1(b) in dark and light shades. Previouswork has demonstrated that the resolvase-mediated res II–res III synapse can be stabilizedwith the help of a secondary synapse formed byFlp or Cre at FRT or loxP sites, respectively, locatedat appropriate distances from res II–res III.10,12 Thehybrid synapse is topologically equivalent to thewell-characterized native resolvase synapse withthree trapped interdomainal supercoil nodes(referred to as 23).17 – 20
Unlike the native FRT sites, the symmetrizedsites are not restricted in the orientation of strandexchange by base-pairing constraints within thespacer. Formally, there are three potential outcomesfor Flp recombination at FRT-Sym from the hybridresolvase/Flp synapse (see Figure 1). Strandexchange in the antiparallel sense will yield a (þ3)trefoil knot as the DNA inversion product.Crossing-over in the parallel sense will give eithera 22 catenane (if the recombination node is þ1)or a 24 catenane (if the recombination node is21) as the deletion product. The change of signfrom 23 in the synapse to þ3 in the product knotis because of the inversion of the darkly shadedDNA domain with respect to the lightly shadedone. We have tested, as described below, which ofthe three possible outcomes depicted here isexperimentally realized.
Note that the resolvase–Flp synapsediagrammed in Figure 1 is the simplest planarprojection of a recombination complex thataccounts for our earlier findings;10 namely, theunique enrichment of the þ 3 trefoil and the 24catenane during resolvase assisted Flp inversionand deletion, respectively (see also Figure 2). Thetopological invariance of the external res II–res IIIsynapse, as inferred from prior results,10 is central
516 Asymmetry in Integrase/Tyrosine Family Recombination

UNCORRECTED PROOF
to interpreting the results in Figure 2. Otherwise,the topology of the recombination products fromthe hybrid synapse would not be unique. Weemphasize this point because resolvase subunitsbound at the crossover sites (res I) also contributeto the functional native resolvase synapse,21,22 andres I is absent from our plasmid substrates. At leastwith the particular FRT–res II–res III configurationsthat we have utilized, absence of res I does notappear to topologically alter the 23 resolvasesynapse.
Topological analyses of products fromresolvase-assisted Flp recombination inplasmid substrates containing wild-type andsymmetrized FRT sites
In the three plasmid substrates (Figure 2), pFRT-Inv, pFRT-Del and pFRT-Sym, the FRT sites havesimilar relative locations with respect to the pairof res II–res III sites. For each substrate, thespacings between res II and FRT (as measuredfrom the midpoint of the two sites) were 133 bp inone FRT–res II–res III cassette and 143 bp in theother. As described previously,10 these distanceswere optimized to minimize random entrapmentof supercoils during Flp recombination, while atthe same time avoiding potential steric clashesbetween resolvase and Flp subunits that mightinterfere with the stable assembly of the hybridsynapse. All the assays shown in Figure 2 werecarried out at the standard pH of 7.5 normally
used for Flp recombination. At this pH, the recom-bination products from negatively supercoiledplasmids cover a relatively wide distribution oftopological complexity.
The FRT sites are in inverted orientation in pFRT-Inv (the inversion susbtrate) and in direct orien-tation in pFRT-Del (the deletion substrate). InpFRT-Sym, with the symmetrized FRT sites, Flprecombination can give inversion as well asdeletion. In control reactions, each of the plasmidswas reacted with Flp in the absence of Tn3resolvase. In the test reactions, each plasmid waspre-incubated with the resolvase protein and thenreacted with Flp.10 The nicked DNA samples wereanalyzed by agarose gel electrophoresis toreveal the topology of the recombination products(Figure 2).
For pFRT-In and pFRT-Del, the major productswith Flp alone were either knots or catenanes,respectively, with a wide distribution of com-plexities (lanes 2 and 5, Figure 2). In the resolvaseplus Flp reaction, the four-noded catenane productfrom pFRT-Del (lane 3, Figure 2) and the three-noded knot product from pFRT-In (lane 6,Figure 2) were uniquely enriched relative to therest of the product ladder. The slight increase inthe bands at positions 5 and 4 in lanes 3 and 6,respectively, would be consistent with a secondrecombination event occurring from the primaryrecombination products (as explained by Graingeet al.10). The fainter bands running between therungs of the major product ladders in lanes 2 and
Figure 1. Plausible outcomes ofrecombination at a pair of sym-metrized FRT sites from a hybridFlp/resolvase synapse. The wild-type and symmetrized FRT sitesdiffer only in the sequence of theirspacer (or core regions; indicatedby bold letters) The Flp bindingelements that flank the spacer inhead to head orientation are high-lighted by the horizontal arrows.Plasmid pFRT-Sym contains twocopies of the Tn3 resolvaseaccessory sites res II–res III in directorientation.10 The symmetrized FRTsites are drawn as the double-headed arrows. In the resolvase/Flp hybrid synapse, the FRT sitescan potentially cross over in threeways: adding no DNA node (anti-parallel mode) or adding a þ1 or a21 DNA node (parallel mode).
GAAGTTCCTATACTTTCTAGAGAATAGGAACTTC
GAAGTTCCTATACTTTCGAAAGAATAGGAACTTC
Wild type FRT site
Symmetrized core FRT site
CTTCAAGGATATGAAAGATCTCTTATCCTTGAAG
CTTCAAGGATATGAAAGCTTTCTTATCCTTGAAG
+3 trefoil
Resolvase/Flp
synapse
Flp-mediated
recombination
0
+1
-1
-2 catenane
-4 catenane
pFRT-Sym
Asymmetry in Integrase/Tyrosine Family Recombination 517

UNCORRECTED PROOF
3, and 5 and 6 were the result of an extra round ofrecombination that the corresponding lowerhierarchy products had gone through. Appropriaterestriction enzyme digestion showed that the DNAin these bands was in the parental configuration, asexpected for two (or even) rounds of recombina-tion (data not shown).
The products of the reaction of pFRT-Sym withFlp (lane 8, Figure 2) consisted of two super-imposed ladders, one with odd-numbered nodesand the other with even-numbered nodes.Neighboring odd and even-numbered bands wereapproximately equal in intensity, suggesting thatthey arose primarily by independent recombina-tion events. Consistent with this inference, restric-tion enzyme digestion of the recombinationreaction revealed approximately equal amounts ofthe inversion and deletion products (data notshown). In the pFRT-Sym/resolvase/Flp reaction(lane 9, Figure 2), the product distribution dis-played the prominent enrichment of the three-
noded knot as the primary recombinant and amodest increase in the four-noded product derivedfrom it by a subsequent recombination event. Thepattern was nearly identical with that seen in thepFRT-Inv/resolvase/Flp reaction (compare lanes 6and 9 in Figure 2).
