
SYAHRUNI HIDAYATULLAH - perpustakaan universitas ...
163
POLA MUTASI GEN MYELOID DIFFERENTIATION FACTOR 88 DAN KADAR TNF-α PADA PENDERITA TUBERKULOSIS PARU Mutation Patterns of Myeloid Differentiation Factor 88 gene and Level TNF-α in the Pulmonary Tuberculosis Patients SYAHRUNI HIDAYATULLAH PROGRAM PASCASARJANA UNIVERSITAS HASANUDDIN MAKASSAR 2013
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of SYAHRUNI HIDAYATULLAH - perpustakaan universitas ...

POLA MUTASI GEN
MYELOID DIFFERENTIATION FACTOR 88
DAN KADAR TNF-α PADA PENDERITA
TUBERKULOSIS PARU
Mutation Patterns of Myeloid Differentiation Factor 88 gene and
Level TNF-α in the Pulmonary Tuberculosis Patients
SYAHRUNI HIDAYATULLAH
PROGRAM PASCASARJANA UNIVERSITAS HASANUDDIN
MAKASSAR 2013

POLA MUTASI GEN
MYELOID DIFFERENTIATION FACTOR 88
DAN KADAR TNF-α PADA PENDERITA
TUBERKULOSIS PARU
Tesis
Sebagai Salah Satu Syarat untuk Mencapai Gelar Magister
Program Studi
Biomedik
Disusun dan diajukan oleh
SYAHRUNI HIDAYATULLAH
kepada
PROGRAM PASCASARJANA UNIVERSITAS HASANUDDIN
MAKASSAR 2013


PERNYATAAN KEASLIAN TESIS
Yang bertanda tangan dibawah ini
Nama : Syahruni Hidayatullah
Nomor Mahasiswa : P1506211005
Program Studi : Biomedik
Menyatakan dengan sebenarnya bahwa tesis yang saya tulis ini
benar-benar merupakan hasil karya saya sendiri, bukan merupakan
pengambilalihan tulisan atau pemikiran orang lain. Apabila di kemudian
hari terbukti atau dapat dibuktikan bahwa sebagian atau keseluruhan tesis
ini hasil karya orang lain, saya bersedia menerima sanksi atas perbuatan
tersebut.
Makassar, 23 Juli 2013
Yang menyatakan
SYAHRUNI HIDAYATULLAH

v
PRAKATA
Puji Syukur penulis panjatkan kehadirat Allah SWT, yang telah
memberikan rahmat, hidayat dan karunia-Nya, sehingga penulis dapat
menyelesaikan tesis ini dengan judul “Pola mutasi Gen Myeloid
Differentiation Factor 88 dan kadar TNF-α pada Penderita Tuberkulosis
Paru ”.
Penulis menyadari sepenuhnya bahwa tesis ini masih jauh dari
kesempurnaan. Oleh karena itu, penulis sangat mengharapkan saran dan
kritik dari semua pihak yang membacanya. Selain itu dalam
menyelesaikan tesis ini, penulis menyadari bahwa hal tersebut tidak
terlepas dari bantuan berbagai pihak baik secara moril maupun secara
materil. Dalam kesempatan ini penulis dengan tulus menyampaikan terima
kasih kepada :
1. Prof. dr. Muh. Nasrum Massi, Ph.D selaku ketua komisi penasehat
dan dr. Uleng Bahrun, Sp.PK (K), Ph.D selaku sekretaris komisi
penasehat atas bantuan dan bimbingannya yang telah diberikan mulai
dari pengembangan minat terhadap permasalahan penelitian ini,
pelaksanaan penelitian sampai pada tahap penulisan tesis ini.
2. Kepada Ketua Program Studi Ilmu Biomedik Prof. dr. Rosdiana
Natsir, Ph.D
3. Kepada seluruh staf dosen Biomedik Konsentrasi Mikrobiologi
Prof. Ahyar Ahmad, Ph.D., Prof. DR. dr. Asaad Maidin, M.Sc, Sp.MK,

vi
dan dr. Rizalinda Sjahril, M.Sc. Ph.D, Prof dr. M. Hatta, Sp.MK,
Prof. drh. Lucia Muslimin, M.Sc, DR. dr. Armyn Nurdin, M.Sc,
DR. dr. Burhanuddin Bahar, M.Kes, yang telah membagikan ilmu
semasa kuliah hingga penulis menyusun dan menyelesaikan tesis ini.
4. Teman – teman seperjuangan di Mikrobiologi angkatan 2011, kakak
Syam Kumaji, Kakak Norma Kambuno, Kakak A. Salsa Anggraini,
kakak Anita, kakak Arniati, kakak Aslim taslim, kakak A.M. Nawir,
Hartati, Alhawaris, Fardi, kakak Sulfiah dan Andini Umiati.
5. Senior – senior di Unit Penelitian Universitas Hasanuddin dan
Laboratorium NHCR (K. Uli, K. Risma, K. Ondo, K. Dina dan Erpi, Zul,
K. Safri) yang selalu mendukung dan membantu saya menyelesaikan
penelitian ini.
Penulis persembahkan tesis ini kepada semua keluarga di Belawa-
Wajo, teristimewa buat Ayahanda dan Ibundaku Drs. Selle Diri dan
Hj. Nurhawati, S.Pd, saudaraku M.Syarief Hidayatullah, SE dan
Suryadi, SE untuk semua doa, masukan, perhatian, bimbingan dan
dukungan sehingga penulis dapat menyelesaikan pendidikan Magister
Kesehatan dalam bidang keahlian Mikrobiologi ini.
Makassar, 23 Juli 2013
Syahruni Hidayatullah

vii
ABSTRAK
SYAHRUNI HIDAYATULLAH. Pola Mutasi Gen Myeloid Differentiation
Factor 88 dan Kadar TNF-α pada Penderita Tuberkulosis Paru. (dibimbing
oleh Muh. Nasrum Massi dan Uleng Bahrun)
Penelitian ini bertujuan untuk menemukan pola mutasi pada gen
Myeloid Differentiation Factor 88 dan kadar TNF-α pada penderita
tuberkulosis paru.
Telah dilakukan penelitian terhadap empat orang penderita
tuberkulosis paru yang berasal dari Rumah Sakit Umum Labuang Baji dan
BBKPM. Metode yang digunakan adalah Metode PCR untuk
mengamplifikasi gen, DNA sekuensing untuk menentukan urutan basa
nukleotida dan ELISA untuk menghitung kadar TNF-α .
Hasil penelitian menunjukkan terdapat 31 titik mutasi yang tersebar
pada 3 daerah yaitu daerah death domain sebanyak 14 titik mutasi,
daerah Intermediated domain sebanyak 3 titik mutasi dan daerah TIR
domain sebanyak 14 titik mutasi. Selain itu ditemukan kadar TNF-α pada
semua sampel berada dibawah cut off. Makna pola substitusi asam amino
dan kadar TNF-α yang diperoleh belum bisa disimpulkan memiliki
keterkaitan dengan respon imun tubuh yang dihasilkan terhadap infeksi
Mycobacterium tuberculosis sehingga perlu dilakukan penelitian lebih
lanjut.
ABSTRACT

viii
SYAHRUNI HIDAYATULLAH. Mutation Patterns of Myeloid Differentiation
Factor 88 gene and TNF-α Level in the Pulmonary Tuberculosis Patients.
(Supervised by Muh. Nasrum Massi and Uleng Bahrun)
This study aims to detect mutation patterns of Myeloid
Differentiation Factor 88 gene and TNF-α levels in patients with pulmonary
tuberculosis.
Has conducted a study of four patients with pulmonary tuberculosis
from Labuang Baji Hospital and BBKPM . The method in this study were
PCR method to amplification of the target gene, DNA sequencing to
determine the nucleotide sequences and ELISA to value the levels of
TNF-α
The results showed there were 31 patterns of mutations in three
regions, death domain region found of 14 patterns of mutations,
intermediated domain region found of 3 patterns of mutations and TIR
domain region found of 14 patterns of mutations. Also found TNF-α levels
in all samples were below the cut-off. However, the meaning of the
variants of the patterns mutations and TNF-α levels found in this study
was still unclear. Therefore, further research should be conducted to find
out the meaning of them.

ix
DAFTAR ISI
Halaman
PRAKATA v
ABSTRAK vii
ABSTRACT viii
DAFTAR ISI ix
DAFTAR TABEL xiii
DAFTAR GAMBAR xiv
DAFTAR LAMPIRAN xv
DAFTAR ARTI LAMBANG DAN SINGKATAN xvi
I. PENDAHULUAN 1
A. Latar Belakang 1
B. Rumusan Masalah 5
C. Tujuan Penelitian 5
D. Manfaat Penelitian 6
II. TINJAUAN PUSTAKA 7
A. Tinjauan Khusus Tuberkulosis 7
1. Sejarah, Definisi, dan Karakteristik Mycobacterium 7 Tuberculosis
2. Klasifikasi Mycobacterium tuberculosis 11

x
3. Epidemiologi Penyakit Tuberkulosis 11
4. Klasifikasi Penyakit Tuberkulosis 13
5. Patogenesis 17
6. Reaksi Imunologik pada Infeksi Tuberkulosis 24
7. Gejala dan Tanda Penyakit 29
B. Sistem Imun Alamiah (Innate Immunity) Terkait 31 Mycobacterium tuberculosis
1. Toll Like Receptors (TLRs) 34
2. NOD Like Receptor (NLRs) 37
3. C-Type Lectins 39
4. DC-SIGN 39
5. DECTIN-1 40
C. Tinjauan Khusus Myeloid Differentiation Factor 88 41
D. Tinjauan Khusus Molekuler 44
1. Penemuan Awal 44
2. Struktur DNA 45
3. Teknik Rekayasa Genetik 47
a. Polymerase Chain Reaction (PCR) 48
b. Teknik DNA Sequencing 52
c. Elektroforesis 56
4. Kode Genetik 58
5. Mutasi 61
E. Tinjauan Umum Sitokin 69
1. Definisi Sitokin 69

xi
2. TNF-α 72
F. Tinjauan Umum ELISA 75
1. Direct ELISA 76
2. Indirect ELISA 77
3. Sandwich ELISA 77
4. Capture ELISA 78
5. Sel ELISA 79
G. Kerangka Konsep 79
H. Definisi Operasional 86
III. METODOLOGI PENELITIAN 88
A. Rancangan Awal 88
B. Waktu dan Tempat Penelitian 88
C. Populasi, Besaran Sampel Penelitian 88
1. Populasi Penelitian 88
2. Sampel Penelitian 88
3. Kriteria Sampel 89
D. Alat dan Bahan Penelitian 90
1. Alat Penelitian 90
2. Bahan Penelitian 90
E. Cara Kerja 91
1. Alokasi Subjek 91
2. Cara Penelitian 91

xii
3. Prosedur Kerja 92
a. Pewarnaan Ziehl Nelseen 92
b. Extraksi DNA 93
c. Amplifikasi DNA 94
d. Elektroforesis gel agarosa 95
e. Purifikasi gel extraksi 96
f. Pemeriksaan sequencing 98
g. Pemeriksaan ELISA 98
F. Pertimbangan Etis 99
G. Analisa Data 100
H. Skema Alur Penelitian 101
IV. HASIL DAN PEMBAHASAN 102
A. Hasil Penelitian 102
B. Pembahasan 108
V. KESIMPULAN DAN SARAN 115
A. Kesimpulan 115
B. Saran 115
DAFTAR PUSTAKA 117
LAMPIRAN 121

xiii
DAFTAR TABEL
Halaman
Tabel 1. Angka Insiden, Prevalensi, dan Kematian TB di Indonesia 13
Tabel 2. Kode Genetik “Universal” 59
Tabel 3. Beberapa Penyimpanan dari Kode Genetik “universal” 61
Tabel 4. Kelompok Sitokin dalam Keluarga 70
Tabel 5. Gambaran Sitokin Imunitas Bawaan dan Dapatan 72
Tabel 6. Karakteristik Sampel 102
Tabel 7. Pola substitusi asam amino region Death domain 104
Tabel 8. Pola substitusi asam amino region Intermediate domain 105
Tabel 9. Pola substitusi asam amino region TIR domain 106
Tabel 10. Substitusi asam amino dengan perubahan sifat 107

xiv
DAFTAR GAMBAR
Halaman
Gambar 1. Bentuk Mycobacterium tuberculosis dengan 8
Perwarnaan Ziehl Neelsen
Gambar 2. Struktur dinding Sel Mycobacterium tuberculosis 9
Gambar 3. Patogenesis Tuberkulosis 24
Gambar 4. Susunan dinding sel Mycobacterium tuberculosis 32
Gambar 5. Pola pengenalan reseptor Mycobacterium tuberculosis 34
dan Downstream signaling pathway
Gambar 6. Jenis PPR selain TLR yang berfungsi dalam pengenalan komponen mikobakteria
Gambar 7. Mapping gen MyD88 42
Gambar 8. Posisi Pembagian daerah domain MyD88 dan 43
Gambar 9. Struktur Kromosom, Gen dan DNA 45
Gambar 10. Struktur pita DNA ganda 46
Gambar 10. Perbedaan Struktur molekul dNTP dan ddNTP 54
Gambar 11. Mutasi gen 68
Gambar 12. Mutasi Kromosom 69
Gambar 13. Direct ELISA 76
Gambar 14. Indirect ELISA 77
Gambar 15. Sandwich ELISA 78
Gambar 16. kerangka Konsep 85
Gambar 17. Kerangka teori 87
Gambar 18. Pemetaan Posisi Substitusi Asam Amino berdasarkan region 103
Gambar 19. Kadar TNF-α pada sampel penderita tuberkulosis 108

xv
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Surat Persetujuan Komisi Etik 122
Lampiran 2. Contoh informed concent 123
Lampiran 3. Sekuens primer 124
Lampiran 4. Data GeneBank 125
Lampiran 5. Hasil Elektroforesis 131
Lampiran 6. Hasil elektroferogram 132
Lampiran 7. Hasil Analisis Bioedit sekuensing exon 1 138
Lampiran 8. Hasil Analisis Bioedit sekuensing exon 2 138
Lampiran 9. Hasil Analisis Bioedit sekuensing exon 3 139
Lampiran 10. Hasil Analisis Bioedit sekuensing exon 4 139
Lampiran 11. Hasil Analisis Bioedit sekuensing exon 5 139
Lampiran 12. Hasil Analisis Bioedit asam amino exon 1 140
Lampiran 13. Hasil Analisis Bioedit asam amino exon 2 140
Lampiran 14. Hasil Analisis Bioedit asam amino exon 3 140
Lampiran 15. Hasil Analisis Bioedit asam amino exon 4 140
Lampiran 16. Hasil Analisis Bioedit asam amino exon 5 140
Lampiran 17. Skema kerja pewarnaan Ziehl Neelsen 141
Lampiran 18. Skema kerja Ekstraksi DNA 142
Lampiran 19. Skema kerja PCR 143
Lampiran 20. Skema kerja ELISA 144
Lampiran 21. Physicochemical Properties Of Amino Acid 145

xvi
DAFTAR ARTI LAMBANG DAN SINGKATAN
Lambang/singkatan Arti dan keterangan
TB Tuberkulosis
HIV Human Immunodeficiency Virus
BTA Basil Tahan Asam
WHO World Health Organization
TLRs Toll-Like Receptors
APC Antigen Presenting cell
Th1 T helper I
TNF-α Tumor Necrosis Factor-α
IL-6 Interleukin-6
PRR Pattern Recognition Receptor
LPS Lipopolisakarida
LTA Asam Lipotechoic
MyD88 Myeloid Differentiation Factor 88
TIRAP TIR domain-containing adaptor protein
TRIF TIR domain-containing adaptor inducing IFN
TRAM Toll receptor-associated Molekul
SPS Sewaktu – Pagi – Sewaktu

xvii
OAT Obat Anti Tuberkulosis
PAMPs Pathogen-Associated Molecular Pattern
NF-kB Necrosis Factor- Kappa Beta
kDa Kilo Dalton
BCG Bacillus-Calmette-Guerin
LAM Lipoarabinomanna
LM Lipomannan
PIM Fosfatidil-myoinositol mannoside
IRF Interferon regulated factor
VDR Vitamim D receptor
SLPI Secretory leukocyte protese inhibitor
Fe2+ Besi
NLRs NOD like receptor
CARD Caspase activation and recruitment domain
T Timin
A Adenin
G Guanin
C Cytosine
DNA Deoxyribonucleic acid

xviii
PCR Polymerase Chain Reaction
dNTP Deoksi ribonukleotida trifosfat
AA Asam Amino

1
BAB I
PENDAHULUAN
A. Latar Belakang Penelitian
Tuberkulosis (TB) adalah penyakit infeksi yang bersifat kronik dan
menular langsung yang disebabkan oleh Mycobacterium tuberculosis
(Hardjoeno dkk, 2007). Mycobacterium tuberculosis dapat menyerang
organ paru-paru (pulmonal TB) dan menyerang organ lain (extrapulmonal
TB) (World Health Organization, 2012). Mycobacterium tuberculosis
menyebar melalui udara (droplet nuclei) saat seorang pasien TB batuk
kemudian percikan ludah yang mengandung bakteri tersebut terhirup oleh
orang lain yang sehat. Masa inkubasinya selama 3-6 bulan (Widoyono,
2008, Jawetz , 2006).
Tuberkulosis (TB) merupakan penyakit infeksi penyebab kematian
manusia terbesar di dunia. Global Report World Health Organization
(WHO) melaporkan bahwa pada tahun 2011 terdapat 9 juta kasus TB baru
di dunia atau sekitar 125 kasus per 100.000 populasi dan 1.4 juta
meninggal (990.000 HIV-negatif dan 430.000 TB-HIV). Diperkirakan
2.621.049 juta merupakan BTA positif (World Health Organization, 2012).
Berdasarkan laporan WHO 2012, Indonesia (0.4 – 0.5 jt) termasuk
dalam 5 negara dengan tingkat insiden tertinggi di dunia dan menduduki
peringkat ke-4 setelah India (2 – 2.5 jt), China (0.9 - 1.1 jt), dan Afrika

2
Selatan (0.4 – 0.6 jt). Di Indonesia dilaporkan pada tahun 2010 terdapat
690.000 kasus TB (289 per 100.000 populasi) dengan 197.797 BTA
positif, 101.750 BTA negatif, 14.054 TB Extrapulmonal, 5.348 kasus
kambuh, dan 318.949 kasus baru (World Health Organization, 2012).
Berdasarkan profil data kesehatan Indonesia tahun 2011, jumlah
penderita TB di Sulawesi Selatan sebanyak 10.970 orang, dimana 8.860
adalah kasus TB BTA positif dengan prevalensi kejadian terbanyak pada
laki-laki (6.352 orang) dibandingkan perempuan (4.618 orang)
(Kementerian Kesehatan Republik Indonesia, 2011).
Seiring dengan perkembangan zaman dan kemajuan teknologi
berbagai upaya telah dilakukan untuk menanggulangi dan menekan laju
pertumbuhan penyakit tuberkulosis paru, mulai dari penemuan jenis terapi
yang baru, metode yang lebih tepat guna sampai berbagai upaya yang
dilakukan para peneliti untuk mempelajari dan mengetahui mekanisme
mulai dari kuman penyebabnya hingga bagaimana mekanisme dari
respon imun hostnya sendiri guna memusnahkan penyakit maupun
menemukan produk yang dapat mencegah seseorang terinfeksi (vaksin
baru). (Nugraha. J, 2012).
Jika ditinjau dari segi hostnya, terdapat suatu mekanisme tubuh
yang dapat berperan dalam melawan kuman M.tuberculosis yang dikenal
degan istilah sistem imun. Kedua sistem imun, baik alami (Innate
immunity) maupun adaptif (adaptive immunity) berperan untuk melawan
kuman M.tuberculosis masuk kedalam tubuh. Sistem imun alami akan

3
mengenali ligan yang terdapat di M. tuberculosis oleh reseptor
pengenalan yang biasa dikenal sebagai Toll-like receptors (TLRs). TLRs
akan merangsang ekspresi gen yang menyandi berbagai molekul yang
berperan pada respon imun. Hal ini terjadi karena sinyal melalui TLR
meningkatkan kemampuan Antigen Precenting Cell (APC) untuk
menginduksi direfensiasi sel T menjadi sel efektor (Th1) untuk
memproduksi sitokin proinflamasi (Jo EK et al 2007, Boedina K.S 2010,
Nugraha. J, 2012).
Aktivasi melalui TLR oleh bagian mikobakterium mengimbas protein
adaptor sitoplasmik MyD88 (Myeloid Differentiation Factor 88) yang
berfungsi untuk mengaktifkan NF-kB dan mensekresi sitokin proinflamasi
(kKleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011)
Peranan MyD88 sangat penting untuk isyarat TLRs. Pada
percobaan dengan mencit yang kekurangan MyD88 sangat rentan
terhadap infeksi M.tuberculosis. sebaliknya di mencit yang kekurangan
TLR2, kerentanan terhadap M.tuberculosis tidak sebesar yang
kekurangan MyD88 dan kerentanan ini bervariasi di berbagai kajian.
Mencit yang kekurangan TLR4 tidak menunjukkan kerentanan yang tinggi
terhadap M.tuberculosis, sedangkan mencit yang kekurangan TLR9 masih
peka terhadap infeksi M.tuberculosis. Maka dapat disimpulkan, bahwa
pada percobaan dengan mencit yang defisien TLR2/TLR4/TLR9
menunjukkan fenotip yang lebih ringan dari pada mencit yang kekurangan
MyD88. Selain itu penelitian yang dilakukan oleh Cecille pada tikus

4
dimana, tikus dengan MyD88 (-/-) menyebabkan produksi sitokin menurun
dan masa hidup juga berkurang bahkan menyebabkan beban infeksi
karena M.tuberkulosis bertambah. Bahkan pada penelitian ini ditemukan
bahwa tikus dengan MyD88 (-/-) sangat rentan terhadap infeksi
M.tuberculosis dan menyebabkan terbentuknya lesi paru granulomatosa
yang lebih luas dibandingkan dengan tikus yang normal. Dari data
tersebut timbul dugaan bahwa selain TLRs, MyD88 juga sangat penting
dalam mengaktivasi sistem imun alami terhadap M.tuberculosis (Geiger
M.A, Levi I, Even R, Smith Y, Bowdish DM, Nussbaum G, Rachmilewitz J.
2012, Burns K, Janssens S, Brissoni B, Olivons N, Beyarert R, Yshopp J.
2003, Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011,
Cecille.et al 2007, Sugawara.I et al 2003).
Terjadinya kecacatan pada suatu gen, organisme disebabkan oleh
banyak faktor, salah satunya adalah terjadi perubahan/mutasi pada tingkat
sekuen basa nukleotidanya karena beberapa hal sehingga menyebabkan
terjadi kesalahan pada proses sintesis protein dan kerusakan gen.
Berdasarkan paparan diatas, peneliti tertarik dan ingin mengetahui
apakah pada penderita tuberkulosis paru terdapat mutasi pada gen
MyD88 dan bagaimana kadar TNF-α.

5
B. Rumusan Masalah
Berdasarkan latar belakang yang telah dijelaskan sebelumnya,
maka rumusan masalah pada penelitian ini adalah :
1. Apakah pada penderita tuberkulosis terdapat mutasi di gen MyD88?
2. Bagaimana pola substitusi asam amino yang terjadi pada gen MyD88
di penderita tuberkulosis paru?
3. Bagaimana kadar TNF-α pada penderita tuberkulosis paru?
C. Tujuan Penelitian
1. Tujuan Umum
Tujuan umum penelitian ini adalah Untuk mendeteksi pola mutasi
pada gen MyD88 dan kadar TNF-α pada penderita tuberkulosis paru.
2. Tujuan Khusus
Tujuan khusus dari penelitian ini adalah :
a. Mengetahui ada tidaknya mutasi yang terjadi pada gen MyD88
di penderita tuberkulosis paru.
b. Mengetahui pola substitusi asam amino yang terjadi pada gen
MyD88 pada penderita tuberkulosis paru
c. Mengetahui kadar TNF-α pada penderita tuberkulosis paru

6
D. Manfaat Penelitian
1. Hasil penelitian ini dapat digunakan sebagai bahan referensi untuk
mengetahui mekanisme tubuh dalam melawan infeksi yang
disebabkan oleh Mycobacterium tuberculosis.
2. Digunakan sebagai data awal yang dapat digunakan untuk menilai
keterlibatan MyD88 terhadap infeksi Mycobacterium tuberculosis.
3. Digunakan sebagai referensi tambahan dalam memecahkan masalah
yang terkait dengan infeksi Mycobacterium tuberculosis.
4. Dapat digunakan sebagai bahan referensi tambahan untuk
membuktikan keterlibatan faktor genetik terhadap keparahan penyakit,
khususnya penyakit yang disebabkan oleh Mycobacterium
tuberculosis

7
BAB II
TINJAUAN PUSTAKA
A. Tinjauan Khusus Tuberkulosis
1. Sejarah, Definisi dan Karakteristik Mycobacterium tuberculosis
Tuberkulosis paru (TB) adalah suatu penyakit infeksi kronik yang
sudah lama dikenal pada manusia yang dbuktikan dengan adanya
penemuan kerusakan tulang vetebrata torak yang khas pada TB dari
kerangka yang digali di Heidelberg dari kuburan zaman neolitikum,
penemuan di mumi yang berasal dari saman Mesir kuno (2000-4000 SM).
Hipokrates telah memperkenalkan terminologi phthisis yang diangkat dari
bahasa Yunani yang menggambarkan tampilan TB paru (Sudoyo A.W,
2009).
Pada tahun 1882 Robert Koch menemukan kuman penyebabnya
semacam bakteri berbentuk batang. Penyakit ini kemudian dinamakan
Tuberkulosis, dan hampir seluruh tubuh manusia dapat terserang tetapi
yang paling banyak menyerang organ paru (Sudoyo A.W, 2009).
Robert Koch mengidentifikasi basil tahan asam Mycobaterium
tuberculosis untuk pertama kalinya sebagai penyebab TB. Kemudian
mendemonstrasikan bahwa basil ini dapat berpindah kepada binatang
yang rentan (Sudoyo A.W, 2009).
Penyakit tuberkulosis adalah penyakit menular langsung yang
disebabkan oleh bakteri M.tuberculosis. Sebagian besar bakteri TB

8
menyerang paru-paru tetapi dapat juga mengenai organ tubuh lainnya
(Depkes RI 2002, Hardjoeno dkk 2007)
Mycobacterium tuberculosis adalah sejenis bakteri berbentuk
batang tipis, lurus atau agak bengkok, bergranula atau tidak mempunyai
selubung, tetapi mempunyai lapisan luar tebal yang terdiri dari lipid
(terutama asam mikolat) berukuran kira - kira 0,5 - 4 m x 0,3 - 0,6 m
Gambar 1. Bentuk Mycobacterium tuberculosis dengan penawaran Ziehl Neelsen
Sumber : www.Mediacastore.com/www.Emedicinehealth.com
Apabila dilihat menggunakan mikroskop elektron, dinding sel
M. tuberculosis terdiri dari 3 lapisan. Secara kimia dinding selnya sangat
kompleks dan mengandung banyak makromolekul lipofil. Tulang
punggung dinding sel mikobakterium terdiri dari dua polimer yaitu
peptidoglikan dan arabinogalaktan. Residu asam muramat dari
peptidoglikan membentuk ikatan fosfodiester dengan residu arabinosa dari
arabinogalaktan, tetapi satu dari sepuluh residu arabinosa terdapat dalam
polimer membentuk ikatan ester dengan asam mikolat. Asam mikolat ini
merupakan salah satu komponen utama kompleks lipid. Beberapa jenis

