![No Representation without Colonisation? (Or, Nature Represents Itself) [author's pre-print]](https://static.fdokumen.com/doc/165x107/63362e6b379741109e00f1ae/no-representation-without-colonisation-or-nature-represents-itself-authors.jpg)
No Representation without Colonisation? (Or, Nature Represents Itself) [author's pre-print]

Subsurface hydrology and degree of burial affectmass loss and invertebrate colonisation of leavesin a woodland stream
DANIELLE C. TILLMAN, ASHLEY H. MOERKE, CARRIE L. ZIEHL and GARY A. LAMBERTI
Department of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556-0369, U.S.A.
SUMMARY
1. In deciduous forest streams, fallen leaves form a large component of the total organic
matter budget, and many leaves become buried within stream sediments. We examined
the processing of buried leaves as compared with those at the surface, and the influence of
subsurface hydrology on processing rates.
2. Leaf packs were secured on the streambed surface or buried 10 cm deep in upwelling
and downwelling reaches of a second-order stream in Michigan, U.S.A. Mass loss and
invertebrate colonisation were measured from October to February.
3. Leaves buried in upwelling reaches lost mass more slowly (exponential decay
coefficient, k ¼ )0.0097) than did leaves from the other treatments (buried downwelling:
)0.017; surface upwelling: )0.022; surface downwelling: )0.021).
4. Initially, more invertebrates colonised surface leaf packs than buried packs. During the
remainder of the study, however, hydrology had a greater effect on invertebrate
abundance than did burial, as more invertebrates were found in packs in downwelling
reaches than in upwelling reaches.
5. Local subsurface hydrology and degree of burial, factors rarely considered in studies of
detritus processing, can significantly influence mass loss and invertebrate colonisation of
fallen leaves in streams. Furthermore, because of slower processing, subsurface zones may
function as organic matter reservoirs that gradually ‘spiral’ carbon to downstream
subsurface and surface habitats.
Keywords: carbon cycling, detritus processing, hyporheic zone, meiofauna, Michigan stream
Introduction
Local vertical hydraulic gradient (VHG) can influence
many stream patterns and processes, including nutri-
ent concentrations (Triska, Duff & Avanzino, 1993;
Valett, 1993; Findlay, 1995), invertebrate distributions
(Williams, 1993; Plenet & Gibert, 1994), metabolism
(Jones, Fisher & Grimm, 1995) and carbon dynamics
(Vervier & Naiman, 1992; Wagner, Schmidt &
Marxsen, 1993; Marmonier et al., 1995). Most studies
of hyporheic carbon dynamics have focused on
dissolved organic carbon (DOC). However, each
autumn in temperate regions, huge quantities of
particulate organic carbon (POC) enter woodland
streams as deciduous leaves (Fisher, 1973), and much
of that carbon may become buried within the sedi-
ments (Smock, Metzler & Gladden, 1989; Metzler &
Smock, 1990). Knowledge of the fate of buried POC is
important for a complete understanding of stream
carbon cycling.
Processing of POC is influenced by a number of
environmental factors (see below and review by
Webster & Benfield, 1986). Conditions can differ
between locations on or within the sediments and
can also vary based on local VHG. For example, faster
leaf processing has been associated with increased
Correspondence: Gary A. Lamberti, Department of Biological
Sciences, University of Notre Dame, Notre Dame, IN 46556-0369,
U.S.A. E-mail: [email protected]
Freshwater Biology (2003) 48, 98–107
98 � 2003 Blackwell Publishing Ltd

oxygen availability (Reed, 1979). Surface water gen-
erally contains more dissolved oxygen (DO) than does
pore water (Hendricks & White, 1991), and DO
concentration is usually higher in downwelling than
in upwelling reaches (Valett, 1993). Faster leaf break-
down has also been associated with higher tempera-
ture (e.g. Rowe et al., 1996). Pore water temperature
generally fluctuates less than surface water tempera-
ture (Silliman & Booth, 1993), and pore water tem-
perature in upwelling reaches tends to be higher than
that in downwelling regions as the air temperature
drops. Thus, in the late autumn and winter, after
deciduous leaf-abscission, the temperature in upwel-
ling reaches can be higher than that in downwelling
reaches. Water nutrient concentrations can also
influence leaf breakdown (Fairchild, Boyle & Robin-
son-Wilson, 1983; Suberkropp & Chauvet, 1995), and
these concentrations often differ between pore and
surface water (Hendricks & White, 1991; Valett, 1993;
Hendricks & White, 1995).
Physical and chemical variability in aquatic envir-
onments also leads to variability in the abundance and
distribution of biota (Dole-Olivier et al., 1994; Boulton
& Stanley, 1995). Because invertebrates are important
processors of leaves in many lotic systems (Anderson
& Sedell, 1979; Kirby, Webster & Benfield, 1983;
Cuffney, Wallace & Lugthart, 1990), spatial hetero-
geneity in invertebrate communities may lead to
heterogeneity in the pattern of leaf processing. Inver-
tebrate abundance and diversity can differ between
assemblages on and within the sediments (Godbout &
Hynes, 1982; Strommer & Smock, 1989; McElravy &
Resh, 1991; Schmid-Araya, 1995; Palmer et al., 2000),
and more individuals and taxa have been found in
downwelling than in upwelling reaches (Creuze des
Chatelliers, 1991; Dole-Olivier, Marmonier & Beffy,
1997).
We conducted a 120-day experiment in a Michigan
stream to investigate how VHG (upwelling or down-
welling) and location within the streambed (buried or
on the surface) affects the breakdown of leaf detritus.