Thus, Flp treats the pFRT-Sym plasmid as aninversion substrate when three resolvase trappedplectonemic nodes are placed outside the Flpsynapse. In the absence of this topological con-straint, Flp is equally good at performing inversionor deletion in pFRT-Sym. Or, recombinationproceeds by the central pathway in Figure 1(b)without the addition of a DNA crossing.
As explained in detail by Grainge et al.,10 a 23synapse would bring two head-to-head FRT sitesin antiparallel alignment, and therefore yield þ3knots by Flp-mediated inversion. The same 23synapse would impose a parallel geometry on twohead-to-tail FRT sites. The ensuing FRT pairing islikely unstable,10 and rearranges to the antiparallel
2345
-
+-
-
+
+resolvase
Flp
4
2
unlinked circle
-
+-
-
+
+ -
+-
-
+
+
product
nodes
open
circle
3
56
79
supercoiled
substratehighly linked
products
1 2 3 4 5 6 7 8 9
pFRT-Del pFRT-Inv pFRT-Sym
resIII II
FRT
site spacing
Figure 2. Recombination reactions from the Flp synapse or Flp/resolvase hybrid synapse in negatively supercoiledplasmids containing wild-type or symmetrized FRT sites. The Flp-resolvase hybrid synapses were established in plas-mids pFRT-Del, pFRT-Inv and pFRT-Sym from a pair of FRT–res II–res III sites diagrammed at the top. The site spacingrefers to the distance between the midpoints of res II and FRT, 143 bp in one cassette and 133 bp in the other. Reactionswere nicked with DNase I,10 and analyzed by electrophoresis in agarose gels. For each reaction set, the left lane was acontrol in the absence of either Flp or resolvase, the middle lane was an Flp-alone reaction, and the right lane was anFlp reaction after the resolvase synapse had been pre-assembled. The numbers against the ladder of bands are theinferred DNA crossings within the knots and catenanes.
518 Asymmetry in Integrase/Tyrosine Family Recombination

UNCORRECTED PROOF
synapse by trapping an extra negative supercoil.The resulting recombination product from pFRT-Del would be a 24 catenane. The energetic costfor sequestering the additional negative node islikely to be minimal in a negatively supercoiledplasmid, accounting for the nearly equal amountsof the þ3 knot and 24 catenane produced frompFRT-In and pFRT-Del, respectively. In principle,the 23 synapse or the 24 synapse would makeno difference to the reactivity of the symmetrizedFRT sites. However, since the pre-arrangedresolvase synapse leads to productive FRT pairingand recombination in pFRT-Sym, there is littleopportunity to sample the 24 synapse. As a result,the þ3 knot is formed in significant excess over the24 catenane.
Flp recombination in tethered parallel andantiparallel FRT sites
The topological interpretations from theresolvase–Flp hybrid synapse may be verified bydetermining the ability of a pair of FRT sites thatare constrained to be either parallel or antiparallelto partake in recombination. The prediction is thatonly the antiparallel sites would be active. The
underlying assumption in these experiments (as isthe case for the topological analyses presented inFigure 2) is that the axes of the DNA partnerswithin the Flp recombination complex are essen-tially coplanar. The crystal structures of Flp andCre recombination complexes9,11 validate thisassumption.
To restrict the relative geometry of the FRT sites,we employed single-stranded T10 tethers (Figure 3)linking two “top strands” (for the antiparallel sub-strate TAP) or a top and a bottom strand (for theparallel substrate TP).23 The cross-over points onthe top and bottom strands are denoted by thefilled circles and squares, respectively. For end-labeled TAP, the radioactive product (RAP) ofintramolecular bottom strand exchange (B1 £ B2),which can occur only from an antiparallel synapse,would be 42 nt long (Figure 3, bottom left). For theTP substrate, the expected labeled recombinantstrand (RP) resulting from an intramolecular paral-lel synapse and exchange of top strands (T1 £ T2)would be 55 nt long (Figure 3, bottom right). How-ever, the recombinants RAP or RP can result fromintermolecular reactions, which Flp is proficient atcarrying out. Intermolecular recombination, beingunconstrained, can occur from parallel synapse or
5’
3’
3’
5’
TTTTTGAGATAGCGAAGTTCCTATACTTTCTAGAGAATAGGAACTTCAGAGCTCTATCGCTTCAAGGATATGAAAGATCTCTTATCCTTGAAGTCTC
TCAGGCGAGAAGTTCCTATTCTCTAGAAAGTATAGGAACTTCCGTCTTTTTAGTCCGCTCTTCAAGGATAAGAGATCTTTCATATCCTTGAAGGCAG
5’
3’
T-10 tether
17 nt29 nt
21 nt 25 nt
T1
T2
B1
B2
B1 x B2 RAP (42 nt)
*
*
Intramolecular recombination
Recombination between tethered antiparallel FRT sites (TAP)
TTTTTCTCTGAAGTTCCTATTCTCTAGAAAGTATAGGAACTTCGCTATCTCGAGACTTCAAGGATAAGAGATCTTTCATATCCTTGAAGCGATAGAG
TCAGGCGAGAAGTTCCTATTCTCTAGAAAGTATAGGAACTTCCGTCTTTTTAGTCCGCTCTTCAAGGATAAGAGATCTTTCATATCCTTGAAGGCAG
T-10 tether
5’
3’
3’
5’
5’
3’
21 nt25 nt
34 nt 78 nt
T1
T2
B1
B2
T1 x T2 RP (55 nt)
*
*
Intramolecular recombination
Recombination between tethered parallel FRT sites (TP)
Figure 3. Recombination in tethered parallel and antiparallel FRT sites. The sequences of the tethered antiparallel(TAP) and parallel (TP) substrates are shown at the top. In the schematics below, the top and bottom strands aredenoted by T and B, respectively. The asterisks denote the radioactive label at a 50 end used to follow recombination.The points of strand breakage and joining on the top strands are indicated by the filled circles; those on the bottomstrands by the filled squares. An intramolecular, antiparallel recombination event between B1 and B2 strands in TAPcan give the radioactive recombinant strand RAP (bottom left). A corresponding parallel recombination in TP willyield RP (bottom right). RAP and RP can also result from unconstrained intermolecular reactions.