9
lemak terdapat pada mikobakterium adalah cord faktor, sulfolipid dan
wax D (Joklik, 1992)
Gambar 2. Struktur dinding sel Mycobacterium tuberculosis Sumber : www.Docstock.com
Asam mikolat merupakan molekul hidrofobik kuat yang membentuk
lapisan lipid mengelilingi organisme dan berperan dalam permeabilitas
permukaan sel. Asam ini juga berfungsi mempertahankan mikobakerium
dari serangan protein kationik, lisozim dan radikal oksigen dalam granula
fagosit serta melindungi mikobakterium ekstrasel dari dekomposisi oleh
komplemen dalam serum. Cord faktor atau trehalosa dimikolat umumnya
dihasilkan oleh galur M. tuberculosis yang virulens. Cord faktor ini bersifat
toksik terhadap sel mamalia dan merupakan inhibitor polimorfonuklear
(PMN) (Todar K, 2008)

10
M. tuberculosis bersifat aerob obligat, sehingga selalu ditemukan
pada lobus paru-paru bagian atas yang teraerasi dengan baik. Basil ini
merupakan parasit intrasel fakultatif yang umumnya terdapat dalam
makrofag. Bakteri ini mempunyai sifat istimewa, yaitu dapat bertahan
terhadap pencucian warna dengan asam alkohol, sehingga sering disebut
basil tahan asam (BTA) serta tahan terhadap zat kimia dan fisik karena
pada dinding sel terdapat kompleks asam mikolat dan lapisan lipid yang
dapat meningkatkan hidrofobisitas permukaan. Kuman tuberkulosis juga
tahan dalam keadaan kering dan dingin, bersifat dorman (Widoyono 2008,
Todar K 2005).
Bakteri tuberkulosis ini mati pada pemanasan 100°C selama 5-10
menit atau pada pemanasan 60°C selama 30 menit, dan dengan alkohol
70 - 95% selama 15-30 detik. Bakteri ini tahan selama 1 – 2 jam di udara
terutama di tempat lembab dan gelap (bisa berbulan - bulan), namun tidak
tahan terhadap sinar matahari langsung (Jawetz et al 2006, Misnadiarly
2006, Sardjono 2008)

11
2. Klasifikasi Mycobacterium tuberculosis
Klasifikasi bakteri berdasarkan buku Bergey's Manual of
Determinative Bacteriology, determinasi bakteri Mycobacterium
tuberculosis adalah sebagai berikut (Williams, L and Wilkins. 1994):
Kingdom : Procaryote
Divisio : Cyanobacteria
Ordo : Actinomycetales
Famili : Mycobacteriaceae
Genus : Mycobacterium
Spesies : Mycobacterium tuberculosis
3. Epidemiologi Penyakit Tuberkulosis
Walaupun pengobatan TB yang efektif sudah tersedia tapi sampai
saat ini TB masih tetap menjadi permasalah kesehatan dunia yang utama.
Pada tahun 1993 WHO mendeklarasikan TB sebagai global public health
emergency (Sudoyo A.W 2009, World Health Organization 2012).
Menurut laporan Global Report World Health Organization (WHO) pada
tahun 2011 terdapat 9 juta kasus TB baru di dunia atau sekitar 125 kasus
per 100.000 populasi dan 1.4 juta meninggal (990.000 HIV-negatif dan
430.000 TB-HIV). Diperkirakan 2.621.049 juta merupakan BTA positif
World Health Organization, 2012)

12
WHO pada tahun 2012 merilis data mengenai negara yang
termasuk kelompok dengan prevalensi yang tinggi dan Indonesia
(0.4-0.5 jt) termasuk dalam 5 negara dengan tingkat insiden tertinggi di
dunia dan menduduki peringkat ke 4 setelah India (2-2.5 jt), China
(0.9-1.1 jt), dan Afrika Selatan (0.4-0.6 jt) (World Health Organization,
2012). Di Indonesia dilaporkan pada tahun 2010 terdapat 690.000 kasus
(289 per 100.000 populasi) dengan kasus TB BTA positif di Indonesia
adalah 197.797, 101.750 adalah TB BTA negatif, 14.054 adalah TB
Extrapulmonal, 5.348 adalah kasus kambuh, dan 318.949 adalah kasus
baru (World Health Organization, 2012). Berdasarkan profil data
kesehatan Indonesia tahun 2011 provinsi dengan angka kejadian TB
tertinggi yaitu provinsi Jawa Barat (62.563 orang) dan provinsis Sulawesi
Selatan sebanyak 10.970 orang, dimana 8.860 adalah kasus TB BTA
positif dengan prevalensi kejadian terbanyak pada laki-laki (6.352)
dibandingkan perempuan (4.618) dengan rentang umur 45 – 54 tahun
(Kementerian Kesehatan Republik Indonesia, 2011).

13
Tabel 1. Angka Insident, Prevalensi dan Kematian TB di Indonesia, 1990 dan 2010
Kasus TB
Tahun 1990
Tahun 2010
Per tahun
Per 100.000
Penduduk
Per Hari
Per Tahun
Per 100.000
penduduk
Per hari
Capaian
(%)
Insiden semua tipe TB
626.867 343 1.717 450.000 189 1.233 44,9
Prevalensi semua tipe TB
809.592 443 ~ 690.000 289 ~ 34,8
Insident kasus baru TB paru BTA positif
282.090 154 773 NA NA NA NA
Kematian 168.956 92 463 64.000 27 175 70,6
*Global Tuberculosis Control WHO Report, 2011
Sumber : Ditjen PP&PL Kemeterian Kesehatan RI, 2011
4. Klasifikasi Penyakit Tuberkulosis
Sampai sekarang ini belum ada kesepakatan diantara para
klinikus, ahli radiologi, ahli patologi, mikrobiologi, dan ahli kesehatan
masyarakat tentang keseragaman klasifikasi tuberkulosis. Namun
beberapa literatur mengklasifikasi tuberkulosis menjadi beberapa
klasifikasi diantaranya (Sudoyo A.W 2009, Depkes RI 2011, Hardjoeno
dkk 2007, Depkes RI 2007) :
4.1 Tuberkulosis menurut Patologis
a. Tuberkulosis primer (Childhood tuberculosis)
b. Tuberkulosis post-primer (Adult tuberculosis)

14
4.2 Tuberkulosis menurut Radiologis
a. Tuberkulosis paru (Koch Pulmonum) aktif
b. Tuberkulosis tidak aktif
c. Tuberkulosis quiescent
d. Tuberkulosis minimal, yaitu terdapat sebagian kecil infiltrat nonka-vitas
pada satu paru maupun kedua paru tetapi jumlahnya tidak melebihi
satu lobus paru
e. Moderately advance tuberculosis, yaitu ada kavitas dengan diameter
tidak lebih dari 4 cm. Jumlah infiltrat bayangan halus tidak lebih dari
satu bagian paru. Bila bayangannya kasar tidak lebih dari sepertiga
bagian satu paru.
f. Far advance tuberculosis, yaitu terdapat infiltrat dan kavitas yang
melebihi keadaan pada moderately advance tuberculosis.
4.3 Tuberkulosis berdasarkan aspek kesehatan masyarakat menurut American Thoracis Society
a. Kategori 0 : Tidak pernah terpapar, dan tidak terinfeksi, riwayat
kontak negatif, tes tuberkulin negatif
b. Kategori I : Terpajan tuberkulosis, tetapi tidak terbukti ada infeksi,
riwayat kontak positif, tes tuberkulin negatif.
c. Kategori II : Terinfeksi tuberkulosi, tetapi tidak aktif, tes tuberkulin
positif, radiologis dan sputum negatif
d. Kategori III : Terinfeksi tuberkulosis dan aktif

15
4.4 Tuberkulosis berdasarkan organ tubuh
a. Tuberkulosis Paru adalah tuberkulosis yang menyerang jaringan
paru, tidak termasuk pleura (selaput paru).
b. Tuberkulosis Ekstra Paru adalah golongan tuberkulosis yang
menyerang organ tubuh lain selain paru, misalnya pleura, selaput
otak, selaput jantung, kelenjar limfe, tulang, kulit, usus, ginjal, alat
kelamin, dan lain-lain. Berdasarkan tingkat keparahan penyakitnya
yaitu :
(1) TB ekstra paru ringan, misalnya : TB kelenjar limfe, sendi, kelenjar
adrenal
(2) TB ekstra paru berat, misalnya : Meningitis, TB usus, TB saluran
kencing, TB alat kelamin
4.5 Tuberkulosis berdasarkan hasil pemeriksaan dahak mikroskopis
a. Tuberkulosis paru BTA positif
- Sekurang - kurangnya 2 dari 3 spesimen dahak SPS (sewaktu-
pagi-sewaktu) hasilnya BTA positif.
- 1 spesimen dahak SPS hasilnya BTA positif dan pada foto rontgen
dada menunjukkan gambaran tuberkulosis aktif.
- 1 spesimen dahak SPS hasilnya BTA positif dan biakan kuman TB
positif

16
- 1 atau lebih spesimen dahak hasilnya positif setelah 3 spesimen
dahak SPS pada pemeriksaan sebelumnya hasilnya BTA negatif
dan tidak ada perbaikan setelah pemberian antibiotik non OAT
b. Tuberkulosis paru BTA negatif
- Pemeriksaan 3 spesimen dahak SPS hasil BTA negatif dan foto
rontgen dada menunjukkan gambaran tuberkulosis aktif.
- TB paru BTA negatif rontgen positif dibagi berdasarkan tingkat
keparahan penyakitnya, yaitu bentuk berat dan ringan. Bentuk
berat bila gambaran foto rontgen dada memperlihatkan gambaran
kerusakan paru yang luas dan keadaan umum penderita buruk
.
4.6 Tuberkulosis berdasarkan riwayat pengobatan sebelumnya
a. Kasus baru adalah pasien yang belum diobati dengan OAT atau
sudah perna menelan OAT kurang dari satu bulan (4 minggu)
b. Kasus kabuh (Relaps) adalah pasien TB yang sebelumnya perna
mendapat pengobatan tuberkulosis dan telah dinyatakan sembuh
atau pengobatan lengkap, didiagnosis kembali dengan BTA positif
(apusan atau kultur)
c. Kasus putus berobat (Default/ Drop Out/ DO) adalah pasien TB yang
telah berobat putus berobat 2 bulan atau lebih dengan BTA positif
d. Kasus gagal (Failure) adalah pasien yang hasil pemeriksaan
dahaknya tetap positif atau kembali menjadi positif pada bulan kelima
atau lebih selama pengobatan.

17
e. Kasus pindahan (Transfer In) adalah pasien yang dipindahkan dari
UPK yang memiliki register TB lain untuk melanjutkan pengobatannya.
f. Kasus lain adalah semua kasus yang tidak memenuhi ketentuan
diatas. Dalam kelompok ini termasuk kasus kronik, yaitu pasien
dengan hasil pemeriksaan masih BTA positif setelah selesai
pengobatan ulangan
5. Patogenesis
a. Tuberkulosis Primer
Penularan TB paru terjadi karena kuman dibatukkan atau
dibersinkan keluar menjadi droplet nucleid dalam udara sekitar kita. Paru
merupakan port d’entree lebih dari 98% kasus infeksi TB. Partikel ini
dapat menetap dalam udara bebas selama 1 – 2 jam, tergantung pada
ada tidaknya sinar ultraviolet, ventilasi yang buruk, dan kelembapan.
Dalam suasana lembab dan gelap kuman dapat bertahan berhari-hari
sampai berbulan-bulan. Bila partikel tersebut terhirup oleh orang sehat
maka partikel tersebut akan menempel pada saluran napas, atau jaringan
paru dan dapat mencapai alveolus. Masuknya M.tuberculosis ini akan
segera diatasi oleh mekanisme imunologis non spesifik dimana kuman
tersebut akan pertama kali di hadapi oleh neutrofil kemudian oleh
makrofag akan memfagositosis kuman M.tuberculosis hingga kuman
mati dan dikeluarkan bersama gerakan silia dengan sekretnya (Sudoyo
dkk 2009, Depker RI 2007).

18
Pada sebagian kecil kasus, makrofag tidak mampu menghacurkan
kuman M.tuberculosis dan kuman akan bereplikasi dan berkembang biak
dalam sito-plasma makrofag. Di sini M.tuberculosis akan terbawa masuk
ke organ tubuh lainnya. Kuman yang bersarang di jaringan paru akan
membentuk koloni tuberkulosis pneumonia kecil yang disebut sarang
primer atau efek primer atau sarang (fokus) GOHN yang dapat terjadi
disetiap bagian jaringan paru (Sudoyo dkk 2009, Depker RI 2007).
Kuman M.tuberculosis akan menyebar melalui saluran
gastrointestinal, jaringan limfe, orofaring, dan kulit, terjadi limfadenofati
regional kemudian bakteri masuk ke dalam vena dan menjalar ke seluruh
organ seperti paru, otak, ginjal, tulang. Bila masuk ke dalam arteri
pulmonalis maka terjadi penjalaran ke seluruh bagian tubuh menjadi TB
milier (Sudoyo dkk 2009, Depker RI 2007).
Dari sarang primer akan timbul peradangan saluran getah bening
menuju hilus (Limfangitis lokal), dan juga dikuti oleh pembesaran kelenjar
getah bening hilus (Limfadenitis regional). Sarang primer limfangitis lokal
digabung dengan limfadenitis regional akan membentuk kompleks primer
(Ranke). Proses ini memerlukan waktu 3 – 8 minggu dengan
pertumbuhan kuman hingga mencapai jumlah 103 – 104 untuk
merangsang sistem imunitas seluler (Sudoyo dkk 2009, Depker RI 2007).
Selama berminggu – minggu awal proses infeksi, terjadi
pertumbuhan logaritmik kuman M.tuberculosis sehingga jaringan tubuh

19
yang awalnya belum tersensitisasi terhadap tuberkulin, mengalami
perkembangan sensitivitas. Pada saat terbentuknya kompleks primer ini,
infeksi TB primer dinyatakan telah terjadi. Hal tersebut ditandai oleh
terbentunya hipersensitivitas terhadap tuberkulo protein, yaitu timbulnya
respon positif terhadap uji tuberkulin. Selama masa inkubasi, uji tuberkulin
masih negatif. Setelah komplek primer terbentuk, imunitas seluler tubuh
terhadap TB telah terbentuk. Pada sebagaian besar individu dengan
sisten imun yang berfungsi baik, begitu sistem imun seluler berkembang,
proliferasi kuman TB terhenti. Namun, sejumlah kecil kuman TB dapat
tetap hidup dalam granuloma. Bila imunitas seluler telah terbentuk, kuman
TB yang masuk ke dalam alveoli akan segera di musnahkan (Sudoyo dkk
2009, Depker RI 2007).
Setelah imunitas seluler terbentuk, fokus primer di jaringan paru
biasanya mengalami resolusi secara sempurna membentuk fibrosis atau
kalsifikasi setelah mengalami nekrosis perkijuan atau enkapsulasi,
kelenjar limfe regional juga akan mengalami fibrosis dan enkapsulasi,
tetapi penyembuhannya biasanya tidak sesempurna fokus primer
dijaringan paru. Kuman M.tuberculosis dapat tetap hidup dan menetap
selama bertahun – tahun dalam jaringan ini dalam bentuk Dorman dan
±10% dapat mengalami reaktivasi kembali (Sudoyo dkk 2009, Depker RI
2007).
Kompleks imun juga dapat mengalami komplikasi. Komplikasi yang
terjadi dapat disebabkan oleh fokus paru atau kelenjar limfe regional.

20
Fokus primer di paru dapat membesar dan menyebabkan pneumonitis
atau pleuritis fokal. Jika terjadi nekrosis perkijuan yang berat, bagian
tengah lesi akan mencair dan keluar melalui bronkus sehingga
meninggalkan rongga dijaringan paru (kavitas). Kelenjar limfe hilus atau
paratrakea yang mulanya berukuran normal saat awal infeksi, akan
membesar karena reaksi inflamasi yang berlanjut. Bronkus dapat
terganggu, obstruksi parsial pada bronkus akibat tekanan eksternal dapat
menyebabkan ateletaksis. Kelenjar yang mengalami inflamasi dan
nekrosis perkijuan dapat merusak dan menimbulkan erosi dinding
bronkus, sehingga menyebabkan TB endobronkial atau membetuk fistual.
Massa kiju dapat menimbulkan obstruksi komplik pada bronkus sehingga
menyebabkan gabungan pneumonitis dan ateletaksis, yang sering
disebut sebagai lesi segmental kolaps-konsolidasi (Sudoyo dkk 2009,
Depker RI 2007).
Selama masa inkubasi, sebelum terbentuknya imunitas seluler,
dapat terjadi penyebaran limfogen dan hematogen. Pada penyebaran
limfogen kuman menyebar ke kelenjar limfe regional membentuk
kompleks primer. Sedangkan penyebaran hematogen, kuman
M.tuberculosis masuk ke dalam sirkulasi darah dan menyebar ke seluruh
organ tubuh. Adanya penyebaran hematogen inilah yang menyebabkan
TB disebut sebagai penyakit sistemik (Sudoyo dkk 2009, Depker RI 2007).
Penyebaran hematogen yang paling sering terjadi adalah bentuk
penyebaran hematogenik tersamar (occult hematogenic spread). Melalui

21
cara ini, kuman M.tuberculosis menyebar secara sporadic dan sedikit demi
sedikit sehingga tidak menimbulkan gejala klinis. Kuman M.tuberculosis
selanjutnya akan mencapai berbagai organ di seluruh tubuh. Organ yang
biasanya dituju adalah organ yang memiliki vaskularisasi yang baik,
misalnya otak, tulang, ginjal, dan paru sendiri, terutama apeks paru atau
lokus atas paru. Di organ tersebut kuman M.tuberculosis akan bereplikasi
dan membentuk koloni kuman sebelum terbentuk imunitas seluler yang
akan membatasi pertumbuhannya (Sudoyo dkk 2009, Depker RI 2007).
Di dalam koloni yang sempat terbentuk dan kemudian dibatasi
pertumbuhannya oleh imunitas seluler, kuman akan tetap hidup dalam
bentuk dormant. Fokus ini umumnya tidak langsung berlanjut menjadi
penyakit, tetapi berpotensi untuk menjadi fokus reaktivasi. Fokus potensial
di apeks paru disebut fokus SIMON. Bertahun – tahun kemudian, bila
daya tahan tubuh menurun, fokus TB ini akan mengalami reaktivasi dan
menjadi penyakit TB di organ terkait, misalnya meningitis, TB tulang dan
lain-lain (Sudoyo dkk 2009, Depker RI 2007).
b. Tuberkulosis Pasca Primer
Kuman yang dormant pada tuberkulosis primer akan muncul
bertahun-tahun kemudian sebagai infeksi endogen menjadi tuberkulosis
dewasa (Tuberculosis post primer). Tuberkulosis sekunder terjadi karena
daya tahan tubuh menurun seperti malnutrisi, alkohol, penyakit malignan,
diabetes, AIDS, gagal ginjal. Tuberkulosis pasca primer dimulai dengan

22
sarang dini yang berlokasi di regio atas paru (bagian apikal-posterior lobus
superior atau inferior). Invasinya adalah ke daerah parenkim paru – paru
dan tidak ke nodus hiler paru (Sudoyo dkk, 2009)
Sarang dini ini mula-mula juga terbentuk sarang pneumonia kecil.
Dalam 3 – 10 minggu sarang ini menjadi tuberkel yakni suatu granuloma
yang terdiri dari sel-sel Histiosit dan sel Datia-Langhans (Sel besar
dengan banyak inti) yang dikeliling oleh sel – sel limfosit dan berbagai
jaringan ikat (Sudoyo dkk, 2009).
TB sekunder juga berasal dari infeksi eksogen dari usia muda
menjadi TB usia tua (elderly tuberculosis) tergantung dari jumlah kuman,
virulensinya dan imunitas pasien. Sarang dini ini bisa menjadi :
- Direabsorbsi kembali dan sembuh tanpa meninggalkan cacat
- Sarang yang mula-mula meluas, tetapi segera menyembuh dengan
dengan serbukan jaringan fibrosis. Ada yang membungkus diri
menjadi keras, menimbulkan perkapuran. Sarang dini yang meluas
sebagai granuloma berkembang menghancurkan jaringan ikat
sekitarnya dan bagian tengahnya mengalami nekrosis, menjadi
lembek membentuk jaringan keju. Bila jaringan keju dibatukkan keluar
akan terjadi kavitas. Kavitas ini mula-mula berdinding tipis, lama-lama
dindingnya menebal karena infiltrasi jaringan fibroblas dalam jumlah
besar, sehingga menjadi kavitas sklerotik (kronik). Terjadinya
perkijuan dan kavitas adalah karena hidrolisis protein lipid dan asam

23
nukleat oleh enzim yang diproduksi oleh makrofag, dan proses yang
berlebihan antara sitokin dengan TNF-nya (Sudoyo dkk, 2009).
Di sini lesi sangat kecil, tetapi berisi bakteri sangat banyak. Kavitas
dapat : a). Meluas kembali dan menimbulkan sarang pneumonia baru. Bila
isi kavitas masuk dalam peredaran darah arteri, maka akan terjadi TB
milier dapat juga masuk dalam paru-paru sebelahnya atau tertelan masuk
dalam lambung ; b). Memadat dan membungkus diri sehingga menjadi
tuberkuloma. Tuberkuloma dapat mengapur atau menyembuh atau dapat
aktif kembali menjadi cair dan menjadi kavitas lagi ; c). Bersih dan
menyembuh (open healed cavity) (Sudoyo dkk, 2009).
Tuberkulosis milier merupakan hasil dari acute generalized
hematogenetic spread dengan jumlah kuman yang besar. Semua
tuberkel yang dihasilkan melalui cara ini akan mempunyai ukuran yang
lebih kurang sama. Istilah milier berasal dari gambaran lesi diseminata
yang menyerupai butir padi-padian/jewarut (millet seed). Secara patologi
anatomi, lesi ini berupa nodul kuning berukuran 1 – 3 mm, yang secara
histologi merupakan granuloma (Sudoyo dkk 2009, Depkes RI 2007)

24
Gambar 3. Patogenesis Tuberkulosis Sumber : http//muqarrabin.wordpress.com
6. Reaksi Imunologik pada Infeksi Tuberkulosis
Terdapat dua macam respon imun pertahanan tubuh terhadap
infeksi tuberkulosis yaitu respon imun selular (sel T dan makrofag yang
teraktivasi) bersama sejumlah sitokin dan respon imun humoral
(antibody-mediated). Respon imun seluler lebih banyak memegang
peranan dalam pertahan tubuh terhadap infeksi tuberkulosis. Respon
imun humoral tidak bersifat protektif tetapi lebih banyak digunakan untuk
membantu menegakkan diagnosis (Bothamley, G.H. 1995).

25
Respon ini di awali dengan diferensiasi limfosit B menjadi satu
populasi sel plasma yang memproduksi dan melepaskan antibodi spesifik
ke dalam darah yang dinamakan imunoglobulin. Imunoglobulin (Ig) di
bentuk oleh sel plasma yang berasal dari ploriferasi sel B akibat adanya
kontak dengan antigen. Antibodi yang terbentuk secara spesifik ini akan
mengikat antigen baru lainnya yang sejenis (Bothamley, G.H. 1995).
Respon imun primer terjadi sewaktu antigen pertama kali masuk ke
dalam tubuh, yang ditandai dengan munculnya IgM beberapa hari setelah
pemaparan. Kadar IgM mencapai puncaknya pada hari ke-7. Pada hari
ke-6 sampai ke-7 setelah pemaparan, IgG dapat terdeteksi dalam serum,
dan IgM mulai berkurang sebelum kadar IgG mencapai puncaknya yaitu
10-14 hari setelah pemaparan antigen. Respon imun sekunder terjadi
apabila pemaparan antigen terjadi untuk yang kedua kalinya, yang di
sebut juga booster. Puncak kadar IgM pada respon sekunder ini
umumnya tidak melebihi puncaknya pada respon primer, sebaliknya kadar
IgG meningkat jauh lebih tinggi dan berlangsung lebih lama. Perbedaan
dalam respon ini di sebabkan adanya sel B dan sel T memory akibat
pemaparan yang pertama (Kardjito,TVM, 1996).
IgG merupakan komponen utama imunoglobulin serum, kadarnya
dalam serum sekitar 13 mg/ml, merupakan 75% dari semua
imunoglobulin. Kadar IgG meninggi pada infeksi kronis dan penyakit
autoimun. Antibodi yang pertama di bentuk dalam respon imun adalah

26
IgM, oleh karena itu kadar IgM yang tinggi merupakan petunjuk adanya
infeksi dini (Kardjito,TVM, 1996).
M. tuberculosis terhirup sehingga masuk ke paru-paru, kemudian di
telan oleh makrofag. Makrofag tersebut mempunyai 3 fungsi utama, yaitu :
1. Memproduksi enzim proteolitik dan metabolit lainnya yang
memperlihatkan efek mycobactericidal.
2. Memproduksi sitokin sebagai respon terhadap M. tuberculosis yakni
IL-1, IL-6, IL-8, IL-10, TNF- TGF-. Sitokin mempunyai efek
imunoregulator yang penting.
3. Untuk memproses dan menyajikan anti gen terhadap limfosist T
(Andersen, A.B., dkk 1994).
Pada tuberkulosis primer, perkembangan infeksi M. tuberculosis
pada target organ tergantung pada derajat aktivitas anti bakteri makrofag
dari sistem imun alamiah serta kecepatan dan kualitas perkembangan
sistem imun yang di dapat. Pada sistem imun alamiah, basil akan di
eliminasi oleh kerjasama antara alveolar makrofag dan sel NK melalui
sitokin yang dihasilkannya yakni TNF- dan INF-. Mekanisme
pertahanan tubuh terhadap infeksi ini terutama dilakukan oleh sel-sel
pertahanan (sel T dan makrofag yang teraktivasi) bersama sejumlah
sitokin. Pada limfonodi regional, terjadi perkembangan respon imun yang
di dapat, yang akan mengenali basil tuberkulosis. Tipe respon imun ini
sangat tergantung pada sitokin yang dihasilkan oleh sistem imun alamiah.