We hypothesised that leaves would be processed (i.e.
lose mass) more rapidly on the surface than when
buried and more rapidly in downwelling than in
upwelling reaches. In addition, we expected inverte-
brate colonisation to differ, based on degree of burial
and local subsurface hydrology, with greater inverte-
brate abundance and diversity in surface than in buried
packs and in downwelling than in upwelling zones.
Methods
Study site
Bertrand Creek (Berrien Co., MI, USA; 41�46¢N,
86�17¢W) is a second-order woodland stream of
about 3 km in length and with a catchment of
5.6 km2. Maple (Acer spp.) and oak (Quercus spp.) are
the most common riparian trees, and the channel has
a dense canopy. The stream drains an area of glacial
till, and sediment grain size is heterogeneous. Bert-
rand Creek cuts through several geological strata
and, as a result, the direction of VHG within the
sediments changes along the stream course. Our
study sites were positioned within four predomin-
antly sandy reaches: two downwelling (positive
VHG) and two upwelling (negative VHG) reaches.
Reaches were approximately 250 m apart and the
substratum was comprised of small gravel and
coarse sand (27%), medium sand (60%), and fine
sand (13%) according to a modified Wentworth scale
(Cummins, 1962). The VHG, determined by ran-
domly placing multiple minipiezometers throughout
the study reaches (Lee & Cherry, 1978), averaged
)0.01 cm cm)1 in downwelling reaches and
0.03 cm cm)1 in upwelling reaches.
Ambient sampling
Surface water, pore water and invertebrate samples
were collected to provide background information for
this study. An Orion 835 DO meter (Orion Research
Incorporated, Boston, MA, USA) was used to measure
temperature and DO concentration in the field. Surface
water and pore water samples (n ¼ 6 per reach) were
collected with a hand vacuum-pump attached to a 0.6-
cm diameter piezometer. Surface water was collected
at mid-depth of the thalweg and sediment pore water
was collected at 12–13 cm deep at randomly selected
locations within the downwelling and upwelling
reaches. Water samples were kept on ice in the dark
until returned to the laboratory for processing. Water
subsamples were analysed for NH4-N, NO3-N, PO4-P,
and DOC. Water subsamples for NH4-N analyses were
frozen until analysed with the manual method of
Solorzano (1969) and Harwood & Kuhn (1970).
Subsamples for NO3-N and PO4-P analyses were
filtered through Gelman 0.45 lm pore size membrane
filters and frozen. NO3-N concentration was measured
using the hydrazine method (Kamphake, Hannah &
Stream subsurface hydrology and leaf breakdown 99
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

Cohen, 1967), and PO4-P concentration with the
manual ascorbic acid method (Murphy & Riley,
1962). For DOC analyses, water was filtered through
preashed Whatman GF ⁄F glass fibre filters (0.7 lm
pore size). Sodium azide (5 lg mL)1 of 0.54 MM) was
added to each sample, and samples were refrigerated
until DOC concentration was determined with a
Shimadzu 5000 Total Organic Carbon Analyser (Shi-
madzu Scientific Instruments, Columbia, MO, USA).
Invertebrates were collected using sediment cores
(7.5 cm diameter) from upwelling and downwelling
reaches (n ¼ 6 per reach). From each core, the sedi-
ments from 0 to 5 cm and 10–15 cm deep were
separated. Magnesium chloride solution was added
to each bag to promote release of fauna from the
sediments (Palmer & Strayer, 1996). Rose Bengal was
also added to stain the invertebrates and ease detec-
tion. Invertebrates were identified at 100· under a
dissecting microscope.
Leaf pack construction, deployment, and analysis
During September–October 1996, falling sugar maple
(Acer saccarhum Marsh) leaves were collected on
fibreglass nets suspended between riparian trees
along Bertrand Creek. These leaves were pooled, air-
dried for about 7 days, randomly divided into
approximately 5-g (dry mass) packs, and their petioles
were secured together with bands of 2-cm width
labelling tape. In order to eliminate any potential
enclosure effects, as reported by Rosemond, Pringle &
Ramırez (1998), the leaf packs were not placed within
a mesh bag or cage. After construction, each pack was
weighed to the nearest 0.01 g and taken to the field in
a separate plastic bag.
The leaf packs were placed in Bertrand Creek at the
end of October, within few days of peak litterfall. In
the field, we carefully removed each pack from its bag
and soaked it in a bucket of stream water for several
minutes. This procedure softened the leaves and
reduced immediate mechanical damage when the
leaves were exposed to the stream current. After the
packs were removed from the plastic bags, the latter
were returned to the laboratory where any leaf pieces
that had broken off were weighed and subtracted from
the initial mass of each pack. Ten packs were tied onto
4-m lengths of monofilament, spaced approximately
30 cm apart. Half of the packs were secured at random
to the main line with 5-cm long tethers, while the other
half of the packs were secured with 15-cm long tethers.
In each of the four reaches, four lines of bags were
secured across the stream on transects approximately
30 cm apart. The lines were placed as close to the
sediment–water interface as possible and secured to
stakes on the bank. The packs with 15-cm long tethers
were buried about 10 cm below the sediment–water
interface, whereas the packs with the 5-cm long tethers
were allowed to rest on the sediment surface.