Asymmetry in Integrase/Tyrosine Family Recombination 519

UNCORRECTED PROOF
antiparallel synapse, regardless of the mode oftethering. Hence, the tethered substrates are usefulonly as long as recombination can be manipulatedto be intramolecular.
In order to oppose the intermolecular contri-bution to recombination, we carried out Flp/TAPor Flp/TP reactions by serially diluting them overa tenfold range (Figure 4). This range was basedon preliminary assays that showed recombinationin the TP substrate to be markedly reduced at ten-fold dilution and virtually undetectable at higherdilutions. Each reaction mixture contained a fixedamount of the tethered substrate (0.02–0.03 pmol).Since there are four Flp-binding elements per sub-strate molecule, this corresponds to roughly0.1 pmol of target for the Flp monomer. The sub-strate concentration in the undiluted reactionmixture (20 ml) was thus approximately 5 nMbinding element. The Flp concentration was main-tained constant at all dilutions at 20 nM (the Kd forFlp binding being in the nanomolar range), whichis equivalent to four Flp monomers per bindingelement in the undiluted sample.
For the TAP substrate, there was little decreasein recombinant production up to fivefold dilutionof the reaction mixture (RAP in lanes 2–4, Figure4). At the tenfold dilution, the RAP outputdropped to approximately 70–75% (lane 5,Figure 4). The effect of dilution was much morestriking for the TP substrate. The production of RPfell to approximately 45%, 20% and 6% at two-,five- and tenfold dilutions, respectively (comparethe levels of RP in lanes 8–10 to that in lane 7,Figure 4).
The sharply contrasting effects of dilution onTAP and TP recombination are readily accom-modated by the functional Flp synapse being anti-parallel and most, if not all, of the product fromTP arising by intermolecular recombination.Assuming synapsis between two Flp-bound TPmolecules to be second-order (and rate-limiting inrecombination), the rate of the intermolecular reac-tion should decrease as the square of the dilutionfactor. The observed data tend towards thispattern. The small discrepancy from the theoreticalprediction could be due to departure from strictMichaelis–Menten kinetics and/or deviationsfrom initial velocity conditions. If all of the recom-bination in the TAP substrate was essentially intra-molecular, dilution should not affect the output ofRAP. The small decrease in RAP at the tenfolddilution may be accounted for partly by the dimin-ution in the intermolecular contribution to totalrecombination. There could be some effect due tothe high molar ratio of Flp monomer to bindingelement at this dilution (40:1).
Regardless of the quantitative subtleties con-sidered above, the indifference of TAP sharply con-trasted by the sensitivity of TP to dilution upholdsthe conclusion that the active Flp synapse isantiparallel.
Recombination in tethered Flp substrates inthe presence of a competing substrate
If recombination in the tethered parallel sub-strate (TP) is possible only in the intermolecularmode, addition of a second, competitor substratemust inhibit this reaction. Conversely, if a largefraction of the recombination events in the tetheredantiparallel substrate (TAP) is intramolecular, itshould be significantly more resistant to thecompetitor.
In Figure 5, the effects of adding increasingamounts of a linear DNA fragment containing anFRT site (LC) to the TAP or the TP reaction areshown. The position of the 50 label (asterisk) in thetethered substrates and the linear competitor areshown schematically at the top of the figure. Theamount of TAP or TP was held constant in eachreaction, while that of LC was increased to givemolar ratios of LC/TAP (or TP) ¼ 0.5, 1, 2, 4, 8and 40 (lanes 8–3 and 16–11, Figure 5). The pro-ducts of interest were RAP and RP (intramolecularor intermolecular recombinants from self reactionsof TAP and TP, respectively) and TAP £ LC and
TAP
RAP
CL
1 x2
x5
x1
0
no p
rote
in
**
TAP TP
+Flp
1 x2
x5
x1
0
no p
rote
in
+Flp
TP
RP
CL
dilution factor
1 2 3 4 5 6 7 8 9 10
Figure 4. Effect of dilution on the yield of recombina-tion product from tethered antiparallel and tetheredparallel substrates. Reactions were done using the sub-strates TAP and TP (see Figure 3) carrying 50 end labels(asterisks). Each incubation mixture within a reactionset with TAP or TP contained the same amount of sub-strate (lanes 2–5), but were diluted differently in therecombination buffer over a tenfold range, as indicated.The concentration of Flp was kept constant in eachreaction at 20 nM. The reactions were fractionated indenaturing polyacrylamide gels, and analyzed by phos-phorimaging. The recombinant products from TAP andTP are marked as RAP and RP, respectively. The bandsdenoted by CL stand for the strand cleavage product byFlp. Heterogeneity in CL was contributed by a mixtureof short DNA-linked Flp peptides resulting from treat-ment with proteinase K.