27
Dominasi produksi sitokin oleh makrofag yang mensekresikan IL-12 akan
merangsang respon sel Th 1, sedangkan bila IL-4 yang lebih banyak
disekresikan oleh sel-T maka akan timbul respon oleh sel Th 2. Tipe
respon imun ini akan menentukan kualitas aktivasi makrofag untuk
mempresentasikan antigen kepada sel-T khususnya melalui jalur MHC
kelas-II (Andersen, A.B., dkk 1994).
Selama imunitas yang di dapat berkembang untuk mempercepat
aktivasi makrofag/monosit, terjadilah bakteremia. Basil menggunakan
makrofag sebagai sarana untuk menyebar dan selanjutnya tumbuh dan
menetap pada sel-sel fagosit di berbagai organ tubuh. Peristiwa ini akan
terjadi bila sel-T spesifik yang teraktivasi pada limfonodi mengalami
resirkulasi dan melewati lesi yang meradang yang selanjutnya akan
membentuk granuloma. Pada peristiwa ini TNF memegang peranan yang
sangat vital. Bila respon imun yang di dapat berkembang tidak adekuat
maka akan timbul manifestasi klinis akibat penyebaran basil yang berupa
tuberkulosis milier atau tuberkulosis meningen. Granuloma merupakan
mekanisme pertahanan utama dengan cara membatasi replikasi bakteri
pada fokus infeksi. Granuloma terutama terdiri atas makrofag dan sel-T.
Selama interaksi antara antigen spesifik dengan sel fagosit yang terinfeksi
pada berbagai organ, sel-T spesifik memproduki IFN- dan mengaktifkan
fungsi anti mikroba makrofag. Dalam granuloma terjadi enkapsulasi yang
di picu oleh fibrosis dan kalsifikasi serta terjadi nekrosis yang menurunkan
pasokan nutrien dan oksigen, sehingga terjadi kematian bakteri. Akan

28
tetapi sering terjadi keadaan di mana basil tidak seluruhnya mati tapi
sebagian masih ada yang hidup dan tetap bertahan dalam bentuk dorman.
Infeksi yang terlokalisir sering tidak menimbulkan gejala klinis dan bisa
bertahan dalam waktu yang lama (Kardjito,TVM, 1996).
Pada tuberkulosis post primer, pertahanan tubuh di dominasi oleh
pembentukan elemen nekrotik yang lebih hebat dari kasus infeksi primer.
Elemen-elemen nekrotik ini akan selalu dikelurkan sehingga akhirnya akan
terbentuk kavitas. Limfadenitis regional jarang terjadi, M. tuberculosis
menetap dalam makrofag dan pertumbuhannya di kontrol dalam fokus-
fokus yang terbentuk. Pembentukan dan kelangsungan hidup granuloma
di kontrol oleh sel-T, di mana komunikasi antara sel-T dan makrofag di
perantarai oleh sitokin. IL-1, TNF-, GM-CSF, TGF-, IL-6, INF- dan
TNF- merupakan sitokin yang mengontrol kelangsungan granuloma,
sebaliknya IL-4, IL-5 dan IL-10 menghambat pembentukan dan
perkembangan granuloma (Kardjito,TVM, 1996).
Proses aktivasi makrofag oleh sitokin merupakan faktor sentral
dalam imunitas terhadap tuberkulosis. Pada sistem ini, INF- telah di
identifikasikan sebagai sitokin utama untuk mengaktivasi makrofag, yang
selanjutnya dapat menghambat pertumbuhan patogen ini. Pembentukan
granuloma dan kavitas di pengaruhi oleh berbagai macam sitokin sebagai
hasil interaksi antara sel-T spesifik, makrofag yang teraktivasi dan
berbagai macam komponen bacterial (Kardjito,TVM, 1996).

29
7. Gejala dan Tanda Penyakit
Gejala penyakit TB dapat dibagi menjadi gejala umum dan gejala
khusus yang timbul sesuai dengan organ yang diserang (Sudoyo A.W
2009, Widoyono 2008, Laban Y 2008).
a. Gejala sistemik/ umum
Keluhan yang dirasakan pasien tuberkulosis dapat bermacam-
macam atau malah banyak pasien ditemukan TB paru tanpa keluhan
sama sekali dalam pemeriksaan kesehatan. Keluhan yang terbanyak
adalah :
- Demam : Biasanya subfebril menyerupai demam influenza tetapi
kadang-kadang panas badan dapat mencapai 40 – 41ºC. Serangan
demam pertama dapat sembuh sebentar, tetapi kemudian dapat
timbul kembali. Begitu seterusnya hilang timbulnya deman influenza
ini, sehingga pasien merasa tidak pernah terbebas dari serangan
demam influenza. Keadaan ini sangat dipengaruhi oleh sistem imun
tubuh pasien dan berat ringannya infeksi kuman tuberkulosis yang
masuk.
- Batuk/ batuk darah : Gejala ini banyak ditemukan. Batuk terjadi
karena adanya iritasi pada bronkus. Batuk ini diperlukan untuk
membuang produk – produk radang keluar. Karena terlibatnya
bronkus pada setiap penyakit tidak sama, mungkin saja batuk baru
ada setelah penyakit berkembang dalam jaringan paru. Sifat batuk
dimulai dari batuk kering (non-produktif) kemudian setelah timbul

30
peradangan menjadi produktif (menghasilkan sputum). Keadaan yang
lanjut adalah berupa batuk darah karena terdapat pembuluh darah
yang pecah. Kebanyakan batuk darah yang terjadi pada tuberkulosis
terjadi pada kavitas, tetapi dapat juga terjadi pada ulkus dinding
bronkus.
- Sesak napas : Pada penyakit yang ringan belum dirasakan sesak
napas. Sesak napas akan ditemukan pada penyakit yang sudah lanjut
yang infiltrasinya sudah meliputi setengah bagian paru-paru.
- Nyeri dada : Gejala ini agak jarang ditemukan. Nyeri dada timbul jika
infiltrasi radang sudah sampai ke pleura sehingga menimbulkan
pleuritis. Terjadin gesekan kedua pleura sewaktu pasien menarik/
melepaskan napasnya.
- Malaise : penyakit tuberkulosis bersifat radang yang menahun. Gejala
malaise sering ditemukan berupa anoreksia tidak ada nafsu makan,
badan makin kurus (berat badan turun), sakit kepala, meriang, nyeri
otot, keringat malam dll. Gejala malaise makin lama akan makin berat
dan terjadi hilang timbul secara teratur.
b. Gejala khusus
- Tergantung dari organ tubuh mana yang terkena, bila terjadi sumbatan
sebagian bronkus (saluran yang menuju ke paru-paru) akibat
penekanan kelenjar getah bening yang membesar, akan menimbulkan
suara "mengi", suara nafas melemah yang disertai sesak

31
- Kalau ada cairan dirongga pleura (pembungkus paru-paru), dapat
disertai dengan keluhan sakit dada
- Bila mengenai organ tulang, maka akan terjadi gejala seperti infeksi
tulang yang pada suatu saat dapat membentuk saluran dan bermuara
pada kulit di atasnya, pada muara ini akan keluar cairan nanah.
Pada anak – anak dapat mengenai otak (lapisan pembungkus
otak) dan disebut dengan meningitis (radang selaput otak), gejalanya
adalah demam tinggi, adanya penurunan kesadaran dan kejang-kejang.
B. SISTEM IMUN TERKAIT PENGENALAN TERHADAP MYCOBACTERIUM TUBERCULOSIS
Sistem imun alami dimulai dari pengenalan oleh TLR dan reseptor
yang lain terhadap molekul ligan yang terdapat di M. Tuberculosis, yang
dilanjutkan oleh jalur pengsignalan dan tindakan lain untuk penyajian
antigen M. tuberculosis di makrofag dan sel dendritik. M. Tuberculosis
merupakan penyakit infeksi yang berada di dalam sel yang tumbuh lambat
dan bertahan hidup di dalam makrofag inang. Di dalam makrofag alveoli,
M. Tuberculosis dikenali oleh TLR khususnya TLR2. (Nugraha J, 2012).
Dinding bakteri M. Tuberculosis mengandung asam mikolik hidrofobik dan
lapisan luar mengandung mengandung glycolipids like (mannose-capped)
lipomannan, mannoglycoproteins

32
Gambar 4. Susunan dinding sel M.tuberculosis
Sumber : Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011
Tahap pengenalan M. Tuberculosis dapat dibagi menjadi 2 yaitu :
1). Fokus ke PRRs dalam pengenalan PAMPs yang spesiifik terhadap
mikobakterium dan menghasilkan sitokin dan mengawali respon imun
adaptif. 2). Reseptor makrofag untuk fagositosis M. tuberculosis yang
terdiri dari reseptor komplemen (CR1, CR3, CR4) reseptor manosa dan
scavenger receptors. Pola reseptor pengenalan mikobakterium dan
downstream signaling pathway digambarkan secara skematis pada
gambar 5.
Mikobakterium dapat dikenali melalui pattern recognition receptor
(PRRs) yang berbeda-beda di inang (Kleinnijenhuis J, Marije O, Leo ABJ,
Mihai GN, Reinout VC, 2011).
Setelah mengenali mikobakterium, signal kaskade dalam sel
diaktivasi dengan penunjukkan aktivasi NF-kB yang mempengaruhi pada
hasil kemokin dan sitokin pro-inflamasi dan anti-inflamasi dimana jenis
kaskade tergantung dengan jenis PRR yang mengenali komponen M.

33
tuberculosis (Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC,
2011).
Ada dua kelompok reseptor membran TLR yang terbagi dalam :
kelompok TLR1, TLR2, TLR4, TLR5, TLR6, TLR10, TLR11, TLR12, dan
TLR13 yang merupakan khas TLR dipermukaan sel; kelompok TLR3,
TLR7 dan TLR9 yang dijumpai pada membran endosome. Aktivasi TLR
tersebut memicu produksi berbagai sitokin seperti IFN-γ, IL-2, IL-6, IL-8,
IL-12, IL-16 dan TNF-α.
Secara umum kaskade aktivasi TLR akan mengikat efektor
kemokin sebagai berikut : IL-12 akan mengikat Th1 dan mengarah ke cell-
mediated immunity yang berfungsi untuk menyerang bakteri dalam sel, IL-
23 akan mengikat Th17 dan mengarah ke sitokin proinflamasi yang
berfungsi untuk menyerang bakteri luar sel, IL-4 akan mengikat Th2 dan
membantu menghasilkan antibodi sel B (Kleinnijenhuis J, Marije O, Leo
ABJ, Mihai GN, Reinout VC, 2011).
Ada empat (4) macam anggota family PRR yaitu TLR (Toll-like-
receptor), NOD-like receptor (NLR), RIG-like receptor (RLR), C-type lectin
receptors (Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC,
2011).
Tempat TLR dan C-type lectin berada di membran sel (plasma),
endosom atau fagosom. Tempat NLR dan PLR berada di dalam sel
(Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011).

34
Gambar 5. Pola pengenalan reseptor M.tuberculosis dan downstream signaling pathway
Sumber : http://www.nature.com
1. Toll Like Receptors (TLRs)
Toll like receptors (TLRs) adalah keluarga patern recognition
receptor (PRR) yang mempunyai hubungan penting antara imunitas
bawaan dan dapatan, yang ,mengikat Pathogen-Associated Molecular
Patterns (PAMPs), yang ditemukan di dalam berbagai jenis organisme.
TLR merupakan protein terikat membran yang berisi leucine-rich
repeat (IL-1) receptor (TIR) di dalam area sitoplasmik yang sangat
terpelihara. TLRs terdapat pada berbagai sel imun, termasuk makrofag,
sel dendritik, sel B, sel T spesifik dan bahkan pada sel non imun seperti
fibroblas dan sel epitel.
Toll, pertama kali teridentifikasi sebagai sebuah sel yang penting
untuk penetapan orientasi dorsal-ventral selama perkembangan embriotik
dalam Drosophila melanogaster, kemudian diketahui memainkan sebuah
peran yang penting di dalam imunitas Drosophila termasuk infeksi jamur.

35
TLR yang terlibat dalam pengenalan M. tuberculosis yaitu TLR2,
TLR4 dan TLR9. TLR2 membentuk heterodimer dengan TLR1 atau TLR6.
Heterodimer tersebut berperan dalam pengenalan glikolipid dinding sel
mikobakterium seperti LAM, LM, glikoprotein 38-kDa dan glikoprotein 19-
kDa. Pengenalan phsophatidylinositol mannoside dilakukan oleh PIM,
pengenalam tricacyl lipoprotein diperankan oleh heterodomer TLR2/TLR1
dan pengenalan diacylated lipopprotein dilakukan oleh heterodimer
TLR2/TLR6. TLR juga berfungsi penting untuk sekresi IL-12 dalam
makrofag, tetapi tidak di dalam sel dendritik (Gerard JN, et all, 2002).
Beberapa PAMPs lain yang dikenali oleh TLR : DNA Genomic dari
galur Mycobacterium bovis, bacillus-Calmette-Gu’erin (BCG),
berkemampuan meningkatkan aktivasi sel NK dan mempengaruhi IFN
tipe I. Motif CpG yang berulang diketahui mengaktivas TLR9.
Lipoarabinomanna (LAM), lipomannan (LM) dan fosfatidil-myoinositol
mannoside (PIM) dikenali oleh TLR2. Lipoprotein 19 kDa dari M.
tuberculosis juga mengaktivasi makrofag lewat TLR2. TLR4 diaktivasi oleh
heat shock protein 60/65 kD. (Nugaha J, 2012).
Aktivasi melalui TLR oleh bagian mikobateria mempengaruhi
protein adaptor sitoplasmik MyD88 (Myeloid differentiation primary factor
88). Fungsi MyD88 akan mengaktivasi NF-kB dan mensekresi sitokin
proinflamasi seperti TNF-α, IL-6, IL-12. TRAM dan TRIF mengaktivasi
IRF-3 (Interferon regulated factor) dan IRF-7 yang akan mempengaruhi

36
pelepasan interferon untuk fungsi perlindungan (Kleinnijenhuis J, Marije O,
Leo ABJ, Mihai GN, Reinout VC, 2011).
Peran MyD88 sangat penting untuk isyarat TLR. Pada percobaan
dengan mencit yang kekurangan MyD88 sangat rentan terhadap infeksi
M.tuberculosis melalui udara. Sebaliknya di mencit yang kekurangan
TLR2, kerentanan terhadap M.tuberculosis tidak sebesar yang
kekurangan MyD88 dan kerentanan ini beragam di berbagai kajian. Mencit
yang kekurangan TLR4 tidak menunjukkan kerentanan yang tinggi
terhadap M.tuberculosis, sedangkan mencit yang kekurangan TLR9 peka
terhadap infeksi M.tuberculosis. mencit yang kekurangan kedua TLR2 dan
TLR9 lebih rentan terhadap infeksi M.tuberculosis. Maka dapat
disimpulkan, bahwa pada pecobaan dengan mencit ada beberapa TLR
yang secara berganda terkait pada pengenalan mikobakterium. Mencit
yang defisien TLR2/TLR4/TLR9 menunjukkan fenotip yang lebih ringan
dari pada mencit yang kekurangan MyD88. Hal ini menimbulkan dugaan
bahwa selain TLR, MyD88 juga sangat penting berperan dalam
mengaktivasi sistem imun alami terhadap infeksi M.tuberculosis (Geiger
M.A, Levi I, Even R, Smith Y, Bowdish DM, Nussbaum G, Rachmilewitz J.
2012, Burns K, Janssens S, Brissoni B, Olivons N, Beyarert R, Yshopp J.
2003, Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011).
Terdapat beberapa mekanisme respon imun alami yang bekerja
sama dengan TLR untuk memusnahkan M.tuberculosis dari dalam host
diantaranya yaitu dengan perangsangan TLR2 mengimbas Cyp21 dan

37
penerima vitamin D/vitamin D receptor (VDR) keduanya terkait peranan
dalam mengimbas cathelicidin yang membunuh langsung mikobakerium.
Ikatan antara SLPI (secretory leukocyte protease inhibitor) oleh TLR
memediasi perusakan dinding sel mikobakterium. TLR4 akan
mempengaruhi Lcn2 dan akan masuk ke dalam sel epitel, mengalveol
epitelan dan menghambat pertumbuhan mikobakterium dengan cara
menghambat penerimaan besi (Fe2+) bakteri (Nugraha J, 2012).
Makrofag memusnahkan mikobakterium yang masuk dengan
mengaktifkan beberapa fungsi efektor, seperti fagosom dan autofagi.
Nramp1 ditunjukkan dimembran fagosom dan mungkin memediasi
pemusnahan mikobakterium dengan cara menghalangi penangkapan Fe2+
oleh bakteri. IFN-γ dan ligand digilir merangsang autofagi dalam
makrofag. Autofagi bertanggung jawab pada pemusnahan mikobakterium
dengan memacu penyatuan fagosom yang mengandung mikobakterium
ke lisosom (Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC,
2011).
2. NOD like receptor (NLRs)
NLR merupakan keluarga protein yang sangat mirip protein plant R
(resistance), yang berperan penting dalam pertahanan terhadap penyakit
di tumbuhan. Keluarga NLR di manusia terdiri dari 20 anggota dengan
susunan yang sederhana. Inti molekul tersebut dibentuk oleh nucleotide-
binding domain, yang diberi nama NACHT (NAIP, CIITA, HET-E dan TP-1)

38
atau NOD (Nucleotide oligomerization domain). Bagian C-terminal
mengandung ulangan seri kaya luecin, yang lebih dahulu mengenali
PAMPs dari penyakit dan mengawali aktivasi molekul. Bagian N-terminal
mengandung bagian efektor dari CARD (caspase activation and
recruitment domain), PYRIN atau BIR (Baculovirus inhibitorof apoptosis
repeat domain). NLR yang mengandung CARD seperti misalnya NOD1
dan NOD2 terlebih dahulu membentuk oligomer dan kemudian
membentuk receptor-interacting protein 2 (RIP2) (atau CARS-containing
kinase-RICK) melalui interaksi CARD-CARD, sehingga menunjukkan
pengerahan NF-kB (Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN,
Reinout VC, 2011).
NOD2 memediasi rangsangan untuk hasilan sitokin proinflamasi
infeksi M.tuberculosis dan NOD2 ini merupakan reseptor untuk
peptidoglikan bakteri yaitu muramyl dipeptide (MDP). NLRP1, NLRP3 dan
IPAF berfungsi sebagai pengimbas penggabungan inflammasom, yang
menuju sekresi IL-18 dan IL-1β lewat caspase-1. Namun kajian terbaru
menunjukkan bahwa mikobanterium dapat menghambat pembentukan
inflammasom dan aktivasi caspase-1, sehingga hasil IL-1β terganggu.
Penghambatan aktivasi caspase-1 ini dilakukan oleh gen Mtb, zmp1 yang
menyanding Zn2+ metalloprotease. Jadi startegi M.tuberculosis untuk
menghindari respon imun yang dimediasi oleh inflammasom
(Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011).

39
3. C-Type Lectins
C-Type Lectin merupakan keluarga PRR yang terlibat dalam
penggenalan sususnan polisakarida dari penyakit. Receptor mannose
(MR, CD206) terdiri dari delapan rantai bagian pegenalan karbohidrat dan
satu bagian yang kaya cystein. MR banyak ditunjukkan di makrofag alveol.
Rangsangan mikobakterium melalui MR menyebabkan hasil sitokin anti-
inflamasi seperti IL-4 dan IL-13, menghambat hasil IL-12 dan kegagalan
mengaktivasi respon oksidatif (Kleinnijenhuis J, Marije O, Leo ABJ, Mihai
GN, Reinout VC, 2011).
Man-LAM dan komponen utama lain dari dinding sel
M.tuberculosis, seperti PIMs (phosphatidylikositol mannosides)
merupakan ligan alami mikobakterium untuk MR (Mannose receptor),
yang asalnya diketahui berfungsi memediasi fagpsitosis dari
mikobakterium. Ikatan M.tuberculosis dengan MR dapat mengimbas
fagositosis, tetapi penyatuan fagosom dengan lisososm terbatas
(Kleinnijenhuis J, Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011).
4. DC-SIGN
Dendritic cell-spesific intercellular molecule-3 grabbing nonintegrin
(DC-SIGN, CD209) berperang penting dalam interaksi antara sel dendritik
dengan M.tuberculosis. Reseptor ini terutama ditunjukkan di sel dendritik
dalam bentuk PRR dan reseptor adhesi, berperan dalam perpindahan sel
dendritik-sel T. DC-SIGN mengenali Man-LAM, Lipomannas dan α-glucan.

40
DC-SIGN memacu respon imun anti-inflamsi dengan cara mematangkan
sel dendritik yang terinfeksi dan mengimbas hasil IL-10 (Kleinnijenhuis J,
Marije O, Leo ABJ, Mihai GN, Reinout VC, 2011).
5. DECTIN-1
Dectin-1 merupakan reseptor yang terdiri dari bagian pengenalan
karbohidrat luar sel dan bagian ITAM dalam sel. Reseptor ini terutama
ditunjukkan di makrofag, sel dendritik, neutrofil dan sel T. Dectin-1
berperang penting dalam pengenalan M.tuberculosis dalam imunitas
alamiah dan mengimbas respon Th1 dan Th17 (Kleinnijenhuis J, Marije O,
Leo ABJ, Mihai GN, Reinout VC, 2011).
Gambar 6. Jenis PRR selain TLR yang berfungsi pada pengenalan komponen mikobakteria
Sumber : Saiga H, Yosuke S, Kiyoshi T, 2011

41
C. TINJAUAN KHUSUS MYELOID DIFFERENTIATION FACTOR 88 (MyD88)
Myeloid Differentiation Factor 88 (MyD88) merupakan gen
pengkode protein adaptor cytosolic yang berperan penting dalan respon
imun alamiah dan dapatan. Protein ini berfungsi sebagai signal transducer
penting pada jalur signal Interleukin-1 dan Toll-like receptor. Regulasi
jalur ini akan mengaktivasi gen numerous proinflamasi. MyD88 selain
terlibat sebagai protein adaptor pada jalur signal Toll-like receptor dan IL-1
pada sistem imun alamia dan dapatan juga dapat mengaktivasi via IRAK1,
IRAK2, IRF7 dan TRAF6, mengaktivasi NF-kB, sekresi sitokin dan
merespon inflammantory.
Menurut Bonnert et al (1997), menemukan overexpression pada
MyD88 menyebabkan peningkatan pada level transkripsi dari interleukin-8
promoter. Muzio et al (1997) melaporkan bahwa domain C-Terminal
MyD88 memiliki kesamaan urutan yang signifikan terhadap domain
sitoplasmik IL1RAP. Mereka memperlihatkan bahwa ekpresi MyD88 yang
kuat menginduksi aktivitas NF-ĸB dengan cara tergantung konsentrasi.
Ditambahkan pula region C-Terminal MyD88 bertindak sebagai inhibitor
domain negatif dari IL1R1/IL1RAP-induce NF-ĸB activity. MyD88
membentuk immunopresicipitable kompleks dengan IL1RAP dan IRAK2.
Medzhitov et al (1998) mendemonstrasikan signal reseptor Toll
manusia mempekerjakan protein adapter, MyD88 dan menginduksi
aktivasi NF-ĸB lewat IRAK (IRAK1) kinase dan protein TRAF6. Kaskade

42
signal TLR memediasi jalur NF-ĸB untuk kekebalan respon Drosophila
dewasa, MyD88 terlibat sebagai adaptor/regulator molekul umum famili
reseptor Toll/ IL1R untuk innate immunity.
MyD88 telah dipetakan, dimana menurut penelitian Hardiman et al
(1997) menjelaskan struktur gen MyD88 pada tikus berada pada
kromosom 9 , menurut urutan coding lengkapnya mencakup 5 exon
sedangkan homolog manusianya dipetakan pada 3p22-p21.3
menggunakan analisis PCR. Menurut Bonnert et al (1997) dengan
menggunakan Fluorescence in situ hubridization untuk mempetakan gen,
dimana gen MyD88 pada manusia terletak di 3p22-3p21.3 yang berada
pada kromosom 3 yang dikode oleh 5 exon dengan ukuran 4.5 kB
Gambar 7. Lokasi gen MyD88
Sumber : http://ghr.nlm.nih.gov/gene/MYD88

43
MyD88 terdiri dari bagian TIR domain pada posisi C-terminal
(sekitar 150 residu asam amino) dan death domain pada posisi N-terminal
(sekitar 90 residu asam amino) serta daerah intermediated domain
(Gambar 8a). Pada respon imun bawaan, TIR domain pada TLR
berfungsi penting dalam pembentukan kompleks inisiasi sinyal, kemudian
akan merekrut IL-1 receptor-associated kinase (IRAK) untuk berinteraksi
dengan death domain. IRAK diaktivasi melalui proses fosforilasi dan
berasosiasi dengan TRAF6 yang akan mengarah pada aktivasi 2 jalur
yang berbeda yaitu aktivasi JNK dan NFĸB (Gambar 8b) (Takeda, K dkk,
2004)
Sumber : Takeda K; Shizuo A, 2004
b
a
Gambar 8 a. Posisi pembagian daerah domain MyD88. b. Interaksi TLR, MyD88
(TIR domain, death domain dan TLR )
Sumber : Takeda K, Shizuo A, 2004

44
D. TINJAUAN KHUSUS MOLEKULER
1. Penemuan Awal
Era penemuan materi genetik dibuka oleh F.Miescher dengan
menggunakan mikroskop sederhana, F.Miescher menyatakan bahwa
bahan aktif yang ada di dalam nukleus disebut sebagai nuclein. Namun
sampai sekarang ini nuclein belum bisa diketahui apakah merupakan
kromosom atau DNA (Fatchiyah dkk, 2011, Yumono T, 2005).
Kromosom merupakan struktur seperti benang pada nukleus sel
eukariot yang nampak pada saat sel mulai membelah. Pada organisme
diploid, kromosom berjumlah diploid (2 set) pada setiap selnya. Kromosom
dapat dibedakan menjadi kromosom autosomal dan kromosom seks.
Kedua set kromosom tersebut membawa gen-gen yang berpasangan,
kecuali pada kromosom-Y (Fatchiyah dkk, 2011, Yumono T, 2005).
Gen adalah unit heriditas suatu organisme hidup. Gen dikode
dalam materi genetik organisme yang disebut molekul DNA, atau RNA.
Ekspresi gen dipengaruhi oleh lingkungan internal atau eksternal seperti
perkembangan fisik atau perilaku dari organisme itu. Gen tersusun atas
urutan basa nukleotida yang terdiri dari daerah yang mengkode suatu
informasi genetis (ekson), daerah yang tidak mengkode suatu informasi
genetis (intron), serta bagian yang mengatur ekpresi gen yaitu sekuens
pengontrol ekspresi gen (regulatory sequence) (Fatchiyah dkk, 2011,
Yumono T, 2005).