A subset of the leaf packs was collected at random on
days 7, 14, 25, 38 and 120 after initial placement of the
packs in the stream. Because more variability in
response variables was expected as the study pro-
gressed (Boulton & Boon, 1991), more packs were
collected later in the study. From each site, three packs
per treatment (buried or surface) were collected on
days 7 and 15, four packs were collected on days 25 and
38, and five packs were collected on day 120. The total
number of packs also varied because of occasional leaf
pack losses. When the packs were collected, a plastic
bag was held open just downstream from each pack, the
tether was cut, and the pack was carefully and quickly
slid into the bag. Before buried packs were collected,
they were carefully exposed by gradually removing
overlying sediment; we saw little evidence that leaf
fragments or invertebrates escaped during this process,
but some loss was possible.
In the laboratory, each leaf pack was gently rinsed
with tap water over a 45-lm mesh sieve, and the
contents of the sieve were then transferred to a plastic
bag and preserved with 70% ethanol. Rose bengal
was also added to each bag. After at least 48 h, the
contents of the bag were transferred to a 45-lm mesh
sieve, rinsed thoroughly to remove excess dye, and
examined under a dissecting microscope at 100·.
Invertebrates collected on days 7, 38 and 120 were
identified to the lowest practical taxonomic level.
The rinsed leaf packs were placed in paper bags,
hung in the laboratory, air-dried to constant mass and
weighed to the nearest 0.01 g. Percent mass remaining
was calculated as
[Final dry mass/(initial dry mass
� pieces broken off in plastic bag)] � 100
Data analyses
Leaf decomposition rates (k) for each of the four
hydrology–depth treatment combinations (down-
100 D.C. Tillman et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

welling buried, downwelling surface, upwelling bur-
ied, upwelling surface) were calculated using an
exponential decay model, Mt ¼ Moe–kt, where Mo is
the initial mass, Mt is the mass remaining at time t,
and t is the time in days (Benfield, 1996). Data for days
7, 14, 25, 38 and 120 were included in the model (day 0
was set at 100% but not sampled with true replicates).
The fit of the lines to the exponential model was
assessed using linear regression, and the slopes of the
lines were statistically compared with ANCOVAANCOVA (Zar,
1996). To examine patterns in leaf mass loss and leaf
pack invertebrate communities at specific periods
during the study (beginning, middle and end), leaf
mass loss data and invertebrate data (expressed both
on a per leaf pack and a per unit mass basis) were
analysed separately by date (days 7, 38 and 120) using
a split-plot ANOVAANOVA (Johnson & Leone, 1977; Steel &
Torrie, 1980). The VHG direction (upwelling or
downwelling) was used as the whole plot factor,
and depth (surface or buried) served as the sub plot
factor. The split-plot ANOVAANOVA is appropriate because
the treatments required experimental units of unequal
sizes (Johnson & Leone, 1977; Steel & Torrie, 1980). In
our case, hydrology treatments required a larger
spatial scale than did depth treatments. Data were
log-transformed when necessary to meet the assump-
tions of ANOVAANOVA, and post hoc comparisons were
performed with Tukey’s multiple comparison tests
(Zar, 1996).
Results
Ambient sampling
Water temperature and concentrations of NH4-N,
PO4-P or DOC did not significantly differ between
surface and pore water or between water from
downwelling and upwelling reaches (Table 1).
However, NO3-N concentration and DO concentra-
tion were significantly lower in upwelling reach
pore water than in downwelling reach pore water
or in any surface water (NO3-N: P < 0.05; DO:
P < 0.001). Invertebrate abundance and richness
were significantly higher in sediments 0–5 cm deep
than in sediments 10–15 cm deep (P < 0.05; Ta-
ble 2). Abundance and richness also were signifi-
cantly higher in downwelling than in upwelling
reaches (P < 0.05).
Leaf breakdown
An exponential decay model regressing each hydrol-
ogy-depth combination against time significantly fit the
leaf breakdown data from each treatment (Table 3).
Decay coefficients ranged from )0.0097 for upwelling
buried packs (slowest processing) to )0.022 for
upwelling surface packs (fastest processing), and the
slopes of the regression lines were not parallel
(F3,127 ¼ 7.43, P ¼ 0.0001; Fig. 1). Mass loss over the
120 days was significantly slower for leaves buried in
Table 1 Mean (1 SE; n ¼ 12) physical and chemical parameters measured in Bertrand Creek for surface water and pore water
(collected from 12 to 13 cm deep in sediments) in downwelling and upwelling reaches
Parameter
Surface water Pore water
Downwelling Upwelling Downwelling Upwelling
Temperature (�C) 10.2 (0.2) 10.2 (0.3) 9.9 (0.3) 10.5 (0.2)
Dissolved oxygen (mg L)1) 11.3 (0.4) 11.6 (0.7) 8.9 (1.0) 2.6 (0.6)
NO3-N (lg L)1) 664 (59) 664 (37) 737 (68) 305 (157)
NH4-N (lg L)1) 40.6 (6.6) 35.4 (3.7) 34.3 (6.7) 36.4 (4.9)
PO4-P (lg L)1) 21.1 (2.5) 21.0 (2.0) 25.3 (1.9) 20.0 (1.4)
DOC (lg L)1) 2.75 (0.85) 3.98 (1.34) 4.10 (1.13) 6.09 (2.48)
Table 2 Mean (1 SE; n ¼ 12) invertebrate density and taxon richness in sediment cores collected from Bertrand Creek
Metric
Surface (0–5 cm) Buried (10–15 cm)
Downwelling Upwelling Downwelling Upwelling
Density (invertebrates dm)3) 87.9 (25.7) 35.2 (7.2) 21.8 (7.0) 8.8 (2.5)
Taxon richness (taxa dm)3) 6.50 (0.86) 4.75 (0.81) 3.50 (0.71) 3.17 (0.68)
Stream subsurface hydrology and leaf breakdown 101
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

upwelling reaches than for buried (P ¼ 0.0087) and
surface leaves (P < 0.0001) in downwelling reaches.