520 Asymmetry in Integrase/Tyrosine Family Recombination
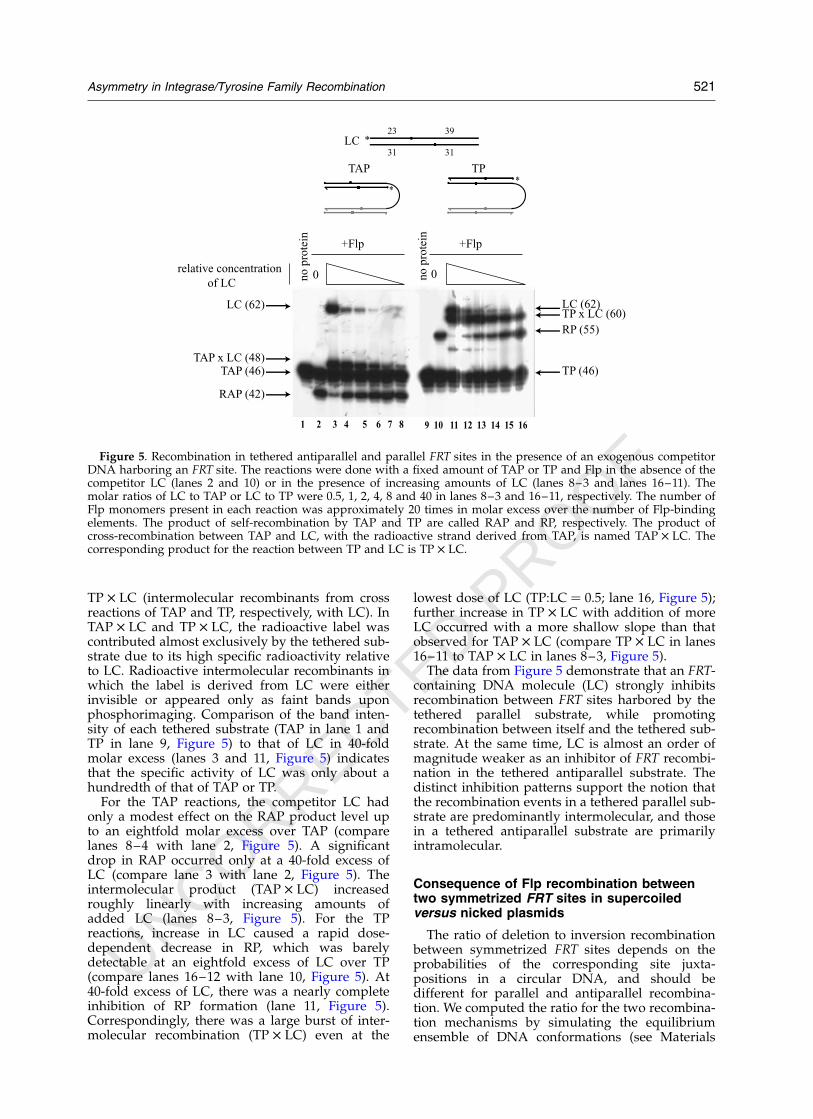
UNCORRECTED PROOF
TP £ LC (intermolecular recombinants from crossreactions of TAP and TP, respectively, with LC). InTAP £ LC and TP £ LC, the radioactive label wascontributed almost exclusively by the tethered sub-strate due to its high specific radioactivity relativeto LC. Radioactive intermolecular recombinants inwhich the label is derived from LC were eitherinvisible or appeared only as faint bands uponphosphorimaging. Comparison of the band inten-sity of each tethered substrate (TAP in lane 1 andTP in lane 9, Figure 5) to that of LC in 40-foldmolar excess (lanes 3 and 11, Figure 5) indicatesthat the specific activity of LC was only about ahundredth of that of TAP or TP.
For the TAP reactions, the competitor LC hadonly a modest effect on the RAP product level upto an eightfold molar excess over TAP (comparelanes 8–4 with lane 2, Figure 5). A significantdrop in RAP occurred only at a 40-fold excess ofLC (compare lane 3 with lane 2, Figure 5). Theintermolecular product (TAP £ LC) increasedroughly linearly with increasing amounts ofadded LC (lanes 8–3, Figure 5). For the TPreactions, increase in LC caused a rapid dose-dependent decrease in RP, which was barelydetectable at an eightfold excess of LC over TP(compare lanes 16–12 with lane 10, Figure 5). At40-fold excess of LC, there was a nearly completeinhibition of RP formation (lane 11, Figure 5).Correspondingly, there was a large burst of inter-molecular recombination (TP £ LC) even at the
lowest dose of LC (TP:LC ¼ 0.5; lane 16, Figure 5);further increase in TP £ LC with addition of moreLC occurred with a more shallow slope than thatobserved for TAP £ LC (compare TP £ LC in lanes16–11 to TAP £ LC in lanes 8–3, Figure 5).
The data from Figure 5 demonstrate that an FRT-containing DNA molecule (LC) strongly inhibitsrecombination between FRT sites harbored by thetethered parallel substrate, while promotingrecombination between itself and the tethered sub-strate. At the same time, LC is almost an order ofmagnitude weaker as an inhibitor of FRT recombi-nation in the tethered antiparallel substrate. Thedistinct inhibition patterns support the notion thatthe recombination events in a tethered parallel sub-strate are predominantly intermolecular, and thosein a tethered antiparallel substrate are primarilyintramolecular.
Consequence of Flp recombination betweentwo symmetrized FRT sites in supercoiledversus nicked plasmids
The ratio of deletion to inversion recombinationbetween symmetrized FRT sites depends on theprobabilities of the corresponding site juxta-positions in a circular DNA, and should bedifferent for parallel and antiparallel recombina-tion. We computed the ratio for the two recombina-tion mechanisms by simulating the equilibriumensemble of DNA conformations (see Materials
TAP (46) TP (46)
LC (62) LC (62)
TAP x LC (48)
RAP (42)
0relative concentration
of LC
RP (55)
0
31 31
23 39
TP x LC (60)
*
TAP*
TP
LC *
no p
rote
in
no p
rote
in
+Flp +Flp
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Figure 5. Recombination in tethered antiparallel and parallel FRT sites in the presence of an exogenous competitorDNA harboring an FRT site. The reactions were done with a fixed amount of TAP or TP and Flp in the absence of thecompetitor LC (lanes 2 and 10) or in the presence of increasing amounts of LC (lanes 8–3 and lanes 16–11). Themolar ratios of LC to TAP or LC to TP were 0.5, 1, 2, 4, 8 and 40 in lanes 8–3 and 16–11, respectively. The number ofFlp monomers present in each reaction was approximately 20 times in molar excess over the number of Flp-bindingelements. The product of self-recombination by TAP and TP are called RAP and RP, respectively. The product ofcross-recombination between TAP and LC, with the radioactive strand derived from TAP, is named TAP £ LC. Thecorresponding product for the reaction between TP and LC is TP £ LC.
Asymmetry in Integrase/Tyrosine Family Recombination 521

UNCORRECTED PROOF
and Methods) and compared it with correspondingexperimental measurements. To extract the ratiofrom the simulated ensemble, we selected confor-mations containing juxtaposed FRT sites in anorientation close to that revealed by the Flp–Holliday junction crystal structure9 (see Figure 6)and determined the recombination products foreach of those conformations (Figure 7(a)).