45
Gambar 9. Struktur kromosom, gen, dan DNA
Sumber : http://khairul-anas.blogspot.com/2012/04/dna-gen-
kromosom.html
2. Struktur DNA
Pada tahun 1953, James Watson dan Francis Crick, berdasarkan
atas data kimia dan fisik, Watson dan Crick membuat model struktur DNA
yang disebut untai ganda (double helix). Untai ganda tersebut tersusun
oleh dua rantai polinukleotida saling berikatan melalui ikatan hidrogen
antara basa adenin (A) dengan Thymine (T), dan antara Guanine (G)
dengan Cytosine (C), dan kedua rantai tersebut mempunyai orientasi yang
berlawanan (antiparalel) : rantai yang satu mempunyai orientasi 5’ → 3’,
sedangkan rantai yang lain berorientasi 3’ → 5’. Spesifisitas pasang basa
semacam ini disebut sebagai komplementaritas. Proporsi basa A dan T
serta G dan C selalu sama sehingga komposisi DNS dapat dinyatakan
Pada kromosom eukoaryotik, gen
tersusun atas ekson dan intron,
gen-gen ini terdapat pada pita
DNA yang diuntai dan
dimampatkan oleh protein histon
dalam bentuk kromosom

46
dengan kandungan G + C yang berkisar dari 26% sampai 74% (hukum
Chargaff) (Fatchiyah dkk, 2011, Yumono T, 2005).
Kerangka gula deoksiribosa dan fosfat yang menyusun DNA
terletak di bagain luar molekul, sedangkan basa purin dan pirimidin
terletak disebelah dalam untaian (helix) dengan diameter untaian DNA
adalah 20 Å (angstrom). Struktur untaian berulang setiap 10 basa atau
dengan kata lain ada 10 pasang basa setiap putaran untaian. Untaian
DNA mempunyai dua lekukan (groove) eksternal yaitu lekukan besar
(major groove) dan lekukan kecil (minor groove). Kedua lekukan tersebut
mempunyai peranan sebagai tempat melekatnya molekul protein tertentu.
(Fatchiyah dkk, 2011, Yumono T, 2005).
Gambar 10. Struktur pita DNA ganda Sumber : http://khairul-anas.blogspot.com/2012/04/dna-gen-
kromosom.html
Ikatan Kedua rantai heliks ganda DNA dapat melakukan replikasi
yang memiliki susunan asam nukleotida yang sama dengan cetakan
aslinya, karena masing-masing untai mengandung sekuens nukleotida

47
yang identik berkomplemen dengan sekuens untai pasangannya. Masing-
masing untai dapat berperan sebagai cetakan untuk sintetis dari untai
komplemen baru yang identik dengan pasangan awalnya (Fatchiyah dkk,
2011).
Beberapa faktor yang menentukan struktur DNA yaitu
Struktur untaian (helix) ditentukan oleh tumpukan (stacking) basa-basa
nukleotida berdekatan yang ada pada satu untai yang terletak pada suau
bidang datar yang tegak lurus terhadap aksis untai ganda. Dan struktur
untai-gandanya ditentukan oleh ikatan hidrogen antara basa-basa yang
berpasangan (Yuwono, T. 2005)
Fungsi dari DNA yang paling penting adalah membawa gen yang
mengandung informasi yang menentukan jenis protein yang harus
disintetis, kapan, dalam tipe sel yang mana, dan berapa banyak jumlah
protein yang harus disintetis (Fatchiyah dkk, 2011).
3. Teknik Rekayasa Genetik
Perkembangan teknologi rekayasa genetika pada dasarnya
didukung oleh perkembangan biologi molekuler. Beberapa teknik utama
yang berhubungan dengan rekayasa senyawa terapetik dan diagnostik,
antara lain polymerase chain reaction (PCR), teknik sequencing DNA,
antibodi monoklonal, teknik pelacakan molekuler, teknik imunoblotting,
teknologi transgenik dan rekayasa protein (Radji M, 2011).

48
a. Polymerase Chain Reaction (PCR)
Polymerase Chain Reaction (PCR) merupakan suatu proses
sintesis enzimatik untuk mengamplifikasi fragmen DNA secara invitro.
PCR pertama kali ditemukan oleh Kary Mullis. Teknik PCR dapat jumlah
fragmen DNA hingga mencapai 106 – 107 kali dalam waktu singkat. Pada
setiap n siklus PCR, akan diperoleh sebanyak 2n kali DNA target.
keberhasilan PCR sangat tergantung pada kemampuan untuk hanya
mengamplifikasi DNA target dan tidak mengamplifikasi DNA non-target.
Proses PCR untuk memperbanyak DNA terdiri dari serangkaian
siklus suhu yang berulang, dimana masing-masing siklus terdiri dari 3
tahap. (i). Tahap denaturasi DNA cetakan pada suhu 94-96ºC, dimana
pada tahap ini terjadi pemisahan DNA heliks ganda menjadi 2 untai
tunggal DNA. (ii). Tahap penempelan (annealing) oligonukleotida primer
pada suhu 45-60ºC, dengan untaian tunggal DNA cetakan pada ujung 3’.
(iii). Tahap elongation, yaitu pemanjangan primer menjadi suatu untai DNA
baru, yang komplementer terhadap masing-masing DNA cetakan untai
tunggal oleh enzim DNA polimerase pada suhu 72ºC (Radji M 2011,
Yuwono T 2006).
Ketiga tahap siklus tersebut diulang sesuai dengan jumlah siklus
amplifikasi. Pada siklus pertama dua untai tunggal DNA cetakan akan
disalin menjadi 2 DNA untai ganda. Pada siklus kedua, 2 DNA cetakan
untai ganda masing-masing akan bertindak sebagai cetakan sehingga

49
pada siklus kedua akan dihasilkan 4 DNA untai ganda. Pada siklus
berikutnya akan dihasilkan jumlah DNA akan disalin menjadi 8 kali, dan
seterusnya (Radji M 2011, Yuwono T 2006).
Pada siklus akhir, DNA cetakan akan digandakan secara
eksponensial sehingga dihasilkan DNA dalam jumlah yang berlipat ganda
hanya dalam waktu yang relatif singkat sekitar 3 – 4 jam (Radji M 2011,
Yuwono T 2006).
Beberapa komponen penting dalam proses PCR terdiri dari :
1) DNA cetakan
Fragmen DNA yang akan dijadikan cetakan untuk menggandakan
DNA biasanya berukuran pendek, kurang dari 1.000 pasang basa
(bp). Hasil amplifikasi yang efisien antara 100 – 400 bp. DNA cetakan
sebaiknya murni, untuk menghasilkan amplikon (hasil amplifikasi
DNA) yang murni. Suspensi DNA yang tidak murni akan
mempengaruhi reaksi amplifikasi dan dapat menghambat kerja enzim
DNA polimerase.
2) Primer
Primer merupakan oligonukleotida rantai pendek yang dibuat secara
sintetik, umumnya sepanjang 15-32 pasang basa. Primer yang
digunakan harus mampu mengenali urutan DNA cetakan yang akan
diamplifikasi.

50
Beberapa hal yang harus diperhatikan dalam perancangan primer
oligonukleotida antara lain adalah : (i). Kandungan GC primer harus
45-60%, dimana ujung 3’ harus terdiri dari basa G atau C, (ii). Harus
dihindar susunan tiga basa berturut – turut terdiri dari G atau C pada
ujung primer, misalnya CCG, CCC, GCG, GGG atau GCC, (iii). Urutan
basa sepanjang primer tidak boleh saling komplementer, sehingga
membentuk dimer atau membentuk ikatan seperti jepitan rambut
(hairpins), (iv). Hindari merancang suatu primer pada daerah repetitif
dan (v). Pemilihan suhu annealing yang tepat.
Penentuan suhu annealing primer oligonukleotida (Tm = melting
temperature) berdasarkan formula :
Tm = [(jumlah A+T) x 2ºC] + [(jumlah C+G) x 4ºC]
3) DNA polimerase
Enzim yang digunakan dalam PCR adalah enzim DNA polimerase
termostabil yang diisolasi dari bakteri Thermus aquaticus, yang hidup
di dalam sumber air panas yang disebut dengan TaqDNA polymerase.
Enzim Taq DNA polimerase tetap aktif pada suhu 100ºC, dengan
aktifitas optimal pada suhu 92-95ºC. Selain itu enzim DNA polimerase
yang termostabil lainnya diisolasi dari bakteri yang hidup pada turbin
kapal selam yang disebut dengan Vent DNA polymerase.
Aktivitas enzim DNA polimerase ini mengkatalisis polimerisasi DNA
berlangsung dari ujung 5’ ke ujung 3’ dan memiliki waktu paruh sekitar

51
40 menit pada suhu 95ºC. Biasanya untuk setiap 100 μL reaksi PCR
ditambahkan 2,0 – 2,5 unut Taq DNA polymerase.
4) Larutan buffer
Larutan buffer untuk reaksi PCR terdiri atas 50mM KCI, 10 mM Tris-Cl
(pH 8,3) dan 1,5 mM MgCl2.
Konsentrasi ion Magnesium dalam larutan buffer merupakan faktor
yang sangat menentukan proses denaturasi, penempelan primer,
aktivitas enzim, ketelitian reaksi, dan spesifisitas produk PCR.
5) Deoksi ribonukleotida trifosfat (dNTPs)
Konsentrasi nukleotida yang terdiri dari dATP, sTTP, dCTP, dan
dGTP, sangat penting untuk memberikan spesifisitas yang tinggi pada
hasil reaksi.
Biasanya konsentrasi masing-masing dNTP adalah 200 μM, pada pH
7,0. Bila konsentrasi terlalu tinggi akan menimbulkan
ketidakseimbangan dengan DNA polimerase. Sedangkan bila
konsentrasi terlalu rendah dapat mempengaruhi ketepatan dan
ketelitian hasil amplifikasi DNA.
6) PCR gene cycle
PCR gene cycle pertama kali dikembangkan oleh perusahaan
PerlinElmen sebagai pemegang paten asli. Pada saat ini telah
dikembangkan dan diproduksi berbagai macam alat PCR gene cycle
dari berbagai perusahaan yang bergerak di bidang bioteknologi. Alat
ini secara meregulasi suhu dan siklus waktu yang dibutuhkan untuk

52
reprodusbilitas dan keakuratan reaksi amplifikasi (Fatchiya dkk 2011,
Radji M 2011, Yuwono T 2006).
b. Teknik Sequencing DNA
Teknik sequencing DNA (DNA sequencing) adalah cara
menentukan urutan basa-basa nukleotida suatu fragmen DNA. Teknik ini
merupakan salah satu teknik yang sangat penting dalam bidang biologi
molekuler, yang dapat dimanfaatkan untuk menentukan urutan basa
nukleotida suatu gen atau sekuen genom total suatu sel atau organisme
(Radji M, 2011).
Teknik pengurutan basa DNA terdiri dari 2 macam cara yang
dikembangkan hampir secara bersamaan yaitu : i). Cara degradasi
kimiawi oleh A.Maxam dan W. Gilbert di Amerika ; ii). Cara terminasi rantai
oleh F. Sanger dan A.R. Coulson di Inggris (Radji M, 2011).
2) Teknik Maxam – Gilbert
Teknik pengurutan basa yang pertama kali dikenal adalah
teknik kimia yang dikembangkan A.Maxam dan W. Gilbert pada tahun
1977. Fragmen DNA yang akan diurutkan dilabel dengan radioaktif
(32P) pada salah satu ujung 5’. Teknik Maxam – Gilbert, dapat
diterapkan baik untuk urutan DNA untai ganda maupun DNA untai
tunggal (Radji M, 2011).
Molekul DNA dipotong terlebih dahulu secara parsial dengan
piperidin. Pengaturan lama inkubasi atau konsentrasi piperidin yang

53
digunakan dapat menghasilkan potongan fragmen DNA dalam
berbagai ukuran. Selanjutnya basa DNA dimodifikasi dengan
menggunakan senyawa kimia tertentu. Dimetilsulfat digunakan untuk
metilasi basa G, asam fosfat untuk menghidrolisis basa A dan G,
sedangkan hidrazin digunakan untuk menghidrolisis basa C dan T.
Dengan demikian akan dihasilkan empat jenis fragmen DNA yang
memiliki ukuran berbeda dengan masing-masing memiliki ujung G,
ujung C, ujung A dan ujung T (Radji M, 2011).
Fragmen-fragmen DNA yang dihasilkan selanjutnya di
elektroforesis dengan gel poliakrilamida. Berdasarkan pola migrasi
pada gel elektroforesis dapat ditentukan urutan basa-basa DNA dari
nmolekul DNA yang ditentukan urutan basa-basanya (Radji M, 2011).
3) Teknik Sanger – Coulson
Teknik Sanger – Coulson merupakan teknik yang saat ini sering
digunakan untuk menentukan urutan basa DNA. Teknik Sanger –
Coulson lebih praktis dari Maxam – Gilbert, sehingga teknik Sanger
lebih banyak dikembangkan dan digunakan untuk sequencing DNA
(Radji M, 2011).
Teknik Sanger – Coulson pada dasarnya memanfaatkan sifat
enzim DNA polimerase yaitu fragmen klenow, yang memiliki
kemampuan mensintesis DNA dengan adanya dNTP dengan ddNTP.
Molekul dNTP, tidak memiliki gugus hidroksil (OH), pada atom C
nomor 2 pada cincin gula pentosa, sedangkan molekul ddNTP, tidak

54
memiliki 2 gugus PH pada posisi atom C nomor 2 dan nomor 3 pada
cincin gula pentosa (Radji M, 2011).
Teknik sequencing DNA menggunakan teknik
dideoksinukleotida dilakukan pada 4 tabung reaksi yag berbeda,
dimana pada setiap tabung reaksi berisi campuran reagen yang terdiri
dari cetakan DNA untai tunggal yang disekuens, primer
oligonukleotida, DNA polimerase, campuran dNTP dan larutan buffer
(Radji M, 2011).
Gambar 11. Perbedaan struktur molekul dNTP dan ddNTP
Sumber : http://khairul-anas.blogspot.com/2012/04/dna-gen-
kromosom.html
DNA cetakan akan disekuensing adalah DNA untai tunggal
sehingga biasanya diklon terlebih dahulu dalam vektor M13.
Sedangkan molekul ddNTP yang dilabeli dengan senyawa radioaktif
Jika ddNTP terikat pada rantai
DNA yang sedang direplikasi,
proses pemanjangan DNA akan
terhenti karea basa DNA lain tidak
dapat bereaksi (tidak memiliki
gugus OH)

55
atau non-radioaktif, hanya ditambahkan pada campuran reaksi yang
sesuai yaitu untuk tabung A hanya ditambahkan ddATP, untuk tabung
C hanya ditambahkan ddCTP, untuk tabung G ditambahkan ddGTP,
sedangkan untuk tabung T ditambahkan ddGTP (Radji M, 2011).
Pada proses pemanjangan untai DNA, selain menggunakan
dNTP, secara acak juga menggunakan ddNTP, maka jika ddNTP
terikat, polimerisasi lebih lanjut basa-basa DNA tidak terjadi atau
terhenti, pada langkah-langkah dasar proses pengurutan basa-basa
DNA (Radji M, 2011).
Disiapkan 4 tabung reaksi yang mengadung masing-masing
DNA untai tunggal yang akan disekuens, primer oligonukleotida,
empat jenis dNTP, enzim DNA polimerase dan larutam buffer.
Kemudian kedalam rabung No. 1 ditambahkan ddGTP, tabung No. 2
ditambahkan ddCTP, tabung No. 3 ditambahkan ddATP, dan tabung
No. 4 ditambahkan ddTTP. DNA cetakan dalam masing-masing
tabung kemudian diamplifikasi secara in vitro dengan teknik PCR.
Dengan demikian dalam setiap reaksi akan dihasilkan sejumlah
fragmen DNA dengan ukuran bervariasi dimana setiap fragmen DNA
tersebut memiliki gugus ddNTP yang telah dilabel dengan radioaktif
pada setiap ujung 3’nya. Untuk mengetahui ukuran fragmen-fragmen
DNA yang terbentuk dilakukan elektroforesis menggunakan gel
poliakrilamida, kemudian dideteksi dengan alat autoradiografi (Radji
M, 2011).

56
Berdasarkan perbedaan – perbedaan migrasi masing-masing
fragmen DNA akan dapat ditentukan urutan basa-basa DNA-nya.
Hasil sequencing DNA merupakan urutan basa yang komplementer
terhadap urutan basa-basa yang terbaca pada hasil elektroforesis
(Radji M, 2011).
Sistem pelabelan yang semula dilakukan dengan radioisotop,
telah dikembangkan dengan sistem pelabelan senyawa fluoresen
yang berbeda warna pendarannya untuk setiap dNTP terminal.
Perbedaan emisi cahaya senyawa fluoresen ketika tereksitasi oleh
sinar laser, akan terbaca dengan jelas oleh alat pendeteksi optik
(detektor) yang terdapat pada mesin DNA sekuensing, sehingga
urutan basa-basa DNA dapat terdeteksi pada sistem elektroforesis
kapiler (Radji M, 2011).
c. Elektroforesis
Elektroforesis adalah suatu teknik pemisahan molekul seluler
berdasarkan atas ukurannya, dengan menggunakan medan listrik yang
dialirkan pada suatu medium yamg mengandung sampel yang akan
dipisahkan. Kecepatan molekul yang bergerak pada medan listrik
tergantung pada muatan, bentuk dan ukuran. Dengan demikian,
elektroforesis dapat digunakan untuk pemisahan makromolekul seperti
DNA yang bermuatan negatif, jika molekul yang bermuatan negatif
dilewatkan melalui suatu medium (gel agarose) kemudian dialiri arus listrik

57
dari satu kutub ke kutub yang berlawanan muatannya, maka molekul
tersebut akan bergerak dari kutub negatif ke kutub positif. Posisi molekul
yang memisah pada gel dapat dideteksi dengan pewarnaan atau
autoradiografi, ataupun dilakukan kuantifikasi dengan densitometer
(Fatchiya dkk 2011, Radji M 2011, Yuwono T 2006, Yuwono T, 2005).
Teknik elektroforesis dapat digunakan untuk analisis DNA, RNA
maupun protein. Elektroforesis DNA dilakukan misalnya untuk
menganalisis fragmen-fargmen DNA hasil pemotongan dengan enzim
restriksi. Fragmen molekul DNA yang telah dipotong-potong dapat
ditentukan ukurannya dengan cara membuat gel agarose, yaitu suatu
bahan semi padat berupa polisakarida yang diekstraksi dari rumput laut.
Gel agarosa dibuat dengan melarutkannya dalam suatu buffer yang
kemudian dipanaskan. Dalam keadaan panas, gel akan dicetak diatas
sebuah lempeng yang terbuat dari Perspex. Sebelum dingin dan
memadat, pada ujung gel dibuat lubang menggunakan lembaran perspex
tipis yang menyerupai sisir. Sisir tersebut ditancapkan pada salah satu
ujung gel yang masih cair. Gel agarose yang sudaj terbentuk akan
dimasukkan ke dalam suatu tangki yang berisi buffer yang sama dengan
yang digunakan untuk membuat gel. Buffer dapat dibuat misalnya dengan
tri-asetat-EDTA (TAE) atau tri-borat-EDTA (TBE) (Yuwono, T, 2005)
Setelah DNA dimasukkan ke dalam lubang sampel, arus listrik
dialirkan. Setelah beberapa waktu gel akan direndam dalam larutan yang
mengandung ethidium bromida. Etidium bromida akan menyisip kedalam

58
DNA, penggunaan etidium bromida dimaksudkan untuk membantu
visualisasi karena etidium bromida dapat memendarkan sinar ultraviolet.
Jika gel disinari dengan UV dari bawah maka akan tanpak citra berupa
pita-pita pada gel (Yuwono, T. 2005).
Selain teknik elektroforesis linear sekarang telah dikembangkan
teknik elektroforesis yang lain seperti teknik pulse field gel electrophoresis
(PFGE), ortogonal field elternation gel electrophoresis (OFAGE),
transverse alternation field electrophoresis (TAFE) (Yuwono, T. 2005).
4. Kode Genetik
Gen tertentu membawa informasi yang dibutuhkan untuk membuat
protein dan informasi inilah yang disebut kode genetik. Kode genetik
adalah cara pengkodean urutan nukleotida pada DNA atau RNA untuk
menentukan urutan asam amino pada saat sintesis protein.
Di alam ada 20 macam asam amino yang umum terdapat di dalam
struktur polipeptida jasad hidup. Masing-masing asam amino mempunyai
kodom yang spesifik sedangkan nukleotida hanya ada empat (4) macam
yaitu A, U, G dan C (Yuwono. T, 2005).
Berbagai penelitian dilakukan untuk memecahkan empat basa
nitrogen yang mampu mengkode 20 asam amino. Jika suatu kodon hanya
terdiri dari dua nukleotide maka hanya akan mendapatkan (42) 16 asam
amino, tetapi apabila kodom disusun oleh 3 nukleotida, maka akan
diperoleh (43) 64 asam amino, sedangkan jumlah asam amino yang umum

59
diketahui pada jasad hidup hanya 20 macam. Beberapa kodon diketahui
mengkode asam amino yang sama. Fenomena ini dikenal sebagai genetic
code redundancy (degeneracy). Oleh karena adanya beberapa kodon
yang berbeda untuk satu asam amino yang sama, maka dikenal 64
macam kodon, tiga diantaranya yaitu TAA (UAA pada mRNA), TAG (UAG
pada mRNA), dan TGA (UGA pada mRNA) tidak menkode asam amino
apapun karena fungsi ketiga kodon tersebut adalah berfungsi sebagai
kodon terminasi (kodon untuk mengakhiri) proses translasi.
Tabel 2. Kode Genetik “ Universal”
Sumber : Yuwono, T. Biologi Molekuler, 2005.

60
Ada beberapa aspek yang perlu diketahui mengenai kode genetik,
yaitu :
a. Kode genetik bersifat tidak saling tumpang-tindih (non-overlapping),
kecuali pada kasus tertentu, misalnya pada bakteriofag ɸX174 yang
mempunyai kodon tumpang-tindih.
b. Tidak ada sela (gap) diantara kodon satu dengan kodong yang lain.
c. Tidak ada koma diantara kodon.
d. Kodon bersifat degenerate, artinya ada beberapa asam amino yang
mempunya lebih dari satu kodon.
e. Secara umum, kodon bersifat hampir universal karena pada beberapa
organel jasad tinggi ada beberapa kodon yang berbeda dari kodon
yang digunakan pada sitoplasma (tabel 3)
Dalam proses translasi, setiap kodon berpasangan dengan
antikodon yang sesuai pada molekul tRNA. Sebagai contoh, kodon
metionin (AUG) mempunyai komplementer dalam bentuk antikodon UAC
yang terdapat pada tRNAmet. Pada waktu tRNA yang membawa asam
amino diikat ke dalam sisi A pada ribosom, maka bagian antikodonnya
berpasangan dengan kodon yang sesuai yang ada pada sisi A tersebut .
namun jika setiap kodon memerlukan tRNA tersendiri, maka diperlukan
banyak tRNA untuk mentranslasikan kode-kode genetik. Oleh keran itu
Francis Crick mengemukanan suatu hipotesis yang dikenal sebagai
hipotesis wobble yang menunjukkan bahwa tRNA yang diperlukan tidak
sebanyak jumlah kodon yang ada (Yuwono, T, 2005)
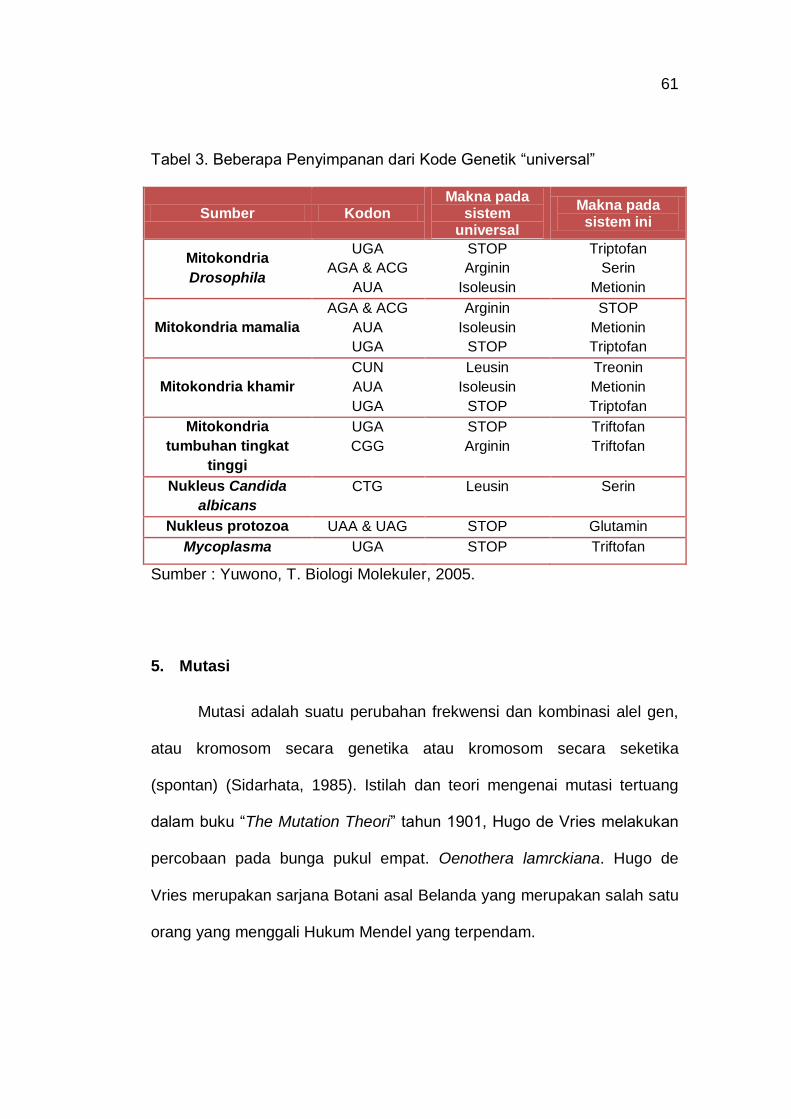
61
Tabel 3. Beberapa Penyimpanan dari Kode Genetik “universal”
Sumber Kodon Makna pada
sistem universal
Makna pada sistem ini
Mitokondria
Drosophila
UGA
AGA & ACG
AUA
STOP
Arginin
Isoleusin
Triptofan
Serin
Metionin
Mitokondria mamalia
AGA & ACG
AUA
UGA
Arginin
Isoleusin
STOP
STOP
Metionin
Triptofan
Mitokondria khamir
CUN
AUA
UGA
Leusin
Isoleusin
STOP
Treonin
Metionin
Triptofan
Mitokondria
tumbuhan tingkat
tinggi
UGA
CGG
STOP
Arginin
Triftofan
Triftofan
Nukleus Candida
albicans
CTG Leusin Serin
Nukleus protozoa UAA & UAG STOP Glutamin
Mycoplasma UGA STOP Triftofan
Sumber : Yuwono, T. Biologi Molekuler, 2005.
5. Mutasi
Mutasi adalah suatu perubahan frekwensi dan kombinasi alel gen,
atau kromosom secara genetika atau kromosom secara seketika
(spontan) (Sidarhata, 1985). Istilah dan teori mengenai mutasi tertuang
dalam buku “The Mutation Theori” tahun 1901, Hugo de Vries melakukan
percobaan pada bunga pukul empat. Oenothera lamrckiana. Hugo de
Vries merupakan sarjana Botani asal Belanda yang merupakan salah satu
orang yang menggali Hukum Mendel yang terpendam.

62
Setiap urutan DNA secara normal tidak akan berubah saat
bereplikasi. Namun, beberapa hal dapat menyebabkan urutan DNA itu
berubah. Perubahan ini disebut juga dengan mutasi. Widodo (2003)
menyebutkan bahwa mutasi merupakan kesalahan di dalam replikasi atau
perbaikan yang dapat memunculkan urutan (sequence) yang baru. Lebih
lanjut Campbell (2002) mengatakan bahwa mutasi adalah perubahan
materi genetik suatu sel atau virus.
Mutasi terjadi pada setiap sel, baik itu sel somatik atau germline.
Mutasi pada sel somatik tidak akan diwariskan. Oleh karena itu, dalam
hubungannya dengan evolusi maka hal tersebut tidak diperhitungkan.
Mutasi merupakan sumber variasi di dalam evolusi (Widodo dkk, 2003).
Ada beberapa macam mutasi yang dikenal sampai saat ini (widodo
dkk, 2003)
a. Berdasarkan tempat terjadinya
1. Mutasi kecil (point mutation)
Mutasi kecil adalah perubahan yang terjadi pada susunan molekul
(DNA) gen. Lokus gen sendiri tetap. Mutasi jenis ini yang menimbulkan
perubahan alel. Mutasi gen diartikan sebagai suatu perubahan fisiokimiawi
gen. Perubahan fisiokimiawi gen yang terjadi antara lain dapat berupa
perubahan atau pergantian pasangan basa. Misalnya pasangan A-T
diganti menjadi G-C.