Split-plot ANOVAANOVA was used to analyse treatment
effects on days 7, 38 and 120, which represented early,
mid- and late processing (Fig. 1). Surface leaves had
significantly more mass remaining after 7 days than
did buried leaves (F1,16 ¼ 31.6, P ¼ 0.0004), but there
was no effect of hydrology on leaf mass at this time
(F1,16 ¼ 0.473, P ¼ 0.50). In contrast, after 38 days
buried leaves had significantly more mass remaining
than did surface leaves (F1,24 ¼ 5.83, P ¼ 0.024), and
leaves in upwelling reaches had more mass remaining
than did leaves in downwelling reaches (F1,24 ¼ 6.90,
P ¼ 0.015). A depth effect was also observed on day
120, with significantly more mass remaining for
buried than for surface leaves (F1,22 ¼ 6.68,
P ¼ 0.017), but hydrology had no significant effect
on leaf mass remaining on day 120 (F1,22 ¼ 2.42,
P ¼ 0.14).
Invertebrate colonisation
For most treatment combinations and dates, Chiro-
nomidae, Nematoda or Rotifera were the most com-
mon invertebrate taxa per unit of leaf mass (Fig. 2).
Taxon richness increased over time (Fig. 3). Inverte-
brates initially colonised surface leaf packs more
rapidly than buried packs. At day 7, surface packs
contained more individuals per unit mass
(F1,15 ¼ 16.3, P ¼ 0.0011; Fig. 2) and more taxa per
unit mass (F1,15 ¼ 31.1, P < 0.0001; Fig. 3) than did
buried packs. At 38 days, the effects of burial on
invertebrate communities were not evident, whereas
the effects of local subsurface hydrology became more
important. Packs in downwelling reaches contained
more individuals per unit mass (F1,20 ¼ 17.2,
P ¼ 0.0005) and more taxa per unit mass
(F1,20 ¼ 6.43, P ¼ 0.020) than did packs in upwelling
reaches. At day 120, packs in downwelling reaches
still contained more individuals per unit mass
(F1,21 ¼ 7.46, P ¼ 0.012), but degree of burial did not
significantly affect invertebrate density (F1,21 ¼ 1.70,
P ¼ 0.21). Neither degree of burial nor hydrology
significantly influenced taxa richness at day 120
(F1,21 ¼ 0.300, P ¼ 0.59; F1,21 ¼ 1.54, P ¼ 0.23). Patterns
Fig. 1 Mean (±1 SE) percent leaf dry mass remaining as a
function of time from initial leaf pack placement in Bertrand
Creek. P-values given for days 7, 38 and 120 are for a split-plot
A N O V AA N O V A, with hydrology as the whole plot and depth as the sub
plot.
Fig. 2 Abundance of most common taxa found in leaf packs in
Bertrand Creek (n ¼ 2–5). P-values are for a split-plot A N O V AA N O V A,
with hydrology as the whole plot and depth as the sub plot.
Error bars represent +1 SE of total abundance. DB, downwelling
buried; DS, downwelling surface; UB, upwelling buried; US,
upwelling surface.
Table 3 Decay coefficients (k), R2 values and regression
P-values of leaf breakdown for the four treatment combinations
over time
Treatment k R2 P
Downwelling buried ) 0.017 0.88 < 0.001
Downwelling surface ) 0.021 0.89 < 0.001
Upwelling buried ) 0.0097 0.62 < 0.001
Upwelling surface ) 0.022 0.93 < 0.001
102 D.C. Tillman et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107
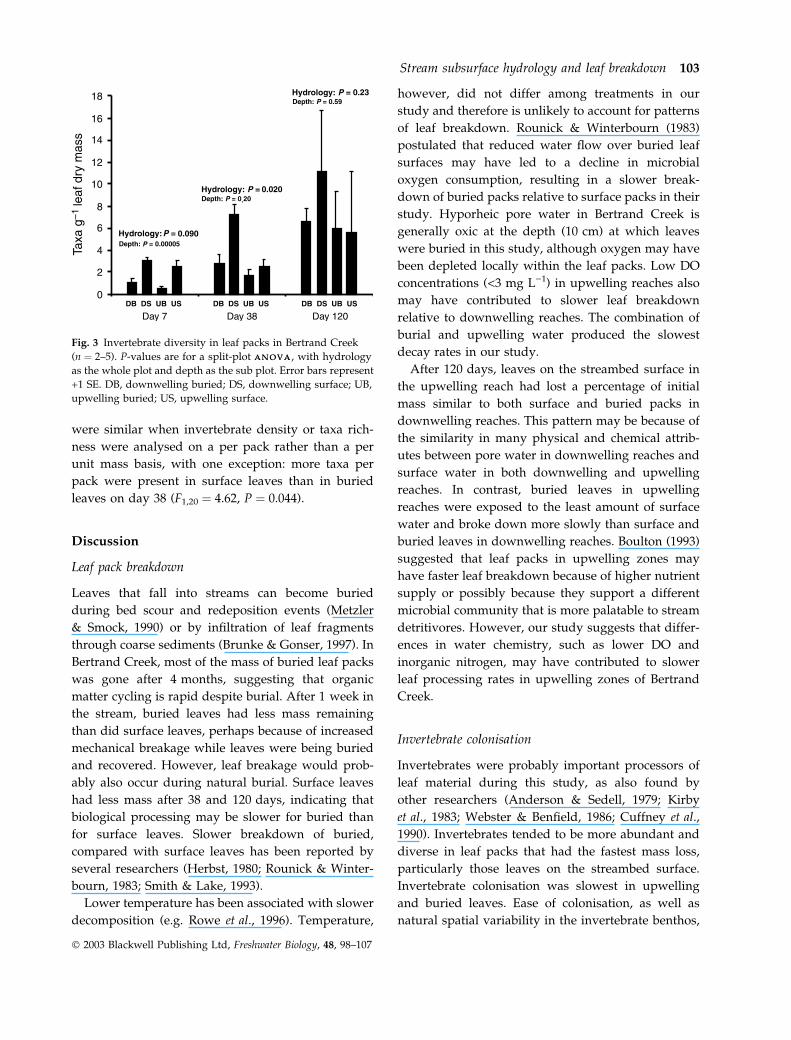
were similar when invertebrate density or taxa rich-
ness were analysed on a per pack rather than a per
unit mass basis, with one exception: more taxa per
pack were present in surface leaves than in buried
leaves on day 38 (F1,20 ¼ 4.62, P ¼ 0.044).