To experimentally measure the deletion toinversion ratio, Flp reactions using pFRT-Symwere digested with Xho I (Figure 7(b) and (c)) todistinguish the inversion (I1, I2) and deletionproducts (D1, D2) from the parental plasmidbands (P1, P2). The shorter inversion product I2,being only approximately 525 bp long, hadmigrated out of the gel boundary. The D1 and I1
Figure 6. Monte Carlo simu-lations of recombination in circularDNA with symmetric FRT sites.(a) The variables that define themutual orientation of the recombi-nation sites (shown in red) residenton the modeled DNA chain areindicated. The relevant details ofthe simulation procedures are out-lined in Materials and Methods,and in the text. Vectors b1 and b2
are perpendicular to a1 and a2,respectively, and are in the planesof the sites. The angles that appearin equation (1) for the potential inthe umbrella method (see ModelingDNA and simulation procedures inMaterials and Methods) are definedas:
w ¼ arccoslða1a2Þl
a1a2
� �; u ¼ arccos
2ðb1b2Þ
b1b2
� �; z ¼ arccos
ðb1rÞ
b1r
� �
Typical simulated conformations for (b) relaxed and (c) supercoiled DNA are shown. In both conformations, therecombination sites are in proper juxtaposition to form the functional synaptic complex.
a
a
b
b
r1
2
1
2
A
B C
I1
P1
P2
D1
D2
1537 bp
XhoI
XhoI
Flp- + - +
XhoI+ + + +
(σ)
A B C
-
0.00 0.02 0.04 0.06
0.3
0.4
0.5
0.6
0.7
Fra
ctio
n of
del
etio
ns
Superhelix density
Figure 7. Preference of deletion over inversion in a nicked circular plasmid containing two symmetric FRT sites.(a) Computed fraction of deletion products from recombination of symmetric FRT sites via parallel and antiparallelsynapses (broken and continuous lines, respectively) are plotted for different superhelical densities. Fluctuationsfrom the mean are shown by the error bars. The values obtained from the experimental data in (b) and (c) are rep-resented by the diamonds. Flp recombination was carried out on (b) the supercoiled form of pFRT-Sym or (c) its DNa-seI pre-nicked form. The control reactions without Flp were run in lanes 1. The designations P, I and D refer to bandscorresponding to parental plasmid, inversion product and the deletion products, respectively. The unmarked bandrunning between D1 and D2 was the result of intermolecular reaction.
522 Asymmetry in Integrase/Tyrosine Family Recombination

UNCORRECTED PROOF
bands were quantified from their ethidiumbromide fluorescence, corrected for their lengthdifference (approximately 3700 bp and 2800 bp)and used to calculate the molar ratio of deletion toinversion. The values for the supercoiled (Figure7(b)) and nicked (Figure 7(c)) substrates(s , 20:06 and 0, respectively) are indicated bythe diamond symbols in Figure 7(a).
Clearly, the observed recombination bias in thenicked plasmid matches that predicted from thesimulations for the antiparallel synapse. This con-cordance between experimental data and the simu-lation results provides another strong argument infavor of antiparallel synapsis.
Topological outcome of recombination atsymmetric loxP sites by the Cre recombinasefrom a 23 synapse arranged by the phage Mutransposition complex
Is the unique product topology obtained fromthe resolvase–Flp synapse a particularity of theresolvase system? Or, will other topologically
defined hybrid synapses yield predictable out-comes during integrase/tyrosine family recombi-nation? To tackle these issues, we chose the well-characterized Cre recombinase as the alternativeto Flp and a DNA synapse assembled by phageMu transposase (MuA) as the alternative to theresolvase synapse.
The plasmid substrate ploxP-Sym, analogous topFRT-Sym (see Figure 1) in its overall organization,contained two symmetrized loxP sites, the targetsfor Cre recombination (Figure 8). Instead of theres II/res III sites, the plasmid housed three appro-priately positioned DNA sites from the phage Mugenome: the left and right att sites (L and R,respectively) and the transposition enhancerelement (E).24 – 26 The att sites contained pointmutations that prevented DNA cleavage by theMuA protein, but still allowed the assembly ofthe LER complex (left end–enhancer–right end),the earliest intermediate characterized in the path-way of Mu transpososome organization.27 Byadapting the topological analyses described byGrainge et al.10 and Kilbride et al.12 to the Mu
CreMuA
HU
DnaseI
Del1
2
3
45
6
-
--- +
+ ++
++
+
+
+
+-- -
--
---
BamHI + EcoRV
OC
SC
L
1 2 3 4
- +1 2 3
MuA/Cre synapseploxP-Sym
A B C
P1
P2
I1
I2
D1
D2
ATAACTTCGTATAATGTATGCTATACGAAGTTAT
Wild type loxP site
Symmetrized core loxP site
TATTGAAGCATATTACATACGATATGCTTCAATA
ATAACTTCGTATAATGTACACTATACGAAGTTATTATTGAAGCATATTACATGTGATATGCTTCAATA
-
+++
Figure 8. Action of Cre on symmetric loxP sites following the assembly of the Mu-LER transposition synapse. TheDNA sequences of the native and symmetrized loxP sites are shown at the top. The core sequence is written in boldletters, and includes the inner 6 bp symmetrized strand-exchange region (spacer). The Cre-binding elements flank thecore in head-to-head orientation. (a) In plasmid ploxP-Sym, the three-crossing Mu-LER synapse and the adjacent loxPsites (two-headed arrows) are represented schematically. (b) Cre reactions carried out in the absence of the Mu-LERsynapse or following its assembly were analyzed by agarose gel electrophoresis after DNase I nicking. The deletioncircles are called Del1 (shown in lanes 2–4) and Del2, which is not seen here, since it migrated beyond the bottomedge of the gel. The symbols SC, OC and L refer to supercoiled, open circular and linear forms of the parent plasmidor its inversion product, respectively. The numbers 2, 3, 4, etc. refer to the complexity of the catenated or knottedrecombination products. (c) The linear DNA fragments I1 and I2 were formed from the inversion product of recombi-nation as a result of digestion with Bam HI and Eco RV. The corresponding fragments formed from the deletion circleswere D1 and D2. The digestion products from the parent plasmid were P1 and P2.
Asymmetry in Integrase/Tyrosine Family Recombination 523

UNCORRECTED PROOF
system, we determined that the Mu-LER synapsetraps three negative supercoil nodes as a result ofthe crossings between the enhancer and the twoatt sites (see Figure 8(a)).16 We could now askwhether Cre recombination at the symmetrizedloxP sites, occurring after the assembly of this 23synapse, was biased towards deletion or inversion,and whether the outcome of such a reaction wastopologically unique.
When the ploxP-Sym plasmid was treated withCre alone, both inversion and deletion resulted.Under the assay conditions employed, deletionwas the preferred reaction (see below and lane 2of Figure 8(c)). Unlike the Flp reaction (see Figure2), the Cre reaction yielded deletion circles thatwere almost exclusively unlinked, as had beenobserved previously.28 However, a trace of thetwo-noded catenane could be detected (lane 2,Figure 8(b)). The larger of the two deletion circles,Del 1, can be seen in lane 2 (Figure 8(b)), whereasthe smaller one, Del 2, overran the bottom edge ofthe gel during electrophoresis. Most of theinversion product, being unknotted, could not bedistinguished from the substrate by electrophoreticmobility; however, a faint band corresponding tothe trefoil was visible (lane 2, Figure 8(b)). Diges-tion of the recombination reaction with Bam HIand Eco RV revealed both the inversion (I1 and I2)and deletion (D1 and D2) products (compare lane2 with lane 1, Figure 8(c)) in addition to theparental plasmid (P1 and P2 in lanes 1–3, Figure8(c)). In ploxP-Sym, recombination was inherentlybiased towards deletion by a factor of approxi-mately 2.5 (compare intensities of D1 to I1 in lane2 of Figure 8(c)C).