63
2. Mutasi besar (gross mutation)
Mutasi besar adalah perubahan yang terjadi pada struktur
kromosom. Istilah khusus mutasi kromosom adalah aberasi.
b. Berdasarkan macam sel yang mengalami mutasi
1. Mutasi somatis (mutasi vegeratif)
Mutasi somatis adalah mutasi yang terjadi pada sel soma, bila
perubahan sel somatis demikian besar, sel-sel dapat mati dan kalau dapat
bertahan hidup memiliki kelainan atau tidak berfungsi secara normal.
2. Mutasi germinal (mutasi gametis/generatif)
Mutasi germinal adalah mutasi yang terjadi pada sel germinal
(terdapat didalam gonad). Hal ini terjadi pada mahluk hidup bersel banyak
dan bukan yang bersel satu.
c. Berdasarkan faktor penyebab mutasi
1. Mutasi alami (spontan)
Mutasi alam adalah mutasi yang terjadi sevara alami (tanpa dibuat
dan disengaja manusia). Penyebab dari mutasi alamiah antara lain: sinar
kosmis, batuan radioaktif, sinar ultraviolet matahari, radiasi ionisasi.
2. Mutasi buatan
Mutasi buatan merupakan mutasi yang sengaja dibuat oleh
manusia, yang biasa diarahkan kepada tujuan-tujuan tertentu. Misalnya
dibidang budidaya, perakitan bibit dan lain-lain.

64
d. Berdasarkan jumlah faktor keturunan
1. Mutasi bertahap (mutasi mikro)
Mutasi mikro adalah mutasi yang terjadi atas satu atau sekelompok
kecil faktor keturunan.
2. Mutasi lompatan (mutasi makro)
Mutasi makro adalah mutasi yang terjadi atas sejumlah besar atau
mungkin seluruh faktor keturunan.
e. Berdasarkan manfaat bagi individu atau populasi yang mengalami mutasi
1. Mutasi yang merugikan
Mutasi yang merugikan adalah mutasi yang berakibat timbulnya ciri
dan kemampuan yang kurang atau tidak adaptip pada individu (populasi)
2. Mutasi yang menguntungkan
Mutasi yang menguntungkan adalah mutasi yang berakibat
timbulnya ciri dan kemampuan yang semakin adaptip pada individu
(populasi), diantara kedua mutasi ini, yang paling banyak terjadi adalah
mutasi yang merugikan akan tetapi ada ruang lingkup mekanisme evolusi,
dampak perubahan karena mutasi efektif adalah mutasi yang
menguntungkan.

65
f. Berdasarkan panjang urutan DNA yang dipengaruhi oleh mutasi
1. Mutasi titik
Mutasi titik merupakan mutasi yang terjadi pada satu nukleotida
tunggal.
2. Mutasi segmental
Mutasi ini terjadi pada beberapa nukleotida.
g. Berdasarkan jenis perubahan yang disebabkan oleh peristiwa mutasi
1. Substitusi
Substitusi merupakan salah satu jenis mutasi yang disebabkan oleh
pergantian satu nukleotida dengan nukleotida yang lain. Menurut Fried
(2005) substitusi merupakan jenis mutasi yang lebih kecil
kemungkinannya dalam mengganggu sintesis protein. Pada substitusi
satu basa digantikan oleh basa lainnya. akibat adanya perubahan kodon
semacam ini adalah asam amino digantikan oleh asam amino lainnya. jika
asam amino yang baru mirip sifatnya dengan asam amino yang aslinya
tidak akan terjadi kerusakan.
Mutasi substitusi dibedakan atas transisi dan transversi. Transisi
adalah perubahan antara A dan G (purin) atau antara C dan T (pirimidin),
dengan kata lain transisi merupakan mutasi yang terjadi karena adanya
pergantian basa purin dengan purin lain, atau antara basa pirimidan
dengan pririmidin. Jenis transisi adalah A→G, G→A, C→T dan T→C.

66
Transversi adalah merupakan perubahan antara suatu purin dengan
pirimidin. Jenis transversi yaitu A→C, A→T, C→A, C→G, T→A, T→C,
G→C dan G→T.
Mutasi yang terjadi pada protein coding dapat pula dibedakan
menjadi beberapa menurut efek yang ditimbulkannya. Ada mutasi
synonymous jika tidak terjadi perubahan apapun pada asam amino yang
ditetapkan. Ada pula mutasi nonsynonymous jika terjadi perubahan asam
amino yang ditetapkan.
Lebih lanjut mutasi nonsynonymous dibedakan menjadi mutasi
missense dan mutasi nonsense. Mutasi missense mengubah kodon yang
dipengaruhi ke dalam suatu kodon asam amino yang kodenya telah
ditetapkan sebelumnya. Mutasi nonsense akan mengubah suatu sense
kodon kedalam sense kodon terminal sehingga translassi akan berakhir
dan menghasilkan protein yang tidak lengkap.
2. Rekombinasi
Selain mutasi, mekanisme lain yang dapat menyebabkan
terjadinyab variasi genetik adalah rekombinan. Rekombinan genetik
adalah proses pertukaran elemen genetik yang dapat terjadi antara
untaian DNA yang berlainan (interstrand) atau antara bagian-bagain gen
yang terletak dala satu untaian DNS (intrastand). Dengan katan lain
rekombinan genetik adalah penggabungan gen dari satu atau lebih sel ke
sel target. ada dua jenis rekombinan yaitu homolog pindah silang

67
(reciprocal recombination) dan konversi gen (nonresiprocal
receombination).
3. Delesi dan Insersi
Delesi dan insersi dapat terjadi karena beberapa mekanisme.
Mekanisme pertama adalah unequal crossing over. Unequal crossing over
antara dua kromosom mengakibatkan delesi suatu segmen DNA pada
satu kromosom dan suatu penambahan timbal balik pada yang lain.
Kesempatan terjadinya unequal crossing over sangat ditingkatkan jika
suatu segmen DNA disalin dalam tandem, sehingga kemungkinan salah
urutan adalah lebih tinggi.
Mekanisme berikutnya adalah delesi dalam rantai yang merupakan
suatu site-spesific recombination yang muncul ketika suatu segmen
verulang berpasangan dengan yang lain memiliki orientasi yang sama
pada ktomatid yang sama, maka sebagai konsekuensi adalah terjadinya
suatu pindah silang intrakromosom.
Mekanisme ketiga adalah replication slippage atau slipped-strand
mispairing. Peristiwa ini terjadi pada daerah DNA repeat/berulang yang
berdekatan. Selama DNA replikasi, slippage dapat terjadi oleh karena
mispairing antara daerah berulang yang berdekatan dan slippage ini dapat
menghasilkan delesi dan duplikasi segmen DNA tergantung kenampakan
slippage pada arag 5→3 atau sebaliknya. Mekanisme keempat yang
bertanggung jawab untuk insersi dan delesi sekuens DNA adalah DNA
transpisition

68
Delesi dan insersi secara bersama dikenal sebagai indels sebagai
singkatan untuk insersi atau delesi. Sebab ketika dua sekuens
dibandingkan, adalah mustahil untuk membahas apakah suatu insersi
telah terjadi atau delesi telah terjadi. (Levine J.S kenneth R.M, 1992)
Gambar 12. Mutasi Gen (Point Mutation) Sumber : http://books.lardbucket.org/books/the-basics-of-general-organic-
and-biological-

69
Gambar 13. Mutasi Kromosom Sumber : http://books.lardbucket.org/books/the-basics-of-general-organic-
and-biological-
E. TINJAUAN UMUM SITOKIN
1. Definisi Sitokin
Sitokin merupakan peptida pengatur (regulator) yang dapat
diproduksi oleh hampir semua jenis sel yang berinti di dalam tubuh, sitokin
dikelompokkan kedalam mediator yang berfungsi dalam komunikasi
antarsel. Sitokin tidak bergantung pada peredaran darah (parenkin dan
autokrin) hal tersebut dikarenakan sitokin mempunyai sel-sel sasaran
yang tidak terlalu jauh letaknya (Subowo, 2009; Boedina K.S, 2010).

70
Menurut Jan Vilcek (1998), sitokin adalah protein regulator yang
dilepaskan oleh sel-sel darah putih (leukosit) dan berbagai jenis sel lain
dalam tubuh, kegiatan pleiotropik sitokin mencakup efek pada sel dari
sistem imun dan modulasi respons radang (Subowo, 2009; Boedina K.S,
2010).
Kelompok sitokin dapat dibedakan dalam keluarga sebagai berikut :
Tabel 4. Kelompok Sitokin dalam Keluarga
Keluarga Perwakilan anggota
Interleukin-2/Interleukin-4 Interleukin-6/Interleukin-12 Interferon-α/β Tumor necrosis factor Interleukin-1 Transforming growth factor-β Chemokines
IL-2 IL-4 IL-5 GM-CSF IL-6 IL-12 IFN-α (banyak jenisnya) IFN-β IFN-ω IFN-ƫ TNFα TNF-β(LT-α) LT-β Fas ligand CD50 ligand TNF releated apoptosis inducing ligand IL-1α IIL-1β IL-1 Receptor antagonist IL-18 TGF-β Bone morphogenetic protein Inhibins Activins C-X-C sub-faily C-C subfamily C subfamily

71
Sitokin menurut Abbas dkk (2007) dapat diklasifikasikan
berdasarkan aktivitas biologik yang utama kedalam 3 kelompok (lihat
tabel 5), yaitu:
a. Mediator dan regulator imunitas bawaan
Kelompok sitokin ini terutama diproduksi oleh fagosit mononuklear
sebagai respon terhadap agen infeksi. PAMP’s seperti lipopolisakarida
(LPS), dsRNA virus, berikatan dengan TLR pada permukaan sel atau
dalam endosom makrofag dan merangsang sintesis dan sekresi beberapa
jenis sitokin sistem imun bawaan yang penting. Sitokin yang sama dapat
pula disekresi oleh makrofag yang diaktivasi oleh limfosit T yang di
stimulasi oleh antigen.
b. Mediator dan regulator imunitas dapatan
Kelompok sitokin ini diproduksi terutama oleh limfosit T sebagai
respons terhadap pengenalan spesifik antigen asing. Beberapa sitokin
yang diproduksi sel T berfungsi terutama untuk mengatur pertumbuhan
dan differensiasi beberapa populasi limfosit
c. Stimulator hemopoesis
Sitokin ini diproduksi oleh sel-sel stroma dalam sumsum tulang,
leukosit dan sel – sel lain dan merangsang pertumbuhan dan diferensiasi
leukosit imatur

72
Tabel 5. Gambaran Sitokin Imunitas Bawaan dan Dapatan
Imunitas bawaan Imunitas dapatan
Contoh Sumber utama Fungsi fisiologik utama Rangsangan Jumlah diproduksi Efek lokal/sistemik Peranan dalam penyakit Inhibitor
TNF, IL-1, IL-12, IFN- Makrofag, sel NK Mediator inflamasi (lokal dan sistemik) LPS(endotoksin), peptidoglikan bakteri, RNA virus, sitokin yang dihasilkan sel T (IFN-) Banyak (dapat dideteksi didalam serum keduanya Penyakit sistemik Kortikosteroid
IL-2, IL-4, IL-5, IFN-γ Limfosit T Mengatur pertumbuhan dan diferensiasi limfosit, aktivasi sel-sel efektor (makrofag, eosinofil, mastosit) Antigen protein Pada umumnya sedikit (tidak terdeteksi didalam serum) Biasanya hanya lokal Kerusakan jaringan lokal Siklosporin
2. TNF (Tumor Necrosis Factor)
Tumor Necrosis Factor (TNF) pada awalnya ditemukan pada tumor
tertentu yang mengalami perdarahan. TNF terutama dihasilkan oleh
makrofag dan sel-sel jenis lain dengan berbagai aktivitas biologik pada
sel-sel sasaran yang termasuk sistem imun maupun bukan (Subowo,
2009).
Tumor Necrosis Factor (TNF) merupakan mediator utama pada
respons inflamasi akut terhadap bakteri gram negatif, dan berperan dalam
respons imun bawaan terhadap berbagai mikroorganisme penyebab
infeksi yang lain, serta bertanggung jawab atas banyak komplikasi
sistemik yang disebabkan infeksi berat.

73
Berbagai efek TNF dengan dengan manifestasi sebagai berikut :
a) Efek sitotoksik
Efek sitotoksit terlihat pada beberapa jenis jaringan tumor yang
mengalami kemunduran dan nekrosis yang disertai perdarahan.
Mekanisme kematian sel tumor in vivo oleh TNF belum jelas, tetapi
yang jelas bahwa kematian sel tumor membutuhkan reseptor untuk
TNF. Kematian sel tumor akan dipercepat jika terdapat hambatan
sintesis protein dalam sel tumor. Tetapi mekanisme kematian sel
tumor secara in vivo bukan pengaruh langsung TNF melainkan secara
tidak langsung. Kemungkinan kematian sel tumor karena terjadinya
nekrosis jaringan tumor sebagai akibat gangguan vaskularisasi untuk
jaringan tumor.
b) Efek radang
TNF sekarang dikenal sebagai mediator utama dalam radang. Pada
percobaan dapat ditunjukkan bahwa TNF yang diperoleh dalam
bentuk murni secara biokimia ternyata bertanggung jawab kepada
aktivitas “cahectin” yang umumnya bekerja pada penderita yang
mengalami infeksi parasit.
c) Efek hemotopoietik
Efek TNF terhadap aktivitas hemotopoietik terlihat dalam bentuk
hambatan pembentukan koloni biakan granulosit-monosit, eritroid dan
koloni sel multi-potensial pada jaringan sumsum tulang manusia.

74
Tetapi sebaliknya pada mencit, TNF meningkatkan sel-sel progenitor
dalam jaringan sumsum tulang pada percobaan in vivo
d) Efek imunologik
Walaupun TNF dalam beberapa aktivitas biologik mirip IL-1, namun
ada beberapa perbedaan dalam mekanisme pengaturan imun. TNF
empunyai aktivitas perangsangan yang multipel terhadap limfosit T
teraktifkan, misalnya respons prolifetif limfosit T terhadap antigen,
peningkatan reseptor untuk IL-2 dan induksi produksi IFNγ. TNF dapat
meningkatkan ekspresi antigen MHC kelas I pada fibroblas dan sel
endotel (Subowo, 2009).
TNF berfungsi untuk meningkatkan ekspresi molekul adhesi yang
memudahkan leukosit melekat pada permukaan endotel, merangsang sel
fagosit mononuklear untuk mensekresi chemokine, serta mengaktifkan
leukosit. Dampak TNF secara sistemik adalah : 1) bersama-sama dengan
IL-1 TNF menimbulkan demam karena TNF berinteraksi dengan sel-sel di
daerah hypothalamus; 2) TNF dapat merangsang fagosit mononuklear
untuk memproduksi IL-1 dan IL-3; 3) merangsang hepatosit untuk
memproduksi protein-protein tertentu; 4) mengaktifkan sistem koagulasi
dengan merubah keseimbangan aktivitas prokoagulan dan antikoagulan
pada endotel vaskuler; 5) menekan aktivitas stem cell dalam sumsum
tulang (Boedina K.S, 2010)
Ada 2 bentuk TNF yaitu TNF-α dan TNF-β . TNF-α diproduksi oleh
berbagai jenis sel termasuk makrofag, sel T, B, NK, astrosit dan Kupfer.

75
Pembentukan terjadi sebagai respon terhadap rangsangan bakteri, virus
dan sitokin, kompleks imun, komponen komplemen C5a dan reactive
oxygen intermediates (ROI). Sebaliknya TNF-β disekresi oleh sel T dan B
teraktivasi, dapat berada dipermukaan sel bila terikat dengan
transmembran LT-β. TNF-α terbukti merupakan modulator respon imun
kuat yang diperantarai induksi molekul adhesi, sitokin lisin dan aktivasi
neutrofil (Subowo, 2009; Boedina K.S, 2010).
F. TINJAUAN KHUSUS ELISA
Pada pengembangan teknologi diagnostik maupun penelitian
banyak pilihan metode yang dapat digunakan sebagai acuan untuk
mendapatkan hasil yang optimal dalam mengukur kadar antibodi didalam
tubuh. ELISA (Enzyme-linked Immunosorbent Assay) merupakan salah
satu metode yang sensitif untuk mendeteksi antibodi, antigen, hormon
maupun bahan toksik (Rantam, F.A, 2003; Handojo, I, 2003).
Metode ELISA pertama kali di perkenalkan oleh Engvall dan
Pelmann (1971) dengan cara mengkonjugasikan enzime dalam
immunoassay. Karena tingkat sensitivitasnya tinggi terutaman untuk tes
serologi pada awal infeksi. Dua macam antibodi yang digunakan dalam
ELISA yaitu antibodi pertama (primary antibody) mengikat pada antigen
dan antibodi kedua (secundary antibody) atau antibodi antiglobulin
mengikat pada antibodi pertama. Antiglobulin ini yang dilabel dengan
enzim seperti horseradish peroxidase, alkali phosphatase yang

76
mempermudah monitor dengan perubahan warna. Adanya reaksi dari
enzim ini secara kuantitatif antibodi pertama dapat dianalisis (Rantam,
F.A, 2003; Handojo, I, 2003).
Berdasarkan pada perkembangan teknologi, maka model ELISA
berkembang antara lain direct ELISA, indirect ELISA, sandwich ELISA,
cupture ELISA, sel ELISA.
1. Direct ELISA
Direct ELISA merupakan salah satu model ELISA yang langsung
diikatkan antara antigen dan antibodi, dimana antibodi harus dilabel
dahulu baru divisualisasikan dengan cara manambahkan substrat.
Kelemahan metode ini adalah diperlukan keahlian dalam melakukan
konjugasi atau melabel antibodi dengan enzim, sehingga tahap ini
memerlukan tahapan purifikasi antibodi dan setelah itu dilanjutkan dengan
pelabelan (Rantam, F.A, 2003; Handojo, I, 2003).
Gambar 14. Direct ELISA
Sumber : www.piercenet.com

77
2. Inderct ELISA
Model ini banyak digunakan di laboratorium, hal tersebut
dikarenakan model ini tidak memerlukan keahlian khusus untuk konjugasi.
Model ini telah dilengkapi dengan konjugat fragmen imunoglobulin anti
imunoglubolin yang akan dideteksi. Model lain yang lebih spesifik yaitu
model indirect ELISA model Fang antibodi, dasar mikroplate telah dilapisi
dengan antibodi selanjutnya ditambahkan antigen dan diinkubasi dalam
waktu 45 menit pada suhu 37 ºC yang selanjutnya ditambahkan antibodi
monoklonal dan akhirnya ditambahkan substrat yang dilabel dengan
enzim alkali fosfatase atau peroksidase (Rantam, F.A, 2003; Handojo, I,
2003).
Gambar 15. Indirect ELISA
Sumber : www.piercenet.com
3. Sandwich ELISA
Sandwich ELISA adalah model tes ELISA yang menggunakan
perangkat tiga macam antibodi. Antibodi pertama biasanya menggunakan
antibodi monoklonal yang dilapiskan pada dasar mikroplate dan

78
selanjutnya direaksikan dengan antigen. Setelah dilakukan pencucian
baru ditambahkan antibodi kedua atau sampel serum yang akan dideteksi
dan selanjutnya direaksikan dengan antibodi ketiga yaitu fragmen
imunoglobulin anti imunglobulin yang akan dideteksi. Model ini memiliki
tingkat sensitivitas dan spesifitas yang lebih tinggi dibanding kedua
metode sebelumnya, sehingga sering digunakan untuk mendeteksi
antigen antibodi pada sampel yang mempunyai konsentrasi rendah
(serum, plasma, cairan serebrospinal, cairan ludah, sekresi air mata)
(Rantam, F.A, 2003; Handojo, I, 2003).
Gambar 16. Sandwich ELISA Sumber : www.piercenet.com
4. Cupture ELISA
Cupture ELISA adalah model yang dikembangkan untuk deteksi
IgM karena untuk mendeteksi IgM sering terjadi faktor negatif yaitu adanya
faktor reumatoid, sehingga sering terjadi positif palsu. Untuk menghindari
hal tersebut maka yang menjadi pelapis (coating) adalah Imunoglobulin M
anti human atau spesies lain. Dengan demikian faktor negatif dapat

79
dihindari karena ikatan spesifik terjadi antara serum yang dideteksi
imunoglobulinnya dengan anti-IgM (Rantam, F.A, 2003; Handojo, I, 2003)
5. Sel ELISA
Sel ELISA dikembangkan untuk mendeteksi antigen atau agen
yang terdapat didalam sel. Sehingga pada model ini tidak diperlukan
pelapisan antigen pada mikroplate tetapi dengan cara fiksasi sel yang
diinokulasikan sampel yang yang dideteksi agennya, kemudian
direaksikan dengan konjugat fragmen imunoglobulin anti imunoglobulin
yang digunakan untuk mendeteksi antigen. Antibodi yang sering
digunakan adalah antibodi monoklonal (Rantam, F.A, 2003; Handojo, I,
2003).
G. KERANGKA KONSEPTUAL
Beberapa hasil penelitian yang mendukung kerangka kosep
penelitian seperti yang dijelaskan di bawah ini :
1. WHO., 2012 : WHO memperkirakan bahwa pada tahun 2011 ada 9
juta kasus TB baru di dunia (125 kasus per 100.000 populasi) dan 1.4
juta meninggal karena infeksi tuberkulosis.
2. WHO., 2012 : Tahun 2011 Indonesia merupakan negara ke 4 yang
memiliki prevalensi penyakit tuberkulosis.

80
3. Johanneke Kleinnijenhuis et al., 2010 : sistem imun bawaan
memainkan peran penting dalam pertahanan tubuh terhadap infeksi
MTB. Beberapa kelas PPR yang terlibat dalam recognition MTB oleh
sistem imun bawaan diantaranya TLRs, CLRs, dan NLRs. Beberapa
keluarga TLR seperti TLR2, TLR4, dan TLR9 serta molekul adaptor
MyD88 berperang lebih penting dalam inisiasi respon imun terhadap
tuberkulosis. Studi epidemologi juga dilakukan dimana
mengungkapkan bahwa terjadinya variasi genetik pada anggota PPR
mempengaruhi tingkat kerentanan penyakit, tingkat keparahan dan
juga berkontribusi pada penciptaan terapi yang baru dan lebih rumit.
4. Y.X Zhang et al., 2011 : Adanya polimorfisme pada gen TIRAP/
MyD88 memungkinkan terjadinya kerentanan terhadap penyakit TB
dan dapat menyebabkan faktor resiko baru terhadap perkembangan
TB. Pada infeksi yang disebabkan oleh TB terdapat dua signal adaptor
TLR4 yang dilalui yaitu MyD88 dan TRIF.
5. Van Lieshout MH et al., 2012 : Penelitian yang dilakukan dengan
tujuan untuk mengetahui peranan dari 2 signal adaptor TLRs yaitu
MyD88 dan TRIF pada pasien dengan kasus hematopoetik sel dan
pneumonia. Dimana MyD88 pada kedua kasus memiliki kontribusi
dalam kelangsungan dan pertahanan tubuh terhadap infeksi bakteri
sedangkan TRIF hanya berkontribusi pada kasus hematopoetik sel.
MyD88 dan TRIF memiliki kontribusi untuk pertahanan tubuh terhadap
berbagai macam infeksi.

81
6. Takeuchi O et al., 2000 : TLR2 mengenali komponen bakteri gram
positif dan TLR4 mengenali komponen LPS gram negatif sedangkan
MyD88 merupakan molekul adaptor yang penting untuk signal TLR.
Tikus dengan defisiensi TLR2 dan MyD88 diinfeksikan dengan
Staphylococcus aureus, dimana tikus dengan defisiensi MyD88 sangat
rentan terhadap S.aureus dengan produksi sitokin tidak ada sama
sekali.
7. Shizuo akira et al., 2003 : TLRs berperan penting dalam mendeteksi
patogen yang menyeran tubuh. TLRs mengenali komponen yang
berbeda yang berasal dari patogen yang diikuti oleh produksi sitokin.
Setelah terjadi stimulasi TLR merekrut IL-1R- associated kinase via
adaptor MyD88 dan menginduksi aktivasi dari NF-kB dan mitogen-
activated protein kinase. Produksi sitokin akan menurun jika pada
MyD88 terjadi kecacatan sehingga dikatakan MyD88 merupakan
signal molekul dari IL-1R yang penting. Pada TLR4 telah ditemukan
molekul baru yang diidentifikasi dapat berpartisipasi pada jalur
MyD88-independent yang dapat menginduksi interferon tipe I (IFNs)
jika terjadi cacat / penurunan fungsi pada MyD88 dependent.
8. Kawait T et al., 1999 : MyD88 sangat penting untuk respon LPS,
namun kemampuan untuk menginduksi LPS tergantung pada ekspresi
gen.
9. Sugawara I et al., 2003 : MyD88 berfungsi untuk memediasi jalur
signal TLR untuk mengaktifkan NF-kB dalam pertahan tubuh terhadap

82
infeksi bakteri terutama M. tuberculosis . Tikus dengan MyD88 (-/-)
sangat rentang terhadap infeksi MTB serta menyebabkan lesi paru
granulomatosa yang lebih luas dibandingkan pada tikus normal.
10. Feng CG et al., 2003 : MyD88 mengatur pertahan tubuh terhadap
infeksi mikobakteria dan memiliki peran dalam menentukan induksi
sitokin proinflamasi dan IFN gamma. Penelitian dilakukan pada tikus
dengan defisiensi MyD88 dan sebagian TLR2 (-/-) dan TLR4 (-/-)
terinfeksi strain M. Avium yang virulen. Dimana MyD88 (-/-) memiliki
kerentanan yang lebih parah dan kelangsungan hidup yang lebih
singkat dibandingan dengan TLR2(-/-) dan TLR4 (-/-) dan memiliki
beban infeksi yang lebih berat terhadap infeksi bakteri serta respon
pembentukan sitokin sangat rendah dan tidak jelas.
11. Eur Bernuth H et al., 2012 : Tikus dengan defisiensi MyD88 dan IRAK-
4 terbukti rentan terhadap 27 jenis bakteri patogen, 8 jenis virus, 17
jenis parasit dan 4 jenis jamur. Pada manusia dengan defisiensi
MyD88 dan IRAK-4 mengalami infeksi dari sejumlah spesies bakteri
walaupun akan menurun seiring bertambahnya usia sehingga dapat
disimpulkan bahwa mutasi MyD88 menyebabkan terjadi berbagai
macam infeksi yang disebabkan oleh mikroorganisme patogen.
12. Chen M et al, 2002 : Penelitian dilakukan pada tikus, dimana tikus
yang defisiensi pada MyD88 menyebabkan 95% meninggal pada hari
ke 10 – 16 dan 95% – 100% tikus yang defisiensi pada TLR2/4
mampu bertahan selama > 7 minggu.