Discussion
Leaf pack breakdown
Leaves that fall into streams can become buried
during bed scour and redeposition events (Metzler
& Smock, 1990) or by infiltration of leaf fragments
through coarse sediments (Brunke & Gonser, 1997). In
Bertrand Creek, most of the mass of buried leaf packs
was gone after 4 months, suggesting that organic
matter cycling is rapid despite burial. After 1 week in
the stream, buried leaves had less mass remaining
than did surface leaves, perhaps because of increased
mechanical breakage while leaves were being buried
and recovered. However, leaf breakage would prob-
ably also occur during natural burial. Surface leaves
had less mass after 38 and 120 days, indicating that
biological processing may be slower for buried than
for surface leaves. Slower breakdown of buried,
compared with surface leaves has been reported by
several researchers (Herbst, 1980; Rounick & Winter-
bourn, 1983; Smith & Lake, 1993).
Lower temperature has been associated with slower
decomposition (e.g. Rowe et al., 1996). Temperature,
however, did not differ among treatments in our
study and therefore is unlikely to account for patterns
of leaf breakdown. Rounick & Winterbourn (1983)
postulated that reduced water flow over buried leaf
surfaces may have led to a decline in microbial
oxygen consumption, resulting in a slower break-
down of buried packs relative to surface packs in their
study. Hyporheic pore water in Bertrand Creek is
generally oxic at the depth (10 cm) at which leaves
were buried in this study, although oxygen may have
been depleted locally within the leaf packs. Low DO
concentrations (<3 mg L)1) in upwelling reaches also
may have contributed to slower leaf breakdown
relative to downwelling reaches. The combination of
burial and upwelling water produced the slowest
decay rates in our study.
After 120 days, leaves on the streambed surface in
the upwelling reach had lost a percentage of initial
mass similar to both surface and buried packs in
downwelling reaches. This pattern may be because of
the similarity in many physical and chemical attrib-
utes between pore water in downwelling reaches and
surface water in both downwelling and upwelling
reaches. In contrast, buried leaves in upwelling
reaches were exposed to the least amount of surface
water and broke down more slowly than surface and
buried leaves in downwelling reaches. Boulton (1993)
suggested that leaf packs in upwelling zones may
have faster leaf breakdown because of higher nutrient
supply or possibly because they support a different
microbial community that is more palatable to stream
detritivores. However, our study suggests that differ-
ences in water chemistry, such as lower DO and
inorganic nitrogen, may have contributed to slower
leaf processing rates in upwelling zones of Bertrand
Creek.
Invertebrate colonisation
Invertebrates were probably important processors of
leaf material during this study, as also found by
other researchers (Anderson & Sedell, 1979; Kirby
et al., 1983; Webster & Benfield, 1986; Cuffney et al.,
1990). Invertebrates tended to be more abundant and
diverse in leaf packs that had the fastest mass loss,
particularly those leaves on the streambed surface.
Invertebrate colonisation was slowest in upwelling
and buried leaves. Ease of colonisation, as well as
natural spatial variability in the invertebrate benthos,
Fig. 3 Invertebrate diversity in leaf packs in Bertrand Creek
(n ¼ 2–5). P-values are for a split-plot A N O V AA N O V A, with hydrology
as the whole plot and depth as the sub plot. Error bars represent
+1 SE. DB, downwelling buried; DS, downwelling surface; UB,
upwelling buried; US, upwelling surface.
Stream subsurface hydrology and leaf breakdown 103
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

may have contributed to these differences. Inverteb-
rate drift would have led to more rapid colonisation
of surface than buried packs. Initially, we found
fewer individuals and taxa in buried packs than in
surface packs, similar to the findings of Rounick &
Winterbourn (1983) in a New Zealand stream. From
core samples, we also found that surface inverte-
brates in Bertrand Creek were generally more
abundant and diverse than invertebrates within the
sediments. Gibert et al. (1990) suggested that hypor-
heic communities may be less diverse than surface
communities for reasons that include reduced pore
space and decreased availability of oxygen and
organic matter.