Cre reaction carried out in the presence of MuAbut without the Escherichia coli HU protein (noLER synapse can be formed under this condition)increased the level of topological complexityamong the recombination products. Now, inaddition to the free deletion circles and the unknot,a ladder of knot bands (3, 5, 7, etc.) and catenanebands (2, 4, 6, etc.) became apparent (lane 3, Figure8(b)). When Cre recombination was performedafter assembly of the LER complex, the single pro-duct species that was most strongly enriched wasthe three-noded knot (lane 4, Figure 8(b)). Theslight increase observed in the two-noded catenaneand the four-noded product can be explained asdue partly to an overall stimulation of Cre recom-bination in the presence of MuA and HU, andpartly to a second round of Cre recombinationoccurring on the initial inversion product. Notethat there was a detectable enhancement of theunlinked deletion circle (Del1) in the Cre reactioncontaining MuA and HU (lane 4, Figure 8(b)) com-pared to the Cre alone reaction (lane 2, Figure 8(b)).The Bam HI plus Eco RV digestion pattern of theMuA and HU-assisted Cre reaction showed astrong preferential increase in inversion (approxi-mately four- to fivefold) relative to the unassistedreaction (compare lane 3 with lane 2, Figure 8(c)).This large shift away from the intrinsic recombina-
tion bias of the Cre alone reaction and the specificenrichment of the three-noded knot are consistentwith the symmetrized loxP sites being treated asan inversion substrate (and not a deletion sub-strate) within the Mu-LER/Cre hybrid synapse.
The complexity of the Cre recombination pro-ducts in the presence of MuA but without HU isinteresting. MuA bound to the att and enhancerDNA may increase the level of random entrapmentof supercoils during the Cre-mediated synapsis ofthe loxP sites. Alternatively, in the absence of HU,a series of topologically distinct but non-functionaltransposition synapses may be formed by randomcollision. Products of subsequent Cre recombina-tion would then reflect this topological diversity.The symmetrized loxP sites, unlike their FRTcounterparts, favor deletion to inversion in theplasmid substrate used in our assays. We do notknow whether the asymmetry in the base-pairsimmediately flanking the strand-exchange regionis responsible for this preferred directionality inrecombination. However, none of our interpret-ations is affected by this bias.
Based on the sum of the topological data, weconclude that Flp and Cre (and, by extension, allmembers of the integrase/tyrosine family) assigna uniform sense of directionality to the target sites,even the symmetric ones, within their synapses.The imposed directionality conforms to antiparal-lel geometry that allows recombination to proceedwithout adding DNA nodes.
Discussion
Using distinct functional recombination assayswith symmetrized as well as sterically constrainedsubstrates, we have determined the geometric dis-position of the two chemically competent recombi-nase monomers within Flp and Cre synapses. Theactive monomers within the recombinase tetramerare diagonally related across the synapse. Thispairwise activation/inactivation at each strandcleavage and exchange step occurs on sym-metrized as well as native recombination sites.Thus, the allosteric control of cleavage is innate tothe protein and occurs even in the absence of adirectionality signal from the DNA sequence.
The integrase/tyrosine family recombinationsynapse: the local geometry of target DNAsites is independent of their globalrelative orientation
The action of Flp/resolvase and Cre/Mu-LERsystems on symmetrized DNA substrates demon-strates that the topology of the recombination pro-ducts is determined solely by the number of DNAcrossings sequestered outside the Flp or Cresynapse. The act of recombination does not add toproduct topology, as would be consistent with anantiparallel synapse.10 These conclusions are sup-ported by the dilution and competition assays
524 Asymmetry in Integrase/Tyrosine Family Recombination

UNCORRECTED PROOF
using tethered antiparallel or parallel FRT sites.They are consistent with the recombination out-comes from a nicked circular plasmid containingsymmetrized FRT sites. When an odd number ofinterdomainal nodes are trapped outside the Flpor Cre synapse, recombination in symmetrizedsites would be channeled towards DNA inversion.When the number of such nodes is even, thereaction would be channeled towards deletion.
The present experimental outcomes are con-sistent with the accepted interpretations of theX-ray structures of Flp–DNA and Cre DNAcomplexes.9,11 However, the specter of a recombina-tion-competent, but parallel orientation of targetsites raised by analyses of FRT and l Int synapsesusing electron microscopy, atomic forcemicroscopy and DNA ligation assays14,15 is causefor some concern that crystal packing forces mighthave selected for the antiparallel synaptic configur-ation and excluded the alternative. The publishedFlp structure does not fully represent the nativesynapse. Because of some rather unexpected reac-tions, all four scissile phosphate positions in thesynapse have been cleaved, and the spacer lengthhas been reduced from the normal 8 bp to 7 bp.9
Assignment of the two “active” and “inactive” Flpmonomers in the structure is possible only byusing the characteristic active-site arrangements inthe Cre structures as a template. The combinedresults from the present study leave little room forany significant contribution from parallel synapseto integrase/tyrosine family recombination. Dis-crepancies in published data may be at leastpartially reconciled by the existence of multiplestates in the pathway to functional synapsis, someof which may have been preferentially sampled ina given protocol.
Although the recombinase protein is the majordeterminant of asymmetry, the DNA substratemay likely facilitate its establishment. In the caseof native recombination sites, preferred bendlocations within the spacer can potentially contri-bute towards asymmetry. At least for some of theintegrase/tyrosine family members, this featuremay dictate the order in which strands are cut andexchanged during recombination. Recently, Lee &Sadowski29 have shown that Cre resolves syntheticHolliday junctions containing the wild-type spacersequence with a particular bias, which may beabolished by symmetrizing the spacer.
Materials and Methods
Synthetic oligonucleotides
Deoxyoligonucleotides (from Integrated DNA tech-nologies, Coralville, IA) were gel-purified, end-labeledand hybridized as described.30
In constructing tethered substrates, the longest strandwas synthesized as two separate oligonucleotides. Theywere ligated with bacteriophage T4 DNA ligase in thepresence of a short junction-complementary splinter
oligonucleotide, purified by denaturing gel electro-phoresis and used for substrate assembly.