83
13. Terence M et al,. 2004 : MyD88 (-/-) mengganggu satu atau lebih
langkah dalam proses seperti fagositosis, namun sekarang telah
diteliti bahwa pada TLR4 terdapat jalur MyD88-independent yang
mungkin dapat memediasi beberapa komponen dan proses imun.
14. Cecille M et al,. 2004 : Tikus dengan MyD88 (-/-) terinfeksi MTB,
dimana tikus dengan MyD88 (-/-) akan mati dalam 4 minggu,
pemberian vaksin BCG untuk perlindungan dari infeksi MTB belum
meningkatkan respon imun terhadap infeksi MTB bahkan signal
MyD88 dihilangkan dengan tujuan untuk menaikkan kekebalan
terhadap respon imun namun hasilnya belum cukup untuk
mengimbangi infeksi. Sehingga disimpulkan bahwa tikus dengan
MyD88 (-/-) tidak mampu mengendalikan infeksi MTB.
15. Beze F et al., 2012 : MyD88 bertanggung jawab terhadap kerentanan
bakteri patogen pada infeksi pernapasan dan MyD88 merupakan
faktor penting untuk kekebalan tubuh terhadap bakteri dan merupakan
target untuk sasaran terapi/ pengobatan.
16. Je EM et al., 2012 : Penelitian dilakukan dengan tujuan untuk melihat
apakah perubahan dari ekpresi protein MyD88 dan mutasi somatik
pada MyD88 berakibat pada keparahan kanker. Hasil penelitian
mengemukakan ekspresi MyD88 meningkat pada sel kanker lambung
dibandingkan dengan sel normal sedangkan menurun pada kanker
kolorektal. Tidak ada mutasi somatik dari gen MyD88 di lambung,
kolorektal, payudara, hepatoseluler, prostat dan karsinoma paru-paru.

84
Kesimpulan bahwa ekspresi MyD88 dalam sel normal berbeda
tergantung pada organ. Data menunjukkan bahwa keuntungan dari
ekspresi MyD88 dalam kanker lambung mungkin memainkan peran
dalam patogenesis kanker dengan mengaktifkan onkogenik
fungsi MyD88.
17. Cecile M et al., 2007 : Respon MyD88 dependent pada sistem host
mengendalikan infeksi akibat M. tuberculosis. Pada kasus MyD88 (-/-)
menyebabkan produksi sitokin berkurang dan masa hidup berkurang
bahkan menyebabkan beban infeksi karena M. tuberculosis
bertambah.
18. Muraille E et al., 2003 : Tikus tanpa MyD88 tidak memiliki proteksi
terhadap penyakit infeksi dan berpengaruh terhadap perkembangan
respon Th2.
19. Nicolle DM et al., 2004 : Tikus dengan defisiensi MyD88 diinjeksikan
dengan BCG bovis dimana kadar sitokin proinflamasi sangat rendah
namun costimulator seperti CD40 dan CD84 stabil dengan
kemampuan untuk merespon sel T masih efisien, namun berakibat
pada kejadian infeksi pneumonia yang kronik.
Berdasarkan pemaparan diatas, maka yang dapat dijadikan obyek
penelitian adalah mengetahui ada tidaknya mutasi pada gen MyD88 di
penderita tuberkulosis paru.
Adapun kerangka konseptual secara skema adalah, sebagai
berikut :

85
Gambar 17. Kerangka Konsep
Mycobacterium
tuberculosis
Penderita
Tuberkulosis
Respon Imun Spesifik
Toll Like
Receptor 4(TLR4)
Myeloid Differentiation
Primary Factor 88
(MyD88)
Induksi NF-kB
Produksi Sitokin
Mutasi
Tidak
ada
Mutasi

86
H. DEFINISI OPERASIONAL
1. Tuberkulosis paru adalah penyakit infeksi paru yang disebabkan oleh
Mycobacterium tuberculosis yang diagnosisnya ditegakkan
berdasarkan hasil pemeriksaan fisik, pemeriksaan BTA, kultur, dan
foto thorax.
2. Mutasi MyD88 adalah keadaan dimana terjadi transisi satu atau lebih
basa nukleotida (A, T, C, atau G) yang menyebabkan berubahnya
urutan asam amino pada gen MyD88 sehingga menghasilkan pola
DNA baru.
3. Polymerase Chain Reaction adalah merupakan suatu proses sintesis
enzimatik untuk mengamplifikasi fragmen DNA secara in vitro.
4. DNA Sequencing adalah suatu metode yang digunakan untuk
menentukan urutan basa-basa nukleotida suatu fragmen DNA.
5. Exon adalah urutan nukleotida yang mengkode asam amino yang
membawa informasi genetik.
6. Intron berasal dari singkatan intragenic regions, yang merupakan
bagian yang tidak dikode oleh prekursor mRNA (pre-mRNA) yang
dibuang sebelum mRNA siap ditranslasi.
7. ELISA (Enzyme-linked Immunosorbent Assay) merupakan metode
yang digunakan untuk mendeteksi antigen dan antibodi.
8. TNF merupakan mediator utama pada respons inflamasi akut
terhadap bakteri gram negatif dan berperan dalam respon imun
bawaan terhadap berbagai mikroorganisme penyebab infeksi.

87
I. KERANGKA TEORI
Penelitian ini didasari oleh teori dari jalur signaling TLR (TLR2/4)
dalam mengaktivasi respon imun terhadap infeksi yang disebabkan oleh
mikroba patogen (M.tuberculosis). adapun kerangka teori tersebut
dijabarkan dalam bagan berikut :
Gambar 18. Kerangka Teori
MyD88-dependent pathway
MyD88-independent pathway
M.Tuberculosis component : LAM, LM, PIMs, STF, 19-kDa Lipoprotein,
MTB HSP65
Activates :
TLR2
TLR4
Signaling
MyD88
Activating
TIRAP
Inducing NF-kB
Promote production of
inflamantory cytokine
Signaling TRAM
Activating TRIF
Inducing IRF3
Eliminate Bacteria &
Other patogen
Promote production of
IFN type I
- IFN beta - IFN alpha
- TNF alpha - IL-6 - IL-12 - IL-8 - IL-1β - RANTES - MIP1β - MIP1α
Faktor Genetik
Influence to

88
BAB III
METODOLOGI PENELITIAN
A. Rancangan Penelitian
Penelitian ini merupakan penelitian yang bersifat deskriptif dengan
menggunakan pendekatan cross sectional study untuk mengetahui mutasi
gen MyD88 dan kadar TNF-α pada penderita tuberkulosis paru.
B. Waktu dan Tempat Penelitian
Pengumpulan sampel penderita tuberkulosis dilakukan di Balai
Besar Kesehatan Paru Masyarakat (BBKPM) dan RS. Labuang Baji
Makassar. Penelitian dilakukan di Unit Penelitian FK Unhas Makassar pda
bulan Maret s.d Juni 2013.
C. Populasi, Besaran Sampel Penelitian
1. Populasi Penelitian
Penderita tuberkulosis yang melakukan pemeriksaan di Balai Besar
Kesehatan Paru Masyarakat (BBKPM) Makassar dan RS. Labuang Baji
Makassar
2. Sampel Penelitian
Sampel adalah sebagian atau wakil dari populasi yang diteliti.
Sampel sebanyak empat (4) orang yang dipilih secara metode non
random sampling yang telah memenuhi kriteria inklusi dan ekslusi. Dari

89
RS. Labuang Baji Makassar dipilih dua (2) sampel dengan status MDR
dan XDR TB dan dari BBKPM dipilih dua (2) sampel dengan status sensitif
TB.
3. Kriteria Sampel
a. Kriteria inklusi
1) Sampel yang berasal dari penderita tuberkulosis paru baik dengan
status sensitif OAT (Obat anti tuberkulosis), MDR (Multi Drug
Resistant) maupun XDR (Extensively Drug Resistant), yang
ditegakkan berdasarkan hasil pemeriksaan sputum BTA (positif),
rontgen (positif), pemeriksaan kultur (tumbuh/positif MTB) dan uji Drug
Susceptibility Test (Sensitif/Resisten).
2) Bersedia ikut penelitian dengan menandatangani informed concent
yang telah dikeluarkan oleh Komisi Etik Fakultas Kedokteran
Universitas Hasanuddin.
b. Kriteria eksklusi
1) Menderita penyakit tuberkulosis dengan komplikasi HIV/AIDS yang
ditegakkan dengan pemeriksaan HIV metode rapid test.
2) Sampel darah lisis.

90
D. Alat dan Bahan Penelitian
1. Alat Penelitian
Mikropipet 100-1000 μL (Eppendorf, German), Mikropipet
10-100 μL (Eppendorf, German), mikropipet 0-10 μL (Eppendorf,
German), sentrifuge (Thermo, German), mikropipet tips, conical tube
15 mL, tube 1.5 mL, tube 0.2 μL, rak tabung, Gel doc XR System (BioRad,
USA), Power pac, Vortex, perangkat elektroforesis (BioRad, USA), Mesin
PCR (Applied Biosystems, USA), Bio Safety Cabinet (ESCO), Elisa
Reader (BioRad, USA).
2. Bahan Penelitian
Darah segar (Antikoagulan EDTA-K3), Alkohol, Handskun, Masker,
Vacutainer set, Tabung EDTA, Pipet volumetri steril, Kit DNA ekstraksi
(Genomic DNA Mini Kit, Geneaid), PCR Mix (Kapa Fast 2G), Primer
MyD88 (Tabel terlampir), Agarosa 2% (Invitrogen), Marker 100 bp
(Generuler, German), Etidium bromida, QIAquick Gel Extraction Kit
(QIAGEN, Hilden, Germany), Proteinase K (Geneaid), Etanol absolut, TBE
(Tris-Borate-EDTA), Kit ELISA TNF-α (R&D System, USA).

91
E. Cara Kerja
1. Alokasi subyek
Penelitian dilakukan pada semua penderita tuberkulosis yang
memenuhi kriteria inklusi yang menjalani pemeriksaan di Balai Besar
Kesehatan Paru Masyarakat Makassar, RS. Labuang Baji Makassar.
2. Cara penelitian
a. Melakukan pencatatan identitas penderita yang memenuhi kriteria
inklusi, diberikan penjelasan lengkap mengenai apa yang akan
dilakukan terhadap mereka dan bila setuju mereka akan mengisi dan
menandatangani informed concent.
b. Pasien menjalani pemeriksaan bakteriologi dengan sampel sputum
c. Pasien menjalani pemeriksaan skrining HIV dengan sampel
plasma/serum.
d. Pasien menjalani pemeriksaan PCR dan PCR sequencing dengan
sampel darah.
e. Pengumpulan Spesimen :
1) Sputum : Diambil sebanyak 3 kali selama 2 hari yang dikenal
dengan istilah S-P-S (sewaktu – Pagi – Sewaktu)
- Sewaktu (Hari pertama) Dahak pasien diambil di laboratorium
sewaktu penderita datang pertama kali.

92
- Pagi (Hari kedua) Sehabis bangun tidur keesokan harinya,
dahak pasien ditampung dalam pot kecil yang kemuadian
dibawah ke laboratorium.
- Sewaktu (Hari kedua) Dahak pasien diambil lagi di laboratorium
saat penderita datang pada hari kedua
2) Darah segar : Subyek penelitian yang memenuhi kriteria inklusi,
dilakukan pengambilan sampel darah EDTA sebanyak 3 cc, untuk
pemeriksaan PCR
3. Prosedur kerja
a. Pewarnaan Ziehl Nelseen
1) Pembuatan hapusan dahak
Ose dipanaskan diatas nyala api spritus sampai merah dan
dibiarkan sampai dingin, lalu bagian dahak yang kental dan berwarna
kuning kehijau-hijauan diambil dan dioleskan secara merata diatas
kaca sediaan, kemudian sediaan dikeringkan di udara terbuka, jangan
biarkan terkena sinar matahari langsung, setalah itu kaca sediaan
difiksasi diatas nyala api spritus dan sediaan yang telah difiksasi
disimpan kedalam kotak sediaan.
2) Pewarnaan sediaan dengan Ziehl Neelsen
Sediaan yang telah difiksasi diletakkan diatas diatas rak
dengan hapusan menghadap keatas, sediaan ditetesi dengan Carbol
Fuchsin 0.3% hingga menutupi seluruh permukaan sediaan, setelah

93
itu sediaan dipanaskan diatas nyala api spritus selama 3-5 menit,
dimana zat warna pertama tidak boleh mendidih. Kemudian sediaan
didiamkan selama 5 menit dan setelah itu sediaan dibilas dengan air
mengalir sampai zat warna yang bebas terbuang, setelah itu sediaan
ditetesi dengan asam alkohol 3% sampai warna merah fuhsin hilang,
selanjutnya sediaan dibilas dengan air mengalir, kemudian sediaan
dietetesi dengan larutan Methylen Blue 0.3% hingga menutupi seluruh
permukaan sediaan. Selanjutnya sediaan dikeringkan diatas rak
pengering di udara terbuka dan setalah kering sediaan dibaca dibawa
mikroskop dengan pembesaran 100x.
b. Extraksi DNA “Genomic DNA Mini Kit”
Extraksi DNA dilakukan sesuai dengan prosedur dari Genomic DNA
Mini Kit (Geneaid). Sebanyak 300 μL darah segar (EDTA-K3) dan 3x
volume sampel RBC Lysis Buffer (900 μL) dicampurkan kedalam tube 1.5
mL (jangan di vortex) kemudian diinkubasi selama 10 menit pada suhu
ruangan. Campuran tersebut disentrifuge kecepatan 3.000 x gravity
selama 5 menit untuk memisahkan pelet dengan supernatan, selanjutnya
supernatan dibuang. Tambahkan 100 μL RBC Lysis Buffer kedalam
suspensi pelet dan campurkan 200 μL GB buffer dengan menggunakan
pipet kemudian inkubasi selama 10 menit, suhu 65ºC, dan goyangkan
setiap 3 menit sekali. Campurkan suspensi tersebut dengan 200 μL
etanol absolute, campur selama 10 detik. Sebanyak 600 μLdimasukkan ke

94
dalam column kemudian disentrifugasi pada kecepatan 16.000 x gravity
selama 5 menit, sehingga larutan keluar dari column dan masuk kedalam
tabung penampung kemudian dibuang, sementara DNA tetap bertahan di
dalam column. Ganti tabung penampung dengan tabung penampung baru
kemudian tambahkan 400 μL W1 Buffer dan sentrifugasi pada kecepatan
16.000 x gravity selama 30 detik, kemudian dibuang. Tambahkan 600 μL
Wash Buffer sentrifugasi pada kecepatan 16.000 x gravity selama 30 detik
lalu buang supernatan dan masukkan kembali column kedalam tabung
penampung. Sentrifugasi kembali dengan kecepatan 16.000 x gravity
selama 3 menit untuk membuang sisa larutan pencuci yang mungkin
masih tertinggal dalam column. Tabung penampung kemudian diganti
dengan tabung eppendorf 1.5 mL steril. Untuk melepaskan DNA yang
terperangkap dalam column ditambahkan 100 μL larutan Elution Buffer
yang telah dipanaskan pada suhu 70 ºC, kemudian sentrifugasi pada
kecepatan 16.000 x gravity selama 30 detik sehingga DNA akan
tertampung dalam tabung steril.
c. Amplifikasi DNA
Amplifikasi atau penggandaan fragmen DNA dilakukan dengan
teknik PCR menggunakan primer spesifik yang hanya dapat
mengamplifikasi DNA MyD88 pada region yang diinginkan.
Primer yang digunakan terdiri dari 13 fragmen yang masing-masing
fragmen akan mengamplifikasi daerah yang sesuai dengan panjang
fragmen berbeda-beda tergantung jenis primernya (Lampiran 1).

95
Proses PCR menggunakan Veriti 96 well PCR (Applied Biosystems,
USA) dengan volume reaksi 25 μL yang mengandung 2,6 μL H2O PCR
grade, 10 μL 2X Kapa Ready Mix, 0,4 μL MgCl2 (25mM), primer forward
dan reverse masing-masing 1 μL, 1 μL DMSO dan DNA hasil ekstraksi
sebanyak 6 μL. Tahap siklus PCR terdiri dari denaturasi, annealing dan
ekstensi dilakukan pada suhu 95 ºC selama 15 detik, * tergantung primer
(Lampiran 1) selama 15 detik dan 68 ºC selama 15 detik dengan 30 siklus.
Siklus diawali dengan tahap initial denaturasi pada suhu 95 ºC selama 3
menit dan diakhiri dengan tahap cooling 14 ºC selama waktu tidak
ditentukan.
d. Elektroforesis gel agarosa
Untuk mengetahui hasil amplifikasi DNA dilakukan proses
elektroforesis terhadap produk PCR pada gel agarosa 2%. Sebanyak 2
gram bubuk agarosa dicampur kedalam 100 μL larutan TBE (tiap liter TBE
mengandung 54,9 g Tris base, 27.5 g asam borat dan 20 mL 0.5 mM
EDTA) kemudian dimasak menggunakan microwave sampai larut dengan
sempurna. Sebanyak 3 μL larutan etidium bromida (10 mg/mL)
ditambahkan kedalam gel agarosa yang masih cair dan dicampur dengan
menggunakan shaker hingga merata, kemudian gel dituangkan ke dalam
cetakan gel yang telah disiapkan yang sebelumnya di pasangkan sisir
diukur keseimbangannya. Gel dibiarkan hingga membeku, kemudian sisir
diangkat sehingga terbentuklah sumur-sumur gel. Gel agarosa bersama

96
dengan cetakannya diangkat kemudian dipindahkan ke dalam container
elektroforesis (BioRad, USA). Larutan TBE kemudian ditambahkan ke
dalam container elektroforesis hingga menutupi permukaan atas gel.
Kemudian kedalam masing-masing sumur, dimasukkan 8 μL produk PCR
karena menggunakan Kapa Ready Mix loading dye sehingga tidak perlu
penambahan loading dye pada saat produk PCR dimasukkan kedalam
sumur agarosa. Pada kedua ujung sumur dimasukkan 5 μL (0.5 μg)
ladder 100 bp sebagai standar ukur DNA. perangkat elektroforesis
dijalankan dengan mengalirkan aliran listrik bertegangan 100 Volt (V) 400
mA selama 75 menit.
Setelah proses elektroforesis gel agarosa selesai, gel kemudian
diangkat dari container dan dimasukkan ke dalam perangkat Gel Doc XR
(BioRad, USA) dan kemudian disinari dengan sinar ultraviolet (UV).
Gambar gel yang terbentuk dibawah sinar UV lalu didokumentasikan.
Hasil positif ditunjukkan apabila terbentuk pita DNA (band) yang
ukurannya sesuai dengan ukuran produk PCR yang diharapkan.
e. Purifikasi gel Extraksi
Pemurnian fragmen DNA hasil amplifikasi dilakukan pada hasil
produk PCR yang mengandung pita DNA nonspesifik, pemurnian
dilakukan dengan cara gel cut extraction. Dibuat gel agarosa seperti cara
diatas dengan menggunakan agarosa dengan konsentrasi 1,5%
menggunakan sisir ukuran besar menampung 40 μL produk PCR. Semua

97
produk PCR yang mengandung pita DNA nonspesifik dimasukkan ke
dalam sumur gel. Elektroforesis dijalankan dengan mengalirakn aliran
listrik bertegangan 100 Volt (V) 400 mA selama 80 menit. Selanjutnya
dibawah sinar UV, dengan membandingkan dengan hasil pita DNA
dengan ladder, gel yang mengandung pita DNA yang sesuai dipotong
dengan menggunakan pisau scalpel dan dimasukkan kedalam tabung
eppendorf 1.5 mL yang beratnya sudah diketahui. Tabung yang telah
berisi potongan gel ditimbang kembali untuk mengetahui selisi berat
antara tabung kosong dan tabung berisi. Potongan gel kemudian di
purifikasi gel cut extraction (QIAquick Gel Extraction Kit) dengan cara
potongan gel ditambahkan dengan larutan buffer QG sebanyak 3x volume
dari selisi berat tabung dan disertai dengan inkubasi pada suhu 50 ºC
selama 10 menit dan tiap 2-3 menit sekali divortex. Selanjutnya
ditambahkan isopropanol sebanyak 1x volume dari selisi berat. Larutan
dipindahkan ke dalam column yang telah dipasangkan pada tabung
penampung, kemudian di sentrifugasi selama 1 menit dengan kecepatan
8.000 revolution per menit (rpm). Cairan yang tertampung pada tabung
penampung kemudian dibuang setalah itu tambahkan 500 μL larutan QC
kemudian sentrifugasi 8.000 rpm selamat 1 menit dan kembali dibuang
yang tertampung. Selanjutnya tambahkan 750 μL buffer PE dan
sentrifugasi kembali dengan kecepatan 13.000 rpm (10.000 x gravity)
selama 1 menit. Untuk langkah selanjutnya pindahkan column kedalam

98
tabung eppendorf 1.5 mL dan tambahkan 50 μL buffer EB (10mM Tris-Cl,
pH 8.5) dan sentrifugasi selama 1 menit dengan kecepatan 13.000 rpm.
f. Pemeriksaan Sequencing
Sequencing dilakukan di Makrogen di Korea dengan
menggunakan metode direct sequencing. Hasil sequencing dalam bentuk
elektroferogram diunduh dan kemudian dilakukan editing dan analisis
menggunakan perangkat lunak BioEdit Sequence Alignment Editor versi
7.0.5.2 dan FinchTv versi 1.4.0.
Untuk mengetahui apakah hasil sekuens membawa mutasi pada
region tertentu, sekuens DNA target disejajarkan dengan sekuens DNA
dari gen MyD88 normal yang diunduh dari GeneBank (NC_000003)
sebagai konsensus
g. Pemeriksaan ELISA
Pengukuran TNF-α dilakukan sesuai prosedur dari Quantikine
ELISA (R&D System, USA). Pertama reagen, standar dan sampel di
siapkan, selanjutnya kedalam masing-masing well dimasukkan 50 μL
reagen Assay Diluent RD1F kemudian ditambah 200 μL Standar dan
sampel kedalam masing-masing sumur sesuai posisi kemudian tutup
sumur dengan strip adhesive dan diinkubasi selama 2 jam pada suhu
ruang. Aspirasi masing-masing sumur dan cuci dengan 400 μL Wash
buffer sebanyak 4X. Selanjutnya tambahkan 200 μL conjugate TNF-α

99
kedalam masing-masing sumur dan tutup dengan strip adhesive kemudian
inkubasi kembali selama 2 jam pada suhu ruang. Aspirasi dan cuci
dengan 400 μL Wash buffer sebanyak 4X. Tambahkan 200 μL Substrat
Solution kedalam masing-masing sumur dan inkubasi selama 20 menit
pada suhu ruang yang terlindung dari cahaya. Selanjutnya tambahkan
50 μL Stop Solution ke dalam masing-masing sumur dan baca pada
panjang gelombang 450 nm (pengerjaan dilakukan secara duplo). Hasil
absorban yang keluar dari mesin Elisa reader kemudian dianalisis
menggunakan software CurveExpert 1.4.
F. Pertimbangan Etis
Penelitian ini direview oleh Komite Etik Penelitian Kesehatan
(KEKP) Fakultas Kedokteran Universitas Hasanuddin untuk kelayakan dan
keamanan subjek penelitian dengan mempertimbahkan prinsip-prinsip:
efek samping perlakuan, kemungkinan adverse event yang akan timbul,
serta memberikan bantuan medis/psikososial jika dibutuhkan. Selain itu,
juga dilakukan antisipasi sebelumnya antara lain :
1. Lembar persetujuan menjadi responden (informed concent)
Respon diberi kesempatan isi lembar permohonan persetujuan.
Apabila responden menolak, maka peneliti tidak memaksa dan tetap
menghormati serta menghargai hak koresponden.

100
2. Tanpa nama (Anonimity)
Peneliti tidak mencantumkan nama responden, tetapi peneliti
menggunakan kode tertentu untuk masing-masing responden. Hal ini
dilakukan untuk menjaga kerahasiaan responden.
3. Kerahasiaan data (confidentiality)
Peneliti menjamin kerahasiaan informasi yang dikumpulkan dari
responden. Data tersebut hanya akan disajikan/dilaporkan pada pihak
yang terkait dengan penelitian.
G. Analisis Data
Data hasil DNA sequencing dalam bentuk elektroferogram
kemudian disejajarkan dengan sekuens normal yang berasal dari
GeneBank (NC_000003) kemudian dianalisis dengan menggunakan
perangkat lunak BioEdit Sequence Alignment Editor versi 7.0.5.2 dan
FinchTv versi 1.4.0. Sedangkan data hasil pengukuran TNF-α dianalsisi
menggunakan perangkat lunak CurveExpert versi 1.4.

101
H. Skema Alur Penelitian
Pasien Tuberkulosis Paru
Darah
(Antikoagulan EDTA-K3)
EKSTRAKSI DNA
PCR
Elektroforesis
Ada Band (Pita) Tidak ada Band
(Pita)
SEQUENCING
Blast
Analisis Data
KESIMPULAN
Purifikasi gel cut Extraksi
Isolasi Serum
ELISA
TNF-α

102
BAB IV
HASIL PENELITIAN DAN PEMBAHASAN
A. HASIL PENELITIAN
1. Karakteristik Subjek Penelitian
Sebagai sampel dari penelitian ini adalah darah dari empat
penderita tuberkulosis paru yang sensitif OAT dan resisten OAT yang
berkunjung ke Balai Besar Kesehatan Paru Makassar dan Rumah sakit
Labuang Baji Makassar. Dari empat sampel tersebut, kemudian
diidentifikasi adanya keberadaan dan pola substitusi asam amino pada
gen MyD88 secara keseluruhan.
Distribusi usia, jenis kelamin, keluhan, dan status pemeriksaan
digambarkan dalam Tabel 6.
Tabel 6. Karakteristik sampel
Sampel Jenis Kelamin Umur (tahun) BTA DST
A
B
C
D
Perempuan
Laki-laki
Laki-laki
Laki-laki
39
40
40
48
Pos 1
Pos 1
Pos 3
Pos 3
Sensitif
Sensitif
H,R,S,E
H,R
Keterangan :
BTA = Basil Tahan Asam R = Rifampisin
DST = Drug susceptibility test S = Sterptomisin
H = Isoniasid (INH) E = Etambutol
Pos = Positif 1 – 3

103
2. Analisis Gen MyD88
Daerah gen MyD88 yang dianalisis dalam penelitian ini adalah
daerah sepanjang 4.5 kB dari region 38179969 sampai region 38184513
yang terdiri atas 3 region yaitu region death domain, intermediate domain
dan TIR domain yang mengkode beberapa asam amino
(http//:ncbi.nlm.nih.gov).
Analisis penentuan variasi nukleotida dilakukan dengan
membandingkan sekuens gen MyD88 masing-masing sampel dengan
sekuens nukleotida MyD88 dari geneBank (Accession Number
NC_000003) yang digunakan sebagai konsensus.
Gambar 18. Pemetaan posisi substitusi asam amino berdasarkan Region
a. Region Death Domain
Ditemukan 14 pola substitusi asam amino pada region death
domain (posisi 54 – 109) dengan distribusi sebagai berikut: sampel A
menunjukkan 3 pola substitusi, sampel B menunjukkan 4 pola substitusi,

104
sampel C menunjukkan 8 pola substitusi dan sampel D menunjukkan 3
pola substitusi. Dari 14 pola substitusi yang ditemukan terdapat 4 pola
substitusi yang terdapat pada daerah yang sama pada beberapa sampel
yaitu pola substitusi S23T, V42A, E66D dan R101P. Pola substitusi asam
amino yang ditemukan pada penelitian ini dapat dilihat pada Tabel 7.
Tabel 7. Pola substitusi asam amino pada region death domain
b. Region Intermediate Domain
intermediate domain mengkode sepanjang 46 asam amino, yaitu
posisi 110 – 155. Pola substitusi asam amino yang ditemukan pada
No Substitusi asam amino Kode subjek
1
2
3
4
5
6
7
8
9
10
11
12
13
14
Pro12Thr
Gly18Arg
Gly20Cys
Ser23Thr
Leu38His
Val42Ala
Ser47Thr
Glu66Asp
Phe69Ser
Glu78Asp
Gly85Ala
Asp89Val
Val99Ala
Arg101Pro
(P12T)
(G18R)
(G20C)
(S23T)
(L38H)
(V42A)
(S47T)
(E66D)
(F69S)
(E78D)
(G85A)
(D89V)
(V99A)
(R101P)
C
C
C
B&C
B
B&C
C
B&C
C
A
D
A
D
A&D

105
daerah intermediate domain yaitu 2 pola substitusi yang menyebabkan
perubahan asam amino dan 1 pola substitusi yang tidak menyebabkan
perubahan asam amino. Pola substitusi tersebut dapat dilihat pada
Tabel 8.
Tabel 8. Pola substitusi asam amino pada intermediate domain
No Substitusi asam amino Kode subjek
1
2
3
Gly110Asp
Leu116Val
Ala146Ala
(G110D)
(L116V)
(A146A)
D
A&D
A
c. Region TIR Domain
TIR domain terdiri dari 136 asam amino (posisi 159 – 269). Pada
region ini ditemukan 14 varian pola substitusi asam amino pada ke-4
sampel dengan distribusi sebagai berikut: 1 pola pada sampel A, 6 pola
substitusi pada sampel B, dan 8 pola substitusi pada sampel D. Pola
substitusi asam amino yang didapat pada penelitian ini dapat dilihat pada
Tabel 9.