In Bertrand Creek sediments, hyporheic inverte-
brates were more abundant and diverse in downwel-
ling than in upwelling reaches, consistent with the
patterns that we observed in leaf packs. Possible
explanations for the greater abundance and diversity
of invertebrates in downwelling reaches include
higher DO (from surface water) and greater food
availability (e.g. periphyton products and leaf frag-
ments drawn into the pore spaces). Previous studies
have shown a positive relationship between the
diversity of interstitial invertebrate and DO concen-
tration (Strommer & Smock, 1989; Plenet & Gibert,
1994). Lenting, Williams & Fraser (1997) found that a
higher quality of hyporheic organic matter (as
measured by the carbon-to-nitrogen ratio) was asso-
ciated with increased invertebrate abundance and
taxon richness. Although leaves are a food resource,
Boulton & Foster (1998) did not find significant
changes in local invertebrate abundance and taxon
richness in streams with buried leaf packs. In Bertrand
Creek, a combination of better physical conditions and
more food resources may have provided invertebrates
with better habitat in downwelling reaches.
Invertebrate taxonomic composition was quite
variable both within and amongst sites. Although
abundant in this study, rotifers and tardigrades have
rarely been reported as common leaf pack inverte-
brates. Other studies have generally used a larger
mesh size than the 45-lm mesh sieve used in this
study to retain invertebrates (Smith & Lake, 1993;
Meegan, Perry & Perry, 1996; Rowe et al., 1996) and
consequently may have missed these small inverte-
brates. Amphipods (Gammarus sp.) were the only
macroinvertebrates commonly found in the leaf
packs, although other large shredders such as cranefly
larvae (Tipula sp.) were present in the stream and
probably contributed to leaf breakdown.
Implications for stream processes
Several factors, including degree of burial, VHG, local
chemistry, water temperature, hydraulics and inver-
tebrate colonisation, probably operate in concert to
determine the breakdown of riparian leaves in
streams. Our study showed that variation in subsur-
face hydrology can affect leaf processing rates, with
faster decomposition occurring in downwelling rea-
ches than in upwelling reaches. Furthermore, burial of
organic matter may reduce the rate of leaf processing
when compared with the surface environment. Thus,
the hyporheic zone may serve as a temporary reser-
voir for organic matter in streams and affect the
overall pattern of stream metabolism.
Because over 20% of autumn leaf input can become
buried in stream sediments (Metzler & Smock, 1990),
processing of buried leaves should be considered in
future studies of stream carbon cycling. Newbold
et al. (1982) noted that ‘the carbon turnover length is
the ratio of the downstream transport flux of carbon
(per unit width of stream) to the respiratory utilisation
of carbon (per unit area of stream bottom)’. Although
our study did not quantify leaf transport, this flux was
probably low because Bertrand Creek is a highly
retentive headwater stream with many debris dams.
Furthermore, Webster et al. (1999) concluded that, in
headwater streams, leaves are generally broken down
near their point of entry into the stream. Our study
indicates that leaf litter carbon may be processed more
rapidly in downwelling reaches than in upwelling
reaches because of faster subsurface processing. As a
consequence, the carbon turnover length may be
shorter and reach efficiency (sensu Newbold et al.,
1982) greater in downwelling than in upwelling
reaches. Although local subsurface hydrology may
affect carbon processing rates, the eventual fate of
the unrespired carbon depends on whether it leaves
the hyporheic zone via scour (i.e. resuspension) or
percolation. If the carbon is not scoured from the
streambed during high flow, it may percolate through
the sediment interstices as fine particulates or follow
hyporheic flow paths in dissolved form (Mathieu,
Essafi & Doledec, 1991; Fiebig, 1995; Marmonier et al.,
1995; Claret et al., 1997). In that case, the faster
decomposition of organic matter in downwelling
104 D.C. Tillman et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

reaches may result in carbon leaving the hyporheic
zone more rapidly in downwelling reaches than in
upwelling reaches. As a result, leaf packs in down-
welling reaches may act as a source of particulate and
dissolved organic carbon to groundwater systems,
which may then be transported to downstream
upwelling areas. In this way, hyporheic zones may
be important sites for a subsurface carbon ‘spiral’,
which can have significant implications for carbon
metabolism in stream ecosystems.
Acknowledgments
We are grateful to Katherine Mosca for her help in the
field and laboratory and to Eric Strauss for statistical
assistance. We also thank the Brazo and White
families for access to the study sites. This research
was supported by an NSF Predoctoral Fellowship to
DCT, and by U.S. Geological Survey Grant no. 1434-
HQ-96-GR-02669 through Purdue University.
References
Anderson N.H. & Sedell J.R. (1979) Detritus processing
by macroinvertebrates in stream ecosystems. Annual
Review of Entomology, 24, 351–377.
Benfield E.F. (1996) Leaf breakdown in stream ecosys-
tems. In: Methods in Stream Ecology (Eds F.R. Hauer &
G.A. Lamberti), pp. 579–590. Academic Press, San
Diego.
Boulton A.J. (1993) Stream ecology and surface-hypor-
heic hydrologic exchange: implications, techniques,
and limitations. Australian Journal of Marine and Fresh-
water Research, 44, 553–564.
Boulton A.J. & Boon P.I. (1991) A review of methodology
used to measure leaf litter decomposition in lotic
environments: time to turn over an old leaf? Australian
Journal of Marine and Freshwater Research, 42, 1–43.
Boulton A.J. & Foster J.G. (1998) Effects of buried leaf
litter and vertical hydrologic exchange on hyporheic
water chemistry and fauna in a gravel-bed river in
northern New South Wales, Australia. Freshwater
Biology, 40, 229–243.
Boulton A.J. & Stanley E.H. (1995) Hyporheic processes
during flooding and drying in a Sonoran Desert
stream. Archiv fur Hydrobiologie, 134, 27–52.
Brunke M. & Gonser T. (1997) The ecological significance
of exchange processes between rivers and ground-
water. Freshwater Biology, 37, 1–33.