Plasmid substrates
The plasmid containing the res II–res III accessory sitesplus the FRT sites with the symmetrized spacer was con-structed using strategies described by Grainge et al.10
The construction of plasmids for carrying out Cre-loxrecombination after the assembly of a transpositionsynapse has been described.16 These plasmids containedthe attL and attR DNA sites from phage Mu in head-to-head orientation along with the transposition enhancersequence included between them. The symmetrizedloxP sites were assembled from synthetic oligonucleo-tides and were inserted on either side of the enhancer.Full details of plasmid construction and plasmidsequences are available from the authors upon request.
Proteins
Flp protein, obtained by affinity chromatography,31
was .95% pure. Cre protein, prepared essentiallyaccording to Abremski et al.,32 was approximately 50%pure. Roughly 85% pure Tn3 resolvase was prepared asdescribed.10 The MuA protein and E. coli HU proteinswere the same as those used by Jiang & Harshey.33
Protein concentrations for the homogeneouspreparations were determined from the correspondingmolar extinction coefficients at 280 nm. For the less purepreparations, aliquots from several dilutions were frac-tionated by SDS-PAGE, stained with Coomassie brilliantblue and quantified by densitometry. Protein amountswere derived from a standard curve prepared similarlywith known amounts of pure bovine serum albumin.
Reactions with resolvase and Flp
Reactions were carried out as described.10 The sub-strate DNA was pre-incubated with Tn3 resolvase forten minutes at 37 8C. The reaction mixtures were trans-ferred to 30 8C, and Flp was added. Incubation was con-tinued at 30 8C for eight minutes. Reactions werestopped by heating at 70 8C for ten minutes.
The incubation time for the Flp assays in the dilutionand competition experiments in Figures 5 and 6 was 20minutes.
Reactions with MuA and Cre
The Mu-LER complex was assembled by incubatingthe substrate with MuA and the E. coli HU proteinunder standard conditions.27 Cre protein was added tothe reaction mixtures, and they were incubated for anadditional 20 minutes. Reactions were terminated byheat inactivation at 70 8C for ten minutes. The Cre alonereactions were carried out in the same buffer (pH 7.6) asthat used to organize the Mu-LER complex.
DNase I nicking and agarose gel electrophoresis
Reactions were nicked by DNase I to remove super-coils, and analyzed by electrophoresis as described.10
Asymmetry in Integrase/Tyrosine Family Recombination 525

UNCORRECTED PROOF
Modeling DNA and simulation procedures
Circular DNA was modeled as a discrete worm-likechain consisting of N rigid cylinders of equal length l,as described.34 The values of the model parameters corre-sponded to a solution of 0.2 M NaCl: DNA persistencelength of 50 nm, torsional rigidity of 3.0 £ 10219 erg cm,and DNA effective diameter of 5 nm.34 The length of astraight segment was equal to 10 nm, so each cylinderof the chain (see Figure 7) corresponded to 30 bp. TwoFRT sites were located at specific positions of the modelchain. The chain length corresponded to 4200 bp, andthe sites were separated by 1440 bp. Each site wasmodeled by two adjacent segments. The circular sub-strate used during the experimental test of the simu-lations was approximately 4200 bp long, and thespacing between the FRT sites was approximately1500 bp. We assumed that each half of an FRT site wasbound by a Flp monomer, and therefore the sites werebent in these complexes.35 The included angle betweenthe two segments modeling an FRT site was set at 908,9
and did not fluctuate during the simulation. The siteswere assumed to be symmetric in this simulation. TheMetropolis procedure was used to simulate the equi-librium ensemble of DNA conformations.34 The numberof chain conformations with the juxtaposed FRT sites inthe mutual orientation that is conducive torecombination9 was quite low in the equilibriumsampling. To circumvent the poor yield, we used abiased sampling procedure, known as the umbrellamethod.36 This procedure is based on adding an artificialpotential to the system to enrich the sampling of desiredstates. Although the potential disturbs the whole distri-bution, it does not affect the conformational distributionamong the states with juxtaposed sites.
We used the following potential:
U ¼ Ar
2q0
r2q2
2rq0
rq0
!exp 2
w2 þ u2 þ z2
2s2
� �; ð1Þ
where r, q, u and z are variables whose definitions areexplained in the legend to Figure 6. The values for theparameters A, q, r0 and s in the computations were: A ¼20kT; q ¼ 1:6; r0 ¼ 0:6l and s ¼ 1; where kT is theBoltzmann temperature factor.
Acknowledgments
The studies described here were supported by grantsfrom the National Institutes of Health to M.J.(GM35654), R.M.H. (GM33247) and A.V. (GM54215).The Robert F. Welch Foundation provided partial sup-port through grants F-1274 to M.J. and F-1531 to R.M.H.
References
1. Grainge, I. & Jayaram, M. (1999). The integrasefamily of recombinases: organization and functionof the active site. Mol. Microbiol. 33, 449–456.
2. Jayaram, M., Grainge, I. & Tribble, G. (2002). Site-specific recombination by the Flp protein ofSaccharomyces cerevisiae. In Mobile DNA II (Craig,N. L., ed.), pp. 192–218, ASM Press, Washington,DC.
3. Van Duyne, G. D. (2002). A structural view of cre-loxsite-specific recombination. Annu. Rev. Biophys.Biomol. Struct. 30, 87–104.
4. Landy, A. (1999). Coming or going: it’s anotherpretty picture for the lambda-Int family album. Proc.Natl Acad. Sci. USA, 96, 7122–7124.
5. Broach, J. R. & Volkert, F. (1991). Circular DNA plas-mids of yeast. In The Molecular and Cellular Biologyof the Yeast Saccharomyces (Broach, J. R., Pringle, J. R.& Jones, E. W., eds), vol. 1, pp. 297–332, Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
6. Austin, S., Ziese, M. & Sternberg, N. (1981). A novelrole for site-specific recombination in the mainten-ance of bacterial replicons. Cell, 25, 729–736.
7. Senecoff, J. F. & Cox, M. M. (1986). Directionality inFLP protein-promoted site-specific recombination ismediated by DNA–DNA pairing. J. Biol. Chem. 261,7380–7386.
8. Hoess, R. H., Wierzbicki, A. & Abremski, A. (1986).The role of the loxP spacer region in P1 site-specificrecombination. Nucl. Acids Res. 14, 2287–2300.