106
Tabel 9. Pola substitusi asam amino pada TIR domain
2. Karakterisasi asam amino yang mengalami substitusi
Pada penelitian ini, ditemukan beberapa asam amino yang
mengalami substitusi. Beberapa diantara substitusi tersebut menyebabkan
perubahan asam amino yang disandi. Asam amino yang mengalami
perubahan menyebabkan terjadi perubahan sifat. Dapat dilihat pada
Tabel 10
No Substitusi asam amino Kode subjek
1
2
3
4
5
6
7
8
9
10
11
12
13
14
Gln197His
Arg209Ser
Val211Ile
Gly214Arg
Cys216Ser
Glu223Ala
Glu226Ala
Leu229Met
Arg231Thr
Arg232Pro
Cys238Ser
Gln271His
Pro276Ala
Glu285Asp
(Q197H)
(R209S)
(V211I)
(G214R)
(C216S)
(E223L)
(E226A)
(L229M)
(R231T)
(R232P)
(C238S)
(Q271H)
(P276A)
(E285D)
D
D
B&D
D
D
D
D
B
B
B
D
A
B
A&B
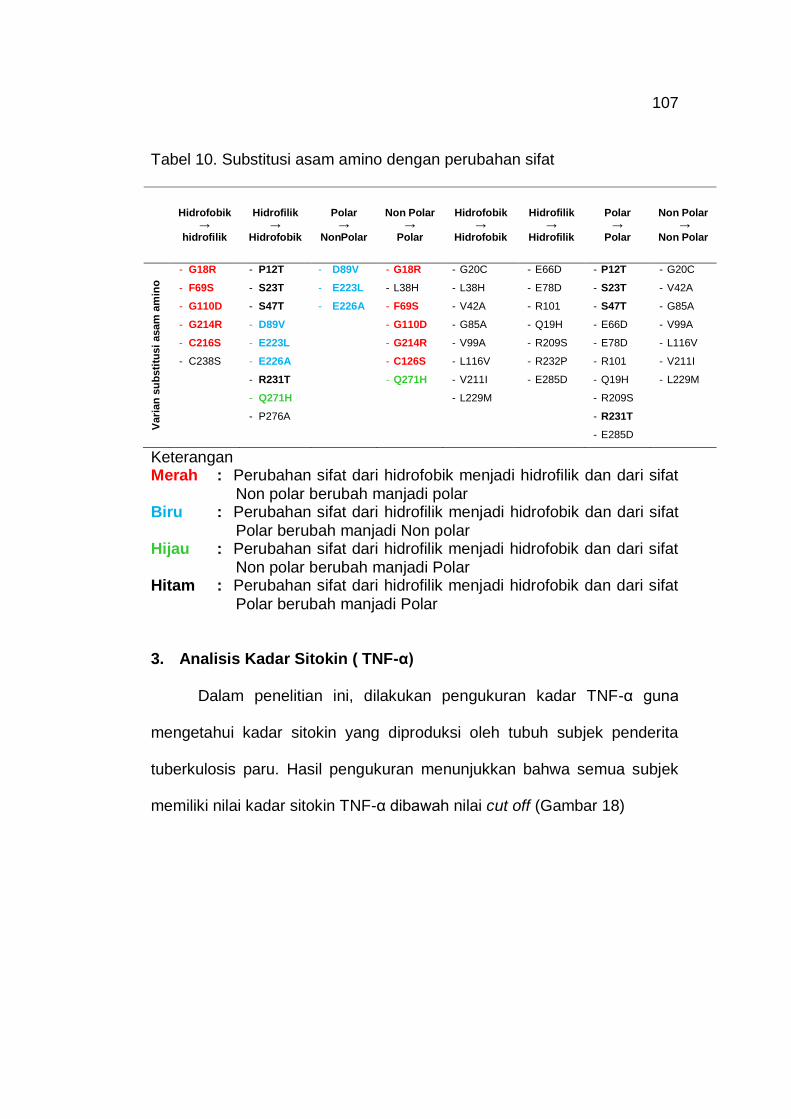
107
Tabel 10. Substitusi asam amino dengan perubahan sifat
Hidrofobik →
hidrofilik
Hidrofilik →
Hidrofobik
Polar →
NonPolar
Non Polar →
Polar
Hidrofobik →
Hidrofobik
Hidrofilik →
Hidrofilik
Polar →
Polar
Non Polar →
Non Polar
Vari
an
su
bsti
tusi
asam
am
ino
- G18R
- F69S
- G110D
- G214R
- C216S
- C238S
- P12T
- S23T
- S47T
- D89V
- E223L
- E226A
- R231T
- Q271H
- P276A
- D89V
- E223L
- E226A
- G18R
- L38H
- F69S
- G110D
- G214R
- C126S
- Q271H
- G20C
- L38H
- V42A
- G85A
- V99A
- L116V
- V211I
- L229M
- E66D
- E78D
- R101
- Q19H
- R209S
- R232P
- E285D
- P12T
- S23T
- S47T
- E66D
- E78D
- R101
- Q19H
- R209S
- R231T
- E285D
- G20C
- V42A
- G85A
- V99A
- L116V
- V211I
- L229M
Keterangan Merah : Perubahan sifat dari hidrofobik menjadi hidrofilik dan dari sifat
Non polar berubah manjadi polar Biru : Perubahan sifat dari hidrofilik menjadi hidrofobik dan dari sifat
Polar berubah manjadi Non polar Hijau : Perubahan sifat dari hidrofilik menjadi hidrofobik dan dari sifat
Non polar berubah manjadi Polar Hitam : Perubahan sifat dari hidrofilik menjadi hidrofobik dan dari sifat
Polar berubah manjadi Polar 3. Analisis Kadar Sitokin ( TNF-α)
Dalam penelitian ini, dilakukan pengukuran kadar TNF-α guna
mengetahui kadar sitokin yang diproduksi oleh tubuh subjek penderita
tuberkulosis paru. Hasil pengukuran menunjukkan bahwa semua subjek
memiliki nilai kadar sitokin TNF-α dibawah nilai cut off (Gambar 18)

108
Gambar 19. Kadar TNF-α pada sampel penderita tuberkulosis
B. PEMBAHASAN
1. Mutasi pada gen Myeloid Differentiation Factor 88 (MyD88)
pola substitusi asam amino yang ditemukan pada penelitian ini
yaitu sebanyak 31 pola mutasi yang tersebar di 3 daerah yaitu 14 pola
substitusi terdapat pada daerah Death domain, 3 pola substitusi pada
daerah Intermediate domain dan 14 pola substitusi pada daerah TIR
domain.
Pada posisi asam amino antara 159 - 318 yang merupakan region
TIR domain ditemukan 14 pola substitusi asam amino dengan pola
substitusi terbanyak ditemukan pada sampel D (8 pola substitusi), sampel
B ditemukan 6 pola substitusi dan sampel A ditemukan 2 pola substitusi.
0
200
400
600
800
1000
1200
1 2 3 4 5 6 7 8 9 10 11 12
TNFα
(p
g/m
L)
Standar Sampel
S1000 S500 S250 S125 S62,5 S31,3 S15,6 A B C D

109
Terdapat 2 pola substitusi sama yang terjadi pada sampel yang berbeda
yaitu V211I dan E285D.
Pada posisi Death domain yaitu posisi antara asam amino 54 – 109
pola substitusi terbanyak ditemukan pada sampel C yaitu sebanyak 7 pola
substitusi, 3 pola substitusi pada sampel A , B dan D. Terdapat 4 pola
substitusi sama yang terjadi pada sampel yang berbeda yaitu S23T,
V42A, E66D dan R101P.
Belum jelas makna dari semua pola substitusi asam amino yang
ditemukan dan apa dampak dari perubahan asam amino tersebut, hal
tersebut diakibatkan karena pola substitusi asam amino yang membawa
mutasi pada gen MyD88 belum pernah dilaporkan pada kasus penyakit
tuberkulosis, baik yang disebabkan oleh spesies Mycobacterium
tuberculosis maupun yang disebabkan oleh golongan mikobakteria non
tuberkulosis.
Beberapa penelitian yang pernah melaporkan varian pola substitusi
asam amino yang terjadi pada gen MyD88 yaitu varian substitusi
Leu265Pro (L265P), namun varian tersebut berkaitan dengan kasus
Waldenstrom macroglobuinemia dan kasus Splenic Marginal Zone
Lymphoma (SMZL) (Poulain S et al,2013 dan Troen G et al, 2012).
Gen MyD88 menyandi 318 asam amino, yang merupakan protein
adaptor sitosolik yang berperan penting pada respon imun bawaan dan
dapatan yang berfungsi dalam jalur signal Toll like receptor dan IL-1
receptor pada respon imun bawaan. Aktivasi melalui IRAK1, IRAK2, IRF7

110
dan TRAF6 akan mengarah pada aktivasi NF-ⱪB, sekresi sitokin dan
respon inflamasi (Ohnishi, H, et all ,2009).
MyD88 terdiri dari N-Terminal death domain (sekitar 99 residu
asam amino) posisi antara 54-109, C-Terminal Toll/Interleukin-1 receptor
(TIR) domain (sekitar 150 residu asam amino) posisi antara 159-318 dan
bagian penghubung yang pendek (Intermediate domain) pada posisi
antara 110-155. Pada respon imun bawaan, TIR domain pada MyD88
memiliki fungsi penting dalam pembentukan kompleks inisiasi sinyal
melibatkan domain sitosol TLRs, di mana domain TIR sitosol LPS
dirangsang oleh TLR4 untuk berinteraksi dengan TIR domain MyD88 dan
bekerja sama dengan protein adaptor lain yang mengandung TIR.
Selanjutnya, signal ditransmisikan ke IL-1 receptor-associated kinase
(IRAK) family. Interaksi antara death domain MyD88 dan IRAK kemudian
diaktifkan oleh jalur fosforilasi dan kemudian berasosiasi dengan TRAF6
untuk mengaktivasi faktor transkripsi NF-ⱪB dan menginduksi JNK (MAP
kinase) untuk mengaktivasi protein AP1 (Gambar 19) (Ohnishi, H el all,
2009; Takeda K et all, 2004).
Untuk mengetahui kemungkinan pola mutasi yang ditemukan pada
penelitian ini memilik makna khususnya terkait dengan jalur signal TLR,
maka perlu dilakukan pengkajian lebih lanjut. Selain itu, pada penelitian ini
juga, belum bisa disimpulkan tentang hubungan keterkaitan antara variasi
genetik dengan tingkat keparahan penyakit tuberkulosis, walaupun
beberapa penelitian dengan menggunakan hewan coba telah

111
membuktikan peran penting MyD88 terhadap infeksi yang disebabkan
oleh M.tuberkulosis.
2. Karakterisasi Asam Amino yang mengalami substitusi
Pola substitusi asam amino yang biasa dilaporkan yang membawa
perubahan yaitu perubahan dari leusin menjadi prolin yang terjadi pada
daerah MyD88 Toll/IL-1 receptor (TIR) domain. Pola mutasi yang
ditemukan pada penelitian ini adalah pola yang tidak lazim, diantaranya
prolin menjadi threonin, glisin menjadi arginin, serin menjadi threonin,
serin menjadi threonin, penil alanin menjadi serin, asam aspartat menjadi
valin, glisin menjadi asam aspartat, glisin menjadi arginin, sistein menjadi
serin, asam glutamat menjadi leusin, asam glutamat menjadi alanin.
Belum jelas apa dampak dari perubahan asam amino tersebut meskipun
terdapat perubahan sifat dari gugus polar menjadi non-polar atau
sebaliknya dan kehilangan sifat hidrofilisitasnya menjadi hidrofobik.
3. Produksi TNF-α
Kadar TNF-α diukur dengan menggunakan metode quantitative
sandwich enzyme immunoassay (ELISA), dasar mikroplate telah dilapisi
dengan antibodi monoklonal yang spesifik terhadap TNF-α. Dari hasil
analisis yang dilakukan menggunakan CurvaExpert 1.4 diperoleh hasil
bahwa empat sampel yang dianalisis menunjukkan kadar TNF-α sangat
rendah yaitu berada dibawah nilai cut off (konsentrasi standar antara

112
1000 – 15,6 pg/mL) dengan distribusi kadar TNF-α sebagai berikut :
sampel A 4,761 pg/mL, sampel B 6,538 pg/mL, C 6,537 pg/mL dan
sampel D 4,281 pg/mL.
Konsentrasi TNF-α yang sangat rendah dapat dikaitkan dengan
mutasi yang ditemukan pada daerah TIR domain dan death domain. Pada
sampel A ditemukan 2 substitusi pada daerah TIR domain, 2 substitusi
pada Intermediate domain dan 3 substitusi pada daerah death domain
dengan kadar TNFα 4,762 pg/mL. Pada sampel B ditemukan 10 substitusi
dengan distribusi : 6 substitusi pada daerah TIR domain dan 4 substitusi
pada daerah death domain dengan kadar TNFα 6,538 pg/mL. Pada
sampel C ditemukan 8 substitusi pada daerah death domain dengan kadar
TNFα 6,537 pg/mL. Sedangkan pada sampel D ditemukan 14 pola
substitusi dengan distribusi : 8 pada daerah TIR, 2 pada daerah
Intermediate domain, dan 3 pola pada daerah Death domain dengan
kadar TNFα yang terukur 4,281 pg/mL.
Data yang diperoleh dapat diindikasikan bahwa adanya
kerusakan/mutasi yang terjadi pada daerah TIR domain maupun death
domain yang merupakan bagian yang berperan dalam penyampaian
signal TLR dan MyD88 menyebabkan signal yang berasal dari MyD88
tidak mampu mengaktivasi NF-ⱪB secara sempurna sehingga produksi
sitokin yang dihasilkan sangat rendah (negatif).

113
Hal ini juga didukung oleh penelitian yang dilakukan oleh Feng CG
et al (2003), menyatakan bahwa MyD88 mengatur pertahanan tubuh
terhadap infeksi mikobakteria dan memiliki peran dalam menentukan
induksi sitokin proinflamasi dan IFN gamma yang dibuktikan dengan
penelitian pada tikus dengan MyD88 (-/-) dan tikus dengan TLR2/TLR4(-/-)
yang diinfeksikan dengan mikobakteria, tikus MyD88 (-/-) memiliki
kerentanan yang lebih parah dan kelangsungan hidup yang lebih singkat
dibandingkan tikus TLR2/TLR4 (-/-) dan dimana beban infeksi lebih berat
dan produksi sitokin sangat rendah.
Aktivasi melalui TLR oleh komponen mikobakteria mempengaruhi
protein adaptor sitoplasmik MyD88 yang merupakan protein adaptor pada
jalur signal Toll like receptors (TLRs) dan reseptors untuk Interleukin (IL)-1
dan IL-8. MyD88 mengatur perakitan multisubunit untuk signal yang
kompleks termasuk IL-1receptor-associated kinase (IRAK)1 dan IRAK4
dan signal Janus kinase/Signal transducer and activator of tanscription
(JAK-STAT) 3 serta jalur aktivasi dan faktor transkripsi seperti canonical
nuclear factor-ĸB (NF- ĸB), aktivator protein 1 (AP-1) dan interferon
regulator faktor (IRF) (O’Neill LA, Bowie AG, 2007).
Beberapa penelitian telah dilakukan untuk membuktikan kaitan
antara gen MyD88 dengan kerentanan terhadap penyakit tuberkulosis
serta keterlibatan gen MyD88 dengan mekanisme imun tubuh terhadap
infeksi tuberkulosis, diantaranya penelitian yang dilakukan oleh Geiger et
al, 2012 dan Kleinnijenhuis et al, 2011, terkait mencit yang kekurangan

114
MyD88, dimana mencit tersebut dilaporkan sangat rentan terhadap infeksi
M.tuberculosis melalui udara. Sebaliknya mencit yang kekurangan TLR
(TLR2,4,9) kerentanannya terhadap M.tuberculosis tidak sebesar yang
kekurangan MyD88.
Belum dapat disimpulkan dengan pasti keterkaitan antara pola
mutasi yang ditemukan dengan kadar TNF-α yang dihasilkan, walaupun
hasil yang diperoleh sesuai pada literatur dan teori yang telah ditemukan
sebelumnya. Hal ini dikarenakan keterbatasan jumlah subyek pada
penelitian ini. Namun dari hasil yang diperoleh dalam penelitian ini, dapat
ditemukan informasi keterkaitan antara faktor genetik dengan respon
imunitas yang dihasilkan tubuh terhadap infeksi yang disebebkan oleh
kuman M.tuberculosis.

115
BAB V
KESIMPULAN DAN SARAN
A. KESIMPULAN
1. Ditemukan 31 pola mutasi dari gen Myeloid Differentiation Factor 88
(total asam amino 318) yang tersebar pada 3 daerah, yaitu daerah
Death domain 14 pola mutasi, daearah Intermediated domain
sebanyak 3 pola mutasi dan daerah TIR domain sebanyak 14 pola
mutasi.
2. Kadar TNF-α pada ke-4 sampel menunjukkan hasil yang sangat
rendah dibawah nilai cut off, namun belum dapat disimpulkan
keterkaitannya dengan pola mutasi gen Myeloid Differentiation Factor
88 yang ditemukan.
B. SARAN
Saran yang dapat direkomendasikan pada penelitian ini adalah:
1. Untuk penelitian selanjutnya, perlu dilakukan penambahan sampel
yang lebih banyak guna menentukan pola mutasi yang paling sering
terjadi pada gen MyD88 dan menghubungkan dengan respon imun
tubuh
2. Perlu dilakukan pengukuran profil produksi sitokin yang lebih
bervariasi untuk menghubungkan mutasi yang terjadi dengan respon
imun yang dihasilkan.

116
3. Perlu dilakukan penelitian dengan membandingkan subjek yang
terinfeksi TB dengan subjek orang sehat pada satu daerah endemik
tuberkulosis untuk membuktikan keterlibatan faktor genetik dengan
keparahan penyakit.

117
DAFTAR PUSTAKA
Abel, B., Thieblemont N., et al. 2002. Toll-like receptor 4 expression is required to control chronic Mycobacterium tuberculosis infection in Mice.Journal of Immunology. 169 :3155 – 62.
Akira, S., and Katuaki, H. 2003. Myeloid Differentiation Factor 88-Dependent and – Independent Pathways in Toll-Like Receptor Signaling. Oxford Journals. 187 : 356 – 363.
Branger, J., Jaklien, C., et al. 2003. Toll-like Receptor4 Plays a Protective Role in Pulmonary Tuberculosis in Mice. International Immunology. 16 : 509 – 516.
Bernuth, H.V., Picard, C., Puel, A., Casanova, J.L. 2012. Experimental and Natural Infections in MyD88- and IRAK-4-deficient Mice and Humans. Eur Journal Immunology. 42 : 3126 – 35.
Bese, F., Chabe, M., and Delhaes, L. 2012. Antibacterial Immunity in Respiratory Tract : What Role for the MyD88 adaptor. Annalec Biologie Clinique Journal. 70 : 245 – 50
Boedina K,S. 2010. Imunologi. Jakarta : Badan Penerbit Fakultas Kedokteran Universitas Indonesia
Bonnert, T.P, Garka,K.E, Parnet, P. Sonodo, G., Testa, J.R, Sims, J.E. 1997. The Cloning and Chracterization of Human MyD88 : a member of an IL-1 receptor related family. FEBS Lett. 402 : 84-81
Burns, K., Janssens, S., Brissoni, B., Olivos, N., Beyaert, R., Tschopp, J. 2003. Inhibition of Interleukin 1 receptor/Toll-Like receptor signaling through the Alternatively Spliced, Short form of MyD88 is due ti its failure to recruit IRAK-4. J. Exp. Med. 197: 263-268
Cecile, M,. Fremond, Togbe, D., et al. 2007. IL-1 Receptor-Mediated Signal Is a Essential Component of MyD88-Dependent Innate Response to Mycobacterium tuberculosis Infection. The Journal of Immunology. 179 : 1178 – 1189.
Cecile, M., Fremond., et al. Fatal Mycobacterium tuberculosis Infection Despite Adaptive Immune Response in the Absence of MyD88. The Journal of Clinical Investigation. 114 : 1790 – 1798.
Chen, M., Aosai, F., et al. 2002. Involvement of MyD88 in Host Defense and The Down –Regulation of Anti-Heat Shock Protein 70 Autoantibody Formation by MyD88 in Toxoplasma gondii Infected Mice. Journal The Parasitology. 88 : 1017 – 9.

118
Ditjen PP&PL Kemeterian Kesehatan RI. 2011. Laporan Situasi Terkini Perkembangan Tuberkulosis di Indonesia. Jakarta : Kementerian Kesehatan Republik Indonesia.
Departemen Kesehatan Republik Indonesia. 2007. Pedoman Nasional Penanggulangan Tuberculosis. Edisi 2. Jakarta : Kemeterian Kesehatan Republik Indonesia.
Departemen Kesehatan Republik Indonesia. 2002. Pedoman Nasional Penanggulangan Tuberkulosis edisi 8. Jakarta : PPTI.
Fatchiyah, Estri, L.A., Sri, W. dan Sri, R. 2011. Biologi Molekuler Prinsip Dasar Analisis. Penerbit Erlangga : Jakarta.
Feng, C.G., Scanga, C.A., et al. 2003. Mice lacking Myeloid Differentiation Factor 88 Dispay Profound Defects in Host Resistance and Immune Responses to Mycobacterium avium Infection not exhibited by Toll Like Receptor 2 (TLR2) and TLR4 deficient Animals. The Journal of Immunology. 171 : 4758 – 64.
Geiger, M.A., Levi., Even R.S., Smith Y., Bowdish DM., Nussbaum G., Rachmilewitz J. 2012. Cell Exposed to Subletal Oxidative Stress Selectively Attact Monocytes/Macrophage Via Scavenger Receptors and MyD88-mediated Sigbaling. Journal Immunologi. 188(3) : 1234 – 44.
Handojo, I. 2003. Pengantar Imunoasai Dasar. Surabaya : Airlangga University Press.
Hardjoeno, Tenri, S., Nurhayana., dkk. 2007. Kumpulan Penyakit Infeksi dan Tes Kultur Sensitivitas Kuman serta Upaya Pengendaliannya. Cahaya Dinan Rucitra : Makassar.
Jawetz, Melnick and Adelberg. 2006. Mikrobiologi Kedokteran. Edisi 20. EGC : Jakarta
Je, E.M., Kim, S.S., Yoo, N.Y., Lee, S.H. 2012. Mutational and Expressional Analyses of MyD88 Gene in Common Solid Cancers. Tumori Journal. 98 : 663 – 9
Jusuf, M. 2001. Genetika I Struktur dan Ekspresi Gen. Sagung Seto. Jakarta
Kawai, T., Adachi, O., Ogawa, T., Takeda, K., Akira, S. 1999. Unresponsiveness of MyD88-deficient Mice to Endotoxin. Journal of Immunity. 11 : 115 – 22.

119
Kementerian Kesehatan Republik Indonesia. 2011. Profil Data Kesehatan Indonesia Tahun 2011. Jakarta : Kementerian Kesehatan Republik Indonesia.
Kleinnijenhuis, J., Marije, O., Leo, A.B.J., Mihai, G.N and Reinout, V.C. 2011. Innate Immune Recognition og Mycobacterium tuberculosis. Clinical and Development Immunology Journal.
Laban Y. 2008. TBC Penyakit dan Cara Pencegahannya. Kanisius : Yogyakarta.
Levine, J.S., Kenneth R.M. 1992. Biology : Discovery Life. Lexintong Mass. United State.
Lieshout, MH.V., Blok, DC., et al. 2012. Differential Roles of MyD88 and TRIF in Hematopoietic and Resident Cells during Murine gram-negative Pneumonia. Journal Infection Dis. 206 : 1415 – 23.
Muraille, E., De, T.C., Brait, M., De, B.P., Leo, O., Carlier, Y. 2003. Genetically Resistant Mice Lacking MyD88-Adapter Protein Display a High Susceptibility to Leishmania Major Infection Associates with a Polarized Th2 Response. The Journal of Immunology. 15 : 4237 – 41
Misnadialy. 2006. Pemeriksaan Laboratorium Tuberkulosis dan Mikobakterium Atipik. Dian Rakyat : Jakarta
Medzhitov, R. 2001. Toll-Like Receptor and Innate Immunity. Nature Reviews Immunology Journal. Volume 1 : 135 - 145.
Medzhitov, R., Preston-Hurlburt, P., Kopp, E., Stadien, A., Chen, C., Ghosh, S., Janeway, C. A. 1998. MyD88 is an adaptor protein in the Toll/IL-1 receptor family signaling pathways. Molec. Cell Journal 2 ; 253-258.
Muzio, M., Ni, J., Feng, F., Dixit, V. M. 1997. IRAK (Pelle) family member IRAK-2 and MyD88 as proximal mediators of IL-1 signaling. Science Medical Journal 278 : 1612-1615.
Najmi, N., Kaur, G., Sharma, SK., Mehra, NK. 2010. Human Toll-like Receptor 4 Polymorphisms TLR4 Asp299Gly and Thr399lle Influence Susceptibility and Severity of Pulmonary Tubercilosis in the Asian Indian Population. Journal Immunology. 76 : 102 – 9.
Nicolle, DM., Pichon, X., Bouchot, A., et al. 2004. Chronic Pneumonia Despite Adaptive Immune Respons to Mycobacterium bovis BCG in MyD88-deficient Mice. Molecular Immunology and Embryology (IEM) Journal. 84 (10) : 1305 – 21

120
Nugraha, J. 2012. Mycobacterium tuberculosis Sistem Imun Alamiah Terkait Penerimanya. Indonesian Journal of Clinical Pathology and Medical Laboratory. 19 : 50 – 45.
O'Neill, L.A.J., and Andrew G. B. 2007. The family of five: TIR-domain-containing adaptors in Toll-like receptor signaling. Natural Reviews Immunology. 7 : 353-364
Ohnishi, H., Hidehito, T,. Zenichiro K., Kenji, E.O., Ailian L., Tekeshi, K., Hidekazu, H., Naomi, K., Masahiro, S. 2009. Structural basis for the multiple interactions of the MyD88 TIR domain in TLR signaling (Online). www.pnas.org/cgi/doi/10.1073/pnas.0812956106.
Poulain, S., Christophe R., Audrey D., Aline R., Charles H., Elisabeth B., et al. 2013. MYD88 L265P mutation in Waldenstrom macroglobulinemia. Blood journal hematology library. 121 : 4504-4511.
Program Pascasarjana UNHAS. 2012. Pedoman Penulisan Tesis dan Disertasi. Edisi 4. Makassar : Univeritas Hasanuddin
Rantam, A.F. 2003. Metode Imunologi. Surabaya : Airlangga University Press.
Radji, M. 2011. Rekayasa Genetika Pengantar untuk Profesi Kesehatan. Sagung Seto : Jakarta.
Retnoningrum, S.D., Roga, F.K., 2004. Mekanisme Resistensi Tingkat
Molekul Terhadap Beberapa Obat Pada Mycobacterium
Tuberculosis. Acta Pharmaceutica Indonesia Journal, (Online). Vol.
29. No. 3: 92-95, diakses 18 Januari 2012.
Runtuwene. R., Damodoro. N., Sekar. S. 2009. Peranan Toll Like
Receptor Terhadap Terjadinya Meningitis Tuberkulosis. Jurnal tidak
diterbitkan. Yogyakarta : Ilmu Penyakit Saraf Fakultas Kedokteran -
UGM
Ryffel, B., Jacobs, M., Parida, S., Botha, T., Togbe, D., Quesniaux, V. 2006. Toll-Like Receptor and Control of Mycobacterium Infection in Mice. Novartis Found Symp. 279 : 127 – 39.
Saiga H., Shimada Y., Takeda K. 2011. Innate Immune Effectores in Mycobacterium tuberculosis Infection. Clinical and Development Immunology.
Sardjono, E., Suradi., Eddy S. 2008. Nilai sensitivitas, Spesifitas Pemeriksaan Serologi TB dan Sputum BTA Mikroskopik pada Klinis Tuberkulosis Paru. Jurnal Tuberkulosis Indonesia. 5 : 24 – 27.