Claret C., Marmonier P., Boissier J.-M., Fontvielle D. &
Blanc P. (1997) Nutrient transfer between parafluvial
interstitial water and river water: influence of gravel
heterogeneity. Freshwater Biology, 37, 657–670.
Creuze des Chatelliers M. (1991) Geomorphological
processes and discontinuities in the macrodistribution
of the interstitial fauna. A working hypothesis. Ver-
handlungen der Internationalen Vereinigung fur Theore-
tische und Angewandte Limnologie, 24, 1609–1612.
Cuffney T.F., Wallace J.B. & Lugthart G.J. (1990)
Experimental evidence quantifying the role of benthic
invertebrates in organic matter dynamics of headwater
streams. Freshwater Biology, 23, 281–299.
Cummins K.W. (1962) An evaluation of some techniques
for the collection and analysis of benthic samples with
special emphasis on lotic waters. American Midland
Naturalist, 67, 477–504.
Dole-Olivier M.-J., Marmonier P. & Beffy J.-L. (1997)
Response of invertebrates to lotic disturbance: is the
hyporheic zone a patchy refugium? Freshwater Biology,
37, 257–276.
Dole-Olivier M.-J., Marmonier P., Creuze des Chatelliers
M. & Martin D. (1994) Interstitial fauna associated with
the alluvial floodplains of the Rhone River (France). In:
Groundwater Ecology (Eds J. Gibert, D.L. Danielopol &
J.A. Stanford), pp. 314–346. Academic Press, San
Diego.
Fairchild J.F., Boyle T.P. & Robinson-Wilson E. (1983)
Microbial action in detrital leaf processing and the
effects of chemical perturbation. Dynamics of Lotic
Ecosystems (Eds T.D. Fontaine & S.M. Bartell), pp. 437–
456. Ann Arbor Science Publishers, Ann Arbor, MI.
Fiebig D.M. (1995) Groundwater discharge and its
contribution of dissolved organic carbon to an upland
stream. Archiv fur Hydrobiologie, 134, 129–155.
Findlay S. (1995) Importance of surface-subsurface
exchange in stream ecosystems: the hyporheic zone.
Limnology and Oceanography, 40, 159–164.
Fisher S.G. (1973) Energy flow in Bear Brook, New
Hampshire: an integrative approach to stream ecosys-
tem metabolism. Ecological Monographs, 43, 421–439.
Gibert J., Dole-Olivier M.-J., Marmonier P. & Vervier P.
(1990) Surface water-groundwater ecotones. In: The
Ecology and Management of Aquatic-Terrestrial Ecotones
(Eds R.J. Naiman & H. Decamps), pp. 199–225. Unesco,
Paris.
Godbout L. & Hynes H.B.N. (1982) The three dimen-
sional distribution of the fauna in a single riffle in a
stream in Ontario. Hydrobiologia, 97, 87–96.
Harwood J.E. & Kuhn A.L. (1970) A colorimetric method
for ammonia in natural waters. Water Research, 4, 805–811.
Hendricks S.P. & White D.S. (1991) Physicochemical
patterns within a hyporheic zone of a northern
Michigan river, with comments on surface water
Stream subsurface hydrology and leaf breakdown 105
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

patterns. Canadian Journal of Fisheries and Aquatic
Sciences, 48, 1645–1654.
Hendricks S.P. & White D.S. (1995) Seasonal biogeo-
chemical patterns in surface water, subsurface hypor-
heic, and riparian ground water in a temperate stream
ecosystem. Archiv fur Hydrobiologie, 134, 459–490.
Herbst G.N. (1980) Effects of burial on food value and
consumption of leaf detritus by aquatic invertebrates
in a lowland forest stream. Oikos, 35, 411–424.
Johnson N.L. & Leone F.C. (1977) Statistics and Experi-
mental Design in Engineering and the Physical Sciences.
John Wiley and Sons, New York.
Jones J.B., Fisher S.G. & Grimm N.B. (1995) Vertical
hydrologic exchange and ecosystem metabolism in a
Sonoran desert stream. Ecology, 76, 942–952.
Kamphake L.J., Hannah S.A. & Cohen J.M. (1967)
Automated analysis for nitrate by hydrazine reduction.
Water Research, 1, 205–216.
Kirby J.M., Webster J.R. & Benfield E.F. (1983) The role of
shredders in detrital dynamics of permanent and
temporary streams. In: Dynamics of Lotic Ecosystems
(Eds T.D. Fontaine III & S.M. Bartell), pp. 425–435. Ann
Arbor Science Publishers, Ann Arbor, MI.
Lee D.R. & Cherry J. (1978) A field exercise on ground-
water flow using seepage meters and mini-piezo-
meters. Journal of Geological Education, 27, 6–10.
Lenting N., Williams D.D. & Fraser B.G. (1997) Qualitative
differences in interstitial organic matter and their
effect on hyporheic colonization. Hydrobiologia, 344,
19–26.
Marmonier P., Fontvielle D., Gibert J. & Vanek V. (1995)
Distribution of dissolved organic carbon and bacteria
at the interface between the Rhone River and its
alluvial aquifer. Journal of the North American Bentholo-
gical Society, 14, 382–392.
Mathieu J., Essafi K. & Doledec S. (1991) Dynamics of
particulate organic matter in bed sediments of two
karst streams. Archiv fur Hydrobiologie, 122, 199–211.
McElravy E.P. & Resh V.H. (1991) Distribution and
seasonal occurrence of the hyporheic fauna in a
northern California stream. Hydrobiologia, 220, 233–246.