9. Chen, Y., Narendra, U., Iype, L. E., Cox, M. M. &Rice, P. A. (2000). Crystal structure of a Flp recombi-nase–Holliday junction complex. Assembly of anactive oligomer by helix swapping. Mol. Cell, 6,885–897.
10. Grainge, I., Buck, D. & Jayaram, M. (2000). Geometryof site alignment during Int family recombination:antiparallel synapsis by the Flp recombinase. J. Mol.Biol. 298, 749–764.
11. Guo, F., Gopaul, D. N. & Van Duyne, G. D. (1997).Structure of Cre recombinase complexed with DNAin a site-specific recombinase synapse. Nature, 389,40–46.
12. Kilbride, E., Boocock, M. R. & Stark, W. M. (1999).Topological selectivity of a hybrid site-specificrecombination system with elements from Tn3 res/resolvase and bacteriophage P1 loxP/Cre. J. Mol.Biol. 289, 1219–1230.
13. Lee, J., Voziyanov, Y., Pathania, S. & Jayaram, M.(1998). Structural alterations and conformationaldynamics in Holliday junctions induced by bindingof a site-specific recombinase. Mol. Cell, 1, 483–493.
14. Cassell, G., Moision, R., Rabani, E. & Segall, A.(1999). The geometry of a synaptic intermediate in apathway of bacteriophage lambda site-specificrecombination. Nucl. Acids Res. 27, 1145–1151.
15. Huffman, K. E. & Levene, S. D. (1999). DNA-sequence asymmetry directs the alignment of recom-biantion sites in the Flp synaptic complex. J. Mol.Biol. 286, 1–13.
16. Pathania, S., Jayaram, M. & Harshey, R. M. (2002).Path of DNA within the Mu transpososome: transpo-sase interactions with two Mu ends and the enhancertrap five DNA supercoils. Cell, 109, 425–436.
17. Grindley, N. D. F. (1994). Resolvase-mediated site-specific recombination. In Nucleic Acids and Mol-ecular Biology (Eckstein, F. & Lilley, D. M. J., eds),vol. 8, pp. 236–267, Springer, Berlin.
18. Wasserman, S. A. & Cozzarelli, N. R. (1985). Determi-nation of the stereostructure of the product of Tn3resolvase by a general method. Proc. Natl Acad. Sci.USA, 82, 1079–1083.
19. Wasserman, S. A., Dungan, J. M. & Cozzarelli, N. R.(1985). Discovery of a predicted knot substantiates amodel for site-specific recombination. Science, 229,171–174.
20. Stark, W. M., Sherratt, D. J. & Boocock, M. R. (1989).Site-specific recombination by Tn3 resolvase:
526 Asymmetry in Integrase/Tyrosine Family Recombination

UNCORRECTED PROOF
topological changes in the forward and reverse reac-tions. Cell, 58, 779–790.
21. Murley, L. L. & Grindley, N. D. F. (1998). Architectureof the gd resolvase synaptasome: oriented hetero-dimers identify interactions essential for synapsisand recombiantion. Cell, 95, 553–562.
22. Sarkis, G. J., Murley, L. L., Leschziner, A. E., Boocock,M. R., Stark, W. M. & Grindley, N. D. F. (2001). Amodel for the gd resolvase synaptic complex. Mol.Cell, 6, 623–631.
23. Kimball, A., Guo, Q., Lu, M., Cunningham, R. P.,Kallenbach, N. R. & Tullius, T. D. (1990). Construc-tion and analysis of parallel and antiparallelHolliday junctions. J. Biol. Chem. 265, 6544–6547.
24. Chaconas, G. & Harshey, R. M. (2002). Transpositionof phage Mu DNA. Mobile DNA II (Craig, N. L.,et al.), ASM Press, Washington, DC in the press.
25. Mizuuchi, M., Baker, T. A. & Mizuuchi, K. (1992).Assembly of the active form of the transposase–MuDNA complex: a critical control point in Mu transpo-sition. Cell, 70, 384–402.
26. Mizuuchi, K. (1992). Transpositional recombination:mechanistic insights from studies of Mu and otherelements. Annu. Rev. Biochem. 61, 1011–1051.
27. Watson, M. A. & Chaconas, G. (1996). Three-sitesynapsis during Mu DNA transposition: a criticalintermediate preceding engagement of the activesite. Cell, 85, 435–445.
28. Abremski, K., Hoess, R. & Sternberg, N. (1983).Studies on the properties of P1 site-specific recombi-nation: evidence for topologically unlinked productsfollowing recombination. Cell, 32, 1301–1311.
29. Lee, L. & Sadowski, P. D. (2001). Directional resol-ution of synthetic Holliday structures by the Crerecombinase. J. Biol. Chem. 276, 1092–1098.
30. Lee, J., Whang, I. & Jayaram, M. (1996). Assemblyand orientation of Flp recombinase active sites ontwo-, three-, and four-armed DNA substrates: impli-cations for a recombination mechanism. J. Mol. Biol.257, 532–549.
31. Lee, J. & Jayaram, M. (1995). Functional roles of indi-vidual recombinase monomers in strand breakageand strand union during site-specific DNA recombi-nation. J. Biol. Chem. 270, 23203–23211.
32. Abremski, K. & Hoess, R. (1984). Bacteriophage P1site-specific recombination. Purification and proper-ties of the Cre recombinase protein. J. Biol. Chem.259, 1509–1514.
33. Jiyang, H. & Harshey, R. M. (1999). Criss-crossedinteractions between the enhancer and the att sitesof phage Mu during DNA transposition. EMBO J.18, 3845–3855.
34. Vologodskii, A. V., Levene, S. D., Klenin, K. V., Frank-Kamenetskii, M. & Cozzarelli, N. R. (1992). Confor-mational and thermodynamic properties of super-coiled DNA. J. Mol. Biol. 227, 1224–1243.
35. Ringrose, L., Lounnas, V., Ehrlich, L., Buchholz, F.,Wade, R. & Stewart, A. F. (1998). Comparative kineticanalysis of FLP and Cre recombinases: mathematicalmodels for DNA binding and recombination. J. Mol.Biol. 284, 363–384.
36. McCammon, J. A. & Harvery, S. C. (1987). Dynamicsof Proteins and Nucleic Acids, Cambridge UniversityPress, New York.
Edited by M. Belfort
(Received 21 February 2002; received in revised form 8 May 2002; accepted 13 May 2002)
Asymmetry in Integrase/Tyrosine Family Recombination 527




