121
Subowo. 2009. Imunobiologi. Edisi 2. Jakarta : Sagung Seto.
Sudoyo A.W. dkk. 2009. Buku Ajar Ilmu Penyakit Dalam. Edisi V. Jilid III. Internal Publishing : Jakarta.
Sugawara, I., Yamada, H., Mizuno, S., Takeda, K., Akira, S. 2003. Mycobacterial Infection in MyD88-deficient Mice. Microbiology Immunology Journal. 47 : 841 – 7.
Syahrini, H. 2008. Tuberkulosis Paru Resisten Ganda. Tesis tidak diterbitkan. Sumatera Utara : Program Spesialis Ilmu Penyakit Dalam FK. Universitas Sumatera Utara.
Takeuchi, O., Hoshino, K., Akira, S., 2000. Cutting Edge : TLR2-deficient and MyD88-deficient Mice are Highly Susceptible to Staphylococcus aureus Infetion. Journal of Immunology. 165 : 5392 – 6.
Takeda, K., Shizuo A., 2004. TLR signaling pathways (Online). www.elsevier.com/locate/ysmim.
Terence, M., Doherty., and Moshe, A. 2004. TB, or not TB : that is the Question does TLR Signaling Hold the Answer. The Journal of Clinical InvestigationI. 114 : 1699 – 1703.
Widoyono. 2008. Penyakit Tropis Epidemiologi, Penularan, Pencegahan dan Pemberantasannya. Penerbit Erlangga : Jakarta.
Williams, L and Wilkins. 1994. Bergey's Manual of Determinative Bacteriology. Lippincott and Wilkins : Philadelphia
World Health Organization. 2012. Global Tuberculosis Report 2012. WHO
Yuwono, T. 2005. Biologi Molekuler. Penerbit Erlangga : Jakarta
Yuwono, T. 2006. Teori dan Aplikasi Polymerase Chain Reaction. Penerbit ANDI : Yogyakarta
Zhang, Y.X., Xue, Y., et al. 2011. Association of TIRAP (MAL) gene polymorfisms with susceptibility to tuberculosis in a Chinese population. Online Journal of Genetics and Molecular Research. 10 : 7 – 15.


123

124
Lampiran 3. Sekuens primer yang digunakan untuk amplifikasi DNA MyD88
Primer Sekuens nukleotida (5’ – 3’) Tm (ºC) Posisi
Fragmen 1F
Fragmen 1R
Fragmen 2F
Fragmen 2R
Fragmen 3F
Fragmen 3R
Fragmen 4F
Fragmen 4R
Fragmen 5F
Fragmen 5R
Fragmen 6F
Fragmen 6R
Fragmen 7F
Fragmen 7R
Fragmen 8F
Fragmen 8R
Fragmen 9F
Fragmen 9R
Fragmen 10F
Fragmen 10R
Fragmen 11F
Fragmen 11R
Fragmen 12F
Fragmen 12R
Fragmen 13F
Fragmen 13R
GAT TCC TCC GTG GAA GAA CTG T
CCC AGC TTG GTA AGC AGC TC
GTC TCC TCC ACA TCC TCC CTT C
TCC CCA GGA CTT GAG CTT TCT A
ACT GCT CGA GCT GCT TAC CAA G
AGG CCA TCA AAG TCA GGT TTG T
CAC ACC TTG CTG AGT TGG AAT C
ACC CAT GCT CTT AAC ACC ACA A
ACT TCT CAG AGC CGT TGA GCT T
GTC AGA GCT CCA TGG GAA GG
TGG TGT TAA GAG CAT GGG TGT T
TCC TGT GGC AAC TTA GTT CAG C
TGC TAG TGA GCT CAT CGA AAA GA
TTG GTG TAG TCG CAG ACA GTG A
CAG GTA AGC TCA ACC CTG CTC T
GTT CCA TGC AGG ACA TGA AAT G
TCA CTG TCT GCG ACT ACA CCA A
CAT CAG TAG GCA TCA CTG TGG A
GGG ATG GGG AGA ACA GAG AGT A
GGG CAG GAG GCT TTT ATA TGG T
CAG TGA CAA GTC CCC AAG AGA C
AAA TTG CTC TGG GAA GGA GAG A
CAT GGG CAC ATA CAG ACT CAC A
CCA GGA TAT AGG AAG AAT GGC AAA
CCC AGG ACA GAC CCT AGG AC
CAG CCC TGC TTA GGA CTG AAC
57.5
57.3
57.1
56.3
57.5
56.8
55.9
54.6
56.5
57.1
55.9
54.6
57.8
674
650
681
595
656
684
683
621
595
639
656
698
483

125
Lampiran 4. Data GeneBank (Accession Number NC_000003)
Homo sapiens chromosome 3, GRCh37.p10 Primary Assembly
Homo sapiens myeloid differentiation primary response 88 (MYD88) position From base :
38179969 to base 38184513
FASTA Graphics
LOCUS NC_000003 4545 bp DNA linear CON 30-OCT-2012
DEFINITION Homo sapiens chromosome 3, GRCh37.p10 Primary Assembly.
ACCESSION NC_000003 REGION: 38179969..38184513 GPC_000000027
VERSION NC_000003.11 GI:224589815
DBLINK BioProject: PRJNA168
KEYWORDS RefSeq.
SOURCE Homo sapiens (human)
ORGANISM Homo sapiens
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi;
Mammalia; Eutheria; Euarchontoglires; Primates; Haplorrhini;
Catarrhini; Hominidae; Homo.
additions
are manually curated by GRC staff. For more information see:
http://genomereference.org.
##Genome-Annotation-Data-START##
Annotation Provider :: NCBI
Annotation Status :: Full annotation
Annotation Version :: Homo sapiens Annotation Release 104
Annotation Pipeline :: NCBI eukaryotic genome annotation pipeline
Annotation Method :: Best-placed RefSeq; Gnomon
Features Annotated :: Gene; mRNA; CDS; ncRNA
##Genome-Annotation-Data-END##
FEATURES Location/Qualifiers
source 1..4545
/organism="Homo sapiens"
/mol_type="genomic DNA"
/db_xref="taxon:9606"
/chromosome="3"
gene 1..4545
/gene="MYD88"
/gene_synonym="MYD88D"
/note=myeloid differentiation primary response gene (88)
Derived by automated computational analysis using gene
prediction method: BestRefseq."
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
mRNA join(1..551,1387..1521,1911..2115,2280..2371,2655..4545)
/gene="MYD88"
/gene_synonym="MYD88D"
/product="myeloid differentiation primary response gene
(88), transcript variant 1"
/note="Derived by automated computational analysis using
gene prediction method: BestRefseq."

126
/transcript_id="NM_001172567.1"
/db_xref="GI:289546502"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
mRNA join(1..551,1387..1521,1911..2091,2280..2371,2655..4545)
/gene="MYD88"
/gene_synonym="MYD88D"
/product="myeloid differentiation primary response gene
(88), transcript variant 2"
/note="Derived by automated computational analysis using
gene prediction method: BestRefseq."
/transcript_id="NM_002468.4"
/db_xref="GI:197276653"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
mRNA join(1..551,1387..1521,2280..2371,2655..4545)
/gene="MYD88"
/gene_synonym="MYD88D"
/product="myeloid differentiation primary response gene
(88), transcript variant 4"
/note="Derived by automated computational analysis using
gene prediction method: BestRefseq."
/transcript_id="NM_001172569.1"
/db_xref="GI:289546652"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
mRNA join(1..551,1911..2091,2280..2371,2655..4545)
/gene="MYD88"
/gene_synonym="MYD88D"
/product="myeloid differentiation primary response gene
(88), transcript variant 3"
/note="Derived by automated computational analysis using
gene prediction method: BestRefseq."
/transcript_id="NM_001172568.1"
/db_xref="GI:289546580"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
mRNA join(1..551,2280..2371,2655..4545)
/gene="MYD88"
/gene_synonym="MYD88D"
/product="myeloid differentiation primary response gene
(88), transcript variant 5"
/note="Derived by automated computational analysis using
gene prediction method: BestRefseq."
/transcript_id="NM_001172566.1"
/db_xref="GI:289546499"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"

127
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
CDS join(185..551,1387..1521,1911..2115,2280..2371,2655..2809)
/gene="MYD88"
/gene_synonym="MYD88D"
/note="isoform 1 is encoded by transcript variant 1;
Derived by automated computational analysis using gene
prediction method: BestRefseq."
/codon_start=1
/product="myeloid differentiation primary response
protein MyD88 isoform 1"
/protein_id="NP_001166038.1"
/db_xref="GI:289546503"
/db_xref="CCDS:CCDS54565.1"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
/translation="MRPDRAEAPGPPAMAAGGPGAGSAAPVSSTSSLPLAALNMRVRR
RLSLFLNVRTQVAADWTALAEEMDFEYLEIRQLETQADPTGRLLDAWQGRPGASVGRL
LELLTKLGRDDVLLELGPSIEEDCQKYILKQQQEEAEKPLQVAAVDSSVPRTAELAGI
TTLDDPLGHMPERFDAFICYCPSDIQFVQEMIRQLEQTNYRLKLCVSDRDVLPGTCVW
SIASELIEKRLARRPRGGCRRMVVVVSDDYLQSKECDFQTKFALSLSPGAHQKRLIPI
KYKAMKKEFPSILRFITVCDYTNPCTKSWFWTRLAKALSLP"
CDS join(185..551,1387..1521,1911..2091,2280..2371,2655..2809)
/gene="MYD88"
/gene_synonym="MYD88D"
/note="isoform 2 is encoded by transcript variant 2;
Derived by automated computational analysis using gene
prediction method: BestRefseq."
/codon_start=1
/product="myeloid differentiation primary response
protein MyD88 isoform 2"
/protein_id="NP_002459.2"
/db_xref="GI:197276654"
/db_xref="CCDS:CCDS2674.2"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
/translation="MRPDRAEAPGPPAMAAGGPGAGSAAPVSSTSSLPLAALNMRVRR
RLSLFLNVRTQVAADWTALAEEMDFEYLEIRQLETQADPTGRLLDAWQGRPGASVGRL
LELLTKLGRDDVLLELGPSIEEDCQKYILKQQQEEAEKPLQVAAVDSSVPRTAELAGI
TTLDDPLGHMPERFDAFICYCPSDIQFVQEMIRQLEQTNYRLKLCVSDRDVLPGTCVW

128
SIASELIEKRCRRMVVVVSDDYLQSKECDFQTKFALSLSPGAHQKRLIPIKYKAMKKE
FPSILRFITVCDYTNPCTKSWFWTRLAKALSLP"
CDS join(185..551,1911..2091,2280..2371,2655..2809)
/gene="MYD88"
/gene_synonym="MYD88D"
/note="isoform 3 is encoded by transcript variant 3;
Derived by automated computational analysis using gene
prediction method: BestRefseq."
/codon_start=1
/product="myeloid differentiation primary response
protein MyD88 isoform 3"
/protein_id="NP_001166039.1"
/db_xref="GI:289546581"
/db_xref="CCDS:CCDS54567.1"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
/translation="MRPDRAEAPGPPAMAAGGPGAGSAAPVSSTSSLPLAALNMRVRR
RLSLFLNVRTQVAADWTALAEEMDFEYLEIRQLETQADPTGRLLDAWQGRPGASVGRL
LELLTKLGRDDVLLELGPSIGHMPERFDAFICYCPSDIQFVQEMIRQLEQTNYRLKLC
VSDRDVLPGTCVWSIASELIEKRCRRMVVVVSDDYLQSKECDFQTKFALSLSPGAHQK
RLIPIKYKAMKKEFPSILRFITVCDYTNPCTKSWFWTRLAKALSLP"
CDS join(185..551,1387..1521,2280..2371,2655..2675)
/gene="MYD88"
/gene_synonym="MYD88D"
/note="isoform 4 is encoded by transcript variant 4;
Derived by automated computational analysis using gene
prediction method: BestRefseq."
/codon_start=1
/product="myeloid differentiation primary response
protein MyD88 isoform 4"
/protein_id="NP_001166040.1"
/db_xref="GI:289546653"
/db_xref="CCDS:CCDS54566.1"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
/translation="MRPDRAEAPGPPAMAAGGPGAGSAAPVSSTSSLPLAALNMRVRR
RLSLFLNVRTQVAADWTALAEEMDFEYLEIRQLETQADPTGRLLDAWQGRPGASVGRL
LELLTKLGRDDVLLELGPSIEEDCQKYILKQQQEEAEKPLQVAAVDSSVPRTAELAGI
TTLDDPLGAAGWWWLSLMITCRARNVTSRPNLHSASLQVPIRSD"

129
CDS join(185..551,2280..2371,2655..2675)
/gene="MYD88"
/gene_synonym="MYD88D"
/note="isoform 5 is encoded by transcript variant 5;
Derived by automated computational analysis using gene
prediction method: BestRefseq."
/codon_start=1
/product="myeloid differentiation primary response
protein MyD88 isoform 5"
/protein_id="NP_001166037.1"
/db_xref="GI:289546500"
/db_xref="CCDS:CCDS54568.1"
/db_xref="GeneID:4615"
/db_xref="HGNC:7562"
/db_xref="HPRD:03703"
/db_xref="MIM:602170"
/translation="MRPDRAEAPGPPAMAAGGPGAGSAAPVSSTSSLPLAALNMRVRR
RLSLFLNVRTQVAADWTALAEEMDFEYLEIRQLETQADPTGRLLDAWQGRPGASVGRL
LELLTKLGRDDVLLELGPSIGAAGWWWLSLMITCRARNVTSRPNLHSASLQVPIRSD"
ORIGIN
1 cccctcagat tcctccgtgg aagaactgtg gctccaccag atcccaaaaa gcaaggcccg
61 tttcctacaa cccccgaagg agggtcgtcc tcactccgcc gccaacctac tagcaccatc
121 accagaccct cgagggcggt gccgtggacc tctccagatc tcaaaaggca gattcctact
181 tcttacgccc cccacatcac ccgcctcgag acctcaaggg tagaggtggg cacccccgcc
241 tccgcacttt tgctcggggc tccagattgt agggcagggc ggcgcttctc ggaaagcgaa
301 agccggcggg gcggggcggg tgccgcagga gaaagaggaa gcgctggcag acaatgcgac
361 ccgaccgcgc tgaggctcca ggaccgcccg ccatggctgc aggaggtccc ggcgcggggt
421 ctgcggcccc ggtctcctcc acatcctccc ttcccctggc tgctctcaac atgcgagtgc
481 ggcgccgcct gtctctgttc ttgaacgtgc ggacacaggt ggcggccgac tggaccgcgc
541 tggcggagga gatggacttt gagtacttgg agatccggca actggagaca caagcggacc
601 ccactggcag gctgctggac gcctggcagg gacgccctgg cgcctctgta ggccgactgc
661 tcgagctgct taccaagctg ggccgcgacg acgtgctgct ggagctggga cccagcattg
721 gtgaggacgt ccccttcctg gcctcgtacc tggggggtga ggaggctgac tttccgcggc
781 ctcagcatcc tgtctcccat ggagagaccc catttcctgc ctcgggggcc cgaagaagcc
841 tgcagaggga gaaccatgcg ggtcccgttc cttcttaata accggtcgcg gttattaaga
901 aggactggag aaaggtccgg ataggcggag atgggaagga agcagcttag gcagaggctt
961 tcaggtaggg ccaggagtca gaatcaggct tctgtggggg catctgggct gtttcaagta
1021 gagcaacagg acaggtgggg cgattgacag tggactgtct tagaaacctc aagtcctggg
1081 gaaatgcagc ccttctttct actcactggc acttacataa tatacatgca taggcgttgg
1141 atacagccgc ccacagacag gcacaccttg ctgagttgga atcactgcac cataaccagt
1201 gggtctcctg agcctttctg gcatgcccag ccccttgctc acatctgccc tggatcccag
1261 aagaagcaga cctaccttgg taccattctt aggatcccta ggaagggaca gagatacaaa
1321 cctgactttg atggccttcc agaaagccag aacaccactg acatcccttt gggtcagtta
1381 gagccagtgg gagctcaact tctcagagcc gttgagcttc gcgtggcacc agtgaactgg
1441 ggaagccctc tagaacaacc cagccagagg aggtgggaca gcggctggat cctgactgtg
1501 ggtaaagagg taggcactcc cagggaggct gctttactct gtctcttccc cacagaggag
1561 gattgccaaa agtatatctt gaagcagcag caggaggagg ctgagaagcc tttacaggtg
1621 gccgctgtag acagcagtgt cccacggaca gcagagctgg cgggcatcac cacacttgat
1681 gaccccctgg gtaagggtcc aatactgttc ccatgggaca ggtggaatag gacattgtgg
1741 tgttaagagc atgggtgttt gaagcagatg ggctgtgaga ccttgggcaa gtcacttaat
1801 ctttctgagc ctcagtttcc tcacctaaga aatggagata atagtcctac ctctggattg
Primer 1 Forward
Primer 1 Reverse & Primer 3 Forward
Primer 2 Forward
Primer 2 Reverse
Primer 4 Forward
Primer 3 reverse
Primer 5 forward
Primer 4 Reverse & Primer 6 Forward

130
1861 ctgtgagatg ctcatgaaat aatgtctgtc tcgtggttaa tccagagctt agcccctgag
1921 gtactcatct ttcctctcct ggaaagggca ctttctctga ggagtatcat cttgggaagg
1981 gtgcagggcc cagggttgcc taggcagggg actcttggct ggatccctcc caagccttcc
2041 catggagctc tgaccaccac ccttgtgctc tgcacccagg gcatatgcct gagcgtttcg
2101 atgccttcat ctgctattgc cccagcgaca tccagtttgt gcaggagatg atccggcaac
2161 tggaacagac aaactatcga ctgaagttgt gtgtgtctga ccgcgatgtc ctgcctggca
2221 cctgtgtctg gtctattgct agtgagctca tcgaaaagag gttggctaga aggccacggg
2281 gtgggtgcgt ggatgcatga agccctgccc tggggtccag atactgggca tctcctccta
2341 gctgtgcact gtccagcctg ggcacagtgg gcccttcctg aagctattcc caggggatat
2401 gctgaactaa gttgccacag gacctgcagc ctgcccactc tcccctaggt gccgccggat
2461 ggtggtggtt gtctctgatg attacctgca gagcaaggaa tgtgacttcc agaccaaatt
2521 tgcactcagc ctctctccag gtaagctcaa ccctgctctg gcaagagaat gagggaatgt
2581 gtaggtgggg cctctggatt gtcagccttc cctccccaag gactgtggat gcagtaccaa
2641 agaactgctg aagatctctg cacacctgag catgtgtgca tgtgtgtgcc tttttgtgtg
2701 agtgaatgtg tgccaggggt acttagatgg gggatggctg ttgttaaccc tggggttgaa
2761 gactgggctt gtcccaccat ggggcaaggg cctgatgcca gcatggcacc ccttggcttg
2821 caggtgccca tcagaagcga ctgatcccca tcaagtacaa ggcaatgaag aaagagttcc
2881 ccagcatcct gaggttcatc actgtctgcg actacaccaa cccctgcacc aaatcttggt
2941 tctggactcg ccttgccaag gccttgtccc tgccctgaag actgttctga ggccctgggt
3001 gtgtgtgtat ctgtctgcct gtccatgtac ttctgccctg cctcctcctt tcgttgtagg
3061 aggaatctgt gctctactta cctctcaatt cctggagatg ccaacttcac agacacgtct
3121 gcagcagctg gacatcacat ttcatgtcct gcatggaacc agtggctgtg agtggcatgt
3181 ccacttgctg gattatcagc caggacacta tagaacagga ccagctgaga ctaagaagga
3241 ccagcagagc cagctcagct ctgagccatt cacacatctt caccctcagt ttcctcactt
3301 gaggagtggg atggggagaa cagagagtag ctgtgtttga atccctgtag gaaatggtga
3361 agcatagctc tgggtctcct gggggagacc aggcttggct gcgggagagc tggctgttgc
3421 tggactacat gctggccact gctgtgacca cgacactgct ggggcagctt cttccacagt
3481 gatgcctact gatgcttcag tgcctctgca caccgcccat tccacttcct ccttccccac
3541 agggcaggtg gggaagcagt ttggcccagc ccaaggagac cccaccttga gccttatttc
3601 ctaatgggtc cacctctcat ctgcatcttt cacacctccc agcttctgcc caaccttcag
3661 cagtgacaag tccccaagag actcgcctga gcagcttggg ctgcttttca tttccacctg
3721 tcaggatgcc tgtggtcatg ctctcagctc cacctggcat gagaagggat cctggcctct
3781 ggcatattca tcaagtatga gttctgggga tgagtcactg taatgatgtg agcagggagc
3841 cttcctccct gggccacctg cagagagctt tcccaccaac tttgtacctt gattgcctta
3901 caaagttatt tgtttacaaa cagcgaccat ataaaagcct cctgccccaa agcttgtggg
3961 cacatgggca catacagact cacatacaga cacacacata tatgtacaga catgtactct
4021 cacacacaca ggcaccagca tacacacgtt tttctaggta cagctcccag gaacagctag
4081 gtgggaaagt cccatcactg agggagccta accatgtccc tgaacaaaaa ttgggcactc
4141 atctattcct tttctcttgt gtccctactc attgaaacca aactctggaa aggacccaat
4201 gtaccagtat ttatacctct aatgaagcac agagagagga agagagctgc ttaaactcac
4261 acaacaatga actgcagaca cagctgttct ctccctctct ccttcccaga gcaatttata
4321 ctttaccctc aggctgtcct ctggggagaa ggtgccatgg tcttaggtgt ctgtgcccca
4381 ggacagaccc taggacccta aatccaatag aaaatgcata tctttgctcc actttcagcc
4441 aggctggagc aaggtacctt ttcttaggat cttgggaggg aatggatgcc cctctctgca
4501 tgatcttgtt gaggcattta gctgccatgc acctgtcccc ctttaatact gggcatttta
4561 aagccatctc aagaggcatc ttctacatgt tttgtacgca ttaaaataat ttcaaagata
4621 tctgagaaaa gccgatattt gccattcttc ctatatcctg gaatatatct tgcatcctga
4681 gtttataata ataaataata ttctaccttg gaaacttgtg tgtgtgttga gtggaagagg
4741 tttggaagca gtaatgtggg taagagaagc tggtccactg ggtgggttcc agcctggatt
4801 tggcacgggc ttcctgaaag ctgggccccc tcctcacagg ttcagtccta agcagggctg
4861 caggcagaac caggaatact gacctcagct acagacactc a
Primer 5 reverse
Primer 7 forward
Primer 6 reverse
Primer 8 forward
Primer 9 forward & Primer 7 reverse
Primer 8 reverse
Primer 10 forward
Primer 9 reverse
Primer 11 forward
Primer 10 reverse
Primer 12 forward
Primer 13 forward
Primer 11 reverse
Primer 12 reverse
Primer 13 reverse

131
Lampiran 5. Hasil Elektroforesis
Keterangan :
P1-A : Sampel kode A untuk primer fragmen 1
P1-B : Sampel kode B untuk primer fragmen 1
P1-C : Sampel kode C untuk primer fragmen 1
P1-D : Sampel kode D untuk primer fragmen 1
P2-A : Sampel kode A untuk primer fragmen 2
P2-B : Sampel kode B untuk primer fragmen 2
P2-C : Sampel kode C untuk primer fragmen 2
P2-D : Sampel kode D untuk primer fragmen 2
P3-A : Sampel kode A untuk primer fragmen 3
P3-B : Sampel kode B untuk primer fragmen 3
P3-C : Sampel kode C untuk primer fragmen 3
P3-D : Sampel kode D untuk primer fragmen 3
P4-A : Sampel kode A untuk primer fragmen 4
P4-B : Sampel kode B untuk primer fragmen 4
P4-C : Sampel kode C untuk primer fragmen 4
P4-D : Sampel kode D untuk primer fragmen 4
P5-A : Sampel kode A untuk primer fragmen 5
P5-B : Sampel kode B untuk primer fragmen 5
674 bp 650 bp
681 bp
595 bp
656 bp
500 bp
500 bp

132
Lampiran 6. Hasil analisis elektroferogram

133

134
135

136

137

138
Lampiran 7. Hasil analisis Bioedit sekuensing gen MyD88 exon 1
Lampiran 8. Hasil analisis Bioedit sekuensing gen MyD88 exon 2

139
Lampiran 9. Hasil analisis Bioedit sekuensing gen MyD88 exon 3
Lampiran 10. Hasil analisis Bioedit sekuensing gen MyD88 exon 4
Lampiran 11. Hasil analisis Bioedit sekuensing gen MyD88 exon 5

140
Lampiran 12. Hasil analisis Bioedit sekuensing asam amino exon 1
Lampiran 13. Hasil analisis Bioedit sekuensing asam amino exon 2
Lampiran 14. Hasil analisis Bioedit sekuensing asam amino exon 3
Lampiran 15. Hasil analisis Bioedit sekuensing asam amino exon 4
Lampiran 16. Hasil analisis Bioedit sekuensing asam amino exon 5
A146A

141
LAMPIRAN 17 SKEMA KERJA PEWARNAAN ZIEHL NEELSEN

142
LAMPIRAN 18. SKEMA KERJA EKSTRAKSI DNA

143
LAMPIRAN 19. SKEMA KERJA PCR

144
LAMPIRAN 20. SKEMA KERJA ELISA

145
LAMPIRAN 21. PHYSICOCHEMICAL PROPERTIES OF AMINO ACID




