Meegan S.K., Perry S.A. & Perry W.B. (1996) Detrital
processing in streams exposed to acidic precipitation
in the Central Appalachian Mountains. Hydrobiologia,
339, 101–110.
Metzler G.M. & Smock L.A. (1990) Storage and dynamics
of subsurface detritus in a sand-bottomed stream.
Canadian Journal of Fisheries and Aquatic Sciences, 47,
588–594.
Murphy J. & Riley J.P. (1962) A modified single solution
method for the determination of phosphate in natural
water. Analytica Chimica Acta, 27, 31–36.
Newbold J.D., Mulholland P.J., Elwood J.W. & O’Neill
R.V. (1982) Organic carbon spiraling in stream ecosys-
tems. Oikos, 38, 266–272.
Palmer M.A., Covich A.P., Lake S. et al. (2000) Linkages
between aquatic sediment biota and life above sedi-
ments as potential drivers of biodiversity and ecolog-
ical processes. Bioscience, 50, 1062–1075.
Palmer M.A. & Strayer D.L. (1996) Meiofauna. In: Methods
in Stream Ecology (Eds F.R. Hauer & G.A. Lamberti), pp.
315–338. Academic Press, San Diego.
Plenet S. & Gibert J. (1994) Invertebrate community
responses to physical and chemical factors at the
river ⁄aquifer interaction zone. I. Upstream from the
city of Lyon. Archiv fur Hydrobiologie, 132, 165–189.
Reed F.C. (1979) Decomposition of Acer rubrum leaves at
three depths in a eutrophic Ohio lake. Hydrobiologia, 64,
195–197.
Rosemond A.D., Pringle C.M. & Ramırez A. (1998)
Macroconsumer effects on insect detritivores and
detritus processing in a tropical stream. Freshwater
Biology, 39, 515–523.
Rounick J.S. & Winterbourn M.J. (1983) Leaf processing
in two contrasting beech forest streams: effects of
physical and biotic factors on litter breakdown. Archiv
fur Hydrobiologie, 96, 448–474.
Rowe J.M., Meegan S.K., Engstrom E.S., Perry S.A. &
Perry W.B. (1996) Comparison of leaf processing rates
under different temperature regimes in three head-
water streams. Freshwater Biology, 36, 277–288.
Schmid-Araya J.M. (1995) Disturbance and population
dynamics of rotifers in bed sediments. Hydrobiologia,
313 ⁄314, 270–290.
Silliman S.E. & Booth D.F. (1993) Analysis of time-series
measurements of sediment temperature for identifica-
tion of gaining vs. losing portions of Juday Creek,
Indiana. Journal of Hydrology, 146, 131–148.
Smith J.J. & Lake P.S. (1993) The breakdown of buried
and surface-placed leaf litter in an upland stream.
Hydrobiologia, 271, 141–148.
Smock L.A., Metzler G.M. & Gladden J.E. (1989) Role
of debris dams in the structure and functioning
of low-gradient headwater streams. Ecology, 70, 764–
775.
Solorzano L. (1969) Determination of ammonia in natural
waters by the phenolypochlorite method. Limnology
and Oceanography, 14, 799–801.
Steel R.G.D. & Torrie J.H. (1980) Principles and Procedures
of Statistics. McGraw-Hill, New York.
Strommer J.L. & Smock L.A. (1989) Vertical distribution
and abundance of riverine invertebrates within the
sandy substrate of a low-gradient headwater stream.
Freshwater Biology, 22, 262–274.
106 D.C. Tillman et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

Suberkropp K. & Chauvet E. (1995) Regulation of leaf
breakdown by fungi in streams: influences of water
chemistry. Ecology, 76, 1433–1445.
Triska F.J., Duff J.H. & Avanzino R.J. (1993) The role of
water exchange between a stream channel and its
hyporheic zone in nitrogen cycling at the terrestrial–
aquatic interface. Hydrobiologia, 251, 167–184.
Valett H.M. (1993) Surface–hyporheic interactions in a
Sonoran Desert stream: hydrologic exchange and diel
periodicity. Hydrobiologia, 259, 133–144.
Vervier P. & Naiman R.J. (1992) Spatial and temporal
fluctuations of dissolved organic carbon in subsurface
flow of the Stillaguamish River (Washington, USA).
Archiv fur Hydrobiologie, 123, 401–412.
Wagner R., Schmidt H.H. & Marxsen J. (1993) The
hyporheic habitat of the Breitenbach, spatial structure
and physicochemical conditions as a basis for benthic
life. Limnologica, 23, 285–294.
Webster J.R. & Benfield E.F. (1986) Vascular plant
breakdown in freshwater ecosystems. Annual Review
of Ecology and Systematics, 17, 567–594.
Webster J.R., Benfield E.F., Ehrman T.P., Schaeffer M.A.,
Tank J.L., Hutchens J.J. & D’Angelo D.J. (1999)
What happens to allochthonous material that falls
into streams? A synthesis of new and published
information from Coweeta. Freshwater Biology, 41,
687–705.
Williams D.D. (1993) Nutrient and flow vector dynamics
at the hyporheic ⁄groundwater interface and their
effects on the interstitial fauna. Hydrobiologia, 251,
185–198.
Zar J.H. (1996) Biostatistical Analysis. Prentice Hall, Upper
Saddle River, NJ.
(Manuscript accepted 24 July 2002)
Stream subsurface hydrology and leaf breakdown 107
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 98–107

Copyright © 2022 FDOKUMEN




















